Embed Size (px)
Citation preview

Stepwise androgen receptor dimerization
Martin E. van Royen1, Wiggert A. van Cappellen2, Carola de Vos1, Adriaan B. Houtsmuller1 and Jan Trapman1,*1Department of Pathology, Josephine Nefkens Institute, Erasmus University Medical Center, PO-Box 2040, 3000 CA Rotterdam, The Netherlands2Department of Reproduction and Development, Erasmus University Medical Center, PO-Box 2040, 3000 CA Rotterdam, The Netherlands
*Author for correspondence ([email protected])
Accepted 15 December 2011Journal of Cell Science 125, 1970–1979� 2012. Published by The Company of Biologists Ltddoi: 10.1242/jcs.096792
SummaryAndrogen-regulated gene expression is a highly coordinated dynamic process mediated by androgen receptor (AR) ligand binding andDNA binding, and by specific AR protein–protein interactions. The latter include DNA-binding domain (D-box) interactions in ARhomodimers, and the interaction of the FQNLF motif in the AR N-terminal domain and the coactivator groove in the ligand-bindingdomain (N/C interaction). We have studied these interactions in AR homodimerization using quantitative imaging techniques. We found
that the initial cytoplasmic intramolecular AR N/C interaction after ligand binding is followed by a D-box-dimerization-dependenttransition to intermolecular N/C interaction in a proportion of nuclear ARs. The consecutive steps leading to homodimerization areinitiated prior to DNA binding. Our data indicate the presence of nuclear pools of both AR homodimers and monomers. On the basis of
AR-regulated reporter assays we propose specificity in regulation of gene expression by AR homodimers and monomers mediated byAR domain interactions. Moreover, our findings elucidate important steps in the spatiotemporal organization of AR intra- and inter-molecular interactions.
Key words: Androgen receptor, Dimerization, N/C interaction, DBD, Quantitative live cell imaging, Target genes
IntroductionRegulation of gene expression is a dynamic process involving
many tightly orchestrated consecutive steps. Androgen-regulated
gene expression is mediated by the androgen receptor (AR). The
AR is a ligand-activated transcription factor and a member of the
steroid receptor (SR) subfamily of nuclear receptors (NRs). Like
all SRs, the AR has a modular structure composed of an N-terminal
domain (NTD), a conserved DNA-binding domain (DBD) and a C-
terminal ligand-binding domain (LBD) (Brinkmann et al., 1989).
Activated ARs regulate genes involved in the development and
maintenance of the male phenotype. AR is also a key factor in
prostate cancer. AR activity is not only regulated by ligand binding
and DNA binding but also by intramolecular interactions between
functional domains, by homodimerization and by interactions
with cofactors. The best-characterized interactions between AR
functional domains are the intra- and intermolecular NTD–LBD
interaction (N/C interaction) that is mediated by the FQNLF motif
in the NTD and the coactivator groove in the LBD, and
the intermolecular DBD–DBD interaction mediated by the
dimerization box (D-box) (Centenera et al., 2008). However,
the spatiotemporal relationship of the different intra- and
intermolecular AR domain interactions in androgen-regulated
gene expression is currently unknown.
Using fluorescence resonance energy transfer (FRET) and
combined fluorescence recovery after photobleaching (FRAP)
and FRET analysis, initial studies on the spatiotemporal
organization of AR protein–protein interactions have been
performed (Schaufele et al., 2005; van Royen et al., 2007).
FRET showed that in the cytoplasm the N/C interaction is in an
intramolecular conformation initiated directly after ligand-
binding and before translocation to the nucleus (Schaufele et al.,
2005; van Royen et al., 2007). In the nucleus, the intramolecular
N/C interaction is followed by an intermolecular N/C interaction
(Schaufele et al., 2005). The N/C interaction preferentially occurs
in mobile ARs and is lost when the AR is bound to DNA (van
Royen et al., 2007). These observations indicate that the AR itself
regulates the time and place of interactions with coregulators by
preventing untimely protein interactions when the AR is mobile,
and allowing coregulator binding when the AR is bound to DNA
(Dubbink et al., 2004; He et al., 2001; van Royen et al., 2007).
The intramolecular and intermolecular N/C interactions are
mediated by binding of the FxxLF peptide motif (FQNLF) in the
AR NTD to the ligand-induced cofactor binding groove in AR
LBD. The phenylalanine residues in the FxxLF motif are
essential for strong N/C interaction and bind deep into the
coactivator groove with van der Waals interactions, whereas
the leucine residue in the peptide motif lies in a shallow ridge on
the surface of the LBD, and the other two amino acid residues are
exposed to the solvent (Dubbink et al., 2004; Hur et al., 2004; van
de Wijngaart et al., 2006). In other SRs, N/C interactions are
absent or weak. In a homodimer, ARs also interact through their
D-boxes in the second zinc finger of the DBD. SR D-box
interactions are sustained by a network of hydrogen bonds
between individual amino acid residues in the D-box and by an
extensive complementary surface. In the AR DBD, a serine
residue at position 597 (S597), which is absent in other SRs,
forms a hydrogen bond and Van der Waals contacts with its
counterpart in the opposing D-box in an AR homodimer (Shaffer
et al., 2004). An additional pair of symmetrical hydrogen bonds is
formed between an alanine at 596 (A596) and a threonine at
602 (T602) in the opposing AR DBD and vice versa. These
interactions result in a relatively strong AR D-box dimerization
interface compared with those of other SRs (Shaffer et al., 2004).
The importance of the D-box in AR function is highlighted by the
1970 Research Article
Journ
alof
Cell
Scie
nce
large number of mutations in this domain found in androgen
insensitivity syndrome (AIS) patients (Centenera et al., 2008)
(http://androgendb.mcgill.ca/).
To study the molecular mechanisms underlying AR
homodimerization and to investigate when and where domain
interactions take place, we used confocal microscopy and
quantitative microscopic techniques to examine cells expressing
functional, single and double YFP- and CFP-tagged wild-type ARs
and appropriate AR mutants. In addition, we investigated the role
that these molecular mechanisms have in differential target gene
expression.
ResultsInteractions of functional domains of single and double
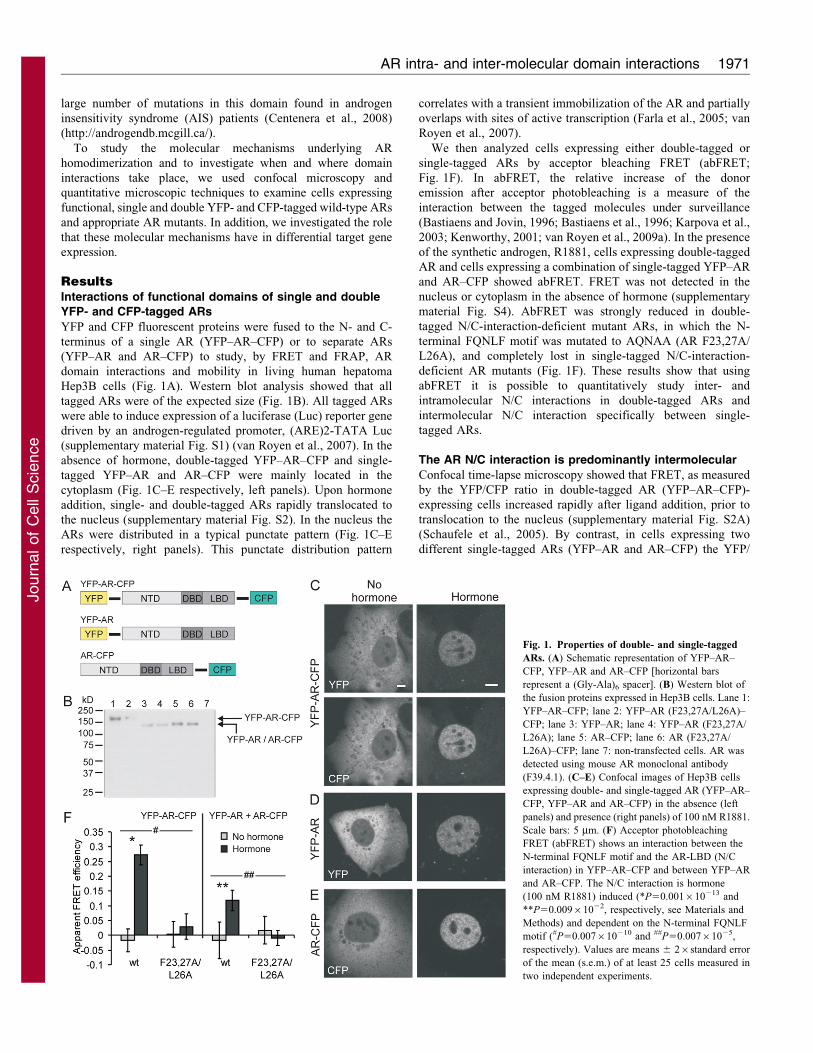
YFP- and CFP-tagged ARs
YFP and CFP fluorescent proteins were fused to the N- and C-
terminus of a single AR (YFP–AR–CFP) or to separate ARs
(YFP–AR and AR–CFP) to study, by FRET and FRAP, AR
domain interactions and mobility in living human hepatoma
Hep3B cells (Fig. 1A). Western blot analysis showed that all
tagged ARs were of the expected size (Fig. 1B). All tagged ARs
were able to induce expression of a luciferase (Luc) reporter gene
driven by an androgen-regulated promoter, (ARE)2-TATA Luc
(supplementary material Fig. S1) (van Royen et al., 2007). In the
absence of hormone, double-tagged YFP–AR–CFP and single-
tagged YFP–AR and AR–CFP were mainly located in the
cytoplasm (Fig. 1C–E respectively, left panels). Upon hormone
addition, single- and double-tagged ARs rapidly translocated to
the nucleus (supplementary material Fig. S2). In the nucleus the
ARs were distributed in a typical punctate pattern (Fig. 1C–E
respectively, right panels). This punctate distribution pattern
correlates with a transient immobilization of the AR and partially
overlaps with sites of active transcription (Farla et al., 2005; van
Royen et al., 2007).
We then analyzed cells expressing either double-tagged or
single-tagged ARs by acceptor bleaching FRET (abFRET;
Fig. 1F). In abFRET, the relative increase of the donor
emission after acceptor photobleaching is a measure of the
interaction between the tagged molecules under surveillance
(Bastiaens and Jovin, 1996; Bastiaens et al., 1996; Karpova et al.,
2003; Kenworthy, 2001; van Royen et al., 2009a). In the presence
of the synthetic androgen, R1881, cells expressing double-tagged
AR and cells expressing a combination of single-tagged YFP–AR
and AR–CFP showed abFRET. FRET was not detected in the
nucleus or cytoplasm in the absence of hormone (supplementary
material Fig. S4). AbFRET was strongly reduced in double-
tagged N/C-interaction-deficient mutant ARs, in which the N-
terminal FQNLF motif was mutated to AQNAA (AR F23,27A/
L26A), and completely lost in single-tagged N/C-interaction-
deficient AR mutants (Fig. 1F). These results show that using
abFRET it is possible to quantitatively study inter- and
intramolecular N/C interactions in double-tagged ARs and
intermolecular N/C interaction specifically between single-
tagged ARs.
The AR N/C interaction is predominantly intermolecular
Confocal time-lapse microscopy showed that FRET, as measured
by the YFP/CFP ratio in double-tagged AR (YFP–AR–CFP)-
expressing cells increased rapidly after ligand addition, prior to
translocation to the nucleus (supplementary material Fig. S2A)
(Schaufele et al., 2005). By contrast, in cells expressing two
different single-tagged ARs (YFP–AR and AR–CFP) the YFP/
Fig. 1. Properties of double- and single-tagged
ARs. (A) Schematic representation of YFP–AR–
CFP, YFP–AR and AR–CFP [horizontal bars
represent a (Gly-Ala)6 spacer]. (B) Western blot of
the fusion proteins expressed in Hep3B cells. Lane 1:
YFP–AR–CFP; lane 2: YFP–AR (F23,27A/L26A)–
CFP; lane 3: YFP–AR; lane 4: YFP–AR (F23,27A/
L26A); lane 5: AR–CFP; lane 6: AR (F23,27A/
L26A)–CFP; lane 7: non-transfected cells. AR was
detected using mouse AR monoclonal antibody
(F39.4.1). (C–E) Confocal images of Hep3B cells
expressing double- and single-tagged AR (YFP–AR–
CFP, YFP–AR and AR–CFP) in the absence (left
panels) and presence (right panels) of 100 nM R1881.
Scale bars: 5 mm. (F) Acceptor photobleaching
FRET (abFRET) shows an interaction between the
N-terminal FQNLF motif and the AR-LBD (N/C
interaction) in YFP–AR–CFP and between YFP–AR
and AR–CFP. The N/C interaction is hormone
(100 nM R1881) induced (*P50.001610213 and
**P50.00961022, respectively, see Materials and
Methods) and dependent on the N-terminal FQNLF
motif (#P50.007610210 and ##P50.00761025,
respectively). Values are means 6 26standard error
of the mean (s.e.m.) of at least 25 cells measured in
two independent experiments.
AR intra- and inter-molecular domain interactions 1971
Journ
alof
Cell
Scie
nce
CFP ratio only increased following translocation of the tagged
ARs to the nucleus (supplementary material Fig. S2B). These
observations indicate that intramolecular N/C interactions were
initiated rapidly after hormone binding, followed by nuclear
translocation and initiation of intermolecular N/C interactions
(see also Schaufele et al., 2005).
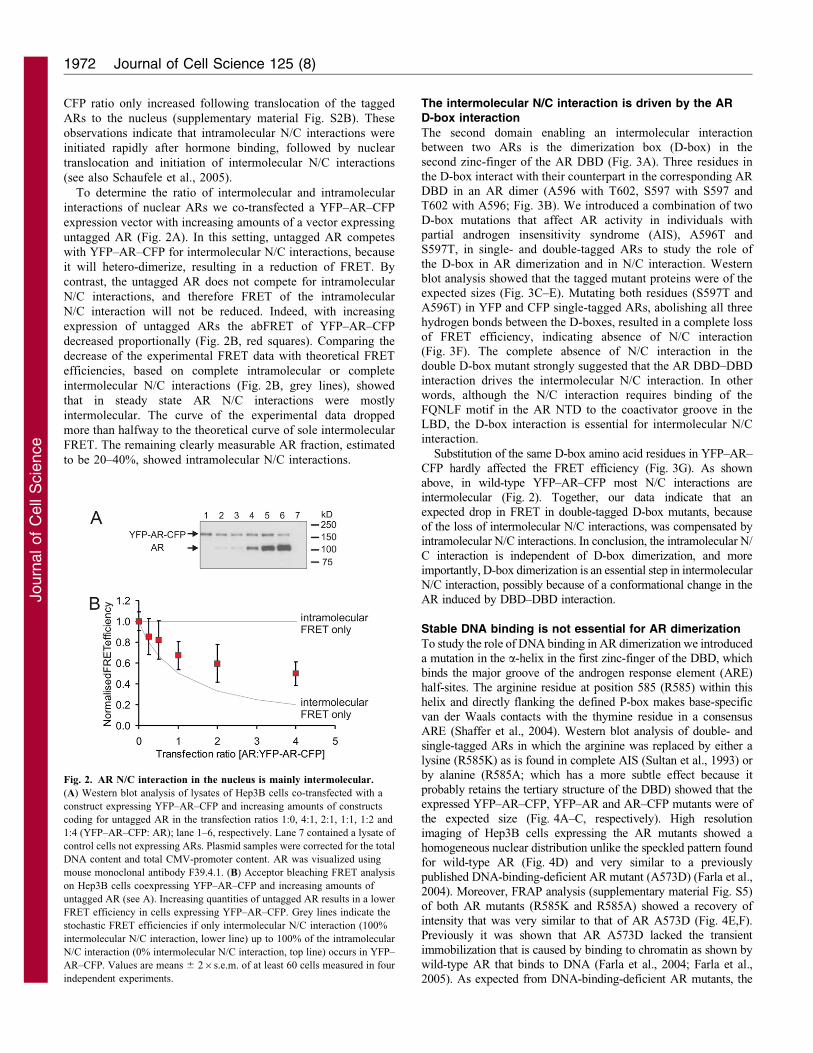
To determine the ratio of intermolecular and intramolecular
interactions of nuclear ARs we co-transfected a YFP–AR–CFP
expression vector with increasing amounts of a vector expressing
untagged AR (Fig. 2A). In this setting, untagged AR competes
with YFP–AR–CFP for intermolecular N/C interactions, because
it will hetero-dimerize, resulting in a reduction of FRET. By
contrast, the untagged AR does not compete for intramolecular
N/C interactions, and therefore FRET of the intramolecular
N/C interaction will not be reduced. Indeed, with increasing
expression of untagged ARs the abFRET of YFP–AR–CFP
decreased proportionally (Fig. 2B, red squares). Comparing the
decrease of the experimental FRET data with theoretical FRET
efficiencies, based on complete intramolecular or complete
intermolecular N/C interactions (Fig. 2B, grey lines), showed
that in steady state AR N/C interactions were mostly
intermolecular. The curve of the experimental data dropped
more than halfway to the theoretical curve of sole intermolecular
FRET. The remaining clearly measurable AR fraction, estimated
to be 20–40%, showed intramolecular N/C interactions.
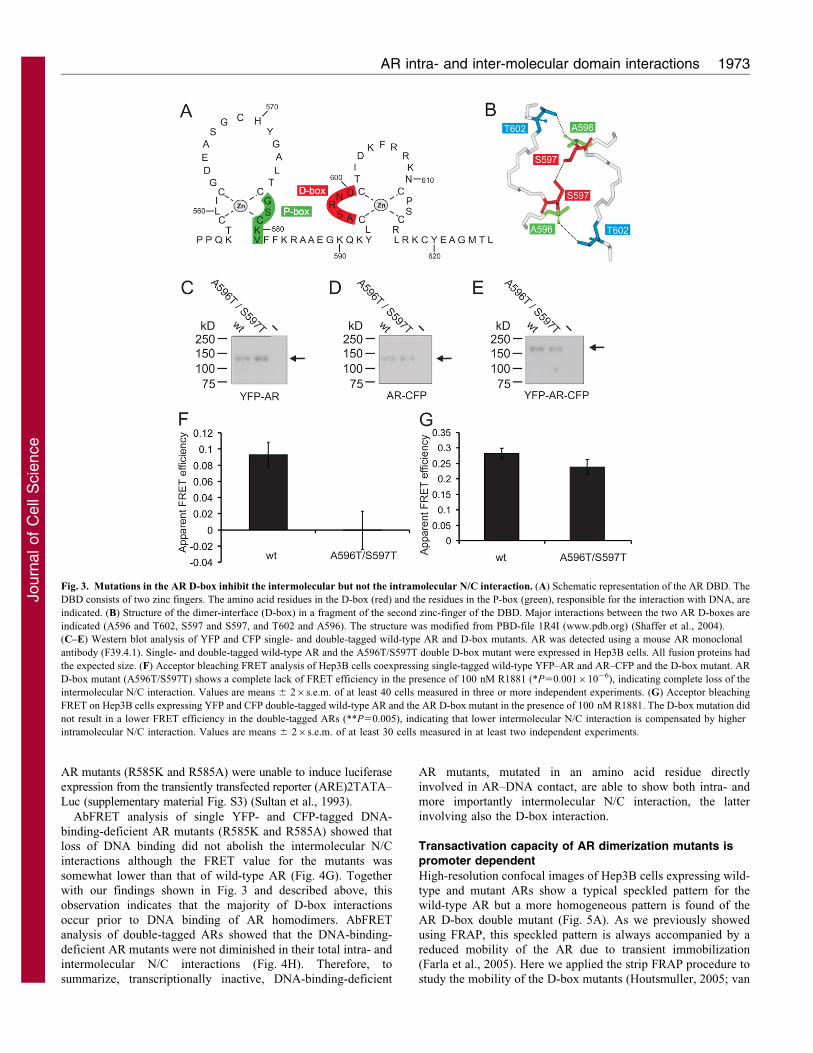
The intermolecular N/C interaction is driven by the ARD-box interaction
The second domain enabling an intermolecular interactionbetween two ARs is the dimerization box (D-box) in thesecond zinc-finger of the AR DBD (Fig. 3A). Three residues in
the D-box interact with their counterpart in the corresponding ARDBD in an AR dimer (A596 with T602, S597 with S597 andT602 with A596; Fig. 3B). We introduced a combination of two
D-box mutations that affect AR activity in individuals withpartial androgen insensitivity syndrome (AIS), A596T andS597T, in single- and double-tagged ARs to study the role of
the D-box in AR dimerization and in N/C interaction. Westernblot analysis showed that the tagged mutant proteins were of theexpected sizes (Fig. 3C–E). Mutating both residues (S597T andA596T) in YFP and CFP single-tagged ARs, abolishing all three
hydrogen bonds between the D-boxes, resulted in a complete lossof FRET efficiency, indicating absence of N/C interaction(Fig. 3F). The complete absence of N/C interaction in the
double D-box mutant strongly suggested that the AR DBD–DBDinteraction drives the intermolecular N/C interaction. In otherwords, although the N/C interaction requires binding of the
FQNLF motif in the AR NTD to the coactivator groove in theLBD, the D-box interaction is essential for intermolecular N/Cinteraction.
Substitution of the same D-box amino acid residues in YFP–AR–
CFP hardly affected the FRET efficiency (Fig. 3G). As shownabove, in wild-type YFP–AR–CFP most N/C interactions areintermolecular (Fig. 2). Together, our data indicate that an
expected drop in FRET in double-tagged D-box mutants, becauseof the loss of intermolecular N/C interactions, was compensated byintramolecular N/C interactions. In conclusion, the intramolecular N/
C interaction is independent of D-box dimerization, and moreimportantly, D-box dimerization is an essential step in intermolecularN/C interaction, possibly because of a conformational change in theAR induced by DBD–DBD interaction.
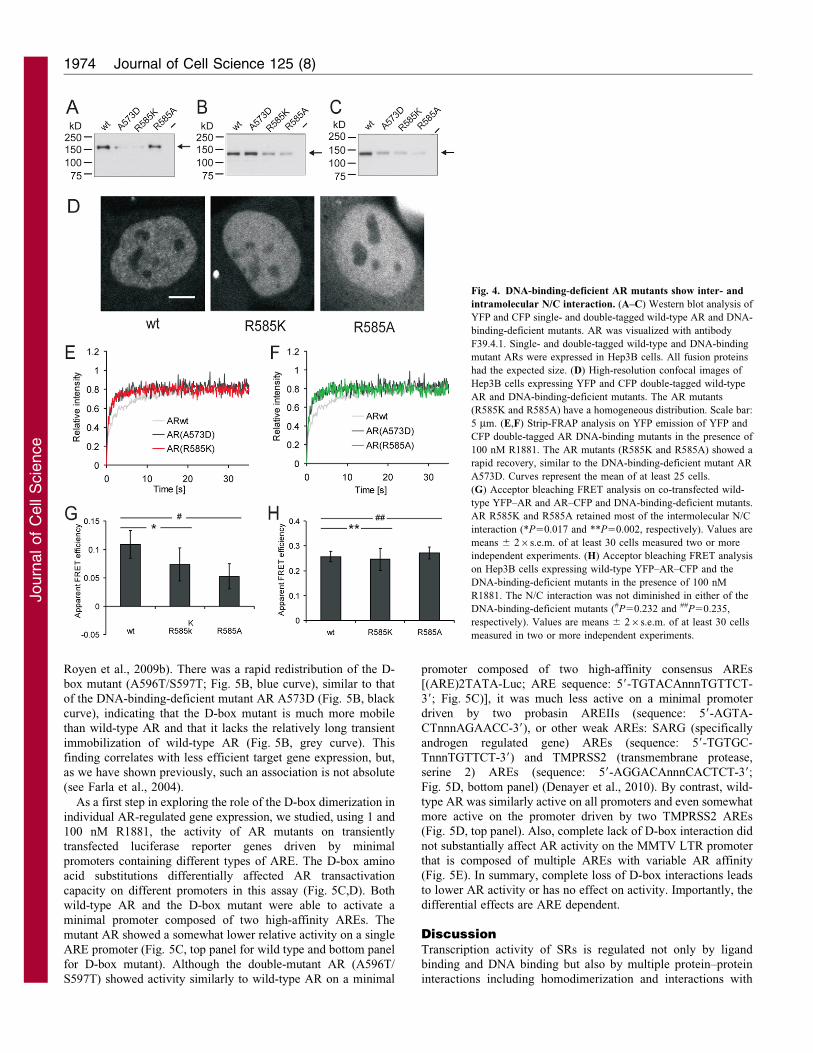
Stable DNA binding is not essential for AR dimerization
To study the role of DNA binding in AR dimerization we introduceda mutation in the a-helix in the first zinc-finger of the DBD, which
binds the major groove of the androgen response element (ARE)half-sites. The arginine residue at position 585 (R585) within thishelix and directly flanking the defined P-box makes base-specific
van der Waals contacts with the thymine residue in a consensusARE (Shaffer et al., 2004). Western blot analysis of double- andsingle-tagged ARs in which the arginine was replaced by either a
lysine (R585K) as is found in complete AIS (Sultan et al., 1993) orby alanine (R585A; which has a more subtle effect because itprobably retains the tertiary structure of the DBD) showed that theexpressed YFP–AR–CFP, YFP–AR and AR–CFP mutants were of
the expected size (Fig. 4A–C, respectively). High resolutionimaging of Hep3B cells expressing the AR mutants showed ahomogeneous nuclear distribution unlike the speckled pattern found
for wild-type AR (Fig. 4D) and very similar to a previouslypublished DNA-binding-deficient AR mutant (A573D) (Farla et al.,2004). Moreover, FRAP analysis (supplementary material Fig. S5)
of both AR mutants (R585K and R585A) showed a recovery ofintensity that was very similar to that of AR A573D (Fig. 4E,F).Previously it was shown that AR A573D lacked the transient
immobilization that is caused by binding to chromatin as shown bywild-type AR that binds to DNA (Farla et al., 2004; Farla et al.,2005). As expected from DNA-binding-deficient AR mutants, the
Fig. 2. AR N/C interaction in the nucleus is mainly intermolecular.
(A) Western blot analysis of lysates of Hep3B cells co-transfected with a
construct expressing YFP–AR–CFP and increasing amounts of constructs
coding for untagged AR in the transfection ratios 1:0, 4:1, 2:1, 1:1, 1:2 and
1:4 (YFP–AR–CFP: AR); lane 1–6, respectively. Lane 7 contained a lysate of
control cells not expressing ARs. Plasmid samples were corrected for the total
DNA content and total CMV-promoter content. AR was visualized using
mouse monoclonal antibody F39.4.1. (B) Acceptor bleaching FRET analysis
on Hep3B cells coexpressing YFP–AR–CFP and increasing amounts of
untagged AR (see A). Increasing quantities of untagged AR results in a lower
FRET efficiency in cells expressing YFP–AR–CFP. Grey lines indicate the
stochastic FRET efficiencies if only intermolecular N/C interaction (100%
intermolecular N/C interaction, lower line) up to 100% of the intramolecular
N/C interaction (0% intermolecular N/C interaction, top line) occurs in YFP–
AR–CFP. Values are means 6 26s.e.m. of at least 60 cells measured in four
independent experiments.
Journal of Cell Science 125 (8)1972
Journ
alof
Cell
Scie
nce
AR mutants (R585K and R585A) were unable to induce luciferase
expression from the transiently transfected reporter (ARE)2TATA–
Luc (supplementary material Fig. S3) (Sultan et al., 1993).
AbFRET analysis of single YFP- and CFP-tagged DNA-
binding-deficient AR mutants (R585K and R585A) showed that
loss of DNA binding did not abolish the intermolecular N/C
interactions although the FRET value for the mutants was
somewhat lower than that of wild-type AR (Fig. 4G). Together
with our findings shown in Fig. 3 and described above, this
observation indicates that the majority of D-box interactions
occur prior to DNA binding of AR homodimers. AbFRET
analysis of double-tagged ARs showed that the DNA-binding-
deficient AR mutants were not diminished in their total intra- and
intermolecular N/C interactions (Fig. 4H). Therefore, to
summarize, transcriptionally inactive, DNA-binding-deficient
AR mutants, mutated in an amino acid residue directly
involved in AR–DNA contact, are able to show both intra- and
more importantly intermolecular N/C interaction, the latter
involving also the D-box interaction.
Transactivation capacity of AR dimerization mutants ispromoter dependent
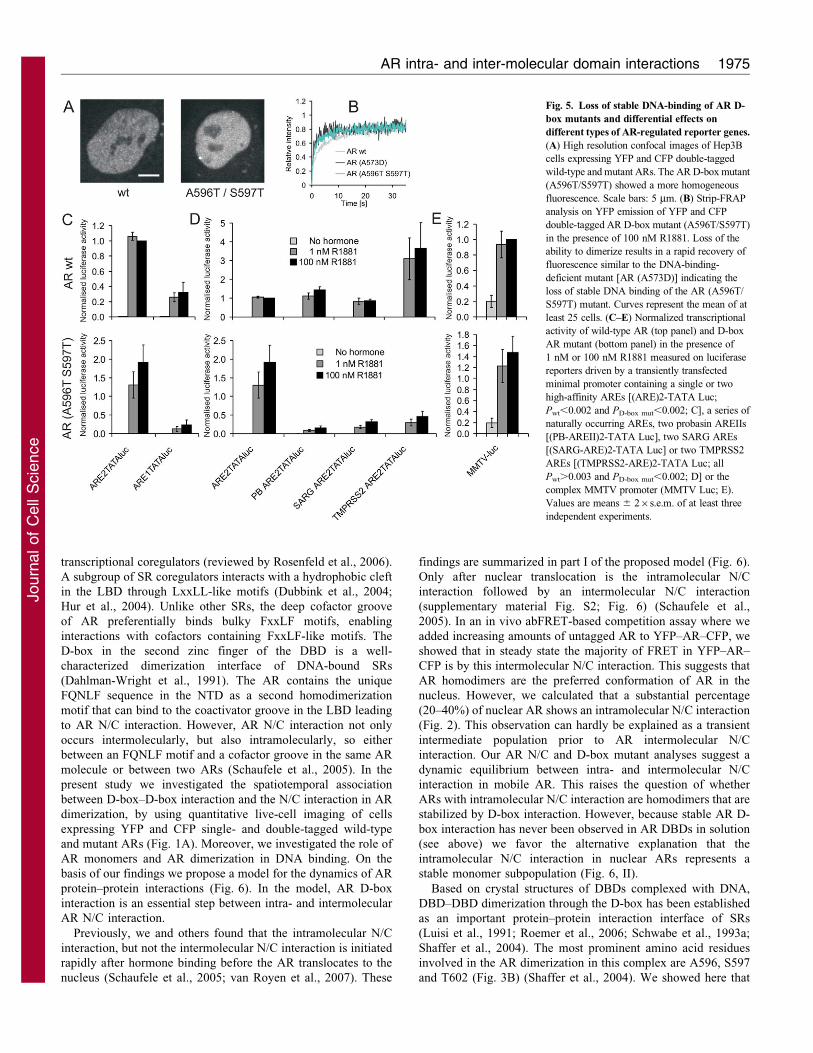
High-resolution confocal images of Hep3B cells expressing wild-
type and mutant ARs show a typical speckled pattern for the
wild-type AR but a more homogeneous pattern is found of the
AR D-box double mutant (Fig. 5A). As we previously showed
using FRAP, this speckled pattern is always accompanied by a
reduced mobility of the AR due to transient immobilization
(Farla et al., 2005). Here we applied the strip FRAP procedure to
study the mobility of the D-box mutants (Houtsmuller, 2005; van
Fig. 3. Mutations in the AR D-box inhibit the intermolecular but not the intramolecular N/C interaction. (A) Schematic representation of the AR DBD. The
DBD consists of two zinc fingers. The amino acid residues in the D-box (red) and the residues in the P-box (green), responsible for the interaction with DNA, are
indicated. (B) Structure of the dimer-interface (D-box) in a fragment of the second zinc-finger of the DBD. Major interactions between the two AR D-boxes are
indicated (A596 and T602, S597 and S597, and T602 and A596). The structure was modified from PBD-file 1R4I (www.pdb.org) (Shaffer et al., 2004).
(C–E) Western blot analysis of YFP and CFP single- and double-tagged wild-type AR and D-box mutants. AR was detected using a mouse AR monoclonal
antibody (F39.4.1). Single- and double-tagged wild-type AR and the A596T/S597T double D-box mutant were expressed in Hep3B cells. All fusion proteins had
the expected size. (F) Acceptor bleaching FRET analysis of Hep3B cells coexpressing single-tagged wild-type YFP–AR and AR–CFP and the D-box mutant. AR
D-box mutant (A596T/S597T) shows a complete lack of FRET efficiency in the presence of 100 nM R1881 (*P50.00161026), indicating complete loss of the
intermolecular N/C interaction. Values are means 6 26s.e.m. of at least 40 cells measured in three or more independent experiments. (G) Acceptor bleaching
FRET on Hep3B cells expressing YFP and CFP double-tagged wild-type AR and the AR D-box mutant in the presence of 100 nM R1881. The D-box mutation did
not result in a lower FRET efficiency in the double-tagged ARs (**P50.005), indicating that lower intermolecular N/C interaction is compensated by higher
intramolecular N/C interaction. Values are means 6 26s.e.m. of at least 30 cells measured in at least two independent experiments.
AR intra- and inter-molecular domain interactions 1973
Journ
alof
Cell
Scie
nce
Royen et al., 2009b). There was a rapid redistribution of the D-
box mutant (A596T/S597T; Fig. 5B, blue curve), similar to that
of the DNA-binding-deficient mutant AR A573D (Fig. 5B, black
curve), indicating that the D-box mutant is much more mobile
than wild-type AR and that it lacks the relatively long transient
immobilization of wild-type AR (Fig. 5B, grey curve). This
finding correlates with less efficient target gene expression, but,
as we have shown previously, such an association is not absolute
(see Farla et al., 2004).
As a first step in exploring the role of the D-box dimerization in
individual AR-regulated gene expression, we studied, using 1 and
100 nM R1881, the activity of AR mutants on transiently
transfected luciferase reporter genes driven by minimal
promoters containing different types of ARE. The D-box amino
acid substitutions differentially affected AR transactivation
capacity on different promoters in this assay (Fig. 5C,D). Both
wild-type AR and the D-box mutant were able to activate a
minimal promoter composed of two high-affinity AREs. The
mutant AR showed a somewhat lower relative activity on a single
ARE promoter (Fig. 5C, top panel for wild type and bottom panel
for D-box mutant). Although the double-mutant AR (A596T/
S597T) showed activity similarly to wild-type AR on a minimal
promoter composed of two high-affinity consensus AREs
[(ARE)2TATA-Luc; ARE sequence: 59-TGTACAnnnTGTTCT-
39; Fig. 5C)], it was much less active on a minimal promoter
driven by two probasin AREIIs (sequence: 59-AGTA-
CTnnnAGAACC-39), or other weak AREs: SARG (specifically
androgen regulated gene) AREs (sequence: 59-TGTGC-
TnnnTGTTCT-39) and TMPRSS2 (transmembrane protease,
serine 2) AREs (sequence: 59-AGGACAnnnCACTCT-39;
Fig. 5D, bottom panel) (Denayer et al., 2010). By contrast, wild-
type AR was similarly active on all promoters and even somewhat
more active on the promoter driven by two TMPRSS2 AREs
(Fig. 5D, top panel). Also, complete lack of D-box interaction did
not substantially affect AR activity on the MMTV LTR promoter
that is composed of multiple AREs with variable AR affinity
(Fig. 5E). In summary, complete loss of D-box interactions leads
to lower AR activity or has no effect on activity. Importantly, the
differential effects are ARE dependent.
DiscussionTranscription activity of SRs is regulated not only by ligand
binding and DNA binding but also by multiple protein–protein
interactions including homodimerization and interactions with
Fig. 4. DNA-binding-deficient AR mutants show inter- and
intramolecular N/C interaction. (A–C) Western blot analysis of
YFP and CFP single- and double-tagged wild-type AR and DNA-
binding-deficient mutants. AR was visualized with antibody
F39.4.1. Single- and double-tagged wild-type and DNA-binding
mutant ARs were expressed in Hep3B cells. All fusion proteins
had the expected size. (D) High-resolution confocal images of
Hep3B cells expressing YFP and CFP double-tagged wild-type
AR and DNA-binding-deficient mutants. The AR mutants
(R585K and R585A) have a homogeneous distribution. Scale bar:
5 mm. (E,F) Strip-FRAP analysis on YFP emission of YFP and
CFP double-tagged AR DNA-binding mutants in the presence of
100 nM R1881. The AR mutants (R585K and R585A) showed a
rapid recovery, similar to the DNA-binding-deficient mutant AR
A573D. Curves represent the mean of at least 25 cells.
(G) Acceptor bleaching FRET analysis on co-transfected wild-
type YFP–AR and AR–CFP and DNA-binding-deficient mutants.
AR R585K and R585A retained most of the intermolecular N/C
interaction (*P50.017 and **P50.002, respectively). Values are
means 6 26s.e.m. of at least 30 cells measured two or more
independent experiments. (H) Acceptor bleaching FRET analysis
on Hep3B cells expressing wild-type YFP–AR–CFP and the
DNA-binding-deficient mutants in the presence of 100 nM
R1881. The N/C interaction was not diminished in either of the
DNA-binding-deficient mutants (#P50.232 and ##P50.235,
respectively). Values are means 6 26s.e.m. of at least 30 cells
measured in two or more independent experiments.
Journal of Cell Science 125 (8)1974
Journ
alof
Cell
Scie
nce
transcriptional coregulators (reviewed by Rosenfeld et al., 2006).
A subgroup of SR coregulators interacts with a hydrophobic cleft
in the LBD through LxxLL-like motifs (Dubbink et al., 2004;
Hur et al., 2004). Unlike other SRs, the deep cofactor groove
of AR preferentially binds bulky FxxLF motifs, enabling
interactions with cofactors containing FxxLF-like motifs. The
D-box in the second zinc finger of the DBD is a well-
characterized dimerization interface of DNA-bound SRs
(Dahlman-Wright et al., 1991). The AR contains the unique
FQNLF sequence in the NTD as a second homodimerization
motif that can bind to the coactivator groove in the LBD leading
to AR N/C interaction. However, AR N/C interaction not only
occurs intermolecularly, but also intramolecularly, so either
between an FQNLF motif and a cofactor groove in the same AR
molecule or between two ARs (Schaufele et al., 2005). In the
present study we investigated the spatiotemporal association
between D-box–D-box interaction and the N/C interaction in AR
dimerization, by using quantitative live-cell imaging of cells
expressing YFP and CFP single- and double-tagged wild-type
and mutant ARs (Fig. 1A). Moreover, we investigated the role of
AR monomers and AR dimerization in DNA binding. On the
basis of our findings we propose a model for the dynamics of AR
protein–protein interactions (Fig. 6). In the model, AR D-box
interaction is an essential step between intra- and intermolecular
AR N/C interaction.
Previously, we and others found that the intramolecular N/C
interaction, but not the intermolecular N/C interaction is initiated
rapidly after hormone binding before the AR translocates to the
nucleus (Schaufele et al., 2005; van Royen et al., 2007). These
findings are summarized in part I of the proposed model (Fig. 6).
Only after nuclear translocation is the intramolecular N/C
interaction followed by an intermolecular N/C interaction
(supplementary material Fig. S2; Fig. 6) (Schaufele et al.,
2005). In an in vivo abFRET-based competition assay where we
added increasing amounts of untagged AR to YFP–AR–CFP, we
showed that in steady state the majority of FRET in YFP–AR–
CFP is by this intermolecular N/C interaction. This suggests that
AR homodimers are the preferred conformation of AR in the
nucleus. However, we calculated that a substantial percentage
(20–40%) of nuclear AR shows an intramolecular N/C interaction
(Fig. 2). This observation can hardly be explained as a transient
intermediate population prior to AR intermolecular N/C
interaction. Our AR N/C and D-box mutant analyses suggest a
dynamic equilibrium between intra- and intermolecular N/C
interaction in mobile AR. This raises the question of whether
ARs with intramolecular N/C interaction are homodimers that are
stabilized by D-box interaction. However, because stable AR D-
box interaction has never been observed in AR DBDs in solution
(see above) we favor the alternative explanation that the
intramolecular N/C interaction in nuclear ARs represents a
stable monomer subpopulation (Fig. 6, II).
Based on crystal structures of DBDs complexed with DNA,
DBD–DBD dimerization through the D-box has been established
as an important protein–protein interaction interface of SRs
(Luisi et al., 1991; Roemer et al., 2006; Schwabe et al., 1993a;
Shaffer et al., 2004). The most prominent amino acid residues
involved in the AR dimerization in this complex are A596, S597
and T602 (Fig. 3B) (Shaffer et al., 2004). We showed here that
Fig. 5. Loss of stable DNA-binding of AR D-
box mutants and differential effects on
different types of AR-regulated reporter genes.
(A) High resolution confocal images of Hep3B
cells expressing YFP and CFP double-tagged
wild-type and mutant ARs. The AR D-box mutant
(A596T/S597T) showed a more homogeneous
fluorescence. Scale bars: 5 mm. (B) Strip-FRAP
analysis on YFP emission of YFP and CFP
double-tagged AR D-box mutant (A596T/S597T)
in the presence of 100 nM R1881. Loss of the
ability to dimerize results in a rapid recovery of
fluorescence similar to the DNA-binding-
deficient mutant [AR (A573D)] indicating the
loss of stable DNA binding of the AR (A596T/
S597T) mutant. Curves represent the mean of at
least 25 cells. (C–E) Normalized transcriptional
activity of wild-type AR (top panel) and D-box
AR mutant (bottom panel) in the presence of
1 nM or 100 nM R1881 measured on luciferase
reporters driven by a transiently transfected
minimal promoter containing a single or two
high-affinity AREs [(ARE)2-TATA Luc;
Pwt,0.002 and PD-box mut,0.002; C], a series of
naturally occurring AREs, two probasin AREIIs
[(PB-AREII)2-TATA Luc], two SARG AREs
[(SARG-ARE)2-TATA Luc] or two TMPRSS2
AREs [(TMPRSS2-ARE)2-TATA Luc; all
Pwt.0.003 and PD-box mut,0.002; D] or the
complex MMTV promoter (MMTV Luc; E).
Values are means 6 26s.e.m. of at least three
independent experiments.
AR intra- and inter-molecular domain interactions 1975
Journ
alof
Cell
Scie
nce

mutation of two of these three amino acid residues completely
abolished intermolecular N/C interaction, most probably because
of complete absence of the D-box interaction. The mutations had
no effect on intramolecular N/C interaction. In fact, in the
absence of intermolecular N/C interaction in D-box mutants, an
increased intramolecular N/C interaction was observed
(Fig. 3F,G). These findings strongly suggest that D-box to D-
box interactions drive the transition from intramolecular AR N/C
interaction to intermolecular N/C interaction in nuclear AR
(Fig. 6, III).
It is not known whether peptide motif interactions other than
D-box interactions and N/C interactions can play a prominent
role in AR dimerization. For AR the evidence for LBD–LBD
interactions, as documented for other SRs, is limited. Although
amino acid residues involved in glucocorticoid receptor (GR)
LBD–LBD interactions are conserved in AR (Centenera et al.,
2008), in crystallographic studies, the isolated AR LBD is present
as a monomer in solution, in contrast to GR, progesterone
receptor (PR) and estrogen receptor (ER) LBDs (Bledsoe et al.,
2002; Matias et al., 2000; Sack et al., 2001; Tanenbaum et al.,
1998; Williams and Sigler, 1998). However, a (weak)
dimerization function in the hinge region as suggested for GR,
or in the C-terminal extension of the AR DBD cannot be
completely excluded (Centenera et al., 2008; Haelens et al., 2003;
Savory et al., 2001).
It has long been disputed whether AR dimerization occurs
before or after DNA binding (Centenera et al., 2008). We
previously showed that the N/C interaction occurs predominantly
when the ARs are mobile and is lost when the ARs are bound to
chromatin (van Royen et al., 2007). Combined with the present
observation that D-box interaction drives the intermolecular N/C
interaction (Fig. 3) this indicates that the D-box interaction
occurs before DNA binding (Fig. 6). These findings are in
contrast to theories based on crystallographic studies which
suggest that separate SR DBDs are monomeric in solution and
show cooperative dimerization when bound to DNA (Freedman
et al., 1988; Hard et al., 1990a; Hard et al., 1990b; Luisi et al.,
1991; Schwabe et al., 1993a; Schwabe et al., 1993b; Shaffer et al.,
2004). However, AR dimerization before DNA binding was
confirmed by experiments carried out with the DNA-binding-
deficient mutants (Fig. 4). Possibly, the stronger D-box to D-box
interaction in AR, compared with other SRs, combined with the
intramolecular N/C interaction, are of crucial importance in this
regard.
The relatively strong dimerization of the AR enables activation
of promoters containing different types of ARE (Fig. 5)
(reviewed by Centenera et al., 2008; Claessens et al., 2008;
Denayer et al., 2010; Shaffer et al., 2004). We showed that ARs
without appropriate D-box interaction cannot activate promoters
driven by two probasin AREIIs, SARG AREs or TMPRSS2
AREs, although a promoter with high affinity AREs can be
stimulated (Fig. 5C–E) (Denayer et al., 2010). On the basis of our
findings it is tempting to speculate that promoters with high
affinity AREs can be activated both by AR homodimers and by
consecutive binding of AR monomers that subsequently dimerize
on the DNA. By contrast, promoters with low-affinity AREs
would preferentially be activated by AR homodimers (Fig. 6,
IV). If this is the case then AR monomers in the nucleus are of
functional importance.
Recently, genome-wide chromatin immunoprecipitation
(ChIP) approaches indicated the presence of thousands or even
tens of thousands of AR binding sites in the human genome (Jia
et al., 2008; Takayama et al., 2011; Wang et al., 2007; Wang
et al., 2009; Yu et al., 2010). Interestingly, the majority of the AR
binding regions found in these studies, and AR binding sites
identified by ChIP in promoter and enhancer regions of
Fig. 6. Schematic representation of the spatiotemporal organization
of domain interactions in AR function. In the absence of ligand the AR
is predominantly located in the cytoplasm. Hormone-binding by the AR
results in a rapid initiation of the intramolecular N/C interaction followed
by nuclear translocation (I). In the nucleus both AR monomers and
dimers form a stable population (II), but the majority of ARs dimerize
through the D-box interaction that drives a transition from intra- to
intermolecular N/C interaction (III). The indicated intermediate
conformations, the AR homodimer with intramolecular N/C interaction
or without the N/C interaction, might be rapidly processed. The AR
dimer is able to stably bind to either a high-affinity ARE (light-green
rectangles) or a more selective low-affinity ARE (dark-green rectangles)
in promoter or enhancer regions of target genes. Two AR monomers are
proposed to bind consecutively to a high-affinity ARE where they
dimerize, but not to a low-affinity ARE (IV). In the DNA-bound AR the
N/C interaction is lost, allowing interactions with coactivators to
initiate transcription.
Journal of Cell Science 125 (8)1976
Journ
alof
Cell
Scie
nce

androgen-regulated genes, apparently contain ARE half-sitemotifs, low-affinity AREs or ARE half-sites with suboptimal
spacing and not obvious high-affinity AR binding motifs (Massieet al., 2007; Wang et al., 2007). Loss of AR dimerization mightdirectly result in less AR binding to these sites (Fig. 5B).
Genome-wide ChIP-seq approaches combined with global geneexpression profiling in cells that exclusively express AR
monomers compared with cells that contains AR mainly in thehomodimer conformation would provide more detailedinformation on the role of monomers in AR-regulated gene
expression. The AR monomer to dimer ratio in a nucleus mightbe a mechanism of regulation of specificity in gene expression.
One obvious parameter that affects the dimerization status of theactivated nuclear AR population, is its concentration. Thishypothesis can well be extended to a role for specific cofactors
that differentially interact with AR monomers and dimers orcofactors that regulate AR dimerization (Bai et al., 2005).
In summary, previous data on the AR intra- and intermolecularN/C interaction lead to a model in which the intramolecular N/Cinteraction is initiated in the cytoplasm directly after hormone
binding, followed by intermolecular N/C interaction in the nucleus(Schaufele et al., 2005). Using quantitative imaging techniques, weelucidated the essential role of D-box dimerization in the transition
from intramolecular to intermolecular N/C interaction (Fig. 6).The D-box dimerization and the shift from intramolecular to
intermolecular N/C interaction might occur as one event or twoseparate events, but both independent of DNA binding. Togetherwith our observations showing that the AR N/C interaction is lost
in DNA-bound AR enabling cofactor interactions (van Royen et al.,2007), data in the present study elucidated the spatiotemporal
relationship of the consecutive AR intra- and intermoleculardomain interactions in living cells (Fig. 6). Moreover, the modelproposes a dynamic equilibrium of AR homodimers and
monomers in the nucleus, which can be an important mechanismof AR-regulated gene expression.
Materials and MethodsConstructs
In all constructs expressing AR fusion proteins the AR was separated from thefluorescent tag by a flexible (Gly-Ala)6 spacer (Farla et al., 2004) indicated by asingle dash. Constructs coding for wild-type and A573D variants of YFP–AR–CFPand AR–CFP were generated as previously described (van Royen et al., 2007). Theconstruct expressing N-terminally YFP-tagged AR was generated by replacingEGFP in pGFP-AR (Farla et al., 2004) with EYFP-C1 (Clontech Laboratories, Inc.,Mountain View, CA). The construct expressing untagged AR was obtained byinserting the AR cDNA from pAR0 (Brinkmann et al., 1989) into pEGFP-C1 fromwhich EGFP was deleted. The F23,27A/L26A mutation of YFP–AR–CFP, AR–CFP and untagged AR was introduced using the QuikChange mutagenesis kit(Stratagene, La Jolla, CA). In YFP–AR–CFP an LBD–CFP fragment was replacedwith an AR–LBD fragment from YFP–AR to obtain YFP–AR (F23,27A/L26A).The DBD mutations R585K, R585A and A596T/S597T were introduced byQuikChange mutagenesis in pYFP-AR-CFP. The LBD mutation E897A inuntagged AR was also generated with QuikChange mutagenesis. Formutagenesis primers see supplementary material Table S1. To generate thesingle-tagged DBD mutant ARs, the AR DBDs of pYFP-AR and pAR-CFP werereplaced with a pYFP-AR-CFP fragment containing the mutant DBD.
The (ARE)2-TATA Luc reporter, containing two high-affinity AREs(underlined in the following sequence: 59-CCGGGAGCTTGTACAGGATG-TTCTGCATGCTCTAGATGTACAGGATGTTCTGGTA-39) was a gift from G.Jenster (Rotterdam, Netherlands). The other reporters were generated by swappingthe ARE fragment in (ARE)2-TATA Luc with a fragment containing a single highaffinity ARE as present in the (ARE)2-TATA Luc reporter (59-CCG-GGAGCTTGTACAGGATGTTCTGCATGCTCTAGAGGTA-39), two probasinAREIIs (59-CCGGGAGCTAGTACTGGAAGAACCGCATGCTCTAGAAGTA-CTGGAAGAACCGGTA-39), two SARG AREs (59-CCGGGAGCTTGTGC-TGGATGTTCTGCATGCTCTAGATGTGCTGGATGTTCTGGTA-39), or twoTMPRSS2 AREs (59-CCGGGAGCTAGGACAGGACACTCTGCATGCTCTAG-AAGGACAGGACACTCTGGTA-39). The MMTV-Luc reporter construct was
described previously (de Ruiter et al., 1995). All new constructs were verified bysequencing. Sizes of expressed ARs were verified by western blotting.
Cell culture, transfection and luciferase assay
For 2 days before microscopic analyses, Hep3B cells, lacking endogenous ARexpression, were grown on glass coverslips in six-well plates in a-MEM(Cambrex, East Rutherford, NJ) supplemented with 5% fetal bovine serum(FBS; HyClone), 2 mM L-glutamine, 100 IU/ml penicillin and 100 mg/mlstreptomycin. At least 4 hours before transfection, the medium was replacedwith medium containing FBS stripped with 5% dextran-coated charcoal (DCC-FBS). Transfections were performed with 1 mg/well AR expression constructs or0.5 mg/well empty YFP or CFP expression vector in FuGENE6 (Roche MolecularBiochemicals, Indianapolis, IN) transfection medium. Four hours aftertransfection, the medium was replaced with 5% DCC-FBS with or without100 nM R1881. In the abFRET competition experiments 1 mg YFP–AR–CFP wasco-transfected with increasing amounts of untagged AR (ratio YFP–AR–CFP: AR1:0, 4:1, 2:1, 1:1, 1:2 and 1:4). Different vector sizes were taken into account. Theamounts of CMV promoters and total transfected DNA were corrected by co-transfecting pcDNA3 (CMV) and pTZ19 vectors.
For the AR transactivation experiments, Hep3B cells were cultured in 24-wellplates in a-MEM supplemented with 5% DCC-FBS in the absence or presence ofR1881 (1 or 100 nM) and transfected using 50 ng AR expression construct and100 ng luciferase reporter construct. After 24 hours, cells were lysed andluciferase activity was measured in a luminometer (GloMax Microplateluminometer; Promega Corporation, Madison, WI).
Western blot analysis
Hep3B cells were cultured and transfected in 6-well plates. After 24 hours, cellswere washed twice in ice-cold PBS and lysed in 200 ml Laemmli sample buffer(50 mM Tris-HCl, pH 6.8, 10% glycerol, 2% SDS, 10 mM DTT and 0.001%Bromophenol Blue). After boiling for 5 minutes, a 5-ml sample was separated on a10% SDS-polyacrylamide gel and blotted to Immobilon-P transfer membrane(Millipore, Billerica, MA). Blots were incubated with anti-AR (1:2000; mousemonoclonal antibody F39.4.1) and subsequently incubated with HRP-conjugatedgoat anti-mouse antibody (DakoCytomation, Glostrup, Denmark). Protein bandswere visualized using Super Signal West Pico Luminol solution (Pierce ChemicalCo., Rockford, IL), followed by exposure to x-ray film.
Confocal imaging, YFP/CFP ratio imaging and abFRET analysis
Immunofluorescence imaging of Hep3B cells expressing tagged ARs wasperformed using a confocal laser-scanning microscope (LSM510; Carl ZeissMicroImaging, Inc., Gottingen, Germany) equipped with a Plan-Neofluar 406/1.3NA oil objective (Carl Zeiss MicroImaging, Inc.) at a lateral resolution of 100 nm.An argon laser was used for excitation of CFP and YFP at 458 and 514 nm,respectively. In all quantitative imaging experiments cells with a physiologicallyrelevant expression level of tagged ARs were selected for analysis (van Royenet al., 2007; van Royen et al., 2009a).
N/C interactions of double-tagged YFP–AR–CFP, or co-transfected YFP–ARand AR–CFP were assessed using YFP/CFP ratio imaging and acceptorphotobleaching FRET (abFRET) (van Royen et al., 2009a and referencestherein). In YFP/CFP ratio imaging cells expressing YFP and CFP double-tagged AR or a combination of YFP–AR and AR–CFP with initially similar signalratios to YFP–AR–CFP were imaged with an interval of 30 seconds using a458 nm excitation at low laser power to avoid monitor bleaching. YFP and CFPemissions were detected using a 560 nm longpass emission filter and a 470–500 nm bandpass emission filter, respectively. The AR N/C interaction wasinitiated by adding R1881 to the cell culture. After subtraction of backgroundFRET was calculated as: IYFP/ICFP. The relative nuclear intensity was determinedsimultaneously using the YFP emission and was calculated as: Inucleus/(Inucleus+Icytoplasm).
In abFRET, YFP and CFP images were collected sequentially beforephotobleaching of the acceptor. CFP was excited at 458 nm at moderate laserpower, and emission was detected using a 470–500 nm bandpass emission filter.YFP was excited at 514 nm at moderate laser power, and emission was detectedusing a 560 nm longpass emission filter. After image collection, YFP in the nucleuswas bleached by scanning a region of ,100 mm2 25 times at 514 nm at high laserpower, covering almost the complete nucleus. After photobleaching, a secondYFP and CFP image pair was collected. Apparent FRET efficiency wasestimated (correcting for the amount of YFP bleached) using the equationabFRET5[(CFPafter2CFPbefore)6YFPbefore]6[(CFPafter6YFPbefore)2 (CFPbefore6YFPafter)]
21, where CFPbefore and YFPbefore are the mean prebleach fluorescenceintensities of CFP and YFP, respectively, in the area to be bleached (after subtractionof background), and CFPafter and YFPafter are the mean postbleach fluorescenceintensities of CFP and YFP, respectively, in the bleached area (Dinant et al., 2008).The apparent FRET efficiency was finally expressed relative to control measurementsin cells expressing either free CFP and YFP (abFRET0) or the CFP–YFP fusionprotein (abFRETCFP–YFP fusion): apparent FRET efficiency5(abFRET2abFRET0)6
AR intra- and inter-molecular domain interactions 1977
Journ
alof
Cell
Scie
nce

(abFRETCFP-YFP fusion2abFRET0)21. For statistical analysis, the abFRET data sets
were compared using the one-tailed Student’s t-test.
FRAP
The mobility of interacting proteins was studied using FRAP (supplementarymaterial Fig. S5) (van Royen et al., 2009b). A narrow strip spanning the nucleuswas scanned at 458 nm excitation (because of simultaneous CFP recording inFRET FRAP) (van Royen et al., 2007) using short intervals (100 ms) at low laserpower (YFP is sufficiently excited at this wavelength) (van Royen et al., 2007).Fluorescence intensity of YFP was recorded using a 560-nm longpass filter. After40 scans, a high-intensity, 100-ms bleach pulse at 514 nm was applied tophotobleach YFP inside the strip. Subsequently, scanning of the bleached strip wascontinued at 458 nm at low laser intensity. The curves were normalized using theequation Inorm5(Iraw2I0)/(Ipre2I0), where Ipre and I0 are the fluorescent intensitiesbefore and immediately after the bleach, respectively.
FundingThis work was supported by the Dutch Cancer Society [grant numberDDHK 2002-2679 to M.v.R.]; and the European Science Foundation[grant number 03-DYNA-F-18 to M.v.R.].
Supplementary material available online at
http://jcs.biologists.org/lookup/suppl/doi:10.1242/jcs.096792/-/DC1
ReferencesBai, S., He, B. and Wilson, E. M. (2005). Melanoma antigen gene protein MAGE-11
regulates androgen receptor function by modulating the interdomain interaction. Mol.
Cell. Biol. 25, 1238-1257.
Bastiaens, P. I. H. and Jovin, T. M. (1996). Microspectroscopic imaging tracks theintracellular processing of a signal transduction protein: fluorescent-labeled proteinkinase C beta I. Proc. Natl. Acad. Sci. USA 93, 8407-8412.
Bastiaens, P. I. H., Majoul, I. V., Verveer, P. J., Soling, H.-D. and Jovin, T. M.
(1996). Imaging the intracellular trafficking and state of the AB5 quaternary structure
of cholera toxin. EMBO J. 15, 4246-4253.
Bledsoe, R. K., Montana, V. G., Stanley, T. B., Delves, C. J., Apolito, C. J., McKee,
D. D., Consler, T. G., Parks, D. J., Stewart, E. L., Willson, T. M. et al. (2002).Crystal structure of the glucocorticoid receptor ligand binding domain reveals a novelmode of receptor dimerization and coactivator recognition. Cell 110, 93-105.
Brinkmann, A. O., Faber, P. W., van Rooij, H. C. J., Kuiper, G. G. J. M., Ris, C.,
Klaassen, P., van der Korput, J. A. G. M., Voorhorst, M. M., van Laar, J. H.,
Mulder, E. et al. (1989). The human androgen receptor: domain structure, genomicorganization and regulation of expression. J. Steroid Biochem. 34, 307-310.
Centenera, M. M., Harris, J. M., Tilley, W. D. and Butler, L. M. (2008). Thecontribution of different androgen receptor domains to receptor dimerization andsignaling. Mol. Endocrinol. 22, 2373-2382.
Claessens, F., Denayer, S., Van Tilborgh, N., Kerkhofs, S., Helsen, C. and Haelens,
A. (2008). Diverse roles of androgen receptor (AR) domains in AR-mediated
signaling. Nucl. Recept. Signal. 6, e008.
Dahlman-Wright, K., Wright, A., Gustafsson, J. A. and Carlstedt-Duke, J. (1991).
Interaction of the glucocorticoid receptor DNA-binding domain with DNA as a dimeris mediated by a short segment of five amino acids. J. Biol. Chem. 266, 3107-3112.
de Ruiter, P. E., Teuwen, R., Trapman, J., Dijkema, R. and Brinkmann, A. O.
(1995). Synergism between androgens and protein kinase-C on androgen-regulatedgene expression. Mol. Cell. Endocrinol. 110, R1-R6.
Denayer, S., Helsen, C., Thorrez, L., Haelens, A. and Claessens, F. (2010). The rulesof DNA recognition by the androgen receptor. Mol. Endocrinol. 24, 898-913.
Dinant, C., van Royen, M. E., Vermeulen, W. and Houtsmuller, A. B. (2008).Fluorescence resonance energy transfer of GFP and YFP by spectral imaging and
quantitative acceptor photobleaching. J. Microsc. 231, 97-104.
Dubbink, H. J., Hersmus, R., Verma, C. S., van der Korput, H. A., Berrevoets,
C. A., van Tol, J., Ziel-van der Made, A. C. J., Brinkmann, A. O., Pike, A. C. W.
and Trapman, J. (2004). Distinct recognition modes of FXXLF and LXXLL motifsby the androgen receptor. Mol. Endocrinol. 18, 2132-2150.
Farla, P., Hersmus, R., Geverts, B., Mari, P. O., Nigg, A. L., Dubbink, H. J.,
Trapman, J. and Houtsmuller, A. B. (2004). The androgen receptor ligand-binding
domain stabilizes DNA binding in living cells. J. Struct. Biol. 147, 50-61.
Farla, P., Hersmus, R., Trapman, J. and Houtsmuller, A. B. (2005). Antiandrogens
prevent stable DNA-binding of the androgen receptor. J. Cell Sci. 118, 4187-4198.
Freedman, L. P., Yamamoto, K. R., Luisi, B. F. and Sigler, P. B. (1988). More fingers
in hand. Cell 54, 444.
Haelens, A., Verrijdt, G., Callewaert, L., Christiaens, V., Schauwaers, K., Peeters,
B., Rombauts, W. and Claessens, F. (2003). DNA recognition by the androgenreceptor: evidence for an alternative DNA-dependent dimerization, and an active roleof sequences flanking the response element on transactivation. Biochem. J. 369, 141-
151.
Hard, T., Dahlman, K., Carlstedt-Duke, J., Gustafsson, J. A. and Rigler, R. (1990a).
Cooperativity and specificity in the interactions between DNA and the glucocorticoidreceptor DNA-binding domain. Biochemistry 29, 5358-5364.
Hard, T., Kellenbach, E., Boelens, R., Maler, B. A., Dahlman, K., Freedman, L. P.,Carlstedt-Duke, J., Yamamoto, K. R., Gustafsson, J. A. and Kaptein, R. (1990b).Solution structure of the glucocorticoid receptor DNA-binding domain. Science 249,157-160.
He, B., Bowen, N. T., Minges, J. T. and Wilson, E. M. (2001). Androgen-inducedNH2- and COOH-terminal Interaction Inhibits p160 coactivator recruitment byactivation function 2. J. Biol. Chem. 276, 42293-42301.
Houtsmuller, A. B. (2005). Fluorescence recovery after photobleaching: application tonuclear proteins. Adv. Biochem. Eng. Biotechnol. 95, 177-199.
Hur, E., Pfaff, S. J., Payne, E. S., Grøn, H., Buehrer, B. M. and Fletterick, R. J.(2004). Recognition and accommodation at the androgen receptor coactivator bindinginterface. PLoS Biol. 2, e274.
Jia, L., Berman, B. P., Jariwala, U., Yan, X., Cogan, J. P., Walters, A., Chen, T.,Buchanan, G., Frenkel, B. and Coetzee, G. A. (2008). Genomic androgen receptor-occupied regions with different functions, defined by histone acetylation, coregulatorsand transcriptional capacity. PLoS ONE 3, e3645.
Karpova, T. S., Baumann, C. T., He, L., Wu, X., Grammer, A., Lipsky, P., Hager,G. L. and McNally, J. G. (2003). Fluorescence resonance energy transfer from cyanto yellow fluorescent protein detected by acceptor photobleaching using confocalmicroscopy and a single laser. J. Microsc. 209, 56-70.
Kenworthy, A. K. (2001). Imaging protein-protein interactions using fluorescenceresonance energy transfer microscopy. Methods 24, 289-296.
Luisi, B. F., Xu, W. X., Otwinowski, Z., Freedman, L. P., Yamamoto, K. R. and
Sigler, P. B. (1991). Crystallographic analysis of the interaction of the glucocorticoidreceptor with DNA. Nature 352, 497-505.
Massie, C. E., Adryan, B., Barbosa-Morais, N. L., Lynch, A. G., Tran, M. G., Neal,
D. E. and Mills, I. G. (2007). New androgen receptor genomic targets show aninteraction with the ETS1 transcription factor. EMBO Rep. 8, 871-878.
Matias, P. M., Donner, P., Coelho, R., Thomaz, M., Peixoto, C., Macedo, S., Otto,N., Joschko, S., Scholz, P., Wegg, A. et al. (2000). Structural evidence for ligandspecificity in the binding domain of the human androgen receptor. Implications forpathogenic gene mutations. J. Biol. Chem. 275, 26164-26171.
Roemer, S. C., Donham, D. C., Sherman, L., Pon, V. H., Edwards, D. P. and
Churchill, M. E. A. (2006). Structure of the progesterone receptor-deoxyribonucleicacid complex: novel interactions required for binding to half-site response elements.Mol. Endocrinol. 20, 3042-3052.
Rosenfeld, M. G., Lunyak, V. V. and Glass, C. K. (2006). Sensors and signals: acoactivator/corepressor/epigenetic code for integrating signal-dependent programs oftranscriptional response. Genes Dev. 20, 1405-1428.
Sack, J. S., Kish, K. F., Wang, C., Attar, R. M., Kiefer, S. E., An, Y., Wu, G. Y.,
Scheffler, J. E., Salvati, M. E., Krystek, S. R., Jr et al. (2001). Crystallographicstructures of the ligand-binding domains of the androgen receptor and its T877Amutant complexed with the natural agonist dihydrotestosterone. Proc. Natl. Acad. Sci.
USA 98, 4904-4909.
Savory, J. G. A., Prefontaine, G. G., Lamprecht, C., Liao, M., Walther, R. F.,
Lefebvre, Y. A. and Hache, R. J. G. (2001). Glucocorticoid receptor homodimersand glucocorticoid-mineralocorticoid receptor heterodimers form in the cytoplasmthrough alternative dimerization interfaces. Mol. Cell. Biol. 21, 781-793.
Schaufele, F., Carbonell, X., Guerbadot, M., Borngraeber, S., Chapman, M. S., Ma,
A. A. K., Miner, J. N. and Diamond, M. I. (2005). The structural basis of androgenreceptor activation: intramolecular and intermolecular amino-carboxy interactions.Proc. Natl. Acad. Sci. USA 102, 9802-9807.
Schwabe, J. W., Chapman, L., Finch, J. T. and Rhodes, D. (1993a). The crystalstructure of the estrogen receptor DNA-binding domain bound to DNA: how receptorsdiscriminate between their response elements. Cell 75, 567-578.
Schwabe, J. W., Chapman, L., Finch, J. T., Rhodes, D. and Neuhaus, D. (1993b).DNA recognition by the oestrogen receptor: from solution to the crystal. Structure 1,187-204.
Shaffer, P. L., Jivan, A., Dollins, D. E., Claessens, F. and Gewirth, D. T. (2004).Structural basis of androgen receptor binding to selective androgen responseelements. Proc. Natl. Acad. Sci. USA 101, 4758-4763.
Sultan, C., Lumbroso, S., Poujol, N., Belon, C., Boudon, C. and Lobaccaro, J. M.(1993). Mutations of androgen receptor gene in androgen insensitivity syndromes.J. Steroid Biochem. Mol. Biol. 46, 519-530.
Takayama, K., Tsutsumi, S., Katayama, S., Okayama, T., Horie-Inoue, K., Ikeda,K., Urano, T., Kawazu, C., Hasegawa, A., Ikeo, K. et al. (2011). Integration of capanalysis of gene expression and chromatin immunoprecipitation analysis on arrayreveals genome-wide androgen receptor signaling in prostate cancer cells. Oncogene
30, 619-630.
Tanenbaum, D. M., Wang, Y., Williams, S. P. and Sigler, P. B. (1998).Crystallographic comparison of the estrogen and progesterone receptor’s ligandbinding domains. Proc. Natl. Acad. Sci. USA 95, 5998-6003.
van de Wijngaart, D. J., van Royen, M. E., Hersmus, R., Pike, A. C. W.,
Houtsmuller, A. B., Jenster, G., Trapman, J. and Dubbink, H. J. (2006). NovelFXXFF and FXXMF motifs in androgen receptor cofactors mediate high affinity andspecific interactions with the ligand-binding domain. J. Biol. Chem. 281, 19407-19416.
van Royen, M. E., Cunha, S. M., Brink, M. C., Mattern, K. A., Nigg, A. L., Dubbink,
H. J., Verschure, P. J., Trapman, J. and Houtsmuller, A. B. (2007).Compartmentalization of androgen receptor protein-protein interactions in livingcells. J. Cell Biol. 177, 63-72.
van Royen, M. E., Dinant, C., Farla, P., Trapman, J. and Houtsmuller, A. B.
(2009a). FRAP and FRET methods to study nuclear receptors in living cells. In The
Journal of Cell Science 125 (8)1978
Journ
alof
Cell
Scie
nce

Nuclear Receptor Superfamily, Vol. 505 (ed. I. J. McEwan), pp. 69-96. Totowa, NJ:Humana Press.
van Royen, M. E., Farla, P., Mattern, K. A., Geverts, B., Trapman, J. andHoutsmuller, A. B. (2009b). FRAP to study nuclear protein dynamics in living cells.In The Nucleus, Vol. 464 (ed. R. Hancock), pp. 363-384. Totowa, NJ: Humana Press.
Wang, Q., Li, W., Liu, X. S., Carroll, J. S., Janne, O. A., Keeton, E. K., Chinnaiyan,A. M., Pienta, K. J. and Brown, M. (2007). A hierarchical network of transcriptionfactors governs androgen receptor-dependent prostate cancer growth. Mol. Cell 27,380-392.
Wang, Q., Li, W., Zhang, Y., Yuan, X., Xu, K., Yu, J., Chen, Z., Beroukhim, R.,Wang, H., Lupien, M. et al. (2009). Androgen receptor regulates a distincttranscription program in androgen-independent prostate cancer. Cell 138, 245-256.
Williams, S. P. and Sigler, P. B. (1998). Atomic structure of progesterone complexedwith its receptor. Nature 393, 392-396.
Yu, J., Yu, J., Mani, R. S., Cao, Q., Brenner, C. J., Cao, X., Wang, X., Wu, L., Li, J.,Hu, M. et al. (2010). An integrated network of androgen receptor, polycomb, andTMPRSS2-ERG gene fusions in prostate cancer progression. Cancer Cell 17, 443-454.
AR intra- and inter-molecular domain interactions 1979
Journ
alof
Cell
Scie
nce





























