Embed Size (px)
Citation preview

UNIVERZITET U NIŠU
PRIRODNO-MATEMATIČKI FAKULTET
DEPARTMAN ZA BIOLOGIJU I EKOLOGIJU
Dragan Vulić
Uticaj pastrmskog ribnjaka u selu Pasjač kod Pirota na
sastav zajednice makrozoobentosa i fiziko-hemijske
karakteristike vode
MASTER RAD
Niš, 2019.

UNIVERZITET U NIŠU
PRIRODNO-MATEMATIČKI FAKULTET
DEPARTMAN ZA BIOLOGIJU I EKOLOGIJU
Uticaj pastrmskog ribnjaka u selu Pasjač kod Pirota na sastav
zajednice makrozoobentosa i fiziko-hemijske karakteristike
vode
MASTER RAD
Kandidat:
Dragan Vulić, 258
Mentor:
Dr Milica Stojković Piperac
Niš, 2019.

UNIVERSITY OF NIŠ
FACULTY OF SCIENCE AND MATHEMATICS
DEPARTMENT OF BIOLOGY AND ECOLOGY
The influence of a trout fish farm in the village of Pasjač near
Pirot on the composition of the macrozoobenthos community
and the physico-chemical characteristics of water
MASTER THESIS
Candidate:
Dragan Vulić, 258
Mentor:
Dr Milica Stojković Piperac
Niš, 2019.

Biografija kandidata
Dragan Vulić, rođen je 20.11.1992. godine u Prokuplju. Osnovnu školu ”Stojan
Novaković” u Gornjoj Draguši završava 2007. godine, nakon toga upisuje Gimnaziju u
Blacu (opšti smer). Prirodno-matematički fakultet u Nišu upisuje 2011. godine na
Departmanu za biologiju i ekologiju. Osnovne akademske studije završava 2016. godine i
iste godine upisuje Master akademske studije, smer Ekologija i zaštita prirode, na istom
fakultetu.

Zahvalnica
Neizmernu zahvalnost dugujem svojoj mentorki Dr Milici Stojković
Piperac na ukazanom poverenju prilikom odabira master teme, pruženoj
pomoći u obradi rezultata istraživanja kao i na nesebičnoj pomoći i korisnim
savetima tokom pisanja ovog rada.
Mojoj porodici dugujem najveću zahvalnost za neizmernu pomoć i
bezrezervnu podršku, a najviše za svu ljubav koju mi pružaju.

Sažetak
Pastrmski ribnjaci vrše direktan uticaj na brdsko-planinske reke preko polutanata
koje direktno oslobađaju u njih.
U ovom radu ispitivane su promene zajednice makrozoobentosa, kao grupe koja se
od svih vodenih organizama najčešće koristi u biomonitoringu tekućih vodenih ekosistema.
Kako bi se analizirao uticaj pastrmskih ribnjaka na organizme makrozoobentosa,
obavljeno je istraživanje kvalitativnog i kvantitativnog sastava zajednice pre i posle uliva
otpadnih voda iz ribnjaka. Takođe, poređene su i vrednosti fiziko-hemijskih karakteristika
vode na kontrolnim i test lokalitetima.
Rezultati ovog rada pokazuju da pastrmski ribnjaci locirani uz gornje tokove i
izvorišnu zonu reka predstavljaju potencijalnu opasnost za remećenje prirodnih staništa ove
grupe vodenih životinja.
Ključne reči: pastrmski ribnjaci, planinski izvor, zajednica makrozoobentosa, kvalitativni i
kvantitativni sastav, fiziko-hemijske karakteristike vode.

Abstract
Trout fish farms directly impact mountain rivers through the pollutants that release
directly into them.
This study examines changes in the macrozoobenthos community, as the most
commonly used group in biomonitoring of aquatic ecosystems.
In order to analyze the impact of trout fish farm on macrozoobenthos, a qualitative
and quantitative composition of the community before and after fish farm effluent input.
Also, the values of physico-chemical characteristics of water at control and test sites were
compared.
The results of this study indicate that trout fish farms located at upstream river
sections and river source present a potential threat to the natural habitats of this group of
aquatic animals.
Keywords: trout fish farms, mountain spring, macrozoobenthos community, qualitative and
quantitative composition, physico-chemical characteristics of water.

Sadržaj
1. Uvod _______________________________________________________________ 1
1.1 Izvori ________________________________________________________________ 2
1.1.1 Zajednica akvatičnih makroinvertebrata izvorišne zone ____________________________3
1.1.2 Fizičke i hemijske karakteristike izvorišne zone ___________________________________4
1.2 Pastrmski ribnjaci i njihov uticaj na izvore i gornje tokove reka _______________ 6
1.3 Akvatične makroinvertebrate kao bioindikatori _____________________________ 8
1.4 Cilj istraživanja _______________________________________________________ 9
2 Materijal i metode ____________________________________________________ 10
2.1 Oblast istraživanja ____________________________________________________ 10
2.2 Uzorkovanje materijala ________________________________________________ 12
2.3 Analiza podataka _____________________________________________________ 13
3 Rezultati ___________________________________________________________ 14
3.1 Taksonomski sastav zajednice makrozoobentosa ___________________________ 14
3.2 Analiza uticaja pastrmskog ribnjaka na zajednicu makrozoobentosa i fizičko-
hemijskih parametara vode u izvorišnoj zoni Pasjačke reke ________________________ 19
4 Diskusija ___________________________________________________________ 21
4.1 Analiza taksonomskog sastava zajednice makrozoobentosa __________________ 21
4.2 Uticaj pastrmskih ribnjaka na zajednicu makrozoobentosa i fizičko-hemijske
parametare vode ____________________________________________________________ 24
5 Zaključak __________________________________________________________ 26
Literatura __________________________________________________________ 27

1
,,Kada izvor presuši, tek onda znamo vrednost vode”– Benjamin Franklin
1. Uvod
Voda je osnova života i sredina u kojoj je život nastao. Voda zauzima 2/3 Zemljine
površine od čega na kopnene vode odlazi svega 0,5% i podeljene su na površinske i
podzemne vode (Simić i Simić, 2009). Kopnene vode su izolovane jedne od drugih za razliku
od mora i okeana, koji predstavljaju kontinuirana vodena prostranstva. Kopnene vode
možemo podeliti i klasifikovati na osnovu različitih kriterijuma i to prema: kretanju (tekuće
i stajaće), sastavu i opštim fizičkim i hemijskim svojstvima (termalne, slatke, zaslanjene,
slane, mineralne, tresavske), poreklu (prirodne i veštačke) i na osnovu trajanja (stalne i
privremene) (Simić i Simić, 2009).
Simić i Simić (2009) kopnene vode prema kretanju dele na:
Tekuće vode (lotički sistemi) kod kojih se celokupna vodena masa neprekidno kreće
u određenom pravcu i po redosledu postanka to su: izvori, potoci, male i velike reke;
Stajaće vode (lentički sistemi) kod kojih vodeni pokreti nisu takvi da postoji
kontinuiran tok u jednom pravcu, već samo talasi i strujenja i tu spadaju: jezera, bare,
lokve, močvare, ritovi i tresave.
Kompleks abiotičkih faktora u kopnenim akvatičnim ekosistemima predstavlja
složen sistem velikog broja uticaja koji svojim delovanjem određuju strukturu i dinamiku
hidrobiocenoza. U važne fizičke abiotičke faktore spadaju: temperatura, svetlosni režim,
vodena kretanja i struktura i sastav sedimenta.
U prirodi nema hemijski čiste vode. Prirodne vode uvek sadrže neke primese, u njoj
se uvek nalaze rastvorene organske i neorganske materije različitog sastava i koncentracije.
Hemijski sastav voda nije svuda isti i one se razlikuju pre svega po prisustvu i koncentraciji

2
rastvorenih gasova, mineralnih materija i organskih materija koje su u obliku koloida,
suspenzija i emulzija. Hemijski sastav voda je rezultat reakcije vode i njene okoline (stena,
zemljišta i atmosfere), ali je takođe i rezultat aktivnosti živih organizama, kao i materija koje
potiču iz antropogenih aktivnosti.
Živi svet akvatičnih ekosistema (biljne i životinjske vrste) grupisan je u populacije,
a populacije su dalje svrstane u hidrobiocenoze čija struktura i dinamika zavise od
specifičnih uslova akvatičnih ekosistema. Hidrobiocenoze su grupisane prema adaptivnim
karakteristikama organizama čije populacije naseljavaju određene ekološke zone i staništa
vodenih ekosistema. Na osnovu toga možemo izdvoljiti sledeće hidrobiocenoze:
Neuston – plutajući organizmi;
Plankton – lebdeći organizmi;
Nekton – plivajući organizmi i
Bentos – organizmi koji naseljavaju dno.
1.1 Izvori
Izvori su mesta prirodnog izlaza podzemne vode na površinu zemlje koji
predstavljaju početne delove potočnih i rečnih tokova. Nastanak odnosno pojavljivanje
samog izvora uslovljeno je postojanjem pukotina u steni koja je prožeta sa sabirnim i
izlaznim područjem podzemnih voda, a takođe zavisi i od konfiguracije terena i nivoa
podzemnih voda. Po svojim fizičko-geografskim, fizičkim i hemijskim osobinama, kao i po
živom svetu, jasno se izdvajaju od ostalog dela toka (Marković, 1998).
Izvori kao ekosistemi su pogodni za istraživanje višeg stupnja ekološke integracije
životnih zajednica, koje čine sve biotičke komponente ekosistema kao najpotpunije ekološke
celine. Kao takve celine, značajne su za proučavanje odnosa između životinjskih zajednica i
ekoloških faktora koji utiču na njihovu raznovrsnost i rasprostranjenost. Nalaze se na prelazu
između podzemne i nadzemne vode predstavljajući zaseban biotop, kojeg nazivamo ekoton.
Pogodna su mesta za proučavanje ekologije i evolucije vrsta kao i biotičkih interakcija i
predstavljaju specifične ekosisteme (Glazier, 1998).

3
Postoje različite podele izvora. Najopštija i najosnovnija podela je prema obliku i
načinu izbijanja vode na površinu. Najčešće se izdvajaju tri tipa izvora:
reokreni tip (tekući izvori) – voda izlazi iz zemlje u mlazu, odmah otiče i gradi vodeni
tok;
limnokreni (lokvičasti izvori) – voda se najpre sakuplja u bazenu u vidu lokve,
odnosno jezera male površine, sa čijih strana ili dna izbija, a potom se razliva preko
ivica bazena u dalji vodeni tok;
helokreni – močvarni, difuzni ili podvirni izvori, u narodu poznati kao pištevine ili
pištoljine, gde voda izvire na većoj površini natapajući zemlju, pri čemu se stvara
zamočvareno zemljište sa koga se formira rečni tok.
Osim ove postoji i podela izvora na osnovu vremena trajanja (stalni, periodični,
povremeni), hidrauličnog mehanizma (gravitacioni i arteški), po hemizmu (slatkovodni i
mineralni), po stepenu mineralizacije, po izdašnosti i sl.
Važna karakteristika izvora je njihova “mozaična struktura“. Uprkos njihovim
obično ograničenim dimenzijama, izvori nisu homogena sredina, već se sastoje od više
mikrostaništa. Ova mozaična struktura sa nekoliko mikrostaništa prvi put je predstavljena
od strane Illies-a i Botosaneanu-a (1963), a kasnije je potvrđena od strane drugih istraživača
(Weigand, 1998). Ova osobina, zajedno sa velikim brojem različitih supstrata koji favorizuju
kolonizaciju i veliku dostupnost hrane, glavni je razlog za visok biodiverzitet i gustinu
populacija koje se nalaze u izvorima (Stanford i sar., 1994).
1.1.1 Zajednica akvatičnih makroinvertebrata izvorišne zone
Sam izvor tekućica (krenon) smatra se posebnom ekološkom celinom koja se pre
svega odlikuje stabilnošću većine ekoloških faktora, pre svega temperature, hemijskog
sastava i dr. Faktori koji mogu da variraju i da time utiču na strukturu životnih zajednica
izvora su pre svega protok vode koji određuje i tip izvora i to na reokreni (tekući) ili
limnokreni (stajaći). Osim karaktera toka vode i prisustvo vegetacije utiče na strukturu
životne zajednice izvora jer menja karakter podloge i količinu svetlosti.

4
Makrozoobentos izvora sastavljen je od relativno malog broja oksifilnih i
hladnostenotermnih predstavnika kao što su: amfipodni rakovi (Gammarus sp., G.
balcanicus, koji su posebno brojni u kraškim izvorima, kakvih je Srbiji veliki broj),
Turbellaria (Crenobia alpina, na većim nadmorskim visinama), Gastropoda (Bythinella
austriaca, veoma sitni oblici, 3-5mm, čija brojnost populacija u nekim izvorima može da
bude veoma velika), Coleoptera (Elmis maugei), Diptera (Diamesa, Bezzia, Dixa),
Plecoptera (Leuctra, Isoperla, Perla, Protonemura), Ephemeroptera (Baetisalpinus, Baetis
sp., Ecdyonuridae), Trichoptera (Drusus, Sericostoma).
1.1.2 Fizičke i hemijske karakteristike izvorišne zone
Glavne fizičke i hemijske karakteristike izvora se mogu rezimirati fokusirajući se na
identifikaciji karakterističnih staništa i indikatora promene životne sredine. Sve faktore koji
su relevantni za karakterizaciju izvora kao staništa trebalo bi, ukoliko je to moguće, istražiti
tokom cele godine ili bar jednom u različitim godišnjim dobima. Godišnji opseg je najbolji
jer sadrži najpouzdanije vrednosti za procenu stabilnosti i karakteristika ne samo za
okruženja izvora, već i za vodonosni sloj (Wilhelm, 1956). Godišnje mere pražnjenja mogu
otkriti važne informacije o dimenzijama i tipologiji vodonosnika. Vrlo često su izvori sa
najvećim i najstabilnijim pražnjenjem iskorišćeni za snabdevanje pitkom vodom.
Temperatura vode zavisi od nekoliko faktora, a najrelevantnije su dubina i brzina fluksa
podzemne vode, dimenzije i srednja visina slivnika.
Izbijajući na površinu gde su respiracija i dekompozicija dominantni metabolički
procesi, izvorne vode bi trebale biti deficitne kiseonikom, a obogaćene ugljen dioksidom.
Međutim, planinski izvori se uglavnom hrane malim i površinskim vodonosnicima, a zonu
iznad izvora često čine velike stene koje određuju visoku poroznost i lak kontakt između
podzemnih voda i atmosfere. Zato su planinski izvori i reokreni izvori često dobro
oksigenisani (Cantonati i sar., 1998; Haseke i Pröll, 2006).
U skoro prirodnim izvorima, provodljivost se uglavnom određuje litološkim
karakteristikama podloge, a naročito njegovoj rastvorljivosti. Na području koje je homogeno
po pitanju geologije, provodljivost podzemnih voda je veća od površinskih voda. Osim toga,
temperatura i provodljivost često su u korelaciji sa nadmorskom visinom, pošto se oba
smanjuju sa višim nadmorskim visinama. Niže temperature odražavaju niže srednje

5
vrednosti u slivu i dolazi do nižih provodljivosti, jer je vreme boravka vode u vodonosniku
kraće što ograničava interakciju između vode i litološke podloge (Cantonati i sar., 1998).
pH se određuje pomoću litologije, sadržaja ugljen dioksida, kiselim zagađivačima
vazdušnog porekla (nitrati, sulfati) i, u izvorima sa sporim tekućim vodama, pomoću
fotosintetske aktivnosti algi, mahovine i viših biljaka. U helo- i reohelokrenim izvorima
organske kiseline takođe mogu biti važne. Dok su izvorina karbonatnoj podlozi dobro
puferirani i imaju prilično stabilan pH, sezonske promene mogu biti izraženije u izvorima sa
slabo mineraliziranom vodom. U slabo puferiranim izvorima, naročito na visokim
nadmorskim visinama, pH može da se smanji tokom perioda kiša (Brehm, 1986).
Neorganski azot u obliku amonijaka obično je prisutan samo u malim količinama u
vodama koje nisu dobro oksigenisane (kao što su helokreni izvori gde se mogu naći veće
vrednosti). Vrednosti veće od 30 µg 1-1 N-NH+
4 su obično zbog organskog zagađenja (Brehm
i Meijering, 1996). Zbog niske sposobnosti apsorpcije mnogih zemljišta, podzemne vode i
izvori često pokazuju veće vrednosti nitrata od površinskih voda istog područja (Maire i
Pomel, 1994). Međutim, vremenske i prostorne promene koncentracije nitrata smatraju se
jednim od glavnih hemijskih indikatora antropogenog uticaja. Oni potiču posebno od
poljoprivrednih aktivnosti u drenažnom bazenu ili od emisija azotnih oksida u saobraćaju.
One utiču na vodene sisteme kao jake kiseline i kao hranjive materije i zbog toga su ključni
faktori za acidifikaciju i eutrofikaciju. Postoje dugotrajne promene koncentracije nitrata u
izvorištima, koje se odnose na povećanje atmosferskih padavina. Izvori su generalno
oligotrofne sredine sa ukupnim vrednostima fosfora nižim od 10 µg 1-1.
Sulfati u izvorištima mogu biti geogeni (oksidacija sulfida) ili antropogeni (sumporni
oksidi industrijskih emisija koji postanu sulfati u atmosferi). Inicijalno sulfati su bili glavni
sastojak kiselih kiša, ali su ih zamijenili nitrati 1980-ih, kada su mnoge zemlje drastično
smanjile emisije sulfata (Psenner i Catalan, 1994). Izvori na silikatnoj podlozi, na koje nisu
značajno delovale kisele kiše obično imaju nisku koncentraciju sulfata. Na karbonatnoj
podlozi mogu se naći vrlo visoke vrednosti sulfata u kojima su u drenažnom bazenu prisutni
gipsni slojevi ili gipsni uglovi.
Silicijum je važan nutrijent za dijatomeje. U izvorima skoro nikada ne dostiže
ograničavajuće vrijednosti (<0.10 mg 1-1) čak i u karbonatnim planinskim područjima. Na
silikatnim podlogama, veće vrednosti su zabeležene u helo- i reohelokrenim izvorima
(Cantonati i sar., 1998).

6
Rastvoreni organski ugljenik (DOC-engl. Dissolved organic carbon) u izvorima
potiče uglavnom od raspadanja organskog materijala u tlu iz drenažnih bazena i od ulaza
kroz površinski oticaj. DOC u izvorima su obično skromni i često manji od 1 mg 1-1.
Sezonske promene su vrlo ograničene i odnose se na sneg i vreme kiše. Još izraženije
promene su uzrokovane antropogenim uticajem (Wilhelm, 1956).
Izvori su kolonizovaniod strane taksona koji se jasno razlikuju od vodenih tokova ili
jezera. Prema Bodon-u i Gaiter-u (1989) i Baldin-u i Gaiter-u (1990), istraživanje
makroinvertebratnih asembleja izvora pruža korisne informacije o različitim aspektima, kao
što su pojave zagađenja, hidraulički režim i najverovatnije poreklo vode.
1.2 Pastrmski ribnjaci i njihov uticaj na izvore i gornje tokove reka
Gajenje riba se najvećim delom obavlja u šaranskim (toplovodnim) i pastrmskim
(hladnovodnim) ribnjacima (preko 95% ukupne količine proizvedene ribe), u malim
količinama u kavezima, ograđenim ili pregrađenim delovima prirodnih i antropogenih voda,
dok je gajenje riba u akvarijumima uglavnom u domenu hobija sa malim brojem
specijalizovanih odgajivačnica i jednim javnim akvarijumom (Marković i Mitrović
Tutundžić, 2003; Marković i Poleksić, 2007; Marković i sar., 2009).
Pastrmska akvakultura se konstantno povećava i pastrmski ribnjaci se obično
izgrađuju u gornjim tokovima reka zato što ovakav tip ribnjaka zahteva nisku temperaturu
vode, oksegisanu i nezagađenu vodu. Pastrmski ribnjaci predstavljaju potencijalnu pretnju
za kvalitet vode gornjih tokova, naročito tokom perioda smanjenog protoka vode
(Boaventura i sar., 1997). Negativni uticaj pastrmskih ribnjaka na tokove je najviše uslovljen
ostacima riblje hrane i fekalnim materijama (Liao, 1970), koje vode ka smanjenju kvaliteta
vode i promenama u strukturi zajednice dna. Prema tome, relativno duga istraživanja koja
su sprovedena u prethodnih 30 godina o uticaju pastrmskih ribnjaka na tokove su se
prvenstveno fokusirala na promene u fiziko-hemijskim parametrima vode i sastavu i
strukturi dna reka. Ova istraživanja su pokazala da su ove promene prvenstveno ispoljile:

7
povećanje koncentracija kako rastvorljivih tako i nerastvorljivih fosfata (Boaventura
i sar., 1997; Muñoz, 1989; Foy i Rosell, 1991; Pulatsu i sar., 2004; Bartoli i sar.,
2007; Oberdorff i Percher, 1994);
povećanje sadržaja azota (najviše u formi NH4 i organskog vezanog azota)
(Boaventura i sar., 1997; Muñoz, 1989; Foy i Rosell, 1991; Selong i Helfrich, 1998;
Bartoli i sar., 2007; Oberdorff i Percher, 1994);
smanjenje koncentracije rastvorenog kiseonika (Muñoz, 1989; Pulatsu i sar., 2004;
Selong i Helfrich, 1998; Oberdorff i Percher, 1994);
i akumulacija suspendovanih čestica na rečnom dnu (Boaventura i sar., 1997).
Uticaj pastrmskih ribnjaka na makrozoobentos (Camargo, 1992a, b; Loch i sar.,
1996) i ihtiofaunu (Oberdorff i Percher, 1994) gornjeg toka, a posebno izvorišne zone nije
detaljno istražen. Generalno, otpadne vode sa pastrmskih ribnjaka uzrokuju opadanje
diverziteta bentosnih makroinvertebrata, zamenom senzitivnih vrsta manje senzitivnim i
promenama u trofičkoj strukturi zbog povećane zastupljenosti kolektora a smanjene
zastupljenosti strugača i sekača (Camargo, 1992a, b; Loch i sar., 1996).
Uticaj ribnjaka je kontrolisan:
veličinom ribnjaka,
prisustvom i tipom sistema za prečišćavanje vode i
sastavom i strukturom riblje hrane koja se koristi.
U cilju definisanja odgovarajućih zakonskih mera koji su potrebni da osiguraju
potpunu zaštitu osetljivih pastrmskih tokova, potrebno je istražiti što je više različitih tipova
pastrmskih ribnjaka. Ovo je naročito važno u zemljama u razvoju sa rapidnim povećanjem
pastrmske akvakulture gde su zakonske mere ove aktivnosti nepostojeće ili neadekvatne.
Broj pastrmskih ribnjaka se u Srbiji u zadnjih 10 godina dupirao, a u budućnosti se
procenjuje da će se povećati za 3 do 5 puta (Marković i sar., 2007). Uprkos povećanom rastu
pastrmskih ribnjaka, postoji samo mali broj istraživanja o uticaju na akvatične ekosisteme
u Srbiji, a zakonske mere o emisiji štetnih supstanci iz pastrmskih ribnjaka su i dalje u fazi
formulisanja.

8
1.3 Akvatične makroinvertebrate kao bioindikatori
Kao najpouzdaniji pokazatelji ekološkog stanja vodenih ekosistema, od svih
slatkovodnih organizama koji se koriste u biomonitoringu, pokazao se makrozoobentos
(Hellawell, 1986; Bonada i sar., 2006; Carter i sar., 2006). Zajednica makroinvertebrata se
zbog niza osobina svrstava u pouzdane pokazatelje dinamike ekoloških uslova koji vladaju
u akvatičnim ekosistemima. Neke od tih osobina, kao i praktični razlozi koji im to
omogućavaju, su (Simić i Simić, 2009):
grupa sadrži mnogo sedentarnih formi;
relativno dug životni ciklus;
dobro razvijena taksonomija;
uzorkovanje i posmatranje je jednostavno;
većina grupa je dobro proučena sa dostupnim ključevima za determinaciju;
veliki broj razvijenih metoda za analizu dobijenih podataka.
U odnosu na osobine koje je neophodno da bioindikatori poseduju, može se reći da
su akvatični makrobeskičmenjaci, kao grupa pogodni za primenu u biomonitoringu
(Paunović, 2007; Marković, 2014; Popović, 2014; Raković, 2015; Zorić, 2015; Tomović,
2015; Atanacković, 2015).

9
1.4 Cilj istraživanja
Osnovni cilj ovog istraživanja je utvrđivanje uticaja pastrmskog ribnjaka u selu
Pasjač na sastav zajednice makrozoobentosa kao i fiziko-hemijske parametare vode pre i
posle ribnjaka u izvorišnoj zoni Pasjačke reke. Osnovni zadaci ovog rada su:
1. utvrđivanje faunističkog sastava makrozoobentosa u izvorišnoj zoni Pasjačke reke;
2. detektovanje promene fiziko-hemijskih parametara vode;
3. detektovanje promene u sastavu zajednice makrobeskičmenjaka.

10
2 Materijal i metode
2.1 Oblast istraživanja
Istraživanje je vršeno na izvoru Kladenac u selu Pasjač, na Suvoj planini koje je 15
km udaljeno od grada Pirota, i na ispustu iz pastrmskog ribnjaka. Izvor se nalazi na
nadmorskoj visini od 698m, sa koordinatama 43° 04′ 33″ SGŠ i 22° 30′ 00″ IGD. Od izvora
na 30 m se nalazi pastrmski ribnjak do kog je voda kanalisana. Ispust vode se nalazi na 10m
od ribnjaka gde voda iz ribnjaka preko cevi otiče dalje i formira Pasjačku reku.
Slika 1. Izvor Kladenac (Autor: Dragan Vulić)

11
Slika 2. Pastrmski ribnjak (Autor: Dragan Vulić)
Slika 3. Ispust iz pastrmskog ribnjaka (Autor: Dragan Vulić)

12
2.2 Uzorkovanje materijala
Prikupljanje uzoraka vršeno je u proleće, jesen i zimu 2018. godine. Istraživanja su
obavljena na po jednom lokalitetu iznad i ispod ispusta otpadne vode ribnjaka, na kojima su
određena po 3 mikrostaništa. Tip mikrostaništa karakteriše sastav podloge, a prilikom
odabira istog se uzorkuje na onim tipovima koji su procentualno najzastupljeniji. Pre
uzorkovanja je izmereno nekoliko abiotičkih parametara in situ korišćenjem uređaja WTW®
Multi 340i probe, i to: temperatura, elektro-provodljivost, pH vrednost, rastvoreni kiseonik
(O2 (mg/l)) i saturacija (O2 [%]). Potom su uzeti uzorci vode u tamnim staklenim flašama
kako bi u labaratoriji odredili biohemijsku potrošnju kiseonika (BPK5)1 (koja je procenjena
korišćenjem standardne metodologije preporučene od strane APHA (1999)). Koncentracija
azota iz nitrata (NO3-N), fosfora iz ortofosfata (PO4-P) i azota iz amonijaka (NH4-N),
izmerena je u laboratriji pomoću portabl fotometra PC MultiDirect Lovibond®.
Prilikom uzorkovanja na samom izvoru i na ispustu iz pastrmskog ribnjaka odabrani
su transekti za uzorkovanje u dužini od 1m. Bentosna zajednica makroinvertebrata je
uzorkovana pomoću kick-net mreže, čije su dimenzije rama 25x25 cm, a promer okaca mreže
je 500μm. Duž svakog transekta su uzeta po tri uzorka. Mreža je postavljana normalno u
odnosu na podlogu, a ispred mreže površina podloge koja odgovara dimenzijama rama je
uzburkavana da bi se uneo podignuti materijal u mrežu. Sav sakupljeni materijal je pakovan
u plastične kese koje su sadržale etikete sa nazivom lokaliteta, datumom i oznakom
poduzorka, ispisane grafitnom olovkom.
Po uzorkovanju uzorci su konzervirani u 70% etil-alkoholu i preneti u laboratoriju
na ispiranje, čišćenje, sortiranje i identifikaciju. Sakupljene jedinke makrozoobentosa su
identifikovane do nivoa roda korišćenjem adekvatnog ključa za determinaciju (Kerovec,
1986).
1 BPK je količina kiseonika koja je potrebna mikroorganizmima da razlože organske materije u vodi na ugljen-dioksid, vodu i minerale.

13
Slika 4. Uzorkovanje makrozoobentosa Kick net mrežom (https://www.google.rs/search)
2.3 Analiza podataka
Sa ciljem grupisanja lokaliteta sa sličnim kvalitativnim i kvantitativnim sastavom
ispitivanih grupa kao i fizičko-hemijskim parametrima vode, u ovom radu korišćena je
nMDS analiza (Nemetričko multidimenzionalno skaliranje eng. Nonmetric
multidimensional scaling). Kao rezultat ove analize dobija se prikaz tačaka (lokaliteta) u
dvodimenzionom prostoru, raspoređenih tako da distanca među njima bude direktno
proporcionalna stepenu sličnosti. Kao mera sličnosti odnosno različitosti u ovom radu
korišćena je Bray-Curtis similarity index u slučaju poređena zajednica i euklidova distanca
kada se ispitivala sličnost lokaliteta u pogledu kvaliteta vode. Parametar „nonmetric stress“
predstavlja stepen neuspešnosti nMDS analize u vizualizaciji. Ukoliko je vrednost stresa
manja od 0.25, da bi dobijeni rezultati su pouzdani. Kako bi se ispitalo da li je kvalitet vode
statistički značajno lošiji na mestu izliva ribnjaka korišćen je Mann Whitny test. Sa ciljem
utvrđivanja kombinacije abiotičkih faktora koji utiču na sastav zajednice
makrobeskičmenjaka, korišćena je multipna regresiona analiza BEST. U ovoj analizi izvdaja
se set sredinskih paratetara, kao nezavisnih promenljivih, koji na najbolji način objašnjava
varijabilnost zajednice makroinvertebrata, kao zavisne promenljive. nMDS i BEST analiza
su urađene u statističkom paketu PRIMER v6, a Mann Whitny test u statističkom paketu
SPSS v19.

14
3 Rezultati
3.1 Taksonomski sastav zajednice makrozoobentosa
Na osnovu prikupljenog materijala iz ispitivanog izvora, na dva istraživana lokaliteta
(UP - izvor, DOWN - ispust) u toku tri sezone (proleće (S), jesen (A) i zima (W)),
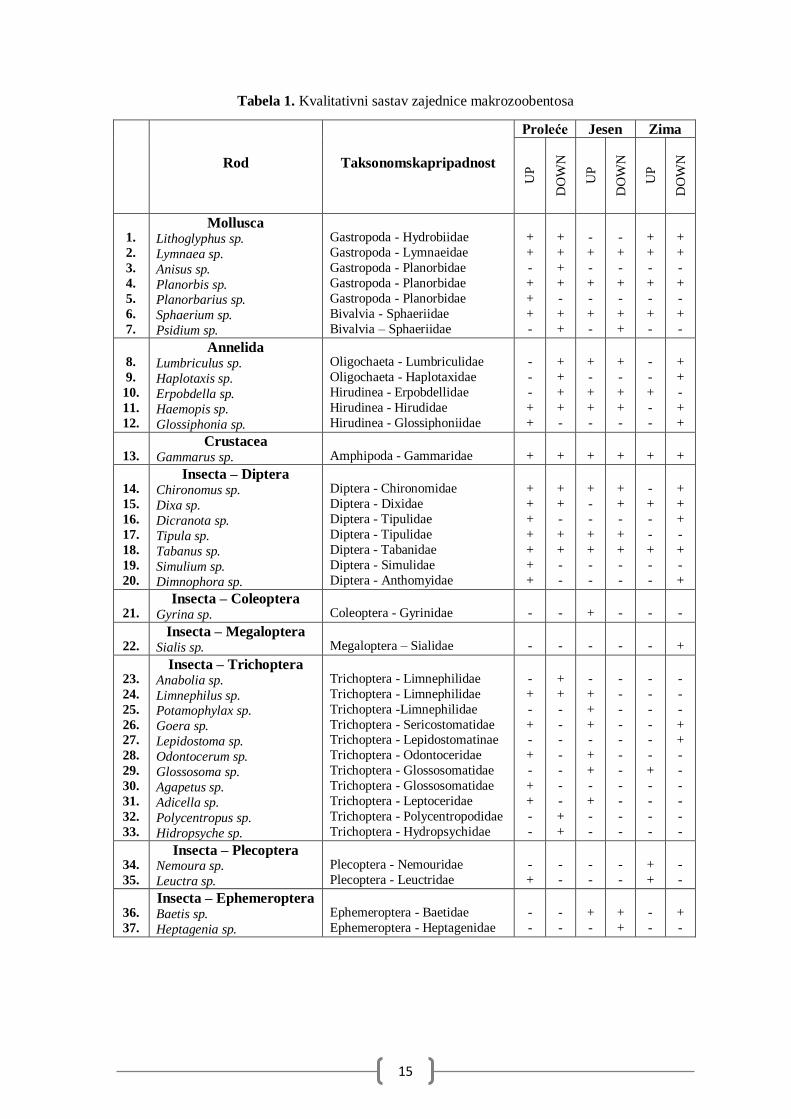
identifikovano je 37 rodova vodenih makrobeskičmenjaka. U Tabeli 1 dat je spisak svih
taksona po lokalitetima i sezonama. Najraznovrsnija grupa su insekti, a među njima se
izdvaljaju Trichoptera i Diptera kao najbrojnije.
U okviru filuma Mollusca određeno je 7 taksona (7 rodova). Tri roda su zabeležena
na oba lokaliteta u svim sezonama i to: Lymnaea, Planorbis i Sphaerium, dok je rod
Planorbarius pronađen samo na izvoru u proleću. Predstavnici filuma Annelida broje 5
taksona. U okviru klase Oligochaeta određena su 2 roda, a u okviru klase Hirudinea 3 roda
gde je rod Haemopis zabeležen u svim sezonama osim na izvoru zimi. Slatkovodni račići iz
reda Amphipoda predstavljeni su samo jednim rodom Gammarus koji je sastavni deo
zajednice makrozobentosa na oba lokaliteta u svim sezonama.
Najveći broj registrovanih taksona je među pripadnicima klase Insecta. U okviru
redova Coleoptera i Megaloptera zabeležen je samo po jedan rod sa po jednom individuom.
U okviru reda Coleoptera rod Gyrina je zabeležen na izvoru u jesen, a u redu Megaloptera
rod Sialis je zabeležen na ispustu zimi. Red Diptera broji 7 rodova među kojima je jedino
rod Tabanus zabeležen na oba lokaliteta u svim sezonama, a rod Chironomus jednino nije
zabeležen na izvoru zimi, a rod Dixa na izvoru u jesen. Rod Simulium je jedino zabeležen na
izvoru u proleće. Najviše zabeleženih rodova je u okviru reda Trichoptera sa čak 10 rodova.
Rodovi Anabolia, Polycentropus i Hidropsyche su zabeleženi samo na ispustu u proleće, rod
Potamophylax je zabeležen samo na izvoru u jesen, rod Lepidostoma samo na ispustu zimi
a rod Agapetus na izvoru u proleće. Redovi Plecoptera i Ephemeroptera su određeni sa po 2
roda. U okviru reda Plecoptera rod Nemoura je zabeležen samo na izvoru zimi, a rod Leuctra
je zabelezen na izvoru u proleće i zimu. U okviru reda Ephemeroptera rod Heptagenia je
zabeležen samo na ispustu u jesen, a rod Baetis je zabelezen na oba lokaliteta u jesen i na
ispustu zimi.
15
Tabela 1. Kvalitativni sastav zajednice makrozoobentosa
Rod
Taksonomskapripadnost
Proleće Jesen Zima
UP
DO
WN
UP
DO
WN
UP
DO
WN
1.
2.
3.
4.
5.
6.
7.
Mollusca Lithoglyphus sp.
Lymnaea sp.
Anisus sp.
Planorbis sp. Planorbarius sp.
Sphaerium sp.
Psidium sp.
Gastropoda - Hydrobiidae
Gastropoda - Lymnaeidae
Gastropoda - Planorbidae
Gastropoda - Planorbidae
Gastropoda - Planorbidae
Bivalvia - Sphaeriidae
Bivalvia – Sphaeriidae
+
+
-
+
+
+
-
+
+
+
+
-
+
+
-
+
-
+
-
+
-
-
+
-
+
-
+
+
+
+
-
+
-
+
-
+
+
-
+
-
+
-
8.
9.
10.
11.
12.
Annelida Lumbriculus sp.
Haplotaxis sp.
Erpobdella sp.
Haemopis sp.
Glossiphonia sp.
Oligochaeta - Lumbriculidae
Oligochaeta - Haplotaxidae
Hirudinea - Erpobdellidae
Hirudinea - Hirudidae
Hirudinea - Glossiphoniidae
-
-
-
+
+
+
+
+
+
-
+
-
+
+
-
+
-
+
+
-
-
-
+
-
-
+
+
-
+
+
13. Crustacea
Gammarus sp.
Amphipoda - Gammaridae
+
+
+
+
+
+
14.
15.
16.
17.
18.
19.
20.
Insecta – Diptera Chironomus sp.
Dixa sp.
Dicranota sp.
Tipula sp.
Tabanus sp.
Simulium sp.
Dimnophora sp.
Diptera - Chironomidae
Diptera - Dixidae
Diptera - Tipulidae
Diptera - Tipulidae
Diptera - Tabanidae
Diptera - Simulidae
Diptera - Anthomyidae
+
+
+
+
+
+
+
+
+
-
+
+
-
-
+
-
-
+
+
-
-
+
+
-
+
+
-
-
-
+
-
-
+
-
-
+
+
+
-
+
-
+
21. Insecta – Coleoptera
Gyrina sp.
Coleoptera - Gyrinidae
-
-
+
-
-
-
22. Insecta – Megaloptera
Sialis sp.
Megaloptera – Sialidae
-
-
-
-
-
+
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
Insecta – Trichoptera Anabolia sp.
Limnephilus sp. Potamophylax sp.
Goera sp.
Lepidostoma sp.
Odontocerum sp.
Glossosoma sp.
Agapetus sp.
Adicella sp.
Polycentropus sp.
Hidropsyche sp.
Trichoptera - Limnephilidae
Trichoptera - Limnephilidae
Trichoptera -Limnephilidae
Trichoptera - Sericostomatidae Trichoptera - Lepidostomatinae
Trichoptera - Odontoceridae
Trichoptera - Glossosomatidae
Trichoptera - Glossosomatidae
Trichoptera - Leptoceridae
Trichoptera - Polycentropodidae
Trichoptera - Hydropsychidae
-
+
-
+ -
+
-
+
+
-
-
+
+
-
- -
-
-
-
-
+
+
-
+
+
+ -
+
+
-
+
-
-
-
-
-
- -
-
-
-
-
-
-
-
-
-
- -
-
+
-
-
-
-
-
-
-
+ +
-
-
-
-
-
-
34.
35.
Insecta – Plecoptera Nemoura sp.
Leuctra sp.
Plecoptera - Nemouridae
Plecoptera - Leuctridae
-
+
-
-
-
-
-
-
+
+
-
-
36.
37.
Insecta – Ephemeroptera Baetis sp.
Heptagenia sp.
Ephemeroptera - Baetidae
Ephemeroptera - Heptagenidae
-
-
-
-
+
-
+
+
-
-
+
-

16
Slika 5. Procentualno učešće jedinki u okviru detektovanih rodova zajednice makrozoobentosa na
izvoru Kladenac u toku svih sezona
Na slici 5 koja predstavlja ukupan broj individua po rodovima na izvoru u toku svih
sezona se vidi da je rod Gammarus najbrojniji sa 62%. Pored njega tu je i rod Lymnaea sa
18% i rod Glossosoma sa 14% dok ostali rodovi imaju veoma mali % udela u ukupnoj
brojnosti individua.
Slika 6. Procentualno učešće jedinki u okviru detektovanih rodova zajednice makrozoobentosa na
ispustu ribnjaka u toku svih sezona
62%18%
14%
2% 1% 3%
UP
Gammarus sp. Lymnaea sp. Glossosoma sp. Chironomus sp. Odontocerum sp. Others
85%
6%
4%2% 3%
DOWN
Gammarus sp. Heptagenia sp. Chironomus sp. Sphaerium sp. Others

17
Na slici 6 mozemo videti da je rod Gammarus zastupljen sa čak 85% od ukupnog
broja individua na ispustu u toku svih sezona. Sa malo većom zastupljenošću se izdvaljaju
rod Heptagenia sa 6% i rod Chironomus sa 4% dok je broj individua ostalih rodova
zanemarljiv.
Slika 7. Sezonska dinamika rodova zajednice makrozoobentosa na izvoru Kladenac toku svih
sezona
Na slici 7 je prikazana relativna brojnost jedinki rodova po sezonama (UP S - izvor
proleće, UP A - izvor jesen i UP W - izvor zima). Na slici 7 se jasno vidi da je rod Gammarus
dominantan u odnosu na ostale rodove u svim sezonama, a naročito tokom jeseni kada je
relativna brojnost ostalih rodova zanemarljiva. Jedino su pored roda Gammarus rodovi
Lymnaea tokom proleća i Glossosoma tokom zime zastupljeni u većoj meri.
Slika 8. Sezonska dinamika rodova zajednice makrozoobentosa na ispustu ribnjaka
0
10
20
30
40
50
60
70
80
90
UP S
UP A
UP W
0
20
40
60
80
100
120
DOWN W
DOWN S
DOWN A

18
Na slici 8 je data relativna brojnost jedinki rodova po sezonama (DOWN W - ispust
zima, DOWN S - ispust proleće i DOWN A - ispust jesen). Kao i na prethodnoj slici i ovde
mozemo videti da je rod Gammarus dominantan samo u još većoj meri nego na izvoru u
svim sezonama.
Gammarus sp
(https://en.wikipedia.orgwi
kiGammarus#mediaFileGa
mmarus_roeselii.jpg)
Heptagenia sp
(https://en.wikipedia.orgwi
kiHeptagenia#mediaFileHe
ptagenia.jpg)
Glossosoma sp
(https://species.wikimedia.
orgwikiGlossosoma#media
FileL3_Dragovištica_3_Gl
ossosoma.jpeg)
Chironomus sp
(https://www.researchgate.netfigureChirono
mus-sp-larvae-were-the-dominant-
Chironomidae-found-in-samples-Image-
source_fig9_307565629
Lymnaea sp
(https://upload.wikimedia.orgwikipediacomm
ons118Lymnaea_sp.jpg)
Slika 9. Najzastupljeniji rodovi na području istraživanja

19
3.2 Analiza uticaja pastrmskog ribnjaka na zajednicu makrozoobentosa i
fizičko-hemijskih parametara vode u izvorišnoj zoni Pasjačke reke
Rezultati nMDS analize pokazali su da se na osnovu kvalitativnog i kvantitativnog
sastava makrozonbentosa mogu izvojiti 3 grupe uzoraka sa sličnošću od 55% (Slika 10).
Slika 10. Grupisanje lokaliteta na osnovu sličnosti u kvalitativnom i kvantitativnom
sastavu makrozoobentosa
U prvoj grupi su se izdvojili uzorci sakupljeni iznad ribnjaka tokom jeseni i zime (UP
A i UP W), dok se uzorak sakupljen iznad ribnjaka tokom proleća (UP S) izdvojio u drugu
grupu. Konačno, svi uzorci sakupljeni ispod uliva ribnjaka (DOWN S, DOWN A i DOWN
W) svrstani su u treću grupu.
Slika 11. Grupisanje lokaliteta na osnovu sličnosti u pogledu kvaliteta vode

20
U pogledu kvaliteta vode, klasifikaciona analiza je izdvojila dve grupe sličnog
sastava lokaliteta (Slika 11). U okviru prve grupe izdvojili su se svi uzorci sakupljeni iznad
ribnjaka tokom proleća, jeseni i zime (UP S, UP A i UP W), dok su se u okviru druge grupe
izdvojili svi uzorci skupljeni ispod uliva ribnjaka (DOWN S, DOWN A i DOWN W).
Mann Whitny test ukazao je koji parametri koji određuju kvalitet vode se statistički
značajno razlikuju pre i posle uliva ribnjaka (Tabela 2).
Tabela 2. Vrednosti fizičko-hemijskih parametara vode ± standardna devijacija na
izvoru i ispustu. Zadeljala slova označavaju da se parametri statistički značajno razlikuju pre
i posle uliva ribnjaka
UP DOWN
Temperatura (⁰C) 9.7±0.1 11.66±1.62
Rastvoreni kiseonik (mg/l) 8.42±0.77 7.87±0.5
Saturacija (%) 79.93±3.09 75.76±5.87
pH 7.05±0.5 7.06±0.4
Elektroprovodljivost (µS/cm) 568±12.28 603±25.51
BPK5 (mg/l) 5.25±0.2 5.69±0.8
N-NO3 1.34±0.05 0.64±0.2
N-NH4 0.48±0.03 0.92±1.2
P-PO4 0.08±0.01 0.49±0.06
Sa ciljem pronalaženja grupe fiziko-hemijskih parametara vode koji na najbolji način
objašnjavaju multidimenzionalnu strukturu zajednice, BEST analiza je pokazala da na
zajednicu makroinvertebrata u najvećem stepenu utiče kombinacija uticaja temperature,
koncetracije azota iz nitrata, koncetracije azota iz amonijaka i fosfora iz ortofosfata i to sa
statistički značajnim koeficijentom Spearman-ove korelacije (BioEnv, r = 0.700, p = 0.03).

21
4 Diskusija
4.1 Analiza taksonomskog sastava zajednice makrozoobentosa
Organizmi makrozoobentosa imaju ključnu ekološku ulogu u mnogim slatkovodnim
ekosistemima (Boulton, 2003), a istovremeno grade najrasprostranjenije i najraznovrsnije
slatkovodne zajednice (Voelz i McArthur, 2000; Strayer, 2006). Unutar ukupnog diverziteta
zajednica ovih organizama svakako dominiraju akvatični insekti, koji čine između 70 i 90%
zajednice i predstavljaju jednu od najbitnijih komponenti bentocenoze tekućica (Cushing i
Allan, 2001). Razlog ovako visokog učešća leži u činjenici da su akvatični insekti
(dominantno njihovi larvalni stadijumi), razvili čitav niz adaptacija koje im omogućavaju da
zauzmu sve raspoložive niše (Cushing i Allan, 2001).
U ovom istraživanju Pasjačke reke, tokom 2018. godine, najveću zastupljenost po
broju rodova ima red Trichoptera, dok su ostali redovi zabeleženi sa manjim brojem rodova.
Po sezonama, najveći broj taksona registrovan je u proleće (27) gde je red Trichoptera
zabeležen sa čak 8 rodova, zatim sledi zima (22) i jesen (21).
Predstavnici reda Trichoptera sreću se na različitim podlogama: peskovitoj,
šljunkovitoj, kamenitoj, između vodenih biljaka, kao i između nataloženih biljnih ostataka.
Od brojnih rodova zabeleženih u ovom istraživanju najčešće se javlja rod Goera koji je
zabeležen u svim sezonama. Na veliki diverzitet Trichoptera u izvorima ukazuju i drugi
autori (Čepić i Marinković-Gospodnetić, 1978; Anderson i Anderson, 1995; Stojanović,
2017; Marković, 1998). Mada treba pomenuti da u uslovima održanja dobrog kvaliteta vode
u brdsko-planinskim potocima i rekama nizvodno od izvora raznovrsnost Trichoptera raste
(Filipović, 1969; Marković i sar., 1996).
U istraživanom izvoru Mollusca su zastupljeni sa dve klase: Gastropoda i Bivalvia.
Gastropoda su većeg diverziteta, tako da su od 7 determinisanih oblika mekušaca 5 iz klase
puževa, a samo 2 iz klase školjki. Od 7 rodova 3 su zabeleženi u svim sezonama na svim
uzorkovanim lokalitetima i to 2 iz klase puževa: Lymnaea i Planorbis, i 1 iz klase školjki:
Sphaerium. Rod Lymnaea je zebeleženna svega 3 izvora zapadne Srbije na razlićitim
podlogama (Marković, 1998). Rod Psidium često nastanjuje izvore. Naseljava mekanu
podlogu, pre svega muljevitu. Može se naći i na kamenitom dnu i to tamo gde se između

22
kamenja i šljunka nataloži mulj. Tokom proučavanja brdsko-planinskih područja Srbije
(Marković, 1998) nalažen je do 1175 metara nadmorske visine. Stojanović (2017) je
zabeležila rod Psidium tokom svog istraživanja kao jedinog predstavnika klase Bivalvia.
Razdeo Annelida je zastupljen sa dve klase: Oligochaeta i Hirudinea. Oligochaeta
nisu od velikog značaja za formiranje bentocenoza izvora. S obzirom da većina akvatičnih
vrsta za ishranu koristi detritus, uglavnom nastanjuju mekanu muljevitu podlogu. S obzirom
na način zivota i optimalne uslove za život maločekinjastih glista u mirnijim vodama,
logično je njihovo češće prisustvo u limnokrenim i holokrenim nego u reokrenim izvorima.
Oligochaeta su zastupljenesa 2 roda: Lumbriculus i Haplotaxis. Ove rodove su zabeležili i
Marković (1998) i Stojanović (2017). Što se tiče pijavica one su zabeležene sa 3 roda:
Erpobdella, Haemopis i Glossiphonia. Hirudinea su najčešće nalažene na izvorima koji služe
za napajanje stoke. Mogu se naći na različitim podlogama, a vrlo često i u slobodnoj vodi.
U izvorima brdsko-planinskih područja Srbije Marković (1998) je takođe zabeležio ova tri
roda, dok je Stojanović (2017) zabeležila samo rod Erpobdella.
Klasa Crustacea je zastupljena samo sa jednim rodom: Gammarus. Rod Gammarus
je prisutan u svim sezonama i na svim ispitivanim lokalitetima, takođe je i najbrojniji rod u
odnosu na ostale rodove (6855 jedinki je izdvojeno u toku istraživanja). Prisustvo roda
Gammarus su zabeležili i Marković (1998), Stojanović (2017), Filipović (1968), Filipović
(1969). Gamarusi nastanjuju mirnije i pliće delove izvorskog biotopa. Najčešće su uz obale,
mada se često nalaze i ispod kamenja, u busenju mahovina i u nataloženom lišću. Na mestima
brze vodene struje jačih izvora – vrela nastanjuju reikofilne mahovine, gde i pored toga što
nemaju nikakve prilagođenosti na opstanak u brzoj vodi, opstaju skrivene u busenje i
zaštićenje od odnošenja vodenom strujom. I pored toga što ih u velikom broju nalazimo u
čistoj vodi planinskih izvora i tekućica, nisu izrazito osetljivi na organsko opterećenje,
ukoliko je voda bogata kiseonikom. Može se reći da povećana količina nekih oblika
organskog opterećenja (biljnih delova, prisustvo uginulih vodenih životinja, pre svega
pripadnika familije Gammaridae) uz dobru aeraciju pospešuje njihov razvoj tako da u
ovakvim uslovima postižu vrlo veliku brojnost. Omnivori su, za ishranu koriste detritus,
suvo i sveže lišće, ali i sveže životinjske leševe, pre svega uginule mlade oblike svoje vrste.
Red Diptera je zabeležen sa 7 rodova gde je jedino rod Tabanus prisutan na svim
lokalitetima u svim sezonama. Pored roda Tabanus najviše su zabeleženi i rodovi:
Chironomus, Dixa i Tipula. Najveći broj oblika Diptera nalazi se na muljevitoj podlozi, kao
i tamo gde se nalaze biljni ostaci. Nalaženi su u izvorima sa čistom vodom, kao i onima gde

23
je konstantovano izrazito prisustvo organskih materija, što govori o širokoj ekološkoj valenci
ove grupe. Pojedini oblici poput roda Simulium za podlogu često biraju i kamenito dno.
Ukoliko se ovo istrazivanje uporedi sa istraživanjem Marković-a (1998), Anderson i
Anderon-a (1995) i Stojanović (2017) uočavaju se slični rezultati u pogledu raznovrsnosti
Diptera. Naime, na svim ispitivanim izvorima Diptera su po diverzitetu prednjačile.
Međutim i pored velike raznovrsnosti, u pogledu kvantitativnog učešća nisu od bitnijeg
značaja za izgradnju bentocenoza izvora. Po značaju za izgradnju zajednica
makroinvertebrata, kao i po frekvenciji javljanja u različitim izvorima, treba izdvojiti
predstavnike rodova: Tipula, Chironomus, Tabanus i Dixa.
Red Coleoptera je zabeležen samo jednim rodom u jesen na izvoru i to rod Gyrina.
Za razliku od ostalih vodenih insekata koji deo života provedu u vodi, vodeni tvrdokrilci
čitav svoj životni ciklus provedu u vodenoj srednini. Sklonište uglavnom nalaze između
kamenja, u udubljenjima na donjoj površini kamenja, kao i u vodenim mahovinama i drugim
vodenim biljkama.
Red Megaloptera je zastupljen rodom Sialis. Oblici Megaloptera uglavnom
nastanjuju muljevito-šljunkovitu podlogu, mada se mogu sresti i ispod kamenja. Ovaj rod je
takođe zabeležen od strane Marković-a (1998) i Stojanović (2017).
Red Plecoptera je zastupljen sa 2 roda: Nemoura i Leuctra. Plecoptera su organizmi
čistih brdsko-planinskih tekućica. U odnosu na izvore u tekućicama se znatno češće sreću, a
i brojnost i raznovrsnost im je veća. Ove konstatacije potvrđuju i rezultati istraživanja drugih
autora (Filipović, 1968; Čepić i Marinković-Gospodnetić, 1978; Anderson i Anderson,
1995). Plecoptera uglavnom naseljavaju kamenito-šljunkovitu podlogu izvora, mada se
jedinke roda Nemoura često nalaze u opalom lišću i drugim biljnim ostacima. Ovi rodovi su
takođe opisani od strane Marković-a (1998) i Stojanović (2017).
Red Ephemeroptera je prisutan sa 2 roda: Baetis i Heptagenia. Oba roda su
pronađena i kod Marković-a (1998) u izvorima brdsko-planinskih područja Srbije.
Ephemeroptera su veoma stara grupa koja potiče još iz mlađeg paleozoika. U pomenutom
istraživanju je prikazano da zastupljenost Ephemeroptera raste od izvora ka zoni gornjeg i
srednjeg toka (gde je najveća), dok se nizvodno u zoni donjeg toka smanjuje. Ovo je takođe
zabeležio i Filipović (1976). Samim tim može se zaključiti da efemeroptere nisu specifičan
činioc bentocenoza izvora što potvrđuju i istraživanja domaćih i stranih autora (Ikonomov,
1959; Filipović, 1968; Matanočkin i sar., 1969; Harrison i Elsworth, 1958, Busnita i sar.,

24
1970; Anderson i Anderson, 1995). Imajući u vidu činjenicu da su izvori u pogledu
koncentracije kiseonika siromašniji u odnosu na vode brdsko-planinskih tekućica, svakako
se u kompleksu ograničavajućih faktora većeg prisustva Ephemeroptera u izvorima nalazi i
količina rastvorenog kiseonika. Ephemeroptere nastanjuju pre svega kamenito-šljunkovitu
podlogu, mada ima i onih koje nastanjuju mahovine, nataloženo lišće ili se zarivaju u meku
muljevitu ili peščanu podlogu.
4.2 Uticaj pastrmskih ribnjaka na zajednicu makrozoobentosa i fizičko-
hemijske parametare vode
Na osnovu rezultata nMDS analize uočavaju se 3 grupe uzoraka na osnovu
kvalitativnog i kvantitativnog sastava makrozoobentosa sa sličnošću od 55% (Slika 10). U
prvoj grupi se vidi da su najsličniji uzorci sakupljeni iznad ribnjaka tokom jeseni i zime kako
po kvalitativnom tako i po kvantitativnom sastavu, dok se uzorak sakupljen tokom proleća
izdvoljio u zasebnu grupu i on ne pokazuje sličnost ni sa jednim uzorkom. Na kraju u trećoj
grupi su se svrstali svi uzorci koji su sakupljeni ispod uliva ribnjaka, što nam govori da su ti
uzorci različiti u odnosu na prve dve grupe po kvalitativnom i kvantitativnom sastavu
makrozoobentosa. Ovakav rezultat potvrđuje inicijalnu pretpostavku da se zajednice
maktrozoobentosa iznad i ispod ispusta ribljaka razlikuju, najverovatnije kao posledica
promene kvaliteta vode. Ovaj zaključak je moguće potkrepiti činjenicom da je nMDS analiza
ukazala na dve grupe sličnih uzoraka na osnovu sličnosti u vrednostima fizičko-hemijskih
parametara (Slika 11), gde su se uzorci iznad ribnjaka jasno odvojili od uzoraka ispod uliva.
Štaviše, Mann Whitny test je pružio preciznu informaciju koji se fizičko-hemijski parametri
vode statistički značajno razlikuju (Tabela2).
Statistički značajno povećanje temperature vode zabeleženo je ispod uliva ribnjaka
u odnosu na izvor gde je bila hladnija za par stepeni, najverovatnije kao posledica povećane
koncentracije organske materije u vodi nakon oslobađanja otpadne vode iz ribnjaka u
vodotok. Elektroprovodljivost je takođe porasla kao i temperatura (na izvoru je manja, a na
ispustu veća), što nam govori da je koncentracija rastvorenih soli viša na ispustu.
Elektroprovodljivost je u direktnoj vezi sa koncentracijom jona u vodi i temperaturom vode,
tako da kako raste temperatura vode i koncentracija jona tako raste i elektroprovodljivost.
Prema rezultatima ranijih studija bilo je očekivano smanjenje koncentracije kiseonika u vodi

25
(Stojanović, 2017). U ovoj studiji, koncentracija rastvorenog kiseonika jeste manja posle
uliva otpadne vode ribnjaka ali ne dovoljno da bi promena bila statstički značajna (Tabela
2). Što se tiče koncentracije azota iz nitrata ona se značajno smanjila na ispustu u odnosu na
izvor, a za razliku od nje koncentracija azota iz amonijaka se statistički značajno povećala
na ispustu u odnosu na izvor. Štaviše, očekivano je da koncentracija amonijaka bude
drastično uvećana posle ispusta ribnjaka (Stojanović, 2017; Milošević, 2018). Takođe,
mnogi autori uočavaju i porast koncentracije fosfora na lokalitetima nizvodno od ispusta
ribnjaka (Carr i Goulder, 1990a, b; Foy i Rosell, 1991; Oberdorff i Porcher, 1994; Adamek
i Sukop, 1996; Boaventura i sar., 1997; Pulatsu i sar., 2004; Bartoli i sar., 2007; Camargo i
Gonzalo, 2007; Camargo i sar., 2011; Guilpart i sar., 2012). U ovom istraživanju registrovan
je porast ukupne koncentracije fosfora iz ortofosfata na ispustu iz ribnjaka. Obzirom da se
na ribnjaku voda ispusta direkto ispušta u recipijent bez prethodnog taloženja, očekivane su
veće koncetracije fosfora iz ortofosfata na lokalitetima nizvodno od ribnjaka.

26
5 Zaključak
Tokom ovog istraživanja na izvoru Kladenac i na ispustu iz pastrmskog ribnjaka
analizirani su uticaji pastrmskog ribnjaka na sastav zajednice makrozoobentosa kao i fizičko-
hemijske parametare vode pre i posle ribnjaka. Tokom istraživanja utvrđeno je sledeće:
Na osnovu prikupljenog materijala iz ispitivanog izvora identifikovano je 37 rodova
vodenih makrobeskičmenjaka. Najraznovrsnija grupa su insekti a među njima se
izdvaljaju Trichoptera i Diptera kao najbrojnije (Trichoptera 10 rodova, Diptera 7
rodova).
Rod Gammarus je najzastupljeniji sa 62% od ukupnog broja individua po rodovima
na izvoru i čak 85% na ispustu u toku svih sezona istraživanja.
Rezultati nMDS analize pokazali su da se na osnovu kvalitativnog i kvantitativnog
sastava makrozonbentosa mogu izvojiti 3 grupe uzoraka sa sličnošću od 55%. U
prvoj grupi su se izdvojili uzorci sakupljeni iznad ribnjaka tokom jeseni i zime (UP
A i UP W), dok se uzorak sakupljen iznad ribnjaka tokom proleća (UP S) izdvojio u
drugu grupu. Konačno, svi uzorci sakupljeni ispod uliva ribnjaka (DOWN S, DOWN
A i DOWN W) svrstani su u treću grupu.
U pogledu kvaliteta vode, klasifikaciona analiza je izdvojila dve grupe sličnog
sastava lokaliteta. U okviru prve grupe izdvojili su se svi uzorci sakupljeni iznad
ribnjaka tokom proleća, jeseni i zime (UP S, UP A i UP W), dok su se u okviru druge
grupe izdvojili svi uzorci skupljeni ispod uliva ribnjaka (DOWN S, DOWN A i
DOWN W).
BEST analiza je pokazala da na zajednicu makroinvertebrata u najvećem stepenu
utiče kombinacija uticaja temperarure, koncetracije azota iz nitrata, koncetracije
azota iz amonijaka i fosfora iz ortofosfata i to sa statistički značajnim koeficijentom
Spearman-ove korelacije (BioEnv, r = 0.700, p = 0.03).

27
Literatura
1. Adamek, Z., Sukop, I. (1996). The impacts of trout farm discharges on benthic
community structure in a small karstic stream. Acta Universitatis Carolinae
Biologica, 40, 3-17.
2. Anderson, M.T., Anderson, H.N., (1995). The insect fauna of spring habitats in
semiarid rangelands in Centrol Oregon. Journal of the Kansas entomologicas society,
68 (2) supl.: 65-76.
3. Atanacković, A. (2015). Akvatične oligohete (Annelida, Clitellata, Oligochaeta) u
različitim tipovima tekućih voda u Srbiji. Doktorska disertacija. Biološki fakultet.
Univerzitet u Beogradu.
4. Baldini, I., Gaiter, S. (1990). Possibili metodologie di valuta zione della vulnerabilita
delle acque sorgive e proposte preliminari per la formulazione di gradienti di rischio
igienico. RivistaI taliana Igiene, 5–6.
5. Bartoli, M., Nizzoli D., Longhi D., Laini A., Viaroli P. (2007). Impact of a trout farm
on the water quality of an Apennine creek from daily budgets of nutrients. – Chem.
Ecol. 23: 1–11.
6. Boaventura, R., Pedro A. M., Coimbra J., Lencastre E. (1997). Trout farm effluents:
characterization and impact on the receiving streams. – Environ. Pollut. 95: 379–387.
7. Bodon, M., Gaiter S. (1989). Considerazioni sulpopolamento macrobentonico delle
acque sorgive. Biologi aAmbientale 2: 5–12.
8. Bonada, N., Rieradevall, M., Prat, N., Resh, V. H. (2006). Benthic macroinvertebrate
assemblages and macrohabitat connectivity in mediterranean-climate streams of
northern California. Journal of the North American Benthological Society 25:32–43.
9. Boulton, A. J. (2003). Parallels and contrasts in the effects of drought on
streammacroinvertebrate assemblages. Freshwater Biology, 48, 1173–1185.
10. Brehm, J. (1986). Der pH-Wert von Buntsandsteinquellen und Bachen. II. Der
Waldquell-pH-Wert nach der Hohenlage der Wasseraustritte und der
Hangneigungsrichtung der Quelleneinzugsgebiete im Knullgebirge. Beitrage
Naturkunde Osthessen 21: 53–66.
11. Brehm, J., M. P. D. Meijering, (1996). Fließgewasserkunde. Einfuhrung in die
Limnologie der Quellen, Bache und Flusse. Quelle & Meyer, Heidelberg, 302 pp.

28
12. Busnita, T., Brezeanu, G., Oltean, M., Popescu-Marinescu, V., Prunescu-Arion, E.,
(1970). Monografiazoneiportilor de fier, Studiulhidrobiologic al Dunariisi al
afluentilursai. EdituraAcademieRebpubliciiSocialiste Romania.
13. Camargo, J. A. (1992a). Structural and trophic alterations in macrobenthic
communities downstream from a fish farm outlet. – Hydrobiologia 242: 41–49.
14. Camargo, J. A. (1992b). Temporal and spatial variations in dominance, diversity and
biotic indices along a limestone stream receiving a trout farm effluent. – Water, Air,
Soil Pollut. 63: 343–359.
15. Camargo, J. A., Gonzalo, C. (2007). Physicochemical and biological changes
downstream from a trout farm outlet: comparing 1986 and 2006 sampling surveys.
Limnetica, 26(2), 405-414.
16. Camargo, J. A., Gonzalo, C., Alonso, Á. (2011). Assessing trout farm pollution by
biological metrics and indices based on aquatic macrophytes and benthic
macroinvertebrates: A case study. Ecological Indicators, 11(3), 911-917.
17. Cantonati, M., F. Corradini, L. Tagnin, (1998). Inquadramentofisico-chimico e
morfologico. In Le sorgentidel Parco Adamello-Brenta (M. Cantonati ed.), Parco
AdamelloBrentaDocumenti 11: 9–54.
18. Carr, O. J., Goulder, R. (1990a). Fish-farm effluents in rivers—I. Effects on bacterial
populations and alkaline phosphatase activity. Water Research, 24(5), 631-638.
19. Carr, O. J., Goulder, R. (1990b). Fish-farm effluents in rivers—II. Effects on
inorganic nutrients, algae and the macrophyteRanunculuspenicillatus. Water
Research, 24(5), 639-647.
20. Carter, J. L., Resh, V. H., Rosenberg, D. M., Reynolds, T. B. (2006). Biomonitoring
in North American rivers: a comparison of methods used for benthic
macroinvertebrates in Canada and the United States. Pages 203–228 in G. Ziglio, M.
Siligardi, and G. Flaim (editors). Biological monitoring of rivers. John Wiley and
Sons, West Sussex, UK
21. Čepić, V., Marinković-Gospodnetić, M., (1978). Zoobentos rijeke Toplice.
Godišnjak Biološkog instituta Univerziteta u Sarajevu, Vol XXXI-1978: 25-32.
22. Cushing, C. E., Allan, D. J. (2001). Streams: their ecology and life. San Diego,
California, USA: Academic Press.
23. Filipović, D., (1968). Limnološka karakteristika izvorskog regiona Lisinskog potoka
na Kopaoniku. Zbornik radova 11, 24: 325-337.

29
24. Filipović, D., (1969). Faunistički sastav izvorskog regiona Crnog Timoka i njegove
karakteristike. III Kongres biologov Jugoslavije, Knjiga plenarnih referatov in
pouzetkov, Ljubljana, 104.
25. Filipović, D., (1976). Istorija tproučavanja Ephemeroptera (Insecta) u našoj zemlji i
rezultati dosadašnjih ispitivanja u Srbiji. Arhiv bioloških nauka, 28 (1-2): 95-101.
26. Foy, R. H., R. Rosell, (1991). Fractionation of phosphorus and nitrogen loadings
from a Northern Ireland fish farm. – Aquaculture 96: 31–42.
27. Guilpart, A., Roussel, J.M., Aubin, J., Caquet, T., Marle, M., Le Bris, H. (2012). The
use of benthic invertebrate community and water quality analyses to assess
ecological consequences of fish farm effluents in rivers. Ecological Indicators, 23,
356-365.
28. Harrison, A.D., Elsworth, J.F., (1958). Hydrobiological studies on the Great Berg
river. Transaction of the royal sociaty of south Africa, Vol XXXV, Part III, Cape
Town.
29. Haseke, H., E. Pro¨ ll, (2006). Das Berchtesgadener Quellwasser und seine Herkunft.
Geologische, physikalisch-chemische und mikrobiologische Parameter. In Gerecke,
R. & H. Franz (eds.), Quellen im National park Berchtesgaden. National park
Berchtesgaden, Forschungsbericht 51.
30. Hellawell, J. M. (1986). Biological indicators of freshwater pollution and
environmental management. Applied Science Publishers, London, 546 pp.
31. Ikonomov, P., (1959). Ephemeroptera na Makedonija. Sistematika i faunistika.
Doktorska disertacija. PMF na Univerzitet Tetovo Skopje.
32. Illies, J., Botosaneanu, L. (1963). Problems, methodes de la classification et de la
zonation ecologique des eauxcourantes, considerees, surtout du point de
vuefaunistique, Mitteilungen der inernational envereinigung fur Theoretische,
Angewandte Limnologie 12: 1-57.
33. Kerovec, M. (1986). Priručnik za upoznavanje beskralješnjaka naših potoka i rijeka.
SNL, Zagreb, Jugoslavija.
34. Liao, P. B., (1970). Pollution potential of salmonid fish hatcheries. – Water and
Sewage Works 117: 291–297.
35. Loch, D. D., J. L. West, D. G. Perlmutter, (1996). The effect of trout farm effluent
on the taxa richness of benthic macroinvertebrates. – Aquaculture 147: 37–55.

30
36. Maire, R. & S. Pomel, 1994. Karst geomorphology and environment. In Gibert, J.,
D.L. Danielopol & J. Stanford, Groundwater Ecology. Academic Press, London,
130–157.
37. Marković, V. (2014). Morfološka varijabilnost i distribucija roda Theodoxus
Montfort, 1819 (Neritomorpha, Gastropoda) u centralnom delu Balkanskog
poluostrva i najužnom obodu Panonske nizije. Doktorska disertacija, Biološki
fakultet, Univerzitet u Begradu.
38. Marković, Z. (1998). Izvori brdsko-planinskih podruja Srbije: ekološka studija
makrozoobentosa. Univerziitet u Beogradu. Biološki fakultet. Beograd.
39. Marković, Z., Mitrović-Tutundžić, V. (2003). Gajenje riba. Zadužbina Andrejević,
Beograd. 137 str.
40. Marković, Z., Mitrović-Tutundžić, V., Savić, I., Ranđelović, N., (1996). Analiza
prelaska izvorske u faunu potoka na primeru vrela reke Banje. Kongres ekologa
Jugoslavije, Zbornik sažetaka, 25, Beograd.
41. Marković, Z., Poleksić, V. (2007). Ribarstvo u Srbiji (Fishery in Serbia). Prof.dr
Zoran Marković, Beograd, 1-239.
42. Matonačkin, I., Pavletić, Z., Habdija, I., Stilinović, B., (1969). Prilog limnologiji
gornjeg toka rijeke Save. Ekologija, Vol 4, No1: 91-124.
43. Milošević, Đ., Stojanović, K., Đurđević, A., Marković, Z., Stojković Piperac, M.,
Živić, M., Živić, I., (2018). The response of chironomid taxonomy- and functional
trait-based metrics to fish farm effluent pollution in lotic systems. Environmental
Pollution 242: 1058-1066
44. Muñoz, M. J., (1989). Indice de calidad y toxicidad de agua para salmonidos. PhD
Thesis No. 77, National Institute for Agrarian Research (INIA), Spain, (in Spanish).
45. Oberdorff, T., J. P. Percher, (1994). An index of biotic integrity to assess biological
impacts of salmonid farm effluents on receiving waters. – Aquaculture 119: 219–
235.
46. Paunović, M. (2007). Struktura zajednica makroinvertebrata kao indikator tipova
tekućih voda Srbije. Doktorska disertacija. Biološki fakultet. Univerzitet u Begradu.
47. Popović, N. (2014). Ekološka analiza zajednica slatkovodnih makrobeskičmenjaka
tri tipa tekućih voda na području Beograda. Doktorska disertacija. Biološki fakultet.
Univerzitet u Begradu.
48. Psenner, R., J. Catalan, (1994). Chemical composition of lakes in crystalline basins:
a combination of atmospheric deposition, geologic background, biological activity

31
and human action. In Margalef, R. (ed.), Limnology Now: A Paradigm of Planetary
Problems. Elsevier Science B.V.: 255–314.
49. Pulatsu, S., Rad, F., G. Köksal, F. Aydin, C. K. Benli, A. Topcu, (2004). The impact
of rainbow trout farm effluents on water quality of Karasu stream, Turkey. – Turk. J.
Fish. Aquat. Sci. 4: 9–15.
50. Raković, M. (2015). Diverzitet mekušaca Dunava (1260-863,5rkm) i taksonomska
analiza rodova Planorbarius, Radix, Physella i Ferrissia (Pulmonata:
Basommatophora). Doktorska disertacija. Biološki fakultet. Univerzitet u Begradu.
51. Selong, J. H., Helfrich, L. A. (1998). Impacts of trout culture effluent of water quality
and biotic communities in Virginia headwater streams. – Prog. Fish. Cult. 60: 247–
262.
52. Simić, V., Simić, S. (2009). Ekologija kopnenih voda (Hidrobiologija I). – Prirodno-
matematički fakultet, Univerzitet u Kragujevcu; Biološki fakultet, Univerzitet u
Beogradu, Kragujevac; Beograd.
53. Stanford, J. A., J. V. Ward, B. K. Ellis, (1994). Ecology of the alluvial aquifers of
the flathead river, Montana (USA). In Gibert, J., D. L. Danielopol& J. Stanford,
Groundwater Ecology. Academic Press, London, 367–390.
54. Stojanović, K., (2017). Uticaj pastrmskih ribnjaka na zajednice makrozoobentosa
tekućica sa posebnim osvrtom na larve roda Baetis (Ephemeroptera, Insecta).
Doktorska disertacija. Biološki fakultet. Univerzitet u Beogradu.
55. Strayer, D. L. (2006). Challenges for freshwater invertebrate conservation. Journal
ofthe North American Benthological Society, 25(2), 271-287.
56. Tomović, J. (2015). Ekologija, biodiverzitet i konzervacija slatkovodnih školjki
familije Unionidae u Srbiji. Doktorska disertacija. Biološki fakultet. Univerzitet u
Beogradu.
57. Voelz, N. J., McArthur, V. J. (2000). An exploration of factors influencing loticinsect
species richness. Biodiversity & Conservation, 9(11), 1543-1570.
58. Weigand, E., (1998). Limnologisch-faunistiche Characterisierung von Karstquellen,
Quellbachen und unterirdischen Gewassernnach Choriotopen und biozonotischen
Gewasserregionen National parko. .Kalkalpen, Osterreich. Unpublished report. 173
pp.
59. Wilhelm, F., (1956). Physikalisch-chemische Untersuchungenan Quellen in den
Bayerischen Alpen und im Alpenvorland. Munchn. geogr. Hft. 10: 1–97.

32
60. Woodley, S. (1993). Monitoring and Measuring Ecosystem Integrity in Canadian
National Parks. S. Woodley, J. Kay and G. Francis (eds.), Ecological Integrity and
the Management of Ecosystems. St-Lucie Press, Florida. p. 155–176
61. Zorić, K. (2015). Invazivnost alohtonih vrsta makroinvertebrata i riba Dunava.
Doktorska disertacija, Biološki fakultet, Univerzitet u Beogradu.

ПРИРОДНO - MАТЕМАТИЧКИ ФАКУЛТЕТ
НИШ
КЉУЧНА ДОКУМЕНТАЦИЈСКА ИНФОРМАЦИЈА
Редни број, РБР:
Идентификациони број, ИБР:
Тип документације, ТД: Монографска
Тип записа, ТЗ: Текстуални / графички
Врста рада, ВР: Мастер рад
Аутор, АУ: Драган Вулић
Ментор, МН: Милица Стојковић Пиперац
Наслов рада, НР: Утицај пастрмског рибњака у селу Пасјач код Пирота на састав
заједнице макрозообентоса и физико-хемијске карактеристике воде
Језик публикације, ЈП: Српски
Језик извода, ЈИ: Енглески
Земља публиковања, ЗП: Р. Србија
Уже географско подручје, УГП: Р. Србија
Година, ГО: 2019
Издавач, ИЗ: ауторски репринт
Место и адреса, МА: Ниш, Вишеградска 33.
Физички опис рада, ФО: (поглавља/страна/ цитата/табела/слика/графика/прилога)
32 стр.; 5 подглавља; 11 слика; 2 табеле
Научна област, НО: биологија
Научна дисциплина, НД: екологија
Предметна одредница/Кључне речи, ПО: pastrmski ribnjaci, planinski izvor, zajednica makrozoobentosa,
kvalitativni i kvantitativni sastav, fiziko-hemijske karakteristike vode.
УДК 639.21:597.552.512+574.587:556.5
Чува се, ЧУ: библиотека
Важна напомена, ВН:
Извод, ИЗ: У овом раду испитиване су промене заједнице макрозообентоса, као
групе која се од свих водених организама најчешће користи у
биомониторингу текућих водених екосистема. Како би се анализирао
утицај пастрмских рибњака на организме макрозообентоса,
обављено је истраживање квалитативног и квантитативног састава
заједнице пре и после улива отпадних вода из рибњака. Такође,
поређене су и вредности физико-хемијских карактеристика воде на
контролним и тест локалитетима. Резултати овог рада показују да
пастрмски рибњаци лоцирани уз горње токове и изворишну зону
река представљају потенцијалну опасност за ремећење природних
станишта ове групе водених животиња.
Датум прихватања теме, ДП: 11.09.2019.
Датум одбране, ДО:
Чланови комисије, КО: Председник: Др Ана Савић, ванредни професор
Члан: Др Саша Станковић, доцент
Члан, ментор: Др Милица Стојковић Пиперац, доцент

ПРИРОДНО - МАТЕМАТИЧКИ ФАКУЛТЕТ
НИШ
KEY WORDS DOCUMENTATION
Accession number, ANO:
Identification number, INO:
Document type, DT: Monograph
Type of record, TR: Textual / graphic
Contents code, CC: Master thesis
Author, AU: Dragan Vulić
Mentor, MN: Milica Stojković Piperac
Title, TI: The influence of a trout fish farm in the village of Pasjač
near Pirot on the composition of the macrozoobenthos
community and the physico-chemical characteristics of
water Language of text, LT: Serbian
Language of abstract, LA: English
Country of publication, CP: Republic of Serbia
Locality of publication, LP: Serbia
Publication year, PY: 2019
Publisher, PB: author’s reprint
Publication place, PP: Niš, Višegradska 33.
Physical description, PD: 32 p. ; 5 chapters; 11 pictures; 2 tabels
Scientific field, SF: Biology
Scientific discipline, SD: Ecology
Subject/Key words, S/KW: trout ponds, mountain spring, macrozoobenthos community, qualitative and
quantitative composition, physico-chemical characteristics of water.
UC 639.21:597.552.512+574.587:556.5
Holding data, HD: library
Note, N:
Abstract, AB: This study examines changes in the macrozoobenthos community,
as the most commonly used group in biomonitoring of aquatic
ecosystems. In order to analyze the impact of trout fish farm on
macrozoobenthos, a qualitative and quantitative composition of the
community before and after fish farm effluent input. Also, the
values of physico-chemical characteristics of water at control and
test sites were compared. The results of this study indicate that
trout fish farms located at upstream river sections and river source
present a potential threat to the natural habitats of this group of
aquatic animals.
Accepted by the Scientific Board on, ASB: 11.09.2019.
Defended on, DE: Defended Board, DB: President: Dr Ana Savić, vanredni profesor
Member: Dr Saša Stanković, docent
Member, Mentor: Dr Milica Stojković Piperac, docent





























