Embed Size (px)
Citation preview

Plant Physiol. (1978) 61, 380-385
Source and Sink Leaf Metabolism in Relation to PhloemTranslocationCARBON PARTITIONING AND ENZYMOLOGY'
Received for publication August 25, 1977 and in revised form November 15, 1977
ROBERT GIAQUINTACentral Research and Development Department, Experimental Station, E. I. du Pont de Nemours andCompany, Wilmington, Delaware 19898
ABSTRACT
The import-export transitio in sugar beet leaves (Bets vu*ans) oc-curred at 40 to 50% leaf expansion and was characterized by loss inassimilate import and Increase in pbotosyntbesis. The metabolsm andpartitoning of assimilated and transocated C were determine during leafdevelopment and related to the traslocaton status of the leaf. The importstage was characterized by C derived from either "C-translocate or 1"C-photosynthate being Incorporated into protein and structural carbohy-drates. Marked changes in the C pr g were temporally correlatedwith the import-export conversion. Exportig leaves did not hydrolyzeaccumulated sucrose and the C derived from CO2 fixation was preferen-tialy incorporated into sucrose. Both source and sink leaves containedsimiar levels of acid invertase and sucrose synthetase activities (sucrosehydrolysis) wbile sucrose pbosphate synthetase (sucrose synthesis) wasdetected only in exporting leaves. The results are discussed in terms ofintracellular compartmentatiom of sucrose and sucrose-metabolizing en-zymes in source and sink leaves.
It is well established that as a young leaf matures its statuschanges from an importer to an exporter of photosynthetic assim-ilates (4, 13, 25). In many dicot species, the import-export transi-tion occurs when the leaf is 40 to 50%o expanded and is character-ized by the rapid loss in the ability to import assimilates with theconcomitant onset of export function. Autoradiographic studies(4, 13, 25) have shown that export initiates at the leaf apex andrapidly develops basipetally. The basipetal source development iscorrelated with the cessation of assimilate import. The physiolog-ical and anatomical events accompanying leaf maturation havebeen studied in most detail in Populus deltoides (2, 3, 13), Cucurbitapepo (25), and Beta vulgaris (4).Many studies on the import-export conversion have been to
distinguish the preparatory events from the causal events ofexport.In this regard, Turgeon and Webb (24, 25) characterized theimport-export conversion during leafdevelopment in C. pepo. Thebasipetal acquisition of export was accompanied by an increasedphotosynthesis rate, basipetal development of mesophyll intercel-lular air spaces, synthesis of the transport sugar, stachyose, andstructural maturation of the minor veins. These studies suggestedthat no one system in the development specifically regulated theonset of export but instead the import-export conversion resultedfrom the integrated maturation ofmany structural and physiolog-
'Contribution No. 2514 from the Central Research and DevelopmentDepartment, Experimental Station, E. 1. du Pont de Nemours and Com-pany, Wilmington, Delaware 19898.
ical systems (25). On the other hand, Fellows and Geiger (4)concluded that many of the observed changes noted during leafmaturation were preparatory in nature. The critical or causalevent for the onset of export was postulated as the attainment ofa sufficient osmotic pressure in the sieve tubes by phloem loadingto produce a mass flow of solute out of the source leaf.Many of the previous studies have been at the whole-plant level
and aimed at elucidating the causal events for export. In thisstudy, the leaf import-export conversion is investigated at thecellular and biochemical level not with the intent of determiningthe causal event(s) of export but rather to elucidate the processesof sink and source leafphysiology in relation to translocation. Themetabolic correlates of leaf maturation were studied both indetached leaves and in the intact translocating sugar beet plant,B. vulgaris. Specifically, the metabolism and partitioning of assim-ilated and translocated C were characterized during leaf devel-opment and related to the translocation status of the leaf. Addi-tionally, the levels ofthe transport sugar, sucrose, and the activitiesof various sucrose-metabolizing enzymes were determined insource and sink leaves. These results are discussed in terms ofintracellular compartmentation of sugars in relation to import andexport.
MATERIALS AND METHODS
B. vulgaris L. (monogerm hybrid, size 3) plants were grown for8 to 10 weeks in a controlled environment under conditionsdescribed previously (8). Four experimental protocols were usedin this study: (a) the relationship between leaf import capacityand photosynthesis was determined during leaf development inthe intact plant to generate a time course for the import-exporttransition in our system; (b) using this time course as a reference,the partitioning of C derived from either ['4CJsucrose or "'CO2fixation into various metabolites was studied to determine whetherany changes in C partitioning or metabolism were temporallycorrelated with the sink-source transition; (c) the activities ofinvertase, sucrose synthetase, and sucrose-P synthetase, enzymesrelated to translocate (sucrose) metabolism, were determined inimporting and exporting leaves; (d) the time course of metabolismof 4C02-derived translocate in sink leaves of intact plants wasdetermined to document the metabolic fate of translocate afterunloading from the phloem.
Assimilate Import and Photosynthesis Determinations. To de-termine the import capacity of leaves at various stages of devel-opment, translocation of "'CO2-derived assimilates was measuredin intact sugar beet plants. Five sugar beet plants were selected foruniformity. The night before the experiment the plants weretrimmed to a simplified source-path-sink system (4) each consist-ing of a fully expanded source leaf of approximately 20- to 22-cmlamina length (2-2.5 dm2), a 15-cm source leaf petiole, and one
380 www.plantphysiol.orgon October 8, 2018 - Published by Downloaded from Copyright © 1978 American Society of Plant Biologists. All rights reserved.
SOURCE AND SINK LEAF METABOLISM
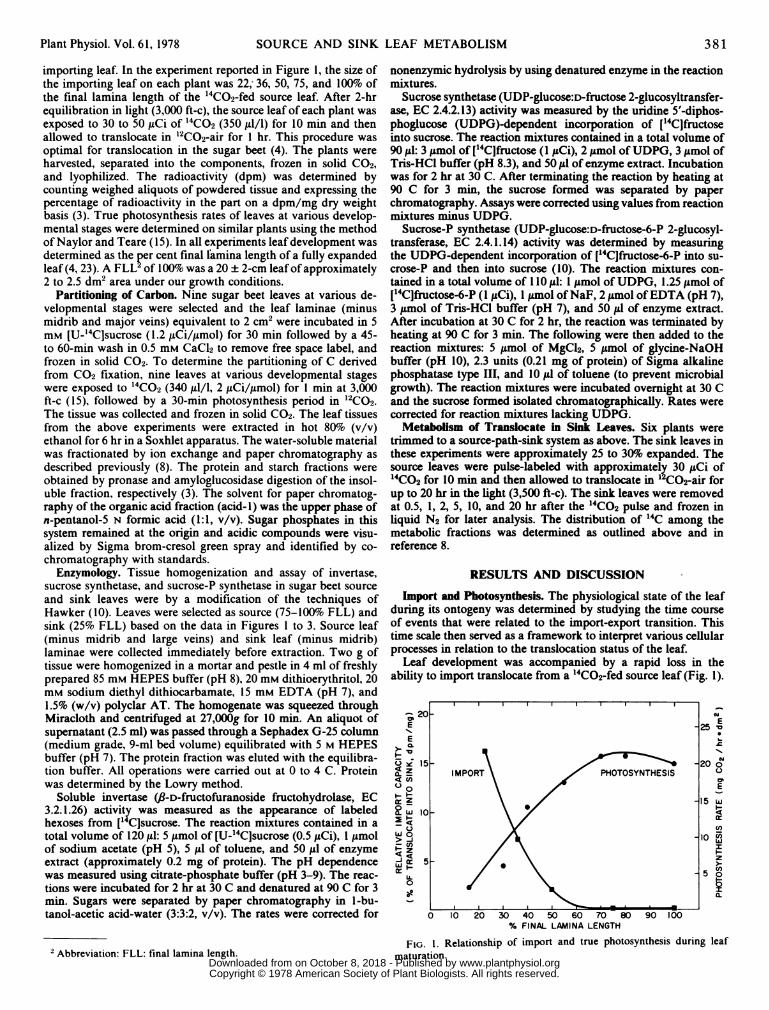
importing leaf. In the experiment reported in Figure 1, the size ofthe importing leaf on each plant was 22, 36, 50, 75, and 10Oo ofthe final lamina length of the '4CO2-fed source leaf. After 2-hrequilibration in light (3,000 ft-c), the source leaf of each plant wasexposed to 30 to 50,ICi of "CO2 (350,ull/1) for 10 min and thenallowed to translocate in '2CO2-air for I hr. This procedure wasoptimal for translocation in the sugar beet (4). The plants wereharvested, separated into the components, frozen in solid C02,and lyophilized. The radioactivity (dpm) was determined bycounting weighed aliquots of powdered tissue and expressing thepercentage of radioactivity in the part on a dpm/mg dry weightbasis (3). True photosynthesis rates of leaves at various develop-mental stages were determined on similar plants using the methodof Naylor and Teare (15). In all experiments leaf development wasdetermined as the per cent final lamina length of a fully expandedleaf (4, 23). A FLL of 100%7o was a 20 ± 2-cm leaf of approximately2 to 2.5 dM2 area under our growth conditions.
Partitioning of Carbon. Nine sugar beet leaves at various de-velopmental stages were selected and the leaf laminae (minusmidrib and major veins) equivalent to 2 cm2 were incubated in 5mM [U-'4C]sucrose (1.2 ,uCi/t,mol) for 30 min followed by a 45-to 60-min wash in 0.5 mM CaCl2 to remove free space label, andfrozen in solid CO2. To determine the partitioning of C derivedfrom CO2 fixation, nine leaves at various developmental stageswere exposed to "CO2 (340 ,ul/l, 2 ,uCi/tmol) for I min at 3,000ft-c (15), followed by a 30-min photosynthesis period in '2CO2.The tissue was collected and frozen in solid CO2. The leaf tissuesfrom the above experiments were extracted in hot 80% (v/v)ethanol for 6 hr in a Soxhlet apparatus. The water-soluble materialwas fractionated by ion exchange and paper chromatography asdescribed previously (8). The protein and starch fractions wereobtained by pronase and amyloglucosidase digestion of the insol-uble fraction, respectively (3). The solvent for paper chromatog-raphy of the organic acid fraction (acid- I) was the upper phase ofn-pentanol-5 N formic acid (1:1, v/v). Sugar phosphates in thissystem remained at the origin and acidic compounds were visu-alized by Sigma brom-cresol green spray and identified by co-chromatography with standards.
Enzymology. Tissue homogenization and assay of invertase,sucrose synthetase, and sucrose-P synthetase in sugar beet sourceand sink leaves were by a modification of the techniques ofHawker (10). Leaves were selected as source (75-100%/o FLL) andsink (25% FLL) based on the data in Figures I to 3. Source leaf(minus midrib and large veins) and sink leaf (minus midrib)laminae were collected immediately before extraction. Two g oftissue were homogenized in a mortar and pestle in 4 ml of freshlyprepared 85 mm HEPES buffer (pH 8), 20 mm dithioerythritol, 20mm sodium diethyl dithiocarbamate, 15 mm EDTA (pH 7), and1.5% (w/v) polyclar AT. The homogenate was squeezed throughMiracloth and centrifuged at 27,000g for 10 min. An aliquot ofsupernatant (2.5 ml) was passed through a Sephadex G-25 column(medium grade, 9-ml bed volume) equilibrated with 5 M HEPESbuffer (pH 7). The protein fraction was eluted with the equilibra-tion buffer. All operations were carried out at 0 to 4 C. Proteinwas determined by the Lowry method.
Soluble invertase (fi-D-fructofuranoside fructohydrolase, EC3.2.1.26) activity was measured as the appearance of labeledhexoses from ["4Cjsucrose. The reaction mixtures contained in atotal volume of 120 Il: 5 ,umol of [U-i4Clsucrose (0.5 ,uCi), I ,umolof sodium acetate (pH 5), 5 ,ul of toluene, and 50 ,il of enzymeextract (approximately 0.2 mg of protein). The pH dependencewas measured using citrate-phosphate buffer (pH 3-9). The reac-tions were incubated for 2 hr at 30 C and denatured at 90 C for 3min. Sugars were separated by paper chromatography in l-bu-tanol-acetic acid-water (3:3:2, v/v). The rates were corrected for
2 Abbreviation: FLL: final lamina length.
nonenzymic hydrolysis by using denatured enzyme in the reactionmixtures.
Sucrose synthetase (UDP-glucose:D-fructose 2-glucosyltransfer-ase, EC 2.4.2.13) activity was measured by the uridine 5'-diphos-phoglucose (UDPG)-dependent incorporation of [i4CJfructoseinto sucrose. The reaction mixtures contained in a total volume of90 il: 3 ,umol of ['4Cjfructose (1 pCi), 2 ,umol of UDPG, 3 ,umol ofTris-HCl buffer (pH 8.3), and 50 ,ul of enzyme extract. Incubationwas for 2 hr at 30 C. After terminating the reaction by heating at90 C for 3 min, the sucrose formed was separated by paperchromatography. Assays were corrected using values from reactionmixtures minus UDPG.
Sucrose-P synthetase (UDP-glucose:D-fructose-6-P 2-glucosyl-transferase, EC 2.4.1.14) activity was determined by measuringthe UDPG-dependent incorporation of 1i4C]fructose-6-P into su-crose-P and then into sucrose (10). The reaction mixtures con-tained in a total volume of 110 .1: 1 ,umol ofUDPG, 1.25 ,umol of['4CJfructose-6-P (1 pCi), 1 pmol ofNaF, 2 pmol ofEDTA (pH 7),3 ,umol of Tris-HCl buffer (pH 7), and 50 ,ul of enzyme extract.After incubation at 30 C for 2 hr, the reaction was terminated byheating at 90 C for 3 min. The following were then added to thereaction mixtures: 5 pmol of MgCl2, 5 ,umol of glycine-NaOHbuffer (pH 10), 2.3 units (0.21 mg of protein) of Sigma alkalinephosphatase type III, and 10 ,u1 of toluene (to prevent microbialgrowth). The reaction mixtures were incubated overnight at 30 Cand the sucrose formed isolated chromatographically. Rates werecorrected for reaction mixtures lacking UDPG.Metabolism of Translocate in Sink Leaves. Six plants were
trimmed to a source-path-sink system as above. The sink leaves inthese experiments were approximately 25 to 30% expanded. Thesource leaves were pulse-labeled with approximately 30 puCi ofi4CO2 for 10 min and then allowed to translocate in '2C02-air forup to 20 hr in the light (3,500 ft-c). The sink leaves were removedat 0.5, 1, 2, 5, 10, and 20 hr after the '4CO2 pulse and frozen inliquid N2 for later analysis. The distribution of 14C among themetabolic fractions was determined as outlined above and inreference 8.
RESULTS AND DISCUSSION
Import and Photosynthesis. The physiological state of the leafduring its ontogeny was determined by studying the time courseof events that were related to the import-export transition. Thistime scale then served as a framework to interpret various cellularprocesses in relation to the translocation status of the leaf.
Leaf development was accompanied by a rapid loss in theability to import translocate from a i4CO2-fed source leaf (Fig. 1).
06
-Cla0 x
W -
aL.
0U 0
cgSR w
2
Jz
4
10 20 30 40 50 60 70 80 90% FINAL LAMINA LENGTH
EX00
N
0
E
I-w
(n0
FIG. 1. Relationship of import and true photosynthesis during leafmaturation.
Plant Physiol. Vol. 61, 1978 381
www.plantphysiol.orgon October 8, 2018 - Published by Downloaded from Copyright © 1978 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 61, 1978
The import phase declined sharply from about 20 to 25% FLL(about 8% final leaf area) and was essentially over by 50% FLL.The time course of loss of ability to import agrees well with thosereported for various species. Fellows and Geiger (4) reported thatmaximum import of translocate in B. vulgaris occurred at 25%FLL and ceased at 45% FLL. Swanson et al. (23) documented a
similar peak in assimilate import at 25% FLL in Phaseolus. Earlierresults for leaves of Glycine max (23) and Cucurbita (24, 25) alsoshow similar trends.Accompanying the loss of ability to import was a marked
increase in true photosynthesis, beginning at or before 15% FLLand reaching a maximum of 20 mg C02/hr idm2 between 65 and100% FLL (Fig. 1). This photosynthetic development is similar tothat reported for other species (2, 24). Thus, decreasing depend-ence on imported assimilates for growth (15-40% FLL) is associ-ated with the increasing ability of the leaf to produce its ownphotoassimilates for maintenance and export. It is important tonote that in sugar beet (4) the loss of import is accompanied bythe onset of export. In sugar beet, export begins at 35 to 40% FLLand reaches a maximum from 50% to full leafexpansion (4). Thus,export temporally coincides with increased photoassimilate pro-duction. It can be inferred from Figure 1 that the plants used hereundergo the import-export transition at 40 to 50% FLL. Using thisas a time course it was possible to characterize various cellparameters in relation to the conversion.
Partitioning of Carbon. Two approaches were used to study Cpartitioning during leafdevelopment. First, [14Cjsucrose, the majortranslocated sugar in sugar beet, was supplied to leaves at variousmaturation stages and its metabolism was followed. Second, thepartitioning of C derived from photosynthetic CO2 fixation wasdetermined to investigate the metabolic fate of in situ photoas-similates during leafdevelopment. Figure 2 shows the distributionof 14C into various metabolites in developing leaves after accu-mulation of externally supplied I14C]sucrose. The import phasewas characterized by a substantial amount of C being partitionedinto the protein (25-30%) and residue (containing starch andstructural carbohydrates) (35%) fractions at the expense of thewater-soluble components (40%). This partitioning of C is alsoreflected in the amino acid and organic acid precursors of thesefractions (Fig. 2B). During leaf maturation a marked change in
<,, 100 A IMPRT LEAF EXPRT 50
°J SOLUBLEa
n2 0 -40 C=
_60 30 tE
W 100 B t20 SOLUBLE FRACTION
EL
C) E-0--1
Flci. 2. Metabolite distribution of '4C-metabolites following ['4CJsu-crose accumulation during leaf expansion.
partitioning occurred with more C being incorporated into thewater-soluble fraction at the expense of secondary biosyntheticproducts. Importantly, the transition in C partitioning occurred at40 to 50% FLL, the stage at which the sink-source leaf conversionbegins. Relatively little free sucrose is found during the importstage. Accompanying the loss of ability to import (and acquisitionof export) the accumulated sucrose becomes less available forbreakdown and subsequent metabolism (Fig. 2B). At full leafexpansion, sucrose accounted for 85 to 90% of the 14C in theneutral fraction. In sink tissue the proportion is reversed. This isconsistent with the translocate atriving at the sink leaf beinghydrolyzed and utilized for metabolism and biosynthesis. As theleaf matures, the need for substrates decreases and more of theassimilates become available for export.
Several questions on the import-export transition warrant in-vestigation. Is leaf maturation accompanied by loss in ability tohydrolyze and metabolize sucrose? Conversely, does the leaf ac-
quire the capacity to synthesize trauslocate as it enters the exportphase? Is there a change in intracellular compartmentation ofeither substrates or enzymes during the import-export conversion?The following experiments address these questions.
Figure 3 shows the partitioning of "CO2-derived photoassimi-lates into various components as a function of leaf development.The 14C distribution in Figure 3, B and C was determined fromthe same leaves used for the photosynthesis measurements inFigure 3A, to reduce plant variability. Similarly to Figure 1,photosynthesis was observed by 20% FLL and reached a maxi-mum from 60 and 100% FLL. During the import stages, '4C is
partitioned between the insoluble and soluble fractions (Fig. 3B)to the same extent as that found following ['4CJsucrose uptake
15
C-)
E 10
I-
80£a-
E
., 60
z
40
Cl)
20
90C-)
' 80
o 70
r 60
C-)50
40) l20 30 40 50 60 70 80 90 100
% FINAL LAMINA LENGTH
50 Uj
I->40 ,r
30 i,0
20 3M
10 t
FIG. 3. Photosynthesis and 14C distribution following '4C02 fixationduring leaf development.
I iASINK - SOURCE
-B I14CO2 DERIVED
A A * H20 SOLUBLE
*
INSOLUBLE
C NEUTRAL FRACTION
SUCROSE
GLUCOSE +FRUCTOSE
I
382 GIAQUINTA
www.plantphysiol.orgon October 8, 2018 - Published by Downloaded from Copyright © 1978 American Society of Plant Biologists. All rights reserved.

SOURCE AND SINK LEAF METABOLISM
into sink leaves (Fig. 2), 60 and 40o, respectively. The dominantform of labeled C reversed during leaf expansion with the transi-tion occurring at 40 to 50%o FLL, a stage consistent with thatobserved in Figures and 2. In the export phase approximately70%o and 30%o of the '4C entered the water-soluble and -insolublefractions, respectively. A marked change in the 14C distributionwas noted in the components of the neutral fraction. Sucrosesynthesized from "CO2 fixation increased during leaf maturationso that by 70 to 100%1o FLL sucrose accounted for 90%o of the "Cin the neutral fraction. Again, it is noteworthy that the markedchange in the dominant species of the neutral fraction occurs at40 to 50% FLL. The increase in sucrose is temporally correlatedwith increase in export. That 50%o of the neutral fraction containedsucrose in importing leaves indicates that this tissue is capable ofsynthesizing sucrose (but on a limited scale, see below) from"CO2. Sucrose has been found in all stages of development inCucurbita (25). However, unlike Cucurbita in which stachyose isthe major translocated sugar, sugar beet translocates mainly su-crose. The increased sucrose synthesized from CO2 during leafexpansion may reflect a transport sucrose pool that is compart-mentalized from the nontransport sucrose found during import.
There is evidence for sucrose compartmentation in varioustissues of exporting leaves (5, 16, 17). We have previously docu-mented (5) the high solute potential of the sieve element-transfercell complex in sugar beet leaves indicating substantial sucroseaccumulation in these cells. In a detailed study of the Vicia sourceleaf, Outlaw and Fisher (16) and Outlaw et al. (17) showedextensive sucrose compartmentation in various cell types includinga strong sucrose partitioning into the minor veins. Furthermore,importing leaves of both sugar beet and bean (4) failed to showphloem loading of either '4CO2-derived or exogenous [14C]sucrose.The increase in sucrose during expansion may, in part, reflect a"histological compartmentation" (17) by phloem loading duringexport. The following data also suggest a more complex partition-ing that involves both an increase in sucrose synthesis and an
intracellular compartmentation of sucrose and/or sucrose-metab-olizing enzymes.Enzymology. Three enzymes are involved in sucrose metabolism
in higher plant cells, sucrose-P synthetase, sucrose synthetase, andinvertase. The latter may be divided into acid and neutral inver-tases based on their pH optima. It is generally accepted thatsucrose-P synthetase (with sucrose-P phosphatase) catalyzes su-
crose synthesis while sucrose synthetase and invertase catalyzesucrose hydrolysis (20). Sucrose synthetase is a reversible enzymeand it may catalyze sucrose synthesis under certain conditions(I 1).The relationship between these enzymes and translocation has
been studied in detail in only one system, the sugar-storing sugar-cane stalks (9). These studies have provided much insight into therole of these enzymes in the mechanism of sugar storage insugarcane. Unfortunately, sugarcane represents a unique systemin which cultivated varieties have been selected because of theirability to store sucrose, and thus mechanisms in this species
probably do not constitute general sink tissue metabolism. Indeed,Giaquinta (8) recently demonstrated that in both the vegetativesink leaf and storage root of sugar beet, sucrose is not extracellu-larly hydrolyzed by a cell wall invertase prior to accumulation as
in sugarcane.In sugarcane, much attention has focused on invertases in
relation to sucrose utilization and storage in the stalks (9). Becauseof the paucity of information on invertase in leaves, their role insucrose metabolism is less clear. The few studies that do exist are
conflicting, showing both increases (14) or decreases (19) in inver-tase activity during leaf maturation. These may reflect differentextraction and assay procedures or species differences.The occurrence of sucrose synthetase and sucrose-P synthetase
is widespread in numerous tissues in many plants (20). The studyby Hawker ( 11) is pertinent to the translocation status of the plant.This study showed that in developing seeds of maize, broad bean,and castor bean, sucrose translocated to the endosperm is con-
verted to hexoses for use in metabolism and biosynthesis. Sucrosesynthetase, which catalyzes cleavage, is high at this stage andsucrose-P synthetase, which catalyzes sucrose synthesis, is low.During seed germination, sucrose-P synthetase activity increasedwhen sucrose synthesis became important and was followed by a
decline in sucrose synthetase activity. More recently, Pollock (18)showed that sucrose-P synthetase activity reached a maximum atligule formation in Lolium leaves and then declined during leafsenescence while sucrose synthetase activity remained unchanged.
Since we have characterized the changes in translocation, pho-tosynthesis, and C partitioning that occur during the import-exporttransition, it was ofinterest to determine activities ofthese enzymesduring the import and export stages. Table I shows the activitiesof invertase, sucrose synthetase, and sucrose-P synthetase in im-porting leaves (15-25% FLL) and fully exporting leaves (75-100%/oFLL). Based on the partitioning data in Figures I to 3, I expectedthat invertase and/or sucrose synthetase activities would decreaseduring leafdevelopment since the leaf loses its ability to hydrolyzesucrose (Fig. 2). (The failure to metabolize sucrose is specificallydue to lack of sucrose hydrolysis since the mature source leavescan extensively metabolize glucose [8].) This was not found to bethe case. Invertase activities were only slightly greater (0-40o) onboth a fresh wt and protein basis in the sink leaves. Also, theinvertase activity from source and sink leaves had a similar Km forsucrose (1.5-2.5 mM). Interestingly, the enzymes capable of hy-drolyzing sucrose are present in both sink and source leaves eventhough source leaves do not break down substantial sucrose. Thisindicates a difference in intracellular compartmentation ofenzymeand/or substrate in sink and source leaves. Importantly, leafdevelopment is not accompanied by the appearance of a neutralor alkaline invertase as in sugarcane stalk (9). The pH dependenceof soluble invertase shows only one peak at pH 5 in both sourceand sink leaves (Fig. 4). This experiment also shows the similarmaximal activities (60 ,umol/hr. g fresh wt) for both tissues.The difference in sucrose hydrolysis in source and sink leaves
does not seem to be due to changes in sucrose synthetase activities
Table I. Invertase, Sucrose Synthetase, and Sucrose Phosphate Synthetase in Sourceand Sink leaves of Beta vulgaris
Enzyme Source Leaf Sink Leaf
pmol/hr.g umol/hr.mg Lmol/hr.g ,amol/hr.mgfr.wt. protein fr. wt. protein
Acid Invertase 44-16 2.8 60-17 3.7Sucrose Synthetase 3.3 0.29 2.5 0.21Sucrose Phosphate
Synthetase 5.0 0.42 N.D. N.D.
N.D. - none detected. Sucrose phosphate synthetase activities were also determined bythe coupled assay method (18). These assays contained in 130 4l: 2 "molUDPG, 5 ,Amol MgC12, 1.25 .Imol 14C fructose-6-phosphate (0.5 WCi), 3 wmol Tris-HCl(pH 7.) and 0.12 mg protein. The rates were 0.62 end 0.15 pmol/hr-mg protein forsource and sink leaves, respectively.
Plant Physiol. Vol. 61, 1978 383
www.plantphysiol.orgon October 8, 2018 - Published by Downloaded from Copyright © 1978 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 61, 1978
70
I.
I-
C
c
o
II-
w
z
30 _
10
3 4 5 6pH
7 8 9
FIG. 4. pH dependence of soluble invertase in the source and sinkleaves. Rates at pH 5 and 6 and 4 ,umol/hr mg protein for sink andsource, respectively.
because of the similar levels found in both tissues (Table I). Thespecific activity of sucrose synthetase was the lowest of the threeenzymes determined in mature leaves.
Sucrose-P synthetase, which catalyzes sucrose synthesis, was notdetected in importing leaves but was present in exporting leaves.Since the limit of resolution for this assay was 0.5 to 1 ,umol/hr gfresh wt, the source leaves contained at least 5 times the activityof this enzyme than did sink leaves. The absolute rates for alithree enzymes agree well with those found in mature leaves ofvarious species (10, 19, 20).The decreased ability to synthesize sucrose from CO2 in sink
leaves (Fig. 3) could be associated with either the much reducedlevels of sucrose-P synthetase or the reversal of sucrose synthetase.Hawker (11) suggested such a reversal ofsucrose synthetase duringseed germination. The additional sucrose synthesized from CO2during the onset of export probably results from the increasedactivities of sucrose-P synthetase found in source leaves. Thissucrose possibly reflects the sucrose pool which is available forphloem loading and translocation, and is somehow isolated fromthe nontransport sucrose pool.Metabolism ofTranslocate In Sink Leaves. Several studies have
documented the differences in '4CO2 fixation patterns in youngand mature leaves (2, 12). Few studies have been directed towarddetermining the initial and secondary products formed from su-crose during translocation into sink leaves. Recently, Dickson andLarson (3), as part of their continuing study of developing poplarleaves (13) have examined the distribution products from bothphotosynthesis and translocation to determine the relative C con-tribution of each during leaf expansion.The partitioning of "C-translocate in importing sugar beet sink
leaves was determined to document the metabolic fate ofincomingtranslocate. The time course of metabolism of 14CO2-derivedtranslocate arriving in the sink leaves of intact sugar beet plantsis shown in Figure 5. Initially, all of the C entering the leaf iswater-soluble (>90%) but after 5 hr, 60 to 65% of the "C has beenincorporated into the starch (25-30%), protein (25-30%), lipid(5%), and residue (5%) fractions (Fig. 5A). These distributionspertain only to the C incorporated into the leaf and not that lostthrough respiration. Both components would be necessary todetermine the total C economy of the leaf. Figure 5B furthershows that the neutral or sugar fraction predominates at the initialstages of arrival and then declines, concomitant with an increasein the acid- (organic acids and sugar monophosphates), basic(amino acids), and acid-2 (phosphoglyceric acid and sugar di-phosphates) fractions. Figure 5, C and D resolves the sugar andacid-l fractions, respectively. Sucrose arrival is followed by hy-drolysis and the appearance of hexoses, with the equilibrium
X OAC,!ID-2 -
t 60 NEUTRAL FRACTION C
CZ SUCROSE GLUCOSE
01-~~~~~~
40
20 ld,0 _FRUCTOSE
C,.80- ACID-I FRACTION D
SUGAR PHOSPHATES 7ALATE
60 1
40
CITRATE
20
1 3 4 5 6 7 8 9 102TIME (hr)
FIG. 5. Partitioning of '4C-translocate after arrival in the sink leaf.
toward glucose, particularly after 1 to 2 hr (Fig. SC). Sugarmonophosphates rapidly decrease in the acid-l fraction followedby a marked increase in malate and, to a lesser extent, citrate.Label in other organic acids such as glycolate, succinate, andfumarate was determined but their total represents only about10% of the acid-l fraction.These data represent the arrival of '4CO2-derived translocate in
sink leaves of different intact plants, and thus the system is notamenable to study over the very short times (seconds to minutes)needed for precursor-product relationships. Nevertheless, the re-sults do give some insight into the general trend of partitioning oftranslocate into various components. Much of the translocate(60-65%) is incorporated into insoluble components within 5 hrafter arrival. Sucrose is readily hydrolyzed to hexoses some ofwhich are phosphorylated and either enter the glycolytic pathwayfor respiration or become precursors for biosynthetic processes.Substantial amounts of "4C enter a malate pool which is notrapidly turning over, suggesting that it represents a storage ratherthan a respiratory pool. High malate levels have been noted inyoung tissues (3).
It is important to note that the distribution of '4CO2-derivedtranslocate in sink leaves of intact plants closely parallels that
....... .SOURCE
-SINK
II I I~~~~~~~~~~~~~~~~~~~~
384 GIAQUINTA
50 _
www.plantphysiol.orgon October 8, 2018 - Published by Downloaded from Copyright © 1978 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 61, 1978 SOURCE AND SINK LEAF METABOLISM 385
found following the accumulation of exogenous ["4C]sucrose insink leaf tissue (Fig. 2). Since the data in Figure 2 represent thatfollowing a 30-min sucrose uptake period followed by a 1-hr washbefore extraction, the "C distribution 2 hr after arrival of trans-locate in sink leaves ofthe intact plant (Fig. 5) should approximatea similar time point in metabolism. The following distributionswere noted from Figure 2 versus Figure 5, respectively: starch andresidue, 34 versus 25%; protein, 25 versus 20%; water-solubles, 40versus 35% (basic, 30 versus 25%; acid, 30 versus 30%; neutral, 40versus 43% [sucrose, 10 versus 25%; and hexoses, 90 versus 75%1).The excellent agreement between values indicates that the eventscharacterized at the tissue level (Fig. 2) translate well to themetabolism of translocate in the intact translocating plant (Fig.5).
Intraceflular Compartmentation. Previous data have shown thatthere is a compartmentation of sucrose within the various cells ofexporting leaves (5, 17). Based on the C partitioning and enzymedata presented here along with some modifications of the schemesof compartmentation proposed for sugarcane stalks (9) and beanpod tissues (21) it is possible to speculate on the intracellularcompartmentation of sugar in sugar beet source and sink leaves.In the sink leaf, sucrose enters the mesophyll without prior extra-cellular hydrolysis (8). Once in the cytoplasm, sucrose is possiblytransported to the vacuole where it is hydrolyzed by acid invertase.The vacuole in this scheme would be a reservoir for hexosesderived from sucrose hydrolysis. Hexoses could then enter thecytoplasm by diffusion (simple or facilitated) in response to aconcentration gradient generated by hexose utilization in thecytoplasm. Thus, the metabolic need of the cell could controlhexose levels and utilization. A diffusional movement of sugarsfrom the vacuole into the cytoplasm has been proposed duringsugar storage in sugarcane cells (9). The nontransport sucrose pool(a possible protective derivative of glucose) synthesized from"CO2 (Fig. 3) presumably occurs in the cytoplasm and may be inreversible equilibrium with the vacuole hexose pool depending onthe requirements of the cell.
Exporting sugar beet leaves have sucrose-P synthetase to syn-thesize substantial sucrose from "CO2 fixation. Based on theenergy-coupling requirements and the number of enzymes andsubstrates needed for sucrose synthesis by this pathway, sucrosesynthesis occurs in the cytoplasm (20). The data indicate twointracellular sucrose pools, one earmarked for export from themesophyll into the apoplast for phloem loading (6, 7), the other anontransport pool which can enter the transport pool dependingon the demand for exported assimilates. Acid invertase is presentand may regulate sucrose breakdown in response to metabolicneed. However, there would be less biosynthetic requirement forglucose in exporting leaves and thus less sucrose would be hydro-lyzed. Evidence suggests that sucrose is both in vacuoles andcytoplasm of exporting leaves (17). Since source leaves containsignificant invertase activity, an intriguing question is what typeof compartmentation exists between the invertase and sucrosepools. The compartmentation may be spatial, kinetic, or regula-tory. The activities of invertase in both source and sink leaves(40-60 ,mol/hr g fresh wt) are capable of hydrolyzing moresucrose than is present in these leaves (5-10,umol/g fresh wt insource leaves), thus further obscuring their function.
Recent studies by Swanson et al. (23) strongly suggest theexistence of two intracellular sucrose pools. Moreover, these stud-ies showed that the nontransport sucrose pool can be drawn uponfor export very rapidly (to. = 9 min) during times of increased
sink demand, suggesting a ready intracellular communicationbetween the pools.
Admittedly, the above scheme is probably an oversimplificationof the compartmentation that occurs in exporting and importingleaves. One particular aspect that has been generally accepted, butnevertheless still unproven, is that acid invertases, because of theirlow pH optima (pH 4-5), are vacuolar enzymes. Recent data haveshown that some classes of enzymes with low pH optima such asacid hydrolases and carboxypeptidases may occur in the cytoplasmof plant cells (G. Wagner, personal communication).
Since the sucrose concentration in the leaf has been shown togovern the translocation rate (1, 22), further study of the eventsregulating intracellular C partitioning is ofparamount importancein understanding the translocation process.
Acknowledgments-The expert technical assistance ofC. Arrabal and the manuscript preparationby D. Bacon are gratefully acknowledged.
LIMRATURE CITMD
1. CHRISTY AL, CA SWANSON 1976 Control of translocation by photosynthesis and carbohydrateconcentrations of the source leaf. In IF Wardlaw, JB Passioura, eds, Transport and TransferProcesses in Plants. Academic Press, New York, pp 329-337
2. DICKMANN DI 1971 Photosynthesis and respiration by developing leaves of cottonwood(PopuhlI deltoides Bans.) Bot Gaz 132: 253-259
3. DICKSON RE, PR LARSON 1975 Incorporation of"C-photosynthate in major chemical fractionsof source and sink leaves of cottonwood. Plant Physiol 56: 185-193
4. FELLOWS RJ, DR GEIGER 1974 Structural and physiological changes in sugar beet leavesduring sink to source conversion. Plant Physiol 54: 877-885
5. GEIGER DR, R GIAQUINTA, SA SOVONICK, RJ FELLOWS 1973 Solute distribution in sugarbeet in relation to phloem loading and translocation. Plant Physiol 52: 585-589
6. GtAQuiNTA R 1976 Evidence for phloem loading from the apoplast. Chemical modification ofmembrane sulfhydryl groups. Plant Physiol 57: 872-875
7. GtAQUINTA R 1977 Possible role of pH gradient and membrane ATPase in the loading ofsucrose in the sieve tubes. Nature 267: 369-370
8. GIAQUINTA R 1977 Sucrose hydrolysis in relation to phloem translocation in Beta vulgaris.Plant Physiol 60: 339-343
9. GLAszIou KT, KR GAYLER 1972 Storage of sugars in stalks of sugarcane. Bot Rev 38:471-49010. HAWKER JS 1967 Activity of uridine diphosphate glucose-D-fructose-6-phosphate-2-glucosyl-
transferase in leaves. Biochem J 105: 943-94611. HAWKER JS 1971 Enzymes concerned with sucrose synthesis and transformations in seeds of
maize, broad bean and castor bean. Phytochemistry 10: 2313-232212. JoYKW 1964Translocation in sugar beet. I. Assimilation of `COs and distribution of materials
from leaves. J Exp Bot 15: 485-49413. LARSON PR, RE DICKSON 1973 Distribution of 14C in developing leaves of Eastern cottonwood
according to phyllotaxy. Planta I 11: 95-11214. MANNING K, GA MAW 1975 Distribution of acid invertase in the tomato plant. Phytochemistry
14: 1965-196915. NAYLOR DG, ID TEARE 1975 An improved, rapid, field method to measure photosynthesis
with "CO2. Agron J 67: 404-40616. OUTLAW WH, DB FISHER 1975 Compartmentation in Viciafaba leaves. I. Kinetics of '4C in
the tissues following pulse labeling. Plant Physiol 55: 699-70317. OUTLAW WH, DB FISHER, AL CHRISTY 1975 Compartmentation in Vicia faba leaves. II.
Kinetics of "4C-sucrose redistribution among individual tissues following pulse labeling.Plant Physiol 55: 704-71 1
18. POLLOCK CJ 1976 Changes in the activity of sucrose-synthesizing enzymes in developing leavesof Lolium temulentum. Plant Sci Lett 7: 27-31
19. POLLOCK CJ, El LLOYD 1977 The distribution of acid invertase in developing leaves of Loliumtemulentum L. Planta 133: 197-200
20. PONTIS HG 1977 Riddle ofsucrose. In DH Northcote, ed, International Review of BiochemistryII, Vol 13. University Park Press, Baltimore, pp 79-117
21. SACHER JA 1966 The regulation of sugar uptake and accumulation in bean pod tissue. PlantPhysiol 11: 181-189
22. SERVAITES JC, DR GEIGER 1974 Effects of light intensity and oxygen in photosynthesis andtranslocation in sugar beet. Plant Physiol 54: 575-578
23. SWANSON CA, J HODDINOTT, JW SIJ 1976 The effect of selected sink parameters on translo-cation rates. In IF Wardlaw, JB Passioura, eds, Transport and Transfer Processes in Plants.Academic Press, New York, pp 347-356
24. TURGEON R, JA WEBB 1975 Leafdevelopment and phloem transport in Cucurbitapepo: carboneconomy. Planta 123: 53-62
25. TURGEON R, JA WEBB 1975 Physiological and structural ontogeny of the source leaf. In SAronoff, J Dainty, PR Gorham, LM Srivastava, CA Swanson, eds, Phloem Transport.Plenum Press, New York, pp 297-313
www.plantphysiol.orgon October 8, 2018 - Published by Downloaded from Copyright © 1978 American Society of Plant Biologists. All rights reserved.





























