Embed Size (px)
Citation preview

Some Aspects of the Foraging Behaviour of Bumble-BeesAuthor(s): Aubrey ManningSource: Behaviour, Vol. 9, No. 2/3 (1956), pp. 164-201Published by: BRILLStable URL: http://www.jstor.org/stable/4532844 .
Accessed: 25/06/2014 10:52
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
BRILL is collaborating with JSTOR to digitize, preserve and extend access to Behaviour.
http://www.jstor.org
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

SOME ASPECTS OF THE FORAGING BEHAVIOUR OF BUMBLE-BEES
by
AUBREY MANNING 1) (Department of Zoology and Comparative Anatomy, Oxford.)
(With 5 Figures) (Rec. 15-IX-I955)
INTRODUCTION
HERMANN MIULLER (I882) wvas the first to discover that honey-bees, when foraging, confine their attention to a relatively small group of flowers and return to them on many successive journeys. He marked bees feeding on Salvias, and found that some of them were still visiting the same plants after ten days. Later GILTAY (1904) found honey-bees returning daily to the same plots of poppies. BONNIER (i906) showed that bees stayed within a small area even if the flower crop was very abundant and continuous. He marked honey-bees feeding on a 5 metre long strip of a long belt of buckwheat, and found that on the next day, only marked bees were feeding on the strip and no other marked ones were more than a few feet from it.
Since then many people have confirmed and extended these observations; FREE (1953) and others have found that bumble-bees forage in the same way, and recently interest has shifted to the study of how a bee's foraging area may vary in size, with the state of the crop and its nectar secretion, and with competition from other bees. Thus SINGH (I950) finds that as a crop wanes the size of foraging area increases, but that individual bees also vary greatly upon the same crop. RIBBANDS (I949) watched bees foraging
i) It is a pleasure to thank Dr N. TINBERGEN for his constant and invaluable help at all stages of this work. 1 am very grateful to Prof. A. C. HARDY, F.R.S. for his hospitality in the Oxford Zoology Department and his continuing interest and encourage- ment. Among my colleagues I must thank especially Dr MARGARET BASroCK for all her assistance Dr M. R. SAMFORD analysed the results statistically. Dr H. PRECHTL kindly prepared the German Summary.
Part of the equipment used was made available by a grant from the Nuffield Foun- dation at the disposal of Dr TINBERGEN, and all the experiments were carried out on the University Estate at Wytham. Finally, I must express my gratitude to the Nature Conservanicy, from whom I held a research grant throughout the period of this work.
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

FORAGING BEHAVIOUR OF BUMBLE-BEES I65
on a specially planted bed of flowers, and could correlate the variations in foraging areas with the number of flowers available, the amount of nectar or pollen they were yielding and also the amount of competition from other bees on the same beds. The size of the areas varied greatly, from a foot or so around a single favoured flower to hundreds of square feet.
Very little is known of how bees learn their foraging areas or how, if at all, they change in their responsiveness to the various stimuli from the flower as a result of their knowledge of its position. After visiting one type of plant for some time, a bee may respond to different features even though it does not know the position of each plant. In this connection some observations of DARWIN (page 224 of 'Cross and self-fertilization of plants', I876) are particularly relevant. He says,
'Improbable as it may appear, they (bees) seem, at least sometimes, to recognize plants even from a distance by their general aspect, in the same manner as we should do. On three occasions I observed bumble-bees flying in a perfectly straight line from a tall larkspur (Delphinium) which was in full flower to another plant of the same species at the distance of fifteen yards which had not as yet a single flower open, and on which the buds showed only a faint tinge of blue. Here neither odour nor the memory of former visits could have come into play, and the tinge of blue was so faint that it could hardly have served as a guide'.
Here is, so far as I know, the first suggestion that bees can become conditioned to the general appearance of a plant as opposed to its flowers. Some further observations by DARWIN in the same chapter give some evidence that a bee's knowledge of the position of individual plants within its general foraging area may be very precise. He says (page 426),
'Memory also comes into play, for, as already remarked, bees know the position of each clump of flowers in a garden. I have repeatedly seen them passing round a corner, but otherwise in ast straight a line as possible, from one plant of Fraxinella and of Linaria to another and distant one of the same species; although, owing to the interven- tion of other plants, the two were not in sight of one another'.
RIBBANDS also mentions how one honey-bee often began its foraging trips on the same individual flower within its foraging area.
There are many fascinating problems of behaviour concerned here. How do bees learn their foraging area, is it a slow process or do they rapidly orientate to flowers as they first find them? How do the effective stimuli from a flower change, firstly as a bee learns its location, and secondly as it reacts to other flowers of the same type whose position it does not know ? To investigate these problems one needs to follow bees foraging from the first days when they begin visiting a particular plant, and to try to analyse the stimuli to which they are reacting at various intervals. Dr N. TINBERGEN told me of some observations he made on bumble-bees
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

i66 A. MIANN1NG
visiting the flowers of the Hound's-tongue (Cynoglossum officinale L.), which showed that the bees sometimes had a very detailed knowledge of the site of inidividual plants, and also that they became conditioned to the general form of the plant. I was able to repeat and extend these observations, and this paper is an account of three seasons' work on the foraging beha- viour of various bumble-bee species on Hound's-tongue. The results throw some light on the questions just outlined.
THIE FORAGING BEHAVIOUR OF BUMBLE-BEES ON
HOUND'S-TONGUE
A description of Hound's-tongue will be a useful preliminary. It is a member of the family Boraginaceae and is a biennial flowering in its second summer. It is a fairly large plant, standing between 2 and 3 feet (60-go cm) high, with usually a single upright stem which bears the grey- green, rather hairy leaves. These are large and close together at the base, becoming smaller and more spaced towards the top of the stem. The flowers are borne in cymes arising from the axils of the upper leaves, and as the flowering season advances, the stalks of both the individual flowers and the inflorescence as a whole, elongate so that they stand out more from the stem and the leaves (see P1. X, Fig. i and 2). The flowers are small, about 8 mm in diameter, and the petals are fused at their bases to form a shallow tube. They hang down slightly, so that from the side the petals are largely obscured by the hairy green sepals. Their colour, at first, is a deep brownish-red with a trace of blue in it, and this latter rapidly in- creases as the flower ages, and after I-2 days the petals are a bright purple. The flowers either fall off or wither, leaving the fertilized seeds surrounded by the calyx.
The flowering season is spread over about a month, from the middle of May, and it is the topmost cymes which flower first; those in the lowest axils ido not develop until those above are finished. 3 or 4 flowers of each inflorescence are out at any one time; about 2 of these will be old flowers, already purple, while the others are young and bright brownish-red. The whole plant, stem and leaves as well as flowers, has a strong mousy smell, especially when bruised, and to the human nose the leaves smell almost as strongly as tfhe flowers. The scent is most probably due to the same substance that gives impure acetamide its characteristic smell.
When visiting a plant a bumble-bee almost always flies to the base of the stem and alights first on the lowest cymes, working up to those at the top before leaving. It clings upside down to the calyces of the group of flowers,
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

FORAGING BEHAVIOUR OF BUMBLE-BEES I67
and thrusts its tongue into each in turn, moving about as is necessary. On any plant bees usually visit all the cymes that have open flowers, and probe all the flowers on each, with the exception of the old purple flowers which they often avoid.
The length of time that a bee spends on a plant varies greatly, and it is almost certainly dependent upon how much nectar the flowers hold. I have hardly investigated this problem, but it is most noticeable that if a new plant, which has not been visited by bees for several hours, is put out, the bee which first visits it spends a very long time on each group of flowers. It is likely, then, that a bee will extract all the nectar that it can from each flower before passing on. The bees can have no means of telling which flowers hold abundant nectar, nor with a fairly compact group of plants, do they appear to memorize well which ones they have recently visited. Sometimes they visit the same plant 2 or 3 times within a few minutes, and from the shortness of their later visits it is clear that hardly any nectar has been secreted during the brief interval since they last visited it, even though, as is most likely, H-ound's-tongue flowers secrete nectar continuously, at least during the day.
Hound's-tongue proved to be an excellent plant with which to study the foraging behaviour of bees, for although it grows in groups so that one can follow bees from plant to plant, the plants are never so close together that there is any doubt as to which plant they are reacting. In the summer of I952 I began watching bumble-bees foraging on such a group.
From these preliminary observations and some simple experiments, three main facts emerged.
I) Individual, marked bumble-bees will spend several days foraging on the same group of plants.
2) They come to learn the sites of individual 'distant' plants, i.e. those separated by IO-I2 feet (3-3.7 m) from any other, with great detail. If such a plant is removed, bees continue, for some time, to fly to the site and hover there.
3) Bumble-bees also become conditioned to the general form of Hound's- tongue plants. They continue to visit plants from which all the flowers have been removed and search in the leaf axils, where flowers are normally borne, as if looking for them. They also frequently fly to and search other plants, such as Ragwort (Senecio jacobea) and Figwort (Scrophularia no- dosa), which are not flowering at the same time, and treat them just as flowerless Hound's-tongue.
It is obvious that conditioning both to place and form play a very large parts when bumble-bees are foraging on Hound's-tongue. The actual flowers
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions
i68 A. MANNING
are unimportant in all but the final stages of the chain of reactions which make up the bee's visit to a plant. As mentioned above Hound's-tongue's flowers are small and inconspicuous and I thought it would be interesting to compare the foraging behaviour of bumble-bees on another plant with very large flowers. In this way one might discover how far Hound's-tongue
24.9
23S
22.
.25
21g
26 20.
29*0
.27
7.: 28
'I 3~ ~~ 126
20 8
50I * 6* 1| ( Approx. scale of yards (2 yd _o.9i m.)
3' Central Area'
2.
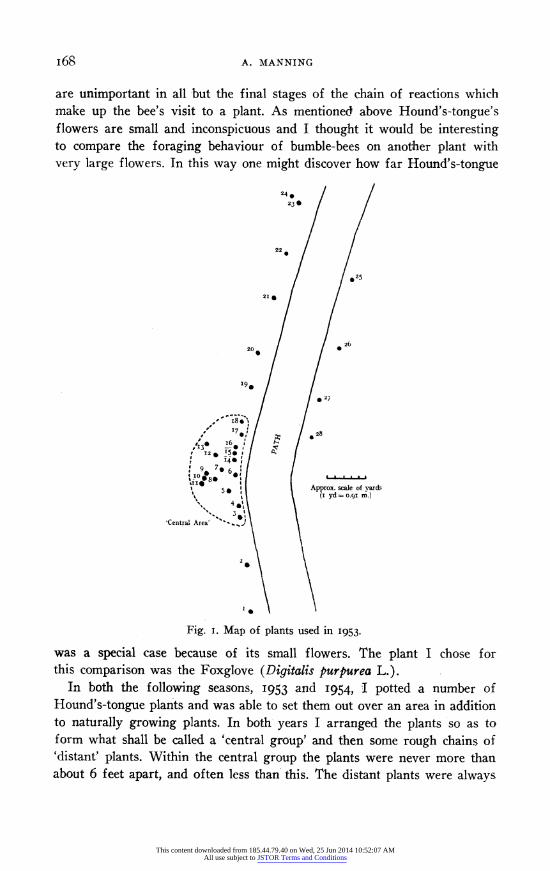
Fig. i. Map of plants used in I953.
was a special case because of its small flowers. The plant I chose for this comparison was the Foxglove (Digitalis purpurea L.).
In both the following seasons, I953 and 1954, I potted a number of Hound's-tongue plants and was able to set them out over an area in addition to naturally growing plants. In both years I arranged the plants so as to form what shall be called a 'central group' and then some rough chains of 'distant' plants. Within the central group the plants were never more than about 6 feet apart, and often less than this. The distant plants were always
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

FORAGING BEHAVIOUR OF BUMBLE-BEES I69
more than IO feet from each other and from any plant in the central group. For illustration a map of the plants used in I953 is given (Fig. I). The great advantage of plants in pots is that they can be moved and replaced at will, and it is easy to put out plants in new positions for a short time. Also one can be certain that no traces of a plant's scent are left behind when it is moved, which could be so if it was just uprooted.
My plan will be to give first an outline account of the general sequence of the bees' foraging behaviour through the season, and then to deal separately with those aspects on which there is most evidence, in greater detail.
THE GENERAL FORAGING CYCLE
At the very beginning of the Hound's-tongue season, as the first flowers appeared, I spent some time each day by the central area where there was the biggest concentration of plants, watching for the first visits of bees. In both years it was noticeable that bees do not find the plants for some time after the first flowers open, even though these earliest flowers, because they open before any of the stalks of the inflorescence have grown, are the most conspicuous because they point directly upwards, see Plate X, Fig. I. For instance, in I953 the first flowers opened on gth May, and by i8th May, II plants had open flowers and these held abundant nectar, yet I saw no bees visiting them until 22nd May. It is, of course, impossible to be certain that no bees had been before, but the copious uncollected nectar in the flowers does not speak of many visits, and certainly no bees had begun to visit the flowers regularly. In 1954 the comparable dates were: first flowers on 14th May, several plants with flowers containing nectar by 20th May, and I first recorded visiting bees on 26th May. According to KNUTH (I906), the flowers are self-fertile if not visited, but in Wytham (where I did most of my work with bees) it is common for the first flowers on the topmost inflorescences to set no seed. This is very rare for the later flowers, and again indicates that the earliest ones were not visited at all while they were out.
The first visits are paid by bees which are flying over the area and hap- pen to pass within a couple of feet of a plant and dip down to alight. They are bees which are feeding regularly on other crops, for they often carry pollen which is not collected from Hound's-tongue. Now these very first reactions must be released by the flowers of the plant, as opposed to its form. I have never seen bees react to plants without some open flowers be f ore they have visited ones with flowers, and bees will make their first visits (so far as I know) to Hound's-tongue plants which have had all
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

170 A. MANNING
their leaves removed, leaving just the flowers and stalk (see P1. XI, Fig. I).
Also during the course of the first few visits, paper models of flowers on a piece of wire are often reacted to before plants without flowers.
While the colour of the flowers must be important in releasing these first reactions, their scent may be involved also, but there is no evidence that it does in fact have any influence on the primary reactions. The paper models have no scent, and plants without open flowers are not visited at the very beginning, although to the human nose the unopened buds smell as strongly as the flowers. Plants with open flowers which are given the wrong scent by adding rosewater, are also visited from the first, but this in itself does not rule out the role of scent as a distance attraction. It is certain that bees which are not conditioned to any particular scent are attracted by a wide range of flower odours.
Usually at its first recorded visit a bee just alights on one or two plants and then flies off. It often has a definite line of flight and only sees plants near this line. I have not been able to catch and mark bees on such visits, because they are normally very brief. The shortness of these early visits is certainly not due to poor nectar supply, but simply because the bees are conditioned to feeding on other plants. Soon, often during the course of the first day, there are some bees which stay on the central group a little longer, and visit perhaps five or six plants-often the same plant twice or three times-before flying off, and these it is sometimes possible to mark. It is reasonable to assume that these bees have been to the plants several times, and sometimes peculiarities in their colouring enable one to fix this as at least their second or third visit.
Very quickly, even by the second recorded visit, bees begin to react occasionally to plants without open flowers and to plants other than Hound's-tongue which have no flowers showing. Thus after a few visits to Hound's-tongue bees become conditioned to the general form of the plant and react to such a form from a distance, using its stimuli in addition to, and often to the exclusion of, those from the flowers. For example, on 2815[54 a Bombus pratorum worker on its second recorded visit to the central area, alighted on 6 plants with open flowers, then went to one with no open flowers, did not alight, went on to another with flowers and then to another without, and then to a small Privet (Ligustrum vulgare) plant to which it reacted briefly, went on to a second Privet plant, and finally flew off. On 22j5[53 a pratorum worker on its fourth recorded visit, alighted on 3 plants with flowers and then reacted to a Stinging Nettle plant, which it searched briefly, exploring the leaf axils where a Hound's- tongue plant would bear its flowers, before flying off.
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

BEHAVIOUR, IX PLATE X
S~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~ .. ..f.. ...
A.-_,Ws A
Fig. i. Fig. 2.
Fig. i. Young Hound's-tongue plant; the peduncles 'have not yet elongated and the few flowers that are open point upwards.
Fig. 2. Mature plant with elongated peduncles and pedicels and the flowers hanging down.
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

BEHAVIOUR, IX PLATE XI
'N' ~ ~ 'M~~~~~~~~~~~~~~~~~~~~~~4
* ~ ~ ~~~~foes removed
Fig. 2. A Foxglove:plant(Digitalispurpurea)withtwoflo : wer s p i
X , :,S ? '., . '''' ' j~~~~~~~~~~~~~N _T
! ; : ' r W i . ~~~ ~~~~~. li i. '
: ' S e !~~~~~~~~~~~~~~~~~~~~~~~~ 1 iS? iW_ a s . ~~~~~~~~~~~~~~~~~~~~-m > . 3...... .. .. ;~~M~
Fig. .Fig. 2.
Fig. I. a (right) Hound's-tongue plant with all leaves removed. b (left) Plant with all f lowers removed.
Fig. 2. A Foxglove plant (Digitalis purpurea) with two flower spikes.
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

FORAGING BEHAVIOUR OF BUMBLE-BEES I71
It is very likely that this conditioning to form enables bees to see plants from a greater distance than if they reacted to the flowers alone. It is dif- ficult to get exact evidence on the distance from which bees can perceive plants: all one can do is put out plants in new positions and try to see from how far bees first react to them directly. Such records at least give one a minimum figure. Thus I have seen direct reactions to a plant from which all the flowers had been removed, (see P1. XI, Fig. ib), from 6' away, but to a plant with some open flowers but no leaves, never from more than 2'. This latter figure is for a plant early in the season before the inflorescence stalks have elongated: later on when they have done so, removing the leaves would probably have less effect, because the numerous long flower stalks and calyces are very consplicuous, though the actual flower petals are almost hidden.
As bees begin to spend more time on the Hound's-tongue plants, their reactions to flowerless plants and to plants of other species become more common. They react to and search briefly a wide range of plants, and the requirements of form, which will be discussed later, are clearly very general. As mentioned in the description, the whole Hound's-tongue plant, apart from the flowers, has the characteristic smell, but it is fairly certain that bees are not reacting to this from any distance, and that form is the U
important stimulus. The evidence against scent can be summarized as foltows:
I. Young, first-year Hound's-tongue plants which have the right scent but the wrong form are not reacted to. They have just a cluster of large radical leaves with no stem.
2. Objects with the wrong form to which I gave the correct scent, by rubbing them with bruised Hound's-tongue plants, were not reacted to, though I did these experiments to only a very liimited extent.
3. Hound's-tongue, and other plants without flowers which are artificially given the wrong scents, are reacted to and searched.
4. Other plants which lack the correct scent, or even have a strong natural scent, e.g. Ragwort (Setnecio jacobea) and Elder (Sambucus nigra), are reacted to and searched.
5. I could detect no influence of wind direction; bees appeared to react just as frequently down wind as up it.
After several days from the first visits some bees begin to spend long periods on the Hound's-tongue, since there are now enough flowers to occupy them. Such bees can be followed for hours on end with very short breaks when they may be visiting other Hound's-tongue plants which I do not know of, or back at their nests disgorging the nectar. The two species Bombus
Behaviour IX I2
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

172 A. MANNING
pratorum and agrorum are the commonest to specialize on Hound's-tongue; ruderarius and lapidarius do so occasionally, and for a few days during the 1953 season one or two honey-bees (Apis mellifera) did so as well. 1)
Throughout, I could detect no differences in the behaviour of the different species and I have always pooled results. Towards the end of the season pratorum males are frequent visitors and, just as with the workers, indivi- duals are very faithful to the group, returning day after day, and apparently spending a large proportion of their time foraging. This fact was rather surprising, for Bombus males are always supposed to be very desultory foragers and to spend most of their time flying from one to another of their scent posts, as described by FRANK (i941) and HAAS (I949) among others. However, I found the same thing with males of ruderatus and hortorum visiting Foxglove, for they too were very regular in their foraging and behaved in just the same manner as workers. So far as I know, no-one has recorded males as ever taking back food to the nest, and it is very unusual for them to return at all after they first leave; thus when they leave the nest I have noticed that they do not orientate as do the young workers.
Soon the bees that are specializing are visiting all the plants of the central group, and they show some signs of knowing their positions; for instance, they may fly very directly around a bush which partially hides a plant (a similar observation to that of DARWIN (1876), which is quoted on p. I65). However, in general their flight is rather wavering as they fly from one plant to another, usually the next nearest, and they frequently react to and search other plants as they do so. It is notable that they will react to the same flowerless plants on the same sites again and again, and clearly they never become negatively conditioned to a form or a site.
When bees are specializing, there begins, sometimes on the first day that a bee specializes, a characteristic piece of behaviour. A bee will visit all of the plants in the central area, some perhaps more than once, and then when leaving a plant at the edge of the group, it flies off fairly slowly and makes wide ranging sweeps over the ground at a height of about I'6"-2' above the ground. It gradually moves off in this fashion in a given direction, but often turns back and doubles over an area again before going on. I think there is little doubt that such a bee is searching for more Hound's- tongue plants. There are three pieces of evidence in favour of this:
i. With a very few exceptions bees on such flights react to and visit any
i) It may be noted here that the behaviour of these honey-bees with respect to Hound's tongue was, so far as I observed, completely similar to bumble-bees; for instance they learnt the sites of individual plants and became conditioned to the general forms of Hound's-tongue.
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

FORAGING BEHAVIOUR OF BUMBLE-BEES 173
Hound's-tongue plants which they come near to and which, so far as I know, they have not visiteid before.
2. Bees on such flights sometimes react to other plants such as Privet and Ash (Fraxinus excelsior) which they often react to on the central area and search as if they were Hound's-tongue.
3. Such bees do not, with rare exceptions, react to other very conspicuous flowers, e.g. Bugle (Ajuga reptans), Ground Ivy (Glechoma hederacea) and Buttercup (Ranunculus sp.), which they pass very close to.
These 'exploration flights' are often very prolonged and frequently my bees were lost to sight still ranging over and among the bracken which surrounded much of the centeral area. It seems as if the bees are seeking to extend their foraging area on Hound's-tongue, possibly because, as more bees feed on the central group, there is less nectar available to each. What- ever their cause, there can be no doubt as to the efficiency of the searching flights. Within the course of a few hours bees have found all the distant plants around the central group, including those hiidden among thick bracken and quite invisible from more than about 2'. Thus on 2615153 a red-marked pratoruin worker was feeding on the central group during the morning. It then began exploration flights, and during the day found for the first time (so far as I know, but there was the confirming evidence that it often made orientation flights after alighting on a plant - see below) all the other distant plants except I and 2 (see map, Fig. I, p.i68). In one flight it left plant I8 and found 19, 20, 2I and 22, 23 and 24; in this it was probably helped by their being in a rough chain along its line of flight-its nest was certainly in this direction from the central group-but even so it made wide detours between every plant, circling over the bracken to either side, and certainly did not take anything like a direct route between them. It appa- rently found each plant by chance, reacting to it when it happened to pass within a few feet and it could see it.
Scent may play a part in helping bees to locate plants, but I have seen bees pass within a f ew inches of a plant, which was probably out of sight because of other vegetation, and not discover it. Also there is no sign that bees are better able to find plants which lie upwind of them.
The effectiveness of exploration flights in enabling bees to increase their foraging area, is greatly enhanced by the fact that in about 75% or more cases, a bee makes a brief, circling, orientation flight when it leaves the newly- discovered plant. These orientation flights will be discussed in more detail later, but it can be saiid here that they enable a bee to fix the site of a plant after one visit, and that subsequent visits with further orientation eventually mean that a bee has a very exact knowledge of the position of all the distant
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

I74 A. MANNING
plants. 'Correlated with this, one finds that a bee has a distinct 'route' which it follows between the distant plants, and that its flight when visiting them is very direct, though it may still make wavering exploration flights to either side of its route on some trips. When visiting distant plants whose posi- tions they know, bees rarely react to plants other than Hound's-tongue which they pass, and on such direct flights they often ignore new Hound's-tongue plants with flowers which are put out right in their path, and may pass within inches of them for several successive trips before alighting. This behaviour is in marked contrast to that when they are visiting plants within the central group. Here, as mentioned before, bees have a very wavering flight; they repeatedly react to and search other plants than Hound's-tongue and any new Hound's-tongue plants put out, are usually visited the first time bees pass near them. Correlated with this behaviour, experiments show that in the central area, bees are relying much more upon direct stimulation from plants, although they have some vague knowledge of the position of plants which they have been visiting for some time.
The bees go on foraging in this way for three weeks or inore, until the plants begin to decline, though I have never been able to follow an indiyi- dual for more than 12 days. The number of bees visiting falls off rapidly towards the end of the flowering period, but some odd bees coiltinue for a long time. Hound's-tongue plants often send out small cymes from the lowest leaf axils as a sort of 'secondary crop'.
Having traced the general foraging cycle, we may now consiider some aspects of the behaviour in more detail. They can be dealt with under the three headings, 'Learning the position of plants', 'Learning the form of plants', and 'The role of stimuli from the flowers' but these are not sharply distinct, and often the results of experiments bear on more than one section and may be mentioned accordingly.
LEARNING THE POSITION OF PLANTS
If a bee finds a new plant with flowers during the course of a searching flight, it very often orientates upon leaving. This takes the form of a brief circling (1-2 secs) around the site, and very frequently the bee alights for a second and even third time, with circlings in between, before it finally flies off. This type of orientation flight resembles in all respects a curtailed form of the thorough orientation which bees make on first emerging after their nest has been moved to a new site.
I had noted that whereas orientation to distant plants was very common, I never saw a bee orientate to a new plant which was put out, close to others in the central area. It looked as if the distance which separated the
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

FORAGING BEHAVIOUR OF BUMBLE-BEES I75
new plant from others which were known, might have an influence on orientation. To test this I put out one or two new plants at varying distances from the central area each day and watched bees that visited them for the first time. Since it seemed likely that only bees which were specializing on Hound's-tongue would orientate, I counted the behaviour of only those bees which visited another plant before or after the new one. By this time a good proportion of the bees were marked, and I discounted bees which I could not identify a second time. The plants were put out at distances of from 3 to 20 feet from the nearest plant in the central group, and at least as far from any distant plant, and I never used the same position more than once. I also tried to make each plant equally conspicuous, and for this reason usually put them out near or along the broad path which skirts the central area (see map, Fig. I, p. I68), where there was no other confusing vegetation.
The results are set out in Table I, and I classified the new plants into those 6 feet or less from the nearest plant known to bees, and those more than 6 feet. This division was chosen because 6 feet is the maximum distance from which I have seen bees fly direct to a new plant, and I thus have no evidence that they can see a plant from more than this distance.
TABLE i
Distance from No. of new No. of bees No. of bees nearest plant plants put out visiting for orientating
Ist time
6' or less Io 2I 21)
More than 6' O I6 3 2)
') Both to a plant 5' from the nearest. 2) 2 bees did not orientate to a plant 7' from the nearest - i did on its second visit.
I bee did not orientate to a plant I2' from the nearest but did on its second visit.
The proportion of bees orientating is significantly greater with distant plants.
(X2 with YATES' correction is 6.493 and P < 0.02.)
Thus it seems that the distance which a new plant is from the central group, which is familiar to the bees, is one factor which causes them to orientate to its position. The conspicuousness of the new plant may well be another factor, but it would be very hard to estimate. As seen from the footnotes to the table, some bees did not orientate until their second visit, so there must be important individual differences as well.
Some bees which make a clear orientation flight after their first visit also show some orientation on subsequent ones, hovering briefly as they
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

I76 A. MANN1\NG
leave. Thus a red-marked pratorum worker on 816/54 found and visited three times a new plant which was i2' from the nearest plant of the central group. My notes run:.
Ist visit. Reacts to plant from about 5', alights, feeds, leaves and circles 2 secs, returns and re-alights, feeds briefly, leaves and circles 3 secs, returns again and alights briefly, leaves finally.
2nd visit. Alights, feeds, leaves and circles i sec, returns and re-alights, feeds, leav-es finally.
3rd visit. Alights, feeds, leaves with a brief, incomplete circling.
It is probable that such repeated orientation flights enable the bee to fix the plant's site with increasing accuracy.
The efficiency of orientation can be judged if one removes any of the distant plants, for bees fly to and hover very accurately over the site for up to IO secs before flying on. Bees will do this even if they have only visited the plant two or three times, and in all of the three cases I have tested, bees returned to the site of a plant which they had visited only once and had orientated upon leaving. Thus on 8,,"6154 a white-marked pratorum worker found a new plant put out 9' from the nearest of the central group: It made a clear orientation flight on leaving, circling and returning three times and then flew off. The plant was then removed, and I watched this bee's behaviour on three successive tripsi to the central area.
On the first trip, Io mins after its visit to the plant, it flew to the ap- proximate site of the plant, circled twice over an area of about 2 square yards, and then returned to the central plants.
On the second trip, 20 mins after the first, it flew to the site, turned immediately and returned.
On the third trip, 50 mins after the second, it flew to the site of the plant, turned immediately and then flew off.
Clearly one visit with orientation can be enough for a bee to fix the site of a plant with fair accuracy:-certainly accurate enough for the bee to fly close enough to see the plant and its memory for the site can persist for three successive visits. After more initial visits to a plant this memory can persist much longer. I have actually counted up to seven successive returns to the site, and many more than this must occur. Two bees were circling briefly on each trip beneath a tree, from which position I had removed a plant two days before.
It is interesting to note that here is another example of the proven 'imprinting' function of an orientation flight to a food source. Most studies of these flights, which have been frequently described for Hymenoptera, but often without critical evidence on their effect, have been concerned with
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

FORAGING BEHAVIOUR OF BUMBLE-BEES I77
orientation to the nest. One of the most complete studies is that of TIN- BERGEN and KRUYT (1938) on the orientation of the hunting-wasp Phi- lanthus to its nest hole. Another comparab'e case of orientation to a feeding place is that of OPFINGER (I93I), who showed that the circling flights made by honeybees when leaving a newly-discovered food dish, served to fix its position for them with respect to their other landmarks.
It was mentioned earlier that bumble-bees do not orientate much to new Hound's-tongue plants which they find in or near the central group, and correspondingly, if such a plant is moved after a bee has visited it two or three times, it shows no sign of remembering the site, and may fly over it en route for another plant without the least check. However, after several days of visiting, bees do show signs of knowing the site to some extent, and this knowledge is probably acquired gradually without any marked orientation flights. Even at this stage, though, it seemed likely that bees visiting in the central area are relying much more upon,direct stimula- tion from the plants than on knowledge of their sites. To ascertain the differences in the stimuli to which bees are responding when visiting central and distant plants, I compared the effect on the number of visits each type of plant received, of first, removing all its flowers, and secondly, removing the plant entirely.
The method was to choose, for each experiment, a pair of plants which bees usually visited in turn. ln the central group these were two adjacent plants, between 3 and 6 feet apart; with the distant plants I chose either one distant plant and the next distant plant to it, or a distant plant and the plant in the central group from which bees usually left when going to visit it. If this latter was the case, then the central plant was always used as the control in the experiments. As far as possible I used different plants in each experiment, but sometimes the same plant was used twice on different days.
Then, in each experiment of the series with central plants, I counted for a control half-hour, the number of visits from all bumble-bees which each of the pair received. Since the chosen plants were next to each other, they received about the same number of visits and bees often flew from one to the other. Next, at the beginning of the so-called 'experimental half-hour', in half the experiments I removed all the flowers from one plant of the pair and left the other as a control. Again the number of visits to the control plant and the flower-less one was counted. In the other half of the experiments one plant of the pair was removed entirely, and in the exper- imental half-hour I counted the visits to the empty site and to the con- trol plant.
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

I 78 A. MANNING
In the other series of experiments with pairs of distant plants, exactly the same procedure was followed. After counting the visits to both for a control half-hour, in half the cases the flowers of one plant were removed, and in the other half, the whole plant.
The relative change between the control and the experimental half-hours, in the number of visits to the experimental plant and the control, gives a measure of the importance of the flowers, form and site of the plant to bees, and comparison of the two series of experiments will reveal any dif- ferences between their reactions to central and distant plants.
Tables 2 and 3 set out the results of these experiments with four pairs of central plants, the plants of each pair being les.s than 6 feet apart.
TABLE 2
Central plants Control '/2-hour Experinmental '2-hour
Plants A & B with flowers Plant A with flowers Plant B without flowers
No. of visits to No. of visits to
lExp. A B A B I 12 I5 I2 13 2 i8 I3 20 II 3 14 I9 12 I5
4 I2 II I2 IO
Total 56 58 56 49
There is no significant reduction in visits to the flowerless plants.
TABLE 3
Central plants Control 'A-hour Experimental '2-hour
Plants A & B with flowers Plant A with flowers Plant B removed entirely
No. of visits to No. of visits to
Exp. A B A B
I II I4 12 6 2 9 i6 II 10 3 13 20 i8 8 4 12 IO I5 6
Total 45 6o 56 30
There is a significant reduction in the number of visits to the empty site as compared with the actual plant.
X2 with YATES' correction - 8.522 and P < O.0I
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

FORAGING BEHAVIOUR OF BUMBLE-BEES
These results indicate that the flowers of central plants are not necessary to attract bees, because of their conditioning to form, and removing them has no effect upon the number of visits. However, the presence of the plant itself is important, for the bees have only a limited knowledge of the site and are relying on direct stimuli from the plant's form to a large extent. The site of a central plant gets only half the visits that the plant does, and on some occasions I was able to note the behaviour of individual bees during the experiments. It seemed that this drop in visits was partly due to some bees not reacting to the site at all, either because they did not remember it or because they were attracted away by the sight of other
plants close by. Also some bees which did react to the site at the beginning of the half-hour, ceased to do so towards the end.
Tables 4 and 5 set out the results of the same experiments with distant
plants, and in order to get* the biggest contrast I worked with plants which were at least 12 feet from the next nearest.
TABLE 4 Distant plants
Control '/2-hour Plants A & B with flowers
No. of visits to
Exp. A B I 7 7
Experimental V2-hour Plant A with flowers
Plant B without flowers
No. of visits to A B 8 9
2 8 io 5 3 ii Io 13 4 9 9 8
Total 35 36 34 There is no significant reduction in visits to the flowerless plants.
TABLE 5 Distant plants
6 IO
9
34
Control ¼2-hour
Plants A & B with flowers Experimental ¼f-hour
Plant A with flowers Plant B removed entirely
No. of visits to No. of visits to
Exp. A B A B 8 8 8 9
2 1 12 13 II
3 9 13 8 Io 4 15 9 II 6
Total 43 42 40 36 There is no significant reduction in visits to the empty site.
179
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

I80 A. MANNING
From these results it is clear that with distant plants the bees have a very detailed knowledge of the position of each plant, and that no direct stimuli at all are nee-ded from the plant to guide the bees to its exact site.
Summing up the results of these experiments, we may say that there are some differences in the stimuli to which bees are responding when visiting a compact group of Hound's-tongue plants, all within 6 feet of the next plant, and when they are visiting the scattered plants around the edge of this group, and all plaints are io-,i2 feet from the next. In the former case they are directed to a great extent by the form of the plants, but have some knowledge of their positions, and this results in their wavering flight and frequent reactions to other plants which have some of the form characters of Hound's-tongue. When visiting the distant plants they rely entirely on their memory of the position of each plant, and this is sufficient to bring them to its exact site without any stimliuli from the plant itself. This again results in their direct flight between these distant plants and in their reacting only rarely to other plants when flying to them.
Of course, for these experiments I deliberately chose the two extremes- plants that were very close together and those over I2 feet apart-and the above conditions probably only apply in their entirety to such extremes. I imagine all intermediate conditions hold for plants at medium distances, but as plants become more separated, bees will probably rely less on direct stimulation and more on memory of position. The earlier experiments on orientation flights suggest that some sort of transition region occurs with plants between about 5 and 8 feet apart, and this may be linked with the fact that this is near to the maximum distance from which a bee can see a plant.
Some confirmation of the above conclusions comes from the amount of time which bees spent searching at the empty site or the flowerless plant in the experiments. Searching behaviour will be dealt with in more detail in the next section, but the results from these experiments seem most ap- propriate here. In both the series of experiments just described, I counted the searching times of each bee that visited the experimental plant or the site. My method was to count mentally, since the use of a stop watch would be very difficult when bees were visiting frequently, and this scale was accurate to about one second, which was quite enough for the purpose. The figures are set out below. Each row represents a single experiment whose number corresponds to those given earlier; the individual numbers represent the searching time, to the nearest second, of each bee as it visited the flower- less plant or the site, in the order which they came. The figure o means that a bee just reacted to the plant or site but did not search it or hover at all, flying off at once.
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

FORAGING BEHAVIOUR OF BUMBLE-BEES i8i
TABLE 6
Searching tiines at flowerless plants in the central group Exp. i: 3 4 2 I 2 I 0 2 I 2 0 2 I
Exp. 2: 2 I I 0 I 0 2 I I 5 I
Exp. 3: 4 2 I 3 I 2 2 0 2 I I 3 5 4 2 Exp. 4: 2 I I 3 2 6 I 5 2 I
Mean: I.8979 Standard error of mean: 0.2004
Standard deziation: 1.403
TABLE 7
Searching tinmes at the sites of plants int the cenitral group
Exp. I: 2 I I I I I Exp. 2: 0 0 I 0 0 I 2 I 2 I
Exp. 3: I 0 0 I 0 3 0 I
Exp. 4: 0 I 0 0 I 0
Mean: 0.767 S.E. of mnean: 0.I388 S.D.: 0.076
TABLE 8
Searchinig times at distant flowerless plants
Exp. i: 5 2 2 3 0 4 3 I I Exp. 2: I 3 0 7 4 I
ExP. 3: I 4 6 I 2 3 0 4 2 3 Exp. 4: 3 I I 3 4 I 3 I 2
Mean: 2.412 S.E. of mwean: 0.2856 S.D.: i.665
TABLE 9
Searching times at the sites of distant plants
Exp. i: 2 2 10 3 2 4 4 6 I Exp. 2: 2 3 4 I 2 2 5 5 I 0 0
Exp. 3: 6 3 4 7 I 5 2 6 6 2 Exp. 4: 5 5 0 3 3 I
M/lean: 3.277 S.E. of mean: 0.3695 S.D.: 2.2I9
Statistical analysis shows that with the central plants, the flowerless plants are searched for significantly longer than the site alone (P < o.Ooi). There is very little hovering over the site of a removed central plant; most of the bees simply fly to the site anrd turn away immediately, a few search briefly in vague circles which are in contrast to the close accuracy, often within a foot or so, of bees hovering at the site of a removed distant plant. This difference in the searching of the plant and the site confirms that bees visiting within the central area are relying primarily upon direct stimulation from the plants. However, when one considers the searching of distant plants, the actual plant is searched no more than the site, indeed the average
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

I 82 A. MANNING
time spent searching the site is greater than that at the flowerless plant, although on these figures not significantly so. (The figures also show that the sites of distant plants are searched significantly longer than those of central plants, P < o.ooi.) Again this confirms the importance attached to the site by bees visiting distant plants, and the fact that the addition of direct stimuli has no enhancing effect. It could be noted that individual bees returned to and searched at the empty sites of, or flowerless, distant plants on each successive trip during the half-hour. There was no detectable falling-off in searching time over this period and it clearly takes much longer for them to become negatively conditioned.
All the evi,dence, then, points to the conclusion that bees are reacting differently when visiting plants which are close to one another, so that from any plant a bee can probably see several others, than when visiting more scattered distant plants where they probably cannot see the next nearest plant when they leave the one they have been visiting. In the former case the bees rely primarily on visual stimuli from the form of plants to direct their flight. Such knowledge of the sites of individual plants which they have, is acquired gradually over the course of many visits, and is probably restricted to a vague sense of the distance to be flown and the direction from one plant to the next. When visiting distant plants, on the other hand, bees do not use direct stimuli from the plant at all, but rely on a vrery exact knowledge of the site of each plant, which is acquired very quickly in the course of one or two marked orientation flights after they find the plant for the first time.
LEARNING THE FORM OF PLANTS
It has been mentioned before how bees visiting Hound's-tongue, as a result of their becoming conditioned to the general form of the plant, will fly to flowerless Hound's-tongue and other plants and 'search' them for flowers. This consists of flying to the base of the stem and hovering up and down at about I-2" from it, often reacting to the bases of the leaves, and thus the bees appear to be treating such plants just as if they were Hound's-tongue, and are searching for flowers in the leaf axils. Now a wide range of different plants growing near Hound's-tongue are searched. I have seen all of the following plants searched 'spontaneously' by bees; that is they have flown to the plants growing naturally without ever having been conditioned to their site from previously visiting Hound's-tongue there.
Rose-bay Willowherb (Chamaenerion angustifolium) Hairy Willowherb (Epilobiumn hirsutum)
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

FORAGING BEHAVIOUR OF BUMBLE-BEES 183
Figwort (Scrophularia nodosa) Foxglove (Digitalis purpurea) Ragwort (Senecio jacobea) Burdock (Arctiumt lappa) Spear Thistle (Cirsium vulgare) Dog's Mercury (Mercurialis perennis) Stinging Nettle (Urtica dioica) Bramble ('Rubus fruticosus') Ash (Fraxinus excelsior) Privet (Ligustrum vulgare) Elder (Sambucus nigra)
A consideration of features which all these plants share and those in which they differ, helps to give an idea of the stimuli which are important to release searching by bees. Features they share are:
I. They all consist of an upright stem bearing leaves along most of its length.
2. They are all fairly large plants-none less than 6" (I5 cm) high. 3. None have flowers at the same time as Hound's-tongue.
Features in which they are idissimilar include:
I. Their leaves are shaped very differently, varying from quite entire, as in Willowherb or Figwort, to much divided as in Ragwort and Thistle.
2. They are many different shades of green. 3. None has a scent resembling Hound's-tongue; most have very little
scent to the human nose, but a few such as Ragwort and Elder have strong and eharacteristic scents.
From the fact that plants which have strong scents quite different from that of Hound's-tongue are searched, it seems that visual features are prob- ably most important in attracting bees to fly to and search flowerless plants. I investigated the effect on searching of grossly altering the scent of flowerless Hound's-tongue plants. Three plants in the central group, adjacent to one another, were chosen, and 1 counted the number of visits each had during a control hour. Then for the experimental period, the flowers of two plants were removed and one of these flowerless plants strongly scented with lavender, which was renewed after half an hour; the third plant was left with flowers as a control. I then counted during the next hour the number of visits each had and the length for which eaeh flowerless plant was searched. The results of 5 such experiments are set out below in the same style as for the previous exepriments, and they show what effect the wrong scent has on, first, the primary reactions to the plant, and secondly, on the length of time that it is searched.
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

I84 A. MANNING
TABLE Io
The eff ect of scent on primnary reactions
Control hour Experimental hour
Plants A, B & C with flowers Plant A with flowers Plants B & C without flowers Plant C given lavender scent
No. of visits to No. of visits to Exp. A B C A B C
I 15 20 I7 20 I8 I5
2 26 24 23 20 17 2I
3 i6 21 30 2I 23 24
4 I0 2I I4 II I7 I4
5 7 II 10 10 I4 20
Total 74 97 94 82 89 94
There is no significant reduction in visits to either normal or scented flowerless plants
TABLE i i
The effect of scent on searching times
Searching times at normal flowerless plants
Exp. I. I 2 I 4 I I 0 3 I 4 I 3 I I 0
4 1 I 2. 0 I I 0 0 I 0 I 2 3 6 3 3 I 0
I 0
3. I 0 I 0 I 0 0 I I 2 I I 0 I I
I 2 I I 0 I I 0
4. 6 2 I 0 I I 3 I I 0 0 I 2 3 0
I 0
5. 2 I I 2 I 2 I 0 I 0 I I 0 2
Mean: 1.225 S.E.: 0. I298 S.D.: i.224
TABLE I2
The effect of scent on searching times
Searching times at flowerless plants scented with lavender
Exp. T. 0 I I I 2 I 4 I 0 0 0 I 0 I I
2. 2 I I 3 0 2 I I 0 4 4 2 I 4 2
I 0 I 0 0 0
3. 1 1 I I 3 2 0 I 2 I 0 I I 0 0
0 0 0 0 3 I 0 2 0
4. 0 2 2 I I 3 3 0 I 0 3 I 2 I 5. I 0 I 4 I 2 I I 0 0 0 0 I 0 I
3 I 2 I 0
Mean: L.II7 S.E.: O.II4 S.D.: i.io8
A statistical analysis shows that there is no significant difference be- tween normal and lavender-scented plants in either case, so it appears that
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

FORAGING BEHAVIOUR OF BUMBLE-BEES 185
altered scent has no effect on primary reactions or searching time. It is most interesting to compare this result with that of altering the scent of a plant with flowers. Almost invariably if a bee reacts to a normal plant with flowers it alights, and thus it is very easy to see if adding some artificial scent to the flowers has any effect. I counted the number of reactions which bees made to a plant in half an hour and how many times they alighted. I then scented both leaves and flowers artificially, in this case with rose. In the following half-hour I again counted the number of reactions and alightings. In both cases, as alighting, I count only an actual probing of at least one flower, and not a mere touching down and then veering away. Table I3 sets out the results of four such experiments.
TABLE I3
Normal plant Rose-scented plant Exp. Reactions Alights Reactions Alights
I I2 I2 13 7 2 15 I5 I9 9 3 I8 i8 15 IO 4 i6 I6 12 4
Total 6i 6i 59 30
There is a significant reduction in the proportion of bees alighting on the scented plants
X2 with YATES' correction is 4.92 and P < o.o5
The effect of the wrong scent in reducing the number of alightings is not very large but it is there-all the experiments go the same way. The effect is certainly very much less than in a very similar experiment I did with Dandelions, giving some flowers lavender or rose scent. This may be due to a combination of reasons. One is probably that the actual flowers of Hound's-tongue because they hang down, are very difficult to scent, and in the experiments quoted above I had to be content with putting the rose water on to the calyces; with the Dandelions I could put scent on the actual flower and its effect was probably more conspicuous as a result. Secondly, of course, scent may be relatively less important as a means of identifying the flowers to bees visiting Hound's-tongue, or it may be suf- ficient for them to be able to detect some Hound's-tongue scent through the rose.
In any case scent has no effect unless associated with the flowers. This bears out in an interesting way the conclusion that during the course of the chain reaction of a bee to a flower, certain stimuli, and particularly scent, may not be responded to, although perceived, early in the chain, but
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

i86 A. MANNING
become important in the final stages as the bee alights on the flower. With Hound's-tongue, after a bee's first few visits, the primary visual reaction from a distance is directed not by the flowers but by the general form of the plant, and presumably the bee *does not 'pass' into the next stage, when it responds to scent stimuli, until it can perceive the flowers. Thus searching of a flowerless plant will not depend upon scent stimuli, for in the abscence of flowers a bee will still be reacting to primarily visual stimuli.
The possession of leaves is essential to release searching. If a plant, stripped of all its leaves so that only the stem remains, is put out on the site of a Hound's-tongue plant which bees have been visiting, it gets no more reactions than would the site alone, and is not searched at all. Thus it can provide none of the stimuli for either a primary reaction or searching. All of the conspicuousness of a plant lies in the projecting leaves, but the exact form of the leaves is not important. Thus a Foxglove plant without flowers is visited and searched just as much Hound's-tongue. Table I4 gives the results of some experiments when a flowerless Foxglove plant is put out on the site of a Hound's-tongue plant in the central group. Another Hound's-tongue plant next to it is used as control.
TABLE 14
Control i2-hour Experimental '12-hour A & B both Hound's-tongue plants with Foxglove plant without flowers
flowers substituted for B
No. of visits to No. of visits to Exp. A B A B
I 9 I2 8 10
2 9 IO 8 IO 3 8 7 IO 9 4 IO 10 8 9
Total 36 39 34 38
There is no significant reduction in visits to the flowerless Foxglove
Clearly bees treat the flowerless Foxglove plant just as if it were Hound's- tongue, and there is no reduction in the number of visits.
Table i 5 sets out the searching times of the Foxglove in the above experiments, and for comparative figures of the searching of flowerless Hound's-tongue plants I have used those given in Table ii, p. 184, which were obtained in a completely comparable way-by setting up a flowerless plant on the site of a plant with flowers, which had been visited by bees.
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

FORAGING BEHAVIOUR OF BUMBLE-BEES I87
TABL,E I5
Searching times at a Foxglove plant without flowers
Exp. I: I I I 2 0 0 2 0 3 I
2: 0 2 I 2 0 2 I 2 I 0
3: 4 2 3 I 2 0 I 0 4 4: 0 2 I I 0 I I 2 I
Mean: I .2432 S.E.: 0. I 768 S.D.: I .075
Analysis shows that there is no significant difference between these figures and those of Table I I, and thus presumably a flowerless Foxglove plant provides very similar stimuli for searching to those of Hound's-tongue itself. It was noticeable that bees only searched among the leaves of the Foxglove where it most resembled Hound's-tongue and did not react to the slender spike where there were no leaves. Even so, to human eyes the colour, shape and number of leaves are quite different in the two plants, but such factors can either not be perceived or have less significance for a bee. I got substantially the same results with bees searching a Figwort plant set up on the site of a Hound's-tongue, but I have not sufficient figures to make them worth quoting.
The form requirements for searching, therefore, appear to be very general, and detail is not necessary, but a plant which gives some 'correct' but at the same time some absolutely 'wrong' stimuli seems to have some inhibitory effect upon searching. I found indications of this when I put out a Foxglove plant with several open flowers and many of the buds already fully coloured. Tables i6 and 17 set out results of four such
experiments.
TABLE i6
Control '/2-hour Experimental '/2-hour
A & B both Hound's-tongue plants with Foxglove plant with flowers flowers substituted for B
No. of visits to No. of visits to Exp. A B A B
I 12 9 I2 6 2 6 10 8 IO 3 15 12 i8 8 4 13 15 8 10
Total 46 46 46 34 Behaviour TX 13
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

I88 A. MANNING
TABLE 17
Searching timies at a Foxglove plant with flowers Exp. I: 0 I I I 0 0
2: I I 2 0 0 I I I I 0
3: I I 0 2 0 0 I 0
4: 0 0 0 I 0 0 0 0 I 0
Mean: 0.5295 S.E. of mean: 0.I039 S.D.: o.6055
The figures of Table i6 do not show any, reduction in the number of primary reactions to the Foxglove with flowers, (x2 0,9762), though some experiments do show quite a large drop. The influence of the actual site, of which bees have some knowledge, is of course there, but the stimuli releasing a primary reaction may be less critical than those for searching. Certainly the Foxglove with flowers is searched much less than either one without flowers (compared with Tabel I5, P < 0.OI), or a flowerless Hound's-tongue plant (compared with Table I I, P < 0.OI). All the searching that the flowering Foxgloves had was at the leafy part of the stem, and on coming close to the flowers the bees flew off; it was very common for them to veer away when they were a few inches from a plant, without searching it at all. The only difference between such a plant and the Foxgloves used in the other experiments, which were searched just as much as Hound's-tongue, was the presence of the large conspicuous flowers, and it seems that the sight of something so obviously 'wrong' inhibits searching.
Searching is probably influenced also by other factors than the direct stimuli from the plant, which have been considered so far. The most obvious is the bees' knowledge of the site of the plant they are searching. All of the experiments in this section have been with plants set up on the site of Hound's-tongue plants in the central area which bees had been visiting. The previous experiments (see 'Tables 6 & 7, pp. i8i), show indications that bees search longer at a distant flowerless plant than at one in the central group, and we know that they have a more exact knowledge of the distant plant's site. I am also convinced that bees search longer at flowerless plants on the site of plants in the central group, than they do at those which are not on the site of plants that they have been visiting, so that their vague knowledge of the site does influence their searching times. Regrettably I have no exact figures on the searching times to flowerless plants which were not on the site of central plants, but on looking through my field notes it is clear that the commonest cases are where bees fly to such plants and search for about i second. 1I once recorded 5 seconds' searching, but this was quite exceptional and the average would, I estimate, be significantly lower than those for plants which were on the former site of others -which bees had visited, given in Table 6.
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

FORAGING BEHAVIOUR OF BUMBLE-BEES I8
The interaction between direct stimuli from the plant and from the site and its effect on searching times, is different for distant plants. Here, as seen from Tables 8 & 9, the knowledge of the site and the relative impor- tance attached to it is much greater. The site alone is searched just as much as a flowerless plant which is put there, and the only effect of the plant is to concentrate a bee's searching towards itself, rather than to the square yard or so which a bee will cover when searching at the site alone. Thus direct stimuli from it do attract bees, but do not lengthen their searching time, and I have sometimes seen that, even when a flowerless plant is pre- sent, bees will not confine their searching to it, but may briefly search other suitable plants which are close by. In the same way, when searching at the site alone, they will react to other plants growing near it.
THE ROLE OF STIMULI FROM THE FLOWERS
In this section I will briefly describe a few incomplete experiments to investigate the stimuli from the flowers of Hound's-tongue-and this term only includes the actual corolla-to which bees are reacting. I tried putting out artificial flowers of various colours, which consisted of small paper squares-at about /4" across-stuck on to wire frames so that about 9-I2'
flowers were mounted close together about I'6"-2' from the ground. The colours used were blue, yellow, green, red and purple-this last most resembling the colour of real flowers to the human eye.
Bees which have been visiting Hound's-tongue for several days and show every sign of, 'specializing' on it, and which therefore are presumably as far conditioned to its flower-colour as any, will react to all of these models and hover briefly in front of them. I have not enough figures to be able to discover any definite preference, but the purple seems to be the most attractive. This is borne out by the fact that if paper models of various colours and real flowers are laid on the ground, the latter and the purple models are the only ones I have seen visited.
This last fact, and some others, indicate that the height of the flowers may be influencing the reactions of bees. Thus while Hound's-tongue is flowering, several other plants such as Bugle, Ground Ivy, Buttercup and Herb Bennet are flowering also beneath it. Bees specializing on Hound's- tongue very rarely react to any of these flowers, but if they are picked and bunches of them suspended about 2' high on sticks, bees do react to them quite often, just as they react to the paper models. Now this may simply be a matter of the increased conspicuousness of the flowers when held up off the ground, but bees react much more often to Hound's-tongue flo- wers laid on the ground, which are very inconspicuous, than they do to the
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

I90 A. MANNING
other flowers when growing naturally a few inches above the ground. It is more likely that bees become conditioned to feeding at a certain height when visiting Hound's-tongue (though this height cannot be at all precise as one plant bears flowers from perhaps 6"-i'6"', I5-45 cm), and that having the correct height may cause flowers of the wrong colours to attract reactions. ,Conversely, the correct colour may make up for incorrect height in the case of Hound's-tongue flowers laid on the ground; in such a position flowers of wrong colours get no reactions.
Clearly much more experiment on the role of flower-colour is needed; but, perhaps correlated with the relative inconspicuousness of the petals, bees specializing on Hound's-tongue seem to be much less 'fixed' on a particular colour than are bees specializing on some other flowers, e.g. Foxglove, to be described next.
It may be noted here that, in spite of their conditioning to the general form of a Hound's-tongue plant, arid the unimportance of the flowers in primary reactions which has been demonstrated, if flowers or models are put out conspicuously, they do attract reactions. This makes it the more likely that one of the reasons for form conditioning is because normally bees are unable to see the flowers until they are close to a plant. The fact that they do not use the flowers as a guide is well illustrated at the end of the flowering season, when some of the larger plants have small clusters of flowers right at the base of their stems. Bees seem to get used to finding flowers there, and I often saw them fly to the bottom of a plant which had no such low flowers. They then have to work their way up the stem until they reach the lowest flowers which the plant has.
From this study, three features of the foraging behaviour of the bumble- bees on Hound's-tongue, stand out as being, perhaps, rather unusual or strangely accentuated, as compared with what we know of their basic reac- tions to flowers. These are:
I. Learning the position of individual plants. 2. 'Exploration flights'. 3. Conditioning to form and searching of flowerless plants and other
plants resembling Hound's-tongue. In these three aspects, bees visiting Hound's-tongue appear to be somewhat
exceptional. The most probable cause of this is that while it is a very attractive plant to bees, on account of its good nectar supply, its flowers are very small and inconspicuous. In a mature plant the flower petals are almost invisible save from below, since the flowers hang down and the calyx hides the petals from the side. Thus a bee cannot rely on direct sight of the flowers to guide it from any distance, and consequently it becomes
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

FORAGING BEHAVIOUR OF BUMBLE-BEES I9I
conditioned to the plant's general form which can guide it from much further. A bee's form discrimination is rather crude, and as a result it reacts to many other plants besides Hound's-tongue, but this is not very disadvantageous since it only searches them briefly and then passes on. Orientation to the site of individual distant plants is necessary, for a bee cannot rely on any direct stimuli to bring it to the plant again, and apparently it is 'worth while' for a bee to include even the most isolated plants within its foraging area.
One of the most interesting ways of checking this explanation is to compare the foraging behaviour of bumble-bees on another plant which has very conspicuous flowers, but where each plant is single and distinct, as is Hound's-tongue. The obvious plant to choose was the Foxglove, which is a very popular plant with bumble-bees.
I shall not describe the whole cycle of foraging behaviour on Foxglove in as much detail as for Hound's-tongue, but will pay especial attention to points where a direct comparison can be made between the two.
THE FORAGING BEHAVIOUR OF BUMBLE-BEES ON FOXGLOVE
The Foxglove-Digitalis purpurea L.,-is a very familiar plant and a short description of it will suffice. It is a member of the Scrophulariaceae and is a biennial, flowering in its second year. The stem grows up to 4' high, of which the lower half bears the dozen or so large, dark green leaves, and the upper part forms the long flower spike, which may bear up to 8o flo- wers and which elongates as the flowers mature from the bottom upwards (see P1. XI, Fig. 2). The flowers have the corolla fused into a broad tube which, when fully open, is about 2" long with a short upper and a longer lower lip. They are reddish-purple in colour with a 'honey-guide' pattern of dark spots inside the lower lip, and have no detectable smell to the human nose.
Foxglove is usually described as a typical 'bumble-bee flower', for only long-tongued bees can reach the nectar, which is secreted into the narrow, extreme base of the corolla tube. Where I worked, Foxglove does not grow naturally, and after I put out some plants, the very long-tongued ruderatus and hortorum were the commonest visitors, but agrorum was fairly com- mon also. In the Forest of Dean, where it grows wild in abundance, I saw terrestris, lucorum and lapidarius visiting it frequently, as well as the above species.
When visiting Foxglove, bumble-bees fly to one of the bottom flowers and, clinging to its lower lip, climb inside the tube; in this they are helped by the long white hairs which grow at the end of the lip. The bees push
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

I92 A. MANNING
their way as far up as their size will allow, and their tongue must reach t-he rest of the way to the nectaries. It is usual for a bee to enter a flower with its tongue already extended-presumably it would have difficulty in extending it within the narrow tube. It may stay anything up to io seconds inside one flower, and then enters another a little further up the spike. It will usually visit most of the open flowers on a plant before passing on, but if, as on very large plants, there is a double row of flowers, it often misses some as it moves up the spike. Bees often force their way into flowers which are not fully open, and in which there is no detectable nectar, and they also react to unopened buds which are fully coloured on the outside.
I imported 25 plants in pots in both seasons, and set them up in much the same arrangement as used for Hound's-tongue. They formed a central group of about I 5 plants, all within 8' (2.4 m) of the next nearest, and about IO more distant plants, some of which were about 30' (9 m) from the central group and each other.
In marked contrast to the condition with Hound's-tongue, bees began visiting the flowers as soon as buds became large, about I " (2.5 cm) long, and fully coloured, bef ore any of them were open. Bees of several species would fly to the buds as they passed and hover for a few seconds. Sometimes they reacted to the junction of the calyx and the corolla, and a few bees even alighted and probed in this crevice. It is possible that they were attracted there by some increased scent stimulus, and it may be that with old flowers where the corolla is loose, bees could reach the nectaries by this way. However, in this case no nectar could be present, and it is more likely that they were attracted by the colour margin which the green calyx makes with the corolla, but scent may be strongest here also.
Bees continue to react to the buds, and as soon as the tube is sufficiently open, force their way into the flowers before any of the stamens are ripe and, so far as I can tell, before any nectar has been secreted. As more flowers on different plants open, some bees begin to spend considerable time moving from plant to plant, reacting to any which have coloured buds. However, I do not think that any bees were entirely constant to Foxglove until all the plants were flowering, for there were not sufficient flowers to satisfy them until then. When all the plants had several open flowers, some bees were virtually constant but occasionally I saw them visit Red Campion (Melawdrium rubrunt) wvhich was growing nearby.
It was noticeable how quickly bees found all of the plants, although- some were partially hidden in thick bracken. On a few occasions I saw 'bees make wavering flights, ranging back and forth, on leaving a plant, and they did not react to other conspicuous flowers which they passed close to. I assume
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

FORAGING BEHAVIOUR OF BUMBLE-BEES
that these were 'exploration flights' of a kind, but I never actually saw a bee discover a new plant on such a flight. New plants which I put out were
rapidly discovered, and several times bees flew direct to such plants from at least I2', SO it is likely that bees can see a Foxglove plant with flowers from twice the distance that they can see a Hound's-tongue plant.
Orientation flights on leaving newly-discovered plants were rare and
irregular, and I never saw any orientation to new plants in the central
group. 7 times I put out plants in new positions 15' or more from the nearest plant that bees were visiting, and saw I6 bees visit them for the first time. Of these only 3 showed any sign of orientating as they left. To a plant 20' distant one bee-a ruderatus male-gave a very full orientation, just as do bees to Hound's-tongue; the other two simply made a brief circle as they left, one from a plant also 20' distant, the other from one I5' away. No bees ever returned to the site of these new plants when they were removed, but unfortunately none of the three bees which did show signs of orientation were seen again the same day. These results are quite incon- clusive, but experiments show that, however obtained, bees do have con- siderable knowledge of the position of each distant plant they visit.
The experiments are exactly parallel to those with Hound's-tongue described on pages 177 to 182. Two adjacent plants in the central group were chosen, and the number of visits each received during a control half-hour was counted. Then in half the experiments the flowers of one plant were removed, while in the other half one of the plants was removed entirely; the number of visits to each was again recorded for half an hour, together with the times for which bees hovered at the flowerless plant, or the site of the plant that was removed. A similar series of experiments was done with two distant plants, more than 20' apart and distant from the central group. The results are set out in Tables 18-25, using just the same notation as in Tables 2-9 for the corresponding Hound's-tongue experiments.
TABLE I8
Central plants Control /2-hour Experimental I2-hour
Plants A & B with flowers Plant A with flowers Plant B without flowers
No. of visits to No. of visits to Exp. A B A B
I 21 I8 20 6 2 II i6 7 3 3 22 26 25 7 4 20 20 17 3
Total 74 80 69 I9 There is a significant reduction in visits to the flowerless plant.
x2 (with YATES' correction) = 20.12 and P < o.ooI.
i93
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

A. MANNING
TABLE I9
Central plants Control /2-hour
Plants A & B with flowers
No. of visits to
Exp. I 2
3 Total
A B
21
25
22
23 29 I8
68 70
Experimental Vz-hour Plant A with flowers
Plant B removed entirely No. of visits to
A B
18 3 30 4 21 4
69 II
There is a significant reduction in visits to the empty site.
x2 with YATES' correction = 28.08 and P < o.ooI.
TABLE 20
Distant plants Control ¼I-hour
Plants A & B with flowers
No. of visits to
Exp. A B
I 7 8 2 8 10 3
Total
Experimental /2-hour Plant A with flowers
Plant B without flowers
No. of visits to A B
6 9
IO 9 IO
24 28
5 6
II
25 22
There is no significant reduction in visits to the flowerless plant x2 with YATES' correction = 0.489
TABLE 2I
Distant plants
Control ¼2-hour Plants A & B with flowers
No. of visits to A B
7 7 I4 14 10 12
Total 3I 33
Experimental ¼2-hour Plant A with flowers
Plant B removed entirely No. of visits to
A B
7 6 10 9 II 10
28 25
There is no significant reduction in visits to the empty site.
X2 with YATES' correction - 0.23
I94
Exp.
I 2
3
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

FORAGING BEHAVIOUR OF BUMBLE-BEES 195
TABLE 22 TABLE 23
Searching tintes at flowerless Searching times at plants in the central group the site of plants in
the central group Exp. I: I 0 I 0 0 0 Exp. i: 0 0 0
2: 0 0 1 2: 1 I I 0
3 I 0 0 1 I 0 0 3 I 0 I 0 4 I D 1
TABLE 24
Searching times at flowerless distant plants Exp. i: 0 0 0 I 0
2: 0 I I 0 0 I
3: 0 I 0 I I I I 0 0 I 0
Mean: 0.45 secs
TABLE 25
Searching times at the site of distant plants Exp. i: 1 2 0 I I 0
2: 0 0 0 0 2 I I I I
3: I 0 I I I I I I I I
Mean: o.8o secs
Thus if a plant in the central group is removed there are very few visits as did the actual plant. It is clear that, just as with Hound's-tongue, bees as did the actual plant. It is clear that, just as with Hound's-tongue, bees have very little knowledge of the sites of central plants, and they are relying upon direct stimuli from the plant to guide them, but they have considerable knowledge of the position of each distant plant, and direct stimuli are quite unnecessary. The great difference in their behaviour on Foxglove as compared with Hound's-tongue is that, while with the latter a plant in the central group with its flowers removed receives as many visits as it does with flowers, a Foxglove plant so treated has hardly any visits at all, no more than does the site alone. Clearly then, bees visiting Foxglove do not become conditioned to the general form of the plant, as do those visiting Hound's-tongue. This conclusion is confirmed by the very short time for which bees which do react to flowerless Foxglove plants, actually search them. In most cases a bee simply flies to the spike and then veers off; occasionally it hovers along the length of the spike for a second, but never more. I never saw a single case of 'spontaneous' searching while bees were visiting Foxglove, -i.e. bees never flew to and searched other flowerless plants as they do when visiting Hound's-tongue, and no Foxglove plant was ever
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

I96 A. MANNING
visited until it had large purple buds. As a consequence, their flight between plants in the central group was very rapid and direct, in contrast to the wavering flight and frequent reactions to other plants, shown by bees visiting a compact group of Hound's-tongue plants.
Within the central group, then, where all the plants are within 6' of the next nearest, bees visiting Foxglove are relying almost entirely upon direct stimuli. Their knowledge of the sites of central plants seems to be even less than that of Hound's-tongue bees, for if one compares the figures in Tables 3 and ig, which give the reduction in visits caused by removing the plants for each type, there is a significantly greater reduction with Foxglove than with Hound's-tongue (x2 with YATES' correction _ 8.o6g, P < o-oi). However, it is obvious that, unlike Hlound's-tongue bees which rely largely upon stimuli from the leafy part of the plant, Foxglove bees orientate entirely by the flowers. As mentioned before, bees can see a Foxglove plant with flowers from at least I2', and obviously in all but the thickest vegetation a spike of flowers will be clearly visible to them over distances of 6' or less, and no other visual guide is necessary. Beyond the range from which they can see the spike of a plant, probably about I5', they, like Hound's-tongue bees, rely on their knowledge of the site of each plant to guide them to it. I got the impression that the Foxglove bees were not as precise in their reactions to the site of distant plants as were Hound's-tongue bees. They hovered over a rather wider area than did the latter, and again this may be linked with the fact that they can normally see the Foxglove plant from so much further than they could the Hound's-tongue. A rough orientation will be quite sufficient to bring them within sight of the plant and enable them to be guided by the flowers.
Although the figures given in Table 25 are too slight for proper analysis, it looks as if bees do not search so long at the site of Foxglove as that of Hound's-tongue (see Table 9, p. i8i). This may also be linked with a poorer knowledge of the site. Alternatively it may be that Foxglove bees are 'used' to reacting directly to the flowers from several feet, and in the absence of such direct stimuli, searching is short because the situation is so obviously 'wrong' to them. Hound's-tongue bees, on the other hand, are not so depen- dent on direct stimuli for finding distant plants since these may not always be available, save from very close quarters, and consequently their absence causes less idisturbance and more prolonged searching.
As when discussing the Hound's-tongue results, I must again stress that with the two categories of 'central' and 'distant' plants, I have picked on the extremes. At intermediate distances, perhaps between IO and I5' with Fox-
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

FORAGING BEHAVIOUR OF BUMBLE-BEES 197
glove, depending upon conspicuousness, knowledge of site may be combined with some direct attraction from the flowers.
The final point of contrast between the behaviour of bees on the two plants concerns their colour conditioning. On page I89 it is described how Hound's-tongue bees will react to several colours if these are displayed at the correct height for the flowers, but that if the height is wrong, only real flowers and what seems to be the best artificial colour are reacted to. I repeated similar tests with Foxglove, putting out various colour models, both on flower spikes and on the ground, and the only colour to which bees reacted was the same purple, already described on p. I89, which is the closest match to Foxglove flowers for the human eye. No other colour, even if displayed at the correct height on the spikes of a deflowered plant, ever attracted bees. It was noticeable that the bees paid little attention to height, for they would react to and enter single flowers laid on he ground, and often did so to the fallen flowers around the base of a plant. However, on the few occasions when bees reacted to flowerless plants on the sites of plants they had been visiting, they only reacted to the spike where the flowers would normally be borne, and did search among the leaves.
We may briefly summarize a comparison between the foraging behaviour of bumble-bees visiting Hound's-tongue and Foxglove as follows.
When visiting plants which are close to one another and within sight, bees on both species are guided by direct stimuli from the plants. With Hound's-tongue bees, these stimuli are chiefly from the general form of the plants, and are independent of the actual flowers, for such bees become conditioned to form very rapidly after their first few visits. This is not the case with Foxglove bees, for with them stimuli from the flowers are quite essential and there i-s no conditioning to form. iCorrespondingly, Fox- glove bees never react to flowerless plants and other species which resemble them in form, as do Hound's-tongue bees.
Bees foraging on both plants are largely guided to more distant plants in their foraging area, beyond visual range, by their knowledge of the site of each individual plant. Hound's-tongue bees appear to have the more precise orientation to distant plants, and this may be correlated with the more common occurrence of clear orientation flights on first finding new distant plants. These flights do occur sometimes with Foxglove bees, but are certain- ly more irregular. Ranging 'exploration flights' are also commoner and more extensive with Hound's-tongue bees. Finally, Foxglove bees appear to be more conditioned to the colour of the flowers than are those visiting Hound's-tongue, which are also influenced by the height of the flowers.
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

198 A. MANNING
DISCUSSION
It is clear that, once bumble-bees have been foraging for some time on a group of plants the stimuli to which they react, may change markedly. Their partially inborn responses to the colour and scent of the flowers, which initially attracted and directed them, may become entirely superseded in importance save at very close range. Conditioned stimuli from the plant's form may take their place, together with an accurate knowledge of the site of individual plants or groups. 'The extent to which these changes occur will vary greatly according to the type of plant; Hound's-tongue and Fox- glove probably represent two extremes and all the differences between the foraging behaviour of bumble-bees on these two plants can be correlated with the different conspicuousness of their flowers. Flowers are a plant's primary means of attracting the visits of bees, and correspondingly we find that most bee-pollinated plants have relatively large, conspicuous ones. How- ever, conspicuousness is only the initial means of attraction; in the end it is probably a good supply of nectar or pollen which makes a flower attractive. Even so, it is necessary for a plant to attract bees in the first place, before they are 'aware' of the food supply and thus the conspicuousness of its flowers, either by colour or scent, cannot be reduced below a certain level.
Once bees have fouild a good crop, it is clear that their behaviour is most flexible, and they can adapt themselves to any peculiarities of the plant concerned. Thus they rapidly become conditioned to the general form of Hound's-tongue, because its flowers are of little use in guiding them, although it is likely that such a resort will be necessary with only very few of the plants upon which they feed. Conversely, bees will not learn any more than is necessary. When the flowers are conspicuous enough, as with Foxglove, they rely entirely upon them and learn nothing of the plant's general form. Between these extremes one must expect to find all intermediates, where both flowers and form play a part.
With both plants studied, bees soon enlarged their foraging area to the maximum extent and, since the number of plants was limited, incorporated all that were available. They learnt the position of each plant which was beyond their visual range when they left the plant next nearest to it, and in this way became completely independant of direct stimuli from the plant itself to guide them very close to it. Learning the position of individual plants may be a slightly anomalous feature of foraging behaviour, which is again a special adaptation to the plants growing separately and often long distances apart. However some kind of orientation to the crop on which they are foraging must be universal among honey- and bumble-bees, for it is known that they nearly always remain constant to a particular group of
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

FORAGING BEHAVIOUR OF BUMBLE-BEES I99
plants even in a dense and continuous crop. This means that to fly to their foraging area they must rely partly upon general landmarks and, to this extent, become independant of stimuli from the actual flowers. It is unknown how bees limit their foraging area on a dense crop. It is probable that they rely on direct stimuli from the flowers, when they are in the area, and do not learn the position of each, although RIBBANDS (I949) gives the case of a bee returning at the beginning of each trip to the same flower in a fairly dense crop of Shirley poppies. On a patchy crop it is very likely that bees do learn something of the location of the clumps of plants which they visit, for each clump may not be directly visible from the others.
The cases described in this paper show how, since bees will adapt their foraging behaviour to meet any requirements of the particular crop, a plant may assume a considerable degree of 'control' in the evolutionary relationship between itself and the bees which pollinate it. Because of its good nectar supply, which makes it ultimately very attractive to bees, Hound's-tongue has a lessened need to develop large and conspicuous flowers. Provided that it can attract quite a few individual bees early in its flowering season, their constancy and other adaptations to offset the inadequacy of its flowers for guidance, will ensure that all the plants over a wide area will be well pollinated. Even so, we find that, presumably because of their inconspicuous- ness, the earliest Hound's-tongue flowers are rarely visited and many not set seed. This delay in the visits of bees after the onset of flowering must be somewhat disadvantageous.
While it is clear that many other factors will be important, it does seem possible that, once a certain level of mutual adaptation had been established between bees and plants, there may have been reduced selective advantage, as far as their pollination was concerned, in the plants developing more conspicuous flowers.
SUMMARY Bumble-bees foraging an Hound's-tongue return to the same plants again and again
for a period of io days or more. When the plants are close together (6' or less apart), they visit them in a rather irregular fashion, and experiments show that, while they have some knowledge of the position of individual plants with respect to other landmarks, bees are responding largely to the general form of the plants. They become conditioned to the form very rapidly after a few visits to plants, when they are responding largely to the flowers. On the other hand, when bees are visiting more distant plants, more than (12') 4 m apart, they are reacting almost entirely to the site of each. This, it has been shown, they fix with great accuracy during one or more brief orientation flights upon leaving the plant after their first few visits to it. They often find distant and partly hidden plants during the course of rambling 'searching flights', which usually start from the central group of plants. After a short time they develop a distinct route among the distant plants, and their flight is usually very direct, in contrast to the
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

2(X) A. MANNING
wavering flight within the central group, where they frequently react to and search for flowers, on other flowerless plants which resemble Hound's-tongue in general form.
It is suggested that much of this behaviour is a direct adaptation to the Hound's-tongue plant because its flowers, though yielding abundant nectar, are so small and inconspic- uous. This is confirmed by a comparison with the behaviour of bees foraging on Fox- glove, which has very large, conspicuous flowers. Here bees rely almost entirely on direct stimuli from the flowers and never become conditioned to the form of the plant or visit plants without flowers. They learn practically nothing of the positions of plants which are close together, but do learn the sites of individual plants when these are more than about 12 feet from the next nearest. Unlike bees visiting Hound's-tongue, Foxglove *bees become firmly conditioned to the colour of the flowers, and do not visit paper models of any colour other than a purple, closely resembling that of the real flowers.
These results show that the foraging behaviour of bumble-bees is most flexible and that they can adapt themselves to suit very different features of the plants they visit. Hound's-tongue and Foxglove probably represent two extremes, and between them there may be many plants where both the flowers and general form play a part in guiding bees. The evolutionary implications of the work are briefly discussed.
REFERENCES
BONNIER, G. (i9o6). Sur la division du travail chez les abeilles. - C. R. Acad. Sci. Paris. 143, p. g4i-6.
DARWIN, C. (I876). The Effects of Cross and Self Fertilisation in the Vegetable King- dom. - Murray, London.
FRANK, A. (ig4i). Eigenartige Flugbahnen bei Hummelmannchen. - Z. vergl. Physiol. 28, P. 467-84.
FREE, J. (I953). Reported in Rep. Rothamsted. exp. Sta. I953. GILTAY, E. (I9o4). Ueber die Bedeutung der Krone bei Bliiten und uiber das Farben-
unterscheidungsverm6gen der Insekten. - Jb. wiss. Bot. 40, p. 368-402. HAAS, A. (i949). Arttypische Flugbahnen von Hummelminnchen. - Z.vergl. Physiol.
3I, p. 28I-307. KNUTH, P. (19o6). Handbook of Flower Pollination (Trans. by J. R. AINSWORTH-DAVIS.)
- 3 vols. Clarendon Press, Oxford. MULLER, H. (I882). Versuche iuber die Farbenliebhaberei der Honigbiene. - Kosmos
I2, P. 273-99. OPFINGER, E. (1931). Ueber die Orientierung der Biene an der Futterquelle. - Z.vergl.
Physiol. I5, P. 43I-87. RIBBANDS, C. R. (i949). The foraging method of individual honeybees. J. Anim.
Ecol. i8, P. 47-66. SINGH, S. (I950). Behaviour studies of honeybees in gathering nectar and pollen. -
Bull. Cornell agric. Exp. Sta. No. 288, p. 1-59. TINBERGEN, N. & W. KRUYT (1938). Ober die Orientierung des Bienenwolfes (Philan-
thus triangulum Fabr.) III. Die Bevorzugung bestimmter Wegmarken. - Z. vergl. Physiol. 20, P. I5I-6i.
ZUSAMMENFASSUNG Bis zu einer Periode von IO und mehr Tagen kehren Hummeln, die an Hundszungen
Futter suchen, immer wieder zu derselben Pflanze zuriick. Wenn die ;Pflanzen nahe bei einander stehen (etwa 2 m oder weniger von einander getrennt), besuchen die Hum- meln diese in ziemlich unregelmassiger Weise. Obwohl die Hummeln etliche Kenntnisse iiber den Standplatz individueller Pflanzen in Bezug zu anderen Landmarken besitzen, reagieren sie hauptsachlich auf die allgemeine Form der Bluten. Sie erlernen die
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions

FORAGING BEHAVIOUR OF BUMBLE-BEES 201
Bliitenform sehr rasch, schon nach wenigen Besuchen, wenn sie in erster Linie auf die Bluten reagieren. Besuchen sie jedoch entfernter stehende Pflanzen (mehr als 4 m weg), so reagieren sie nahezu ganzlich auf den Platz einer jeden. Wiihrend eines oder mehrerer kurzer Orientierungsfliige fixieren sie diesen mit grosser Genauigkeit, nach- dem sie die Pflanze nach dem ersten Besuch verlassen haben. Oft finden sie entfernte und teilweise versteckte Pflanzen, wenn sie auf ,,Suchfligen" umherschweifen. Dabei starten sie gewohnlich von einer zentralen Pflanzengruppe. Nach kurzer Zeit ent- wickeln sie eine distinkte Route zwischen den entfernten Pflanze. Sie fliegen dann gewohnlich direkt, im Unterschied zu dem unentschlossenen Fliegen in der zen- tralen Gruppe, und reagieren dann haufig auf bliitenlose Pflanzen und suchen nach Bluten, wenn diese Pflanzen der allgemeinen Form der Hundszungen ihneln.
Wir nehmen an, dass es sich bei vielen von derartigen Verhaltensweisen um eine direkte Adaptation an Hundszungen handelt, da deren Bliuten klein und unmerkbar sind, obwohl sie eine Fuille von Nektar abgeben. Dies konnte in Versuchen mit Hummeln bestatigt werden, die an Fingerhiiten (Digitalis) Futter suchten, die ja grosse, auf- fallige Bliiten besitzen. Dabei verliessen sich die Hummeln fast ganzlich auf die Bliutenform und lernten niemals die Form der Pflanze. Auch besuchten sie niemals blhitenlose Pflanzen. Sie lernten praktisch nichts uber die Position eng benachbart stehender Pflanzen, doch lernten sie die Lage individueller Pflanzen, wenn diese mehr als 4 m von der nachsten entfernt war. Im Gegensatz zu Hundszungenbesuchern, erlernten Fingerhutbesucher sehr fest die Bliitenfarbe und besuchten keine Papiermodelle mit irgend einer anderen Farbe als Purpur, obwohl sie den echten Bliiten sehr ahnelten.
Diese Ergebnisse zeigen die Plastizitat des Futtersuchverhaltens der Hummeln. Sie konnen sich an sehr verschiedenen Merkmalen der besuchten Pflanzen anpassen. Hunds- zunge und Fingerhut stellen wahrscheinlich zwei Extreme dar. Zwischen diesen k6nnen sehr viele Pflanzen stehen, bei denen sowohl die Bliuten wie die Gestalt der Pflanze die Hummeln lenken. Die stammesgeschichtlichen Aspekte dieser Untersuchung werden kurz gestreift.
This content downloaded from 185.44.79.40 on Wed, 25 Jun 2014 10:52:07 AMAll use subject to JSTOR Terms and Conditions





























