Embed Size (px)
Citation preview

Role of liver in regulating
distribution and excretion of manganese1p2
P. S. PAPAVASILIOU, S. T. MILLER, AND G. C. COTZIAS Medical Research Center, Brookhaven National Laboratory, Upton, New York
PAPAVASILIOU, P. S., S. T. MILLER, AND G. C. COTZIAS. Role
of liver in regulating distribution and excretion of manganese. Am. J. Physiol. 2 I-I (I) : -2 I 1-2 16. I g66.-Rectal obstruction abol- ished the total-body loss of Mns4 in both manganese-loaded and jaundiced rats. Bile duct obstruction only diminished the excre- tion of the radioisotope, proving that several gastrointestinal routes excrete manganese. Excretion was exceedingly fast if the isotope was injected intraportally, but then the effect of bile ligation became enhanced. Furthermore, biliary ligation in- duced a rise followed by a decline in the concentration of Mns4 in the liver, whereas that of the natural metal (MnS5) increased. Ancillary experimentation linked these phenomena to absorp- tion and rapid reexcretion of the metal. This enterohepatic circulation participated in regulating the excretion of man- ganese : The customary acceleration of excretion following moderate loads with manganese was abolished by blocking the bile duct. Acceleration could be induced in bile-ligated animals provided they were overloaded with metal. The sum of the data indicated that bile formation constitutes the main regula- tory route under ordinary conditions, but, under conditions of overloading, auxiliary gastrointestinal routes participate.
isotopes; neutron activation analysis; homeostatic regulation; bile
S ELECTIVE EXCRETION by the kidney is a major mecha- nism by which the concentrations of many minerals are regulated in mammalian tissues, but significant excretion also occurs via the gastrointestinal tract. The latter route is especially important in the elimination of alkaline earths and metals of the periodic table’s first transition group, of which one, the essential element manganese, appears to be almost totally excreted into the gastro- intestinal lumen (8).
Mammalian liver and pancreas, both of which origi- nate from the primordial gut (I, I g), have been reported to excrete manganese (6, 13). It iS not known however, whether one or both of these.organs regulate the concen- tration of this metal in mammalian tissues, and if so, whether the gut participates in this regulation.
Received for publication 8 October 1965. 1 This work was supported by the U.S. Atomic Energy Com-
mission and, in part, by National Institutes of Health Grant OH 00159-03.
2 A preliminary report has been published (I I).
The accompanying papers (5, 15) indicated that the turnover of manganese depends extensively on the dietary supply of this metal, whereas the tissue concen- trations tend to remain stable primarily by virtue of ex- cretion of excess metal. The present studies examine the effect of obstruction of the rectum and of the bile flow on the distribution and excretion of this nutrient. The find- ings suggest the existence of a rapid enterohepatic circulation which regulates the excretion of manganese as one among several gastrointestinal routes.
METHODS
Animals. Male, albino Sprague Dawley rats, weighing 200-250 g, were observed for 1-2 weeks prior to experi- mentation. They were housed in steel cages, some of which permitted separate collection of urine and feces. Purina laboratory chow (32.5 E.cg Mn5S/g) and tap water were offered ad lib., except that some groups were fed milk containing approximately 36 rug Mn55/liter as de- termined by neutron activation analysis (I o, I 2, I 8). Manganese sulfate was added as noted. The results on 325 of the animals studied are reported.
Operations. Laparotomies were performed under ether anesthesia. In some groups, the common bile duct was ligated with 5-o silk at the juncture of the middle to the proximal third; in a few of these, double ligatures were applied also to the lower segment of the rectum. In another series, the liver was prevented from receiving MnE4++ by clamping the hepatic artery and the portal vein with a polyvinyl-tipped mosquito clamp for the IO min which followed the intravenous injection of the isotope (2 and Fig. 4). The control animals of this series received the Mnb4 after the release of the clamp, at which time the viscera had begun resuming their normal appearance.
All but two animals were decapitated by guillotine. Surgery seemed to affect the distribution of manganese and it was suspected that the agonal stages might do so as well. To vary the agonal stages one animal was killed by electrocution and one by oxygen deprivation.
Isotopes. The Mn56SO4. Hz0 (J. T. Baker Chemical Co.) was added to the milk and was given by stomach tube or injected intravenously in 0.1 ml 0.9 % NaCl solution. In one experiment this salt was first incubated
211
by 10.220.33.2 on May 4, 2017
http://ajplegacy.physiology.org/D
ownloaded from

212 PAPAVASILIOU, MILLER, AND COTZIAS
!J
3 .4 c P .3
- z
05 * A
.2
g .I
‘0 40 80 120 160 801 I I I I J 6420246
m g m (WET wt)LlVER cm .D . . 3 . . .
g si: 4 l l l l , , l l :ctF, l , l I, , ,I , 6420246 0 4 8 I2 I6 20 32 36 40
cm MINUTES
FIG. I. A : determination of Mnss in 8 samples of varying ,weight from the same rat liver. (Note that wet weights are reported in contrast to Figs. 5 and 6.) B: change of counting rate of tagged rat, when the midpoint of its trunk was moved parallel to the crystal. IOO% Represents the counting rate when the midpoint was over the center. C: same as B but for the kidney of this animal. D: change in total-body counting rate with passing time in a rat in- jected with Mn s4+f. The first point (I 00%) was determined at the completion of the injection of Mns4++.
with rat serum aerobically at 20 C for I hr and the serum was injected intravenously.
The Mn54C12 (T1iz- 3 14 days) was the carrier-free isotope used in earlier work (5, 15). Standardized doses of I .o-I .5 PC in o. I ml 0.9 % NaCl were injected.
The operated animals were given the radioisotope either into the vena cava or into the portal vein; the intact animals were injected into a leg or a tail vein. Ani- mals were excluded if the success of these steps was in doubt.
Neutron activation analysis. The methodology has been discussed earlier ( I o, I 2, I 8). The precautions in obtain- ing the samples and the reasons for not computing “spe- cific activities” of Mn54 are indicated in the preceding paper (15). Figure I, curve A, shows that the measure- ment of Mn56 was independent of sample size, the vari- ance of which had been suspected to affect the metal’s activation in the nuclear reactor.
Measurement of radioactivity. One-half hour or 2 hr after injection of the isotope each rat was placed in a Bollman cage (4) which held the midpoint of the animal’s trunk fixed 6 cm above the center of a flat NaI-TI crystal measuring I I x 8.8 cm. Figure I, curve B, shows the diminution in recorded radioactivity when the animal’s midpoint was moved from the center of this crystal. The crystal was connected to the circuitry discussed else- where (9). The total-body radioactivity and the Mnb4 of the tissues were counted in the manner stated in the second paper of this series (I 5). There was some de- pendence of the counts, recorded on the position of a , given organ in relation to the center of the crystal (Fig. I, curve C). Hence this center was utilized as much as possible for the measurement of all radioactivities. Because the various organs approximated a point source better than did intact animals, the total-body radio- activity was again (5) invariably lower by I O-I 7 % than the sum of its fractions recovered with isolated organs.
z 80
W > 70
3
? 60
5
s 50 2 L I- &
40
15 l I I I I I I I
0 1 2 3 4 5 6 7 TIME, DAYS
FIG. 2. Total-body retention curves of rats injected with Mns4++ as indicated.
By comparison to the position of animals or organs over the crystal, the changes in counting geometry which fol- lowed the distribution of the injected tracer in a given animal were minor indeed (Fig. I, curve D).
Typical /wotocoZ. A rat was weighed, anesthetized, and operated on according to one of the procedures discussed above. The tracer was then injected intravenously and the animal was placed over the crystal 20 min later. The first count served as the IOO % retention for the subse- quent ones and the decline of the radioactivity was plotted semilogarithmically as a function of time. After a period of observation, the animal was decapitated and autopsied: the liver, kidneys, thoracic and abdominal viscera, and the remainder of the carcass were dissected and counted separately on the apparatus used for the intact animal. I f the bile duct was ruptured, the animal was excluded from the study.
RESULTS
Abolition of the excretion of Mnb4 by rectal ligation. Nor- mally, Mn54 is excreted into the gastrointestinal tract (8), but this remained to be shown in jaundiced and in manganese-loaded animals. Groups of four rats with rectal obstruction were given Mn54 intravenously and manganous sulfate by stomach tube (3.2, 6.4, and I 2.8 mg Mn++/animal). The total-body radioactivitv remained unchanged during the subsequent 5 days of observation. A similar almost total retention of the iso- tope was observed in four animals after both the common bile duct and the rectum were ligated. Analyses of the urine of these latter animals and analyses of both urine and feces from three animals with biliary ligation showed that the urine contained almost no measurable radio- activity, whereas the fecal Mn54 accounted for the total- body loss.
by 10.220.33.2 on May 4, 2017
http://ajplegacy.physiology.org/D
ownloaded from

REGULATION OF MANGANESE EXCRETION BY LIVER *r3
IOL 1 1 11 Illq 1 1 II,lll, 1 1 l l llll l l , I ,lll , I I I Ili
0
I
&---- - - -
v-w -e- -
- - - ‘p--,-a
0 0
.ot L 1 l 1111111 l I I l lllll HOURS
I I I I I1111 I I I I lllll l I I l llll
.Ol .I I IO 100 1000 FIG. 4. Changes in the ratio of concentration of Mns4 in liver
and carcass of rats as a function of time. A clamp had been applied to the hepatic vessels of all animals. Intravenous injection of Mns4++ was given to the test animals before, and to the control animals after the release of the clamp.
4 In
2 0 I I I I
0 2 4 6 8 IO transport of Mn .54 from the tissues to the liver might assume the form of a wave when the tracer’s flow into
TIME 1 DAYS)
FIG. 3 animals
Comparison of the retention curves of h/ins4 in livers of ndicated. Vertical lines denote the total envelope of the i the bile was blocked. Hence a different method of block-
ing was tried: A single intravenous injection of Mns4 was given to each of 34 rats either before or after the applica- tion of a clamp to the hepatic blood vessels for the IO
min during which go % of the isotope is cleared (2 and Fig. 4). Five minutes later, the livers of the clamped ani- mals contained 2-3.5 % of the injected Mn54, whereas those of the corresponding controls contained 32 %. Eleven days later, the respective values were I 8 and 24 %. These results are summarized in Fig. 4 as liver-to-carcass ratios (14, I 5). The total-body retention of Mns4 was measured in the animals sacrificed on the 5th and I I th days (4 groups of 3 animals each). The animals that had received the isotope with the clamp on showed identical total-body retention curves of Mn54, as had those with biliary ligation (Fig. 2, curve C). Those that had re- ceived the isotope after the release of the clamp behaved identically to the sham-operated animals (Fig. 2, curve D). Evidently, some Mns4 was being excreted via extra- hepatic routes even when the bile duct was patent; transport from the tissues to the liver accounted only for the rise shown in Fig. 2, but not for the fall.
Diminished excretion of Mn54 following biliary ligation. The previous experiments confirmed the gastrointestinal ex- cretion of manganese. The contribution of the bile was studied thereafter by means of biliary ligation in animals receivin .g the isotope into either a peripheral or the portal vein. The total-body retention curves suing four groups of three animals are shown
of the en- Fig. 2. in
Biliary ligation diminished but did not abolish the excre- tion of this isotope whether or not it had been injected intravenously or intraportally. The two routes of injec- tion reflected themselves in the relative rates by which this isotope became excreted : intraportal injection was followed by more rapid excretion or by more marked retention, depending on whether the bile duct was open or shut. These experiments indicated that, although the liver was only one of the excretory routes, it excreted its isotope very rapidly, possibly because it received signifi- cant amounts of metal from the gut.
The diminished excretion of Mns4 following biliary ligation was expected to produce a rise of the isotope’s concentration in the liver. The unexpected findings and their explanation are dealt with below.
A second possibility was that cellul .ar damage (3) di- mini .shed the manganese uptake and thus induced the decline under investigation. This was tested in 18 bile- ligated, 13 sham-operated, and 6 control animals, all of which were given I PC Mn54 intravenously 2 hr prior to
Fifteen animals with biliary obstruction were dis- sected at various intervals in parallel with 2 control groups of 15 laparotomized and 15 intact animals. Figure 3 shows that the radioactivity rose sharply and then declined in the livers of obstructed animals. This differentiated the obstructed group from its two control groups. Surprisingly, the livers of all operated animals differed from the nonoperated ones. The effects of sur- gery were not restricted to the liver, since the kidneys of these animals showed a significantly higher rate of loss than did the nonoperated controls. The other tissues tested did not show striking differences among these groups.
- sacrifice. Sacrifice took place after the following time intervals: 2 hr (6 ligated, 6 sham operated, 6 controls) ; I day (6 ligated, 2 sham operated); 5 days (6 ligated, 5 sham operated). The percent uptake by the liver was computed for each animal. The data were paired on all permutations and subjected to statistical analysis (t test). No significant differences were found between groups: the ligated group as a whole showed liver up- takes with a mean and standard deviation of 26.5 =t 8 while the corresponding numbers for the other two groups were 29. I rt 7 and 21.4 rt 3, respectively. I. Now it became probable that manganese absorbed from the diet accumulated in the obstructed liver and
Several possible mechanisms could account for the striking rise and subsequent fall of the radioactivity in these livers. The first to be investigated was that the
by 10.220.33.2 on May 4, 2017
http://ajplegacy.physiology.org/D
ownloaded from
PAPAVASILIOU, MILLER, AND COTZIAS
F!i > i E 0 \
3 DAYS AFTER OPERATION
A ’ ’ ’ ’ ’ ’ ’ ’ ’ ’ + 0 I 2 3 4 5 6 7 8 9 IO
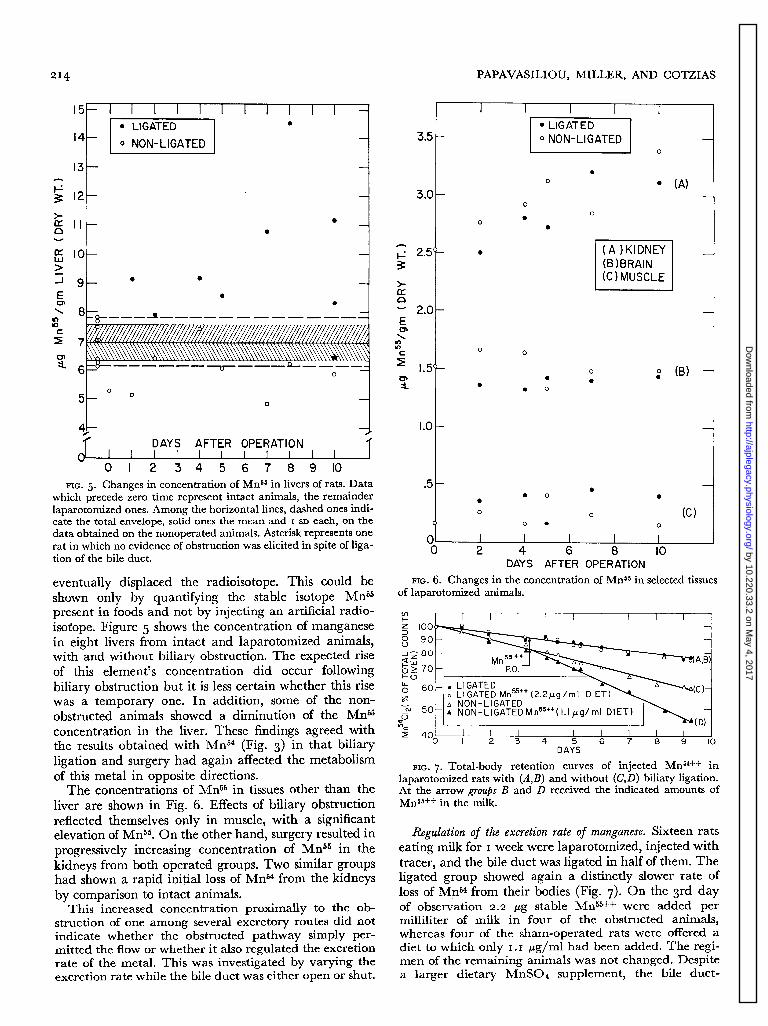
FIG. 5. Changes in concentration of Mns6 in livers of rats. Data which precede zero time represent intact animals, the remainder laparotomized ones. Among the horizontal lines, dashed ones indi- cate the total envelope, solid ones the mean and I SD each, on the data obtained on the nonoperated animals. Asterisk represents one rat in which no evidence of obstruction was elicited in spite of liga- tion of the bile duct.
eventually displaced the radioisotope. This could be shown only by quantifying the stable isotope MrP present in foods and not by injecting an artificial radio- isotope. Figure 5 shows the concentration of manganese in eight livers from intact and laparotomized animals, with and without biliary obstruction. The expected rise of this element’s concentration did occur following biliary obstruction but it is less certain whether this rise was a temporary one. In addition, some of the non- obstructed animals showed a diminution of the MrP concentration in the liver. These findings agreed with the results obtained with Mn54 (Fig. 3) in that biliary ligation and surgery had again affected the metabolism of this metal in opposite directions.
The concentrations of MrP in tissues other than the liver are shown in Fig. 6. Effects of biliary obstruction reflected themselves only in muscle, with a significant elevation of Mn 56. On the other hand, surgery resulted in progressively increasing concentration of Mnss in the kidneys from both operated- groups. Two similar groups had shown a rapid initial loss of Mnb4 from the kidneys by comparison to intact animals.
This increased concentration proximally to the ob- struction of one among several excretory routes did not indicate whether the obstructed pathway simply per- mitted the flow or whether it also regulated the excretion rate of the metal. This was investigated by varying the excretion rate while the bile duct was either open or shut.
2 2.5” 3
&
z 2.0- E ? z c = l.5CL T
1.0 -
.5 -
0
0
0 0 (A)
0
0 0
0 0
: (B)
0
0 (Cl
0 0 0
0' I I I I I 0 2 4 6 8 IO
DAYS AFTER OPERATION FIG. 6. Changes in the concentration of Mn66 in selected tissues
of laparotomized animals.
r r I I I I I I I I I I
LL 0 60 8 7J 50 u
x- 5 401 I I I I I I I I I I
0 I 2 3 4 DA&
6 7 8 9 10
FIG. 7. Total-body retention curves of injected Mn64++ in laparotomized rats with (A,B) and without (C,O) biliary ligation. At the arrow groups B and D received the indicated amounts of M#++ in the milk.
Regulation of the excretion rate of manganese. Sixteen rats eating milk for I week were laparotomized, injected with tracer, and the bile duct was ligated in half of them. The ligated group showed again a distinctly slower rate of loss of R/Ins4 from their bodies (Fig. 7). On the 3rd day of observation 2.2 pg stable Mn55++ were added per milliliter of milk in four of the obstructed animals, whereas four of the sham-operated rats were offered a diet to which only I. I pg/m.l had been added. The regi- men of the remaining animals was not changed. Despite a larger dietary MnSO4 supplement, the bile duct-
by 10.220.33.2 on May 4, 2017
http://ajplegacy.physiology.org/D
ownloaded from

REGULATION OF MANGANESE EXCRETION BY LIVER 215
TABLE I. Distribution of injected dose of Mn54 at end of experiment summarized in Fig. 7, as percent of initial dose given
Liver Viscera carcass Biliary obstruction 45 (44-48) 21 (20-22)
(milk) 22 (q-24)
Biliary obstruction 27 (26-q) 42 (41-43) (milk + MnSOJ
23 (q-26)
No biliary obstruction 27 (q-29) Ig (18-20) (milk)
23 (21-24)
No biliary obstruction (6-14) 20 9 (17-25) 23 (21-25) (milk + MnS04)
Values are means, with ranges in parentheses. Under viscera there is listed the sum of the determinations of the radioactivity in the gastrointestinal tract, kidneys, spleen, pancreas, testes, adrenals, and thoracic organs for each animal.
TABLE 2. Percents of total-body MnS4 remaining 5 days after injection as a function of single oral doses of manganese carrier
No. of NLnpf Md6 An-
Given, pg imals Mean Range imals Mean Range
Ligated Nonligated
Controls 11 *o-80.5 79-5 79 9 75.5 75 *o-76 -0 5x10 3 78.0 75 l 2-80.5 3 57.0 51.2-63.2 1x102 2 82.5 81.5-83.5 56.0 4g.5-63.2 3 2XI02 3 76.0 66.4-85 .o 47.2-58.6
3 52.7 4x10~ 3 86.0 78.6- 3 59.8 52.0-67.5
8x10~ 3 66.8 62 .o .o-72 3 40.5
26.2-56.0
1.6X103 6 54*2 46.0-63.0 6 20.8 19.5-22.1
46.0 3.2X103 1 3 21.5 ‘7*5-25.5 6.4X103 6 46.5 38-5-55.0 7 21.8 I I .5-32.5
I .28X104 5 32*5 28.2-37.0 6 16.5 9.2-24.0
0 I 2 3 4 5 6 7 8
FIG. 8. Total-body retention curves of injected MrP4+t in lap- arotomized rats with (A,B,C) and without (D,E,F) biliary obstruc- tion. At the arrow all animals were injected intravenously with the indicated amounts of Mns5++.
of as well as to parenterally injected carrier. manganese Administration of doses of graded metal, orally and parenterally, were preferable to fractionation of the Mn54 between feces and intestinal wall, since washings ex- tracted significant amounts of radioactivity.
The amount of administered carrier was orally varied as follows: Rats maintained on milk for I week were di- vided into sham-operated and ligated groups. These were injected iv with calibrated doses of Mn54 and were
obstructed animals showed no discernible increase in the
(Fig. 7h
rate of total-body loss of Mn54, whereas a marked en- hancement of the excretion was exhibited by the controls
This striking retention could have been a reflection of diminished dietary (manganese) intake rather than of excretory consequences of biliary ligation. Therefore evi- dence for isotopic exchange was sought in the distribution of the radioisotope among the organs of these rats
further divided into subgroups of by stomach tube I ml of water
total-body radioactivities for 5 days. These radioactivities are shown as percents of the six animals killed 2 hr after tagging, in Table 2. Biliary ligation resulted in slower excretion of Mn54 regardless of how much natural manganese was given.
The above experiments would have proved that the
MnS04 solution and were sacrificed after recording their
three. Each was or graded amou
given nts of
(Table I ). Again (2, 5), the carcasses of all these animals bile flow can be bypassed under conditions of heavy showed similar retention of this isotope regardless of manganese loading, if the following were true: that the jaundice or dietary manganese. However, the percent of isotope lost by these animals had been actually excreted, the Mn54 administered was smaller in the liver, after the rather than exchanged across their intestinal mucosas, manganous supplement was given. Biliary obstruction Hence, in the experiments shown in Fig. 8, the manga- again induced a relative increase of the radioactivity re- nous salt was injected intravenously to eliminate isotopic
exchange across the gut. maining in the liver, regardless of the manganous supple- ment. The pooled viscera showed the same percent retention of the administered Mn54 in all groups, except for the one that had both biliary obstruction and the manganous supplement : here the -viscera (including the gastrointestinal tract) contained the balance of the radioactivity lost by the livers.
These findings ruled out diminished food intake as the
The rate of loss of MnS4 by the animals with biliary ligation was similar to the corresponding ones on the milk diet (Fig. 7), whereas the sham-operated controls showed a faster rate of loss. The obstructed ones showed accelerated excretion of Mn54 only with doses higher than 50 pg Mn55* /animal, whereas the controls re- sponded to all doses given. When 200 pg Mn55++ were
only reason for the insensitivity of the Mn54 turnover to given intravenously with rat serum or saline as the manganese loading. Still, this insensitivity could have vehicle, the responses obtained were less pronounced been feigned by diminished mobility of the intestines of than those found in the corresponding animals receiving these jaundiced rats. I f this were the case, the ligated 400 pug Mn55++ (Fig. 8) but the vehicle induced no animals should remain insensitive to even larger amounts differences.
by 10.220.33.2 on May 4, 2017
http://ajplegacy.physiology.org/D
ownloaded from

216 PAPAVASILIOU, MILLER, AND COTZIAS
DISCUSSION
The first demonstration to emerge of this work was that the intestine and some of its tributaries constitute a system of multiple excretory routes for manganese. This system contrasted to the kidney which excreted negligible amounts of metal even during jaundice and after loading with manganese. The routes which did excrete this metal have as their common origin the primordial gut. This embryological consideration evoked interest in the rela- tive effectiveness of these routes, as discussed in a com- panion paper (2).
Additional findings pertained to several effects of surgery. These were generally opposite in direction to those induced by biliary obstruction. Hence, in the pres- ent experiments, the normal contribution of the liver to the excretion of manganese had been underrated because of the surgery employed. While attempting to explain these manifestations of stress, one might note the effects of administering cortisol or ACTH (I 5), since all were in the same direction.
The biphasic change of the concentration of Mns4 in obstructed livers became important after it led us to show the central role of the liver in the distribution of this metal: the initial rise indicated movement of manga- nese from the tissues to the liver, which was confirmed by excluding the liver from receiving initially the in- jected radioisotope. Transport in the opposite direction was shown by the evidence summarized in Table I. In addition, some of the absorbed metal must be normally
REFERENCES
I. AREY, L. B., Developmental Anatomy: A ‘Textbook and Laboratory Manual of Embryology (5th ed.). Philadelphia : Saunders, I 948, p. 226.
2. BERTINCHAMPS, A. J., S. T. MILLER, AND G. C. COTZIAS. In- terdependence of routes excreting manganese. Am. J. Physiol. 21 I : 217-224, 1966.
3. BIRNS, M., B. MASEK, AND 0. AUERBACH. The effects of ex- perimental acute biliary obstruction and release on the rat liver. Am. J. Pathol. 40: 95-I I I, 1962.
4. BOLLMAN, J. L. A cage which limits the activity of rats. J. Lab. Clin. Med. 33: 1348-1352, 1948.
5. BRITTON, A. A., AND G. C. COTZIAS. Dependence of manganese turnover on intake. Am. J. Physiol. 2 I I : 203-206, I 966.
6. BURNETT, W. T., JR., R. R. BIGELOW, A. W. KIMBALL, AND C. W. SHEPARD. Radiomanganese studies on the mouse, rat and pancreatic fistula dog. Am. J. Physiol. 168: 620-625, 1952.
7. COTZIAS, G. C., D. C. BORG, AND A. J. BERTINCHAMPS. Clini- cal experiences with manganese. In: Metal Binding in Medicine, edited by M. Seven. Philadelphia: Lippincott, I 960, p. 50.
8. COTZIAS, G. C., AND J. J. GREENOUGH. The high specificity of the manganese pathway through the body. J. Clin. Invest. 37: I 298-I 305, 1958.
g. COTZIAS, G. C., S. T. MILLER, AND J. EDWARDS. J. Lab. Clin. Med. Neutron activation analysis: the stability of manganese concentrations in human blood and serum. In press.
IO. COTZIAS, G. C., AND -Pi’ S. PAPAVASILIOU. State of binding of natural manganese in human cerebrospinal fluid, blood and plasma. Nature I 95 : 823-824, I 962.
I I. COTZIAS, G. C., AND P. S. PAPAVASILIOU. Primordial homeo-
transported promptly into the bile, in view of the decline of radioactivity and increased manganese concentration which followed biliary obstruction (Figs. 3 and 5).
Several intriguing observations pertained to the regu- lation of excretion: rectal obstruction abolished the ex- cretion, whereas biliary obstruction impaired primarily the sensitivity with which excretion was normally regu- lated. Furthermore, preventing the liver from receiving the initial bulk of tracer did not prevent the animals from excreting it, although their livers were accumulating instead of losing radioactivity. Consequently, altho wh these results proved the existence of more than one ex- cretory route (6, 13), they raised valid questions regard- ing the identity of the tissues drained by each.
The sum of these and some earlier findings (7) indi- cates that the absorbed manganese reaches the liver, becomes localized in its mitochondria (14, 16, I 7, 20),
and, although some metal becomes distributed to the tissues, a significant fraction is discharged into the bile. It remains to be seen to what extent this circulation is completed by reabsorption, whether its pump is lo- cated in the mitochondria and, if so, of which tissues. The latter is worth ascertaining since the liver was shown by these investigations to be instrumental in the homeo- stasis of this metal.
The authors thank Mr. Alexander R. Nicholson for securing data on distribution and turnover and for their statistical analysis, and Miss Judith Edwards for analysis of some of the tissues.
stasis in a mammal as shown by the control of manganese. Nature 201 : 828-829, 1964.
I 2. COTZIAS, G. C., P. S. PAPAVASILIOU, AND S. T. MILLER. Neu- tron activation analysis : clinical and biological studies of manganese. Colloq. Intern. Biol. Saclay, 3” p. 287-306, 1963.
13. GREENBERG, D. M., D. H. COPP, AND E. M. CUTHBERTSON. Studies in mineral metabolism with the aid of artificial radio- active isotopes. VII. The distribution and excretion, particu- larly by way of the bile, of iron, cobalt and manganese. J. Biol. Chem- I47 : 749-756, 19430
14. HUGHES, E. R., AND G. C. COTZIAS. Adrenocorticosteroid hor- mones and manganese metabolism. Am. J. Physiol. 201 : I 06 I - 1064, 1961.
15. HUGHES, E. R., S. T. MILLER, AND G. C. COTZIAS. Tissue con- centrations of manganese and adrenal function. Am. J. Physiol. 21 I : 207-210, 1966.
16. LINDBERG, O., AND L. ERNSTER. Manganese, a cofactor of oxi- dative phosphorylation. Nature I 73 : I 038-I 039, 1954.
17. MAYNARD, L. S., AND G. C. COTZIAS. The partition of man- ganese among organs and intracellular organelles of the rat. J. Biol. Chem. 214: 489-495, 1955.
18. PAPAVASILIOU, P. S., AND G. C. COTZIAS. Neutron activation analysis : the determination of manganese. J. Biol. Chem. 236 : 2365-2369, 1961.
19. PATTEN, B. M. Foundations of Embryology. New York : McGraw,
1958, P* 431* 20. ZETTERSTR~M, R., AND L. ERNSTER. Bilirubin, an uncoupler of
oxidation phosphorylation in isolated mitochondria. Nature 178: ‘335-1337, ‘956*
by 10.220.33.2 on May 4, 2017
http://ajplegacy.physiology.org/D
ownloaded from





























