Embed Size (px)
Citation preview

REPLIKACJA, NAPRAWA i REKOMBINACJA DNA
1) Replikacja DNA
2) Replikacja całych chromosomów
3) Replikacja telomerów
4) Naprawa DNA przy jego syntezie
5) Naprawa DNA poza jego syntezą
6) Naprawa DNA – system SOS
7) Rekombinacja homologiczna DNA

1) Replikacja DNA – konieczność odsłonięcia jednoniciowego DNA
Rozdzielenie podwójnej nici powoduje odsłonięcie ‘lepkich’
pojedynczych nici, które maja tendencję do ponownego łączenia się.
Zapobiegają temu białka SSBs (single strand binding).

1) Replikacja DNA – inna przeszkodą jest skręcenie helisy
Nić helisy jest skręcona i to skręcenie się nasila w miarę przesuwania się
kompleksu replikacyjnego. Rozwiązaniem jest działanie topoizomerazy,
która nacina jedną z nici, przepuszcza drugą przez powstałą przerwę i
doprowadza do powtórnego złączenia.
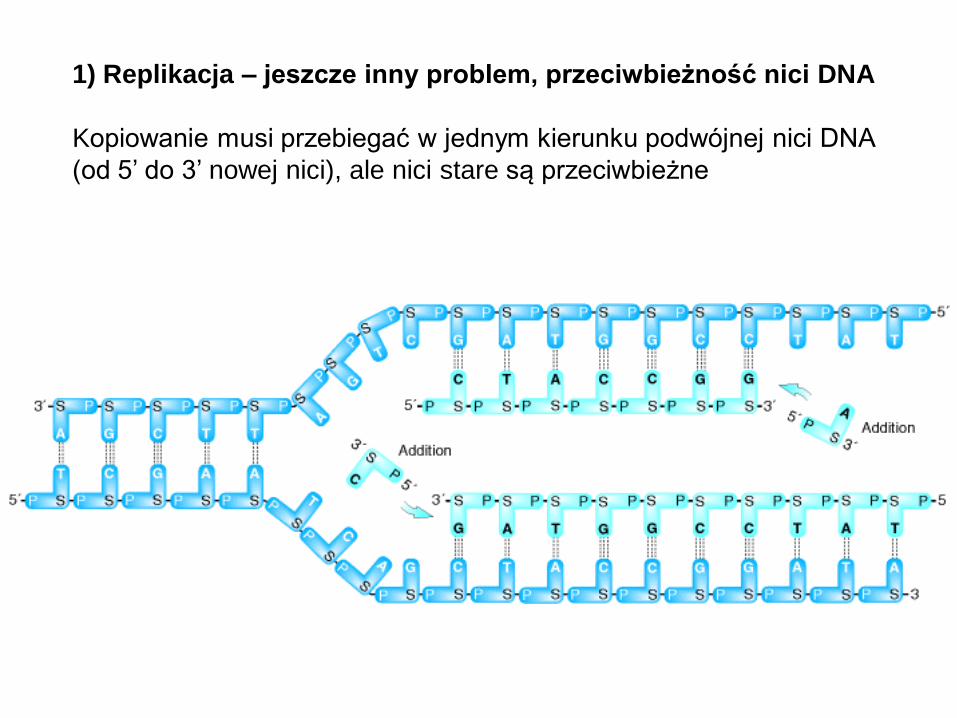
1) Replikacja – jeszcze inny problem, przeciwbieżność nici DNA
Kopiowanie musi przebiegać w jednym kierunku podwójnej nici DNA
(od 5’ do 3’ nowej nici), ale nici stare są przeciwbieżne

1) Replikacja DNA
– rozwiązaniem jest
„normalna” replikacja w
przypadku nici “wiodącej” i
odcinkowa replikacja
(fragmenty Okazaki)
“spóźniającej się” nici DNA
(polimeraza RNA może
syntetyzować krótkie odcinki
na pojedynczej nici DNA,
polimeraza DNA może wtedy
zacząć od nici podwójnej)

1) Replikacja DNA
- helikaza – rozdziela starą podwójną nić DNA
- polimeraza DNA –dokłada komplementarne zasady do starej nici DNA
- topoizomeraza – likwiduje naprężenia powstające
- primaza RNA – tworzy krótkie komplementarne odcinki RNA
- ligaza DNA – łączy fragmenty Okazaki w ciągłą nową nić

2) Replikacja chromosomu bakteryjnego
- chromosomy bakterii (a także plazmidy i koliste genomy wirusów) mają
jeden początek replikacji
eukarionty

2) Replikacja chromosomu eukariotycznego
U organizmów eukariotycznych występuje wiele początków replikacji w
każdym z chromosomów

3) Replikacja telomerów
Przy replikacji liniowych chromosomów eukariontów na ostatnim fragmencie
nici opóźniającej się (lagging) nie można utworzyć starteru RNA i dlatego ta
nić skracałaby się o ten fragment po replikacji.
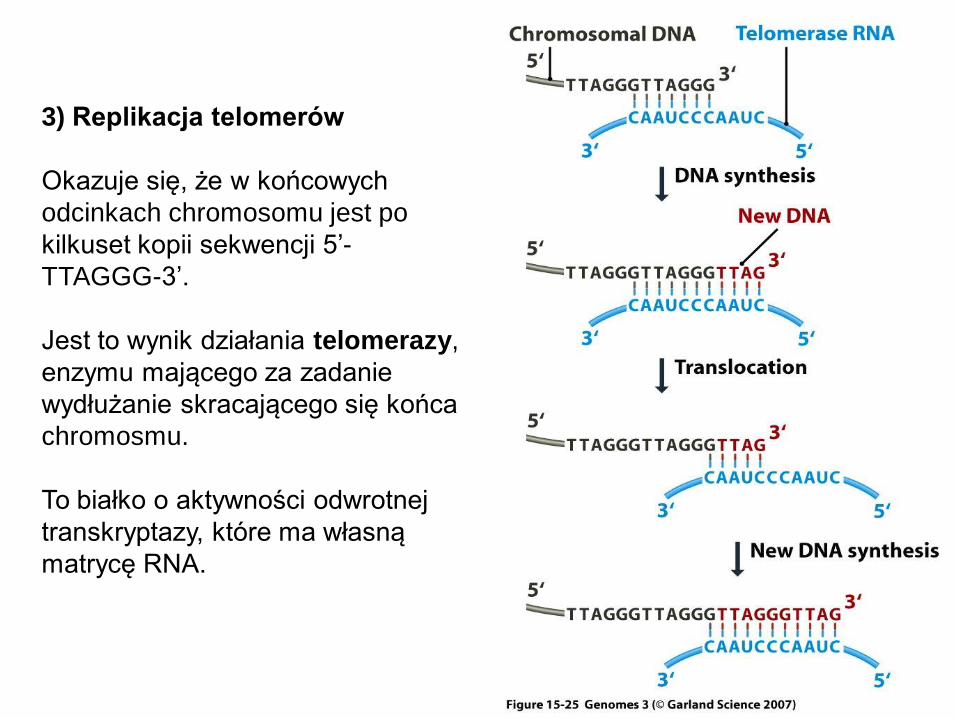
3) Replikacja telomerów
Okazuje się, że w końcowych
odcinkach chromosomu jest po
kilkuset kopii sekwencji 5’-
TTAGGG-3’.
Jest to wynik działania telomerazy,
enzymu mającego za zadanie
wydłużanie skracającego się końca
chromosmu.
To białko o aktywności odwrotnej
transkryptazy, które ma własną
matrycę RNA.

3) Telomery a starzenie się
komórek
Telomeraza jest aktywna w
komórkach linii generatywnej i
komórkach progenitorowych
(stem cells). W tkankach
normalnie nie działa.
Dlatego tam telomery się
skracają i jest to zegar starzenia
się. Po odliczonej liczbie
podziałów, zwykle
kilkudziesięciu, komórka nie
może się dalej dzielić.

4) Naprawa DNA przy jego
syntezie
Polimeraza DNA działa tak by
nie popełniać błędów. Selekcja
nukleotydów jest imponująco
dokładna, pomyłki występują
raz na 10,000 sparowań.

4) Naprawa DNA przy jego
syntezie
Jeśli polimeraza DNA się
pomyli, to potrafi sama
naprawić swój błąd ponieważ
jest też egzonukleazą
Ta forma naprawy nazywana
jest naprawą korekcyjną
(proofreading). Pomyłki
następują tu około 1/1000.

4) Naprawa DNA tuż po
jego syntezie –
mismatch repair
Tuż po syntezie można
jeszcze rozróżnić nić starą
od nowe.
U bakterii tylko stara nić
jest zmetylowana (rycina).
U eukariontów
rozróznianie jest inne i
gorzej poznane.
Można wtedy usuwać
niedopasowania
(mismatches) tylko z
nowej nici.

4) Naprawa DNA tuż po jego
syntezie – mismatch repair –
naprawa niedopasowań
U bakterii dwa białka, MutH i MutS,
skanują DNA.
Po natrafieniu na źle sparowane
nukleotydy przywołują egzonukleazę
wycinającą fragment nowej nici i
polimerazę, która zapełnia ubytek.
Białka homologiczne do MutS i MutH
wykrywające złe sparowania są także
u eukariontów.
Ta naprawa pozostawia około 1/1000
niepoprawnych sparowań.

4) Naprawa DNA przy jego syntezie – efekt łączny
Badania kultur bakteryjnych w laboratorium wykazały, że:
- przy dobieraniu nukleotydów tempo błędu wynosi 1 x 10-4
- z tego po naprawie korekcyjnej pozostaje 1 x 10-3
- z tego po naprawie niedopasowań pozostaje 1 x 10-3
Łącznie daje to tempo błędów zaledwie 1 x 10-10 na nukleotyd
Jak to się ma do tempa mutacji u człowieka szacowanego na
podstawie porównywania sekwencji rodziców i dzieci?

Bakteryjne tempo 10-10 na jeden nukleotyd na jeden podział komórkowy
to około 10-8 na jeden nukleotyd na jedno pokolenie ludzkie zakładając
około 102 podziałów komórkowych w linii generatywnej człowieka …
- zgadza się z danymi z sekwencjonowania -
…. czyli większość mutacji powstaje przy powielaniu DNA

5) Naprawa DNA poza jego syntezą
DNA w komórce nie dzielącej się może zostać uszkodzone w pojedynczym
nukleotydzie w taki sposób, który nie wymaga (A) lub wymaga (B) wycięcia
w starej nici. Niedopasowanie (C) z definicji wymaga wycięcia. Pęknięcie
obu nici (D) wymaga złączenia, co może odbywać się bez względu na
sekwencję pękniętych końców (nonhomologous).

5) Naprawa DNA poza jego syntezą – naprawa bezpośrednia
Naprawa bezpośrednia to np. naprawa pęknięcia (nick) jednej nici DNA
przez ligazę.

5) Naprawa DNA poza jego syntezą – wycięcie jednego nukleotydu
Uszkodzona zasada jest odnajdywana przez glikozylazę DNA i odcinana od cukru.
Następnie cały nukleotyd, ale tylko ten jeden, jest usuwany a potem zapełniany
prawidłowym.

5) Naprawa DNA poza jego syntezą – wycięcie wielu nukleotydów
Gdy uszkodzenie lub brak sparowania powoduje lokalne zaburzenie struktury
helisy, najpierw następuje rozdzielenie nici (melting) przez helikazę. Potem nić
zawierająca uszkodzenie jest wycinana, polimeraza dobudowuje drugą nić, a
ligaza łączy ją kowalencyjnie.

5) Naprawa DNA poza jego syntezą – łączenie końców
Gdy podwójna nić pęknie
istnieją systemy
szybkiego łączenia
wolnych końców.
U eukariontów
pośredniczy w tym białko
Ku i szereg innych
białek.
W ten sposób złączeniu
ulec mogą końce, które
niedawno się przełamały
lub nie odpowiadające
sobie (nonhomologous).

6) Naprawa DNA – bakteryjny system SOS
Gdy w czasie replikacji
‘normalna’ polimeraza III
natrafi na zbyt uszkodzone
DNA nie może posuwać się
dalej.
Wtedy przywoływane są
białka RecA, które pozwalają
na odłączenie polimerazy III i
umożliwiają pracę mniej
wymagającej polimerazie V.
Synteza postępuje, ale z
błędami.
U eukariontów także istnieją
takie ‘ratunkowe’ polimerazy
pomostowe, zdolne do
przejścia przez uszkodzone
DNA.
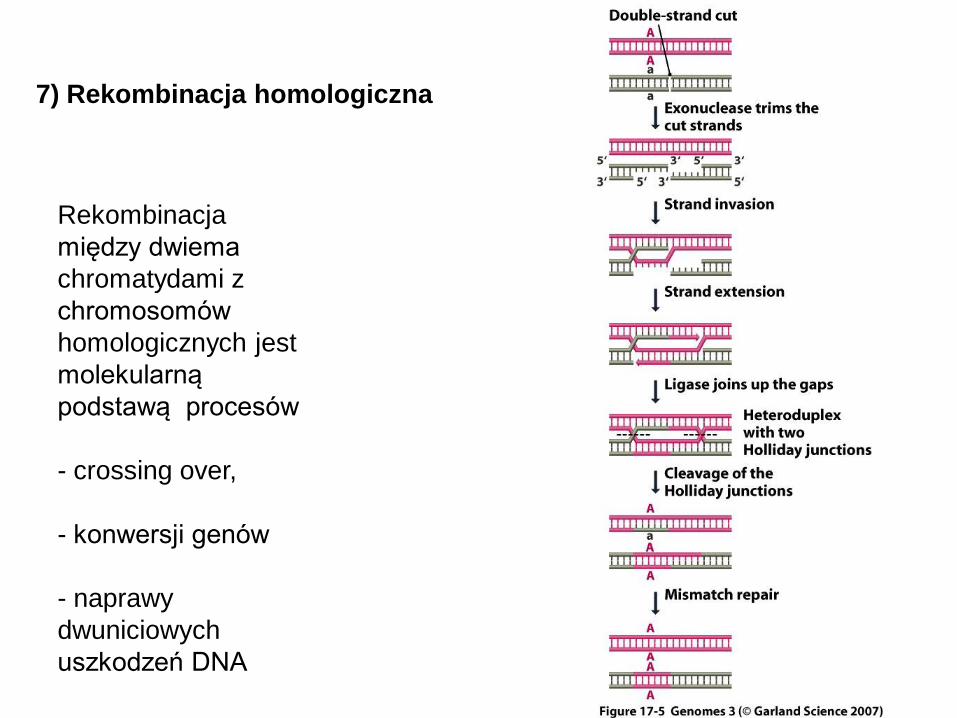
Rekombinacja
między dwiema
chromatydami z
chromosomów
homologicznych jest
molekularną
podstawą procesów
- crossing over,
- konwersji genów
- naprawy
dwuniciowych
uszkodzeń DNA
7) Rekombinacja homologiczna
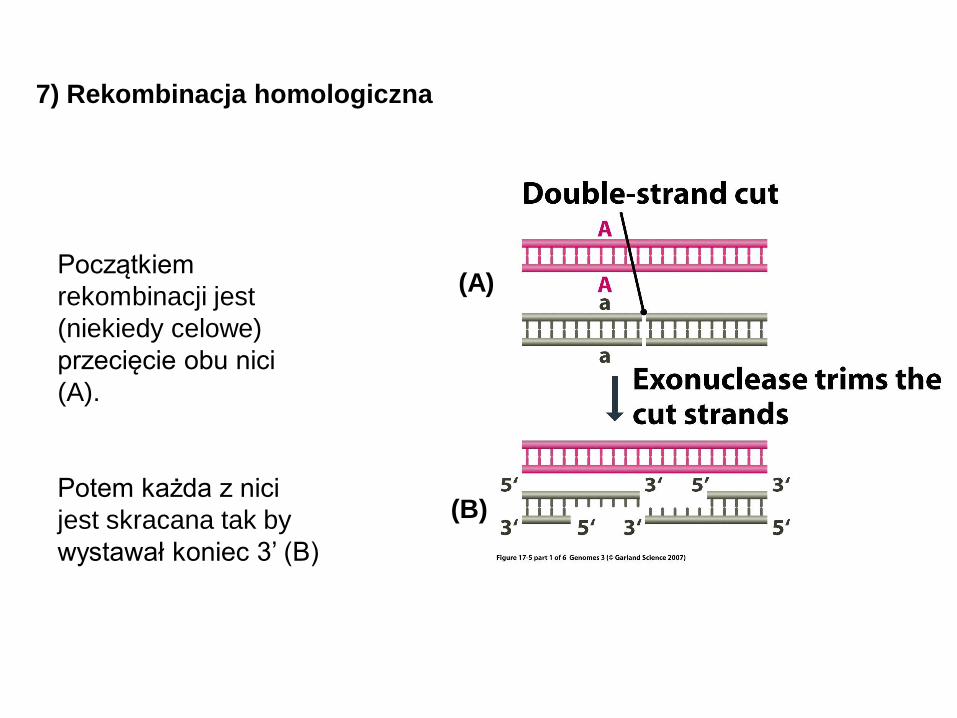
Początkiem
rekombinacji jest
(niekiedy celowe)
przecięcie obu nici
(A).
Potem każda z nici
jest skracana tak by
wystawał koniec 3’ (B)
7) Rekombinacja homologiczna
(A)
(B)

Następnie jeden z
końców 3’ dokonuje
inwazji powodując
miejscowe rozejście się
w drugiej nici połączone
z odszukaniem
sekwencji
homologicznej (C). Ten
koniec 3’ ma już
matrycę.
Rozejście przesuwa się
i drugi koniec 3’ zyskuje
matrycę. Oba końce 3’
mogą się wydłużać (D).
(C)
7) Rekombinacja homologiczna
(B)
(C)
(D)

Po zapełnieniu odcinków
jednoniciowych drugą nicią
tworzy się struktura z
dwóch różnych chromatyd
(heterodupleks)
Połączone są dwoma
przejścia nici między
chromatydami
homologicznymi - Holliday
junctions (E)
(D)
(E)
7) Rekombinacja homologiczna

Heterodukpleks musi
być przecięty.
Możliwe są różne
kombinacje cięć (F)
Prowadzą one albo
tylko do lokalnej
wymiany materiału
genetycznego
(konwersja genu) lub
do wymiany ramion
chromatyd (cross-
over)
konwersja
cross-over
(E)
(F)
7) Rekombinacja homologiczna

Powtórzenie całego szlaku.
Ten proces jest inicjowany
celowo na początku mejozy
by chromosomy
homologiczne na pewno się
odnalazły.
Jest także używany do
reperacji dwuniciowych
uszkodzeń DNA (pęknięcia,
połączenia kowalencyjne)
by zreperować poprawnie,
korzystając z sekwencji
innej chromatydy.
7) Rekombinacja homologiczna







![Zapobieganie i naprawa szkód w środowisku [tryb zgodności]ochronaprzyrody.gdos.gov.pl/files/...i_naprawa_szkod_w_srodowisku.pdf · ZAPOBIEGANIE I NAPRAWA SZKÓD W ŚRODOWISKU Edyta](https://img.dokumen.tips/doc/110x75/5c7593f209d3f25d028b557e/zapobieganie-i-naprawa-szkod-w-srodowisku-tryb-zgodnosci-zapobieganie-i.jpg)
![Badanie i naprawa elektronicznych elementów układów ... › images › dokumenty › dydaktyka › ... · 724[O2].Z2.02 Badanie i naprawa elementów elektrycznych i elektronicznych](https://img.dokumen.tips/doc/110x75/5f0bbf4f7e708231d43204d0/badanie-i-naprawa-elektronicznych-elementw-ukadw-a-images-a-dokumenty.jpg)





















