Embed Size (px)
Citation preview

A
sttaa©
K
1
ppTOctati[stswtds
0d
J. of Supercritical Fluids 38 (2006) 354–372
Sterilization using high-pressure carbon dioxide
Jian Zhang a, Thomas A. Davis a, Michael A. Matthews a,∗,Michael J. Drews b, Martine LaBerge c, Yuehuei H. An d
a Department of Chemical Engineering, University of South Carolina, Columbia, SC 29208, USAb School of Materials Science and Engineering, Clemson University, Clemson, SC 29634, USA
c Department of Bioengineering, Clemson University, Clemson, SC 29634, USAd Orthopaedic Research Laboratories, Medical University of South Carolina, Charleston, SC 29403, USA
Received 26 January 2005; received in revised form 7 May 2005; accepted 31 May 2005
bstract
Sterility is required for medical devices use in invasive medical procedures, and for some situations in the food industry. Sterilization of heat-ensitive or porous materials or devices, such as endoscopes, porous implants, liquid foodstuff, and liquid medicine, poses a challenge to current
echnologies. There has been a steady interest in using high-pressure carbon dioxide as a process medium for new sterilization technology. Amonghe potential advantages are that CO2 may sterilize at low temperatures. This paper is a review of the technical and patent literature, includingnalysis of the microorganisms studied, important operating parameters, and deactivation mechanisms. The current research status and challengesre summarized at the end of this paper.2006 Elsevier B.V. All rights reserved.
cmalomliot[ts
hb(
eywords: Sterilization; High-pressure carbon dioxide
. Introduction
Sterilization of implants is crucial to prevent infectingatients. In the United States, over 600,000 arthroplasties areerformed each year [1], of which 0.6–2.3% result in infection.his can cause physical injury or even death to the patients [2].ther widely used medical devices, such as endoscopes, can also
ause infection if not properly sterilized between uses. Disinfec-ion of heat-sensitive biomaterials, especially polymers, presentschallenge to current sterilization technology. In medical prac-
ice the standard sterilization methods include steam, gamma-rradiation, ethylene oxide, and hydrogen peroxide sterilization3,4]. Each method has drawbacks in certain applications, asummarized in Table 1. Steam sterilization is the most commonechnique because of its low cost and effectiveness. However,team sterilization operates at 121 ◦C so heat-sensitive materialsill be damaged or destroyed [4]. Additionally, steam steriliza-
ion may deposit an oxide layer onto metallic devices, whichecreases the biocompatibility of the treated implants [5]. Unliketeam sterilization, �-irradiation and ethylene oxide sterilization
∗ Corresponding author. Tel.: +1 803 777 0556; fax: +1 803 777 8265.E-mail address: [email protected] (M.A. Matthews).
bhrrrb(
896-8446/$ – see front matter © 2006 Elsevier B.V. All rights reserved.oi:10.1016/j.supflu.2005.05.005
an be applied to heat-labile materials. However, �-irradiationay cause changes in shear and tensile strength, elastic modulus,
nd transparency of medical polymers [4]. For example, long-ived free radicals generated by �-irradiation cause progressivexidation, breaking of polymer chains, and deterioration ofechanical properties in ultra high molecular weight polyethy-
ene (UHMWPE) [6]. Ethylene oxide is flammable and toxic,s a known carcinogen and can cause hemolysis [4]. Ethylenexide sterilization can also chemically alter material proper-ies, such as the molecular weight of biodegradable polymers4]. Because of limitations with current sterilization techniques,he next generations of polymeric medical devices and heat-ensitive biomaterials demand new sterilization methods [7].
The biocidal effects of high-pressure carbon dioxide (CO2)ave been quantified on various species of bacteria and haveeen summarized elsewhere by Spilimbergo and Bertucco2003) [8]. Using CO2 as a sterilant has several potentialenefits. First, CO2 is not flammable and is non-toxic; the chiefazard in its use is asphyxiation. Unlike ethylene oxide, CO2equires no special handling or ventilation, and leaves no toxic
esidues. Second, CO2 is inert in most situations so it does noteact with polymers, which alleviates the aging problem causedy �-irradiation. Next, CO2 has a low critical temperature31.1 ◦C) [9]. This is only slightly above room temperature,
J. Zhang et al. / J. of Supercritical Fluids 38 (2006) 354–372 355
Table 1Advantages and limitations of sterilization methods
Effects Steam EtO �-Irradiation UV CO2
Sterilization of inside surfaces Yes Yes Yes No YesSSE
sobab[(qFsAeareciwsa
2
cwN9oHarmgs1prt
arapstaT
iseewtttge
rIstait(
3
pn(afW2mhaftsesblpPmt
urface cleaning Nourface contamination, decreased biocompatibility Yesffects on mechanical properties of polymers Yes
o thermal degradation is not a problem when a process isperated around the critical temperature. Because of theseenefits CO2 has been proposed for use in other biomaterialpplications such as incorporating bioactive ingredients intoiodegradable polymers [10] and producing enzyme particles11]. Moreover, in the supercritical state, CO2 has low viscosity3–7 × 10−5 N s m−2) and zero surface tension [9], so it canuickly penetrate complex structures and porous materials.inally, CO2 is inexpensive and readily available, which makeswitching to CO2-based sterilization economically feasible.
preliminary economic analysis has been conducted tostimate the feasibility of this novel technique [3]. Taking intoccount the low operating temperature and absence of toxicesidue, CO2-based sterilization may be superior to steam andthylene oxide sterilization for certain applications, and beompetitive with hydrogen peroxide sterilization. In short, CO2s inexpensive, non-toxic, non-flammable, physiologically safe,ith a low critical temperature, therefore, a high-pressure CO2
terilization technique could be an option for heat-sensitivend/or porous medical devices and biomaterials sterilization.
. Early work
In 1951, Fraser proposed a novel technique to collect theontents of Escherichia coli by bursting cells in liquid cultureith a sudden release of pressurized (1.7–6.2 MPa) gases (Ar,2, N2O, and CO2) [12]. E. coli deactivation rates between5 and 99% were achieved with 3.4 MPa of CO2. This levelf deactivation is acceptable for collecting cell contents.owever, in terms of sterilization, 99% deactivation is only
pproximately a 2-log reduction, far below the >6-log reductionequired for medical sterilization [4]. The concept of treatingicroorganisms with fast depressurization of high-pressure
ases was further examined by Foster et al. in 1962 [13]. Sixpecies of microorganisms were processed with nitrogen at2 MPa in a specially designed apparatus with instantaneousressure release. The experiments achieved up to 58.9% cellupture. Electron microscope images of treated cells were usedo support Fraser’s theory of cell rupture.
In the late 1980’s, strict regulations on ethylene oxidend radiation sterilization in Japan inspired several Japaneseesearchers to explore the use of high-pressure gas treatments an alternative for sterilizing biological products (e.g. plasmaowder) [14] and heat-labile compounds [15], and for pre-
erving foods [16]. These studies focused on CO2 because ofhe benefits mentioned above. Both entrainers (or, co-solvents)nd water content, were examined by Kamihira et al. [15] andaniguchi et al. [14], and were shown to be beneficial for steril-at
[
No No No YesYes No No NoYes Yes No No
zation. Bacterial endospores (Bacillus subtilis and Geobacillustearothermophilus spores) were the test organisms. However,ven after adding 0.5 wt.% acetic acid or 2 wt.% ethanol as anntrainer, only 57 and 38% of G. stearothermophilus sporesere destroyed, respectively [15]. These results demonstrate
hat bacterial spores are highly resistant to high-pressure CO2reatment. Another major contribution from these studies washe postulation of a physiological deactivation mechanism, sug-esting that cell deactivation was due to enzyme deactivation andxtraction of cytoplasmic materials by high-pressure CO2 [15].
In subsequent years more than 50 journal papers and researcheports have been published on high-pressure CO2 sterilization.n those publications, 36 species of microorganisms have beentudied (Tables 3–6) under a wide range of operating condi-ions (Table 7). A number of experimental techniques have beendopted to characterize morphological or structural changesnduced by high-pressure CO2 treatment (Table 10), and newheories of the deactivation mechanisms have been proposedTable 9).
. Gases evaluated for sterilization
Though the majority of publications have focused on high-ressure CO2, some studies exploited other gases, includingitrogen, nitrous oxide, ethylene, argon, and tetrafluoroethaneTFE) (Table 2). Fraser treated E. coli with Ar, N2, N2O, and CO2t 37 ◦C and a pressure between 1.7 and 6.2 MPa for 5 min andound that the other three gases were less effective than CO2 [12].
ei et al. showed that CO2 treatment at 35 ◦C, 5.5–6.2 MPa, forh completely deactivated L. monocytogenes, while N2 treat-ent at the same condition had no effect [17]. Similar results
ave been reported by Nakamura et al. [16], Enomoto et al. [18],nd Dillow et al. [4]. Dillow et al. concluded that using N2 farrom the critical point (Tc = −147 ◦C, Pc = 3.39 MPa) sacrificedhe unique properties (gas-like diffusivity and liquid-like den-ity) of a supercritical fluid. They found CO2 to be much moreffective than N2 at the same conditions of temperature and pres-ure [4]. Not only is the vicinity to a critical point important,ut also the chemical properties of the gas are important. Dil-ow et al. tested sterilization effects of TFE, which has a criticaloint (Tc = 328 K, Pc = 4.06 MPa) similar to CO2 (Tc = 304.13 K,c = 7.38 MPa), but different chemical properties (dipoleoment, DCO2 = 0 D, DTFE = 1.80 ± 0.22 D; solubility parame-
er, δCO2 = 7.0, δTFE = 13.6) [4]. At the same reduced temperature
nd reduced pressure as successful CO2 experiments, no reduc-ion of viable cells was observed after TFE treatment.Another gas having a biocidal effect is N2O. Castor and Hong19] reported that supercritical N2O is more effective than N2

356 J. Zhang et al. / J. of Supercritical Fluids 38 (2006) 354–372
Table 2Properties of gases used for sterilization
Gas Tca (K) Pc
a (MPa) Dipole momenta (D) δ at 25 ◦C (MPa1/2) Solubility in H2O at 25 ◦Cand 1 atm (mol/mol)
Ref.
Carbon Dioxide 304.13 7.375 0 12.3b 6.15 × 10−4a Most of the referencesArgon 150.87 4.898 0 10.9b 2.519 × 10−5a [12,18]Nitrogen 126.21 3.39 0 5.3 b 1.183 × 10−5a [4,13,16–19,28,43,51,53]Nitrous oxide 309.57 7.255 0.16 NA 4.376 × 10−4a [12,18,19]TFE 328c 4.065 1.80 13.6c 2.646 × 10−4d [4]
a CRC Handbook of Chemistry and Physics (84th ed.).nd ed
iTsbE4vTcoda
4t
4
bssaer
bpsw[iImaaottts
4
gr
F
b CRC Handbook of Solubility Parameters and Other Cohesion Parameters (2c Dillow et al. [4].d MSDS of 1,1,1,2-tetrafluoroethane.
n recovering nucleic acids from vegetative bacteria and yeast.his is possibly because the high density and low polarity ofupercritical N2O could favor solubilizing lipids and hydropho-ic compounds in the cell wall and the cytoplasmic membrane.nomoto et al. reported biocidal effects of N2O on yeast cells atMPa, 40 ◦C after 4 h exposure. However, N2O failed to deacti-ate B. megaterium spores at 6 MPa, 60 ◦C even after 24 h [18].he biocidal effect of N2O may be a result of proximity to theritical point and its solubility in water. The critical parametersf N2O are very close to those of CO2; N2O only has a smallipole moment, while CO2 has a zero dipole moment; both havecomparatively high solubility in water.
. Microorganisms used in high-pressure CO2reatment
.1. Overview
The papers reviewed herein cover 12 species of gram-positiveacteria, 10 species of gram-negative bacteria, spores of eightpecies of bacteria, and eight species of fungi (fungus and fungal
pores). Fig. 1 shows the distributions of the number of speciesnd the number of publications in each category. Studies on veg-tative bacteria account for approximately 60% of all the studieseported, in terms of both the number of species and the num-tGpc
ig. 1. Distribution of the number of species (gram-positive bacteria, gram-negative b
.).
er of publications. This is in accordance with the fact that foodoisoning [20] and foodborne diseases [21] are caused by non-porulating vegetative bacteria. The objectives of those studiesere either to preserve foods [22], to improve product quality
22], or to recover bioproducts [23]. Different target microorgan-sms have been studied, depending on the practical application.n the area of food spoilage, pathogenic bacteria such as Listeriaonocytogenes [17,24–27], Staphylococcus aureas [4,15,20,28]
nd Salmonella typhimurium [17,21,29] are of interest. Hong etl. treated Lactobacillus plantarum in order to solve the problemf over-acidification caused by proliferation of L. plantarum inhe late stage of kimchi production [30,31]. Bacterial spores arehe least frequently studied type of organism. Fewer than 20% ofhe studies are dedicated to treatment of spores, possibly becausepore deactivation is not required for food preservation.
.2. Treatment of vegetative bacteria
Traditionally, bacteria are categorized into two majorroups, gram-positive and -negative bacteria, according to theiresponses to the gram stain [32]. The different response to
he gram stain derives from differing peptidoglycan content.ram-positive cell walls are simple in structure, but have thickeptidoglycan layers (10–20 layers thick, as much as 90% of theell wall), which make the cell walls strong and robust. How-acteria, bacterial spores, and fungi) and the number of publications by species.
J. Zhang et al. / J. of Supercritical Fluids 38 (2006) 354–372 357
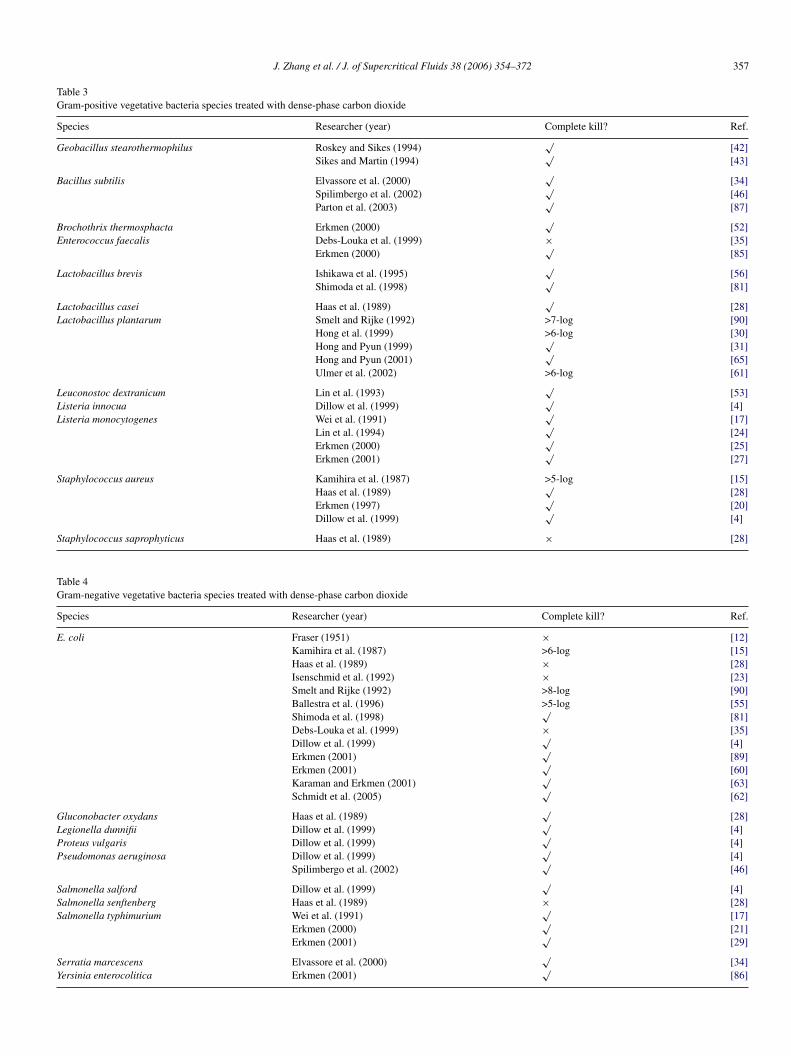
Table 3Gram-positive vegetative bacteria species treated with dense-phase carbon dioxide
Species Researcher (year) Complete kill? Ref.
Geobacillus stearothermophilus Roskey and Sikes (1994)√
[42]Sikes and Martin (1994)
√[43]
Bacillus subtilis Elvassore et al. (2000)√
[34]Spilimbergo et al. (2002)
√[46]
Parton et al. (2003)√
[87]
Brochothrix thermosphacta Erkmen (2000)√
[52]Enterococcus faecalis Debs-Louka et al. (1999) × [35]
Erkmen (2000)√
[85]
Lactobacillus brevis Ishikawa et al. (1995)√
[56]Shimoda et al. (1998)
√[81]
Lactobacillus casei Haas et al. (1989)√
[28]Lactobacillus plantarum Smelt and Rijke (1992) >7-log [90]
Hong et al. (1999) >6-log [30]Hong and Pyun (1999)
√[31]
Hong and Pyun (2001)√
[65]Ulmer et al. (2002) >6-log [61]
Leuconostoc dextranicum Lin et al. (1993)√
[53]Listeria innocua Dillow et al. (1999)
√[4]
Listeria monocytogenes Wei et al. (1991)√
[17]Lin et al. (1994)
√[24]
Erkmen (2000)√
[25]Erkmen (2001)
√[27]
Staphylococcus aureus Kamihira et al. (1987) >5-log [15]Haas et al. (1989)
√[28]
S
TG
S
E
GLPP
SSS
SY
Erkmen (1997)Dillow et al. (1999)
taphylococcus saprophyticus Haas et al. (1989)
able 4ram-negative vegetative bacteria species treated with dense-phase carbon dioxide
pecies Researcher (year)
. coli Fraser (1951)Kamihira et al. (1987)Haas et al. (1989)Isenschmid et al. (1992)Smelt and Rijke (1992)Ballestra et al. (1996)Shimoda et al. (1998)Debs-Louka et al. (1999)Dillow et al. (1999)Erkmen (2001)Erkmen (2001)Karaman and Erkmen (2001)Schmidt et al. (2005)
luconobacter oxydans Haas et al. (1989)egionella dunnifii Dillow et al. (1999)roteus vulgaris Dillow et al. (1999)seudomonas aeruginosa Dillow et al. (1999)
Spilimbergo et al. (2002)
almonella salford Dillow et al. (1999)almonella senftenberg Haas et al. (1989)almonella typhimurium Wei et al. (1991)
Erkmen (2000)Erkmen (2001)
erratia marcescens Elvassore et al. (2000)ersinia enterocolitica Erkmen (2001)
√[20]√[4]
× [28]
Complete kill? Ref.
× [12]>6-log [15]× [28]× [23]>8-log [90]>5-log [55]√
[81]× [35]√
[4]√[89]√[60]√[63]√[62]
√[28]√[4]√[4]√[4]√[46]
√[4]
× [28]√[17]√[21]√[29]
√[34]√[86]

3 ritical Fluids 38 (2006) 354–372
em1ap
sgt1iasttbEic2swsbCgbLrBrS2
phvnsfb5t
bstctfm
4
oo
acCo1carogosc
rossoatw
h1ot[p
Vt[
58 J. Zhang et al. / J. of Superc
ver, gram-negative cells have complex cell wall structures butuch thinner peptidoglycan layers (only 1–2 layers thick, about
0% of the cell wall) [32,33]. Therefore, the gram-positive cellsre stronger, less likely to be broken mechanically, and are lessermeable than the gram-negative cells [34].
Both gram-positive and -negative bacteria have beenubjected to CO2 treatment (Tables 3 and 4). Generally,ram-positive bacteria have been more difficult to deactivatehan gram-negative bacteria. In a semi-continuous apparatus,07 colony forming unit (cfu) per ml of gram-positive B. subtilisn phosphate buffered saline (PBS) was completely deactivatedfter 2.5 min exposure under 7.4 MPa CO2 at 38 ◦C, while theame concentration of gram-negative S. marcescens was deac-ivated at 0 min (the time when the apparatus is pressurized tohe desired pressure) under the same conditions [34]. The sameehavior was seen by Dillow et al. [4] and Debs-Louka et al. [35]lectron microscope images provide more information regard-
ng the relationship between the robustness of gram-positiveell walls and reduced cell wall damage. With CO2 treatment at5 ◦C, 20.5 MPa for 1 h, SEM images of E. coli (gram-negative),howed partially damaged cell walls and some ruptured cells,hile the images of S. aureus (gram-positive) bacterium,
howed no cell wall damage [4]. However, gram-negativeacteria are not always more susceptible to high-pressureO2 than gram-positive bacteria. Dillow et al. examined tworam-positive (L. innocua, S. aureus) and five gram-negativeacteria species (S. salford, P. aeruginosa, E. coli, P. vulgaris,. dunnifii). Generally, the gram-positive species showedesistance higher, or at least equal to, the gram-negative species.ut S. salford, a gram-negative bacterium, showed only 3-log
eduction, while the two gram-positive bacteria, L. innocua and. aureus were reduced by 9- and 7-log, respectively, at 34 ◦C,0.5 MPa, 0.6 h, with six pressure cycles [4].
Although there is a difference in sensitivity between the gram-ositive and -negative vegetative bacteria, both are susceptible toigh-pressure CO2 treatment. Twenty out of twenty-two testedegetative species were completely deactivated at some combi-ation of temperature, pressure, time, etc. (Tables 3 and 4). Thepecies that were not completely deactivated, Salmonella sen-tenberg and Staphylococcus saprophyticus, have been studiedy only one group [28], and the pressures used were only 6.2 and.5 MPa. It is possible that the pressures in these studies wereoo low to give complete deactivation.
In conclusion, high-pressure CO2 treatment of vegetativeacteria has been largely successful, with over 90% of the testedpecies completely deactivated. However, success with vegeta-ive cells does not guarantee that high-pressure CO2 treatmentan be used for sterilization. Sterilization is defined as deactiva-ion of ALL living microorganisms including the most resistantorm of bacteria — endospores [36]. At least 106 cfu/ml sporesust be completely deactivated in order to claim sterilization [4].
.3. Treatment of endospores
A spore (or endospore) is the highly resistant dormant formf various bacilli and clostridia. Sporulation of vegetative cellsccurs under harsh environments such as poor nutrition. Spores
avt[
Fig. 2. Major structures of a B. subtilis spore (Driks [37]).
re highly resistant to heat, UV radiation, free radicals, andhemicals because of their unique structures (Fig. 2) [32,37].ompared to a vegetative cell which contains on the orderf 80–90% water, the spore core is highly dehydrated (only0–25% water content), making it very resistant to heat andhemicals [32]. The Ca2+ dipicolinic acid complex and smallcid-soluble protein (SASP), which bind to DNA, increase sporeesistance to heat, desiccation, and UV radiation [32,38,82]. Theutside of the spore core is a thick, loosely cross-linked peptido-lycan layer called the spore cortex, which prevents hydrationf the spore core [32]. The outmost structure is the multilayeredpore coat, which is a permeability barrier to chemicals such ashloroform and lysozyme [37].
Because spores are highly resistant to heat, chemicals andadiation, extreme temperatures (121 ◦C steam), UV radiation,r highly oxidative chemicals, e.g. ethylene oxide, are used forterilization. Spore survivability is the standard assay to testterilization equipment [36]. The most frequently used modelrganisms are G. stearothermophilus, which is used to test steamnd hydrogen peroxide sterilizers, B. atrophaeus, which is usedo test dry heat and ethylene oxide sterilizers and B. pumilus,hich is used to test radiation sterilizers.Spores have not been studied extensively in the presence of
igh-pressure CO2. Only eight species have been investigated in4 publications (Table 5). Most of the studies on spores reportnly experimental data and do not address mechanistic ques-ions [4,15,39,40–43]. Only Enomoto et al. [18], Ballestra et al.44], Watanabe et al. [45], and Spilimbergo et al. [46,47] discussossible deactivation mechanisms.
Spores are highly resistant to high-pressure CO2 treatment.egetative G. stearothermophilus cells were reduced by more
han 6-log after 1.5-h exposure to CO2 at 2.75 MPa and 25 ◦C42]. However, even with 2-h exposure to pure CO2 at 20 MPand 35 ◦C, 80% of G. stearothermophilus spores remained
iable. Even with the addition of ethanol or acetic acid, lesshan 60% of the G. stearothermophilus spores were deactivated15].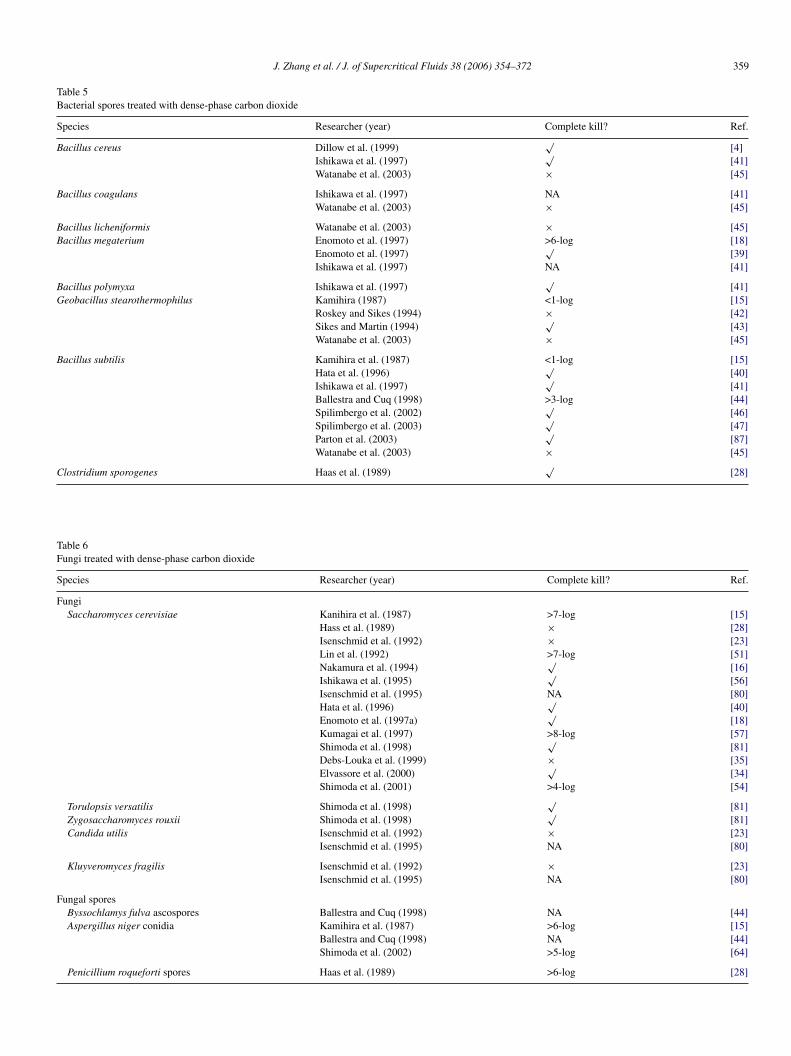
J. Zhang et al. / J. of Supercritical Fluids 38 (2006) 354–372 359
Table 5Bacterial spores treated with dense-phase carbon dioxide
Species Researcher (year) Complete kill? Ref.
Bacillus cereus Dillow et al. (1999)√
[4]Ishikawa et al. (1997)
√[41]
Watanabe et al. (2003) × [45]
Bacillus coagulans Ishikawa et al. (1997) NA [41]Watanabe et al. (2003) × [45]
Bacillus licheniformis Watanabe et al. (2003) × [45]Bacillus megaterium Enomoto et al. (1997) >6-log [18]
Enomoto et al. (1997)√
[39]Ishikawa et al. (1997) NA [41]
Bacillus polymyxa Ishikawa et al. (1997)√
[41]Geobacillus stearothermophilus Kamihira (1987) <1-log [15]
Roskey and Sikes (1994) × [42]Sikes and Martin (1994)
√[43]
Watanabe et al. (2003) × [45]
Bacillus subtilis Kamihira et al. (1987) <1-log [15]Hata et al. (1996)
√[40]
Ishikawa et al. (1997)√
[41]Ballestra and Cuq (1998) >3-log [44]Spilimbergo et al. (2002)
√[46]
Spilimbergo et al. (2003)√
[47]Parton et al. (2003)
√[87]
Watanabe et al. (2003) × [45]
C
TF
S
F
F
lostridium sporogenes Haas et al. (1989)
able 6ungi treated with dense-phase carbon dioxide
pecies Researcher (year)
ungiSaccharomyces cerevisiae Kanihira et al. (1987)
Hass et al. (1989)Isenschmid et al. (1992Lin et al. (1992)Nakamura et al. (1994)Ishikawa et al. (1995)Isenschmid et al. (1995Hata et al. (1996)Enomoto et al. (1997a)Kumagai et al. (1997)Shimoda et al. (1998)Debs-Louka et al. (199Elvassore et al. (2000)Shimoda et al. (2001)
Torulopsis versatilis Shimoda et al. (1998)Zygosaccharomyces rouxii Shimoda et al. (1998)Candida utilis Isenschmid et al. (1992
Isenschmid et al. (1995
Kluyveromyces fragilis Isenschmid et al. (1992Isenschmid et al. (1995
ungal sporesByssochlamys fulva ascospores Ballestra and Cuq (199Aspergillus niger conidia Kamihira et al. (1987)
Ballestra and Cuq (199Shimoda et al. (2002)
Penicillium roqueforti spores Haas et al. (1989)
√[28]
Complete kill? Ref.
>7-log [15]× [28]
) × [23]>7-log [51]√
[16]√[56]
) NA [80]√[40]√[18]
>8-log [57]√[81]
9) × [35]√[34]
>4-log [54]√
[81]√[81]
) × [23]) NA [80]
) × [23]) NA [80]
8) NA [44]>6-log [15]
8) NA [44]>5-log [64]
>6-log [28]

3 ritica
epa5mSsfr[3Ht
r9[spfoapaoh
tautoAdma�
ta[
4
Ctr
5
cWCMs
cT
5
aatcaft1htheo
iebdttdipmH3pt3
tesso[oste
gdpnso
60 J. Zhang et al. / J. of Superc
To achieve greater deactivation, several approaches have beenmployed, including increasing treatment time, raising tem-erature, and using pressure cycling. Enomoto et al. achievedpproximately 7-log reduction of B. megaterium spore with a0-h treatment at 7.8 MPa and 60 ◦C [39]. Clearly, a 50-h treat-ent would be problematic for a practical sterilization process.pilimbergo et al. reported that only a 0.9-log reduction of B.ubtilis spore can be achieved with a treatment at 12 MPa, 54 ◦C,or 24 h [46]. However, at 75 ◦C and 7 MPa, greater than 7-logeduction of B. subtilis spores was observed after a 2-h treatment47]. Ballestra et al. observed a biocidal effect of approximately.5-log reduction at 5 MPa for 1 h at temperatures at 80 ◦C [44].ata et al. reported a 6-log reduction of B. subtilis spore with
reatment at 70 ◦C, 20 MPa for 10 h.By increasing temperature and/or the pressure, less time is
equired to achieve a 6-log reduction. Only 2 h were needed at0 ◦C, 6 MPa to achieve a 6-log reduction of B. subtilis spores40]. The lowest temperature with significant deactivation of B.ubtilis spores was reported by Ishikawa et al. [41]. They accom-lished a 6-log reduction of B. subtilis spores at 55 ◦C, 30 MPaor 60 min using their micro-bubble method, in which numer-us CO2 micro-bubbles were formed by feeding CO2 throughstainless steel filter (10 �m pore size) from the bottom of a
ressure chamber. Because these experiments were conductedt moderate to high temperatures, these authors concluded thatnly with a combination of high-pressure CO2 and at least mildeat can spores be deactivated [46].
Long treatment time and high temperatures are two poten-ial problems of the CO2 sterilization technique. Even thoughhigh degree of deactivation of spores has been realized, this
sually requires more than 10 h, which is not competitive withhe average time of 10–15 min for steam sterilization. On thether hand, ethylene oxide processes require a 15-h cycle [3].dditionally, the high temperatures used (55–90 ◦C) can easilyamage heat-sensitive materials. Pressure cycling is a promisingethod to enhance deactivation while lowering the temperature
nd time requirements. With pressure cycling of 30 cycles/h,P = 8 MPa, at 36 ◦C for 30 min, a 3.5-log reduction of B. sub-
ilis spores was achieved. Without pressure cycling, a treatmentt 36 ◦C, 7.5 MPa for 24 h only resulted in 0.5-log reduction46].
.4. Treatment of fungi
A few investigators have studied the effect of high-pressureO2 on fungi and fungal spores. These microorganisms are not
he focus of this review, but for completeness, Table 6 shows theeferences for these studies.
. Experimental parameters
Bacterial cells and spores are complex chemical systemsomposed of various kinds of organic and inorganic components.
hen one also considers the variety of growth media, high-phaseO2 processing becomes a very complicated process indeed.any factors have been studied, including temperature, pres-
ure, depressurization rate, pressure cycling, treatment time, cell
slcl
l Fluids 38 (2006) 354–372
oncentration, cell growth phase, agitation, media, and entrainer.he influence of these factors will be reviewed below.
.1. Effects of temperature, pressure, and state
Temperature and pressure are the most important factorsffecting growth of microorganisms. Each microorganism hasspecies-specific maximum temperature. Above that tempera-
ure, proteins denature, cytoplasmic membranes collapse, andells lyses and are deactivated [32]. A wide range of temper-tures has been employed for high-pressure CO2 treatment,rom 0 ◦C [16] to 100 ◦C [42,43]. Bacteria are more resistanto pressure than to temperature. A hydrostatic pressure between00 and 1000 MPa is required to deactivate bacteria [48]. Highydrostatic pressure processes have been reviewed by the Insti-ute of Food Technologists [49] and Cheftel [48]. However, ifigh-pressure CO2 is used, the pressure requirement can be low-red below 20 MPa (Table 7). The highest pressure reported isnly 33 MPa [50].
Generally, deactivation is more pronounced with increas-ng temperature [21,51]. It is believed that higher temperaturenhances deactivation by (a) increasing the fluidity of cell mem-ranes, making them easier to penetrate, and (b) increasing theiffusivity of CO2 [22]. Therefore, higher temperatures reducehe duration of the first stage of deactivation [21], which ishought to be diffusion-controlled (see Section 5.6 for a detailediscussing of two-stage kinetics). Higher temperatures alsoncrease the rate in the second stage [52]. However, higher tem-eratures may reduce the ability of CO2 to extract low-volatilityaterials and decrease CO2 solubility in aqueous media [53].ong and Pyun reported that deactivation of L. plantarum at0 ◦C, 7 MPa was better than that at 40 ◦C, 7 MPa [31]. Thisrobably is the result of higher density at 30 ◦C (0.27 g/ml) thanhat at 40 ◦C (0.20 g/ml); higher CO2 solubility in the media at0 ◦C than that at 40 ◦C.
High pressure facilitates solubilization in water and pene-ration through cell walls, and increases density and thereforextraction power [24]. All these factors are thought to inten-ify the deactivation process. Experiments by Debs-Louka et al.howed a pressure threshold below which no deactivation wasbserved. This pressure threshold varies with bacterial species35]. The D-value (the time needed to achieve 1-log reduction)f S. cerevisiae showed a steep decrease with increase in pres-ure from 4 to 10 MPa [54]. The duration of the earlier stage andhe inactivation rate of the second stage have been found to bextremely sensitive to pressure [52,55].
Depending on temperature and pressure, CO2 exists in theas, liquid or supercritical fluid state. Physical properties such asensity, diffusivity, solubility in aqueous solution, and extractionower vary dramatically around the critical point [9]. Unfortu-ately, even though several studies have covered two or threetates (Table 7), only a few authors discussed the physical statef CO2 and its possible effect on sterilization [15,53,56]. The
upercritical state is characterized by gas-like diffusivity andiquid-like density. The gas-like diffusivity allows supercriti-al CO2 to quickly diffuse through complex matrices; and theiquid-like density confers high extraction power [9]. Because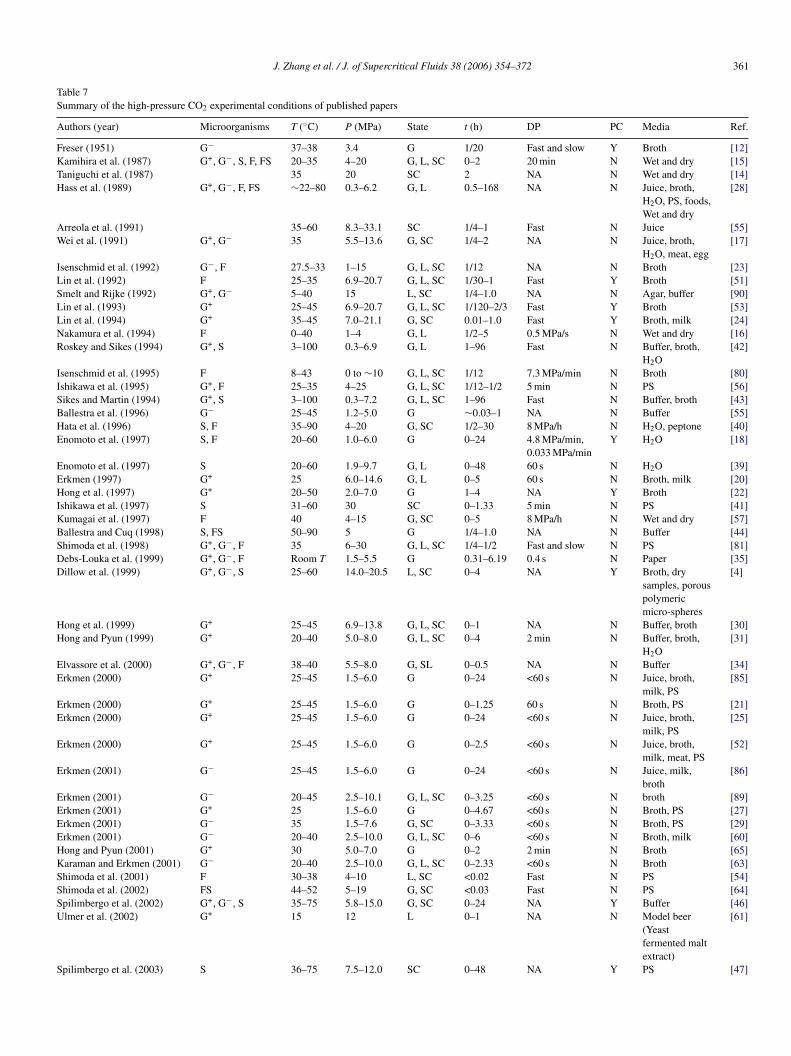
J. Zhang et al. / J. of Supercritical Fluids 38 (2006) 354–372 361
Table 7Summary of the high-pressure CO2 experimental conditions of published papers
Authors (year) Microorganisms T (◦C) P (MPa) State t (h) DP PC Media Ref.
Freser (1951) G− 37–38 3.4 G 1/20 Fast and slow Y Broth [12]Kamihira et al. (1987) G+, G−, S, F, FS 20–35 4–20 G, L, SC 0–2 20 min N Wet and dry [15]Taniguchi et al. (1987) 35 20 SC 2 NA N Wet and dry [14]Hass et al. (1989) G+, G−, F, FS ∼22–80 0.3–6.2 G, L 0.5–168 NA N Juice, broth,
H2O, PS, foods,Wet and dry
[28]
Arreola et al. (1991) 35–60 8.3–33.1 SC 1/4–1 Fast N Juice [55]Wei et al. (1991) G+, G− 35 5.5–13.6 G, SC 1/4–2 NA N Juice, broth,
H2O, meat, egg[17]
Isenschmid et al. (1992) G−, F 27.5–33 1–15 G, L, SC 1/12 NA N Broth [23]Lin et al. (1992) F 25–35 6.9–20.7 G, L, SC 1/30–1 Fast Y Broth [51]Smelt and Rijke (1992) G+, G− 5–40 15 L, SC 1/4–1.0 NA N Agar, buffer [90]Lin et al. (1993) G+ 25–45 6.9–20.7 G, L, SC 1/120–2/3 Fast Y Broth [53]Lin et al. (1994) G+ 35–45 7.0–21.1 G, SC 0.01–1.0 Fast Y Broth, milk [24]Nakamura et al. (1994) F 0–40 1–4 G, L 1/2–5 0.5 MPa/s N Wet and dry [16]Roskey and Sikes (1994) G+, S 3–100 0.3–6.9 G, L 1–96 Fast N Buffer, broth,
H2O[42]
Isenschmid et al. (1995) F 8–43 0 to ∼10 G, L, SC 1/12 7.3 MPa/min N Broth [80]Ishikawa et al. (1995) G+, F 25–35 4–25 G, L, SC 1/12–1/2 5 min N PS [56]Sikes and Martin (1994) G+, S 3–100 0.3–7.2 G, L, SC 1–96 Fast N Buffer, broth [43]Ballestra et al. (1996) G− 25–45 1.2–5.0 G ∼0.03–1 NA N Buffer [55]Hata et al. (1996) S, F 35–90 4–20 G, SC 1/2–30 8 MPa/h N H2O, peptone [40]Enomoto et al. (1997) S, F 20–60 1.0–6.0 G 0–24 4.8 MPa/min,
0.033 MPa/minY H2O [18]
Enomoto et al. (1997) S 20–60 1.9–9.7 G, L 0–48 60 s N H2O [39]Erkmen (1997) G+ 25 6.0–14.6 G, L 0–5 60 s N Broth, milk [20]Hong et al. (1997) G+ 20–50 2.0–7.0 G 1–4 NA Y Broth [22]Ishikawa et al. (1997) S 31–60 30 SC 0–1.33 5 min N PS [41]Kumagai et al. (1997) F 40 4–15 G, SC 0–5 8 MPa/h N Wet and dry [57]Ballestra and Cuq (1998) S, FS 50–90 5 G 1/4–1.0 NA N Buffer [44]Shimoda et al. (1998) G+, G−, F 35 6–30 G, L, SC 1/4–1/2 Fast and slow N PS [81]Debs-Louka et al. (1999) G+, G−, F Room T 1.5–5.5 G 0.31–6.19 0.4 s N Paper [35]Dillow et al. (1999) G+, G−, S 25–60 14.0–20.5 L, SC 0–4 NA Y Broth, dry
samples, porouspolymericmicro-spheres
[4]
Hong et al. (1999) G+ 25–45 6.9–13.8 G, L, SC 0–1 NA N Buffer, broth [30]Hong and Pyun (1999) G+ 20–40 5.0–8.0 G, L, SC 0–4 2 min N Buffer, broth,
H2O[31]
Elvassore et al. (2000) G+, G−, F 38–40 5.5–8.0 G, SL 0–0.5 NA N Buffer [34]Erkmen (2000) G+ 25–45 1.5–6.0 G 0–24 <60 s N Juice, broth,
milk, PS[85]
Erkmen (2000) G+ 25–45 1.5–6.0 G 0–1.25 60 s N Broth, PS [21]Erkmen (2000) G+ 25–45 1.5–6.0 G 0–24 <60 s N Juice, broth,
milk, PS[25]
Erkmen (2000) G+ 25–45 1.5–6.0 G 0–2.5 <60 s N Juice, broth,milk, meat, PS
[52]
Erkmen (2001) G− 25–45 1.5–6.0 G 0–24 <60 s N Juice, milk,broth
[86]
Erkmen (2001) G− 20–45 2.5–10.1 G, L, SC 0–3.25 <60 s N broth [89]Erkmen (2001) G+ 25 1.5–6.0 G 0–4.67 <60 s N Broth, PS [27]Erkmen (2001) G− 35 1.5–7.6 G, SC 0–3.33 <60 s N Broth, PS [29]Erkmen (2001) G− 20–40 2.5–10.0 G, L, SC 0–6 <60 s N Broth, milk [60]Hong and Pyun (2001) G+ 30 5.0–7.0 G 0–2 2 min N Broth [65]Karaman and Erkmen (2001) G− 20–40 2.5–10.0 G, L, SC 0–2.33 <60 s N Broth [63]Shimoda et al. (2001) F 30–38 4–10 L, SC <0.02 Fast N PS [54]Shimoda et al. (2002) FS 44–52 5–19 G, SC <0.03 Fast N PS [64]Spilimbergo et al. (2002) G+, G−, S 35–75 5.8–15.0 G, SC 0–24 NA Y Buffer [46]Ulmer et al. (2002) G+ 15 12 L 0–1 NA N Model beer
(Yeastfermented maltextract)
[61]
Spilimbergo et al. (2003) S 36–75 7.5–12.0 SC 0–48 NA Y PS [47]

362 J. Zhang et al. / J. of Supercritical Fluids 38 (2006) 354–372
Table 7 (Continued )
Authors (year) Microorganisms T (◦C) P (MPa) State t (h) DP PC Media Ref.
Parton et al. (2003) G+, S 40–60 8.9–10.0 SC 0–6.5 NA N Juice [87]Watanabe et al. (2003) S 35–95 30.0 SC 0.5–2.0 2 min N H2O [45]S L
G ; DP:
tbccItagrsfnp[tpmbt
5
istwcmifAbresidctnctdK
bss
cop
tscei
5
oa[ac[pDw2ts
rsp[c2
tfbpsapttb
H
chmidt et al. (2005) G− 20 7.0
: gram-positive and -negative bacteria; F: fungus; FS: fungal spores; S: spores
he penetration of CO2 through the cell envelope is believed toe the controlling step in deactivation, and the extraction of cellontents is a possible deactivation mechanism [51], supercriti-al CO2 may be superior to both gaseous and liquid CO2 [53].shikawa et al. [56] and Kamihira et al. [15] compared steriliza-ion with gaseous, liquid and supercritical CO2 and showed thatt the same density, the supercritical state is superior to bothas and liquid. Liquid CO2 with density of 0.9 g/ml at 25 ◦Ceduced L. brevis by ∼2.7-log; while supercritical CO2 with theame density at 35 ◦C reduced L. brevis by 6-log. Hata et al.ound that the sterilization rate constant k increases infinitelyear the critical pressure [40]. Dillow et al. believed that theroximity to the critical point determines the deactivation effect4]. However, in experiments by Enomoto et al. on B. mega-erium spores at 60 ◦C, 24 h treatment, they found an optimumressure in the sub-critical region at 6 MPa, which gave a maxi-um degree of deactivation. The reason for this was thought to
e the aggregation of spores under high pressure, which madehem more resistant to high-pressure CO2 [39].
.2. Effect of depressurization rate
The effect of depressurization rate is frequently addressedn the literature on high-pressure CO2 treatment. A fast depres-urization rate is believed to burst cells and/or enhance massransfer across cell membranes. Fraser proposed that the cellsere mechanically ruptured by the fast expansion of CO2 within
ells during flash discharge of pressure [12]. The cell ruptureechanism is also supported by Nakamura et al. with their exper-
ments on yeasts [16], which always gave higher deactivation ataster depressurization rates than at lower depressurization rates.lso, SEM images of treated cells showed deformed and evenroken cells. However, Nakamura et al. proposed that the cellupture might not be the only cause of deactivation. The earli-st papers either used flash (rapid) depressurization or did notpecify the depressurization rate [13–15,28]. This became anssue when Arreola et al. questioned the effectiveness of flashischarge [50]. Arreola et al. recovered large numbers of intactells from rapidly depressurized samples in the early stage ofheir experiments. However, experiments by Arreola et al. areot convincing because CO2 might have not accumulated suffi-iently in the cells due to short exposure. To identify the effect ofhe depressurization rate, other researchers employed rather slowepressurization rate, (e.g. 8 MPa/h in experiment conducted byumagai et al. [57]) and still achieved high deactivation.
Carbon dioxide penetration through the cell wall is thought toe the rate-controlling step. If the mass transfer rate is improved,o is the deactivation effect [22]. Fast pressurization and depres-urization are believed to enhance the transfer of CO2 into the
Ladt
0.25 NA N cotton [62]
depressurization rate; PC: pressure cycling.
ells and extraction of materials from the cells, so Lin et al. rec-mmended using pressure cycling to enhance the deactivationrocess [53,58].
Enomoto et al. observed nearly the same amount of pro-ein release from treatments with either explosive decompres-ion (50 s) or slow decompression (1.5–2 h), which neither theell rupture theory nor the enhanced mass transfer theory canxplain. Therefore, Enomoto et al. postulate that the depressur-zation rate is not essential in cell deactivation [18].
.3. Effect of pressure cycling
Pressure cycling involves repeated release and compressionf CO2. There are two theories of how a pressure cycling oper-tion can enhance deactivation: an enhanced cell rupture theory12] and an enhanced mass transfer theory [4,22,30,31]. Fraserchieved more than 90% cell rupture only after two pressureycles, at least a 15% increase from a single pressurization step12]. Lin et al. claimed a higher degree of deactivation using aressure cycling method, but no data were presented [24,53].illow et al. also observed significant increase in log-reductionith pressure cycling. For example, with a treatment at 34 ◦C,0.5 MPa and total treatment time 0.6 h, the degree of inactiva-ion jumped from 3-log with three pressure cycles to 9-log withix pressure cycles [4].
Pressure cycling also enhanced the deactivation of bacte-ial spores. Spilimbergo et al. achieved 3.5-log reduction of B.ubtilis spores by cycling pressure 30 times in 30 min, over aressure range from 8 MPa to atmospheric pressure at 36 ◦C46]. At 60 ◦C, Dillow et al. successfully deactivated 8-log B.ereus spores with six pressure cycles during a 4-h operation at0.5 MPa [4].
The enhanced deactivation of bacterial spores may not only behe result of an enhanced rupture rate or an enhanced mass trans-er rate, but also could be the result of spore germination inducedy high-pressure [46,47]. It is known that very high hydrostaticressure (100–600 MPa) can germinate bacterial spores (e.g. B.ubtilis). The mechanism is not clear, but may be a result ofctivating germinant receptors and/or cutting short germinationathways [59]. It is speculated that cycling the pressure severalimes will increase the efficiency of pressure-induced germina-ion [46,47]. However, whether CO2 at 30 MPa can germinateacterial spores is still unknown.
The literature on pressure cycling with CO2 is not unanimous.ong et al. cycled CO2 pressure above MRS broth suspension of
actobacillus sp., but observed the same amount of deactivations with static CO2 pressure [22]. Enomoto et al. observed lowereactivation with pressure cycling. With the same total treatmentime of 4 h, the log reduction of baker’s yeast decreased from
ritica
7c
5
ibvaarb[fd
Luaagaat6roaowdo[avfa
dbHitCmc
EdpltsbiK
we[c3wa
5
sbhSfl
if<st(ce[
ltssc3
5ata
5
(atto
t(vat
J. Zhang et al. / J. of Superc
-log without pressure cycling to 4-log with a frequency of 1ycle/h [18].
.4. Growth medium
The majority of studies have used aqueous cell suspensions,ncluding distilled water [17], physiological saline [29], PBSuffer solution [34], milk [60], fruit juices [50], beer [61], andarious kinds of culture broth [31]. Non-aqueous media havelso been studied, including meat [52], egg [17], cotton [62],nd filter paper [35]. Another method of controlling the envi-onment around the microorganism is to control water contenty drying wet cells [15] or re-hydrating dry material with water14] (Table 7). The medium used affects factors such as acidity,at/oil content and water content, all of which may influence theeactivation process.
Acidity may or may not promote deactivation of bacteria.actobacillus, for example, is acid tolerant [22]. Such bacteriase homeostasis to keep a pH difference between the cytoplasmnd the medium, so they can survive in acidic conditions. Hongnd Pyun observed little deactivation of L. plantarum with nitro-en at 7 MPa and 30 ◦C over a 0.1 M citrate buffer suspensiont pH 3.5 [31]. However, more than 7-log reduction has beenchieved by high-pressure CO2 treatment at the same tempera-ure and pressure over a spore suspension in distilled water at pH.0. The contrast between the ineffectiveness of an acidic envi-onment and nitrogen with the effectiveness of CO2 treatmentf an un-acidified medium indicates that the internal pH drop isfactor in cell deactivation. Low internal pH can disturb home-stasis and proton force [31], and can precipitate the enzymesith acidic isoelectric points [55]. Even though acidity aloneoes not deactivate some microorganisms, low pH does havether effects on cells, such as weakening spore resistance to heat42] and attenuating resistance to destruction [22]. Therefore,cidity can act synergistically with high-pressure CO2 in deacti-ation [28]. For example, Gluconobacter oxydans was reducedrom 3 × 105 cfu/ml to 0, 25, 270, and 270 cfu/ml at pH 2, 3, 4,nd 5, respectively, with a treatment at 5.5 MPa, 22 ◦C for 2 h.
Media containing fat or oil have been shown to protect againstense-phase CO2 treatment. For example, L. monocytogenes inroth was completely deactivated at 13.7 MPa, 35 ◦C over 2 h.owever, under exactly the same conditions, L. monocytogenes
noculated in whole egg increased two-fold [17]. It is believedhat fat or oil contents increase cell resistance to dense-phaseO2 treatment by changing the structures of the cell wall andembranes [20,24,31], or by depressing CO2 penetration into
ells [52].Water is regarded as necessary to the biocidal effect of CO2.
xperiments on plasma powders of different water content [14],ry and wet cells (70–90% water content) [15,16,57], and filteraper [35] have proven that cells with low water content showedow or no deactivation. For example, yeast cells with morehan 70% water content were completely deactivated, while the
amples with less than 40% water content were only reducedy ∼0.5-log (40 ◦C, 4 MPa, 5 h) [16]. Water swells the cells,ncreases the permeability of cell walls, and hydrates CO2 [4,53].umagai et al. noted the high correlation between unfreezablesm[r
l Fluids 38 (2006) 354–372 363
ater and deactivation, and proposed that it is not the water innvironment, but water absorbed by the cell that is important57]. However, there is one exception. Dillow et al. achievedomplete inactivation of both dry and wet E. coli at 14 MPa and4 ◦C after 30 and 60 min, respectively. The kinetic curves ofet cells and dry cells were different, which probably indicatesdifferent deactivation mechanism for wet and dry cells [4].
.5. Additives
Additives (co-solvents) have been studied extensively inupercritical fluid extraction and other operations, but have noteen widely used in high-pressure CO2 sterilization. Only Kami-ira et al. [15], Taniguchi et al. [14], Sikes and Martin [43], andchmidt et al. [62] have reported on the use of additives in theuid phase.
Kamihira et al. used 2 wt.% ethanol or 0.5 wt.% acetic acidn supercritical CO2. These additives in CO2 at 35 ◦C, 20 MPa,or 2-h, increased the deactivation of dry A. niger conidia from1-log (pure CO2) to >6-log. However, the deactivation of G.tearothermophilus spores increased only from 20 to ∼50% athe same conditions [15]. Taniguchi et al. also used ethanol1.9–4.5 wt.%) and acetic acid (0.41–0.72 wt.%) with super-ritical CO2 to process plasma powder and observed similarnhancements due to acetic acid, but ethanol was not effective14].
Sikes and Martin examined the effectiveness of egg whiteysozyme, an enzyme that hydrolyzes the peptidoglycan struc-ure which is the main component of the bacterial cell wall andpore cortex. With 300 ppm egg-white lysozyme added to a G.tearothermophilus spore suspension (not added in CO2 phase),omplete deactivation of spores was achieved within 2 h and7 ◦C with only 1.37 MPa CO2 overpressure [43].
Schmidt et al. disinfected E. coli on cotton by mixing eithervol.% 2-propanol or commercial disinfectants with CO2 in anutoclave. E. coli was completely killed by two of the formula-ions, 2-propanol or Irgasan DP 300 (active ingredients Triclosannd 2′,4,4′-trichloro-2-hydroxy diphenylether) [62].
.6. Treatment time and deactivation kinetics
Treatment times in the literature vary from a few minutese.g. 5 min [23]) to several days (e.g. 100 h [43]). One wouldnticipate that deactivation increases with treatment time, buthe specific time required to complete deactivation depends onhe category of the microorganism (bacteria, fungus), the formf microorganism (vegetative, spore) and treatment conditions.
Lin et al. measured the ratio of living to dead cells as a func-ion of treatment time and observed a two-stage kinetic curveTable 9) [51]. The first stage was characterized by a slow deacti-ation rate, and the second stage by fast linear deactivation. Lin etl. believed that the first stage is the slow penetration of CO2 intohe cell walls, and is the controlling step of deactivation. In the
econd stage, CO2 extracts vital components from cytoplasm orembranes and thus “lethally disturbs or unbalances” the cells51]. Two-stage kinetics were also observed by several otheresearchers (Fig. 3(a) and (b)) [20–22,25,44,55]. Some studies
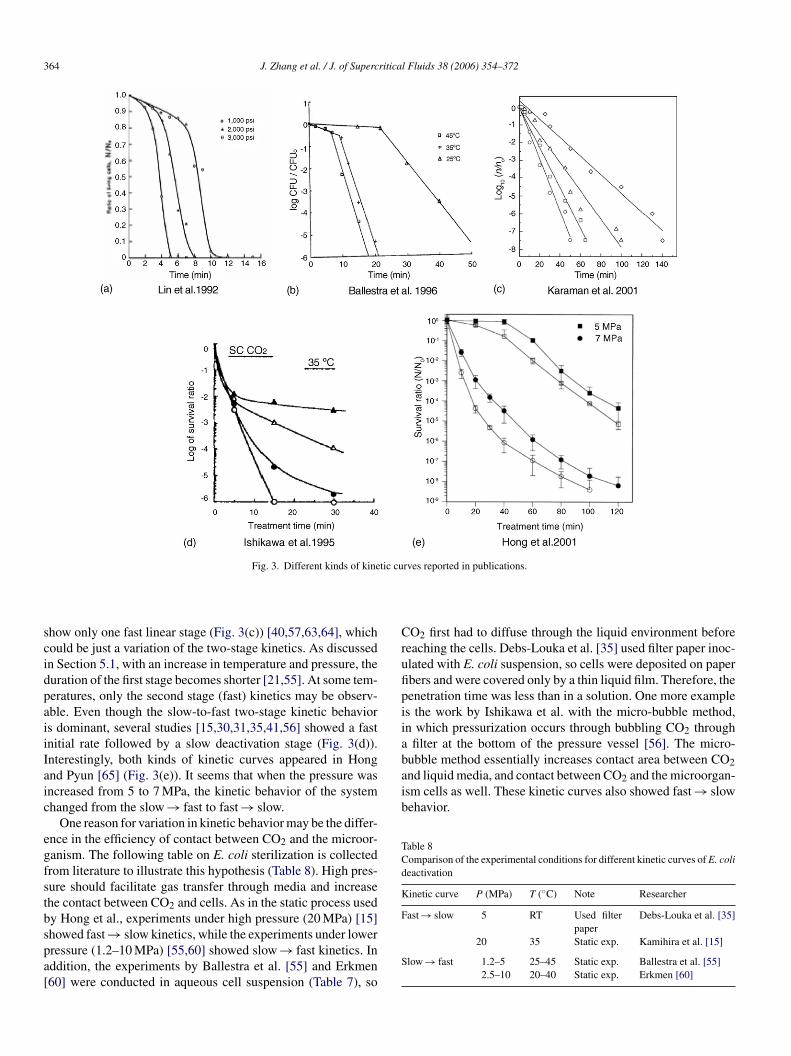
364 J. Zhang et al. / J. of Supercritical Fluids 38 (2006) 354–372
tic cu
scidpaiiIaic
egfstbspa[
Crufipiiabubble method essentially increases contact area between CO2and liquid media, and contact between CO2 and the microorgan-ism cells as well. These kinetic curves also showed fast → slowbehavior.
Table 8Comparison of the experimental conditions for different kinetic curves of E. colideactivation
Kinetic curve P (MPa) T (◦C) Note Researcher
Fast → slow 5 RT Used filter Debs-Louka et al. [35]
Fig. 3. Different kinds of kine
how only one fast linear stage (Fig. 3(c)) [40,57,63,64], whichould be just a variation of the two-stage kinetics. As discussedn Section 5.1, with an increase in temperature and pressure, theuration of the first stage becomes shorter [21,55]. At some tem-eratures, only the second stage (fast) kinetics may be observ-ble. Even though the slow-to-fast two-stage kinetic behaviors dominant, several studies [15,30,31,35,41,56] showed a fastnitial rate followed by a slow deactivation stage (Fig. 3(d)).nterestingly, both kinds of kinetic curves appeared in Hongnd Pyun [65] (Fig. 3(e)). It seems that when the pressure wasncreased from 5 to 7 MPa, the kinetic behavior of the systemhanged from the slow → fast to fast → slow.
One reason for variation in kinetic behavior may be the differ-nce in the efficiency of contact between CO2 and the microor-anism. The following table on E. coli sterilization is collectedrom literature to illustrate this hypothesis (Table 8). High pres-ure should facilitate gas transfer through media and increasehe contact between CO2 and cells. As in the static process usedy Hong et al., experiments under high pressure (20 MPa) [15]
howed fast → slow kinetics, while the experiments under lowerressure (1.2–10 MPa) [55,60] showed slow → fast kinetics. Inddition, the experiments by Ballestra et al. [55] and Erkmen60] were conducted in aqueous cell suspension (Table 7), soS
rves reported in publications.
O2 first had to diffuse through the liquid environment beforeeaching the cells. Debs-Louka et al. [35] used filter paper inoc-lated with E. coli suspension, so cells were deposited on paperbers and were covered only by a thin liquid film. Therefore, theenetration time was less than in a solution. One more examples the work by Ishikawa et al. with the micro-bubble method,n which pressurization occurs through bubbling CO2 through
filter at the bottom of the pressure vessel [56]. The micro-
paper20 35 Static exp. Kamihira et al. [15]
low → fast 1.2–5 25–45 Static exp. Ballestra et al. [55]2.5–10 20–40 Static exp. Erkmen [60]

ritica
6
ims
tHssss[sa1tEnmbdwtacewpscpt(scpec
Hm
5aarsAspBm(it0boasn[
7
dewma
ne[
J. Zhang et al. / J. of Superc
. Patents
Several patents have been filed using CO2 in various steril-zation applications, including both solid (soft tissues, polymers,
edical instruments) and liquid (foodstuffs and medicines) sub-trates.
Pilz and Rupp (US 4,263,253) is the first U.S. patent men-ioning supercritical fluids in the sterilization of solids [66].owever, in this patent the actual sterilization was achieved by
terile filtration, and the supercritical fluids were only used asolvents. Fischer and Muller (US 5,043,280) proposed use of aupercritical gas in an integrated method of manufacturing andterilizing polymeric particles embedded with active ingredient67]. This patent reported complete deactivation of 104 cfu ofeveral species of microorganism per gram of poly-lactide withsupercritical CO2 treatment at 50 ◦C, 14 MPa, for 12 h or 40 ◦C,4 MPa, for 0.5 h. The microorganisms tested included B. sub-ilis spores, B. stearothermophilus spores, E. coli, Salmonelladinburgh, S. aureus, P. aeruginosa, Candida albicans, and A.iger. Dillow et al. (US 6,149,864) patented a supercritical fluidethod for sterilization, especially for polymeric materials for
iomedical applications [68]. Solid PLGA micro-spheres withiameters of 1, 7, and 20 �m, and porous PLGA micro-spheresith a diameter of 10 �m were inoculated with E. coli and
reated with supercritical CO2. The presence of polymer did notffect the deactivation kinetics. No morphological or chemicalhanges were observed in the polymer micro-spheres. Poddevint al. (US 6,120,558) described a SCF method to simultaneouslyash, clean, desize and disinfect textile structures containing aoly(�-hydroxy-acids) [69]. Kanno (US 6,610,251) claimed aterilization method for medical instruments using supercriti-al fluids, including ethylene, ethanol, nitrogen oxide, ethane,ropane, and carbon dioxide [70]. However, the objective ofhis patent targets only bacteria and viruses. Kafesjian et al.application number: US 2001-981274) have filed a patent on aupercritical fluid process for tissue preparation [71]. This patent
laims that infectious materials including bacteria, virus, andrions can be removed from soft-tissue by supercritical fluidxtraction. The recommended supercritical fluids include super-ritical fluorocarbons, alkanes, nitrous oxide, nitrogen, and CO2.bEpd
Fig. 4. Proposed mechanisms of deactivatio
l Fluids 38 (2006) 354–372 365
owever, it is unlikely that supercritical fluids can extract wholeicroorganisms.Osajima et al. (US 5,520,943 [72], US 5,667,835 [73], US
,704,276 [74], US 5,869,123 [75], US 6,616,849 [76]) filedseries of patents on the use of SCF to deactivate enzymes
nd microorganisms in liquids, and to deodorize liquid mate-ials, including liquid foodstuffs and medicines with eitheremi-continuous [72,73] or continuous operations [74–76].t 50 ◦C, 30 MPa, 106 cfu/ml of eight species of Bacillus
pores, including B. cereus, B. subtilis, B. megaterium, B.olymyxa, B. coagulans, B. circulans, B. licheniformis, and. macerans, were completely deactivated with 80 min treat-ent using a semi-continuous micro-bubble method [73]. Sims
US 6,331,272) designed a new membrane contactor to ster-lize liquids [77]. The liquid or compressed CO2 was con-acted in pores of the membrane. The Wildasin et al. (WO203816) patent uses gaseous or liquid CO2 to reduce micro-ial and/or enzymatic activity in a liquid produce such asrange juice [78]. Malchesky (US 6,558,622) has designedcleaning and antimicrobial decontamination process using
ub-critical fluids, including carbon dioxide, argon, xenon,itrous oxide, oxygen, helium, and mixtures of these gases79].
. Deactivation mechanism
Ever since the discovery of CO2-based sterilization in 1951,etermining the true deactivation mechanisms has been of inter-st. The chief mechanisms have been mentioned previously andill be discussed in depth in this section. There are two generalechanisms promoted in the literature, mechanical cell rupture,
nd physiological deactivation (Fig. 4 and Table 9).Physical cell rupture was the earliest proposed mecha-
ism. Cells were presumed to rupture because of the explosivexpansion of high-pressure CO2 during flash depressurization12]. This mechanism was supported by microscopy images of
urst cells [13]. However, based on electron microscopy [16],nomoto et al. proposed that, the rupture of cells happens duringressurization when the cells are swollen by CO2, rather thanuring the depressurization stage [18].n mechanisms of vegetative bacteria.
366 J. Zhang et al. / J. of Supercritical Fluids 38 (2006) 354–372
Table 9Summary of the mechanisms and kinetics of published papers
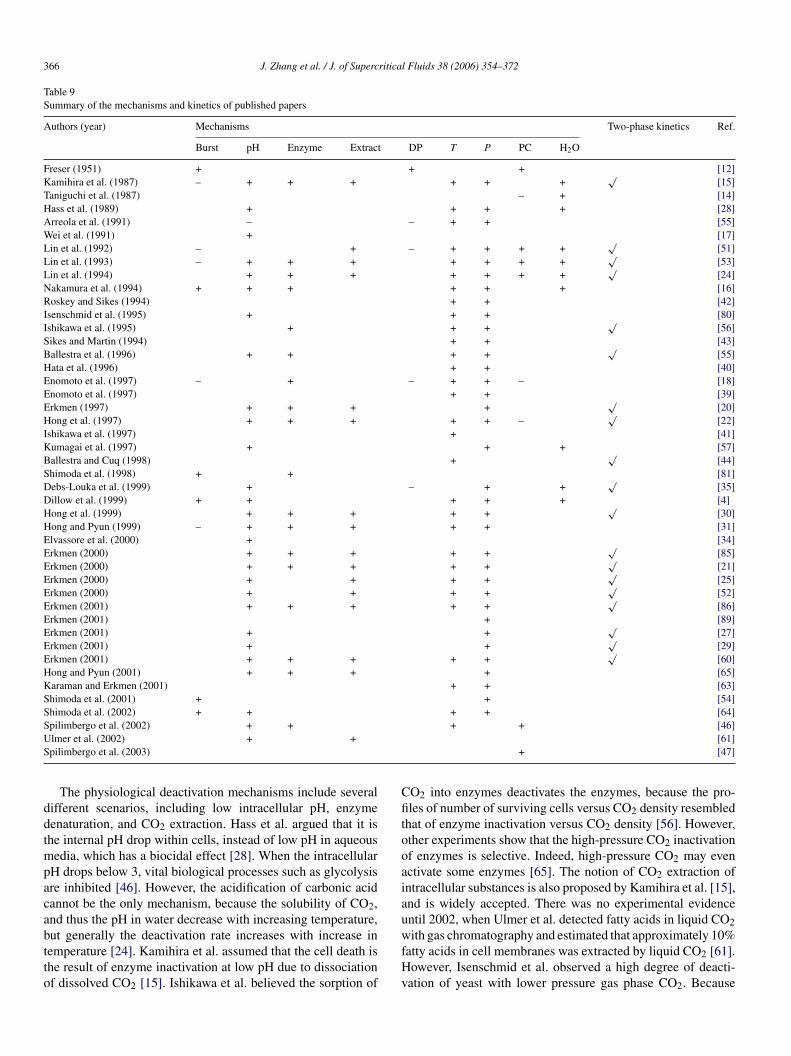
Authors (year) Mechanisms Two-phase kinetics Ref.
Burst pH Enzyme Extract DP T P PC H2O
Freser (1951) + + + [12]Kamihira et al. (1987) – + + + + + +
√[15]
Taniguchi et al. (1987) – + [14]Hass et al. (1989) + + + + [28]Arreola et al. (1991) – – + + [55]Wei et al. (1991) + [17]Lin et al. (1992) – + – + + + +
√[51]
Lin et al. (1993) – + + + + + + +√
[53]Lin et al. (1994) + + + + + + +
√[24]
Nakamura et al. (1994) + + + + + + [16]Roskey and Sikes (1994) + + [42]Isenschmid et al. (1995) + + + [80]Ishikawa et al. (1995) + + +
√[56]
Sikes and Martin (1994) + + [43]Ballestra et al. (1996) + + + +
√[55]
Hata et al. (1996) + + [40]Enomoto et al. (1997) – + – + + – [18]Enomoto et al. (1997) + + [39]Erkmen (1997) + + + +
√[20]
Hong et al. (1997) + + + + + –√
[22]Ishikawa et al. (1997) + [41]Kumagai et al. (1997) + + + [57]Ballestra and Cuq (1998) +
√[44]
Shimoda et al. (1998) + + [81]Debs-Louka et al. (1999) + – + +
√[35]
Dillow et al. (1999) + + + + + [4]Hong et al. (1999) + + + + +
√[30]
Hong and Pyun (1999) – + + + + + [31]Elvassore et al. (2000) + [34]Erkmen (2000) + + + + +
√[85]
Erkmen (2000) + + + + +√
[21]Erkmen (2000) + + + +
√[25]
Erkmen (2000) + + + +√
[52]Erkmen (2001) + + + + +
√[86]
Erkmen (2001) + [89]Erkmen (2001) + +
√[27]
Erkmen (2001) + +√
[29]Erkmen (2001) + + + + +
√[60]
Hong and Pyun (2001) + + + + [65]Karaman and Erkmen (2001) + + [63]Shimoda et al. (2001) + + [54]Shimoda et al. (2002) + + + + [64]SUS
ddtmpacabtto
Cfitooaiau
pilimbergo et al. (2002) + +lmer et al. (2002) + +pilimbergo et al. (2003)
The physiological deactivation mechanisms include severalifferent scenarios, including low intracellular pH, enzymeenaturation, and CO2 extraction. Hass et al. argued that it ishe internal pH drop within cells, instead of low pH in aqueous
edia, which has a biocidal effect [28]. When the intracellularH drops below 3, vital biological processes such as glycolysisre inhibited [46]. However, the acidification of carbonic acidannot be the only mechanism, because the solubility of CO2,nd thus the pH in water decrease with increasing temperature,
ut generally the deactivation rate increases with increase inemperature [24]. Kamihira et al. assumed that the cell death ishe result of enzyme inactivation at low pH due to dissociationf dissolved CO2 [15]. Ishikawa et al. believed the sorption ofwfHv
+ + [46][61]
+ [47]
O2 into enzymes deactivates the enzymes, because the pro-les of number of surviving cells versus CO2 density resembled
hat of enzyme inactivation versus CO2 density [56]. However,ther experiments show that the high-pressure CO2 inactivationf enzymes is selective. Indeed, high-pressure CO2 may evenctivate some enzymes [65]. The notion of CO2 extraction ofntracellular substances is also proposed by Kamihira et al. [15],nd is widely accepted. There was no experimental evidencentil 2002, when Ulmer et al. detected fatty acids in liquid CO2
ith gas chromatography and estimated that approximately 10%atty acids in cell membranes was extracted by liquid CO2 [61].owever, Isenschmid et al. observed a high degree of deacti-ation of yeast with lower pressure gas phase CO2. Because

ritica
gb
ptNceplowidesofoR[nptm
mHLaist
pstahhaba(aSmatms
8
kb
ciTaav[ti
tesCosais
htbapa
aavaaastp
cpmwpr[aea
opdva
J. Zhang et al. / J. of Superc
aseous CO2 is a very poor solvent, extraction could not haveeen the main mechanism in their experiments with yeast [80].
The relative importance of the cell rupture mechanism andhysiological mechanisms is extensively debated. Generally,hese two mechanisms are not regarded as mutually exclusive.akamura et al. observed morphological changes in some yeast
ells, such as wrinkles, holes, or completely broken cells afterxplosive decompression [16]. They speculated that at leastart of the inactivation is the result of explosive decompressioneading to cell rupture. Ballestra et al. indicate that both physi-logical and cell rupture mechanisms are important; their SEMork showed that more than 25% of the E. coli cell walls were
ntact, though other cell walls were deformed, after explosiveecompression [55]. Enomoto et al. compared the deactivationffects and protein release from both fast (4.8 MPa/min) andlow (0.033 MPa/min) decompression experiments. The levelsf deactivation and protein release were often nearly the same forast and slow decompression, and they concluded that the effectf fast decompression is not as great as suggested by others.ather, much of the deactivation occurs during pressurization
18]. Some researchers believe that cell rupture is not a mecha-ism of deactivation at all. Lin et al. found only a low level ofroteins in L. dextranicum cell supernatant, so they concludedhat the cell rupture did not occur significantly at their experi-
ental conditions [53].Some other researchers have proposed that the deactivation
echanisms are species-specific or vary with operation modes.ong and Pyun observed deformation in yeast, but not in treated. plantarum [31]. By comparing results between continuousnd batch systems, Shimoda et al. proposed that the mechanisms process-mode-dependent, that is, the mechanism for a batchystem is enzyme deactivation, while the mechanism for a con-inuous system is cell rupture [81].
It should be emphasized that all of the mechanisms pro-osed above have been based on treating vegetative cells, notpores. Enomoto et al. speculates that spores will be subjecto different mechanisms than vegetative cells [18]. Ballestra etl. concluded that the B. subtilis spores were first activated byeat (80–90 ◦C), and then destroyed by the synergistic effect ofeat and pressure [44]. Spilimbergo et al. [46,47] and Watan-be et al. [45] believed that the spores might be first germinatedy the application of pressure, then deactivated. Spilimbergo etl. processed B. cereus spores with pulsed electric field (PEF)4.5–25 kV/cm) followed by 15- and 24-h SC CO2 treatmentst 40 ◦C, 20 MPa. A synergistic effect between the PEF andC CO2 was observed. SEM images showed a distinct defor-ation (wrinkling and possibly cracking) of the spore surface
fter applying the PEF. Spores treated only with PEF showed lit-le deactivation, while the higher deactivation from PEF + CO2
ight be the result of enhanced penetration of CO2 into thepores [83].
. Experimental techniques
Several techniques have been used to quantify the level ofilling, and to characterize changes in integrity of the cell mem-rane, the permeability of the cell wall, and morphological
a
ce
l Fluids 38 (2006) 354–372 367
hanges (Table 10). The most common technique for determin-ng deactivation rates is the standard agar plate counting method.he treated cell suspension is serially diluted and inoculated togar plates. The number of colonies on each Petri dish is countedfter incubation for a fixed time. Then the colony count is con-erted to the cell concentration in the original cell suspension84]. Plate counting is an easy and effective way to quantifyhe number of viable cells, but does not reveal any mechanisticnformation.
To visually observe structure changes in cells, scanning elec-ron microscopy (SEM) [4,15,16,31,55,81] and transmissionlectron microscopy (TEM) [31] have been adopted to observeurface and internal structure changes induced by high-pressureO2 treatment. Burst cells (Fig. 5(a) left), wrinkles and holesn the cell surface (Fig. 5(a) right (b) left), as well as intact cellurfaces (Fig. 5(b) right) have been observed with SEM. Hongnd Pyun (1999) is the only study that used TEM [31], reveal-ng internal structure changes, including increased periplasmicpace and fractures in membranes (Fig. 5(c)).
UV absorbance [42,65,81] and protein analysis [18,51,53,58]ave been used to determine loss of cell content as a result of CO2reatment of wet cell slurries. The idea is that, if the cell mem-ranes are damaged, internal components such as lipids, aminocids, and peptides will be detectable in the media [65]. Theresence of these materials can be detected by UV absorbancend protein analysis.
All UV studies have reported an increase in absorbancefter CO2 treatment. Roskey et al. reported an increase in UVbsorbance in the range of 210–240 nm from a pressure-treatedegetative G. stearothermophilus cell suspension [42]. Hongnd Pyun measured steep increases in UV absorbance at 260nd 280 nm from a L. plantarum suspension [65]. Shimoda etl. also reported a “great increase” in UV absorbance from cellolution treated with a continuous CO2 process, but no quanti-ative data were given [81]. It should be noted that these studiesrovide no specific identification of the UV-absorbing species.
Proteins have been analyzed by Castor et al. using a bicin-honinic acid assay kit [19], by Enomoto et al. using the Lowryrotein assay [18], and by Lin et al. without specifying theethod employed [51,53,58]. Unlike absorbance experiments,hich showed release of UV-absorbing materials after high-ressure CO2 treatment, only Enomoto et al. identified proteinelease from baker’s yeast after high-pressure CO2 treatment18]. One would expect that cell rupture would release both UV-bsorbing compounds as well as proteins. There is no reasonablexplanation for the apparent inconsistency between UV resultsnd protein results.
Investigators have frequently attempted to relate the decreasef pH in the medium to the level of deactivation. In most cases,H was measured after depressurization [20,35,52,85,86], whichoes not give the pH during the CO2 treatment, because CO2 willaporize and pH will increase after depressurization. Hass et al.dopted a modified high-pressure pH meter in their experiments,
nd measured pH up to 6.2 MPa pressure [28].Other assays have provided useful information. Coulterounting gives the number and size of bacterial cells [23,80],nzyme activity assays reveal the effects of CO2 on enzymes

368 J. Zhang et al. / J. of Supercritical Fluids 38 (2006) 354–372
Table 10Summary of the measurement methods used in published papers
Authors (year) Plate TEM SEM UV Protein pH Enzyme Other Ref.
Freser (1951)√
[12]Kamihira et al. (1987)
√ √[15]
Hass et al. (1989)√ √
[28]Arreola et al. (1991)
√[55]
Wei et al. (1991)√
[17]Isenschmid et al. (1992)
√ √Coulter counter [23]
Lin et al. (1992)√ √
[51]Lin et al. (1993)
√ √[53]
Lin et al. (1994)√
[24]Nakamura et al. (1994)
√ √[16]
Roskey and Sikes (1994)√ √
[42]Isenschmid et al. (1995)
√ √Coulter counter [80]
Sikes and Martin (1994)√
[43]Ballestra et al. (1996)
√ √ √[55]
Hata et al. (1996)√
[40]Enomoto et al. (1997)
√ √[18]
Enomoto et al. (1997)√
[39]Erkmen (1997)
√ √[20]
Hong et al. (1997)√
[22]Ishikawa et al. (1997)
√[41]
Kumagai et al. (1997)√
Gravimeter, DSC [57]Ballestra and Cuq (1998)
√[44]
Shimoda et al. (1998)√ √ √
[81]Debs-Louka et al. (1999)
√ √[35]
Dillow et al. (1999)√ √
[4]Hong et al. (1999)
√[30]
Hong and Pyun (1999)√ √ √
[31]Elvassore et al. (2000)
√[34]
Erkmen (2000)√ √
[85]Erkmen (2000)
√ √[21]
Erkmen (2000)√ √
[25]Erkmen (2000)
√ √[52]
Erkmen (2001)√ √
[86]Erkmen (2001)
√[89]
Erkmen (2001)√
[27]Erkmen (2001)
√ √[29]
Erkmen (2001)√ √
[60]Hong and Pyun (2001)
√ √ √ √Glucose assay, K and Mg ionassay, proton permeabilityassay, constituitive enzymeactivity assay, H+-ATPaseactivity assay
[65]
Karaman and Erkmen (2001)√
[63]Shimoda et al. (2001)
√[54]
Shimoda et al. (2002)√
[64]Spilimbergo et al. (2002)
√[46]
Ulmer et al. (2002)√
Membrane permeability; GC [61]Spilimbergo et al. (2003)
√[47]
Parton et al. (2003)√ √
Phloxine B stain [87]Watanabe et al. (2003)
√[45]
S√
[HouacpL
9
f
chmidt et al. (2005)
55], and dye uptake helps reveal membrane damage [65,87].ong and Pyun provided the most comprehensive studyf membrane damage and enzyme inactivation [65]. Theysed a traditional plate counting technique, as well as UV
bsorbance, proton permeability, K and Mg ion assay, glu-ose assay, and enzyme activity assay. Their experiments sup-orted the idea of “irreversible cellular membrane damage” to. plantarum.ncee
[62]
. Modeling work
The majority of publications report experimental data; only aew publications propose models of either kinetic or thermody-
amic aspects (Table 11). The kinetics of deactivation are usuallyharacterized by a first-order expression. For example, Karamant al. correlated the deactivation rate constant k for a first-orderxpression [63]. However, Peleg pointed out that survival curves
J. Zhang et al. / J. of Supercritical Fluids 38 (2006) 354–372 369
opic
aie
L
ELm
[
l
T
Fig. 5. Electron microsc
re usually non-linear, so it is not proper to express the kinet-cs with a first-order relationship. Peleg proposed the followingmpirical expression for an isobaric survival curve [88]:
og reduction = b(P)tn(p)
rkmen correlated data from dense-phase CO2 treatment of. monocytogenes with different sigmoid models and recom-ended the three-parameter-modified Gompertz model below
μ
Nt
images of treated cells.
26]:
og N = log N0 + A exp{
−exp[μe
A(λ − t) + 1
]}
he parameter μ is correlated with an Arrhenius model [89]:( )
= A exp −EμRT
0 and N are the number of cells initial and surviving, respec-ively, μ the inactivation rate, λ the phase of disappearance (time

370 J. Zhang et al. / J. of Supercritica
Table 11Reported modeling work
KineticsFirst-order linear Karaman et al. [63]Empirical non-linear Peleg [88]Sigmoid models Erkmen [26]Empirical eq. from the response surface model Debs-Louka et al. [35]Mass transfer model for CO2 diffusionthrough the medium
Elvassore et al. [34]
T
fo
al
wctc
bpc
1
pc
1
2
3
Ci
1
2
3
A
cn
R
hermodynamicspH drop and CO2 solubility in phospholipid Spilimbergo et al. [46]
or complete inactivation), and A is the lower asymptote valuef log (N/N0).
Debs-Louka et al. used response surface models to generatepolynomial relationship between pressure, treatment time and
og reduction [35]. Their expressions are totally empirical.To compare different operation modes, Elvassore proved
ith simple time-dependent mass transfer models that a semi-ontinuous device takes at least one order of magnitude lessime to reach the same CO2 concentration in the water phase,ompared with a batch operation device [34].
There is only one report of thermodynamic modeling. Spilim-ergo et al. reported the pH drop and CO2 solubility in phos-holipids with thermodynamic modeling, but no details of thealculation were given [46].
0. Current research status and challenges
After a comprehensive review of prior research on high-ressure CO2 treatment of microorganisms, the status of theurrent research is summarized as follows:
. vegetative bacteria have been extensively studied and are sus-ceptible to high-pressure CO2 treatment;
. the effects of temperature, pressure and medium on deacti-vation of vegetative cells can be substantial, but there is notyet a clear understanding of these effects;
. several deactivation mechanisms have been proposed. Whilethere are some common features, there are also points ofdisagreement. It is difficult to acquire unambiguous data thatconfirm a given mechanism. It may be that different speciesreact differently to CO2, and that the precise mechanistic pathvaries by organism.
In order to address the goal of developing a high-pressureO2 sterilization method, there are several challenges remain-
ng:
. True sterilization requires killing of spores, but data on theeffect of CO2 on bacterial spores is quite limited. The exper-imental conditions reported for killing spores appear to betoo extreme (high temperature, high pressure, long treatment
time) to be competitive with other sterilization techniques.A key question is how to adapt methods, such as pressurecycling and additives, to achieve sterilization at milder pro-cess conditions.[
[
l Fluids 38 (2006) 354–372
. Some data are available on the effects of pressure cycling andthe use additives; these methods have promise for enhancingthe deactivation process and leading to milder conditions.
. The weakest links in knowledge are mechanistic studies andmathematical modeling for process design. These are essen-tial to understanding the fundamentals of the high-pressureCO2 treatment process.
The mechanisms of vegetative cell deactivation are not yetclear, although several mechanisms have been proposed. Themechanisms of spore deactivation have scarcely been studied.In addition, the mechanisms of vegetative cell deactivationmay not be applicable to spore deactivation. For example,because the spore core is highly dehydrated, it is less likelythat high-pressure CO2 can lower the internal pH within thecore.
Mathematical modeling of CO2 sterilization also needsto be strengthened because it is important for elucidatingthe mechanisms, and for process optimization. On the onehand, no modeling work has been done to test the assumedmechanism of CO2 extraction of cell contents; on the otherhand, mathematical modeling can provide information on themaximum amount of additives in CO2 under the operatingconditions, which is important for process optimization.
cknowledgement
This work is supported by NIH/National Institute of Biomedi-al Imaging and Bioengineering, Bioengineering Research Part-ership (NIH BRP Grant R01 EB55201).
eferences
[1] A. Praemer, S. Furner, D.P. Rice, Musculoskeletal Conditions in the UnitedStates, American Academy of Orthopaedic, USA, 1992.
[2] Y.H. An, R.J. Friedman, Handbook of Bacterial Adhesion: Principles,Methods, and Applications, Humana Press, Totowa, NJ, 2000.
[3] M.A. Matthews, L.S. Warner, H. Kaiser, Exploring the feasibility of usingdense-phase carbon dioxide for sterilization, Med. Device Diagn. Ind. 5(2001) 140.
[4] A.K. Dillow, F. Dehghani, J.S. Hrkach, N.R. Foster, R. Langer, Bacterialinactivation by using near- and supercritical carbon dioxide, Proc. Natl.Acad. Sci. U.S.A. 96 (1999) 10344.
[5] J. Lausmaa, B. Kasemo, S. Hansson, Accelerated oxide growth on titaniumimplants during autoclaving caused by fluorine contamination, Biomateri-als 6 (1985) 23.
[6] V. Premnath, W.H. Harris, M. Jasty, E.W. Merrill, Gamma sterilizationof UHMWPE articular implants: an analysis of the oxidation problem,Biomaterials 17 (1996) 1741.
[7] M. Moisan, J. Barbeau, S. Moreau, J. Pelletier, M. Tabrizian, L.H. Yahia,Low-temperature sterilization using gas plasmas: a review of the experi-ments and an analysis of the inactivation mechanisms, Int. J. Pharm. 226(2001) 1.
[8] S. Spilimbergo, A. Bertucco, Non-thermal bacteria inactivation with denseCO2, Biotechnol. Bioeng. 84 (2003) 627.
[9] M. McHugh, V. Krukonis, Supercritical Fluid Extraction,Butterworth–Heinemann, USA, 1993.
10] A.I. Cooper, Recent developments in materials synthesis and processingusing supercritical CO2, Adv. Mater. 13 (2001) 1111.
11] S. Moshashaee, M. Bisrat, R.T. Forbes, H. Nyqvist, P. York, Supercriticalfluid processing of proteins. Part I. Lysozyme precipitation from organicsolution, Eur. J. Pharm. Sci. 11 (2000) 239.

ritica
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
J. Zhang et al. / J. of Superc
12] D. Fraser, Bursting bacteria by release of gas pressure, Nature 167 (1951)33.
13] J.W. Foster, R.M. Cowan, T.A. Maag, Rupture of bacteria by explosivedecompression, J. Bacteriol. 83 (1962) 330.
14] M. Taniguchi, H. Suzuki, M. Sato, T. Kobayashi, Sterilization of plasmapowder by treatment with supercritical carbon dioxide, Agric. Biol. Chem.51 (1987) 3425.
15] M. Kamihira, M. Taniguchi, T. Kobayashi, Sterilization of microorgan-isms with supercritical carbon dioxide, Agric. Biol. Chem. 51 (1987)407.
16] K. Nakamura, A. Enomoto, H. Fukushima, K. Nagai, M. Hakoda, Dis-ruption of microbial-cells by the flash discharge of high-pressure carbon-dioxide, Biosci. Biotechnol. Biochem. 58 (1994) 1297.
17] C.I. Wei, M.O. Balaban, S.Y. Fernando, A.J. Peplow, Bacterial effect ofhigh-pressure CO2 treatment on foods spiked with Listeria or Salmonella,J. Food Protect. 54 (1991) 189.
18] A. Enomoto, K. Nakamura, K. Nagai, T. Hashimoto, M. Hakoda, Inacti-vation of food microorganisms by high-pressure carbon dioxide treatmentwith or without explosive decompression, Biosci. Biotechnol. Biochem. 61(1997) 1133.
19] T.P. Castor, G.T. Hong, Critical fluid disruption of microbial cells, in: Pro-ceedings of 2nd International Symposium on Supercritical Fluids, 1991, p.139.
20] O. Erkmen, Antimicrobial effect of pressurized carbon dioxide on Staphy-lococcus aureus in broth and milk, Food Sci. Technol.-Lebensm.-Wiss.Technol. 30 (1997) 826.
21] O. Erkmen, Inactivation of Salmonella typhimurium by high pressure car-bon dioxide, Food Microbiol. 17 (2000) 225.
22] S.I. Hong, W.S. Park, Y.R. Pyun, Inactivation of Lactobacillus sp. fromkimchi by high pressure carbon dioxide, Food Sci. Technol.-Lebensm.-Wiss. Technol. 30 (1997) 681.
23] A. Isenschmid, I.W. Marison, U. Von Stockar, Effect of a near-criticaland supercritical fluid on the viability ratio of microbial cells, ProgressBiotechnol. 8 (1992) 407.
24] H.M. Lin, N.J. Cao, L.F. Chen, Antimicrobial effect of pressurized carbondioxide on Listeria monocytogenes, J. Food Sci. 59 (1994) 657.
25] O. Erkmen, Effect of carbon dioxide pressure on Listeria monocyto-genes in physiological saline and foods, Food Microbiol. 17 (2000)589.
26] O. Erkmen, Predictive modelling of Listeria monocytogenes inactivationunder high pressure carbon dioxide, Lebensm.-Wiss. Technol.-Food Sci.Technol. 33 (2000) 514.
27] O. Erkmen, Kinetic analysis of Listeria monocytogenes inactivation by highpressure carbon dioxide, J. Food Eng. 47 (2001) 7.
28] G.J. Haas, H.E. Prescott, E. Dudley, R. Dik, C. Hintlian, L. Keane, Inacti-vation of microorganisms by carbon dioxide under pressure, J. Food Safety9 (1989) 253.
29] O. Erkmen, H. Karaman, Kinetic studies on the high pressure carbon diox-ide inactivation of Salmonella typhimurium, J. Food Eng. 50 (2001) 25.
30] S.I. Hong, W.S. Park, Y.R. Pyun, Non-thermal inactivation of Lactobacillusplantarum as influenced by pressure and temperature of pressurized carbondioxide, Int. J. Food Sci. Technol. 34 (1999) 125.
31] S.I. Hong, Y.R. Pyun, Inactivation kinetics of Lactobacillus plantarum byhigh pressure carbon dioxide, J. Food Sci. 64 (1999) 728.
32] M.T. Madigan, J.M. Martinko, J. Parker, Brock Biology of Microorganisms,Prentice Hall, USA, 2002.
33] A.L. Sonenshein, J.A. Hoch, R. Losick, Bacillus subtilis and its ClosestRelatives — From Genes to Cells, ASM Press, USA, 2001.
34] N. Elvassore, S. Sartorello, S. Spilimbergo, A. Bertucco, Micro-organismsinactivation by supercritical CO2 in a semi-continuous process 2 (2000)773.
35] E. Debs-Louka, N. Louka, G. Abraham, V. Chabot, K. Allaf, Effect ofcompressed carbon dioxide on microbial cell viability, Appl. Environ.
Microbiol. 65 (1999) 626.36] G.J. Tortora, B.R. Funke, C.L. Case, Microbiology, An Introduction, Pear-son Education, USA, 2002.
37] A. Driks, Bacillus subtilis spore coat, Microbiol. Mol. Biol. Rev. 63 (1999)1.
[
[
l Fluids 38 (2006) 354–372 371
38] T.A. Slieman, W.L. Nicholson, Role of dipicolinic acid in survival ofBacillus subtilis spores exposed to artificial and solar UV radiation, Appl.Environ. Microbiol. 67 (2001) 1274.
39] A. Enomoto, K. Nakamura, M. Hakoda, N. Amaya, Lethal effect of high-pressure carbon dioxide on a bacterial spore, J. Ferment. Bioeng. 83 (1997)305.
40] C. Hata, H. Kumagai, K. Nakamura, Rate analysis of the sterilization ofmicrobial cells in high pressure carbon dioxide, Food Sci. Technol. Int. 2(1996) 229.
41] H. Ishikawa, M. Shimoda, K. Tamaya, A. Yonekura, T. Kawano, Y.Osajima, Inactivation of Bacillus spores by the supercritical carbondioxide micro-bubble method, Biosci. Biotechnol. Biochem. 61 (1997)1022.
42] C.T. Roskey, A. Sikes, Effect of hyperbaric carbon dioxide on sporesand vegetative cells of Bacillus stearothermophilus, NATICK/TR-94/019(1994).
43] A. Sikes, C. Martin, Control of thermophilic spore activity with pressurizedCO2 and egg-white lysozyme, Report, NATICK/TR-95/020 (1995).
44] P. Ballestra, J.L. Cuq, Influence of pressurized carbon dioxide on thethermal inactivation of bacterial and fungal spores, Food Sci. Technol.-Lebensm.-Wiss. Technol. 31 (1998) 84.
45] T. Watanabe, S. Furukawa, J. Hirata, T. Koyama, H. Ogihara, M. Yamasaki,Inactivation of Geobacillus stearothermophilus spores by high-pressurecarbon dioxide treatment, Appl. Environ. Microbiol. 69 (2003) 7124.
46] S. Spilimbergo, N. Elvassore, A. Bertucco, Microbial inactivation by high-pressure, J. Supercrit. Fluids 22 (2002) 55.
47] S. Spilimbergo, A. Bertucco, F.M. Lauro, G. Bertoloni, Inactivation ofBacillus subtilis spores by supercritical CO2 treatment, Innov. Food Sci.Emerg. Technol. 4 (2003) 161.
48] J.C. Cheftel, Review: high-pressure, microbial inactivation and food preser-vation, Food Sci. Technol. Int. 1 (1995) 75.
49] Institute of Food Technologists, Special supplement: kinetics of microbialinactivation for alternative food processing technologies — IFT’s responseto task order #1, US Food and Drug Administration: how to quantify thedestruction kinetics of alternative processing technologies, J. Food Sci.(2000) 4.
50] A.G. Arreola, M.O. Balaban, C.I. Wei, A. Peplow, M. Marshall, J. Cornell,Effect of supercritical carbon dioxide on microbial populations in singlestrength orange juice, J. Food Qual. 14 (1991) 275.
51] H.M. Lin, Z.Y. Yang, L.F. Chen, Inactivation of Saccharomyces cerevisiaeby supercritical and subcritical carbon dioxide, Biotechnol. Progress 8(1992) 458.
52] O. Erkmen, Antimicrobial effect of pressurised carbon dioxide on Bro-chothrix thermosphacta in broth and foods, J. Sci. Food Agric. 80 (2000)1365.
53] H.M. Lin, Z.Y. Yang, L.F. Chen, Inactivation of Leuconostoc dextranicumwith carbon dioxide under pressure, Chem. Eng. J. 52 (1993) B29.
54] M. Shimoda, J. Cocunubo-Castellanos, H. Kago, M. Miyake, Y. Osajima,I. Hayakawa, The influence of dissolved CO2 concentration on the deathkinetics of Saccharomyces cerevisiae, J. Appl. Microbiol. 91 (2001) 306.
55] P. Ballestra, A.A. Dasilva, J.L. Cuq, Inactivation of Escherichia coli bycarbon dioxide under pressure, J. Food Sci. 61 (1996) 829.
56] H. Ishikawa, H. Shimoda, H. Shiratsuchi, Y. Osajima, Sterilization ofmicroorganisms by the supercritical carbon dioxide micro-bubble method,Biosci. Biotechnol. Biochem. 59 (1995) 1949.
57] H. Kumagai, C. Hata, K. Nakamura, CO2 sorption by microbial cells andsterilization by high-pressure CO2, Biosci. Biotechnol. Biochem. 61 (1997)931.
58] H.M. Lin, E.T. Lin, Inactivation of microorganisms with supercritical fluids,Front. Foods Food Ingred. 1 (1993) 253 (Science for the Food Industry ofthe 21st Century).
59] E.Y. Wuytack, J. Soons, F. Poschet, C.W. Michiels, Comparative studyof pressure- and nutrient-induced germination of Bacillus subtilis spores,
Appl. Environ. Microbiol. 66 (2000) 257.60] O. Erkmen, Effects of high-pressure carbon dioxide on Escherichia coli innutrient broth and milk, Int. J. Food Microbiol. 65 (2001) 131.
61] H.M. Ulmer, D. Burger, M.G. Ganzle, H. Engelhardt, R.F. Vogel, Effectof compressed gases on the high pressure inactivation of Lactobacillus

3 ritica
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
72 J. Zhang et al. / J. of Superc
plantarum TMW 1.460, Progress Biotechnol. 19 (2002) 317 (Trends inHigh Pressure Bioscience and Biotechnology).
62] A. Schmidt, K. Beermann, E. Bach, E. Schollmeyer, Disinfection of textilematerials contaminated with E. coli in liquid carbon dioxide, J. Clean. Prod.13 (2005) 881.
63] H. Karaman, O. Erkmen, High carbon dioxide pressure inactivation kineticsof Escherichia coli in broth, Food Microbiol. 18 (2001) 11.
64] M. Shimoda, H. Kago, N. Kojima, M. Miyake, Y. Osajima, I. Hayakawa,Accelerated death kinetics of Aspergillus niger spores under high-pressurecarbonation, Appl. Environ. Microbiol. 68 (2002) 4162.
65] S.I. Hong, Y.R. Pyun, Membrane damage and enzyme inactivation of Lacto-bacillus plantarum by high pressure CO2 treatment, Int. J. Food Microbiol.63 (2001) 19.
66] V. Pilz, R. Rupp, Process for rendering solids sterile, United States Patent,US 4,263,253 (1981).
67] W. Fischer, B.W. Muller, Method and apparatus for the manufacture of aproduct having a substance embedded in a carrier, United States Patent, US5,043,280 (1991).
68] A.K. Dillow, R.S. Langer, N. Foster, J.S. Hrkach, Supercritical fluid steril-ization method, United States Patent, US 6,149,864 (2000).
69] N. Poddevin, J. Fages, R. Guidoin, Method for manufacturing and treatingtextiles, United States Patent, US 6,120,558 (2000).
70] M. Kanno, Method of sterilizing medical instruments, United States Patent,US 6,610,251 B1 (2003).
71] R. Kafesjian, M.J.R. Howanec, Supercritical fluid extraction process fortissue preparation, United States Patent Application Publication, Pub. No.US 2003/072677 A1 (2003).
72] Y. Osajima, M. Shimoda, T. Kawano, Method for modifying the quality ofliquid foodstuff, United States Patent, US 5,520,943 (1996).
73] Y. Osajima, M. Shimoda, T. Kawano, Method for inactivating enzymes,microorganisms, and spores in a liquid foodstuff, United States Patent, US5,667,835 (1997).
74] Y. Osajima, M. Shimoda, T. Kawano, K. Okubo, System for processing liq-uid foodstuff or liquid medicine with a supercritical fluid of carbon dioxide,United States Patent, US 5,704,276 (1998).
75] Y. Osajima, M. Shimoda, T. Kawano, K. Okubo, System for processing liq-
uid foodstuff or liquid medicine with a supercritical fluid of carbon dioxide,United States Patent, US 5,869,123 (1999).76] Y. Osajima, M. Shimoda, M. Takada, M. Miyake, Method of and systemfor continuously processing liquid materials, and the product processedthereby, United States Patent, US 6,616,849 B1 (2003).
[
[
l Fluids 38 (2006) 354–372
77] M. Sims, Method and membrane system for sterilizing and preservingliquids using carbon dioxide, United States Patent, US 6,331,272 B1(2001).
78] R.E. Wildasin, J. Forbes, R. Robey, A.J. Paradis, Treating liquid food andother products using carbon dioxide, International Patent, WO 02/03816A1 (2002).
79] P.S. Malchesky, Sub-critical fluid cleaning and antimicrobial decontam-ination system and process, United States Patent, US 6,558,622 B1(2003).
80] A. Isenschmid, I.W. Marison, U. Vonstockar, The influence of pressure andtemperature of compressed CO2 on the survival of yeast cells, J. Biotechnol.39 (1995) 229.
81] M. Shimoda, Y. Yamamoto, J. Cocunubo-Castellanos, H. Tonoike, T.Kawano, H. Ishikawa, Y. Osajima, Antimicrobial effects of pressuredcarbon dioxide in a continuous flow system, J. Food Sci. 63 (1998)709.
82] C.G. Mallidis, J. Scholefield, The release of dipicolinic acid during heat-ing and its relation to the heat destruction of Bacillus stearothermophilusspores, J. Appl. Bacteriol. 59 (1985) 479.
83] S. Spilimbergo, F. Dehghani, A. Bertucco, N.R. Foster, Inactivation ofbacteria and spores by pulse electric field and high pressure CO2 at lowtemperature, Biotechnol. Bioeng. 82 (2003) 118.
84] J.G. Cappuccino, N. Sherman, Microbiology: A Laboratory Manual, Ben-jamin Cummings, USA, 2001.
85] O. Erkmen, Antimicrobial effect of pressurised carbon dioxide on Entero-coccus faecalis in physiological saline and foods, J. Sci. Food Agric. 80(2000) 465.
86] O. Erkmen, Note. Antimicrobial effect of pressurized carbon dioxide onYersinia enterocolitica in broth and foods, Food Sci. Technol. Int. 7 (2001)245.
87] T. Parton, E. Rocchi, S. Spilimbergo, N. Elvassore, A. Bertucco, Ster-ilization of fruit juice by high pressure CO2, advances in high pressurebioscience and biotechnology II, in: Proceedings of the Second Interna-tional Conference on High Pressure Bioscience and Biotechnology (2003)419.
88] M. Peleg, Simulation of E-coli inactivation by carbon dioxide under pres-
sure, J. Food Sci. 67 (2002) 896.89] O. Erkmen, Mathematical modeling of Escherichia coli inactivation underhigh-pressure carbon dioxide, J. Biosci. Bioeng. 92 (2001) 39.
90] J.P.P.M. Smelt, G.G.F. Rijke, High pressure treatment as a tool for pasteur-ization of foods, High Pressure Biotechnol. 224 (1992) 361.