Embed Size (px)
Citation preview

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, researchlibraries, and research funders in the common goal of maximizing access to critical research.
STATUS AND DISTRIBUTION OF THE ALLIGATOR SNAPPING TURTLE,MACROCHELYS TEMMINCKII, IN OKLAHOMAAuthor(s): J. Daren Riedle, Paul A. Shipman, Stanley F. Fox, and David M. Leslie Jr.Source: The Southwestern Naturalist, 50(1):79-84. 2005.Published By: Southwestern Association of NaturalistsDOI: http://dx.doi.org/10.1894/0038-4909(2005)050<0079:SADOTA>2.0.CO;2URL: http://www.bioone.org/doi/full/10.1894/0038-4909%282005%29050%3C0079%3ASADOTA%3E2.0.CO%3B2
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, andenvironmental sciences. BioOne provides a sustainable online platform for over 170 journals and books publishedby nonprofit societies, associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance ofBioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiriesor rights and permissions requests should be directed to the individual publisher as copyright holder.

THE SOUTHWESTERN NATURALIST 50(1):65–106 MARCH 2005
NOTES
SAGUARO (CARNEGIEA GIGANTEA, CACTACEAE) GROWTH RATE OVERITS AMERICAN RANGE AND THE LINK TO SUMMER PRECIPITATION
TALY DAWN DREZNER*
Department of Geography, Bolton 410, P.O. Box 413, University of Wisconsin–Milwaukee,Milwaukee, WI 53201-0413
*Correspondent: [email protected]
ABSTRACT Using an index of saguaro (Carnegiea gigantea) growth rate, relative growth was com-pared among multiple saguaro populations and mapped across the American range of the species.Relative growth at 10 sites was strongly linked to summer precipitation. This is the first study ofrelative growth and the first to link growth rate to environmental variables in multiple populations.
RESUMEN Usando un ındice de la tasa de crecimiento del saguaro (Carnegiea gigantea), se com-paro el crecimiento relativo entre poblaciones multiples de saguaro y se ilustro a traves del rangoamericano de la especie. El crecimiento relativo en 10 sitios fue fuertemente ligado a la precipi-tacion de verano. Este es el primer estudio de crecimiento relativo y el primero en ligar la tasade crecimiento a variables ambientales en poblaciones multiples.
Saguaro (Carnegiea gigantea) growth rates aregenerally consistent relative to height (e.g.,slow growth at youth, highest growth rates be-tween 3 and 4 m in height, and declininggrowth rates following) (Steenbergh andLowe, 1983; Turner, 1990; Niklas and Buch-man, 1994). Actual growth, however, varies bylocale (Steenbergh and Lowe, 1983). Drezner(2003a) recently developed a new method toestimate saguaro growth rate and age in anypopulation. By assuming a fixed growth pat-tern by height, a local growth factor can becomputed from repeat sampling of individuals.This value serves as an index, and growth ratescan then be compared between populations(Drezner, 2003a). The value of the factor atSaguaro National Park, East was set to 1.0, bydefinition, and values were calculated for Sa-guaro National Park, West (factor 5 0.743)and Organ Pipe Cactus National Monument(factor 5 0.617) (Drezner, 2003a). Thus, forexample, a saguaro that grows 10 cm per yearat a given height at Saguaro National Park,East will only grow 7.43 cm at that same heightat Saguaro National Park, West and 6.17 cm atOrgan Pipe Cactus National Monument.
The first objective of this study was to com-pare saguaro growth rates across multiple lo-cations that are geographically distant. In 2000,I used a telescoping leveling rod to collect dataon saguaro trunk height for 16 to 21 saguarosof varying heights and number of branches atseveral locales, all in flat areas (growth ratevaries by slope and aspect; Pierson and Turner,1998); washes were excluded (McAuliffe andJanzen, 1986; Drezner, 2003b). I revisited 2 ofthese locales in 2002 (see also Drezner andBalling, 2002), and I revisited 5 others in 2003.
I calculated the factor for the 7 resampledlocales by the procedure outlined in Drezner(2003a). The Drezner (2003a) technique isbased on a single general formula that modelssaguaro growth over its life, with a multiplica-tive adjustment factor for relative growth ateach site. This local adjustment factor (or in-dex value) can be calculated through repeatsampling of individuals and fitting height andgrowth at that height to the general growthcurve. Height and growth are fitted to thegrowth curve by minimizing the sum ofsquares, the same principle used for fitting aregression line (Drezner, 2003a).

66 vol. 50, no. 1The Southwestern Naturalist
For the first time, saguaro growth in differ-ent populations (with unique population struc-tures) can be quantified and compared acrossthe range of the species. A preliminary map ofsaguaro growth rates over its Arizona range isshown in Fig. 1. Saguaros generally exhibit rel-atively fast growth rates in the southeast, andgrowth rates decline northward and westwardacross their American range.
A second objective of this study was to ex-plore the relationship between saguaro growthrate and climate over its geographic range inArizona. I calculated 62 monthly climate vari-ables. These were based on 30 years of data(1968 through 1997) from 47 first-order me-teorological stations and the cooperative ob-serving network (National Climate Data Cen-ter, Summary of the Day database) rangingfrom the USA–Mexico border to 34.58N lati-tude and from the Arizona–California borderto 1108W longitude and 4 stations in eastern-most California. Although summer precipita-tion is highly variable over the short term (e.g.,1 rain event), the data for precipitation are re-liable when calculated for a 30-year period.The 62 variables used were lowest maximummonthly temperature and lowest minimumtemperature in December, in January, in Feb-ruary, and in all 3 months combined (8 vari-ables); mean minimum and mean maximummonthly temperature for these same months(8 variables); mean maximum monthly tem-perature in May through September (5 vari-ables); mean minimum monthly temperaturein May through September (5 variables); high-est maximum monthly temperature in Maythrough September (5 variables); highest min-imum monthly temperature in May throughSeptember (5 variables); highest maximumtemperature observed from June to August (1variable); maximum monthly precipitation andaverage monthly precipitation (24 variables);and mean annual precipitation (1 variable).These were interpolated to the 10 sampled lo-cales based on the methods described in Drez-ner (2003b), with adjustments from Steen-bergh and Lowe (1983). A complete list of themeteorological stations and their locations isprovided in Drezner and Garrity (2003). Usingthe Kolmogorov-Smirnov normality test, all var-iables were normally distributed at P . 0.01,and most were normal at P . 0.05.
With growth factor as the dependent vari-
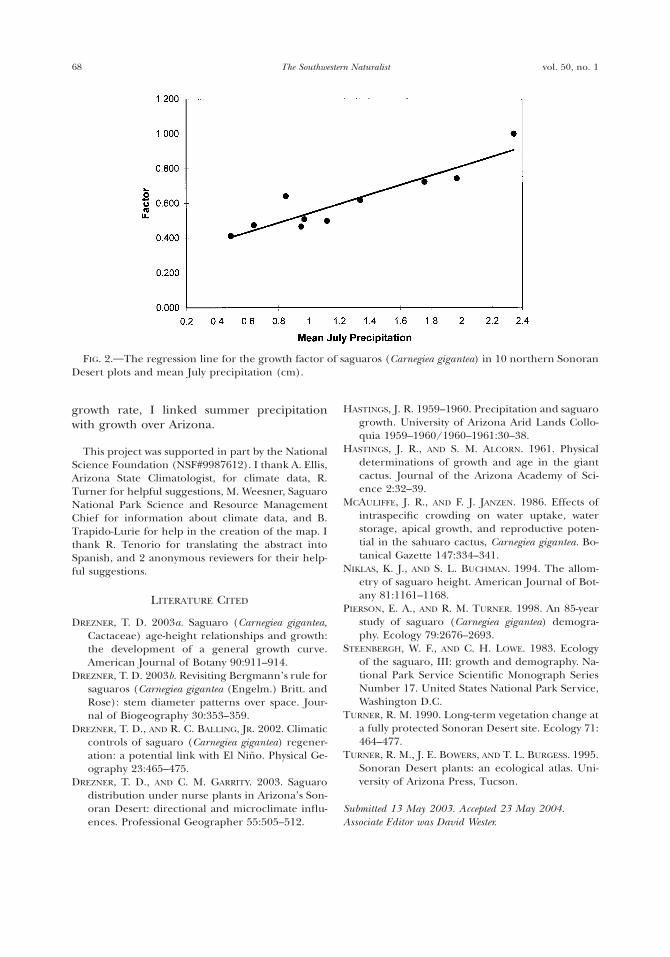
able, stepwise regression yielded one variable,mean July precipitation (P , 0.001; r2 5 0.84;Fig. 2). The resulting formula based on these10 locales is: growth factor 5 0.2705 1 0.2713(mean July precipitation).
One could indirectly estimate the growthfactor if mean July precipitation is available. Itis important to note, however, that the growthfactor is an estimate, and relying on summerprecipitation data to deduce growth rate mightcompound the error and should be discour-aged. The growth factor should be calculatedfrom direct field observation. It is clear thatwhere July rain is high, growth rates are high,and areas that receive less July rainfall supportslower growing saguaro populations. The pop-ulations sampled in this study exhibited sub-stantial variations in growth rate (Fig. 1). Forexample, a 5-m tall saguaro at the westernmostsite (Fig. 1) with a factor of 0.41 is estimatedto be 135 years old (e.g., Drezner and Balling,2002), while at Saguaro National Park, East, a5-m tall saguaro is estimated to be only 59 yearsold (Steenbergh and Lowe, 1983). Thus, if nofield data are available, mean July precipitationwill likely provide a better estimate than thedata of Steenbergh and Lowe (1983) from Sa-guaro National Park (which receives largequantities of summer rain); this is most notablytrue for sites far from Saguaro National Parkand the high summer rainfall belt. Nonethe-less, extreme caution should be used in inter-preting any results based on mean July precip-itation data rather than direct field observa-tion of populations.
Others have observed, through intensive di-rect measurement over a summer season, thatsaguaros grow in response to rainfall and dur-ing the summer (Hastings, 1959–1960; Has-tings and Alcorn, 1961; Steenbergh and Lowe,1983). These observations have all been madeat Saguaro National Park. The present studyquantified growth rate at multiple locales overthe northern Sonoran Desert. For the firsttime, a quantitative comparison of growth ratein different populations in different parts ofthe range of the species has been conducted,and a preliminary trend map is shown that de-picts saguaro growth patterns over the north-ern portion of their range. Finally, after ananalysis that included many possible environ-mental variables that might be linked with

March 2005 67Notes
FIG. 1 The location and growth factor of the 10 populations of saguaro (Carnegiea gigantea) sampled,including Saguaro National Park East (SNP-E), Saguaro National Park West (SNP-W), and Organ Pipe CactusNational Monument (OPCNM). The growth factor values for SNP-W, SNP-E, and OPCNM were derivedusing the data of Steenbergh and Lowe (1983) (Drezner, 2003a). The saguaro range is based on Turner etal. (1995).
68 vol. 50, no. 1The Southwestern Naturalist
FIG. 2. The regression line for the growth factor of saguaros (Carnegiea gigantea) in 10 northern SonoranDesert plots and mean July precipitation (cm).
growth rate, I linked summer precipitationwith growth over Arizona.
This project was supported in part by the NationalScience Foundation (NSF#9987612). I thank A. Ellis,Arizona State Climatologist, for climate data, R.Turner for helpful suggestions, M. Weesner, SaguaroNational Park Science and Resource ManagementChief for information about climate data, and B.Trapido-Lurie for help in the creation of the map. Ithank R. Tenorio for translating the abstract intoSpanish, and 2 anonymous reviewers for their help-ful suggestions.
LITERATURE CITED
DREZNER, T. D. 2003a. Saguaro (Carnegiea gigantea,Cactaceae) age-height relationships and growth:the development of a general growth curve.American Journal of Botany 90:911–914.
DREZNER, T. D. 2003b. Revisiting Bergmann’s rule forsaguaros (Carnegiea gigantea (Engelm.) Britt. andRose): stem diameter patterns over space. Jour-nal of Biogeography 30:353–359.
DREZNER, T. D., AND R. C. BALLING, JR. 2002. Climaticcontrols of saguaro (Carnegiea gigantea) regener-ation: a potential link with El Nino. Physical Ge-ography 23:465–475.
DREZNER, T. D., AND C. M. GARRITY. 2003. Saguarodistribution under nurse plants in Arizona’s Son-oran Desert: directional and microclimate influ-ences. Professional Geographer 55:505–512.
HASTINGS, J. R. 1959–1960. Precipitation and saguarogrowth. University of Arizona Arid Lands Collo-quia 1959–1960/1960–1961:30–38.
HASTINGS, J. R., AND S. M. ALCORN. 1961. Physicaldeterminations of growth and age in the giantcactus. Journal of the Arizona Academy of Sci-ence 2:32–39.
MCAULIFFE, J. R., AND F. J. JANZEN. 1986. Effects ofintraspecific crowding on water uptake, waterstorage, apical growth, and reproductive poten-tial in the sahuaro cactus, Carnegiea gigantea. Bo-tanical Gazette 147:334–341.
NIKLAS, K. J., AND S. L. BUCHMAN. 1994. The allom-etry of saguaro height. American Journal of Bot-any 81:1161–1168.
PIERSON, E. A., AND R. M. TURNER. 1998. An 85-yearstudy of saguaro (Carnegiea gigantea) demogra-phy. Ecology 79:2676–2693.
STEENBERGH, W. F., AND C. H. LOWE. 1983. Ecologyof the saguaro, III: growth and demography. Na-tional Park Service Scientific Monograph SeriesNumber 17. United States National Park Service,Washington D.C.
TURNER, R. M. 1990. Long-term vegetation change ata fully protected Sonoran Desert site. Ecology 71:464–477.
TURNER, R. M., J. E. BOWERS, AND T. L. BURGESS. 1995.Sonoran Desert plants: an ecological atlas. Uni-versity of Arizona Press, Tucson.
Submitted 13 May 2003. Accepted 23 May 2004.Associate Editor was David Wester.

March 2005 69Notes
PROBABLE HYBRIDIZATION IN THE ACACIA CONSTRICTA SPECIESGROUP (FABACEAE: MIMOSIDEAE)
H. DAVID CLARKE, JOHN E. EBINGER,* AND DAVID S. SEIGLER
Department of Biology, University of North Carolina-Asheville, Asheville, NC 28804 (HDC)Emeritus Professor of Botany, Eastern Illinois University, Charleston, IL 61920 (JEE)
Department of Plant Biology, University of Illinois, Urbana, IL 61801 (DSS)*Correspondent: [email protected]
ABSTRACT Principal components analyses (PCA) suggest that Acacia constricta and A. neovernicosararely hybridize, 1 lone probable hybrid being reported from Durango, Mexico. In contrast, hy-brids of A. neovernicosa and A. schottii seem to be more common. Probable hybrids involving these2 species were found in Brewster and Presidio counties, Texas, all within the known geographicrange of A. schottii.
RESUMEN Analisis de componentes principales (PCA) sugiere que Acacia constricta y A. neover-nicosa raramente sufren hibridizacion, solo se registra 1 probable hıbrido de Durango, Mexico.En contraste, hıbridos de A. neovernicosa y A. schottii parecen ser mas frecuentes. Hıbridos proba-bles de estas dos especies se encontraron en los condados de Brewster y Presidio, Texas, todosadentro del rango conocido de A. schottii.
The 5 members of the Acacia constricta spe-cies group occur in North America from south-western Texas and southern New Mexico andArizona, south through the deserts of northernand central Mexico. Shrubs, small trees, orrarely prostrate shrubs, these taxa have stipularspines, small bipinnately compound leaves with1 to 8 pairs of pinnae, leaflets mostly less than6 mm long, and globose inflorescences withsmall yellow flowers with numerous separatestamens. Members of this group are separatedfrom other members of Acacia subgenus Acaciaby the presence of involucral bracts near themiddle of the peduncles, a chartaceous peri-carpic strip lining the inner fruit valves, andoblong mottled seeds with small oval to U-shaped pleurograms. Included in this groupare A. biaciculata, A. constricta, A. glandulifera,A. neovernicosa, and A. schottii (Clarke et al.,1990).
During recent studies, occasional specimenswere encountered that did not fit the expectedrange of variation for some of the taxa includ-ed in this species group. These individualswere scored for many of the characters used inthe principal components analysis (PCA) ofthe original study of this species group byClarke et al. (1990) (Table 1). Data from spec-imens of the putative parental taxa, selected on
both a geographical and morphological basis,as well as specimens that other investigatorshad considered to be hybrids (based on fieldor annotation labels), were included in theseanalyses. Because some multistate characterswere included in the original analyses, PCAsalso were run using only continuous measure-ment characters. To avoid use of multistatecharacters in these analyses, a resemblance co-efficient for mixed data types was calculated(Legendre and Legendre, 1983; Podani, 1999)and the resulting matrix subjected to principalcoordinates analyses (PCoA).
A total of 62 specimens was selected for anal-ysis. Abbreviations used here for herbaria fol-low Holmgren et al. (1990). Each specimenwas scored for 13 vegetative and 4 fruit char-acters (Table 1). Measurement characters weredetermined for each specimen (3 or moremeasurements) and plotted to confirm thatgaps indeed exist in the character states em-ployed. Only primary leaves, that is, those as-sociated with nodes on elongated brancheswere used for measurement of leaf and leafletcharacters. Many individuals of these speciesdevelop numerous leaves on short shoots (spurbranches) in the axes of primary leaves. Theseleaves, designated as short-shoot leaves, aresmaller, have fewer pinna pairs, few leaflets per

70 vol. 50, no. 1The Southwestern Naturalist
TABLE 1—Characters scored for a principal components analysis involving Acacia constricta, A. neovernicosa,A. schottii, and the probable hybrids of these taxa, and their means and standard deviations.
Character Mean SD
1 Twig coating (TWI) (1 5 not glutinous, 2 5 glutinous)2 Petiole length (PTL) (mm)3 Petiole pubescence (PTP) (1 5 puberulent, 2 5 glabrous or nearly so)4 Petiole gland diameter (GLA) (mm)5 Rachis length (RAC) (mm)
1.406.681.480.369.23
0.491.840.500.179.52
6 Pinna pairs per leaf (PRS)7 Pinna length (PLN) (mm)8 Leaflet pairs per pinna (LPR)9 Leaflet arrangement (LAR) (1 5 opposite to subopposite, 2 5 alternate)
10 Leaflet length (LLN) (mm)
2.6011.306.711.322.80
1.484.682.230.471.07
11 Leaflet width (LWH) (mm)12 Leaflet cross section (CS) (1 5 flat, 2 5 subterete)13 Distance between leaflets (LDS) (mm)14 Fruit width (FWH) (mm)15 Fruit coating (FCO) (1 5 not glutinous, 2 5 glutinous)
0.731.291.654.601.73
0.240.460.971.430.45
16 Fruit glands (FGI) (1 5 absent, 2 5 sessile, 3 5 stalked)17 Fruit constrictions (FCT) (1 5 constrictions, 2 5 rarely constricted)
1.731.19
0.450.40
pinna, and smaller leaflets than leaves associ-ated with elongated stems.
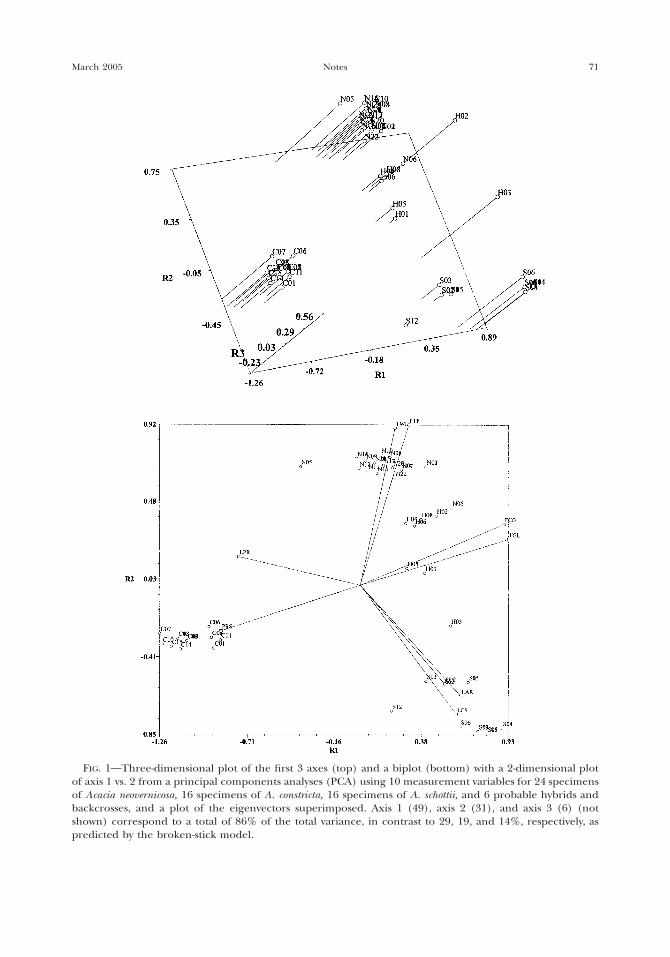
For principal components analyses (PCA),the data were first standardized and a corre-lation matrix, eigenvalues, and eigenvectorscalculated using NTSYS-pc version 2.1 (Rohlf,2000). Eigenvectors were scaled by the squareroot of l. The axes were rotated and the re-sulting loading values plotted as both 2-dimen-sional and 3-dimensional plots (Fig. 1).
To carry out PCoA, Gower’s resemblance co-efficients were calculated (Legendre and Le-gendre, 1983; Podani, 1999; Dickinson, 2000).The nature of each character was designated(binary, multistate, and quantitative descrip-tors) and all characters were weighted equally(Dickinson, 2000). The data matrix derivedwas transformed by the DCENTER algorithmusing distances squared, and eigenvectors andeigenvalues calculated with NTSYS-pc version2.1 (Rohlf, 2000). Eigenvectors were scaled bythe square root of l. The resulting loading val-ues were plotted as both 2-dimensional and 3-dimensional plots (Fig. 2).
A PCA using only measurement characters(Fig. 1), a PCA based on all 17 characters (notshown), a PCoA based on Gower’s similaritycoefficients for A. constricta, A. neovernicosa, A.schottii, and all of the possible and proposedhybrids (Fig. 2), as well as PCAs including A.neovernicosa and A. schottii with the putative hy-
brids (not shown) and including A. constrictaand A. neovernicosa with the respective putativehybrid (not shown), all suggested that hybrid-ization occurs between these species, based onintermediacy of purported hybrid operationaltaxonomic units in the analyses. This overallintermediacy of presumed hybrid individualsoccurred despite considerable variation andlack of intermediacy in some of the individualcharacters observed. Moreover, the positionsof these individuals were generally more vari-able than those of the putative parents in thevarious analyses conducted.
In the PCA with only measurement charac-ters (Fig. 1a, b), the first 2 characters account-ed for 80% of the total variance. Leaf widthand distance between leaflets (characters 11and 13) were most important for determiningthe component score on the first axis, petioleand leaflet length (characters 2 and 10) weremost important for the second axis, and thenumber of leaflet pairs per pinna (character8) for the third axis (Table 1). In a biplot withthe corresponding vectors, individuals of A.schottii (S) were differentiated from others bythe pinna and leaflet length, fruit width, anddistance between leaflets. The petiole lengthdifferentiated individuals of A. schottii and A.constricta (C) from those of A. neovernicosa (N).Individuals of A. constricta were distinct fromothers in the petiolar gland diameter, rachis
March 2005 71Notes
FIG. 1 Three-dimensional plot of the first 3 axes (top) and a biplot (bottom) with a 2-dimensional plotof axis 1 vs. 2 from a principal components analyses (PCA) using 10 measurement variables for 24 specimensof Acacia neovernicosa, 16 specimens of A. constricta, 16 specimens of A. schottii, and 6 probable hybrids andbackcrosses, and a plot of the eigenvectors superimposed. Axis 1 (49), axis 2 (31), and axis 3 (6) (notshown) correspond to a total of 86% of the total variance, in contrast to 29, 19, and 14%, respectively, aspredicted by the broken-stick model.

72 vol. 50, no. 1The Southwestern Naturalist
FIG. 2 Three-dimensional plot of the first 3 axes (top) and 2-dimensional plot (bottom) of axis 1 vs. 2from a principal coordinates analysis (PCoA) employing Gower’s similarity coefficients based on 17 variablesfor 24 specimens of Acacia neovernicosa, 16 specimens of A. constricta, 16 specimens of A. schottii, and 6probable hybrids and backcrosses. Axis 1 (21), axis 2 (17), and axis 3 (6) (not shown) correspond to a totalof 44% of the total variance, in contrast to 8, 9, and 5%, respectively, as predicted by the broken-stick model.

March 2005 73Notes
length, number of leaflet pairs per pinna, andleaflet width.
For the PCA based on 17 characters, the first2 principal components accounted for 79% ofthe total variance. Leaf width, leaflet arrange-ment, and distance between leaflets (charac-ters 11, 9, and 13) were most important fordetermining the component score of the firstaxis, whereas petiole pubescence, presence orabsence of a glutinous twig coating, and peti-ole length (characters 3, 1, and 2) were mostimportant for determining the second axis(Table 1).
To avoid the problem of using both contin-uous and multistate or binary characters inPCA, a PCoA using Gower’s resemblance co-efficients was performed (Legendre and Le-gendre, 1983; Podani, 1999; Dickinson, 2000).The resulting loading values are presented ina 2-dimensional and 3-dimensional plots (Fig.2). The parental species and putative hybridswere more clearly defined by this approachthan in the plot based on only measurementcharacters.
Acacia constricta 3 A. neovernicosa: Acacia neov-ernicosa is now rarely confused with A. constric-ta, but originally was considered a part of thattaxon (Standley, 1919; Benson, 1943). Easilyseparated, A. neovernicosa has glutinous leavesand twigs, is mostly glabrous throughout, andhas 1 to 2 (rarely 3) pairs of pinnae, whereasA. constricta lacks glutinous leaves and twigs,generally is lightly pubescent, and has leaveswith 3 to 8 pairs of pinnae. Specimens of A.constricta are consistently cyanogenic, those ofA. neovernicosa only rarely so (Seigler et al.,1976; Clarke et al., 1990). In addition, thechromosome numbers differ, 2n 5 52 for A.constricta and 2n 5 26 for A. neovernicosa (Turn-er and Fearing, 1960). Acacia neovernicosa isusually found on gypseous soil and generallyflowers 1 to 2 weeks before A. constricta (Turn-er, 1959). Isely (1969) mentioned that thesespecies are not separated geographically, or ap-parently ecologically, but he found little evi-dence of hybrids. The PCA and PCoA analysesof this study also suggested that hybrids occuronly rarely between these 2 species (Fig. 1). Ofthe 10 specimens that were suggested to be ofhybrid origin by information on the label oron annotation labels, only 1 specimen fell out-side the cluster of either species. This speci-men (N05) had large leaflets (2 to 3.5 mm
long), petioles more than 10 mm long, andfruits without glands, all traits of typical A. con-stricta. It was similar to A. neovernicosa in havinga glutinous covering on the stem and fruits,and mostly only 1 pair of pinnae per leaf. Thelabel information for this specimen is: MEXI-CO: Durango: Camp O, Mezquital, ca. 0.5 mileSW of town along Rıo Mezquital, elevation1,460 m, 15 November 1970, O. H. Soule 2059(MO).
Acacia neovernicosa 3 A. schottii: According toIsely (1969), A. schottii shows close affinities toA. constricta and A. neovernicosa. More recently,Clarke et al. (1990) found some specimensfrom Brewster and Presidio counties, Texas,that were intermediate in leaflet characteristicsbetween A. schottii and A. neovernicosa and werepresumed to be of hybrid origin.
Usually A. schottii is easily distinguished fromA. constricta and A. neovernicosa by its linear, su-bulate, alternate, and widely spaced leafletsthat give the foliage a feathery appearance.This species is apparently quite successful in itsrestricted range, where A. schottii is a commonspecies, often occurring in large, relativelypure thickets (Turner, 1959). These areas ofthorn-scrub vegetation are located on lime-stone or gypsum derived soils, where A. constric-ta and A. neovernicosa are common. Acacia schot-tii is particularly common in the southern por-tion of the Big Bend region of Texas and pos-sibly occurs to the south in adjacent Coahuila,Mexico, although the authors have seen nospecimens of this taxon from that region. Toour knowledge, the chromosome number forA. schottii has not been published.
Both PCA and PCoA analyses suggested thatthe hybrid A. neovernicosa 3 A. schottii is occa-sionally encountered in Brewster and Presidiocounties, Texas, within the known geographicrange of A. schottii. Most hybrid individuals in-cluded in the PCA were probably F1 hybrids,although others (H02, H07, and H08) couldbe possible backcrosses to A. schottii (Figs. 1and 2). Most of the putative hybrids were in-termediate in leaflet length and width, thenumber of leaflet pairs per pinna, the distancebetween leaflets along the pinna rachis, and infruit width (Table 1). In general, most individ-uals were similar to A. neovernicosa in havingglutinous material on some parts of the plantand flat leaflets, and similar to A. schottii in hav-

74 vol. 50, no. 1The Southwestern Naturalist
ing puberulent petioles and rachises and alter-nately arranged leaflets.
The probable hybrids are listed below, alongwith label information. Three specimens (H02,H07, and H08) might represent backcrossesand, although similar to other probable hy-brids, had leaves with longer rachises and pin-nae, more typical of A. schottii. UNITEDSTATES: Texas: Brewster County: clay-slatehills, near Lajitas, 27 June 1978, A. M. Powell3333 (SRSC, TEX); limestone soil 2 miles E ofPacksaddle Mountain, elevation 1,070 m, 12June 1949, B. L. Turner 1072 (SRSC); lime-stone hills at Boquillas, Big Bend NationalPark, elevation 985 m, 5 August 1966, B. H.Warnock 20896 (SRSC); sandy soil near AdobeWall Spring, elevation 1,020 m, 19 June 1949,B. H. Warnock, B. L. Turner, and J. O. Parks1134 (SRSC). Presidio County: gravel hills, ca.5 miles N of Lajitas, 7 July 1984, A. M. Powelland M. L. Powell 4406 (SRSC).
We wish to thank an anonymous referee and T. A.Dickinson who provided valuable suggestions and aBASIC program used to calculate Gower’s resem-blance coefficients. In addition, we thank the cura-tors of the Missouri Botanical Garden, University ofTexas, and Sul Ross State University for the loan ofspecimens used in this study. Financial support wasprovided through grant NSF DEB 04-5803.
LITERATURE CITED
BENSON, L. 1943. Revisions of status of southwesterndesert trees and shrubs. American Journal of Bot-any 30:230–240.
CLARKE, H. D., D. S. SEIGLER, AND J. E. EBINGER. 1990.Acacia constricta (Fabaceae: Mimosoideae) and re-lated species from the southwestern U.S. andMexico. American Journal of Botany 77:305–315.
DICKINSON, T. A. 2000. Program Gower6, BASIC soft-ware for calculation of Gower’s coefficients.Made available by the author.
HOLMGREN, P. K., N. H. HOLMGREN, AND L. C. BAR-NETT, editors. 1990. Index Herbariorum. Part I:the herbaria of the world, eighth edition. NewYork Botanical Garden, Bronx, New York. Updat-ed in: http://www.nybg.org/bsci/ih/ih.html.
ISELY, D. 1969. Legumes of the United States: I. Na-tive Acacia. Sida 3:365–386.
LEGENDRE, L., AND P. LEGENDRE. 1983. Numericalecology. Elsevier Scientific Publishing, Amster-dam, The Netherlands.
PODANI, J. 1999. Extending Gower’s general coeffi-cient of similarity to ordinal characters. Taxon48:331–340.
ROHLF, F. J. 2000. Numerical Taxonomy and Multi-variate Analysis Systems (NTSYSpc), version 2.1.Exeter Software, Setauket, New York.
SEIGLER, D. S., J. E. DUNN, AND E. E. CONN. 1976.Acacipetalin in Acacia constricta from NorthAmerica. Phytochemistry 15:219–200.
STANDLEY, P. C. 1919. New Mimosaceae. Contribu-tion from the United States National Herbarium20:184–191.
TURNER, B. L. 1959. The legumes of Texas. Univer-sity of Texas Press, Austin.
TURNER, B. L., AND O. S. FEARING. 1960. Chromo-some numbers in the Leguminosae. III. Speciesof the southwestern United States and Mexico.American Journal of Botany 47:603–608.
Submitted 4 October 2002. Accepted 19 April 2004.Associate Editor was Chris Lauver.
TEMPORAL ASSESSMENT OF A WEST TEXAS STREAMFISH ASSEMBLAGE
TIMOTHY H. BONNER,* CHAD THOMAS, CASEY S. WILLIAMS, AND JOHN P. KARGES
Department of Biology/Aquatic Station, Texas State University-San Marcos, San Marcos, TX 78666 (THB, CT, CSW)The Nature Conservancy, P.O. Box 2078, Fort Davis, TX 79734 (JPK)
*Correspondent: [email protected]
ABSTRACT We compared fish survey data across 3 collection periods (1952 to 1968, 1976 to1994, and 2001 to 2002) from Independence Creek (Rio Grande drainage, Terrell County, Texas)to assess long-term changes in fish assemblage structure. The number of species collected declinedfrom 28 in the 1952 to 1968 collection period, to 26 (plus 1 hybrid) in the 1974 to 1994 collectionperiod, and to 23 (plus 1 hybrid) in the 2001 to 2002 collection period. Species loss included 5

March 2005 75Notes
native fishes that currently persist in the adjoining Pecos River and 4 species that were non-nativeto the lower Pecos River. Species additions included species native to the Pecos River that rarelyinhabit smaller tributary streams and species that were introduced into the Pecos River. Twentyspecies were collected consistently from 1952 through 2002. Unweighted averages of abundantspecies across collection periods were Dionda episcopa (32%), Gambusia spp. (23%), Cyprinella pro-serpina (11%), C. lutrensis (10%), and Notropis amabilis (9%). Although apparent extirpations andadditions of several rare species (,1% in relative abundance each) were noted, the IndependenceCreek fish assemblage remained similar during the last 50 years, in contrast to the general trendof native species reductions in much of the Rio Grande drainage.
RESUMEN Comparamos los datos de estudios pesqueros en el arroyo Independencia (cuencadel rıo Grande, condado de Terrell, Texas), a lo largo de tres perıodos de colecta (1952 a 1968,1976 a 1994 y 2001 a 2002), para evaluar los cambios a largo plazo en la estructura del ensamblajede peces. El numero de especies colectadas declino de 28 en el periodo 1952 a 1968, a 26 (mas1 hıbrido) en el periodo 1974 a 1994 y a 23 (mas 1 hıbrido) en el periodo 2001 a 2002. La perdidade especies incluyo a 5 especies de peces nativas que actualmente persisten en el adyacente rıoPecos y 4 especies no nativas de la parte baja del mismo rıo. El aumento de especies incluyoespecies nativas del rıo Pecos que rara vez habitan en los pequenos arroyos tributarios y especiesque fueron introducidas al rıo Pecos. Veinte especies fueron colectadas consistentemente, de 1952a 2002. Los promedios no ajustados de las especies abundantes a lo largo de los periodos decolecta fueron Dionda episcopa (32%), Gambusia spp. (23%), Cyprinella proserpina (11%), C. lutrensis(10%), y Notropis amabilis (9%). Aun cuando hubo erradicaciones y aumentos aparentes de variasespecies raras (,1% de abundancia relativa de cada una), el ensamblaje de peces del arroyoIndependencia se mantuvo similar durante los ultimos 50 anos, en contraste con la tendenciageneral en la reduccion de especies nativas en la mayor parte de la cuenca del rıo Grande.
In the lower Pecos River (Carlsbad, NewMexico, to the Rio Grande), the number ofnative fishes has been reduced by half becauseof anthropogenic modifications, including in-troduced species, reduced water quality, anddiminished spring and stream flows (Hoags-trom, 2003). However, the largest tributary ofthe lower Pecos River, Independence Creek,has been minimally impacted by human activ-ities and continues to support many of the fish-es native to the lower Pecos River and RioGrande drainages (Karges, 2003). The Inde-pendence Creek fish assemblage includes 2state-listed (Texas) threatened species (Cypri-nella proserpina and Etheostoma grahami), 1 spe-cies (Notropis jemezanus) considered threatenedin Texas by Hubbs et al. (1991), 1 species (Ic-talurus lupus) considered of special concern inTexas by Hubbs et al. (1991), and 2 species(Dionda episcopa and Notropis amabilis) endemicto the Edwards Plateau of Texas and RioGrande drainages. In 1998, The Nature Con-servancy purchased 8,000 ha that included 13km of Independence Creek and CarolineSpring, a primary contributor of freshwater tothe creek. The purpose of our study was tocompare historical fish survey data with thatfrom recent surveys to assess long-term chang-
es in the fish assemblage of IndependenceCreek.
Independence Creek drains 1,935 km2 oflimestone-based Ector Rock Outcrop soils com-posed of stony loams, stony clay loams, androck outcrops (Turner and Fox, 1974). The pe-rennial portion of Independence Creek isabout 16 km long and is sustained largely byVanderbeek Spring and Caroline Spring, withseveral peripheral springs contributing lesserflow (Brune, 1981). Throughout its course, In-dependence Creek is dominated by runs andpools, with few riffles and backwater areas.Stream width ranges from 6 to 17 m, currentvelocity ranges from 5 to 45 cm/s, and depthranges from 18 to 48 cm. Dominant substratesare cobble and gravel, with a few reaches ofbedrock and cemented gravel. Dominantstreamside plant associations are classified asWalnut-Desert Willow, Saltcedar, and Sawgrass-Willow (Webster, 1950).
Information on the fish fauna of Indepen-dence Creek was obtained from Tulane Muse-um of Natural History (1952 to 1976); L.Campbell (1959, Basic survey and inventory ofspecies present in the Pecos River of Texas,Texas Game and Fish Commission, Austin); N.Valdez (1994, Composition and structure of

76 vol. 50, no. 1The Southwestern Naturalist
TABLE 1—Relative abundance (%) of species from3 collection periods in Independence Creek, Texas.The letter ‘‘I’’ denotes nonnative species in Inde-pendence Creek. The letter ‘‘P’’ denotes fishes thatwere present but not counted.
Species
Relative abundance
1952–1968
1974–1994
2001–2002
Lepisosteus osseusDorosoma cepedianumCampostoma anomalum (I)Cyprinella lutrensisCyprinella proserpina
0.02
0.0421.37.1
0.07
7.012.7
P
0.511.7
Cyprinella venusta (I)Cyprinus carpio (I)Dionda episcopaMacrhybopsis aestivalisNotropis amabilis
0.010.1
36.10.024.0
0.05
30.80.45.2
0.2P
29.7
19.1Notropis braytoniNotropis jemezanusNotropis stramineusPimephales vigilaxCarpiodes carpio
0.5
0.020.20.1
8.70.9
0.70.02
Moxostoma congestumAstyanax mexicanusIctalurus sp.Ictalurus lupusIctalurus punctatus
0.053.60.012.9
0.11.6
0.50.5
0.41.7
0.8P
Pylodictis olivarisCyprinodon variegatus 3
C. pecosensis (I)Fundulus zebrinusLucania parvaGambusia spp.
5.10.2
13.2
0.04
0.050.020.4
26.8
0.05
0.50.50.5
28.7Menidia beryllina (I)Lepomis auritus (I)Lepomis cyanellusLepomis macrochirusLepomis megalotis
0.10.90.041.2
0.020.40.090.021.1
2.50.10.070.5
Micropterus salmoidesPomoxis annularis (I)Etheostoma grahamiCichlasoma cyanoguttatum
0.20.020.52.3
0.2
0.41.3
0.3
1.21.0
Number of individualsSpecies richnessSpecies diversity (H)Evenness (E)
8,388281.970.25
5,514271.970.26
4,062241.790.25
fish assemblages of Chandler IndependenceCreek Preserve, Texas A&M University, unpub-lished report); G. W. Linam and L. J. Kleinsas-ser (1996, Relationship between fishes and wa-ter quality in the Pecos River, Texas, TexasParks and Wildlife Department, Austin); G. P.Garrett (1997, Chihuahuan Desert fishes statussurvey, Texas Parks and Wildlife Department,Ingram); and G. W. Linam, L. J. Kleinsasser,and K. B. Mayes (2002, Regionalization of theindex of biotic integrity for Texas streams, Tex-as Parks and Wildlife Department, Austin).Collectively, these records were from 1952through 1994. Specimens were collected withseines or electrofishing gear, but effort andhabitat types sampled were not always record-ed. We assumed that species composition andabundances obtained during these collectionsaccurately represented the IndependenceCreek fish assemblage at the time and locationof sampling.
Our collections were made from December2001 through July 2002 at 7 sites on Indepen-dence Creek from its headwaters at Vander-beek Spring to its confluence with the PecosRiver. Fishes were collected with seines and abackpack electrofisher (Smith-Root Model 12-B), identified, enumerated, and released, ex-cept for voucher specimens. Voucher speci-mens were anesthetized in tricaine methanesulphonate and preserved in 10% formalin.Field identification of 3 previously reportedGambusia (G. affinis, G. geiseri, and G. speciosa)was complicated by hybridization and nomen-clatorial confusion between G. affinis and G.speciosa (G. P. Garrett, pers. comm.). There-fore, all Gambusia were treated as 1 taxon.
Assemblage composition and relative abun-dances were compared among 3 collection pe-riods: 1952 to 1968, 1974 to 1994, and 2001 to2002. Shannon-Weiner index (H; natural base)for species diversity, evenness (Buzas and Gib-son’s E; Hayek and Buzas, 1997), and Renko-nen similarity index (Krebs, 1989) were usedto quantify and compare assemblages acrosstime.
Thirty-three species and 1 hybrid were takenfrom Independence Creek from 1952 through2002 (Table 1). The number of species col-lected declined from 28 in the 1952 to 1968collection period, to 26 (plus 1 hybrid) in the1974 to 1994 collection period, and to 23 (plus
1 hybrid) in the 2001 to 2002 collection peri-od. Species diversity was 1.97 from the 1952 to1968 and 1974 to 1994 collection periods, andslightly lower (1.79) for the 2001 to 2002 col-lection period. Evenness values (range 0.25 to0.26) were not substantially different among

March 2005 77Notes
collection periods. Assemblage similarity was68% between 1952 to 1968 and 1974 to 1994collection periods, and 79% between 1974 to1994 and 2001 to 2002 collection periods. Al-though assemblages were similar among collec-tion periods, some notable changes in assem-blage composition and species abundancewere apparent.
Of 8 species not taken during 2001 to 2002,3 (Lepomis microlophus, Pomoxis annularis, andpossibly Campostoma anomalum) are not nativeto the lower Pecos River (Hubbs et al, 1991;Hoagstrom, 2003), and 5 native fishes (Lepisos-teus osseus, Macrhybopsis aestivalis, Notropis stra-mineus, N. braytoni, and Pimephales vigilax) per-sist in the lower Pecos River near Indepen-dence Creek (Hoagstrom, 2003). These nativefishes normally inhabit larger rivers with siltand sand substrate, such as the lower PecosRiver, and marginally or temporarily inhabitsmaller tributaries with cobble and gravel sub-strate, such as Independence Creek (Pflieger,1975; Robison and Buchanan, 1988; Edwardsand Contreras-Balderas, 1991; Page and Burr,1991; Rhodes and Hubbs, 1992). Thus, re-duced species richness in the IndependenceCreek fish assemblage through time was notattributed to native species extirpations.
Six species (Dorosoma cepedianum, N. jemezan-us, I. punctatus, Pylodictis olivaris, Cyprinodon var-iegatus 3 C. pecosensis, and Menidia beryllina)not taken during 1952 to 1968 were found dur-ing 1974 to 2002. Four species (D. cepedianum,N. jemezanus, I. punctatus, and P. olivaris) arenative to the lower Pecos River, although I.punctatus might have derived from individualsstocked for sportfishing purposes in CarolineSpring (Karges, 2003). Cyprinodon variegatus(and its subsequent hybridization with C. peco-sensis) and M. beryllina occurrences are attri-buted to incidental stockings in the late 1950sor early 1960s in the lower Pecos River (Minck-ley, 1965; Stevenson and Buchanan, 1973).
Eighteen species were present during eachcollection period. Two species (Cyprinus carpioand Carpiodes carpio) were not collected during1964 through 1974. Among these 20 species, 4(Cyprinella venusta, Cyprinus carpio, L. auritus,and 1 of the Gambusia) are not native to thelower Pecos River (Hubbs et al., 1991; Hoags-trom, 2003). Average percent abundancesacross all 3 periods were D. episcopa (32%),Gambusia (23%), Cyprinella proserpina (11%), C.
lutrensis (10%), and N. amabilis (9%) for themost common fishes. Abundant species withthe greatest change in relative abundancethrough time were C. lutrensis (decreased from21 to 0.5%), N. amabilis (increased from 4% to19%), and Gambusia (increased from 13 to29%), although theses changes might be par-tially attributed to previous collections takenmainly from lower reaches, whereas 2001 to2002 collections were evenly distributedthroughout Independence Creek. In 2001 to2002 collections, C. lutrensis was abundant onlyin the lower reach of Independence Creek.
Among species of conservation concern, C.proserpina, E. grahami, and I. lupus were takenduring each collection period. Cyprinella proser-pina was the most abundant state-listed threat-ened species in Independence Creek and wastaken from all sampling locations in 2001 to2002 collections. Etheostoma grahami and I. lu-pus were relatively uncommon (,3%) but per-sistent in Independence Creek. Among habi-tats, E. grahami was abundant in riffles and I.lupus was abundant in deep runs in the upperreaches of Independence Creek. Notropis jeme-zanus has not been collected in IndependenceCreek since 1991 or in the lower Pecos Riversince 1987 (Hoagstrom, 2003). Dionda episcopaand N. amabilis, species endemic to the Ed-wards Plateau and Rio Grande drainage, com-posed 49% of the current fish assemblage inIndependence Creek.
Although apparent extirpations and addi-tions of several uncommon species (,1% inrelative abundance each) were noted, the In-dependence Creek fish assemblage remainedsimilar during the last 50 years. This is in con-trast to other lotic environments in the RioGrande and Gulf slope drainages of Texas,where 80% of 129 fish assemblages were sub-stantially changed during a 30-year period (An-derson et al., 1995). Reservoir construction,dredging, pollution, salination, and exotic in-troductions were responsible for changes in60% of the assemblages characterized as al-tered by Anderson et al. (1995). Although hab-itats of Independence Creek have been modi-fied by water diversions for irrigation, im-poundment of Caroline Spring, and nonnativefish introductions, fish composition and abun-dance have not substantially changed. The per-sistence of a largely intact native fish assem-blage in Independence Creek is a contrast to

78 vol. 50, no. 1The Southwestern Naturalist
the general trend of native species reductionsin much of the Rio Grande drainage (Edwardsand Contreas-Bladeras, 1991; Platania, 1991;Edwards et al., 2003; Hoagstrom, 2003).
Texas Parks and Wildlife Department, The NatureConservancy, and Texas State University-San Marcos,Department of Biology, provided funding for thisproject. J. C. Wrinkle (Lower Pecos Project Director,The Nature Conservancy) provided access on Inde-pendence Creek. N. G. Martinez, J. M. Watson, C. A.Hooker, and C. Hartl assisted with fieldwork. G. W.Linam, G. P. Garrett, and an anonymous reviewerprovided helpful comments that improved this man-uscript. D. G. Solıs provided resumen translation.
LITERATURE CITED
ANDERSON, A. A., C. HUBBS, K. G. WINEMILLER, AND
R. J. EDWARDS. 1995. Texas freshwater fish assem-blage following three decades of environmentalchange. Southwestern Naturalist 40:314–321.
BRUNE, G. M. 1981. Springs of Texas, volume 1.Branch-Smith, Inc. Fort Worth, Texas.
EDWARDS, R. J., AND S. CONTRERAS-BALDERAS. 1991.Historical changes in the ichthyofauna of the low-er Rio Grande (Rio Bravo Del Norte), Texas andMexico. Southwestern Naturalist 36:201–212.
EDWARDS, R. J., G. P. GARRETT, AND E. MARSH-MAT-THEWS. 2003. Fish assemblages of the Rio Con-chos basin, Mexico, with emphasis on their con-servation and status. In: G. P. Garrett and N. L.Allen, editors. Aquatic fauna of the NorthernChihuahuan Desert. Special Publications Num-ber 46, Museum, Texas Tech University, Lub-bock. Pp. 75–89.
HAYEK, L. C., AND M. A. BUZAS. 1997. Surveying nat-ural populations. Columbia University Press, NewYork.
HOAGSTROM, C. W. 2003. Historical and recent fishfauna of the lower Pecos River. In: G. P. Garrettand N. L. Allen, editors. Aquatic fauna of theNorthern Chihuahuan Desert. Special Publica-tions Number 46, Museum, Texas Tech Univer-sity, Lubbock. Pp. 91–109.
HUBBS, C., R. J. EDWARDS, AND G. P. GARRETT. 1991.An annotated checklist of the freshwater fishes ofTexas, with keys to identification of species. TexasJournal of Science, Supplement 43:1–56.
KARGES, J. 2003. Aquatic conservation and The Na-ture Conservancy of West Texas. In: G. P. Garrettand N. L. Allen, editors. Aquatic fauna of theNorthern Chihuahuan Desert. Special Publica-tions Number 46, Museum, Texas Tech Univer-sity, Lubbock. Pp. 141–150.
KREBS, C. J. 1989. Ecological methodology.HarperCollins, New York.
MINCKLEY, W. L. 1965. Records of atherinid fishes atinland localities in Texas and northern Mexico.Great Basin Naturalist 25:73–76.
PAGE, L. M., AND B. M. BURR. 1991. A field guide tofreshwater fishes of North America north of Mex-ico. Houghton Mifflin Company, Boston, Massa-chusetts.
PFLIEGER, W. L. 1975. The fishes of Missouri. Missou-ri Department of Conservation, Jefferson City.
PLATANIA, S. P. 1991. Fishes of the Rio Chama andupper Rio Grande, New Mexico, with prelimi-nary comments on their longitudinal distribu-tion. Southwestern Naturalist 36:186–193.
RHODES, K., AND C. HUBBS. 1992. Recovery of PecosRiver fishes from a red tide fish kill. Southwest-ern Naturalist 37:178–187.
ROBISON, H. W., AND T. M. BUCHANAN. 1988. Fishesof Arkansas. University of Arkansas Press, Fayette-ville.
STEVENSON, M. M., AND T. M. BUCHANAN. 1973. Ananalysis of hybridization between the cyprino-dont fishes Cyprinodon variegatus and C. elegans.Copeia 1973:682–692.
TURNER, A. J., AND R. E. FOX. 1974. Soil survey ofTerrell County, Texas. United States Departmentof Agriculture, Soil Conservation Service.
WEBSTER, G. L. 1950. Observations on the vegetationand summer flora of the Stockton Plateau innortheastern Terrell County, Texas. Texas Jour-nal of Science 2:234–242.
Submitted 21 January 2003. Accepted 10 May 2004.Associate Editor was David Propst.

March 2005 79Notes
STATUS AND DISTRIBUTION OF THE ALLIGATOR SNAPPING TURTLE,MACROCHELYS TEMMINCKII, IN OKLAHOMA
J. DAREN RIEDLE,* PAUL A. SHIPMAN, STANLEY F. FOX, AND DAVID M. LESLIE, JR.
Oklahoma Cooperative Fish and Wildlife Research Unit, Department of Zoology, Oklahoma State University,Stillwater, OK 74078 (JDR, PAS)
Department of Zoology, Oklahoma State University, Stillwater, OK 74078 (SFF)United States Geological Survey, Oklahoma Cooperative Fish and Wildlife Research Unit, Oklahoma State University,
Stillwater, OK 74078 (DML)Present address of JDR: Arizona Game and Fish Department, Nongame Branch, 2221 West Greenway Road,
Phoenix, AZ 85023Present address of PAS: Department of Biological Sciences, Rochester Institute of Technology, 85 Lomb Memorial Drive,
Rochester, NY 14623*Correspondent: [email protected]
ABSTRACT Although historic records of the alligator snapping turtle, Macrochelys temminckii,show a past distribution throughout eastern Oklahoma, little is known about the current statusand distribution of this species in the state. In 1997, surveys were initiated to identify extantpopulations of M. temminckii and assess their relative densities and viability. We surveyed 67 sitesin 15 counties, with a total effort of 1,085 net nights. A total of 63 M. temminckii was captured at11 sites, which are only in the southeastern quarter of the state and occur only in protected orisolated locations. Because of this apparent decrease of the range of this species in Oklahoma andbecause so few sites exhibited capture rates high enough to suggest possible healthy populations,we conclude that dramatic population declines of M. temminckii have taken place in Oklahoma.Possible reasons for these declines include overharvest and habitat alteration.
RESUMEN Aunque los registros historicos de la tortuga Macrochelys temminckii demuestran unadistribucion anterior en todo el este de Oklahoma, no se sabe mucho del estatus y distribucionactual de la especie en el estado. En 1997, se iniciaron muestras para identificar poblacionesactuales de M. temminckii y para evaluar sus abundancias relativas y viabilidad. Muestreamos 67sitios en 15 condados con un esfuerzo total de 1,085 red-noches. Un total de 63 M. temminckii fuecapturado en 11 sitios, que se ubican solamente en el cuarto sureste del estado y se encuentransolamente en localidades protegidas o aisladas. Debido a esta aparente disminucion de la distri-bucion de esta especie en Oklahoma y debido a que solo unos cuantos sitios mostraron tasas decaptura tan altas para sugerir poblaciones saludables, concluimos que declines dramaticos de M.temminckii han ocurrido en Oklahoma. Las razones posibles para los declines incluyen la caserıaexcesiva y la alteracion del habitat.
The alligator snapping turtle, Macrochelystemminckii, is the largest freshwater turtle inNorth America, with males attaining a cara-pace length of 80 cm and a live mass of 113 kg(Pritchard, 1989). Adults exhibit sexual dimor-phism; females reach a maximum live mass ofonly 35 kg (Pritchard, 1989). Little informa-tion exists on the biology of M. temminckii. Prit-chard (1989) and Ernst et al. (1994) suggestedthat M. temminckii populations have declinedthroughout its range. Overharvesting and hab-itat alteration have been listed as the primarycauses (Reed et al., 2002). In 1984, the UnitedStates Fish and Wildlife Service proposed M.
temminckii for listing as a threatened species.However, the request for listing was precludeddue to a lack of ecological information aboutthe species. The status of the species was re-viewed again in 1991, but no further actionswere taken (United States Fish and WildlifeService, 1991). At the state level, M. temminckiiis afforded some protection in all states inwhich it occurs, except Louisiana (Roman andBowen, 2000). Macrochelys temminckii currentlyis listed as a species of special concern inOklahoma.
Macrochelys temminckii is confined to river sys-tems that drain into the Gulf of Mexico. The

80 vol. 50, no. 1The Southwestern Naturalist
FIG. 1 Historic distribution of the alligator snapping turtle, Macrochelys temminckii, in Oklahoma.
species occurs as far north as Kansas and Illi-nois (Galbreath, 1961; Clarke, 1981) and fromthe Florida Panhandle to eastern Texas andOklahoma (Conant and Collins, 1991). Histor-ically, M. temminckii occurred throughout theeastern one-third of Oklahoma (Glass, 1949;Webb, 1970; Black, 1982; Carpenter and Kru-pa, 1989; and Heck, 1999; Fig. 1). Historicalaccounts of M. temminckii are based on singleindividuals, so information on distribution anddemography of M. temminckii in Oklahoma ismeager. Our objectives were to: 1) identify ex-tant populations of M. temminckii in Oklahoma;2) assess their relative densities and viability;and 3) capture, permanently mark, and releaseall specimens for subsequent population mon-itoring.
We sampled sites throughout the easternone-third of Oklahoma from May through Au-gust 1997 to 1999, with supplemental samplingof 2 sites in July 2000. Many of those sites wereat or near historic sites of occurrence for thespecies in Oklahoma. We surveyed a variety ofhabitats to adequately sample all possible hab-itats in which M. temminckii might occur.
We sampled sites using commercial hoopnets that were 2.1 m in length and constructed
of 4 hoops (1.05 m in diameter) covered with2.5-cm square-mesh net. Nets were set up-stream from submerged structures, such astrees and log jams, and were baited with freshfish suspended by a piece of twine on the hoopfarthest from the opening of the trap. We setnets in the late afternoon or evening andchecked them the following morning.
We surveyed 67 sites in 15 counties (Fig. 2).Some sites were surveyed more than once dueto the presence of M. temminckii or if seeminglygood habitat was present. Our total trappingeffort was 1,085 net nights (1 net per night 51 net night). From 1997 to 1999, we made 69captures of 63 individuals of M. temminckii(plus 8 more captures in July 2000 from Se-quoyah National Wildlife Refuge) at 11 sites(Table 1; Fig. 3).
Macrochelys temminckii was once distributedthroughout all the major river systems in east-ern Oklahoma but was captured in our studyat only 11 of the 67 sites sampled within thathistoric range. This suggests a dramatic declinein numbers of M. temminckii in the state. Cur-rently known populations seem to be restrictedto a few remote or protected locations in thesoutheastern one-quarter of Oklahoma. Of

March 2005 81Notes
FIG. 2 Sites sampled for Macrochelys temminckii in Oklahoma between 1997 and 1999. Some points rep-resent more than 1 site due to close proximity of sample sites.
TABLE 1—Alligator snapping turtle (Macrochelys temminckii) capture rates in Oklahoma by site.
Site CountyNumber of
catpuresNet
nights
Capture rate(number ofturtles pernet night)
Little RiverKiamichi River*Dirty Creek**Hezekiah Creek**
McCurtainPushmatahaMuskogeeSequoyah
3273
167343717
0.0180.0590.1200.180
Big Vian Creek***Little Vian CreekDutchess CreekMill Creek
SequoyahSequoyahMcIntoshMcIntosh
242648
126649
13
0.2000.4100.4440.620
* Represents 1 site on the Kiamichi River and 1 site on its tributary, Mill Creek.** Resampled July 2000, bringing total number of M. temminckii captures to 77.*** Represents 2 sites on Big Vian Creek and 1 site on Horton Slough.
those populations, only the sites at Eufala Res-ervoir (Mill Creek and Dutchess Creek) andSequoyah National Wildlife Refuge (Big VianCreek, Little Vian Creek, Dirty Creek, HortonSlough, and Hezekiah Creek) yielded capturerates high enough to suggest viable popula-tions (Table 1). Macrochelys temminckii seems tohave been extirpated from the northeastern
one-quarter of the state. The possible reasonsfor this decline are habitat alteration and deg-radation and historical incidental and illegalharvest.
There are several forms of habitat alterationthat might have a negative effect on M. tem-minckii in Oklahoma. The Verdigris River hasbeen channelized for navigation throughout

82 vol. 50, no. 1The Southwestern Naturalist
FIG. 3 Distribution of Macrochelys temminckii in Oklahoma based on the 1997 to 1999 survey. Some pointsrepresent more than 1 site due to close proximity of sample sites.
much of Oklahoma. This manipulation of theriver channel turns a low-energy, meandering,aquatic system with high habitat diversity intoa higher energy system with low habitat diver-sity that is vastly different from the habitat pre-ferred by M. temminckii (Shipman, 1993; Molland Moll, 2000).
Macrochelys temminckii is exclusively aquatic,except for females during egg laying (Prit-chard, 1989). An impoundment, such as a damor a lock, would block movement of individualsupstream or downstream of the structure. TheArkansas, Caney, Verdigris, and Neosho riversseem to be the major dispersal pathways for M.temminckii throughout the central and north-ern parts of its range in Oklahoma. The seriesof locks and dams along the Arkansas, Caney,and Verdigris rivers might be the main imped-iment to the dispersal of individuals into thenorthern reaches of Oklahoma rivers andstreams.
Thermal alteration of aquatic environments,such as hypolimnetic release of cold water, alsomight be responsible for the decrease in M.temminckii abundance in Oklahoma. TheMountain Fork River in McCurtain County is
managed as a coldwater stream for trout fish-ing. Summer water temperatures taken duringthe study varied between 178 and 218C. Littlework has been done with the thermal require-ments of M. temminckii, but Allen and Neill(1950) noted that they refuse food at temper-atures ,188C. Based on our observations, thethermal environment in rivers such as theMountain Fork is suboptimal for M. temminckii.A 36.4-kg M. temminckii was captured on theMountain Fork River in 1993 by anglers (Ship-man, Fox, and Riedle, pers. obser.), but no in-dividuals were captured on the Mountain Forkduring our survey. Heck (1998) reported a de-cline in the number of M. temminckii observedon the Mountain Fork River since the con-struction of the Broken Bow Dam in 1969; hislast M. temminckii reported from the MountainFork River was from 1995.
Heck (1998) listed several sources of pollu-tion on the Little River that might have con-tributed to the decline of M. temminckii overthe last 30 y. Sources include sewage discharge,runoff from chicken farms, wastewater dis-charge from chicken processing plants, chem-ical runoff, and soil erosion from commercial

March 2005 83Notes
timber harvest. Large amounts of wastewaterdischarge were noted on the Little River southof Broken Bow, McCurtain County, in 1998.Turtle captures at both the Little River andMountain Fork River were not only especiallylow for M. temminckii, but for all aquatic turtlesnative to that area of Oklahoma (Riedle,2001).
The primary forms of harvest of M. tem-minckii include incidental and illegal capture.Most incidental captures are those on trotlinesand limb lines set by fishermen for catfish.Shipman and Riedle (1994) identified unat-tended limb lines and trotlines as a primarythreat to turtles on the Saint Francis River insoutheastern Missouri. Shipman et al. (1991)reported a specimen caught on a limb line 32km north of the Oklahoma border on a trib-utary of the Verdigris River in Kansas. Heck(1998) listed several accounts of M. temminckiicaptures on limb lines and trotlines in Mc-Curtain County, Oklahoma.
Due to its large adult size, M. temminckii hasbeen harvested historically throughout itsrange as a source of meat for personal andcommercial use (Pritchard, 1989). Sloan et al.(1995) reported 17,117 kg live-weight of M.temminckii purchased by a single buyer in Lou-isiana between 1984 and 1986. Turtles werehistorically taken for this market from Florida,Georgia, Mississippi, Arkansas, Texas, and pos-sibly Oklahoma (Pritchard, 1989). The major-ity of historical records for Oklahoma (Glass,1949; Webb, 1970; Black, 1982; Carpenter andKrupa, 1989; Heck, 1998) were based on indi-viduals taken by fishermen, and all were keptby the fishermen themselves or donated to pri-vate or public collections. Commercial harvestin Louisiana is still ongoing, even though M.temminckii is protected in surrounding states.
Although little information is available onthe historical status of M. temminckii, it seemsthat the species has suffered a severe declinein Oklahoma. Current populations occur in ar-eas that are difficult to access and are affordedsome protection from harvest. In Missouri,Shipman and Riedle (1994) found that M. tem-minckii was absent or rare from sites that hadseemingly suitable habitat but were in closeproximity to historic and current turtle meatmarkets. This also seems to be true inOklahoma. Due to the many river impound-ments in Oklahoma, recruitment upstream
and downstream from current populationsmight be nonexistent. We recommend that thefuture management of this large aquatic turtlespecies in Oklahoma include: 1) continuedmonitoring of the species in the state, 2) con-tinued protection of known populations fromincidental and illegal take, and 3) reestablish-ment of extirpated populations.
The project was funded by the Federal Aid, En-dangered Species Act under Project E-40 of theOklahoma Department of Wildlife Conservationand Oklahoma State University, with additional fi-nancial and logistical support from Sequoyah Na-tional Wildlife Refuge and the Oklahoma Coopera-tive Fish and Wildlife Research Unit (OklahomaState University, Oklahoma Department of WildlifeConservation, United States Geological Survey Bio-logical Resources Division, and the Wildlife Manage-ment Institute cooperating). We also want to thankour field assistants and volunteers K. Cole, C. Finch-er, C. O’Melia, M. O’Melia, R. Stark, T. Talbot, andM. Stangland-Willis for all their help in ensuring thesuccess of this project.
LITERATURE CITED
ALLEN, E. R., AND W. T. NEILL. 1950. The alligatorsnapping turtle, Macroclemys temminckii, in Flori-da. Special Publication Number 4, Ross Allen’sReptile Institute, Silver Springs, Florida.
BLACK, J. H. 1982. An annotated bibliography to ar-ticles, notes and photographs on reptiles and am-phibians appearing in Oklahoma Game and FishNews, Oklahoma Wildlife, and OutdoorOklahoma. Oklahoma Herpetological Society,Special Publication Number 2.
CARPENTER, C. C., AND J. J. KRUPA. 1989. Oklahomaherpetology: an annotated bibliography. Univer-sity of Oklahoma Press, Norman.
CLARKE, R. F. 1981. A record of the alligator snap-ping turtle, Macroclemys temminckii (Testudines:Chelydridae), in Kansas. Transactions of the Kan-sas Academy of Science 84:59–60.
CONANT, R., AND J. T. COLLINS. 1998. A field guide toreptiles and amphibians: eastern and centralNorth America, third edition. Houghton MifflinCompany, Boston, Massachusetts.
ERNST, C. H., J. E. LOVICH, AND R. W. BARBOUR. 1994.Turtles of the United States and Canada. Smith-sonian Institution Press, Washington, D.C.
GALBREATH, E. C. 1961. Two alligator snappers, Ma-croclemys temminckii, from southern Illinois. Trans-actions of the Illinois State Academy of Science54:134–135.
GLASS, P. B. 1949. Macroclemys temminckii inOklahoma. Copeia 1949:138–141.

84 vol. 50, no. 1The Southwestern Naturalist
HECK, B. A. 1998. The alligator snapping turtle (Ma-croclemys temminckii) in southeast Oklahoma. Pro-ceedings of the Oklahoma Academy of Science78:53–58.
MOLL, E. O., AND D. MOLL. 2000. Conservation ofriver turtles. In: M. W. Klemens, editor. Turtleconservation. Smithsonian Institution Press,Washington, D.C. Pp. 126–155.
PRITCHARD, P. C. H. 1989. The alligator snapping tur-tle: biology and conservation. Milwaukee PublicMuseum, Milwaukee, Wisconsin.
REED, R. N., J. CONGDON, AND J. W. GIBBONS. 2002.The alligator snapping turtle [Macrochelys (Macro-clemys) temminckii]: a review of ecology, life his-tory, and conservation, with demographic analy-ses of the sustainability of take from wild popu-lations. Report, Division of Scientific Authority,United States Fish and Wildlife Service, Aiken,South Carolina.
RIEDLE, J. D. 2001. The ecology of the alligator snap-ping turtle, Macrochelys temminckii, in Oklahoma.Unpublished M.S. thesis, Oklahoma State Uni-versity, Stillwater.
ROMAN, J., AND B. W. BOWEN. 2000. The mock turtlesyndrome: genetic identification of turtle meat
purchased in the south-eastern United States ofAmerica. Animal Conservation 3:61–65.
SHIPMAN, P. A. 1993. Alligator snapping turtle, habi-tat selection, movements, and natural history insoutheast Kansas. Unpublished M.S. thesis, Em-poria State University, Emporia, Kansas.
SHIPMAN, P. A., D. R. EDDS, AND D. BLEX. 1991. Re-port on the recapture of an alligator snappingturtle (Macroclemys temminckii) in Kansas. KansasHerpetological Society Newsletter 85:8–9.
SHIPMAN, P. A., AND D. RIEDLE. 1994. Alligator snap-ping turtle, Macroclemys temminckii, trap, mark andrelease project 1994. Final Report, Missouri De-partment of Conservation, Jefferson City.
SLOAN, K. N., K. A. BUHLMANN, AND J. E. LOVICH.1995. Stomach contents of commercially harvest-ed adult alligator snapping turtles, Macroclemystemminckii. Chelonian Conservation and Biology2:96–99.
WEBB, R. G. 1970. Reptiles of Oklahoma. Universityof Oklahoma Press, Norman.
Submitted 8 January 2004. Accepted 17 May 2004.Associate Editor was Geoffrey C. Carpenter.
BLACK-NECKED STILT (HIMANTOPUS MEXICANUS) BREEDING RANGEEXTENSION IN MEXICO
ARMANDO J. CONTRERAS-BALDERAS*
Laboratorio de Ornitologıa, Facultad de Ciencias Biologicas, Universidad Autonoma de Nuevo Leon, A.P. 425,San Nicolas de los Garza, N. L., Mexico 66450
*Correspondent: [email protected]
ABSTRACT This report documents a breeding range extension of the black-necked stilt (Himan-topus mexicanus) in Coahuila, Mexico.
RESUMEN Se amplia la distribucion del area de reproduccion del candelero americano (Hi-mantopus mexicanus) en el estado de Coahuila, Mexico.
On 6 May 2002, I found a pair of black-necked stilts (Himantopus mexicanus) nesting atVenustiano Carranza (Don Martin) Reservoiron the Rio Sabinas (278319130N, 1008379570W).The nest was on the ground between rocks(Fig. 1). On 6 May, it had 3 eggs, and the nextday had 4 eggs. The 4 eggs averaged 42.7 331.6 mm. Six other pairs of black-necked stiltsalso were observed in the immediate area, but
I found no other nests. Friedmann et al.(1950) and Urban (1959) did not list this spe-cies for Coahuila. The American Ornitholo-gists’ Union (1998) and Howell and Webb(1995) showed the species as a non-breedingvisitor in southern Coahuila. Therefore, theseobservations document a northern extensionof the known breeding range in the interior ofnorthern Mexico and begin to fill the gap in

March 2005 85Notes
FIG. 1 Photos of black-necked stilt nest at DonMartin Reservoir, Coahuila, Mexico: top, 6 May2002; bottom, 7 May 2002.
the known breeding range between Mexicoand United States (Robinson et al., 1999).
LITERATURE CITED
AMERICAN ORNITHOLOGISTS’ UNION. 1998. Check-listof North American birds. American Ornitholo-gists’ Union, Washington, D.C.
FRIEDMANN, R., L. GRISCOM, AND R. T. MOORE. 1950.Distributional check-list of the birds of Mexico,part I. Pacific Coast Avifauna 29:1–202.
HOWELL, S. N. G., AND S. WEBB. 1995. A guide to thebirds of Mexico and northern Central America.Oxford University Press, New York.
ROBINSON, J. A., J. M. REED, J. P. SKORUPA, AND L. W.ORING. 1999. Black-necked stilt (Himantopus mex-icanus). In: A. Poole and F. Gill, editors. Thebirds of North America, number 449. Birds ofNorth America, Inc., Philadelphia, Pennsylvania.
URBAN, E. K. 1959. Birds from Coahuila, Mexico.University of Kansas, Publications of the Museumof Natural History 11(8):443–516
Submitted 9 December 2003. Accepted 22 March 2004.Associate Editor was Timothy Brush.
ATY PICAL PAIR-BONDING BEHAVIOR AMONG GOLDEN-FRONTEDWOODPECKERS (MELANERPES AURIFRONS)
MICHAEL S. HUSAK*
Department of Biology, Angelo State University, San Angelo, TX 76909Present Address: Department of Biological Sciences, Cameron University, 2800 W Gore Boulevard, Lawton, OK 73505
*Correspondent: [email protected]
ABSTRACT While conducting a study of golden-fronted woodpecker (Melanerpes aurifrons) ter-ritorial behavior and site fidelity in west-central Texas in 1996, I observed a color-banded maleinteracting with 2 females in a manner consistent with pair-bonding. This behavior, which waspreviously unreported for this species, was observed late in the breeding season and continuedthrough the following winter. No nesting attempts were observed, but the male and females didexhibit typical mate behavior, including the exchanging of mate-greeting calls and displays. Thefemales occupied separate, adjacent territories, both of which the male regularly occupied. Thechronology of events and potential factors influencing such behavior are discussed.
RESUMEN Durante un estudio sobre el comportamiento territorial y fidelidad al sitio del car-pintero cheje (Melanerpes aurifrons) en la parte central del oeste de Texas en 1996, observe laasociacion de un macho marcado con anillos de color con 2 hembras en una manera consistentecon la formacion de la pareja. Este comportamiento, que no se habıa registrado previamente enesta especie, se observo tarde en la temporada de apareamiento y continuo hasta el siguiente

86 vol. 50, no. 1The Southwestern Naturalist
invierno. No se observaron tentativas de nidificar, pero el macho y las hembras mostraron com-portamiento tıpico de parejas, incluyendo el intercambio de vocalizaciones de reconocimiento ycortejo. Las hembras ocuparon territorios separados y adyacentes, los cuales fueron tambien ocu-pados regularmente por el macho. Se discuten la cronologıa de eventos y los factores probablesque influencian dicho comportamiento.
As a family, woodpeckers are overwhelming-ly monogamous breeders (Short, 1982), oftenmaintaining individual pair bonds betweenbreeding seasons (Winkler and Christie, 2002).Even among cooperatively breeding species,extra-pair copulations are rare ( Jackson,1994). The scattered records of polygamy inthe family have predominantly been of classicalpolyandry (e.g., Willimont et al., 1991; Kotaka,1998; Wiebe, 2002). Ligon (1993) suggestedthat male participation was essential for nestsuccess among woodpeckers, and thus the lackof classical polygny. However, recent observa-tions suggest that polygyny is possible (e.g.,Wiktander et al., 2000; Conner et al., 2001).
Woodpeckers can exhibit considerable sea-sonal variation in social interactions and tol-erances (Short, 1982). Members of the NewWorld genus Melanerpes, in particular, exhibit adiversity of social interactions, ranging fromseasonal solitary to communal and cooperativebehavior (Short, 1982; Winkler and Christie,2002).
Golden-fronted woodpeckers (Melanerpes au-rifrons) are the least studied melanerpinewoodpecker in the United States, but as far asis known, they are monogamous and aggres-sively defend breeding territories against con-specifics (Husak and Maxwell, 1998). Duringthe non-breeding seasons, social interactionsare variable. Individuals can remain with mateson a territory, occupy individual territories,wander, make use of communal foraginggrounds in which conspecifics are mildly tol-erated in times of concentrated food abun-dance, or a combination of these behaviors(Husak, 2000). Here I report on a previouslyundescribed behavior for golden-frontedwoodpeckers: a male sharing a foraging terri-tory and maintaining pair bonds with multiplefemales during the late summer, fall, and earlywinter of 1996.
All observations were made during studies ofgolden-fronted woodpecker territorial behaviorand site fidelity at San Angelo State Park, TomGreen County, Texas (318319N, 1008339W), con-ducted from January 1995 through May 1998.
The 410-ha site was located along the NorthConcho River and included the North ConchoRiver Day Use Area, Bald Eagle Creek Camp-ground, and Wildlife Management Area 8. Veg-etation of the site was predominantly mesquite(Prosopis glandulosa) brushland of varying den-sity, associated with narrow bands of riparianwoodlands dominated by pecan (Carya illinoien-sis) along 3 waterways that pass through thearea. For details of the study area, see Husak(2000). Approximately 70% of all individuals onthe study site were color-banded. Two bandswere placed on each leg of adult and juvenilewoodpeckers that were subsequently known by4-letter codes representing their color combi-nation. Resident individuals that were not band-ed could readily be identified by unique varia-tions in the color of the nape and nasal tuftsand the unique pattern of white bands on theouter rectrices.
Male RSRS was banded as an after-hatch-yearbird in October 1995. He remained on thestudy site and attempted to breed with a color-banded female (YGYG) in spring 1996. Aftertheir nest failed on 17 June, the pair beganexploring new cavities and renewed courtshipbehavior, including mutual tapping, mounting,reverse mounting, and cavity excavation. How-ever, YGYG abandoned the study area duringthe last week of June before a second nestcould be initiated.
Within a week of the disappearance ofYGYG, an unbanded female of unknown originwas observed foraging without incident in theterritory of RSRS. By the middle of July, RSRSand the new female were observed movingabout the territory as a pair. The pair regularlyforaged in the same tree, actively defendedboundaries together, and maintained vocalcontact when apart. They also regularly ex-changed mate-greeting displays, includinghead-swinging, bill-pointing, and bill-waving(see Husak, 1996, for descriptions of displays).Neither copulation nor nesting was observed.
In late August, female YGYG returned to thestudy site for the first time since June. Follow-ing her return, YGYG and the unbanded fe-

March 2005 87Notes
male were observed fighting near the center ofthe territory of the male. By 3 September,YGYG occupied a territory adjacent to the east-ern edge of the territory of the unbanded fe-male, where a service road served as a bound-ary between the 2 females. For the following 6weeks, the 2 females maintained well-definedterritories, and the male divided his time be-tween the 2 territories. The male regularly ex-changed vocalizations and mate-greeting dis-plays with both females, and actively assistedboth females in defending their territoryboundaries from other intruding conspecifics.However, the male aided neither when the fe-males engaged in aggressive encounters withone another.
From mid October until mid November, ter-ritorial behavior greatly decreased on the studysite as large numbers of golden-fronted wood-peckers from adjacent areas began foragingalong waterways, exploiting the ripening pe-cans (Husak, 2000). RSRS, YGYG, and the un-banded female occupied home ranges consis-tent with previous territories described above.The unbanded female was last seen in the areaon 30 October; the reasons for her disappear-ance are unknown. By 3 November, anotherfemale (YSSS) was observed occupying muchof the former territory of the absent unbandedfemale. This female was submissive to RSRS,and within several days seemed to be acceptedby him and behaved much as a mate would,exchanging calls and displays, and foraging inclose proximity to one another. Once com-munal foraging came to an end in mid Novem-ber, females YGYG and YSSS began defendingtheir respective territories against one another,as well as other conspecifics, and continued ex-changing mate displays with RSRS. None ofthese individuals remained on the study site bythe end of winter, preventing further observa-tion.
Although nesting attempts were not ob-served, this does not necessarily mean that in-dividuals were not exhibiting pair-bonding be-havior for future reproductive potential. Gold-en-fronted woodpecker pair-bonding behaviordoes intensify during late February and March,but pair formation has been observed year-round (Husak and Maxwell, 1998). Too little isknown about the earlier history and ultimatefate of these individuals to allow conclusivestatements about why such behavior was ob-
served; however, the novelty of such observa-tions warrants documentation and invites fu-ture consideration. During the course of field-work, this was the only case of such interac-tions observed, suggesting it is rare amonggolden-fronted woodpeckers. However, it un-derscores the plasticity of behavior and socialinteraction within this poorly known species.
I thank J. Husak, C. Adkins, and C. Weaver forassistance with banding of golden-fronted wood-peckers. R. Conner, A. Husak, T. Maxwell, U. Wik-tander, and an anonymous reviewer provided nu-merous helpful comments that improved earlier ver-sions of this manuscript. I am most grateful to I.Vilchez Ramirez for Spanish translation of the ab-stract.
LITERATURE CITED
CONNER, R. N., J. R. MCCORMICK, R. R. SCHAEFER, D.SAENZ, AND D. C. RUDOLPH. 2001. A red-cockadedwoodpecker group with two simultaneous nesttrees. Wilson Bulletin 113:101–104.
HUSAK, M. S. 1996. Breeding season displays of thegolden-fronted woodpecker. Southwestern Natu-ralist 41:441–442.
HUSAK, M. S. 2000. Seasonal variation in territorialbehavior of the golden-fronted woodpecker inwest-central Texas. Southwestern Naturalist 45:30–38.
HUSAK, M. S., AND T. C. MAXWELL. 1998. Golden-fronted woodpecker (Melanerpes aurifrons). In: A.Poole and F. Gill, editors. The birds of NorthAmerica, number 373. Birds of North America,Inc., Philadelphia, Pennsylvania.
JACKSON, J. A. 1994. Red-cockaded woodpecker (Pi-coides borealis). In: A. Poole and F. Gill, editors.The Birds of North America, number 85. Acad-emy of Natural Sciences, Philadelphia, Pennsyl-vania, and American Ornithologists’ Union,Washington, D.C.
KOTAKA, N. 1998. Classical polyandry in the greatspotted woodpecker Dendrocopos major. Ibis 140:335–336.
LIGON, J. D. 1993. The role of phylogenetic historyin the evolution of contemporary avian matingand parental care systems. In: D. M. Powers, ed-itor. Current ornithology, volume 10. PlenumPress, New York. Pp. 1–46.
SHORT, L. L. 1982. Woodpeckers of the world. Del-aware Museum of Natural History MonographSeries, Number 4.
WIEBE, K. L. 2002. First reported case of classicalpolyandry in a North American woodpecker, thenorthern flicker. Wilson Bulletin 114:401–403.
WIKTANDER, U. O., O. OLSSON, AND S. G. NILSSON.

88 vol. 50, no. 1The Southwestern Naturalist
2000. Parental care and social mating system inthe lesser spotted woodpecker Dendrocopos minor.Journal of Avian Biology 31:447–456.
WILLIMONT, L. A., J. A. JACKSON, AND B. J. S. JACKSON.1991. Classical polyandry in the West Indianwoodpecker on Abaco, Bahamas. Wilson Bulletin103:124–125.
WINKLER, H., AND D. A. CHRISTIE. 2002. Family Pici-
dae (woodpeckers). In: J. del Hoyo, A. Elliott,and J. Sargatal, editors. Handbook of birds of theworld, volume 7. Jacamars to woodpeckers. LynxEdicions, Barcelona, Spain. Pp. 296–555.
Submitted 14 October 2003. Accepted 29 June 2004.Associate Editor was Timothy Brush.
ECTOPARASITES AND FOOD HABITS OF ELLIOT’S SHORT-TAILEDSHREW, BLARINA HYLOPHAGA
CHRISTOPHER M. RITZI,* BRIAN C. BARTELS, AND DALE W. SPARKS
Department of Ecology and Organismal Biology, Indiana State University, Terre Haute, IN 47809 (CMR, DWS)Department of Biological Sciences and Sternberg Museum of Natural History, Fort Hays State University,
Hays, KS 67601 (BCB)Present address of CMR: Department of Biology, Sul Ross State University, Alpine, TX 79832
*Correspondent: [email protected]
ABSTRACT The natural history of Elliot’s short-tailed shrew (Blarina hylophaga) has remainedvirtually unknown since it was recognized as a distinct species in 1981. For this study, we examined26 specimens of B. hylophaga for ectoparasites. We found 5 species of fleas, 1 species of tick, and17 species of mites. We also examined stomach contents of 30 specimens of B. hylophaga anddiscovered that insects (particularly beetles), slugs, and spiders were the most common food items.To determine the degree of similarity between B. hylophaga and the 2 other widely distributedshort-tailed shrews, we compared these data to the published ectoparasite and food habits datafor B. carolinensis and B. brevicauda. We found that both the ectoparasitic fauna and the diet of B.hylophaga broadly overlap those of its congeners. We also noted, however, that each species ofBlarina had parasites that were more closely associated with it than with the other 2 species ofBlarina. In addition, the diet of B. hylophaga consists of harder-bodied foods, containing morebeetles, fewer earthworms, and less fungi than that of the other 2 species.
RESUMEN La historia natural de la musarana Blarina hylophaga ha quedado virtualmente des-conocida desde que fue reconocida como una especie distinta en 1981. En este estudio exami-namos los ectoparasitos de 26 B. hylophaga. Encontramos 5 especies de pulga, 1 especie de gar-rapata, y 17 especies de acaro. Tambien examinamos los contenidos estomacales de 30 especı-menes de B. hylophaga y se descubrio que los insectos (particularmente escarabajos), babosas yaranas fueron la comida mas comun. Para determinar el grado de semejanza entre B. hylophaga ylas otras dos musaranas ampliamente distribuidas, comparamos estos datos con los publicados deectoparasitos y habitos alimenticios de B. carolinensis y B. brevicauda. Encontramos que ambos lafauna de ectoparasitos y la dieta de B. hylophaga se sobrelapan ampliamente con las de sus con-generes. Sin embargo, notamos tambien que cada especie de Blarina tuvo parasitos que estancercanamente asociados con la misma que con las otras 2 especies de Blarina. Ademas, la dietade B. hylophaga consiste de comida mas dura, conteniendo mas escarabajos, menos lombrices ymenos hongos que la de las otras 2 especies.
Taxonomic studies of Blarina during the past3 decades (Genoways and Choate, 1972;
George et al., 1981; George et al., 1982; Han-dley and Varn, 1994) have led to a split of the

March 2005 89Notes
northern short-tailed shrew, B. brevicauda, into3 species. As revised, the northern short-tailedshrew ranges from New Brunswick west to Sas-katchewan and south along the Appalachiansto Georgia in the east and to southern Ne-braska on the Great Plains. The southernshort-tailed shrew, B. carolinensis, occurs pri-marily on the Coastal Plain in the Southeast,and Elliot’s short-tailed shrew, B. hylophaga, isdistributed from southern Iowa through west-ern Kansas and south to Texas.
Unfortunately, this increase in the numberof species recognized has not led to detailednatural history studies of the re-elevated spe-cies (Genoways and Choate, 1998). Most of theavailable natural history data is based on themost widespread species, B. brevicauda, withsome information also available for B. caroli-nensis (Mumford and Whitaker, 1982; Geno-ways and Choate, 1998). The purposes of thisstudy were to conduct the first detailed studyof the diet and ectoparasites of B. hylophaga,and to compare these results with what isknown about B. brevicauda and B. carolinensis.
We obtained B. hylophaga for this study from2 sources. Seven shrews were unprepared spec-imens stored in the freezers of the SternbergMuseum of Natural History (formerly the Mu-seum of the High Plains, MHP) since as earlyas 1993; the other 23 specimens were collectedin summer 2002. All shrews were collected dur-ing the months of May through Septemberfrom north-central Kansas using line transectsof both Sherman live traps and Museum Spe-cials, plus pitfalls set against drift fences. Uponcapture, shrews were killed, individuallybagged, and stored as frozen carcasses at MHPuntil examination. All shrews were taken in ac-cordance with the American Society of Mam-malogists guidelines for animal care and use(Animal Care and Use Committee, 1998).
Twenty-six shrews were processed for ecto-parasites using the washing technique outlinedby Ritzi and Whitaker (2003). Hosts wereplaced in a glass jar with soapy water to cover,shaken vigorously for 1 to 2 minutes, and re-moved from the wash after being rinsed with70% ethanol. The wash was then vacuum fil-tered using filter paper and a Buchner funnel.While the wash was filtering, the jar and lidwere rinsed (into the funnel) with ethanol tomaximize ectoparasite recovery and preventcontamination between hosts. The filter papers
then were dried and examined for ectopara-sites using a 7–303 stereomicroscope, with allparasites tabulated by group. Representativevoucher specimens were slide-mounted in PVAmedium and identified using a combination ofkeys, including Krantz (1986), Whitaker(1982), Lukoschus et al. (1988), and Fain et al.(1991). Finally, shrews were examined under astereomicroscope to determine if any parasitesremained attached after the wash. Although noadditional ectoparasite species were recoveredat this time, the value of microscopy in locatingparasites embedded or firmly attached to thehost is well documented (Whitaker et al., 1993;Ritzi and Sparks, 2002). Voucher specimens ofboth parasites and shrews were deposited atboth MHP and the Indiana State UniversityVertebrate Collection.
Parasites obtained from B. hylophaga werecompared with data from B. brevicauda (Whi-taker and Mumford, 1972; Mumford and Whi-taker, 1982; Pascal, 1984) and B. carolinensis(Pascal, 1984; Whitaker et al., 1994) by usingChi-square analysis of 2 3 3 contingency tables(Zar, 1996). Parasitological terms follow thosesuggested in Bush et al. (1997); prevalencepertains to the percentage of hosts infestedwith a particular parasite, and mean intensityrefers to the average number of a particularparasite species per infested host.
The diet of 30 shrews was studied using thesame techniques as Whitaker et al. (1994).Briefly, we removed the stomachs and openedthem into watch glasses. Stomach contentswere removed, suspended in 75% ethanol, andexamined using a 7–303 stereomicroscope.The percentage of each food item in eachstomach was then estimated visually. Data weresummarized in terms of percent volume (theaverage percent of the diet of each shrew) andpercent frequency (percent of shrews that hadeaten a particular food item). We then used aMANOVA followed by a series of Student-New-man-Keuls multiple range tests to compare thediet of B. hylophaga in this study to data col-lected previously for B. brevicauda (Mumfordand Whitaker, 1982) and B. carolinensis (Whi-taker et al., 1994).
Ectoparasites Only 2 species of parasite areknown for B. hylophaga, a laelapid mite, Eulae-laps stabularis, from Kansas ( Jameson, 1947),and a flea, Stenoponia americana, from Kansas(Poorbaugh and Gier, 1961). Both records
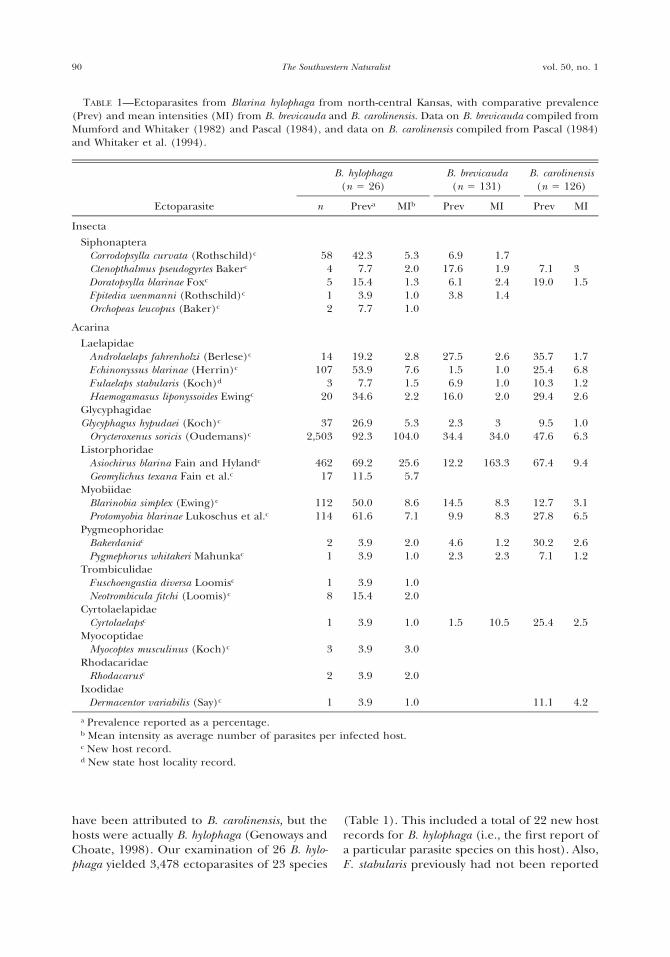
90 vol. 50, no. 1The Southwestern Naturalist
TABLE 1—Ectoparasites from Blarina hylophaga from north-central Kansas, with comparative prevalence(Prev) and mean intensities (MI) from B. brevicauda and B. carolinensis. Data on B. brevicauda compiled fromMumford and Whitaker (1982) and Pascal (1984), and data on B. carolinensis compiled from Pascal (1984)and Whitaker et al. (1994).
Ectoparasite
B. hylophaga(n 5 26)
n Preva MIb
B. brevicauda(n 5 131)
Prev MI
B. carolinensis(n 5 126)
Prev MI
Insecta
SiphonapteraCorrodopsylla curvata (Rothschild)c
Ctenopthalmus pseudogyrtes Bakerc
Doratopsylla blarinae Foxc
Epitedia wenmanni (Rothschild)c
Orchopeas leucopus (Baker)c
584512
42.37.7
15.43.97.7
5.32.01.31.01.0
6.917.66.13.8
1.71.92.41.4
7.119.0
31.5
Acarina
LaelapidaeAndrolaelaps fahrenholzi (Berlese)c
Echinonyssus blarinae (Herrin)c
Eulaelaps stabularis (Koch)d
Haemogamasus liponyssoides Ewingc
14107
320
19.253.97.7
34.6
2.87.61.52.2
27.51.56.9
16.0
2.61.01.02.0
35.725.410.329.4
1.76.81.22.6
GlycyphagidaeGlycyphagus hypudaei (Koch)c
Orycteroxenus soricis (Oudemans)c37
2,50326.992.3
5.3104.0
2.334.4
334.0
9.547.6
1.06.3
ListorphoridaeAsiochirus blarina Fain and Hylandc
Geomylichus texana Fain et al.c46217
69.211.5
25.65.7
12.2 163.3 67.4 9.4
MyobiidaeBlarinobia simplex (Ewing)c
Protomyobia blarinae Lukoschus et al.c112114
50.061.6
8.67.1
14.59.9
8.38.3
12.727.8
3.16.5
PygmeophoridaeBakerdaniac
Pygmephorus whitakeri Mahunkac21
3.93.9
2.01.0
4.62.3
1.22.3
30.27.1
2.61.2
TrombiculidaeEuschoengastia diversa Loomisc
Neotrombicula fitchi (Loomis)c18
3.915.4
1.02.0
CyrtolaelapidaeCyrtolaelapsc 1 3.9 1.0 1.5 10.5 25.4 2.5
MyocoptidaeMyocoptes musculinus (Koch)c 3 3.9 3.0
RhodacaridaeRhodacarusc 2 3.9 2.0
IxodidaeDermacentor variabilis (Say)c 1 3.9 1.0 11.1 4.2
a Prevalence reported as a percentage.b Mean intensity as average number of parasites per infected host.c New host record.d New state host locality record.
have been attributed to B. carolinensis, but thehosts were actually B. hylophaga (Genoways andChoate, 1998). Our examination of 26 B. hylo-phaga yielded 3,478 ectoparasites of 23 species
(Table 1). This included a total of 22 new hostrecords for B. hylophaga (i.e., the first report ofa particular parasite species on this host). Also,E. stabularis previously had not been reported

March 2005 91Notes
TABLE 2—Food habits of 25 Elliot’s short-tailedshrews (Blarina hylophaga) from north-central Kan-sas. All animals (Metazoa) are adults unless other-wise indicated. Volume (Vol.) is the average per-centage of the stomach contents for each item;frequency (Freq.) is the percentage of shrews whosestomachs contained a food item.
FoodVol.(%)
Freq.(%)
Metazoa (total)Arthropoda (total)
Insecta (total)Insecta (unknown)Coleoptera (total)
97.071.861.64.6
50.3
100100961696
Coleoptera (larvae)Coleoptera (unknown)
CarabidaeScarabidae (total)Scarabidae (unknown)
17.03.6
14.114.012.6
3216365252
CotinisPhanaeus
CurculionidaeLepidopteraDiptera
0.21.21.6
trace0.4
44848
IsopteraOrthoptera (Gryllidae)Homoptera (Cicadellidae)
DraeculaecephalisHymenoptera (Formicidae)
2.42.4
0.60.8
48
44
Arachnida (Araneae)Isopoda
Mollusca (Gastropoda)Chordata (Rodentia)
Peromyscus maniculatus?
7.33.0
21.2
4.0
324
24
4Plantae (total)
Forbs (unknown)Poaceae
Fungi (total)Basidiomycota (unknown)Zygomycota (cf. Endogone)
1.00.60.42.01.20.8
1288
1288
from B. hylophaga from Kansas and, thus, is anew host-locality record for the state. We didnot recover S. americana.
The 24 species of ectoparasites now knownfrom B. hylophaga contrast with 44 species as-sociated with B. carolinensis and 120 species as-sociated with B. brevicauda. Of these 24 para-sites, B. hylophaga shares 16 with B. carolinensisand 21 with B. brevicauda. Fifteen species ofparasites are shared by all 3 host species. Theonly parasites recovered exclusively from B. hy-lophaga were Geomylichus texana, Neotrombiculafitchi, and Myocoptes musculinus. Both N. fitchiand M. musculinus commonly are associatedwith other hosts native to the Great Plains, andin our opinion, they represent accidental oc-currences. The 17 specimens of G. texana col-lected from 3 shrews captured in Rooks Coun-ty, Kansas, are more difficult to explain. Listro-phorid mites are highly co-evolved with theirhosts, and host switching typically results inspeciation events (Fain, 1994). The specimensof G. texana recovered during this study aremorphologically identical to museum speci-mens obtained from Ord’s kangaroo rat (Di-podomys ordii). Resolving this relationshipshould be a goal of future studies.
Although the 3 species of Blarina share manyparasites, there is interspecific variation in thecomposition of these communities. In partic-ular, each species of Blarina tends to be infest-ed with different flea species: Ctenophthalmuspseudagyrtes on B. brevicauda (x2
(df52) 5 39.7, P, 0.001), Doratopsylla blarinae on B. carolinensis(x2
(df52) 5 6.5, P 5 0.05), and Corrodopsylla cur-vata on B. hylophaga (x2
(df52) 5 459.7, P ,0.001). Laelapid mites also provide evidence ofinterspecific variation. Echinonyssus blarinae wasmore often associated with B. hylophaga (x2
(df52)
5 386.2, P , 0.001) than the other 2 speciesof Blarina, whereas H. liponyssoides was foundmore often on B. carolinensis than expected(x2
(df52) 5 24.0, P , 0.001). The glycyphagidmites Glycyphagus hypudaei and Orycteroxenus sor-icis were both significantly more common onB. hylophaga (x2
(df52) 5 273.5, P , 0.001 andx2
(df52) 5 12,598.2, P , 0.001, respectively)than on the other 2 hosts, as were the myobiidmites, Blarinobia simplex (x2
(df52) 5 302.5, P ,0.001) and Protomyobia blarinae (x2
(df52) 5 1,843,P , 0.001). Finally, the listrophorid mite Asi-ochirus blarina was found in large numbers onB. brevicauda, moderate numbers on B. hylopha-
ga, and low numbers on B. carolinensis (x2(df52)
5 928.4, P , 0.001). The other mites and in-sects found on all 3 hosts were distributedequally across the 3 species.
Diet Twenty-five stomachs contained food,whereas 5 were empty and excluded from fur-ther analysis (Table 2). Insects (61.6% volume,100% occurrence), especially beetles (50.3%and 96%, respectively), were the most com-mon food, followed by slugs (21.2% and 24%,respectively) and spiders (7.3% and 32%, re-spectively).
The diet of B. hylophaga was similar in most

92 vol. 50, no. 1The Southwestern Naturalist
TABLE 3—Summarized food habits for 3 species ofBlarina reported as untransformed mean percentvolumes of each food group. Data for this summaryare available in Mumford and Whitaker (1982) forB. brevicauda, Whitaker et al. (1994) for B. carolinen-sis, and Table 2 for B. hylophaga. Significantly distinctgroups (Student-Newman-Keuls multiple range tests,a 5 0.05) are denoted by letters (a, b, c) followingpercent volumes. Percent values with multiple lettersrepresent cases where a value was indistinguishablefrom more than 1 other species.
Food item
Volume (%)
B. brevi-cauda
(n 5 125)
B. caro-linensis
(n 5 41)
B. hylo-phaga
(n 5 25)
ColeopteraLarvaAdult
5.6 a8.7 a
8.2 ab12.0 a
17.0 b33.3 b
LepidopteraLarva
OrthopteraOther ArthropodaAnnelida
8.28.1
17.835.7 a
3.72.1
22.714.8 b
0.02.4
19.10.0 c
GastropodaVertebrataPlantaeFungi
8.60.13.63.6 a
18.01.01.2
16.3 b
21.24.01.02.0 a
respects to that of the other species of Blarina,consisting primarily of animal matter, althoughthe shrews occasionally take small amounts offungi and higher plants. To make direct com-parisons among and between the 3 species ofshrews, we summarized the data on food habitsfor the 3 species into 10 categories (Table 3).A MANOVA revealed that the overall diet ofthe 3 shrews varied significantly between spe-cies (l 5 0.581, P , 0.001). A series of Student-Newman-Keuls multiple range tests indicatedthat the 3 species all varied significantly (a 50.05) from one another in terms of the per-cent volume of earthworm in the diet. Earth-worms were the most common individual fooditem for B. brevicauda in Indiana, while wefailed to record any in the diet of B. hylophagafrom Kansas. Blarina hylophaga consumed moreadult beetles than the other species, and morebeetle larvae than B. brevicauda. Blarina caroli-nensis consumed more fungi than the otherspecies. These observations suggest that hard-ness of the diet of these species range from B.hylophaga, which focuses on hard-bodied food
items, to B. brevicauda, which consumes rela-tively soft food items. Studies of shrews in ge-nus Sorex from the Pacific Northwest (Carrawayand Verts, 1994; Carraway et al., 1996) have re-vealed relationships between the diets of theseshrews and their ability to generate bite force.Thus, we suggest mensural data collected dur-ing earlier taxonomic studies of Blarina be re-examined in light of the functional morphol-ogy of the jaws.
We thank Fort Hays State University, the Stern-berg Museum of Natural History, and Indiana StateUniversity for enabling this research. J. R. Choate, J.R. Thomasson, and E. Gillock provided logisticalsupport and access to specimens. Thanks go to G.Liggett and C. Liggett for housing CMR and DWSwhile conducting this study at the Sternberg Muse-um of Natural History. Thanks to J. R Choate, J. O.Whitaker, Jr., and an anonymous reviewer for im-proving earlier versions of this manuscript, and to J.H. Magers for aid in composing the Spanish resu-men. This study was funded primarily by a grant toDWS and CMR from the Indiana Academy of Sci-ence. Additional funding was provided to CMR fromthe Indiana State University Office of SponsoredPrograms Student Research Scholarship, and the In-diana State University Graduate Research Fund.DWS was supported during this work by a grant fromthe Indianapolis International Airport.
LITERATURE CITED
ANIMAL CARE AND USE COMMITTEE. 1998. Guidelinesfor the capture, handling, and care of mammalsas approved by the American Society of Mam-malogists. Journal of Mammalogy 79:1416–1431.
BUSH, A. O., K. D. LAFFERTY, J. M. LOTZ, AND A. W.SHOSTAK. 1997. Parasitology meets ecology on itsown terms: Margolis et al. revisited. Journal ofParasitology 83:575–583.
CARRAWAY, L. J., AND B. J. VERTS. 1994. Relationshipof mandibular morphology to relative bite-forcein some western Sorex. Carnegie Museum of Nat-ural History Special Publications 18:201–210.
CARRAWAY, L. J., B. J. VERTS, M. L. JONES, AND J. O.WHITAKER, JR. 1996. A search for age-relatedchanges in bite force and diet in shrews. Ameri-can Midland Naturalist 135:231–240.
FAIN, A. 1994. Adaptation, specificity and host-para-site coevolution in mites (Acari). InternationalJournal of Parasitology 24:1273–1283.
FAIN, A., J. O. WHITAKER, JR., AND H. H. THOMAS.1991. Two new fur-mites of the genus GeomylichusFain, 1970 (Acari, Listrophoridae) from kanga-roo rats, Dipodomys spp., from the U.S.A. Inter-national Journal of Acarology 17:175–180.

March 2005 93Notes
GENOWAYS, H. H., AND J. R. CHOATE. 1972. A multi-variate analysis of systematic relationships amongpopulations of the short-tailed shrew (genus Blar-ina) in Nebraska. Systematic Zoology 21:106–116.
GENOWAYS, H. H., AND J. R. CHOATE. 1998. Naturalhistory of the southern short-tailed shrew, Blarinacarolinensis. Occasional Papers of the Museum ofSouthwestern Biology 8:1–43.
GEORGE, S. B., J. R. CHOATE, AND H. H. GENOWAYS.1981. Distribution and taxonomic status of Blari-na hylophaga Elliot (Insectivora: Soricidae). An-nals of the Carnegie Museum, 50:493–513.
GEORGE, S. B., H. H. GENOWAYS, J. R. CHOATE, AND
R. J. BAKER. 1982. Karyotypic relationships withinthe short-tailed shrews, genus Blarina. Journal ofMammalogy, 63:639–645.
HANDLEY, C. O., JR., AND M. VARN. 1994. Identifica-tion of the Carolinian shrews of Bachman 1837.In: J. F. Merritt, G. L. Kirkland, Jr., and R. K.Rose, editors. Advances in the biology of shrews.Special Publication 18, Carnegie Museum of Nat-ural History, Pittsburgh, Pennsylvania. Pp. 393–406.
JAMESON, E. W., JR. 1947. Natural history of the prai-rie vole (mammalian genus Microtus). Universityof Kansas Publication, Museum of Natural His-tory 1:125–151.
KRANTZ, G. W. 1986. A manual of acarology, secondedition. Oregon State University Book Stores,Inc., Corvallis.
LUKOSCHUS, F. S., G. J. JEUCKEN, AND J. O. WHITAKER,JR. 1988. A review of the Protomyobia americanagroup (Acarina: Prostigmata: Myobiidae) with de-scriptions of Protomyobia panamensis n. sp. and Pro-tomyobia blarinae n. sp. Journal of Parasitology 74:305–316.
MUMFORD, R. E., AND WHITAKER, J. O., JR. 1982. Mam-mals of Indiana. Indiana University Press, Bloo-mington.
PASCAL, D. D., JR. 1984. A taxonomic study of Mid-western short-tailed shrews (genus Blarina) withemphasis upon ectoparasites. Unpublished Ph.D.dissertation, Indiana State University, TerreHaute.
POORBAUGH, J. H., AND H. T. GIER. 1961. Fleas (Si-phonaptera) of small mammals in Kansas. Jour-nal of the Kansas Entomological Society 34:198–204.
RITZI, C. M., AND D. W. SPARKS. 2002. New ectopar-asite records for the rock squirrel, Spermophilusvariegatus grammurus, in Socorro County, NewMexico. Southwestern Entomologist 27:257–262.
RITZI, C. M., AND J. O. WHITAKER, JR. 2003. Ectopar-asites of small mammals from the NewportChemical Depot, Vermillion County, Indiana.Northeastern Naturalist 10:149–158.
WHITAKER, J. O., JR. 1982. Ectoparasites of mammalsof Indiana. Monograph Number 4, Indiana Acad-emy of Science.
WHITAKER, J. O., JR., G. D. HARTMAN, AND R. HEIN.1994. Food and ectoparasites of the southernshort-tailed shrew, Blarina carolinensis (Mamma-lia: Soricidae), from South Carolina. Brimleyana21:97–105.
WHITAKER, J. O., JR., AND R. E. MUMFORD. 1972. Foodand ectoparasites of Indiana shrews. Journal ofMammalogy 53:329–335.
WHITAKER, J. O., JR., W. J. WRENN, AND R. E. LEWIS.1993. Parasites. In: H. H. Genoways and J. H.Brown, editors. Biology of the Heteromyidae.Special Publication Number 10, American Soci-ety of Mammalogists. Pp. 386–478.
ZAR, J. H. 1996. Biostatistical analysis, third edition.Prentice Hall, Upper Saddle River, New Jersey.
Submitted 11 November 2003. Accepted 2 April 2004.Associate Editor was Cody W. Edwards.

94 vol. 50, no. 1The Southwestern Naturalist
NEST RECORD OF SOREX MILLERI FROMMADERAS DEL CARMEN, MEXICO
JONAS A. DELGADILLO VILLALOBOS, BONNIE REYNOLDS MCKINNEY,*FELICIANAO HEREDIA PINEDA, AND SANTIAGO GIBERT ISERN
Proyecto El Carmen, Cemex Central S.A. de C.V., Ave. Constitucıon 444 Pte. 64000 Monterrey, Nuevo Leon, Mexico(JDV, BRM, FHP, SGI)
Present address of BRM: Proyecto El Carmen, P.O. Box 420608, Del Rio, TX 78842*Correspondent: [email protected]
ABSTRACT We report on the first documented nest of Miller’s shrew (Sorex milleri) from theMaderas del Carmen, Coahuila, Mexico. We located 7 nests in mesic montane forest at 2,400 to2,700 m elevation during October and November 2003.
RESUMEN Reportamos el primer nido documentado de la musarana (Sorex milleri), de Maderasdel Carmen, Coahuila, Mexico. Se localizo un total de 7 nidos en las mesetas boscosas humedasde 2,400 hasta 2,700 m de elevacion durante octubre y noviembre del 2003.
We report on the first documented nest ofMiller’s shrew (Sorex milleri) and provide addi-tional vouchered records of the occurrence ofthis species in Maderas del Carmen, Coahuila,Mexico. Little information on the life historyof this species is known for northern Coahuila.While collecting mammals during October andNovember 1940, F. W. Miller obtained 3 spec-imens of Miller’s shrew, including the typespecimen from the Maderas del Carmen atCampo Madera, an abandoned lumber campin a mesic montane forest dominated by Doug-las-fir (Pseudotsuga menziesii), Coahuila fir (Abiescoahuilensis), and southwestern white pine (Pi-nus strobiformis) ( Jackson, 1947; Baker, 1956).Baker (1956) captured 23 Miller’s shrews insoutheastern Coahuila about 20 km east of SanAntonio de las Alazanas at an elevation of2,805 m in an association of Douglas-fir, pine,and aspen (Populus tremuloides). Findley (1955)compared the Miller’s shrew that Jackson col-lected to other shrews and considered the Mill-er’s shrew to be a relict population of S. ciner-eus isolated in the Sierra Madre Oriental innortheastern Mexico. Miller’s shrew has beendocumented from Cerro Potosı in the adjacentstate of Nuevo Leon, which is part of the SierraMadre Oriental (Baker, 1956). Miller’s shrew iscurrently listed as ‘‘protected and endemic’’ inthe NORMA Oficial Mexicana (NOM, 2002).
Maderas del Carmen is a mountain range lo-cated 60 km south of Big Bend National Park
in western Texas and 165 km northwest ofMuzquiz, Coahuila, Mexico (298049060N,1028379170W). Maderas del Carmen is a typicalsky island surrounded by Chihuahuan Desertwith a range in elevation from 900 m to 2,700m. Dominant trees above 2,000 m are fir, pine-oak, oak-pine, and pine associations.
In October 2003, while we were conductingsmall mammal surveys as part of the baselineinventory of the Maderas del Carmen, wefound an active Miller’s shrew nest at CuadroPelota (298569940N, 1028359140W). The nestwas located in a forest opening with decayingtree trunks and logs surrounded by mesicmontane forest at an elevation of 2,400 m.Ground cover consisted of grasses, includingpoverty oat grass (Danthonia spicata), tobosagrass (Hilaria mutica), bull muhly (Muhlenbergiaemersleyi), and Pringle needlegrass (Piptochae-tum pringlei), with scattered pines, Douglas-fir,and silverleaf oak (Quercus hypoleucoides). Lo-cated at the entrance of a Botta’s pocket go-pher (Thomomys bottae) burrow under a rottinglog 25 cm in diameter, nest measurement fromthe outside edges was 10 cm in diameter acrossthe top, which was open and flush with theunderside of the log. The nest showed visiblesigns of wear and was a cup-like, loosely con-structed mass of grass blades. Pine needles, bitsof bark, and oak leaves were around the bot-tom of the nest. The nest contained a side en-trance hole that was 19 mm in diameter.

March 2005 95Notes
The nest contained 1 adult Miller’s shrew,which fled into the Botta’s pocket gopher tun-nel, and 6 young that scrambled from the nestwhen disturbed. We hand-captured the young,photographed them, and retained 2 for vouch-er specimens (Proyecto El Carmen, Registrode Coleccion Cientifica, Clave: NL-MAM-139-11-02; 046, 047). The adult Miller’s shrew wasbrownish gray, with an elongated nose, and waslarger than the young. The young had theireyes closed, their elongated noses were fleshypink, the pelage was short, soft, and chocolatebrown, the tail was short and hairless. Whenthe vouchers were prepared, we noted theskulls of the 2 specimens were not ossified. Thetotal length, tail, hindfoot, and ear of the 2voucher specimens were: 82, 83 mm; 28, 29mm; 9, 9 mm; and 4, 4 mm, respectively. Othersmall mammals collected in the same localitywere Botta’s pocket gopher, yellow-nosed cot-ton rat (Sigmodon ochrognathus ochrognathus),brush mouse (Peromyscus boylii), and westernharvest mouse (Reithrodontomys megalotis).
On the same day, 2 additional nests werefound at Mesa Bonita (238009670N,1028369760W), 12 km north of Cuadro Pelotaat 2,700 m in similar habitat. Four additionalunoccupied nests were located on 8 November2003 at Mesa Bonita. These 6 nests were con-structed in the same manner with the samenest materials, and all were under decayinglogs in old Botta’s pocket gopher tunnels.These nests were in various stages of disarrayfrom use, and 1 nest contained several smallfeathers.
Intense baseline inventory for 2 years (2001to 2003) in the Maderas del Carmen revealedthat the desert shrew (Notiosorex crawfordi) in-
habits the lower desert elevations to the highergrasslands that are interspersed with beakedyucca (Yucca rostrata), sotol (Dasylirion leiophyl-lum), and junipers (Juniperus) from 1,000 to1,415 m, and the Miller’s shrew inhabits themesic montane forest from 1,900 m to 2,700 min the Maderas del Carmen in northern Coa-huila, Mexico.
This research project was funded by Cemex andconducted as part of the Proyecto El Carmen withinthe Greater Maderas Del Carmen Ecosystem. Wegreatly appreciate the unwavering support of Ce-mex, which has made a long-term commitment tothe conservation of wildlife in Mexico. We thank L.Harveson and R. Valdez for reviewing the manu-script. We especially thank R. Baker for his insightfulsuggestions and inspiration, and in recognition ofhis studies on the mammals of Coahuila.
LITERATURE CITED
BAKER, R. H. 1956. Mammals of Coahuila, Mexico.University of Kansas Publication, Museum of Nat-ural History 9:125–335.
FINDLEY, J. S. 1955. Taxonomy and distribution ofsome American shrews. University of Kansas Pub-lication, Museum of Natural History 7:613–618.
JACKSON, H. H. T. 1947. A new shrew (genus Sorex)from Coahuila. Proceedings of the Biological So-ciety of Washington 60:131–132.
NOM. 2002. NORMA Oficial Mexicana. NOM-059-ECOL-2001, Proteccion ambiental nativas deMexico de flora y fauna silvestres-Categorıas deriegso y especificaciones para su inclusion, exclu-sion o cambio-Lista de especies en riesgo. Segun-da Seccion, Secretaria de Medio Ambiente y Re-cursos Naturales 2:1–81.
Submitted 9 December 2003. Accepted 30 April 2004.Associate Editor was Cody W. Edwards.

96 vol. 50, no. 1The Southwestern Naturalist
THREE NEW RECORDS OF BATS FROM EL SALVADOR
JAMES G. OWEN
Universidad Salvadorena ‘‘Alberto Masferrer,’’ A.P. 2053, San Salvador, El Salvador, Central America*Correspondent: [email protected]
ABSTRACT I report distributional records that document the presence of 3 additional speciesof bats (Carollia sowelli, Lasiurus ega, and Eptesicus furinalis) for the fauna of El Salvador, CentralAmerica.
RESUMEN Se reportan registros de distribucion que documentan la presencia de 3 especiesadicionales de murcielagos (Carollia sowelli, Lasiurus ega, y Eptesicus furinalis) para la fauna de ElSalvador, Centroamerica.
Geographical range maps in Hall (1981)and Reid (1997) tacitly predict the occurrenceof Carollia sowelli (Sowell’s short-tailed bat),Lasiurus ega (southern yellow bat), and Eptesi-cus furinalis (Argentine brown bat) in El Sal-vador. Although each of these species is widelydistributed in Central America (Hall, 1981;Reid, 1997), none has actually been docu-mented from this country. I report the firstspecimens of these bats from El Salvador.These 3 records bring the total number of spe-cies of bats reported for El Salvador to 62 (Ta-ble 1), representing about 50% of the totalmammalian fauna of the country (Felten,1955, 1956a, 1956b, 1956c, 1957; Burt and Stir-ton, 1961; Davis, 1968; Jones and Bleier, 1974;Hellebuyck et al., 1985; Owen et al., 1991,1993; Engstrom et al., 1994).
I examined specimens cited in this report,and they are now deposited in the followinginstitutions: Carnegie Museum of Natural His-tory (CM), Museo de Historia Natural de ElSalvador (MUHNES), Texas Cooperative Wild-life Collection (TCWC), and the Museum ofTexas Tech University (TTU). Capitalized lifezones are those of Holdridge (1975). Eleva-tions and geographical coordinates are thoserecorded by the original collectors, when avail-able, otherwise I estimated them using topo-graphical maps at a scale of 1 to 50,000, pub-lished by the Salvadoran Ministry of PublicWorks.
A collection of bats of the genus Carolliafrom El Salvador includes the species C. perspi-cillata, C. sowelli, and C. subrufa. Specimens ofC. sowelli from western Panama northward, in-
cluding El Salvador, were previously referredto C. brevicauda (Pine, 1972). Central Americanpopulations of C. brevicauda are now thoughtto be restricted to eastern Panama (Wright etal., 1999; Baker et al., 2002). Carollia sowelli isrepresented by 6 males (CM 114837–114841,114843) and 6 females (CM 114836, 114842,114844–144847) from northwestern El Salva-dor, 3.4 km S La Palma, Department of Chal-atenango (148179N, 898099W; 980 m elevation;Subtropical Very Moist Forest). The samplesize, mean, and range for length of forearmand greatest length of skull, respectively, are: n5 12, mean 5 40.5 mm, range 5 39.8 to 41.4mm; n 5 12, mean 5 22.5 mm, range 5 22.3to 22.9 mm. These measurements, as well asdistinctive characteristics of the pelage, matchthose presented by Pine (1972) and Owen etal. (1984) for Middle American C. sowelli,which they recorded as C. brevicauda.
At the latitude of northwestern El Salvador,C. sowelli has been collected from the Atlanticversant (Nueva Ocotepeque, Honduras, 840 melevation) down to the Caribbean lowlands(Pine, 1972; Hall, 1981). This report docu-ments its occurrence at about the same lati-tude as Nueva Ocotepeque from the Pacificversant as well. This species might occur acrossmuch of northern El Salvador, where it prob-ably is restricted to humid montane localities.In the area of the Gulf of Fonseca, C. sowelli isknown from the Pacific lowlands of Honduras,6 km east of El Amatillo, El Salvador, 60 melevation (Pine, 1972; Hall, 1981). I predictthat this species also occurs in the Pacific low-lands of El Salvador, in the eastern Depart-ment of La Union.

March 2005 97Notes
TABLE 1—Species of bats collected in El Salvador.
Taxon1,2,3 Common name4
Emballonuridae
Balantiopteryx plicata Gray sac-winged bagPeropteryx macrotis Lesser dog-like batRhynchonycteris naso Proboscis batSaccopteryx bilineata Greater sac-winged batS. leptura Lesser sac-winged bat
Noctilionidae
Noctilio albiventris Lesser bulldog batN. leporinus Greater bulldog bat
Mormoopidae
Mormoops megalophylla Ghost-faced batPteronotus davyi Davy’s naked-backed batP. gymnonotus Big naked-backed batP. parnellii Parnell’s mustached batP. personatus Wagner’s mustached bat
Phyllostomidae
MicronycterinaeMicronycteris microtis Little big-eared bat
DesmodontinaeDesmodus rotundus Vampire batDiaemus youngi White-winged vampire
batDiphylla ecaudata Hairy-legged vampire bat
LonchorhininaeLonchorhina aurita Tomes’ sword-nosed bat
PhyllostominaeChrotopterus auritus Big-eared woolly batMacrophyllum macro-
phyllum Long-legged batPhyllostomus discolor Pale spear-nosed batTrachops cirrhosus Fringe-lipped bat
GlossophaginaeAnoura geoffroyi Geoffroy’s tailless batChoeroniscus godmani Godman’s long-tailed batGlossophaga commissar-
isiCommissariss’ long-
tongued batG. leachii Gray long-tongued batG. soricina Pallas’ long-tongued batLeptonycteris curasoae Southern long-nosed bat
CarolliinaeCarollia perspicillata Seba’s short-tailed batC. sowelli Sowell’s short-tailed batC. subrufa Gray short-tailed bat
StenodermatinaeArtibeus inopinatus Honduran fruit-eating
batA. intermedius Intermediate fruit-eating
batA. jamaicensis Jamaican fruit-eating batA. lituratus Great fruit-eating batCenturio senex Wrinkle-faced batChiroderma salvini Salvin’s big-eyed bat
TABLE 1—Continued.
Taxon1,2,3 Common name4
C. villosum Hairy big-eyed batDermanura azteca Aztec fruit-eating batD. phaeotis Pygmy fruit-eating batD. tolteca Toltec fruit-eating batEnchisthenes hartii Velvety fruit-eating batPlatyrrhinus helleri Heller’s broad-nosed batSturnira lilium Little yellow-shouldered
batS. ludovici Highland yellow-shoul-
dered batUroderma bilobatum Tent-making batU. magnirostrum Brown tent-making bat
Natalidae
Natalus stramineus Mexican funnel-earedbat
Vespertilionidae
Eptesicus furinalis Argentine brown batE. fuscus Big brown batLasiurus blossevillii Western red batL. ega Southern yellow batL. intermedius Northern yellow batMyotis elegans Elegant myotisM. keaysi Hairy-legged myotisM. nigricans Black myotisM. velifer Cave myotisRhogeessa tumida Black-winged little yellow
bat
Molossidae
Eumops auripendulus Black bonneted batE. underwoodi Underwood’s bonneted
batMolossus rufus Black mastiff batM. molossus Pallas’ mastiff batNyctinomops laticauda-
tus Broad-eared bat
1 Subfamilies of the family Phyllostomidae arethose proposed by Baker et al. (2003).
2 Myotis fortidens, the cinnamon myotis, has beencollected from the Pacific lowlands of Guatemala,about 23 kilometers from the border of El Salvador(Dolan and Carter, 1979) and by Dickerman et al.(1981) at Montufar, Guatemala, near the southwest-ern border of El Salvador. Based on geographicaland habitat considerations, M. fortidens probably alsooccurs in the adjacent lowlands of El Salvador.
3 Molecular data suggest that A. intermedius mightnot be specifically distinct from A. lituratus (Lim etal., 2004).
4 Common names mostly follow Wilson and Cole(2000).

98 vol. 50, no. 1The Southwestern Naturalist
On 27 February 1967, a female Lasiurus ega(TCWC 19749) was collected from north-cen-tral El Salvador 20 km W Chalatenango, De-partment of Chalatenango (148069N, 898069W;250 m elevation; Subtropical Moist Forest).The capture of this specimen in winter sug-gests that L. ega has migratory movements, atleast on a local geographical scale, similar tothose of other North American lasiurines. Las-iurus ega resembles L. intermedius, but in areasof geographical overlap, L. ega is smaller (Halland Jones, 1961). Length of forearm of thisspecimen is 47.97 mm, which is well below themean and range of 2 specimens of L. interme-dius (1 from western El Salvador and 1 syntop-ically with this specimen of L. ega) n 5 2, mean5 54.95 mm, range 5 53.04 and 56.86 mm.
Lasiurus ega occurs from lowlands (Handley,1976) to elevations of up to about 2,300 m(Genoways and Jones, 1968). Based on therange of elevations from which specimens ofthis species have been collected and based onits wide geographical distribution in CentralAmerica (Dickerman et al., 1981; Hall, 1981;Dinerstein, 1985; McCarthy, 1987), I predictthat L. ega occurs throughout El Salvador, ex-cept in the highest cloud forest.
Specimens of Eptesicus furinalis have beencollected at 8 scattered localities in central ElSalvador as follows: 1 male (TTU 13371) and3 females (TTU 13372, 13373, 13374) 1.9 kmW Suchitoto, Department of Cuscatlan(138569N, 898039W; 420 m elevation; Subtropi-cal Moist Forest); 2 males (TTU 13370, 13381)and 1 female (TTU 13369) 13.5 km NW Colon,Department of La Libertad (138489N, 898249W;460 m elevation; Subtropical Moist Forest); 2females (TTU 17128, 17129) 3.2 km E Usulu-tan, Department of Usulutan (138209N,888249W; 80 m elevation; Subtropical MoistForest); 1 female (CM 115632) 10 km S Zaca-tecoluca, Hacienda Escuintla, Department ofLa Paz (138249N, 888549W; 5 m elevation; Sub-tropical Moist Forest); 1 male (MUHNES1123) and 1 female (MUHNES 1124) Nancu-chiname National Park, Department of Usu-lutan (138229N, 888439W; 8 m elevation; Sub-tropical Moist Forest); 1 male (TCWC 19733)20 km W Chalatenango, Department of Chal-atenango (148069N, 898069W; 250 m elevation;Subtropical Moist Forest); 2 males (TTU 6407864079) Deininger National Park, Departmentof La Libertad (138299N, 898169W; 20 m ele-
vation; Subtropical Moist Forest); and 1 female(TTU 64080) 4.5 km NW San Luıs, Depart-ment of La Paz (138299N, 898079W; 76 m ele-vation; Subtropical Moist Forest).
The subspecies from El Salvador, E. f. gau-meri, is widely distributed in Central America(Dickerman et al., 1981; Hall, 1981; McCarthy,1987) and is considered to be a lowland form,mainly occurring at elevations ,1,000 m (Da-vis, 1966). These data suggest that E. furinalisis distributed throughout El Salvador from thePacific lowlands up to elevations of about 1,000m.
I thank D. Aguilar, R. J. Baker, D. Schlitter, and J.R. Wible for permission to examine specimens un-der their care. D. Wilson and an anonymous review-er made comments that improved this paper. Spec-imens, herein cited as new records for El Salvador,were originally collected in the field by R. J. Baker,W. J. Bleier, R. K. LaVal, and J. G. Owen. Collectingpermits for Owen were provided by A. Sanchez, Na-tional Parks and Wildlife, Salvadoran Ministry of Ag-riculture.
LITERATURE CITED
BAKER, R. J., S. R. HOOFER, C. A. PORTER, AND R. A.VAN DEN BUSSCHE. 2003. Diversification amongNew World leaf-nosed bats: an evolutionary hy-pothesis and classification inferred from digen-omic congruence of DNA sequence. OccasionalPapers, Museum of Texas Tech University 230:1–32.
BAKER, R. J., S. SOLARI, AND F. G. HOFFMAN. 2002. Anew Central American species from the Carolliabrevicauda complex. Occasional Papers, Museumof Texas Tech University 217:1–11.
BURT, W. H., AND R. A. STIRTON. 1961. The mammalsof El Salvador. Miscellaneous Publications, Mu-seum of Zoology, University of Michigan 117:1–69.
DAVIS, W. B. 1966. Review of South American bats ofthe genus Eptesicus. Southwestern Naturalist 11:245–274.
DAVIS, W. B. 1968. Review of the genus Uroderma(Chiroptera). Journal of Mammalogy 49:676–698.
DICKERMAN, R. W., K. F. KOOPMAN, AND C. SEYMOUR.1981. Notes on bats from the Pacific lowlands ofGuatemala. Journal of Mammalogy 62:406–411.
DINERSTEIN, E. 1985. First records of Lasiurus casta-neus and Antrozous dubiaquercus from Costa Rica.Journal of Mammalogy 66:411–412.
DOLAND, P. G., AND D. C. CARTER. 1979. Distributionnotes and records for Middle American Chirop-tera. Journal of Mammalogy 60:644–649.

March 2005 99Notes
ENGSTROM, M. D., B. K. LIM, AND F. A. REID. 1994.Two small mammals new to the fauna of El Sal-vador. Southwestern Naturalist 39:281–306.
FELTEN, H. 1955. Fledermause (Mammalia, Chirop-tera) aus El Salvador. Teil 1. SenckenbergianaBiologica 36:271–85.
FELTEN, H. 1956a. Fledermause (Mammalia, Chirop-tera) aus El Salvador. Teil 2. SenckenbergianaBiologica 37:69–86.
FELTEN, H. 1956b. Fledermause (Mammalia, Chirop-tera) aus El Salvador. Teil 3. SenckenbergianaBiologica 37:179–212.
FELTEN, H. 1956c. Fledermause (Mammalia, Chirop-tera) aus El Salvador. Teil 4. SenckenbergianaBiologica 37:341–367.
FELTEN, H. 1957. Fledermause (Mammalia, Chirop-tera) aus El Salvador. Teil 5. SenckenbergianaBiologica 38:1–22.
GENOWAYS, H. H., AND J. K. JONES, JR. 1968. Notes onbats from the Mexican State of Zacatecas. Journalof Mammalogy 49:743–745.
HALL, E. R. 1981. The mammals of North America,second edition. John Wiley and Sons, New York.
HALL, E. R., AND J. K. JONES, JR. 1961. North Ameri-can yellow bats, ‘‘Dasypterus,’’ and a list of thenamed kinds of the genus Lasiurus Gray. Univer-sity of Kansas Publications, Museum of NaturalHistory 14:73–94.
HANDLEY, C. O., JR. 1976. Mammals of the Smithson-ian Venezuelan Project. Brigham Young Univer-sity Science Bulletin, Biological Series 20:1–91.
HELLEBUYCK, V., J. R. TAMSITT, AND J. G. HARTMAN.1985. Records of bats new to El Salvador. Journalof Mammalogy 66:783–788.
HOLDRIDGE, L. R. 1975. Mapa ecologica de El Salva-dor: memoria explicativa. Ministerio de Agricul-tura y Ganaderıa, San Salvador, El Salvador.
JONES, J. K., JR., AND W. J. BLEIER. 1974. Sanborn’slong-tongued bat, Leptonycteris sanborni, in El Sal-vador. Mammalia 38:144–145.
LIM, B. K., M. D. ENGSTROM, T. E. LEE, JR., J. C. PAT-TON, AND J. W. BICKHAM. 2004. Molecular differ-entiation of large species of fruit-eating bats (Ar-tibeus) and phylogenetic relationships based onthe cytochrome b gene. Acta Chiropterologica 6:1–12.
MCCARTHY, T. J. 1987. Distributional records of batsfrom the Caribbean lowlands of Belize and adja-cent Guatemala and Mexico. Fieldiana Zoology,New Series 39:137–162.
OWEN, J. G., J. ARROYO-CABRALES, AND J. K. JONES, JR.1993. First record of Noctilio albiventris (Chirop-tera, Noctilionidae) in El Salvador. Texas Journalof Science 45:273–274.
OWEN, J. G., J. K. JONES, JR., AND R. J. BAKER. 1991.Annotated checklist of land mammals of El Sal-vador. Occasional Papers, Museum of Texas TechUniversity 139:1–17.
OWEN, J. G., D. J. SCHMIDLY, AND W. B. DAVIS. 1984.A morphometric analysis of three species of Car-ollia (Chiroptera, Glossophaginae) from MiddleAmerica. Mammalia 48:85–93.
PINE, R. H. 1972. The bats of the genus Carollia.Technical Monograph, Texas Agricultural Exper-iment Station, Texas A&M University, 8:1–125.
REID, F. A. 1997. A field guide to the mammals ofCentral America & southeast Mexico. OxfordUniversity Press, New York.
WILSON, D. E., AND F. R. COLE. 2000. Common namesof mammals of the world. Smithsonian Institu-tion Press, Washington.
WRIGHT, A. J., R. A. VAN DEN BUSSCHE, B. K. LIM, M.D. ENGSTROM, AND R. J. BAKER. 1999. Systematicsof the genera Carollia and Rhinophylla based onthe cytochrome-b gene. Journal of Mammalogy80:1202–1213.
Submitted 25 February 2003. Accepted 5 April 2004.Associate Editor was Cheri A. Jones.

100 vol. 50, no. 1The Southwestern Naturalist
RANGE EXPANSION OF RED FOXES IN NORTHWESTERN TEXAS ANDNORTHEASTERN NEW MEXICO
JAN F. KAMLER, WARREN B. BALLARD,* ROBERT L. HARRISON, AND C. GREGORY SCHMITT
Department of Range, Wildlife, and Fisheries Management, Box 42125, Texas Tech University,Lubbock, TX 79409 (JFK, WBB)
Department of Biology, University of New Mexico, Albuquerque, NM 87131 (RLH)New Mexico Department of Game and Fish, Box 25112, Santa Fe, NM 87504 (CGS)
Present address of JFK: Wildlife Conservation Research Unit, Department of Zoology, South Parks Road,Oxford, 0X1 3PS, United Kingdom
*Correspondent: [email protected]
ABSTRACT We recently collected specimens of red foxes (Vulpes vulpes) in northwestern Texasand northeastern New Mexico that extend the known range of this species in both states. In Texas,the new record likely represents a western range expansion of introduced red foxes that hadspread across the state during the past 100 years. In New Mexico, the new record likely representsa westward expansion from Texas, or possibly an eastward or northward expansion from previouslydocumented populations in the state.
RESUMEN Recientemente colectamos especımenes de zorra roja (Vulpes vulpes) en el noroestede Texas y noreste de Nuevo Mexico, con lo que se extiende la distribucion conocida de estaespecie en ambos estados. El nuevo registro en Texas probablemente representa la expansionhacia el oeste de zorras rojas introducidas que se han extendido a traves del estado en los ultimos100 anos. El nuevo registro en Nuevo Mexico probablemente representa la expansion hacia eloeste desde Texas, o posiblemente la expansion hacia el este o norte de poblaciones previamentedocumentadas en el estado.
Red foxes (Vulpes vulpes) are common faunain most areas of North America. However, redfoxes are absent from many semi-arid regions,including lowland areas of the southwesternUnited States (Hall and Kelson, 1959; Samueland Nelson, 1982; Voigt, 1987). In Texas andNew Mexico, red foxes reportedly occur inmost areas of both states, except for large por-tions of the shortgrass prairie region in ex-treme western Texas and eastern New Mexico(Findley et al., 1975; Davis and Schmidly, 1994;Choate, 1997).
In Texas, red foxes are considered nonna-tive, as they did not historically occur in thestate (Davis and Schmidly, 1994). Starting in1895, red foxes (probably from states to theeast) were introduced for sport in the centraland eastern portions of Texas, where they es-tablished populations by the 1940s (Strecker,1926; Taylor and Davis, 1947; Davis andSchmidly, 1994). By the 1960s, red foxesspread north and west in Texas, and were re-ported as far north as Swisher County, and asfar west as Hale, Lubbock, and Upton counties
(Packard and Bowers, 1970; Davis, 1974). Innorthwestern Texas, the western limit of redfoxes was reported as the central Panhandle(Davis and Schmidly, 1994; Choate, 1997).
In September 2000, we collected remains ofa road-killed red fox in Stratford, in westernSherman County, Texas (TTU #85507, TheMuseum, Texas Tech University, Lubbock).This record extends the known range of redfoxes approximately 75 km west than previous-ly reported in the northern Texas Panhandle(Davis and Schmidly, 1994). We also observedred foxes in some towns of adjacent countiesto the west and south of Sherman County, sug-gesting red foxes likely occur throughout theentire western part of the Texas Panhandle.
In New Mexico, all records and specimensof red foxes (n . 30) before 1950 were frommontane habitats and valleys throughout cen-tral and western New Mexico (Townsend,1893; Bailey, 1932; Halloran, 1946; Clothier,1957; Hall and Kelson, 1959; Findley et al.,1975), indicating native red foxes historicallyoccupied the western two-thirds of the state.

March 2005 101Notes
For example, red foxes were reported from theSan Juan and Sangre de Cristo mountainsacross northern New Mexico (Bailey, 1932; Fin-dley et al., 1975), southwards as far as the SanAndres (Halloran, 1946; Hall and Kelson,1959; Findley et al., 1975) and Mogollon (Bai-ley, 1932) mountains in southern and westernNew Mexico, respectively. Historically, red fox-es were rare or absent from the plains of east-ern New Mexico. A red fox captured by a trap-per in Chaves County in 1958 was the first re-cord from eastern New Mexico (Findley et al.,1975). The trapper had never seen a red foxin the region and was not sure of its identity(Findley et al., 1975). No additional recordswere reported in eastern New Mexico until 2red fox specimens were collected in the late1980s near Portales, Roosevelt County (#9707and #10533; Natural History Museum, EasternNew Mexico University, Portales).
In May 1998, we collected a red fox speci-men near Clayton, Union County, in north-eastern New Mexico (Museum of Southwest-ern Biology, University of New Mexico, Albu-querque; specimen number not yet assigned).This record is approximately 200 km east ofprevious records from the mountains of north-ern New Mexico (Hall and Kelson, 1959; Fin-dley et al., 1975; Samuel and Nelson, 1982).This specimen also is 250 km north of red fox-es collected from the plains of east-central NewMexico (Roosevelt County). The specimen wecollected might represent a westward expan-sion from Texas, an eastward expansion fromexisting populations in the mountains ofnorthern New Mexico, or a northward expan-sion from red foxes in east-central New Mexico(which had an unknown source). Red foxeshave been harvested by trappers in all countiesof eastern New Mexico during the past 20 years(unpublished harvest data, New Mexico De-partment of Game and Fish, Albuquerque),suggesting red foxes now occur throughouteastern New Mexico.
We thank R. Gilliland, D. Anderson, C. Cosper,and F. Pronger for providing information concern-ing red foxes. This is Texas Tech University, Collegeof Agricultural Sciences and Natural Resources pub-lication T-9-847.
LITERATURE CITED
BAILEY, V. 1932. Mammals of New Mexico. NorthAmerican Fauna 53:1–412.
CHOATE, L. L. 1997. The mammals of the Llano Es-tacado. Special Publications of The Museum,Texas Tech University, Number 40.
CLOTHIER, R. 1957. Distribution of the mammals ofthe Sandia and Manzano mountains, New Mexi-co. Unpublished Ph.D. dissertation, University ofNew Mexico, Albuquerque.
DAVIS, W. B. 1974. The mammals of Texas. TexasParks and Wildlife Press, Austin.
DAVIS, W. B., AND D. J. SCHMIDLY. 1994. The mam-mals of Texas. Texas Parks and Wildlife Press,Austin.
FINDLEY, J. S., A. H. HARRIS, D. E. WILSON, AND C.JONES. 1975. Mammals of New Mexico. Universityof New Mexico Press, Albuquerque.
HALL, E. R., AND K. R. KELSON. 1959. The mammalsof North America. Ronald Press Company, NewYork.
HALLORAN, A. 1946. The carnivores of the San An-dres Mountains, New Mexico. Journal of Mam-malogy 27:154–161.
PACKARD, R. L., AND J. H. BOWERS. 1970. Distribution-al notes on some foxes from western Texas andeastern New Mexico. Southwestern Naturalist 14:450–451.
SAMUEL, D. E., AND B. B. NELSON. 1982. Foxes. In: J.A. Chapman and G. A. Feldhamer, editors. Wildmammals of North America: biology, manage-ment, and economics. Johns Hopkins UniversityPress, Baltimore, Maryland. Pp. 475–490.
STRECKER, J. K. 1926. A check-list of the mammals ofTexas. Baylor Bulletin (Baylor University) 29:1–48.
TAYLOR, W. P., AND W. B. DAVIS. 1947. The mammalsof Texas. Bulletin of Texas Game, Fish and OysterCommission 27:1–79.
TOWNSEND, C. H. T. 1893. On the life zones of theOrgan Mountains and adjacent region in south-ern New Mexico, with notes on the fauna of therange. Science 22:313–315.
VOIGT, D. R. 1987. Red fox. In: M. Novak, J. A. Baker,M. E. Obbard, and B. Mallock, editors. Wild fur-bearer management and conservation in NorthAmerica. Ontario Ministry of Natural Resources,Toronto, Canada. Pp. 379–392.
Submitted 15 May 2003. Accepted 2 April 2004.Associate Editor was Cody W. Edwards.

102 vol. 50, no. 1The Southwestern Naturalist
OCCURRENCE OF JAGUAR (PANTHERA ONCA) IN SINALOA, MEXICO
CARLOS J. NAVARRO-SERMENT, CARLOS A. LOPEZ-GONZALEZ, AND
JUAN-PABLO GALLO-REYNOSO*
Centro de Investigacion en Alimentacion y Desarrollo, A.C. Unidad Guaymas, Carretera a Varadero Nacional,Km. 6.6. Col. Las Playitas, Guaymas, Sonora 85480, Mexico (CJNS, JPGR)
Escuela de Biologıa, Universidad Autonoma de Queretaro, Cerro de las Campanas s/n,Queretaro, 76010, Mexico (CALG)
*Correspondent: [email protected]
ABSTRACT Little is known about the distribution of jaguar (Panthera onca) in Sinaloa. We pro-vide current information about the distribution of this species in this state. Different areas of thestate were visited, and people were interviewed from May 2000 to May 2002. We obtained 57records, of which 41 are new; records were most abundant in the southern half of the state. Theextensive areas still covered by tropical deciduous forest in good condition are important for theconservation of the jaguar. The density of prey species for jaguar seems to be high according tolocal residents. Free-ranging cattle also are distributed in all areas of the state, and predation oncattle is common. Ranchers regularly kill jaguars using poisons (strychnine), dogs, steel traps, andshooting. The abundance of records suggests that the jaguar still exists in Sinaloa, especially inthe mountainous areas, but there is an urgent need to conduct additional studies to determineits actual status.
RESUMEN Se sabe poco de la distribucion del jaguar (Panthera onca) en Sinaloa. Proveemosinformacion actual de la distribucion de esta especie en este estado. Se visitaron diferentes areasdel estado y se entrevisto a los lugarenos de mayo del 2000 al mayo del 2002. Se presentan 57registros de los cuales 41 son nuevos registros; los registros fueron mas abundantes en la mitadsur del estado. Las extensas areas todavıa cubiertas por bosque tropical caducifolio en buenacondicion son importantes para la conservacion del jaguar. La densidad de las especies que con-forman las presas del jaguar son aparentemente altas de acuerdo con las personas entrevistadas.El ganado vacuno tambien se encuentra distribuido por todo el estado, y la depredacion delganado parece ser comun. Los rancheros regularmente matan a los jaguares usando venenos(estricnina), perros, trampas de acero y disparandoles. La abundancia de registros sugiere que eljaguar aun existe en Sinaloa, especialmente en la zona serrana, pero hay una necesidad urgentede conducir mas estudios para determinar su estado actual.
The largest felid in the New World, the jag-uar (Panthera onca) historically ranged fromthe southwestern United States to northern Ar-gentina; however, habitat destruction andhunting have reduced its former range (Sey-mour, 1989) to less than 37% of its extent inca. 1900 (Sanderson et al., 2002). In north-western Mexico, it still occurs in the state ofSonora, where at least 3 sub-populations havebeen identified in recent years (Lopez-Gonza-lez and Brown, 2002). Just south of Sonora and500 km south of the USA-Mexico border, thestate of Sinaloa constitutes the link betweenpopulations in Sonora and more southerlypopulations near the Mexican Pacific coast. Le-opold (1959:529) reported that ‘‘the highestdensities of jaguars noted in the course of this
survey [of Mexican wildlife] were along theheavily forested flatlands and foothills of south-ern Sinaloa, the swamps of coastal Nayarit, theremaining uncut forests along the Gulf coastas far east as central Campeche, and the greatrain forests of northern Chiapas.’’ It is a wide-spread belief that the range of the jaguar inMexico has been reduced so much that it isonly possible to find important populations inthe southeastern states (e.g., Tellez-Giron andLopez-Forment, 1995; Aranda, 1996). Howev-er, that belief possibly reflects the lack of re-search and available information rather thanthe lack of jaguars, as was discussed during thelast attempt to gather information on currentdistribution in 1999 during the ‘‘Jaguar in theNew Millennium’’ workshop (Sanderson et al.,

March 2005 103Notes
FIG. 1 Approximate locations of the recent re-cords of jaguar (Panthera onca) obtained by us (blackdots) and cities where interviews were made (opencircles).
2002). It was shown at that workshop that in alarge area of western Mexico, of which Sinaloarepresented a considerable portion, the statusof the jaguar remained largely unknown (San-derson et al., 2002). The tropical deciduousforests that cover much of the sierras of Sina-loa are part of the 18% of the historic rangeof the jaguar on which its status remains un-known, and where the development of surveysis considered a priority (Sanderson et al.,2002). To better understand the current situ-ation of the jaguar in Sinaloa, we undertook aseries of field trips, reviewed literature ac-counts of the species, and summarized pub-lished information (Appendix 1).
We conducted 60 interviews opportunistical-ly from the summer of 2000 to May 2002. In-terviewees included local ranchers, local live-stock association officials, hunting club mem-bers, and employees of local tanneries fromthose areas where jaguars were known or sus-pected to exist, especially in mountainouscountry. We tried to obtain physical records(photographs, skins, or bones) of hunted jag-uars, as well as basic information regarding killsites (such as location, date, and type of vege-tation). We visited 8 towns and cities (El Fuer-te, Badiraguato, San Ignacio, Concordia, Chu-paderos, Copala, Mazatlan, and Teacapan; Fig.1) and summarized previously published rec-ords from Sinaloa.
We obtained 57 records of individual jag-uars, of which 41 are first reported here (Ap-pendix 1). Jaguar records were abundant inthe southern half of the state. The local peopleconsidered the species a common and regularmember of the local fauna throughout themountainous sierra. The jaguar seems to bemost common in the San Ignacio and Concor-dia municipalities; however, in those 2 munic-ipalities the search effort was more extensive.More research is needed in northern Sinaloa.
During the course of this survey, only 1 (pos-sibly 2) records were found from higher ele-vations in oak-pine (Pinus-Quercus) forest and1 record from riparian vegetation. Most occur-rences were from the tropical deciduous forestthat originally occupied most of the lowlandsin the state and still covers much of the sierra.Tropical deciduous forest currently covers40.09% of the area in the state, whereas oakand pine forests cover 14.71% of Sinaloa (INE-GI, 2000). Today, most of the coastal plain(34.72% of Sinaloa) is being transformed foragriculture, aquaculture, or human settlement(INEGI, 2000), and few adequate habitatpatches remain for jaguars there. However, 2recent records occurred close to the coast, ap-proximately 80 km north of the city of Maza-tlan; one was within the Mesa de Cacaxtla Nat-ural Protected Area. Although much impactedby aquaculture operations or human settle-ments, mangrove swamps still represent 7.36%of the area in the state (INEGI, 2000).
Predation on livestock by jaguars seems to bea common event. Cattle traditionally are al-lowed to roam over vast expanses of land. Lo-cal ranchers kill jaguars regularly using differ-ent methods, including poison (strychnine),dogs, and steel traps, or by shooting wheneverthey have a chance encounter. Steel traps forlarge cats can be purchased readily (prices cur-rently range from 400 to 600 pesos, aboutUS$40 to US$60) at many local hardwarestores, despite the fact that jaguars and moun-tain lions (Felis concolor) have been legally pro-tected in Mexico since 1986 and are includedin the ‘‘Norma Oficial Mexicana’’ (NOM-059-ECOL-1994) (SEMARNAP, 1994), the federallist of protected animals and plants. In someareas, landowners pay bounties of up to 5,000pesos (about US$500) for killing a trouble-some jaguar in cattle areas or, in some places,for any jaguar. To determine whether a jaguar

104 vol. 50, no. 1The Southwestern Naturalist
FIG. 2 Jaguar (Panthera onca) hunted near Co-pala, in the municipality of Concordia, in the early1990s. This specimen might be 1 of the 2 ‘‘tigre pin-ta menuda’’ reported north of South America.
or a mountain lion killed cattle, ranchers ob-serve where the predator started to eat thecow. Mountain lions kill cattle by biting thethroat, producing asphyxia; they begin to eatthe hindquarters and cover the carcass withbranches and dirt. Jaguars kill by biting thecervical vertebrae or the skull. Jaguars dragtheir prey by the muzzle, which causes thetongue to protrude. Jaguars begin eating thetongue, the muscles of the chest, or the mus-cles of the face, and do not cover the carcass.Both felines remove the stomach and intes-tines. In 2 interviews, ranchers saw a jaguar eat-ing a dead cow. These methods to identifywhich feline killed cattle also are used in So-nora (Brown and Lopez-Gonzalez, 2001).
Whenever a ‘‘tigre’’ (jaguar) is killed, it isskinned and the hide normally is sold, usuallyto people from larger cities, such as Mazatlan,Concordia, Culiacan, and Escuinapa, or totourists. Ranchers usually sell the skins for1,500 to 3,000 pesos (about US$150 to 300).No evidence of current sport hunting of jag-uars was found during this survey, but somemight still occur.
Extensive areas of tropical deciduous forestremain along the Sinaloan sierras. Densities ofjaguar prey, such as armadillo (Dasypus novem-cinctus), coatimundi (Nasua narica), collaredpeccary (Tayassu tajacu), and white-tailed deer(Odocoileus virginianus), seemed to be high, ac-cording to most people, and free-ranging do-mestic animals were widespread. A new, unde-scribed population of European wild boar orferal hog (Sus scrofa) might be established inthe Concordia municipality, as suggested by aset of tusks from the area. European wild boarswere introduced into the buffer zone of LaMichilıa Biosphere Reserve, Durango, in 1989(Weber, 1995), about 160 km from Concordia.Hogs might provide jaguars another prey item,because jaguars frequently prey on several pigspecies, both wild and domestic (Leopold,1959; Seymour, 1989; Brown and Lopez-Gon-zalez, 2000). The widespread practice of drugharvest and traffic during recent years mighthave benefited jaguars, because fewer sporthunters and foreigners visit the area.
A specimen (Fig. 2) hunted near Copala, inthe municipality of Concordia, in the early1990s, has unusually small, broken rosetteswithout interior spots. This variation is knownas ‘‘tigre pinta menuda,’’ or small-spotted jag-
uar (Brown and Lopez-Gonzalez, 2001). Thisspecimen and the 1 described by Alessio-Ro-bles (2002) from Campeche are the only 2such specimens reported north of South Amer-ica.
The abundance of recent records suggeststhat a jaguar population still exists in Sinaloa,especially throughout the sierra, and more sur-veys urgently are needed to better understandits current status.
We acknowledge the help of S. Vizcarra, S. Vizcar-ra, Jr., A. van der Heiden, H. Plascencia, A. van derHeiden, D. Castro, J. Calderon, E. Castro, A. Ruız-Luna, and C. Puente, all of whom provided valuableinformation and assistance in the field. V. Gehr-mann created the map. C. J. Navarro took the pho-tograph of the ‘‘tigre pinta menuda’’ (Fig. 2). Partof this study was supported by grants from theNorthern Jaguar Project and the Minority Interna-tional Research Training University of CaliforniaSanta Cruz Centro de Investigacion en Alimenta-cion y Desarrollo, Unidad Guaymas. The manuscriptimproved thanks to the help of D. Valenzuela.
LITERATURE CITED
ALESSIO-ROBLES, M. 2002. Un jaguar en el Silencio.Estafeta. Impresos Naucalpan. San Andres AtotoNumero 12. Colonia San Andres Atoto. MexicoD. F., 53500, Mexico.
ARANDA, M. 1996. Distribucion y abundancia del jag-uar, Panthera onca (Carnivora: Felidae) en el es-

March 2005 105Notes
tado de Chiapas, Mexico. Acta Zoologica Mexi-cana 68:45–52.
ARMSTRONG, D. M., J. K. JONES, JR., AND E. C. BIRNEY.1972. Mammals from the Mexican state of Sina-loa. III. Carnivora and Artiodactyla. Journal ofMammalogy 53:48–61.
BROWN, D. E., AND C. A. LOPEZ-GONZALEZ. 2000.Notes on the occurrences of jaguars in Arizonaand New Mexico. Southwestern Naturalist 45:537–546.
BROWN, D. E., AND C. A. LOPEZ-GONZALEZ. 2001. Bor-derland jaguars. University of Utah Press, SaltLake City.
INSTITUTO NACIONAL DE ESTADıSTICA GEOGRAFıA E IN-FORMATICA (INEGI). 2000. Anuario estadıstico delestado de Sinaloa, Mexico. www.inegi.gob.mx(Accessed: July 2000).
LEOPOLD, A. S. 1959. Wildlife of Mexico. Universityof California Press, Berkeley.
LOPEZ-GONZALEZ, C. A., AND D. E. BROWN. 2002. Dis-tribucion y estado de conservacion actuales deljaguar en el noroeste de Mexico. In: R. A. Me-dellın, C. Equihua, C. L. B. Chetkiewicz, P. G.Crawshaw, A. Rabinowitz, K. H. Redford, J. G.Robinson, E. Sanderson, and A. Taber, compilers.El jaguar en el nuevo milenio. Fondo de CulturaEconomica Universidad Nacional Autonoma deMexico Wildlife Conservation Society. Mexico,D. F. Pp. 379–391.
LOPEZ-GONZALEZ, C. A., D. E. BROWN, AND G. LOR-ENZANA. 2000. El jaguar en Sonora, ¿desapare-ciendo o solamente desconocido? Especies. Re-vista sobre Conservacion y Biodiversidad. 9(3):19–23.
MCCURDY, R. 1981. Life of the greatest guide; houndstories and others of Dale Lee. Blue River Graph-ics, Phoenix, Arizona.
SANDERSON, E. W., C. L. B. CHETKIEWICZ, R. A. MED-ELLıN, A. RABINOWITZ, K. H. REDFORD, J. G. ROB-INSON, AND A. B. TABER. 2002. Prioridades geo-graficas para la conservacion del jaguar. In: R. A.Medellın, C. Equihua, C. L. B. Chetkiewicz, P. G.Crawshaw, A. Rabinowitz, K. H. Redford, J. G.Robinson, E. Sanderson, and A. Taber, compilers.El jaguar en el nuevo milenio. Fondo de CulturaEconomica Universidad Nacional Autonoma deMexico Wildlife Conservation Society. Mexico,D. F. Pp. 601–628.
SECRETARıA DEL MEDIO AMBIENTE RECURSOS NATURA-LES Y PESCA (SEMARNAP). 1994. Norma OficialMexicana NOM-059-ECOL-1994, que determinalas especies y subespecies de Flora y Fauna silves-tres terrestres y acuaticas en peligro de extincion,amenazadas, raras y las sujetas a proteccion es-pecial y que establece especificaciones para suproteccion. Diario Oficial de la Federacion.www.semarnat.gob.mx (Accessed: July 2000).
SEYMOUR, K. L. 1989. Panthera onca. Mammalian Spe-cies 340:1–9.
TELLEZ-GIRON, G., AND W. LOPEZ-FORMENT. 1995.Panthera onca veracrucis (Carnivora: Felidae) enQueretaro, Mexico. Revista Mexicana de Masto-zoologıa 1:73–75.
WEBER, M. 1995. La introduccion del jabalı europeoen la Reserva de la Biosfera La Michilıa, Duran-go: implicaciones ecologicas y epidemiologicas.Revista Mexicana de Mastozoologıa 1:69–73.
Submitted 7 March 2003. Accepted 5 July 2004.Associate Editor was Cheri A. Jones.
APPENDIX 1 Records of jaguar from Sinaloa, Mex-ico, in reverse chronological order (1857 through2002). We have included the date of the kill, loca-tion (geographic coordinates are approximate), bi-otic community, sex (if known), documentation, andcitation, if the record was previously published.
JULY 2002, Concordia municipality, tropical decid-uous forest, female and cub (sex unknown), huntedby local ranchers (A. van der Heiden, pers. comm.,2002). MAY 2002, between Rancho Coyote and Ma-gistral, Concordia municipality (238219490N,1068599450W), tropical deciduous forest, sex un-known, dead livestock seen by local ranchers andpositively identified as jaguar kill. APRIL 2002, ‘‘LosLlanitos,’’ ca. 2 km from El Marmol, Mazatlan mu-nicipality (238329190N, 1068359580W), tropical decid-uous forest, sex unknown, hunted by local people(A. Ruız-Luna, pers. comm., May 2002). APRIL 2002,ca. San Juan, San Ignacio municipality (238569430N,1068209150W), tropical deciduous forest, sex un-known, dead livestock seen by local ranchers andpositively identified as jaguar kill. MARCH 2002, ‘‘DosArroyos,’’ Badiraguato municipality (258169320N,1078249470W), tropical deciduous forest, sex un-known, hunted by local rancher. MARCH 2002, Ma-gistral, Concordia municipality (238219410N,1068599160W), tropical deciduous forest, young in-dividual stoned to death by local ranchers, sex un-known. MARCH 2002, El Habal, ca. Copala, Concor-dia municipality (238239580N, 1058569070W), tropicaldeciduous forest, sex unknown, hunted by localrancher.
1998–2002, Concordia municipality, tropical de-ciduous forest, skins of 4 individuals seen by us atMazatlan, sexes unknown.
SUMMER 2001, San Isidro Ranch, Concordia munic-ipality (238219510N, 1058599590W), tropical deciduousforest, male (?), footprints seen by local rancher. DE-CEMBER 2001, ‘‘La Laguna,’’ ca. Durango Ranch, Con-cordia municipality (238209570N, 1058549140W), trop-ical deciduous forest, female (?), poisoned by localranchers. NOVEMBER 2001, Rancho Coyote, Concordiamunicipality (238219540N, 1058599580W), tropical de-ciduous forest, sex unknown, heard by local ranchers.

106 vol. 50, no. 1The Southwestern Naturalist
Killed a goat at the ranch that night. FALL 2001,‘‘southern Sinaloa,’’ male and female, captive cubs inprivate home (A. van der Heiden, pers. comm.,2002). 2001, Coacoyol, San Ignacio municipality(238579420N, 1068299340W), tropical deciduous forest,sex unknown, shot by local rancher as livestock-killer.2001, Mesa de Cacaxtla, San Ignacio municipality(238399380N, 1068429590W), tropical deciduous forest,sex unknown, hunted by local ranchers. 2001, Sahui-tapa, ca. San Ignacio, San Ignacio municipality(248009000N, 1068299470W), tropical deciduous forest,2 individuals shot by local ranchers, sexes unknown.
SUMMER 2000, ‘‘Cerro El Pirame,’’ Concordia mu-nicipality (238219500N, 1058599560W), tropical decid-uous forest, male shot by local rancher as livestock-killer. Skull collected and deposited at Centro de In-vestigacion en Alimentacion y Desarrollo, Guaymascollection (CIAD-070502-1). SUMMER 2000, El Tule,San Ignacio municipality (238499360N, 1068259320W),tropical deciduous forest, 2 individuals separately shotby local ranchers while feeding on dead cows, 1 maleand an individual of unknown sex. LATE 2000, ca.Coyotitan, San Ignacio municipality (238479530N,1068349580W), tropical deciduous forest, sex un-known, killed by car while crossing the road at night,skin in possession of the former mayor of San Ignacio.2000, Coacoyol, San Ignacio municipality(238579420N, 1068299340W), tropical deciduous forest,sex unknown, shot by local rancher.
1999, ‘‘Cerro El Elefante,’’ Concordia municipal-ity (238199000N, 1058599580W), tropical deciduousforest, sexes unknown, 2 cubs captured alive by localrancher. 1992, ca. Mesillas, Concordia municipality(238179440N, 1068049130W), tropical deciduous for-est, male, attracted by commercial predator-call tapeand shot as livestock killer, skin seen by us in Con-cordia. EARLY 1990s, ca. Copala, Concordia munici-pality (23823 580N, 1058569000W), tropical deciduousforest, sex unknown, skin seen by us in Copala. 1990,Baroten, ca. 3.5 Km S of El Fuerte, El Fuerte munic-ipality (268239450N, 1088359550W), riparian vegeta-tion along El Fuerte river, sex unknown, hunted bylocal people and seen by local veterinarian.
LATE 1980s, EARLY 1990s. El Tule, ca. Escuinapa, Es-cuinapa municipality, tropical deciduous forest, sexesunknown, 3 individuals seen on different occasionsquietly following people along forest trails at night.
1980s, ca. Los Angeles ranch, Escuinapa municipality(228399560N, 1058489140W), tropical deciduous forest-mangrove swamps, sexes unknown, 3 individualshunted by local people, skins seen by us. 1970s. PlayaBrujas, ca. 23 km NW of Mazatlan, Mazatlan munici-pality, tropical deciduous forest, 1 individual of un-known sex seen walking on the beach. 1962, SierraMadre NE of Matantan near the Durango–Sinaloaboundary, oak-pine forest (?), sex unknown, skull ofindividual shot purchased in Rosario (Armstrong etal., 1972). 1960’s, Sierra de Surutato, Badiraguato mu-nicipality (258489370N, 1078339280W), oak-pine forest,male, dead individual seen by the president of theBadiraguato local livestock association. 1940s, NE ofSan Ignacio, San Ignacio municipality, tropical decid-uous forest, male shot by hunting party (McCurdy,1981).
PRIOR TO 1959, near Santiago, San Ignacio munic-ipality, tropical deciduous forest, male (?), shot bysport hunters (Leopold, 1959). PRIOR TO 1959, vicin-ity of San Ignacio, San Ignacio municipality, tropicaldeciduous forest, sex unknown (Leopold, 1959).1937 or 1938, Mesa de Platanos, San Ignacio munic-ipality, tropical deciduous forest, male and female, 2individuals shot by hunting party (McCurdy, 1981).SUMMER 1935 or 1936, Los Frailes, San Ignacio mu-nicipality, tropical deciduous forest, male shot byhunting party (McCurdy, 1981). 1935 or 1936, SanJuan, 10 miles S of San Ignacio, San Ignacio munic-ipality, tropical deciduous forest, 2 females shot byhunting party (McCurdy, 1981). PRIOR TO 1933, Es-cuinapa, Escuinapa municipality, tropical deciduousforest, sex unknown (Nelson and Goldman, 1933, asreported by Armstrong et al. 1972).
1905–1906, Escuinapa, Escuinapa municipality,tropical deciduous forest, 1 male, 3 females, and 1immature female ( J. H. Batty, as reported by Brownand Lopez-Gonzalez, 2001). 1901, Escuinapa, Escui-napa municipality, tropical deciduous forest, male( J. H. Batty, as reported by Brown and Lopez-Gon-zalez, 2001). Prior to 1901, Cacalotan, Rosario mu-nicipality, tropical deciduous forest, sex unknown(Mearns, 1901, as reported by Armstrong et al.1972).
1857, Mazatlan, tropical deciduous forest, sex un-known, type specimen of Panthera onca hernandesii(Gray, 1858, as reported by Armstrong et al. 1972).













![The Status of Snapping Turtles in the Coastal Plain of ......Dermatemys mawii [7], the pig-nosed turtle, Carettochelys insculpta [8] and the alligator snapping turtle, Macrochelys](https://img.dokumen.tips/doc/110x75/5edac4fefa3b3a5ad21692a4/the-status-of-snapping-turtles-in-the-coastal-plain-of-dermatemys-mawii.jpg)















