Embed Size (px)
Citation preview

8i
STRUCTURAL CHANGES IN THE SUPRARENALGLAND OF THE MOUSE DURING PREGNANCY
BY Y. TAMURA,Animal Breeding Research Department, University of Edinburgh.
(With Plates VII and VIII.)
(Received \%ih January 1926.)
CONTENTS.PAGE
1. Introduction 812. Material and methods 823. Description of experiments 824. Discussion 875. Summary 916. References 91
1. INTRODUCTION.
GUIEYSSE (1889) observed that the suprarenal gland of the guinea-pig underwentenlargement during pregnancy, this increase chiefly affecting the zona fasciculata.This observation was confirmed by Kolmer (1912) and extended to the case of therabbit by Kolde (1913) and others. According to Guieysse the hypertrophy of thezona fasciculata in the guinea-pig is due to an enlargement of the cells themselvesand not to an increase in their number. He also states that there is no enlargementof the zona glomerulosa or of the medulla.
An increase in the weight of the suprarenal during pregnancy was noted in therat by Herring (1920) and in the guinea-pig by Verdozzi (1917) and Castaldi(1922). Donaldson (1924) states that there is no change in the weight of the adrenalin the healthy albino rat during the phase of pregnancy lactation. Masui andTamura (1924) have shown that the suprarenal of the mouse increases in size atthe beginning of pregnancy, but that at the same time degeneration of the zonareticularis sets in and proceeds until this zone has completely disappeared so thattowards the end of pregnancy the gland as a whole is greatly reduced in size,although the zona fasciculata itself has increased slightly.
The present work was undertaken in order to examine the effect of unilateralovariotomy upon the suprarenal. Observations were made on unilaterally ovario-tomised mice during pregnancy. The results obtained show that unilateral ovario-tomy is not followed by any change in the histological structure of the adrenal(apart from such variation which is also found in normal non-pregnant females),^Btha t during pregnancy in the unilaterally ovariotomised female both adrenalsundergo similar modifications in structure identical with those that occur in thenormal pregnant female.
BJEB-Ivi 6

82 Y. TAMURA
2. MATERIAL AND METHODS.Ordinary mixed tame mice were used as material. Each experimental
was composed of individuals from one and the same litter, and virgin females froma few litters were kept as controls. Unilateral ovariotomy was performed underether anaesthesia at three, five, seven and eight weeks after birth. The operatedand control animals from each experimental group were kept in the same box. Theovariotomised animals were mated at nine weeks and killed with chloroform at thefirst, second or third pregnancy. They were weighed and measured immediatelyafter killing; the ovaries and other endocrine organs were removed and fixed.
Allen-Bouin, Regaud, and Chura's solutions were used as fixative agents. Thelast solution gave good results for the purposes of examination of mitochondria,granules and chromosomes. The thickness was 6/x. for structure and 3 or 4/a. forgranules examination. The stains used were Delafield and Heidenhain's haema-toxylin for structure, Champy-Kulle's acid-fuchsin-toloidin-aurantia for mito-chondria and granules, and osmic acid for lipoid and fatty substances. The uterusand its contents were weighed; then all foetuses were weighed and their measure-ments taken. It is impossible by examination to determine with exactitude thestage which pregnancy has reached in the mouse. If the length of the foetus isused as a standard for classification, the whole period of pregnancy may be con-veniently divided into the following five stages:
Stage I. Before the formation of the placenta, i.e. the stage of fixation of thefertilised egg, or the coelomic stage.
Stage II. From the formation of the first traces of placenta to the stage whenthe length of the foetus is 5 mm.
Stage III. The length of foetus from 6 to 12 mm.Stage IV. The length of foetus from 13 to 20 mm.Stage V. The length of foetus from 21 mm. and over.In order to determine the range of variation in the length of the foetus during
Stage V, measurements were taken of young at birth and lengths from 21 mm. to27 mm. were obtained. To determine the relative sizes of the different areas of theadrenal a drawing was made of the largest section (fixed in Allen-Bouin solutionfor as short a time as possible in order to prevent shrinkage, and stained withordinary Delafield haematoxylin) and the outline of each zone was traced with theaid of a planimeter.
My thanks are due to Dr F. A. E. Crew for providing accommodation andexperimental material and also for his valuable help and constructive criticism duringthe course of this study.
3. DESCRIPTION OF EXPERIMENTS.Masui and Tamura (1924) have described in detail both histologically and
cytologically the structure of the suprarenal cortex; it is not necessary thereforein this paper to deal specially with this. The changes in the histological strud^Pbof the adrenal during pregnancy will be described with reference to the five stageswhich have been defined.

Structural Changes in Suprarenal Gland of Mouse during Pregnancy 83
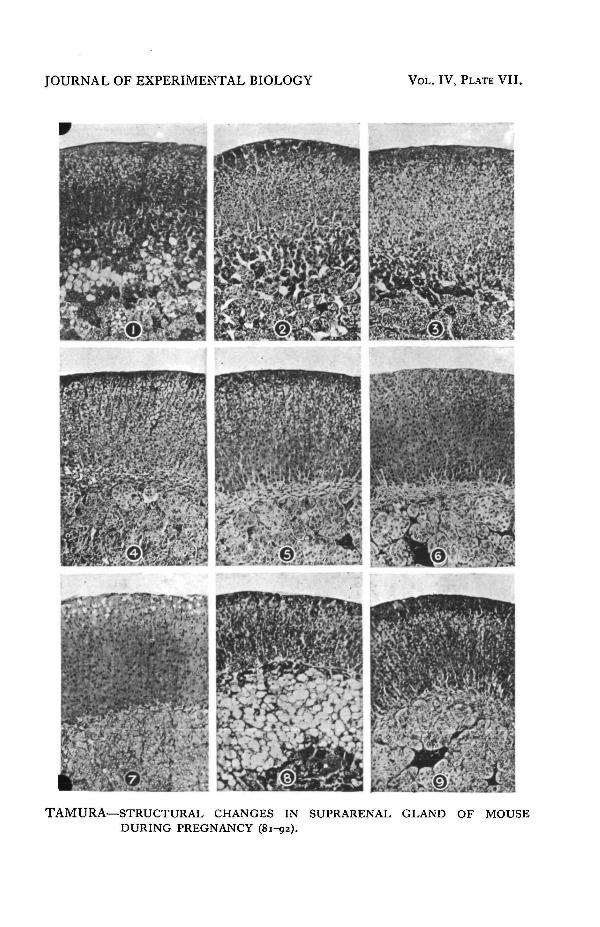
I (PI. VII, figs, i, 2 and 3). During this stage no change in the relative^ of the areas of all zones either in the cortex or in the medulla is to be observed,
as is shown in Tables I and II. The histological structure of the zones, however,shows modification, that of the zona reticularis most and that of the zona fasciculataleast.
Table I.Control
Ex. 6, No. 3Ex. 37, No. 2Ex. g, No. 2Ex. 9, No. 3Ex. 26, No. 2No., i 9No. 2 9No. 3 9No. 4 9No. 5 9
Average
Area in mm.2
Zonaglomerulosa
0-2240-1780-18402240-13201660-1450-1180-1320-1320-164
Zonafasciculata
092605310-540O-4730-526O-OII0881038206250-3950589
Zonareticularis
O-3540-42205290-302O-4730 7 1 00-3420-4800-4540-381O-445
Medulla
022702040289O-2IO0250017802890-1910184O-I540-217
Total
i-73i[•335[•542[•2891-381•665
1-657•171395065
1-423
Group 1
Table II. Stage I. (0-5 mm.)
No. 16 9 p.No. 21 9 p.Ex. 8, No. 2No. 17 9 p.No. 10 9 p.Ex. 23, No. 3Ex. 31, No. 1Ex. 16, No. 3Ex. 21, No. 3
Average
Area in mm.2
Zonaglomerulosa
0-1910-15101930229017601830256016802140196
Zonafasciculata
0-3950-5260531o-6n06580532063905260-736O-573
Zonareticularis
0-5720-5520-56902700549O'433O-3540270
O-339O-434
Medulla
018302240-2460-3020-19201680-263025602820235
Total
341•453•539•412•575•316•512•220•57i438
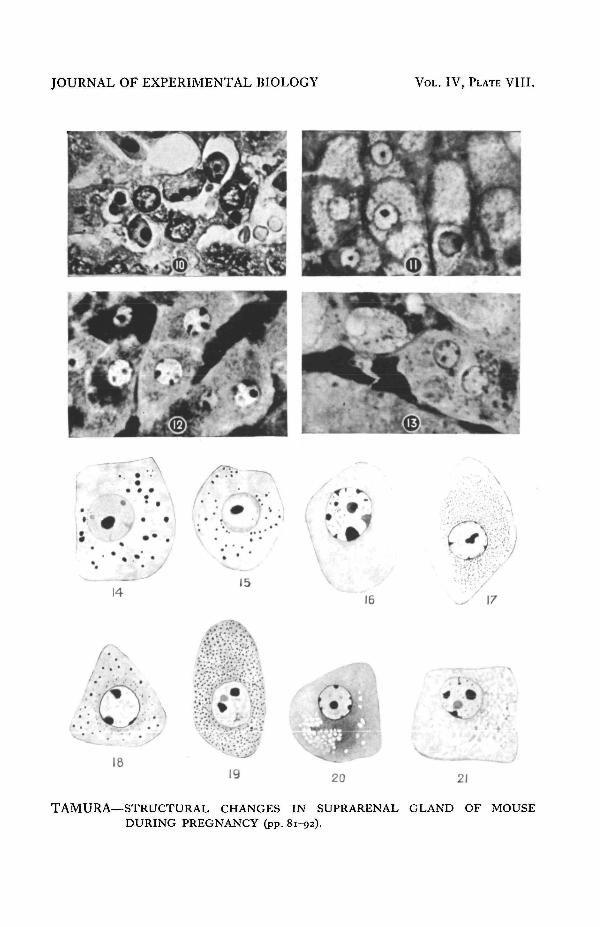
The cells of the zona glomerulosa have become active, some showing mitosis,others becoming enlarged in size whilst minute granules are present as in the glandof the normal non-pregnant female. In the zona fasciculata two types of cellsappear. One is the typical vacuolated fasciculata cells (PI. VIII, figs. 11 and21) and these are present in large numbers throughout the zone; the others aremuch smaller, clear, non-vacuolated cells which lie at the border line betweenthis zone and the zona reticularis. These cells are similar in structure to those ofthe zona reticularis. Signs of mitosis are visible in their nuclei. The region ofthese cells will be termed zona gestationis. The zona reticularis shows a remark-a^Bchange due to a rapid degeneration of the tissues (PI. VIII, fig. 10). Thisdegeneration begins along the whole of the border-line of the zona fasciculataand proceeds centripetally; at an early stage of degeneration a layer composed of
6-2

84 Y. TAMURA
contracted cells, packed closely together, can be seen between the two zones(PI. VII, figs. 2 and 3). In some cases this process is far more advanced tha^P^others; the zona reticularis has, for the most part, degenerated into large vacuolesor connective tissue and is represented by a thin layer of cells adjacent to themedulla, which still retain their original character. The cells of this layer are reducedin size and have irregularly-shaped nuclei with one or more very clear nucleoliand a small quantity of chromatin granules. Cells containing secretory granulesare, when present, very few in number and the size of the granules is smaller thanthat of those in the non-pregnant female. Thus, although at this period the zonareticularis is present, the characteristic reticularis cells are almost absent, thisfinding being in agreement with that of Masui and Tamura.
In the medulla a few mitotic figures are present, but no other changes can beseen.
Stage II (PI. VII, fig. 4). The total area of the section is reduced by about18 per cent, and a remarkable change in the relative sizes of the zonae fasciculataand reticularis has occurred; the latter zone is much reduced in size owing to theabsorption of the tissue, which is the principal cause of the shrinkage of the sectionas a whole. In the zona glomerulosa no change is to be observed as is shown inTables II and III. The area of the zona fasciculata shows an absolute increase byabout 20 per cent., i.e. a relative increase (estimated in relation to the total area ofthe cortex) of about 56 per cent. On the other hand, the area of the zona reticularishas decreased by 84 per cent, in absolute, and 78 per cent, in relative value.
Notwithstanding the remarkable change in the relative size of the area, thehistological structure of the zonae glomerulosa and fasciculata is almost the sameas in the former stage. The smaller, clear and non-vacuolated cells (zona gestationis)of the zona fasciculata have increased in number towards the periphery; amongthem small cells having large clear nuclei and a relatively scanty protoplasm arealso visible. The zona reticularis has either almost or entirely disappeared and isreplaced by connective tissue. When the zona reticularis has degenerated, it doesnot surround the medulla as in the normal gland, but becomes compressed alongthe abscissa of the section and accumulates in elongated processes at each side ofthe medulla along its ordinate. No noticeable change has occurred in the medullaalthough its area has increased slightly. Several mitotic figures are observed asin the former stage.
Stage HI (PI. VII, fig. 5). There is no observable change in the relativesizes of the different zones when compared with those of the previous stage (seeTables III and IV); the zona reticularis has increased by 37 per cent.; this is causedby the presence of large vacuoles (the so-called zona spongiosa). On histologicalexamination the zona glomerulosa is seen to be still developing and mitotic figuresin considerable numbers are visible, mostly in cases when the length of the foetusis 8 mm. and over. Granules, which were described in the previous stage are againfound, though they are now of varying sizes.
The structure of the zona fasciculata in this stage may be of two differenttypes; in one type almost the whole of the area is occupied by typical vacuolated

Structural Changes in Suprarenal Gland of Mouse during Pregnancy 85
f^rciculata cells (as in the former stages) and the vacuoles near the centre area^Ptler than those in the peripheral region. In the second type the central portionis occupied by the clear cells (PI. VIII, fig. 16) described in the previous stage,which are now more numerous. A few mitotic figures at varying stages are found.
Table III.Group 2
Ex. 19, No. 5No. 6 ? p.Ex. 21, No. 4No. 11 9 p.Ex. 21, No. 2
Average
Area in mm.2
Zonaglomerulosa
0-2170097O-I2IO-2IOOI590-161
Zonafasciculata
092104710-710O-5560-7740686
Zonareticularis
0-10500360-050o-o6o0-0850-067
Medulla
042002700-256o-i8o0-2150268
Total
1663087411371-0061-2231-182
Table IV. Stage III. (6-12 mm.)
No. 9 $ p. (6)No. 18 9 p. (6)No. 1 ? p. (7)Ex. 19, No. 6 (8)Ex. 31, No. 2 (8)No. 8 ? p. (9)No. 2 ? p. (9)No. 13 $ p. (10)No. 12 9 p. (10)No. 22 9 p. (io)No. 23 9 p. (10)No. n 2 9 p . (10)No. 7 9 p. (n-5)Ex. 34, No. 4 (12)Ex. ai , No. 1 (12)
Average
Area in mm.2
Zonaglomerulosa
0-2150-1510-1320-20901790-1670-1320-1710122O-22IOI58OO70O-I2I029502350-171
Zonafasciculata
0826o-6io0634083404600-82305840640
°'5470711O'4340682080608640-8090684
Zonareticularis
0-1090-1720-046008800950-0600-06001970-091011302430-0280-085O-IOI0-1140-107
Medulla
02540-1980-232°-4i40-19203510164O-2IIO-249O-29O0-I5I0-20603600-2610-2890-255
Total
1-4041-1311-0441-5450-9261 40109401-2191-0091-335098609861-3721-5011-4471-217
It is noteworthy that when these latter cells are present the vacuolated cellsin the peripheral portion are polyhedral in shape and the nuclei in some cases showdegeneration. The relative size of the area of the medulla in this stage shows adecrease which, being but slight, may be due to fluctuation. Mitotic figures aremore numerous, beginning from about the 8 mm. stage.
Stage IV (PI. VII, figs. 6, 7 and 8). There is no observable change in therelative size of the zona fasciculata, while that of the zona glomerulosa has in-creased by about 26 per cent, as compared with that of former stages, and that ofth^zona reticularis by about 46 per cent.^ B n microscopical examination it is seen that the zona glomerulosa is remarkably
changed. During the previous stage the minute granules were seen to be increasingin size; at this stage, mostly at from about 16 mm., they are seen to be typical
86 Y. TAMURA
secretory granules (PI. VIII, figs. 14 and 15) having a structure identical withthat described by Masui and Tamura (1924) in the cells of the zona reticul^^The maximal secretory activity is found at about 17 mm. As observed in the zonareticularis of the normal non-pregnant female, very big vacuoles or a network ofconnective tissue (PI. VIII,fig. 13) in the cells are also found; the nuclei in thesecases are similar to those of the secretory cells. As shown in Table IV, the areaof this zone takes up on an average about 21 per cent, of the cortex and at itsmaximal 36 per cent.
In the zona fasciculata at this stage the clear cells (PI. VIII, fig. 12), mentionedabove, appear in all cases, although their number varies. In some cases they occupythe whole area of this zone; in others they press the vacuolated cells towardsthe periphery, so that the latter become diminished in size and rectangular orelliptical in shape, and their nuclei are irregularly shaped.
Some of the clear cells, mostly those situated in the peripheral region, show(PI. VIII, figs. 18 and 19) "siderophile bodies" (secretory granules) of varioussizes, while those situated in the central portion do not contain any granules ormitochondria.
Ex. 22, No. 3 (12-5)Ex. 21, No. s (13)No. 25? p. (13)No. 26? p. (13)Ex. 19, No. 3 (14)Ex. 8, No. 1 (14)No. i s ? p. (14)Ex. 34, No. 2 (16)Ex. 6, No. 12 (16)No. 4 ? p. (17)Ex. S3, No. 2 (17)Ex. 30, No. 2 (17-5)No. 27 ? p. (18)Ex. 16, No. 1 (20)
Average
Table V. Stage IV. 13-20 mm/
Area in mm.2
Zonaglomerulosa
023301580-224O2I2OI95O-2I30-1140296O-229O-3520-191Q-3430-1780-2840230
Zonafasciculata
0622O-5390-848O-5570-707O-57905460-94007940822O-555O-5590-59209880689
Zonareticularis
0-1140-127010501840-26402390-080O-I550-1890016O-55500460506O-2OIo-i 98
Medulla
02800-18903090-2500-2250-151°-25702990-2740-368024703890-3020-2610272
Total
1249[•013[•486[•203[•391[•182O-9971-6901-4861-5581-5481-3371-5781-7341389
At this stage the size of the zona reticularis varies very considerably; in one caseit was equal to that of the zona fasciculata (PI. VII, fig. 8), though it consistedof nothing but big vacuoles and a network of connective tissue. In another caseit occupied but 1 per cent, of the cortex (PI. VII, fig. 7) and had the form of a thinline of connective tissue.
No marked change is found in the medulla as regards its microscopical structure;its area is found to be increased by about 6 per cent. The mitotic figures are stillincreasing in number.
Stage V (PI. VII, fig. 9). At this stage the relative size of the areas of the ^glomerulosa, fasciculata, and reticularis have decreased by 17, 5 and 56 per cent,respectively. On histological examination the secretory activity in the zona glomeru-

Structural Changes in Suprarenal Gland of Mouse during Pregnancy 87
is not visible, although in a few cases secretory granules are still observed.P^ise , in the zona fasciculata a remarkable change has occurred. In cases
where the typical fasciculata vacuolated cells which have been pushed towards theperiphery remain, they become enlarged. In other cases, where no fasciculata cellswere present, they appear and the zona gestationis is seen to undergo degenera-tion (PI. VII, fig. 9). Its cells become greatly vacuolated, in some instances theentire cell protoplasm being reduced to a network. The degeneration proceedsfrom the periphery. In some cases the zona disappears entirely and typical fascicu-lata cells occupy the whole area. The proportion of the area of the medulla increasesby 14 per cent, in absolute, and by 30 per cent, in relative, value to the entire areaof the gland.
Table VI. Stage V. (21 mm. and over.)
No. 3 $ p. (21)Ex. 16, No. 2 (21)Ex. io, ? p. (21)Ex. 9, No. 1 (24)Ex. 25, No. 4 (24)Ex. 14, ? p. (24)No. 5 ? p. (24)No. 52 $2 (24)Ex. 6, No. 2 (24)
Average
Area in mm.2
Zonaglomerulosa
0-1000-2340-32102180-1630-167O-22OOIOI0-1890-190
Zonafasciculata
O-53405180-90206580-602°|960-676073606390651
Zonareticularis
O-O2O006601970068O-I55006300750-07000700-087
Medulla
O-3740266051303000-19202980307040801930317
Total
•028•084
[•933•244• 1 1 2•124•278•3i5•091
•245
4. DISCUSSION.
(1) Hypertrophy of the zona fasciculata.
Guieysse stated that in the guinea-pig the hypertrophy of the zona fasciculatais the result of an enlargement of the cells themselves and not of an increase intheir number. However, in the mouse two types of hypertrophy may be observed.One is similar to that described by Guieysse in the guinea-pig. It takes place ata very early period of pregnancy and the cells increase in diameter by about 38 percent., the largest cell increasing from 16/i to 22/4. Guieysse maintained thathypertrophy implies hyperfunctioning of the zona fasciculata, even though he wasunable to demonstrate how this occurred. However, it is reasonable to assume thatin the mouse this hypertrophy is only compensatory following a shrinkage causedby rapid degeneration and complete disappearance of the zona reticularis.
An examination of Tables II, III, V and VI will show that the average area ofthe zona fasciculata in Stage II shows a very great increase as compared with Stage I;and that in Stage V a decrease, as compared with Stage IV. However, the initialincrease is much greater than the subsequent decrease. This is probably due to thef^Bthat the increase in size of the zona fasciculata is permitted by a very rapiddegeneration of the zona reticularis and is thus a process of compensatory hyper-trophy. In the later stage (V), when the zona gestationis has ceased to develop

88 Y. TAMURA
and begins to degenerate, this process is slow and therefore the increasearea of the typical vacuolated fasciculata cell is very gradual.
Another type of the hypertrophy of the zona fasciculata is that caused by thedevelopment of the zona gestationis within the zona fasciculata. The new zoneincreases gradually in size and at Stage IV occupies almost the whole area of thezona fasciculata; at Stage V it ceases to develop and begins to degenerate and con-sequently the area of the zona fasciculata becomes reduced. As shown in Table V,the area of the zona fasciculata is greatest at Stage IV.
(2) The zona gestationis.
Origin. As stated above, when at an early period of pregnancy the zona reti-cularis begins to degenerate, there appears on the border line between it and thezona fasciculata a layer of degenerated cells, which clearly separates these twozones. At the same time one can identify at the inner margin of the zona fasciculataclear cells which increase gradually in number and in the third quarter of preg-nancy (i.e. 17 mm. in length) occupy almost the whole area of the zona fasciculata,while during the last quarter they become reduced in number. As stated above,the vacuoles of the cells in the inner portion of the zona fasciculata are smaller insize than those of the outer one. Mitotic figures are present in the cells up to themiddle of pregnancy.
From these facts it is seen that the zona gestationis arises on the border linebetween the zona fasciculata and the zona reticularis. In a non-pregnant femalethe cells of this zone cannot be distinguished from those of the zona reticularis,but the occurrence of pregnancy stimulates their activity and produces structuralchanges.
Structure. At the early period of pregnancy the cells do not show any signsof secretory activity and have a very clear protoplasm and nucleus (PI. VIII, fig. 16).Some of them show mitotic figures which increase gradually in number up to thebeginning of Stage IV, when minute granules appear which are arranged irregularlyaround the nucleus (PI. VIII, fig. 17); at the 17 mm. stage some cells showtypical secretory granules (PI. VIII, figs. 18 and 19), while at Stage V (21 mm.and over) the granules change into vacuoles (PI. VIII, fig. 20) which are similarto those of typical fasciculata cells. It is noteworthy that the change begins alongthe whole outer margin of the zone and proceeds inwards.
Kolmer (1912) states that "Alle Rinderelemente aus der Keimschicht derglomerulosa und zwar erst durch Amitose, dann durch Mitose hervorgehen."However, the course of the development and degeneration of the zona gestationisis not in agreement with this hypothesis, and it is more reasonable to assume thatevery zone has an independent origin, as well as developmental and functionalactivity.
(3) Zona reticularis.
The degeneration of this zone was noted by Kolmer (1912), andTamura (1924); and its complete disappearance by the latter workers. The resultsof the present investigation confirm the observation that it disappears entirely and

Structural Changes in Suprarenal Gland of Mouse during Pregnancy 89
loudly at a very early period of pregnancy. During Stage IV large vacuoles taket ^ m e site of this zone (PI. VII, fig. 8). This "zona spongiosa," according toGuieysse and others, is a characteristic feature of the suprarenal gland duringpregnancy in the guinea-pig. In the mouse it is present only in the earlier partof Stage I and in Stage IV, and is not characteristic. The vacuoles are filledwith a fatty substance which stains black with osmic acid.
(4) Zona glomerulosa.
According to Guieysse, no hypertrophy occurs in this zone. However, in thecourse of this investigation, it was observed at certain periods that an enlargementof this zone occurred resulting from an increase in number of the constituent cellsand a heightened activity. During Stage IV there is an average hypertrophy ofabout 29 per cent, with a maximum of 80 per cent.
The presence of mitotic figures was noted by Kolmer (1912). In the courseof the present investigation these were observed at an early period of pregnancy,mostly at the beginning of Stage IV, while no mitotic figures were visible at Stage V.As stated previously, cells of the zona glomerulosa are most active during the closingperiod of Stage IV.
(5) Medulla.
According to Guieysse there is no hypertrophy in the medulla of the guinea-pigduring pregnancy. Masui and Tamura reported that it atrophied in the mouseat the closing period of pregnancy. In the present investigation a marked increaseof the area of the medulla was observed; this process is slow in certain stages, buttowards the end of pregnancy there is a sudden acceleration so that in certain casesthe medulla occupies as much as 26 per cent, of the total area, thus representingan increase of 46 per cent, over that which obtains at the beginning of pregnancywhen the medulla is but 16 per cent, of the whole area of the gland. A few mitoticfigures were observed during Stage II, and a considerably larger number in Stage IV.This suggests that there occurs a hypertrophy of the medulla during Stages IV and V.
(6) Area.
In a recent paper, Donaldson {1924) comes to the conclusion that "there isno change in the weight of the adrenal of the healthy albino rat during the preg-nancy lactation period," although he observed a slight increase in weight in thefirst quarter, an observation also made by Verdozzi and Castaldi in the case of theguinea-pig. Similarly, Herring (1920) observed in the rat an increase in the weightof the adrenal at the close of pregnancy. It is not reasonable to hold that there mustbe a correlation between the size of the gland and the degree of its functionalactivity, as suggested by Riddle (1923) in the pigeon,.especially as in the case of thesuprarenal gland of the mammal there are several distinct zones.^_There can be no doubt that the zona reticularis during pregnancy in the mouse^Pbmposed of nothing but connective tissue or big vacuoles in which fatty sub-stances are present. Thus the increase in the weight of a gland cannot be consideredas indicative of an increase in its functional activity. A gland may have a very large

90 Y. TAMURA
total area, yet if the area taken up by the degenerated zona reticularis is deducfrom it, it will be seen that the area occupied by functional tissues is actually s ^than in the normal gland.
(7) Functional activity.
Guieysse, who observed vacuoles and siderophile bodies in the adrenal of theguinea-pig, regards them as signs of secretory activity of the gland. He suggeststhat in the guinea-pig the secretory activity is cyclic, increasing till the middle ofpregnancy, and followed by a resting stage until parturition.
But in the present investigation vacuoles are present in great numbers inStage I; in Stage II they are reduced in number and are replaced by the cells ofthe zona gestationis which at this stage show no sign of secretory activity. Asstated above, this process is very gradual. During Stage IV the vacuoles are almostabsent. The cells of the zona gestationis as well as those of the zona glomerulosaexhibit the maximal degree of secretory activity, containing large numbers of sidero-phile bodies (referred to by Masui and Tamura as secretory granules).
Mahnert (1920) assumed that during pregnancy the suprarenal is in an exhaustedcondition. In explanation of this assumption he says, " Geht aus der Lipoidverar-mung dieser Organe bei den Schwangerschaftstoxikosen." On histological ex-amination there can be no doubt that the lipoids are scanty throughout pregnancyexcept at a very early stage, so that the assumption of Mahnert is probably true.If the functional activity during pregnancy is noted, it will be seen to increaseslowly until the third quarter and to decrease during the last quarter.
Watrin (1914, 1919) attempted to discover the cause of the hypertrophy of thesuprarenal in the rabbit and after numerous experiments concluded that hyper-trophy is due to the presence of the fertilised ovum before fixation and of thefoetal placenta. The results of the present investigation are such as tend to corro-borate this suggestion.
Mahnert considers that during pregnancy the suprarenal is functioning abnor-mally and that this disfunctioning (or malfunctioning) of the endocrine organsduring pregnancy is caused by the toxins of the foetal elements of the placenta.Further, he agrees with others that the disfunctioning of the ovary is primary andthat of the adrenal secondary.
Current opinion concerning the relationship of the suprarenal ovary can bestated as follows:
1. The zona reticularis is hyperfunctioning at the period of "heat." (Masuiand Tamura.)
2. During pregnancy, when the ovary and adrenals are in the disfunctioningstage, the zona reticularis degenerates and disappears. (Tamura in this paper.)
3. The zona reticularis reappears when the ovary regains its normal functioning.(Masui and Tamura.) ^ A
4. In the blood of the gonadectomised female there is present an enzyme whichstimulates the development of the ovary. (Long and Evans, and Goto.)

Structural Changes in Suprarenal Gland of Mouse during Pregnancy 91
Complete gonadectomy in the female has no effect on the structure of therenal. (Masui and Tamura.)
The results of the present investigation are such as tend to confirm Mahnert'sfirst suggestion, but in them there is no evidence which permits a decision as towhether the disfunctioning of the suprarenal in relation to that of the ovary isprimary or secondary.
5. SUMMARY.1. In the mouse there is no correlation between body weight, body length,
and the size of the suprarenal gland. The size of this gland is not always indicativeof its functional activity.
2. Unilateral ovariotomy has no effect on the functional activity of the supra-renal gland.
3. During pregnancy the gland becomes reduced in area.4. The hypertrophy of the zona glomerulosa occurs during pregnancy and at
the 17 mm. stage it reaches the maximal secretory activity.5. Observations on the hypertrophy of the zona fasciculata are in agreement
with those of other workers. This hypertrophy may be caused by either an enlarge-ment of cells or an increase of their number. In the latter case it is due to theappearance of a new zone, zona gestationis. This zone reaches its maximal activityat about 17 mm., and at this stage so-called siderophile bodies (secretory granules)are visible in great numbers.
6. Zona reticularis degenerates rapidly at an early period (before formationof placenta) of pregnancy, and at the middle of pregnancy the site of this zone isoccupied by vacuoles.
7. The hypertrophy of the medulla occurs at the closing period of pregnancy.8. Mitotic figures were observed in every zone except zona reticularis at an
early period of pregnancy.9. The secretory activity of this gland increases slowly and reaches its maximum
at about the third quarter of pregnancy, followed by a partial decrease during thelast quarter.
6. REFERENCES.CASTALDI (1922). Arch. d. fisiol. 20, 33. (Quoted after Donaldson.)DONALDSON, J. E. (1924). " The Influence of Pregnancy and Lactation on the Weight of the Adrenal
Gland in the Albino Rat." Amer. Journ. Physiol. 68, 517-522.GOTO, N. (1923). " Studies on the Internal Secretion of Ovary applying to Parabiose in the Albino
Rat, I." Japan. Journ. Gynec. 18, 3.(1923). "Studies on the Internal Secretion of Ovary applying to Parabiose in the Albino
Rat, II ." Ibid.GUIEYSSE, M. A. (1889). "La capsule surrenale chez la femelle du cobaye en gestation." C.R. Soc.
Biol. 51, 898-900.(1901). "La capsule surrenale du cobaye." Journ. Anat. and Physiol. 37, 312-435.
HERRING (1920). "The Effect of Pregnancy on the Various Organs of the White Rat." B.M.J.^ P t 11, 886.
PE, W. (1013). " Veranderungen der Nebenniere bei Schwangerschaft und nach Kastration."Arch.f. Gyn. 99, 272-283.
KOLMER, W. (1912). "Beziehungen von Nebennieren und Geschlechtsfunktion." Pfl. Arch.Physiol. 144.

92 Y. TAMURA
LONG, J. A. and EVANS, H. M. (1922). "The Oestrous Cycle in the Rat and its Associated Pmena." Memo. Univer. Calif. 16.
MAHNERT, A. (1920). "Weitere Beitrage zum Studium der Dysfunction endokriner DriisenSchwangerschaft." Arch.f. Gyn. 113, 472-489.
MASUI, K. and TAMURA, Y. (1924). "The Effect of Gonadectomy on the Structure of the SuprarenalGlands of Mice, with Special Reference to the Functional Relation between this Gland and theSex Gland of the Female." Japan. Journ. Zootech. Sci. 1, 55-79.
RIDDLE, O. (1923). " Studies on the Physiology of Reproduction in Birds. XIV. Suprarenal Hyper-trophy coincident with Ovulation." Amer. Journ. Physiol. 66, 322—339.
VERDOZZI, 0 . (1917). "Capsules surrenales et allaitement." Arch. ital. biol. 66, 121. (Quotedafter Donaldson.)
WATRIN, J. (1914). " L'hypertrophie des capsules surre'nales, au cours de la gestation, est-ellesous la d£pendance du corps jaune?" C.R. Soc. Biol. 77, 142-143.
(1914). "Le corps jaune 'sensibilise' les capsules surrgnales a 1'action des facteurs quide'terminent leur hypertrophie gravidique." Ibid. pp. 207-209.
(1914). "L'ceuf f£cond£ conditionne, avant sa fixation, l'hypertrophie des capsulessurrenales chez la lapine." Ibid. pp. 321—323.
(1919). "L'hypertrophie des capsules surre'nales chez la lapine gestante ne doit pas fitreattribute a la presence du foetus." Ibid, v, 82, 1405-1407.
DESCRIPTION OF PLATES VII AND VIII.
PLATE VII
FIG. 1. Adrenal of the mouse at a very early stage of pregnancy (Stage I).FIGS. 2 and 3. The same (Stage I).FIG. 4. The same (Stage II), showing degenerated zona reticularis.FIG. 5. The same during Stage III.FIG. 6. The same at the beginning of Stage IV.FIG. 7. The same, showing zona gestationis occupying the entire site of zonafasciculata, and vacuoles in zona glomerulosa.FIG. 8. The same, showing large vacuoles in the site of zona reticularis.FIG. 9. The same during Stage V.
PLATE VIII
FIG. 10. Degenerating cells in zona reticularis during Stage I.FIG. 11. Typical vacuolated cells of the zona fasciculata.FIG. 12. Cells of zona gestationis (from Fig. 7).FIG. 13. Cells of zona glomerulosa during Stage IV (from Fig. 7).FIGS. 14 and 15. A cell of zona glomerulosa showing granules (from Fig. 13).FIG. 16. A cell of zona gestationis during Stage III.FIGS. 17, 18 and 19. A cell of zona gestationis during Stage IV.FIGS. 20 and 21. The same during Stage V.
JOURNAL OF EXPERIMENTAL BIOLOGY VOL. IV, PLATE VII.
hm§m^§M
TAMURA—STRUCTURAL CHANGES IN SUPRARENAL GLAND OF MOUSEDURING PREGNANCY (81-92).

JOURNAL OF EXPERIMENTAL BIOLOGY VOL. IV, PLATE VIII.
14
(••
18
15
:*r i%
19
16
20
'm• <
TAMURA—STRUCTURAL CHANGES IN SUPRARENAL GLANDDURING PREGNANCY (pp. 81-92).
21
OF MOUSE






























