Embed Size (px)
Citation preview

Behavioural Brain Research 99 (1999) 169–179
Research report
An electrophysiological characterization of ventral tegmental areadopaminergic neurons during differential pavlovian fear conditioning
in the awake rabbit
Fay A. Guarraci *, Bruce S. Kapp
Department of Psychology, Uni6ersity of Vermont, Burlington, VT 05405, USA
Received 10 October 1997; received in revised form 29 April 1998; accepted 22 May 1998
Abstract
Recent research has suggested that the mesencephalic dopaminergic (DA) system is activated by stress. For example, alterationsin DA metabolites have been found in the ventral tegmental area (VTA) following footshock and immobilization in the rat [15,37].Furthermore, this activation appears selective to DA neurons within the VTA since no changes were observed within thesubstantia nigra [15,16]. While this research suggests that DA neurons in the VTA are activated by aversive events, there has beena paucity of electrophysiological research designed to examine the sensory response characteristics of these DA neurons, and inparticular their response to stimuli which predict aversive events. The present study was conducted to investigate the responsecharacteristics of DA neurons within the VTA of the awake rabbit to acoustic stimuli which, via Pavlovian aversive conditioningprocedures, came to predict the occurrence of a mild shock to the pinna. 45% of the neurons meeting pre-established criteria forDA neurons demonstrated either significant excitation or inhibition to conditioned aversive stimuli. These neurons respondeddifferentially to CS+ and CS− presentations. Some of these neurons (65%) demonstrated a greater increase in activity duringthe CS+ compared to the CS− , some (22%) demonstrated a greater decrease in activity during the CS+ compared to the CS−and some (13%) demonstrated a greater increase in activity during the CS− compared to the CS+ . Further, conditioned heartrate responses in the rabbits occurred during the recording of a majority of these neurons. These overall results suggest thatconditioned aversive stimuli can affect the firing of VTA DA neurons and that these neurons comprise a heterogenous populationwith respect to their response profiles. © 1999 Elsevier Science B.V. All rights reserved.
Keywords: Ventral tegmental area; Dopamine; Pavlovian fear conditioning; Extracellular single cell recordings
1. Introduction
The mesencephalic dopaminergic (DA) system hasbeen identified as a crucial neural system mediating therewarding properties of various incentive stimuli, in-cluding food and water in deprived animals [2,21,49],sex [12,34], electrical brain stimulation [36] and drugs ofabuse (e.g. opiates and psychomotor stimulants[17,45,46]). Additional research has suggested that this
system is also activated by the stress of aversive events.This activation has been inferred from alterations inpostmortem or in vivo levels of dopamine and itsmetabolites. For example, increased DA release and/oraltered levels of DA metabolites have been reported innumerous brain areas following a variety of aversiveevents including footshock, immobilization, swimstress, and conditioned aversive stimuli. These areasinclude the ventral tegmental area (VTA) [15,16,25], themedial prefrontal cortex (mPFC) [1,9,18,22,27], the nu-cleus accumbens [1,9,18,22] and the amygdala [10,22].Contradictory results, however, have also been reported
* Corresponding author. Tel.: +1 802 6562670; Fax: +1 8026568786; E-mail: [email protected]
0166-4328/99/$ - see front matter © 1999 Elsevier Science B.V. All rights reserved.PII S0166-4328(98)00102-8

F.A. Guarraci, B.S. Kapp / Beha6ioural Brain Research 99 (1999) 169–179170
that demonstrate no evidence of altered dopamine re-lease or metabolism in each of these areas in responseto some of these aversive events [1,5,6,9,22,27,33,37].
These disparate results may be a function of thepotential problems inherent in the techniques used inthese studies to infer activation of DA neurons frommetabolic by-products. For example, while some DAmetabolites may be more accurate measures of do-pamine release [47], others may be more representativeof recently formed intracellular pools [41]. Further, theextent to which enhanced dopamine release ormetabolism is observed in response to a particularaversive event will depend upon the exact time thatneurochemical samples are collected. Indeed, Commis-siong [11] has argued that changes in DA metabolitesmay have been falsely equated with changes in DAneuronal activity, and therefore electrophysiologicalmonitoring of neuronal activity should be used toprovide converging evidence to compliment researchconducted using metabolic by-products.
While electrophysiological research has been directedat characterizing the response of DA neurons to re-warding stimuli [26,28,30,38–40], little research hasbeen directed towards characterizing their response toaversive events, and in particular conditioned stimulipredictive of aversive events. In one related study,Trulson and Preussler [44] characterized the response ofVTA DA neurons in the cat to a tone which was pairedwith a corneal airpuff. They reported that a majority ofthese neurons demonstrated enhanced activity duringtone presentations when compared to activity during aperiod of quiet waking. Although these data suggestthat these neurons are activated by a conditioned aver-sive stimulus, an important question remains unan-swered from this experiment. Since the activity of theseneurons was not examined in response to presentationsof a second tone of a different frequency that had notbeen paired with the airpuff, it is not possible todetermine the extent to which this activity simplyreflects the sensory responsiveness of these neuronsrather than a specific response to the conditioned aver-sive characteristics of the stimulus. This is an importantdistinction in light of the data demonstrating that mes-encephalic DA neurons are responsive to acoustic, vi-sual and somatosensory stimuli [7,19,29,42,43].
Due to the lack of definitive results of previousexperiments, the present study was conducted to char-acterize the response of individual DA neurons in theVTA of the rabbit to conditioned acoustic stimuli pre-dictive of an aversive event. A Pavlovian differentialfear conditioning paradigm was used to distinguishnonspecific neuronal responses to acoustic stimuli fromspecific responses to the aversive characteristics of con-ditioned acoustic stimuli.
2. Method
2.1. Animals
Eight experimentally naive female New Zealand rab-bits (Oryctolagus cuniculus) weighing from 2.3 to 2.6 kgat the start of the experiment were used. They weremaintained on a 12:12 h light/dark cycle and given foodand water ad libitum. Principles for the care and use oflaboratory animals in research, as outlined in the Guidefor the Care and Use of Laboratory Animals (NationalInstitutes of Health, 1985), were strictly followed. Allprocedures were approved by the University of Ver-mont Animal Care and Use Committee.
2.2. Surgical procedure
Following application of topical lidocaine (4%) to theregion of the marginal ear vein, each rabbit was anes-thetized with an injection of PromAce (acepromazine, 4mg/kg) followed by sodium pentobarbital (30–60 mg/kg, i.v.). Each rabbit was mounted into a Kopfstereotaxic instrument fitted with a rabbit head holder.Using aseptic surgical procedures, a midline incisionwas made over the skull and holes were drilled toprovide access to the brain for the recording of singleneurons and for the placement of an indifferent elec-trode. Additional holes were drilled into the skull forthe placement of five stainless steel anchor screws, oneof which served as a ground. Nylon bolts for headimmobilization were attached to the skull withcyanoacrylate cement. The indifferent electrode wascomprised of a 0.01 cm in diameter stainless steel wireinsulated except at the cross-sectional tip. The exposedtip was inserted approximately 0.5 mm into the neocor-tex at the following coordinates: 6.0 mm posterior tobregma, 3.0 mm lateral to the midline suture. For thepurpose of single neuronal recording, a stainless steelelectrode well, designed to accommodate a miniaturehydraulic microdrive (Trent Wells), was positioned overa 3.0 mm in diameter hole in the skull (5.5 mm poste-rior from bregma and centered on the midline) toprovide stereotaxic access to the VTA. The well wassecured in place with cyanoacrylate cement, swabbedwith antibiotic (Bacitracin) and sealed with a nylonplug. The exposed skull was then covered with dentalcement. Each rabbit was closely monitored followingsurgery until it regained postural control. Each rabbitwas monitored daily thereafter for a seven day postsur-gical recovery period. Behavioral experiments thencommenced.
2.3. Apparatus
During the experiment, each rabbit was placed in aPlexiglas restrainer with an adjustable headstock and

F.A. Guarraci, B.S. Kapp / Beha6ioural Brain Research 99 (1999) 169–179 171
padded backplate. The restrainer was placed within ashielded, ventilated and sound-attenuating recordingchamber (Industrial Acoustics) that was equipped witha 24 cm in diameter speaker situated 24 cm in front ofthe rabbit. A modified stereotaxic frame was used toimmobilize the rabbit’s head. Responses of single neu-rons were amplified by a high-impedance, differentialpreamplifier (World Precision, model DAM-6A), cou-pled with a main amplifier (Grass Instruments model7P511). The bandwidth was 0.1–10 kHz. Responsesfrom single neurons were isolated, fed into an audiomonitor and a window discriminator (Frederick Haer)and displayed on a storage oscilloscope (Tektonixmodel 5103N). Two independent output channels of aGrass S88 stimulator were used to administer the un-conditioned stimulus (US) to the pinna during behav-ioral conditioning and to produce anodal d.c. markerlesions (100 mA anodal d.c. for 5.0 s) at the end ofselected, daily recording sessions. Acoustic stimuli weregenerated with two separate voltage controlled genera-tors (Wavetek model 111). Neuronal responses, verbalcommentary, heart rate and event markers wererecorded by a multichannel FM tape recorder (A.R.Vetter model 420). The neuronal responses were fedinto a laboratory minicomputer (ABLE 40, New Eng-land Digital) for online data analysis. This computeralso controlled the presentation of stimuli during theconditioning procedure.
2.4. Beha6ioral training
Following surgical recovery, each rabbit was placedinto the restrainer within the recording chamber for six,daily, 1 h sessions, and the head was immobilizedduring the last two sessions by attachment of the nylonbolts to the modified stereotaxic frame. On the dayfollowing the last habituation session, a Pavlovian dif-ferential conditioning procedure was administered dur-ing which conditioned decelerative heart rate changes(i.e. bradycardia) were measured to demonstrate theability of the rabbit to learn that an acoustic stimuluspredicts the occurrence of an aversive event. The orient-ing phase of the procedure consisted of 18 pseudoran-dom presentations each of two tones of differingfrequency (500 and 1300 Hz, 80 dB, 5.0 s). They wereadministered so that no more than two consecutivepresentations of either tone was presented. A variable60 s intertrial interval was used. These trials werenecessary in order to habituate the bradycardic orient-ing response, a decrease in heart rate commonly ob-served in rabbits in response to the presentation of anovel stimulus. The conditioning phase then beganduring which 24 pseudorandom presentations of eachtone were administered with the offset of one tone(CS+ ) being coincident with the onset of the US (500ms, 1.0 mA, 60 Hz stimulus train applied to the pinna).
The second tone (CS− ) never was followed by the US.The US was administered via two stainless steel wireloops that pierced the pinna and were spaced approxi-mately 2.0 cm apart. They were inserted under locallidocaine anesthetic 24 h prior to behavioral condition-ing. The designated tone frequencies for the CS+ andCS− were counter-balanced among rabbits. To opti-mize conditioned discrimination between the CS+ andthe CS− , a second differential conditioning session,again consisting of 24 presentations each of the CS+and the CS− , was administered 24 h later. The secondday of behavioral conditioning was followed by dailyelectrophysiological recording sessions.
For the recording of heart rate, electrodes were posi-tioned by threading a stainless steel wire subcuta-neously using a sterile 21 gauge hypodermic needle. Thefree ends of the wire were then twisted together to forma loop to which the heart rate leads were attached. Theinsertion procedure for the heart rate electrodes pro-duces no movement of, nor discomfort to, the rabbit,and the electrodes were removed immediately after theexperimental session.
2.5. Electrophysiological recording procedure
Recordings from single neurons were made via pla-tinized, tungsten microelectrodes (4 MV tip impedance;Frederick Haer). The microelectrodes were advancedtoward the VTA using a hydraulic microdrive. To aidin identifying DA neurons, occasional novel acousticstimuli (e.g. brief white noise bursts) were presented tothe rabbit during neuronal recordings, since DA neu-rons have been reported to be responsive to novelacoustic stimuli [19,42,43]. Successive electrode penetra-tions took place on subsequent days and were spaced0.2–1.0 mm apart. The criteria for successful isolationof a putative DA neuron was as follows: (a) waveformconstancy during the observation period, (b) a 3:1signal-to-noise ratio to permit sufficient waveform am-plitude to enable reliable and unambiguous discrimina-tion, (c) a waveform duration of 2.0 ms or longer, (d) afiring frequency of between 1.0 and 12 Hz and (e) burstfiring with descending amplitude spikes [20,48].
Following isolation, an interspike interval histogramwas generated over a 30 s period in order to character-ize rates and patterns of spontaneous activity. Six pre-sentations of each of the CS+ and CS− were thenpresented in a pseudorandom sequence using a variable60 s intertrial interval. Each trial consisted of a 5.0 spre-CS baseline period followed by a 5.0 s CS period.Electrophysiological recordings were conducted duringan extinction phase (i.e. in the absence of US presenta-tions) to provide a more stable recording environmentfree of shock artifact. Additional blocks of conditioningtrials (six CS+ and six CS− ) in which the CS+ wasfollowed by the US were administered as needed to

F.A. Guarraci, B.S. Kapp / Beha6ioural Brain Research 99 (1999) 169–179172
maintain conditioned discrimination across recordingsessions. These trials were administered following thecompletion of recordings from an individual neuron.
2.6. Histology
Each rabbit was euthanized with an overdose ofsodium pentobarbital (Somlethal; 150 mg/kg, i.v.) andperfused with physiological saline followed by a 10%formol-saline solution. Seventy micron frozen sectionswere taken through the rostral-caudal extent of theVTA. Sections were stained with thionin to determinethe location of recording sites. The location of thesesites was determined microscopically by comparing mi-crodrive coordinates with the relative electrode tractpositions and marker lesions.
2.7. Data analysis
2.7.1. Heart rate conditioningTo determine the magnitude of the heart rate change
to presentations of each CS, the number of beatsoccurring during the 5.0 s CS period was comparedwith the number of beats during the 5.0 s pre-CSperiod. The difference was expressed as a percentchange from the pre-CS baseline period. Mean changesin heart rate during each daily session of 24 CS+ andCS− trials were calculated and data were evaluated oneach of two days of conditioning using a two-factor(CS-TYPE X DAYS) within-subjects analysis of vari-ance. During single unit recording sessions, the reliabil-ity of differential changes in heart rate during blocks ofeach of the CS+ and the CS− presentations wereevaluated using two-tailed t-tests for related measures.
2.7.2. Electrophysiological dataPeristimulus time histograms were generated for each
block of CS+ and CS− trials administered during therecording sessions. The number of spikes occurring inconsecutive 10, 100 ms, or 1.0 s bins of the 5.0 s pre-CSand CS periods were calculated. Neuronal responseswere analyzed in several ways. First, the mean andstandard deviation for the pre-CS baseline period werederived and standard scores were calculated and usedto detect significant changes in spike discharge rateduring stimulus presentations. Calculations were basedon 10 ms and 1.0 s bins, and on the entire 5.0 s CSperiod, depending on the response parameter in ques-tion, such as onset latency or response duration. Stan-dard scores exceeding 91.96 (95% confidence interval)were considered to represent a statistically significantchange in neuronal activity during a block of CS+ andCS− trials. Second, to statistically characterize thedifferential responsiveness of a neuron to CS+ versusCS− presentations, mean neuronal responses duringthe entire 5.0 s presentations were compared usingtwo-tailed t-tests for related measures.
A final analysis was used to determine the relation-ship between changes in neuronal firing rates andchanges in heart rate responses. The degree to whichthese two measures varied on a trial-by-trial basis wasdetermined by calculating the correlation between neu-ronal activity and heart rate changes to each CS presen-tation during all CS+ and CS− trials.
3. Results
3.1. Heart rate conditioning
Single unit activity was recorded from histologicallyverified VTA neurons in six rabbits. The electrodeplacements in the remaining two rabbits were anteriorto the VTA and these animals were excluded from thedata analysis. Of the six rabbits included in the dataanalysis, all developed reliable differential heart rateresponses to the CS+ and the CS− as shown in Fig.1. A two-factor (CS-TYPE X DAYS) within-subjectsanalysis of variance (ANOVA) revealed a significantCS-TYPE effect (F(1,5)=14.3, pB0.05) and a signifi-cant CS-TYPE X DAYS interaction (F(1,5)=14.77,pB0.01). The DAYS factor was not significant. Thesignificant CS-TYPE and CS-TYPE X DAYS interac-tion demonstrated that bradycardic heart rate responsesemerged to the CS+ , and that these responses werestatistically distinguishable from those that emerged tothe CS− . Further t-tests revealed that heart rate re-sponses during the CS+ were not significantly differ-ent from heart rate responses during the CS− on thefirst day of conditioning. However, on the second dayof conditioning, heart rate responses during the CS+were significantly decelerated when compared to heartrate responses during the CS− (t(5)=4.65, pB0.005).
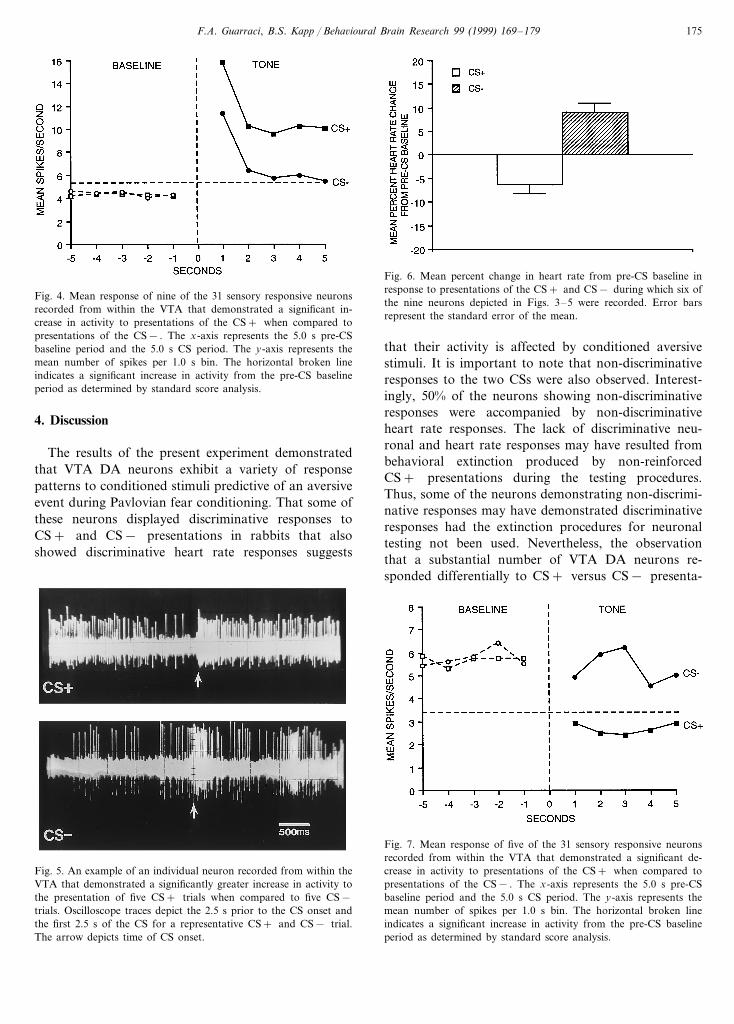
Fig. 1. The mean percent change in heart rate to CS presentationsduring the two days of conditioning for the six rabbits from whichrecordings were made in the VTA. Note the emergence of theconditioned bradycardic response to presentations of the CS+ . Errorbars represent standard error of the mean.

F.A. Guarraci, B.S. Kapp / Beha6ioural Brain Research 99 (1999) 169–179 173
Fig. 2. (A). Oscilloscope traces (four overlays) of a representativeneuron recorded from within the VTA. Note the long duration of theaction potential (\3.0 ms). (B) An oscilloscope trace depicting thespontaneous discharges of the neuron in (A). Note high-frequencybursts (brackets; mean interspike interval approximately 60 ms) inter-spersed with single spike discharges (asterisks), characteristics of VTADA neurons.
duration revealed Z scores ranging from 2.10 to 10.03(pB0.05). For eight of these 15 neurons, there was alsoa significant increase in activity during CS− presenta-tions (Z scores: 2.03–2.70, pB0.05).
3.2.2.1.2. Differential responsi6eness. Individual t-tests for related measures revealed that there was asignificantly greater response to presentations of theCS+ when compared to the CS− for nine of these 15neurons (t : 2.26–4.13, pB0.05). The mean data forthese nine neurons are presented in Fig. 4. As is appar-ent from this figure, the significant increase in activityduring CS+ presentations occurred during the entire5.0 s period. Standard score analyses revealed a signifi-cant increase in activity between the first and eighth 10ms bin (0–80 ms) for these nine neurons following theonset of either the CS+ or the CS− with a meanlatency of 22.6 ms (Z scores: 2.02–9.01, pB0.05).Furthermore, four of these nine neurons also demon-strated a significant correlation between neuronal activ-ity and heart rate changes on a trial-by-trial basis (r :−0.58, −0.59, −0.60 and −0.67, pB0.05). An ex-ample of recordings from one of these neurons duringrepresentative CS+ and CS− presentations is shownin Fig. 5.
3.2.2.1.3. Heart rate conditioning. Differentially con-ditioned bradycardic responses were observed in re-sponse to CS presentations during the recording ofeight of these 15 neurons. This was demonstrated bysignificantly greater bradycardic responses during CS+trials than during CS− trials (t : 3.98–7.10, pB0.05).Of the nine neurons which demonstrated significantlygreater activity during the CS+ than during the CS− ,six were also accompanied by significantly greaterbradycardic responses during CS+ trials than duringCS− trials. The mean heart rate data from CS+ andCS− trials presented during the recording of these sixneurons are shown in Fig. 6. Differential bradycardicresponses were not observed in response to the CS+versus the CS− for three of the six neurons that didnot respond differentially to the conditioned stimuli.
3.2.2.2. Type II3.2.2.2.1. CS responsi6eness. Type II neurons (n=5)
demonstrated a statistically significant decrease in activ-ity to presentations of the CS+ . Standard score analy-ses for the entire 5.0 s CS duration revealed Z scoresranging from −2.24 to −2.94 (pB0.05) during CS+presentations for these neurons. There was no signifi-cant change in the activity of these neurons to CS−presentations.
3.2.2.2.2. Differential responsi6eness. Individual t-tests for related measures revealed that there was sig-nificantly less activity during CS+ presentations thanduring CS− presentations for these neurons (t : 2.98–10.97, pB0.05). The mean data for these neurons arepresented in Fig. 7. An example of recordings from one
3.2. Electrophysiological data
3.2.1. Characteristics of 6entral tegmental areadopaminergic neurons
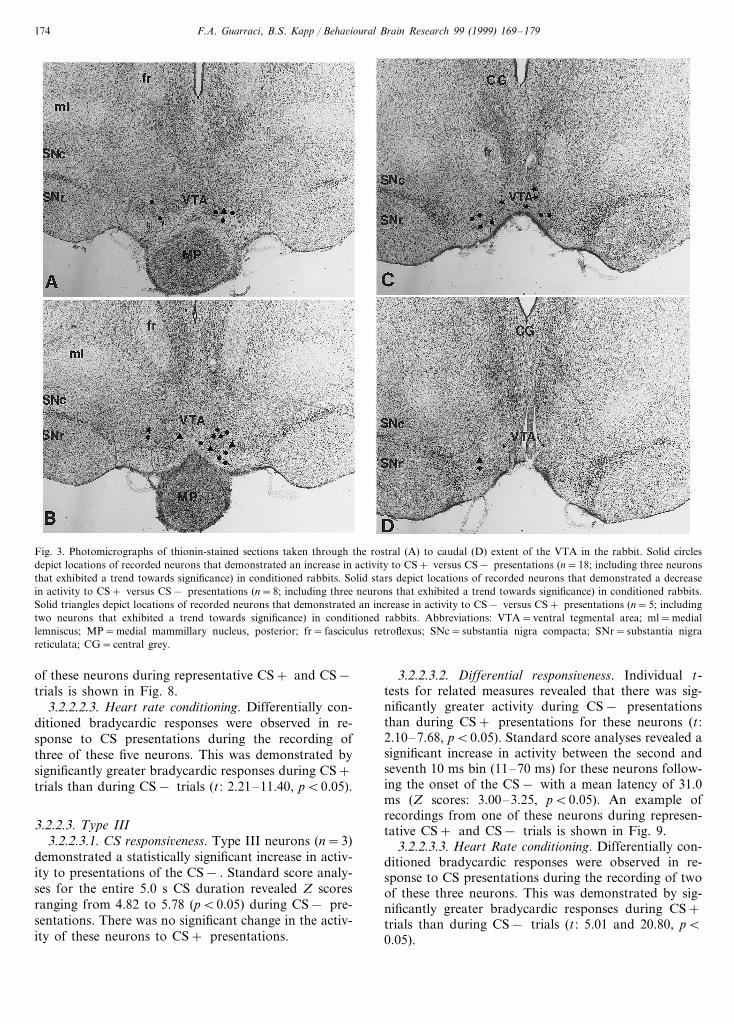
Extracellular single unit recordings were made from51 neurons which (a) met the electrophysiological crite-ria for VTA DA neurons, as described previously andshown in Fig. 2(A) and (B), (b) were located within theVTA based on histological analysis and (c) were testedfor responsiveness to at least four presentations of eachof the CS+ and CS− . Of these 51 neurons, 31 wereresponsive to the conditioned stimuli, and their re-sponses were subjected to statistical analyses. Each ofthese neurons was classified into one of three categoriesdescribed below. The remaining 20 neurons were notresponsive to CS presentations as determined by a lackof responsiveness to at least two presentations of theCS+ and the CS− . Fig. 3 depicts the location of the31 CS-responsive neurons in coronal, thionin-stainedsections through the rostral-caudal extent of the VTA.As depicted in Fig. 3, these CS-responsive neurons wereall within the DA neuron-rich region of the VTA.
3.2.2. Description of neuronal types (I–III)
3.2.2.1. Type I3.2.2.1.1. CS responsi6eness. Type I neurons (n=15),
which were the most frequently encountered CS-re-sponsive neurons, demonstrated a statistically signifi-cant increase in activity to presentations of the CS+ .Standard score analyses for the entire 5.0 s CS+
F.A. Guarraci, B.S. Kapp / Beha6ioural Brain Research 99 (1999) 169–179174
Fig. 3. Photomicrographs of thionin-stained sections taken through the rostral (A) to caudal (D) extent of the VTA in the rabbit. Solid circlesdepict locations of recorded neurons that demonstrated an increase in activity to CS+ versus CS− presentations (n=18; including three neuronsthat exhibited a trend towards significance) in conditioned rabbits. Solid stars depict locations of recorded neurons that demonstrated a decreasein activity to CS+ versus CS− presentations (n=8; including three neurons that exhibited a trend towards significance) in conditioned rabbits.Solid triangles depict locations of recorded neurons that demonstrated an increase in activity to CS− versus CS+ presentations (n=5; includingtwo neurons that exhibited a trend towards significance) in conditioned rabbits. Abbreviations: VTA=ventral tegmental area; ml=mediallemniscus; MP=medial mammillary nucleus, posterior; fr= fasciculus retroflexus; SNc=substantia nigra compacta; SNr=substantia nigrareticulata; CG=central grey.
of these neurons during representative CS+ and CS−trials is shown in Fig. 8.
3.2.2.2.3. Heart rate conditioning. Differentially con-ditioned bradycardic responses were observed in re-sponse to CS presentations during the recording ofthree of these five neurons. This was demonstrated bysignificantly greater bradycardic responses during CS+trials than during CS− trials (t : 2.21–11.40, pB0.05).
3.2.2.3. Type III3.2.2.3.1. CS responsi6eness. Type III neurons (n=3)
demonstrated a statistically significant increase in activ-ity to presentations of the CS− . Standard score analy-ses for the entire 5.0 s CS duration revealed Z scoresranging from 4.82 to 5.78 (pB0.05) during CS− pre-sentations. There was no significant change in the activ-ity of these neurons to CS+ presentations.
3.2.2.3.2. Differential responsi6eness. Individual t-tests for related measures revealed that there was sig-nificantly greater activity during CS− presentationsthan during CS+ presentations for these neurons (t :2.10–7.68, pB0.05). Standard score analyses revealed asignificant increase in activity between the second andseventh 10 ms bin (11–70 ms) for these neurons follow-ing the onset of the CS− with a mean latency of 31.0ms (Z scores: 3.00–3.25, pB0.05). An example ofrecordings from one of these neurons during represen-tative CS+ and CS− trials is shown in Fig. 9.
3.2.2.3.3. Heart Rate conditioning. Differentially con-ditioned bradycardic responses were observed in re-sponse to CS presentations during the recording of twoof these three neurons. This was demonstrated by sig-nificantly greater bradycardic responses during CS+trials than during CS− trials (t : 5.01 and 20.80, pB0.05).
F.A. Guarraci, B.S. Kapp / Beha6ioural Brain Research 99 (1999) 169–179 175
Fig. 4. Mean response of nine of the 31 sensory responsive neuronsrecorded from within the VTA that demonstrated a significant in-crease in activity to presentations of the CS+ when compared topresentations of the CS− . The x-axis represents the 5.0 s pre-CSbaseline period and the 5.0 s CS period. The y-axis represents themean number of spikes per 1.0 s bin. The horizontal broken lineindicates a significant increase in activity from the pre-CS baselineperiod as determined by standard score analysis.
Fig. 6. Mean percent change in heart rate from pre-CS baseline inresponse to presentations of the CS+ and CS− during which six ofthe nine neurons depicted in Figs. 3–5 were recorded. Error barsrepresent the standard error of the mean.
that their activity is affected by conditioned aversivestimuli. It is important to note that non-discriminativeresponses to the two CSs were also observed. Interest-ingly, 50% of the neurons showing non-discriminativeresponses were accompanied by non-discriminativeheart rate responses. The lack of discriminative neu-ronal and heart rate responses may have resulted frombehavioral extinction produced by non-reinforcedCS+ presentations during the testing procedures.Thus, some of the neurons demonstrating non-discrimi-native responses may have demonstrated discriminativeresponses had the extinction procedures for neuronaltesting not been used. Nevertheless, the observationthat a substantial number of VTA DA neurons re-sponded differentially to CS+ versus CS− presenta-
4. Discussion
The results of the present experiment demonstratedthat VTA DA neurons exhibit a variety of responsepatterns to conditioned stimuli predictive of an aversiveevent during Pavlovian fear conditioning. That some ofthese neurons displayed discriminative responses toCS+ and CS− presentations in rabbits that alsoshowed discriminative heart rate responses suggests
Fig. 5. An example of an individual neuron recorded from within theVTA that demonstrated a significantly greater increase in activity tothe presentation of five CS+ trials when compared to five CS−trials. Oscilloscope traces depict the 2.5 s prior to the CS onset andthe first 2.5 s of the CS for a representative CS+ and CS− trial.The arrow depicts time of CS onset.
Fig. 7. Mean response of five of the 31 sensory responsive neuronsrecorded from within the VTA that demonstrated a significant de-crease in activity to presentations of the CS+ when compared topresentations of the CS− . The x-axis represents the 5.0 s pre-CSbaseline period and the 5.0 s CS period. The y-axis represents themean number of spikes per 1.0 s bin. The horizontal broken lineindicates a significant increase in activity from the pre-CS baselineperiod as determined by standard score analysis.

F.A. Guarraci, B.S. Kapp / Beha6ioural Brain Research 99 (1999) 169–179176
Fig. 8. An example of an individual neuron recorded from within theVTA that demonstrated a significant decrease in activity to thepresentation of five CS+ trials when compared to four CS− trials.Oscilloscope traces depict the 2.5 s prior to the CS onset and the first2.5 s of the CS for a representative CS+ and CS− trial. The arrowdepicts time of CS onset.
The present data are also consistent with and extendthe findings of Trulson and Preussler [44], who charac-terized the response of VTA DA neurons in the catduring presentations of a tone that had previously beenpaired with an aversive airpuff. The use of a differentialconditioning paradigm and the demonstration of con-current discriminative heart rate and neuronal respond-ing in the present study suggests that the responsivenessof at least some VTA DA neurons reflects a response tothe acquired motivational significance of the condi-tioned stimuli and not merely the sensory responsive-ness of these neurons.
Recently, however, Mirenowicz and Schultz [32], us-ing an avoidance paradigm, reported that very fewmesencephalic DA neurons (15%) demonstrated en-hanced activity in response to conditioned stimuli sig-nalling ‘non-noxious’ airpuff presentations delivered tothe hand, or hypertonic saline solution delivered intothe mouth, in two well-trained monkeys. Most of theneurons, however were excited by primary and condi-tioned appetitive stimuli, leading to the conclusion thatactivation of mesencephalic DA neurons convey infor-mation primarily about the appetitive value of primaryand conditioned environmental stimuli. A number ofreasons may account for why VTA DA neurons wereencountered which were excited by conditioned aversivestimuli in the present study but very few were encoun-tered by Mirenowicz and Schultz [32]. First, the dis-crepancy may lie in the differences between the twotasks. Airpuff delivery to the hand or oral injections ofhypertonic saline in monkeys may not have been suffi-ciently noxious to condition significant levels of fear tothe environmental stimuli signaling their occurrence.This is of considerable importance since the magnitudeof a stressor is related to the magnitude of dopamineactivation in specific terminal regions, as well as to howwide-spread dopamine activation is in multiple DAterminal regions as measured with in vivo and in vitroprocedures [23]. Had Mirenowicz and Schultz [32] useda more noxious US, substantial fear may have beenconditioned to the environmental stimuli, yieldinggreater numbers of neurons excited by the conditionedstimuli. Second, Mirenowicz and Schultz [32] recordedfrom mesencephalic DA neurons in well-trained mon-keys during an instrumental avoidance task. Generally,well-trained subjects in such a task completely avoidUS presentations by responding appropriately duringthe CS, and it has been repeatedly demonstrated thatCS presentations to well-trained animals in instrumen-tal avoidance conditioning paradigms elicit little signsof overt fear [31]. Mirenowicz and Schultz [32] mayhave encountered a smaller percentage of neuronswhich responded to conditioned stimuli predictive of anairpuff or orally administered hypertonic saline solutionbecause of a lack of substantial fear elicited by theseconditioned stimuli in the instrumental avoidance task.
tions during differential Pavlovian fear conditioningsuggests that the release of dopamine is altered in VTAterminal fields when these neurons are affected byfear-arousing stimuli. These data are consistent with alarge body of research illustrating changes in dopaminerelease and metabolism in various terminal regions,particularly in the mPFC, in response to both condi-tioned and unconditioned stressors.
Fig. 9. An example of an individual neuron recorded from within theVTA that demonstrated a significantly greater increase in activity tothe presentation of six CS− trials when compared to five CS+trials. Oscilloscope traces depict the 5.0 s prior to the CS onset andthe 5.0 s of the CS for a representative CS+ and CS− trial. Thearrow depicts time of CS onset.

F.A. Guarraci, B.S. Kapp / Beha6ioural Brain Research 99 (1999) 169–179 177
The higher percentage of recorded neurons that wereencountered which were responsive to conditioned aver-sive stimuli in the present study may have been due togreater fear elicited by these stimuli.
It is important to note that several additional VTADA response patterns were encountered in the presentstudy. The first of these was characterized by differen-tial decreases in neuronal activity during CS presenta-tions, with a significantly greater decrease to the CS+than to the CS− . This response pattern is similar to apattern encountered by Mirenowicz and Schultz [32].They reported neurons (30%) which demonstrated adecrease in activity in response to conditioned stimulithat signaled the occurrence of either an airpuff ororally administered hypertonic saline. However, theextent to which the response pattern of the populationencountered by Mirenowicz and Schultz [32] is identicalto that encountered in the present study is unknownsince a differential Pavlovian conditioning procedurewas not used in the Mirenowicz and Schultz [32] study,and, therefore, the differential responsiveness of theneurons in that study was not examined.
A second pattern was characterized by differentialincreases in neuronal activity in response to CS presen-tations, but significantly greater increases to CS− thanto CS+ presentations. This response pattern mayreflect the activation of VTA DA neurons to the moti-vational significance of the CS− as a safety signal.However, the interpretation that this response patternreflects the acquired ‘safety’ properties of the CS−should be considered speculative since the present studydid not rigorously test for the acquired ‘safety’ proper-ties of the CS− .
A final pattern was characterized by similar increasesin neuronal activity to both conditioned stimuli (i.e. theCS+ and the CS− ) even though a statistically signifi-cant, discriminative bradycardic response was observed.The non-discriminative response of these neurons mayreflect their activation by some common, as yet unde-termined, property of the conditioned stimuli.
That a population of DA neurons in the VTA wasexcited by a fear-arousing CS in the present researchmay suggest the contribution of these neurons to theelicitation and/or expression of a central state of fear.Evidence consistent with this suggestion comes fromthree lines of research in the rat. First, lesions of theVTA affect a number of fear-motivated behaviors. Forexample, 6-hydroxydopamine (6-OHDA) lesions com-pletely block conditioned avoidance behavior [35],whereas excitotoxic lesions block the potentiation ofacoustic startle by a conditioned fear-arousing stimulusin the fear-potentiated startle paradigm [4]. Second,electrical stimulation of the VTA facilitates acousticstartle [4]. Third, blockade of dopamine transmissionwith systemic injections of DA antagonists has beenshown to disrupt conditioned avoidance and fear po-
tentiated startle [3,8,13,14]. More recent evidence sug-gests that the avoidance impairments followingadministration of DA antagonists are most likely afunction of antagonist-induced enhancement of species-specific defensive freezing behavior which could poten-tially compete with the conditioned active avoidanceresponse, thereby creating impaired avoidance [3]. Insupport of this interpretation are the findings thatfreezing behavior during contextual fear conditioning isattenuated by systemic injections of the non-specificagonist apomorphine or by a combined injection ofquinpirole, a D2 agonist and SKF 38393, a D1 agonist[24]. These findings suggest that the VTA contributesimportantly to some aspects of fear behavior, and theresults of the present study are consistent with thissuggestion.
Given the present results demonstrating that motiva-tionally significant conditioned aversive stimuli modu-late the activity of VTA DA neurons, the questionarises concerning the source of VTA afferents whichcontribute to this modulation. Future research specifi-cally designed to identify and characterize the afferentmodulation of VTA DA neurons during fear condition-ing is necessary to determine (a) if different afferents tothe VTA contribute differentially to the various re-sponse patterns exhibited by VTA DA neurons ob-served in the present study and (b) the functionalsignificance of these patterns during Pavlovian fearconditioning.
Acknowledgements
This research was supported by National ScienceFoundation grant c IBN 9319699 awarded to BruceS. Kapp.
References
[1] Abercrombie ED, Keefe KA, Di Frischia DS, Zigmond MJ.Difference on in vivo dopamine release in striatum, nucleusaccumbens and medial frontal cortex. J Neurochem1989;52:1655–8.
[2] Agmo A, Federman I, Navarro V, Padua M, Velazquez C.Reward and reinforcement produced by drinking water: role ofopioids and dopamine receptor subtype. Pharmacol BiochemBehav 1993;46:183–94.
[3] Blackburn JR, Phillips AG. Enhancement of freezing behaviorby metoclopramide: Implications for neuroleptic-induced avoid-ance deficits. Pharmacol Biochem Behav 1990;35:685–91.
[4] Borowski TB, Kokkinidis L. Contribution of ventral tegmentalarea dopamine neurons to expression of conditioned fear: effectsof electrical stimulation, excitotoxin lesions, and quinpirole infu-sion on potentiated startle in rats. Behav Neurosci1996;110:1349–64.
[5] Cabib S, Puglisi-Allegra S. Opposite responses of mesolimbicdopamine system to controllable and uncontrollable aversiveexperiences. J Neurosci 1994;14:3333–40.

F.A. Guarraci, B.S. Kapp / Beha6ioural Brain Research 99 (1999) 169–179178
[6] Cenci MA, Kalen P, Mandel RJ, Bjorklund A. Regional dif-ferences in the regulation of dopamine and noradrenaline re-lease in medial frontal cortex, nucleus accumbens andcaudate-putamen: a microdialysis study in the rat. Brain Res1992;581:217–28.
[7] Chiodo LA, Antelman SM, Caggiula AR, Lineberry CG. Sen-sory stimuli alter the discharge rate of dopamine neurons: evi-dence for two functional types of dopamine cells in thesubstantial nigra. Brain Res 1980;189:544–9.
[8] Chipkin RE, Iorio LC, Coffin VL, Mc Quade RD, Berger JG,Barnett A. Pharmacological profile of SCH39166: A dopamineD1 selective benzoaphthazepine with potential antipsychotic ac-tivity. J Pharmacol Exp Ther 1988;247:1093–102.
[9] Claustre Y, Rivy JP, Dennis T, Scatton B. Pharmacologicalstudies on stress-induced increase in frontal cortical dopaminemetabolism in the rat. J Pharmacol Exp Ther 1986;238:693–9.
[10] Coco ML, Kuhn CM, Ely TD, Kilts CD. Selective activationof mesoamygdaloid dopamine neurons by conditioned stress:Attenuation by diazepam. Brain Res 1992;590:39–47.
[11] Commissiong JW. Monoamine metabolites: Their relationshipand lack of relationship to monoaminergic neuronal activity.Biochem Pharmacol 1985;34:1127–31.
[12] Damsma G, Pfaus JC, Wenkstern D, Phillips AG, Fibiger HC.Sexual behavior increases dopamine transmission in the nucleusaccumbens and striatum of male rats: comparison with noveltyand locomotion. Behav Neurosci 1992;106:181–91.
[13] Davidson AB, Weidley E. Differential effects of neurolepticsand other psychotropic agents on acquisition of avoidance inrats. Life Sci 1976;18:1279–85.
[14] Davis M. Pharmacological analysis of fear-potentiated startle.Braz J Med Biol Res 1993;26:235–60.
[15] Deutch AY, Tam SY, Roth RH. Footshock and conditionedstress increase 3,4-dihydroxyphenylacetic acid (DOPAC) in theventral tegmental area but not substantia nigra. Brain Res1985;333:143–6.
[16] Deutch AY, Lee MC, Gillham MH, Cameron DA, GoldsteinM, Iadarola MJ. Stress selectively increases Fos protein indopamine neurons innervating the prefrontal cortex. CerebCortex 1991;1:273–92.
[17] DiChiara G, Imperato A. Drugs abused by humans preferen-tially increase synaptic dopamine concentrations in themesolimbic system of freely moving rats. Proc Natl Acad SciUSA 1988;85:5474–8.
[18] Fadda F, Argiolas A, Melis MR, Tissari AH, Onali PL, GessaGL. Stress-induced increase in 3,4-dihydroxyphenylacetic acid(DOPAC) levels in the cerebral cortex and in n. accumbens:reversal by diazepam. Life Sci 1978;23:2219–24.
[19] Freeman AS, Bunney BS. Activity of A9 and A10 dopaminer-gic neurons in unrestrained rats: further characterization andeffects of apomorphine and cholecystokinin. Brain Res1987;405:46–55.
[20] Grenhoff J, Ugedo L, Svensson TH. Firing patterns of mid-brain dopamine neurons: difference between A9 and A10 cells.Acta Physiol Scand 1988;134:127–32.
[21] Heffner TG, Hartman JA, Seiden LS. Feeding increases do-pamine metabolism in the rat brain. Science 1980;208:1168–70.
[22] Herman JP, Guillonneau D, Dantzer R, Scatton B, Semerd-jian-Rouquier L, Le Moal M. Differential effects of ines-capable footshocks and of stimuli previously paired withinescapable footshocks on dopamine turnover in cortical andlimbic areas of the rat. Life Sci 1982;30:2207–14.
[23] Inoue T, Tsuchiya K, Koyama T. Regional changes in do-pamine and serotonin activation with various intensity of phys-ical and psychological stress in the rat brain. PharmacolBiochem Behav 1994;49:911–20.
[24] Kamei H, Kameyama T, Nabeshima T. Activation of bothdopamine D1 and D2 receptors necessary for amelioration ofconditioned fear stress. Eur J Pharmacol 1995;273:229–33.
[25] Kaneyuki H, Yokoo H, Tsuda A, Yoshida M, Mizuki Y,Yamada M, Tanaka M. Psychological stress increases do-pamine turnover selectively in mesoprefrontal dopamine neu-rons of rats: reversal by diazepam. Brain Res 1991;557:154–61.
[26] Kosobud AEK, Harris GC, Chapin JK. Behavioral associa-tions of neuronal activity in the ventral tegmental area of therat. J Neurosci 1994;14:7117–29.
[27] Lavielle S, Tassin JP, Thierry AM, Blanc DH, Barthelemy C,Glowinski J. Blockade by benzodiazepines of the selective highincrease in dopamine turnover induced by stress in the meso-cortical dopaminergic neurons of the rat. Brain Res1978;168:585–94.
[28] Ljungberg T, Apicella P, Schultz W. Response of monkey do-pamine neurons during learning of behavioral reactions. J Neu-rophysiol 1992;67:145–63.
[29] Maeda H, Mogenson GJ. Effects of peripheral stimulation onthe activity of neurons in the ventral tegmental area, substantianigra and midbrain reticular formation. Brain Res Bull1982;8:7–14.
[30] Miller JD, Sanghera MK, German DC. Mesencephalic do-paminergic unit activity in the behaviorally conditioned rat.Life Sci 1981;29:1255–63.
[31] Mineka S, Gino A. Dissociation between conditioned emo-tional response and extended avoidance performance. LearningMotivat 1980;11:476–502.
[32] Mirenowicz J, Schultz W. Preferential activation of midbraindopamine neurons by appetitive rather than aversive stimuli.Nature 1996;379:449–51.
[33] Pei Q, Zetterstrom T, Fillenz M. Tail pinch-induced changes inthe turnover and release of dopamine and 5-hydroxytryptaminein different brain regions of the rat. Neuroscience 1990;35:133–8.
[34] Pfaus JG, Damsma G, Nomikos GG, Wenkstern DG, BlahaCD, Phillips AG, Fibiger HC. Sexual behavior enhances cen-tral dopamine transmission in the male rat. Brain Res1990;530:345–8.
[35] Oades RD, Rivet JM, Taghzouti K, Kharouby M, Simon H,Le Moal M. Catecholamines and conditioned blocking: effectsof ventral tegmental, septal and frontal 6-hydroxydopaminelesions in rats. Brain Res 1987;406:136–46.
[36] Olds ME, Fobes JL. The central basis of motivation: intracra-nial self-stimulation studies. Annu Rev Psychol 1981;32:523–74.
[37] Roth RH, Tam SY, Ida Y, Yang JX, Deutch AY. Stress andthe mesocorticolimbic dopamine systems. In: Kalivas PW, Ne-meroff CB, editors, Mesocorticolimbic Dopamine System vol.537. New York: New York Academy of Sciences, 1988:138–147.
[38] Schultz W, Apicella P, Ljungberg T. Response of monkey do-pamine neurons to reward and conditioned stimuli during suc-cessive steps of learning a delayed response task. J Neurosci1993;13:900–13.
[39] Schultz W, Romo R. Dopamine neurons of the monkey mid-brain: Contingencies of response to active touch during self-ini-tiated arm movements. J Neurophysiol 1990;63:592–606.
[40] Schultz W, Romo R. Dopamine neurons of the monkey mid-brain: Contingencies of response to stimuli eliciting immediatebehavioral reactions. J Neurophysiol 1990;63:607–24.
[41] Soares-da-Silva P, Garrett MC. A kinetic study of the rate offormation of dopamine, 3,4-dihydroxyphenylacetic acid (DO-PAC) and homovanillic acid (HVA) in the brain of the rat:Implications for the origin of DOPAC. Neuropharmacology1990;29:869–74.

F.A. Guarraci, B.S. Kapp / Beha6ioural Brain Research 99 (1999) 169–179 179
[42] Steinfels GF, Heym J, Strecker RE, Jacobs BL. Behavioralcorrelates of dopaminergic unit activity in freely moving cats.Brain Res 1983;258:217–28.
[43] Strecker RE, Jacobs BL. Substantia nigra dopaminergic unitactivity in behaving cats: Effect of arousal on spontaneousdischarge and sensory evoked activity. Brain Res1985;361:339–50.
[44] Trulson ME, Preussler DW. Dopamine-containing ventral teg-mental area neurons in freely moving cats: activity duringsleep-waking cycle and effects of stress. Exp Neurol1984;83:367–77.
[45] Wise RA. Brain neuronal systems mediating reward processes.In: Smith JE, Lane JD, editors, The Neurobiology of OpiateReward Processes. New York: Elsevier, 1983:405–437.
[46] Wise RA, Rompre PP. Brain dopamine and reward. Annu RevPsychol 1989;40:191–225.
[47] Wood PL, Nair NPV, Bozarth M. Striatal 3-methoxytyramineas an index of dopamine release: Effects of electrical stimula-tion. Neurosci Lett 1982;32:291–4.
[48] Yim CY, Mogenson GJ. Electrophysiological studies of neu-rons in the ventral tegmental area of Tsai. Brain Res1980;181:310–3131.
[49] Yoshida M, Yokoo H, Mizoguchi K, Kawahara H, Tsuda A,Nishikawa T, Tanaka M. Eating and drinking cause increaseddopamine release in the nucleus accumbens and ventral teg-mental area in the rat: measurement by in vivo microdialysis.Neurosci Lett 1992;139:73–6.
.





























