Embed Size (px)
Citation preview

THE JOURNAL OF COMPARATIVE NEUROLOGY 25535-49 (1987)
Relationship Between Astrocytes, Ganglion Cells and Vasculature of the Retina
JONATHAN STONE AND ZOFIA DREHER School of Anatomy, University of New South Wales, Sydney, Australia
ABSTRACT We have studied the distribution of astrocytes in the ganglion cell and
nerve fibre layers of the retina in cat, rat, rabbit, and possum using anti- serum and a monoclonal antibody against glial fibrillary acidic protein (GFAP) and o w own monoclonal antibody against glial filaments. The dis- tribution of retinal astrocytes appears to be strongly determined by the vasculature of the retina; astrocytes are absent from almost all the retina of the possum and from the avascular regions of the rabbit retina. In the cat and rabbit, retinal astrocytes also show a strong affinity for the bundles of ganglion cell axons found at the inner surface of the retina.
Retinal astrocytes do not invest the somas of ganglion cells, and even in areas of retina in which they are numerous, they are sharply confined to the layer of ganglion cell axons. It is suggested that retinal astrocytes are “immigrant” fibrous astrocytes that enter the retina with its vasculature.
Key words: astrocyte origin, neuroglia, blood vessels, Mueller cells, axon bundles
The presence of astrocytes in the nerve fibre and ganglion cell layers of the retina has been established for some time, and the affinity of astrocyte processes for blood vessels of the central nervous system has also long been recognised. The pattern of vascularisation of the mammalian retina and the variation of that pattern between species provide an opportunity to examine the structural relationships of astrocytes in added detail. The retinas of some mammals, such as the possum (Freeman and Tancred, ’78), contain virtually no blood vessels, receiving nutrients by diffusion from the choroid. These retinas are perhaps the only parts of the mammalian central nervous system not vascularised (i.e., lacking penetrating vessels) in the adult. In other mammals, such as the rabbit (Davis ’29), the retina is vascularised only over a central portion, near the optic disc. The retina is fully vascularised in the rat and cat, but even in these retinas the vessels are strictly segregated to partic- ular retinal layers. Vessels larger than capillaries are found only in the nerve fibre and ganglion cell layers, and only capillaries extend into the inner plexiform and inner nu- clear layers (Michaelson, ’54). The outer plexiform and outer nuclear layers are free of vessels in, probably, all mammals.
In addition, the ganglion cell layer of the retina in many species contains large concentrations of cells whose axons are sequestered to the more superficial layer of axon bun- dles. This concentration and separation allow an assess- ment of the relationship between retinal astrocytes and the different portions of ganglion cells.
Our observations lead us to suggest that the spread of astrocytes in the retina is facilitated primarily by their
0 1987 ALAN R. LISS, INC.
affinity for vessels and secondarily by their affinity for axon bundles. Their spread appears to be strongly inhibited by the somas and (perhaps) dendrites of ganglion cells. Retinal astrocytes may be Ymmigrant” fibrous astrocytes that fol- low blood vessels into the retina from the optic nerve. A brief report of some of the present findings has been pre- sented (Stone and Dreher, ’85).
MATERIALS AND METHODS Material
Eyes were obtained from common cats, from albino (Wis- tar) and hooded rats, from New Zealand albino rabbits, and from a brush-tailed possum (Trichosurus vulpecula). The possum eyes were kindly supplied by Dr. K.J. Sanderson. In all cases the eyes were removed after the animal was killed by an overdose of anaesthetic (halothane or Nembu- tal). In most cases the animal was perfused perimortem with 4% paraformaldehyde in a 0.1 M phosphate buffer at pH 7.4.
Preparation of monoclonal antibodies Cerebella from two adult rats were homogenized without
fixation in approximately 6 ml of physiological saline. The cell suspension so obtained was used as the antigen. For primary immunisation the cell suspension was mixed with an equal volume of complete Freund’s adjuvant (Common- wealth Serum Laboratories). Mice (BalbiC females, 8 weeks
Accepted July 11,1986.

36
old) each received an intraperitoneal injection of 0.4 ml of the diluted suspension. Secondary immunisations were per- formed 3 weeks later and again 3 days before the cell fusion. On each occasion each mouse received an intraperi- toneal injection of 0.4 ml of the original suspension (i.e., not diluted with adjuvant).
The mice were killed by cervical dislocation and their spleens were removed. Spleen cells were put into suspen- sion and fused with NS-1 myeloma cells by using 50% polyethyleneglycol (MW 4,000, Merck) as a fusing agent. The fused cells were cultured in Dulbecco's MEM (Gibco) containing 15% foetal calf serum, 10-4M hypoxanthine, 4 x 10-7M aminopterin, and 1.6 x 10-5M thymidine. Super- natants from resultant hybridomas were screened for anti- body activity by indirect immunofluorescence staining of sections of rat cerebellum and retina. Cell lines of interest were cloned three times by the technique of limiting dilutions.
The effectiveness of the antibodies was enhanced by the production of antibody-rich ascites fluid. Mice were pre- treated by intraperitoneal injection of 0.5 ml pristane (Sigma) and 5 days later hybridoma cells (lo7 cells/animal) were injected. After collection, the ascites fluid was clari- fied by centrifugation and used for all tests at optimal dilution.
Screening and staining Both the screening of supernatants and the assessment of
the specificity of antibodies obtained were performed with indirect immunofluorescence on frozen sections, employing an intermediary biotidstreptavidin complex (Amersham).
Tissue (cerebellum or eyes) was fixed by vascular perfu- sion with 4% paraformaldehyde in 0.1 M phosphate buffer, preceded by phosphate buffer (both at pH 7.4). The tissues were promptly dissected free and postfixed for 1 hour in the same solution. They were then placed in 1 M sucrose in phosphate buffer at 4°C until they sank (usually over- night). They were frozen by contact with a metal plate held on the surface of liquid nitrogen and then sectioned at 10 pm in a cryotome.
The following protocol gave reliable detection of antibody binding on sectioned tissue, with dilutions of ascites fluid of up to lop3. All reagents were diluted in phosphate- buffered saline (PBS) at pH 7.4 containing 1% (wh) bovine serum albumin. Each section was:
J. STONE AND Z. DREHER
1. Air-dried on a gelatinised slide, kept at -20°C until needed, and then washed (three changes each of 30 seconds) in PBS.
2. Wiped free of excess buffer, covered with the solution of primary antibody, and incubated at 37°C for 1 hour. The volume used was that sufficient to cover the section.
3. Washed (3 x 30 seconds) in PBS, wiped free of excess buffer, covered with biotinylated antimouse Ig whole anti- body (from sheep, 1 5 0 dilution), and incubated at 37°C for 30 minutes.
4. Washed again in PBS (3 x 30 seconds), covered with fluorescein-streptavidin (1: 100 dilution), and incubated at room temperature for 15 minutes.
5 . Washed in PBS (3 x 30 seconds) and mounted in a 1:2 mixture of PBS and glycerol.
Immunocytochemistry of wholemount
mounted retinas. Each retina or piece of retina was: The above procedure proved readily adaptable to whole-
1. Washed in PBS containing 1 % Triton (2 X 10 minutes). Each was placed free-floating PBS, and washing was facili- tated by placing the dish on a gently rocking surface.
2. Placed in sufficient primary antibody to allow full immersion and incubated overnight at 4°C.
3. Washed in PBS, as above. 4. Immersed in biotinylated antimouse Ig whole antibody
5. Washed again in PBS, as above. 6. Immersed in fluoresceine streptavidin (1:lOO dilution)
for 30 minutes at room temperature. 7. Washed in PBS, as above, spread carefully on a glass
slide, and mounted in glycero1:PBS.
(150 dilution) for 2 hours at 37°C.
In many staining runs we compared the staining obtained with and without pretreatment with Triton (1% in all buff- ers and antibodies), but we noted no consistent improve- ment.
RESULTS Identification of astrocytes
Astrocytes were made visible using an anti-GFAP rabbit serum, an anti-GFAP monoclonal antibody (Labsystems), and a monoclonal antibody developed in this laboratory (designated MA1). The appearance of astrocytes was essen- tially identical with the three antibodies (compare, for ex- ample, Fig. 1D and E); MA1 may bind to the intracellular filaments that contain GFAP. It demonstrated astrocyte morphology in detail equivalent to that obtained with anti- GFAP antibodies, and demonstrated Miiller cells more clearly.
Astrocytes were originally named from the starlike shape created by their radiating processes, and some recent stud- ies (e.g., Ogden, '78) have continued to identify them by their overall morphology. In most recent studies, however, astrocytes have been characterised by different criteria, reviewed in Korr ('86): by their soma and nuclear morphol- ogy as seen in Nissl-stained sections; by their cytoplasmic content of filaments and organelles as seen in the electron microscope (for retinal astrocytes see, for example, Hogan and Feeney, '63-c; Ikui et al., '76; Bussow, '80); and by their content of intermediate filaments, especially vimentin (con- cerning retinal astrocytes see Shaw and Weber, '84; Schnitzer, '85) and GFAP (Bignami and Dahl, '79; Shaw and Weber, '84; Bjorklund and Dahl, '85; Bjorklund et al. '85). Eng ('85) has concluded, for example, that GFAP is the
Fig. 1. Astrocytes in cat retina demonstrated in wholemount prepara- tions. A. Near the edge of the retina astrocytes are found in considerable numbers. They spread in the innermost retinal layers, with their processes overlapping. B. Individual astrocytes have many profuse, radially oriented, and sparsely branched processes, which give the cell a starlike shape. C. Individual astrocytes may wrap both blood vessels and axons bundles. For example, processes of one astrocyte (upper left arrow) run along the axon bundle that crosses this photomicrograph from top to bottom. The same cell sends processes to contact (lower right arrow) a blood vessel that curves obliquely from upper right to lower left. Note that this vessel, located in the fibre raphe temporal to the area centralis, is only partially invested by astrocytes. D. Astrocyte processes wrapping a relatively large, branching vessel. The processes also run along fibre bundles. Where they approach the vessel at right angles they wrap around it without changing direction. Where the processes follow axon bundles running parallel to the vessel, short branches extend from them to wrap around the vessel. E. The pro- cesses wrapping larger vessels often show expanded tips. F. Astrocyte pro- cesses wrapping axon bundles and blood vessels. Note again the strong tendency for processes to extend along bundles, and around vessels. Anti- bodies: A-D were obtained with the monoclonal anti-GFAP E and F were obtained with monoclonal antibody MA1. Magnification: A- x 87.5, B,C,E- x350, D,F-x175.

Figure 1


RELATIONSHIPS OF RETINAL ASTROCYTES 39
Fig. 10. Individual processes do not spread significantly along the vessel.
Relationship to fibre bundles. Astrocytes investing the bundles of axons in the nerve fibre layer of retina show a strong tendency to spread along the length of the bundle (Fig. 1C-F). As a consequence, the pattern of axon bundles across the surface of the retina was readily apparent in the pattern of astrocytes (Fig. 2). The bundles can be traced to the disc from the edge of the retina, from the area centralis (Fig. 3A), and from the raphe of axons that runs temporal and superior from the area centralis (Figs. 2, 3B,C).
Where bundles are sparse the relationship between an individual astrocyte and a bundle can be seen (Fig. lC , arrowed). Processes stream along the length of axon bun- dles in a relation quite distinct from that between these processes and vessels. The processes wrap around vessels but along bundles.
Relationship to nerve cell somas. Although astrocytes invest much of the intraretinal length of ganglion cell axons, they do not invest their somas. Indeed, retinal astro- cytes seem to avoid the cell bodies and dendrites of all nerve cells, being absent from layers of the retina external to the axon layer, and from most regions of the ganglion cell layer. The only region of the retina in which they are present in substantial numbers in the ganglion cell layer is near the edge of the retina (Fig. lA), where the ganglion cells are very sparse.
Observations of as- trocytes in the nonuniform regions of the retina give addi- tional information on the constraints that determine their distribution. At the optic disc, for example, the plexus of astrocytes seems particularly dense, presumably in re- sponse to the concentration of vessels and axon bundles a t the disc.
At the area centralis, by contrast, no astrocytes are found (Figs. 2, 3A). Astrocytes spread along the axon bundles that run from the area centralis to the disc, but neither the ganglion cell somas which concentrate there nor the plexus of capillaries which spreads just deep to the concen- tration of ganglion cells at this site (Stone, '81, Fig. 3) attracts astrocytes. Astrocytes are apparently not needed for the function of this most specialised region of the retina.
At the edge of the retina, equally intriguingly, astrocytes are found in considerable numbers (Fig. lA,B), extending to within a few microns of the edge of the retina. As already noted, these very peripheral astrocytes show the classic asteroid shape, undistorted by vessel or bundle. The significance of this lack of commitment is considered in the Discussion.
Axons from ganglion cells in retina temporal to the area centralis do not course over the area centralis on their way to the disc, but arch below or above it. Axons from ganglion cells above a watershed or raphe arch above the area cen- tralis; those from ganglion cells below the watershed arch below (Bishop et al. '62; Murakami et al. '82). Astrocytes follow these axon bundles, but close to the area centralis only a few scattered astrocytes are found within the raphe (Figs. 2, 3A). Further peripherally, more and more astro- cytes extend into the raphe (Fig. 3A-C). As already noted, a few relatively large vessels cross the raphe, but they do not attract astrocytes into the region (Fig. 1C). The signif- icance of these observations is also considered in the
Relationship to retinal landmarks.
major protein of intermediate filaments of differentiated astrocytes. It remains untested whether the populations of cells distinguished by these several criteria are identical, or close to identical. AS Korr ('86) noted, this uncertainty needs to be recognised in relating new findings to old. In this paper we use the term astrocytes to refer to cells that (1) were immunoreactive to anti-GFAP antibodies or to MA1 and (2) showed a starlike morphology or some recog- nisable modification of it.
Cat Appearance of individual astrocytes. Astrocytes with the
characteristic starlike shape were best seen in peripheral regions of the retina (Figs. 1A-C, 3B,C). Their somas are small (< 10 pm in diameter) and inconspicuous. Their pro- cesses are profuse, little branched, and radial in orienta- tion, generating an asteroid appearance. Where processes from neighbouring cells overlap they intertwine freely. They resemble the fibrous form of astrocyte distinguished in the brain, lacking the vellate processes considered characteris- tic of the protoplasmic form (see, for example, Figs. 8 and 9 in Varon and Somjen, '79).
In other areas of retina, the asteroid shape of astrocytes is distorted and lost as processes wrap around blood vessels and fibre bundles (Fig. 1D-F). Both large vessels and large bundles attract astrocytes in such numbers that individual cells contributing to the astrocyte network cannot be distin- guished. In favourable situations, however, it is apparent that individual cells are free to invest both vessels and bundles. In some situations, different processes of one cell (arrows in Fig. 1C) could be seen passing to vessels and bundles. In other circumstances (Fig. lD-F), the same pro- cesses could be seen passing from bundle to vessel.
Distribution over retina Relationship to blood vessels. The larger blood vessels of
the cat retina are, with a few exceptions discussed below, strongly invested by processes with GFAP-like immunore- activity (Fig. 1D-F). Occasionally both vessels of a vein-ar- tery pair were seen to be invested. Vessels as small as capillaries were not observed to be invested by these processes.
The geometry of the investment is of interest. Where astrocyte processes follow an axon bundle across a vessel, they wrap around the vessels without deviation (Fig. lD,F). Where the processes follow a bundle parallel to a vessel, short processes diverge from the bundle to wrap part way around the vessel (Fig. 1D-F), often ending in small expan- sions (Fig. 1E). Thus astrocyte processes wrap around vessels.
Not all larger-than-capillary vessels in cat retina are strongly invested by astrocytes. Where vessels pass across the astrocyte-sparse raphe region (described below, Fig. lC), they are invested only very partially and the contribution of a single astrocyte to a vessel could be seen (mowed in
Fig. 2. Astrocytes in area centralis region of cat retina. The montage shows the area centralis region of the right retina of a cat. The asterisk indicates the location of the peak of ganglion cell density, estimated from previous analyses (e.g., Stone, '65). The borders of the montage are approx- imately vertical and horizontal on the retina; the arrow points to the optic disk. Astrocytes follow the axon bundles toward their cell bodies and are present in considerable numbers where the axon layer is thick. Where it is thin, as at the area centralis and in the raphe region temporal to the area centralis, astrocytes are found only in small numbers. T indicates temporal, N nasal. Antibody: monoclonal anti-GFAP. Magnification: X87.5.
Discussion. Comparison with Miiller cells. The distribution of reti-
nal astrocytes differs markedly from that of the radial glia

Figure 3

RELATIONSHIPS OF RETINAL ASTROCYTES 41
not being attracted to axon bundles in the nerve fibre layer of the retina. Only near the disc, where the axon bundles are thickest, was there any tendency for astrocytes to fol- low axon bundles (Fig. 5A,B).
Relation to nerue cells. As in the cat, astrocyte processes do not invest the somas of ganglion cells, or of other nerve cells. They are sharply confined to the innermost layers of the retina.
Relation to retinal landmarks. As in the cat, astrocytes are numerous near the optic disc. Presumably because there is no discontinuity in the axon layer or blood vessel pattern associated with the region of maximal ganglion density in the rat retina, the distribution of astrocytes is continuous over that region. Astrocytes do extend to the edge of the retina, although their numbers there are not as high as in the cat.
Distribution across thickness of retina As in the cat, astrocytes are confined to the innermost lamina of the retina, the axon layer Fig. 5C). In freshly fixed tissue the processes of Mueller cells could be seen spanning the width of the retina, both in sections and at the broken edge of a wholemount preparation (Fig. 5C,D).
Rabbit Topographg. The rabbit retina is distinctive in several
ways. First, the major nonuniformity in the ganglion cell layer is a narrow ridge of high ganglion cell density that stretches horizontally across the retina, as indicated in Figure 6. This ridge has recently been termed the visual streak (Hughes, '71; Provis, '79). Because of this concentra- tion, the convergence of ganglion cell axons on the optic disc also has a strong horizontal orientation. Further, many of the axons become myelinated before they reach the optic disc, creating a horizontally oriented white band on the retina, apparent in the living eye. It was this white band that Davis ('29) originally referred to as the visual streak. In addition, the retina is vascularised over the area of the white band, but not in more superior or inferior retina.
Figure 7A shows a region at the inferior margin of the vascularised area of a Nissl-stained rabbit retina. The most distal vessels loop at this margin (Fig. 7A, arrows) and turn back toward the optic disc. Fibre bundles, on the other hand, originate in more peripheral, nonvascular regions of the retina, such as the region of high ganglion cell density (the visual streak) apparent at the bottom of Figure 7A. Figure 7B and C compares sections of vascular and avas- cular regions of the rabbit retina. In C the layer of myelin- ated axons (labelled a) is as thick as the remainder of the retina.
Distribution of astrocytes. With anti-GFAP antibodies and MA1, astrocytes can be seen to wrap both axon bun- dles and vessels. As reported by Schnitzer ('85), the astro- cytes are sharply confined to the vascularised area of retina, and a border between astrocyte-rich and astrocyte- free areas of retina is very obvious (Fig. 8A,B).
Relation to axon bundles. As in the cat retina, astro- cytes show a strong affinity for axon bundles, streaming along them and providing a strong display of the pattern of bundles in the axon layer (Fig. 8A). They are so densely arranged that it is difficult to see the contribution made by an individual astrocyte to the overall pattern, At the edge of the astrocyte-rich area, however, some impression can be gained of the cell bodies and processes of individual astrocytes (Fig. 8A,B). Astrocyte somas are small (< 10 pm in diameter) and inconspicuous, and give rise to un-
of the retina, the Muller cells. The inner feet of Miiller cells stain with both anti-GFAP antibodies and with MA1- more effectively with the latter. In wholemount prepara- tions, the inner feet are seen (Fig. 3E,F) to form a regular mosaic that extends, apparently unbroken, across the ret- ina. They do not show the variations in distribution appar- ent among astrocytes. Figure 3E shows astrocyte processes streaming freely just internal to the inner feet of Muller cells. The distributions of the two cell classes seem independent.
Distribution of astrocytes across thickness of retina Pro- cesses demonstrated in cat retina with MA1 take two forms (Fig. 3D,E). Many spread along the inner surface of the retina, close to the inner limiting membrane; these are the processes of astrocytes. In addition, the inner feet of Muller cells appear as conical elements with their base against the inner limiting membrane. Occasionally, the somas of the Miiller cells could be detected (arrowed in Fig. 3F). The staining of the Muller cells was particularly sensitive to delay after fixation and freezing. Even in the freshest material, however, the outer lengths of Miiller cells were not stained. The Miiller cells are seen with particular clar- ity at the broken edge of whole-mounted retinas (Fig. 3F), perhaps because the cells remain unsectioned.
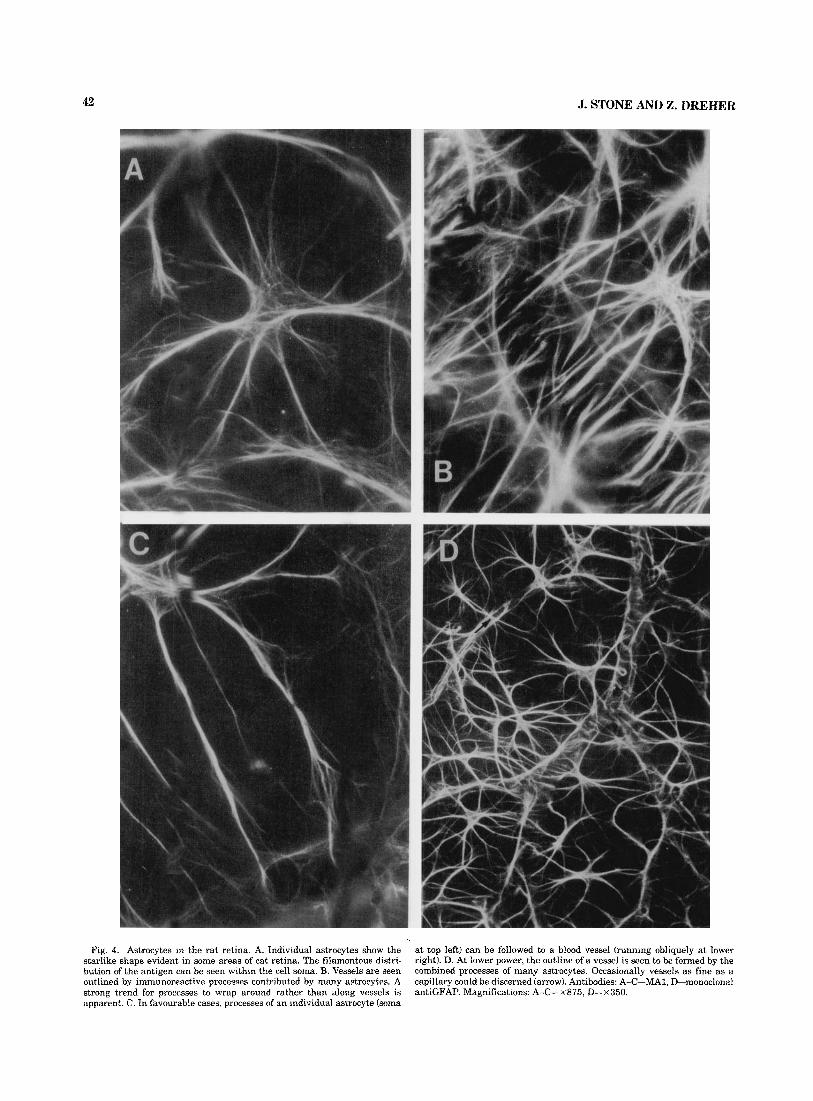
Rat Appearance of individual astrocytes. Individual astro-
cytes in rat retina (Fig. 4A) are very similar t o those of the cat. Their processes are also profuse and radiate from the cell soma, creating a starIike shape and resembling the fibrous rather than the protoplasmic astrocytes of the brain. Their somas are small (< 10 pm in diameter) and inconspi- cuous. We did not note any differences between the retinas of albino and pigmented animals in the morphology or distribution of astrocytes.
Distribution over the retina Relation to blood uessels. The relationship of astrocytes
to blood vessels in rat retina is very similar to that seen in the cat. Astrocytes show a strong affinity for vessels larger than capillaries, their processes tending to wrap around the vessels (Fig. 4B,C). All vessels radiating from the disc were invested by astrocytes, indicating that both veins and arter- ies attract astrocytes. Vessels of capillary size were occa- sionally seen to attract astrocyte processes (Fig. 4D, arrow), but only in the nerve fibre layer.
Relation to oxon bundles. Astrocytes of the rat retina differ from those of cat (above), rabbit (below), and human (Provis, Penfold, and Stone, unpublished observation) in
Fig. 3. Astrocytes and Mueller cells in cat retina. A. The area centralis region. Note that astrocytes extend along axon bundles toward the area centralis and above and below it. Astrocytes are not found, however, in the region of peak ganglion cell density, indicated by the asterisk. B. In the fibre raphe, 3.3 mm temporal to the area centralis, astrocytes are found in a sparse array. C. In the fibre raphe 8 mm temporal to the area centralis, astrocytes are present in considerable numbers. Estimated from maps of ganglion cell distribution in cat retina, the density of ganglion cells in A reaches about 8,000/mm2, 500-1,000/mmz in B, and 300-500/mm2 in C. D. The inner half of the thickness of the retina is traversed by the radially oriented inner feet of Mueller cells. E. The inner feet of Mueller cells form a fine, unbroken mosaic. Astrocytes appear to run freely between the end feet and the inner surface of the retina. F. At the broken edge of a whole- mount preparation the pads formed by individual Mueller cells are seen, together with their inner process stretching across thickness of the retina. The arrow points to what may be the soma of a Mueller cell. Antibodies: A- C, monoclonal anti-GFAP; D-F, MA1. Magnifications: A-C- ~ 8 7 . 5 , D,E- X 350, F-X500 (approximate).
42 J. STONE AND Z. DREHER
Fig. 4. Astrocytes in the rat retina. A. Individual astrocytes show the starlike shape evident in some areas of cat retina. The filamentous distri- bution of the antigen can be seen within the cell soma. B. Vessels are seen outlined by immunoreactive processes contributed by many astrocytes. A strong trend for processes to wrap around rather than along vessels is apparent. C. In favourable cases, processes of an individual astrocyte (soma
at top left) can be followed to a blood vessel (running obliquely at lower right). D. At lower power, the outline of a vessel is seen to be formed by the combined processes of many astrocytes. Occasionally vessels as fine as a capillary could be discerned (arrow). Antibodies: A-C-MA1, D-monoclonal antiGFAP. Magnifications: A-C-X875, D-x350.

43 RELATIONSHIPS OF RETINAL ASTROCYTES
branched, radiating processes. The processes were often seen to terminate in small pads (Fig. 8B), which are pre- sumably apposed to distal blood vessels or to axon bundles.
The failure of astrocytes to follow axon bundles into avascular areas of retina has already been noted (above; Stone and Dreher, '85; Schnitzer, '85). It deserves empha- sis: the axon bundles continue without break into more peripheral retina. Presumably, the glial class investing them changes abruptly from astrocytes to Miiller cells.
Relationship to blood vessels. As in the cat and rat, the blood vessels in rabbit retina larger than capillaries at- tract a substantial investment of GFAP-positive processes (Fig. 8D). It was always more difficult, however, to trace processes from astrocyte somas to blood vessels, partly because of the density of astrocytes in this region and partly because the blood vessels were situated slightly deep to the axon bundles.
Distribution across thickness of the retina. In sections, the processes of astrocytes are seen (confirming Schnitzer, '85) to be restricted to the axon layer of retina (Fig. 8C), as in the cat and rat. The border of the region containing astrocytes was easy to identify in sections; close to the bottom of Figure 8C, for example, the astrocyte processes prominent a t the inner surface of the retina fade away. The inner feet of Miiller cells were often apparent, partic- ularly when MA1 was used. The Miiller cells appear to be distributed relatively evenly across the astrocyte border and across the whole retina (again confirming Schnitzer, '85).
Possum In the adult possum, retinal vasculature is confined to
the optic disc and a narrow annulus immediately around it (Freeman and Tancred, '78; Fig. 7D). Correspondingly GFAP+ve processes were found only within the disc and immediately adjacent to it. Most immunoreactive elements were fibres oriented circumferentially within the disc and radially just outside it (Fig. 7E). These circumferential and radial orientations are apparent in the nuclei stained with cresyl-violet material as well (Fig. 7D). More than a few hundred microns from the margin of the disc, no GFAP+ve material was apparent.
DISCUSSION Ideas of the function of astrocytes and of macroglia in
general have multiplied in recent years (reviewed in Varon and Somjen, '79). Astrocytes contribute to the glial invest- ment of blood vessels and to the glia limitans at the pial surface of the brain. They react to tissue damage by hyper- trophy and phagocytosis, and perhaps by proliferation. Ma- croglia, presumably including astrocytes, are involved in the uptake and recycling of transmitters and are also con- sidered to act as K+ sinks during neural activity. Further, studies of the embryology of neuroglia indicate that cells of the astrocyte line form the radial glia of the telecephalon, which guide migrating neuroblasts to their place in the cortical plate (Schmechel and Rakic, '79; Rakic, '81) before transforming into adult astrocytes. Some of the present observations, in particular the absence of astrocytes from avascular regions of retina, emphasise the vascular orien- tation of astrocyte function. Overall, however, the present observations raise two possibilities we did not anticipate: that retinal astrocytes invade the retina from the optic nerve, and that, even where they are present in consider-
able numbers, they are not essential to the function of the retina.
Morphology of retinal astrocytes In all three species (rabbit, cat, rat) in which astrocytes
were recognised in the retina, they resembled the fibrous astrocytes of brain tissue, with small (10 pm or less in diameter) somas and radiating processes that lacked the vellate structures characteristic to protoplasmic astrocytes. As Ogden ('78) noted in the primate retina, the processes of many astrocytes in all three species vary from this shape in that they are moulded to axon bundles or vessels. Ogden concluded that most (93%) of the astrocytes he observed in Golgi-impregnated wholemount preparations of the mon- key retina seemed to be related to be either axon bundles or vessels; only a small minority seemed to related to both. Our observations in the cat, however, indicate that in this species at least individual astrocytes invest both vessels and axon bundles. Different processes of one cell may pass to axons or fibre bundles (Fig. lC), or the same processes may pass from axon bundle to vessel (Fig. 1E).
Retinal distribution of astrocytes: Previous observations
Ikui et al. ('76) have noted an absence of astrocytes from the ora serrata and foveal regions of monkey retina, basing the identification of astrocyte processes and their distinc- tion from Miiller cell processes on the filament and organ- elle content of their cytoplasm. The absence of astrocytes from the centre of the foveal region correlates well with the absence of blood vessels from the outer plexiform and outer nuclear layers of all primate retinas; these are the only layer present at the foveola. Ikui et al.'s observation prob- ably referred also to the concentration of ganglion cells around the foveola. If so it is matched by the present obser- vation that retinal astrocytes are not found in the concen- tration of ganglion cells at the cat area centralis. In the rat and cat, whose retinas resemble the primate in being vas- cularised to their edge, astrocytes do extend to the edge of the retina. Our observations in the rabbit and possum go beyond Ikui et al.'s in indicating an absence of astrocytes from large areas of retina in which the full neuronal ma- chinery of the retina is present, but no blood vessels.
Ogden ('78) has noted the concentration of astrocytes in the fibre bundles of the retina, an observation strongly confirmed here. Kondo et al. ('84) have noted that the pro- cesses that wrap capillaries in the middle layers of the retina are derived from Muller cells rather than astrocytes. This idea is also confirmed by the present study.
Schnitzer ('85) has recently and independently described the restriction of astrocytes to the "medullary ray" region of the rabbit and, within that region, to the innermost layers of the retina.
The GFAP-like immunoreactivity of retinal glia In retinal astrocytes, GFAP-like immunoreactivity has
been observed consistently in a range of species (e.g., Big- nami and Dahl, '79; Shaw and Weber, '84; Bjorklund and Dahl, '85; Bjorklund et al., '85). In Miiller cells, however, early reports suggested an absence of GFAP-like immuno- reactivity (reviewed in Shaw and Weber, '841, while more recent work with a greater range of antibodies has reported its clear presence (Bignami, '84; Bjorklund and Dahl, '85; Bjorklund et al, '85). In our material Miiller cells could not

44 J. STONE AND Z. DREHER
Figure 5

RELATIONSHIPS OF RETINAL ASTROCYTES 45
- - - -
- Visual strea u - Visual streak
5rnm
Fig. 6. The vascular region of the rabbit retina. The drawing shows the outline of a wholemount preparation of a rabbit retina (kindly provided by J.M. Provis). The retina was cut radially in several places to facilitate flattening. The optic disc is represented, as is the distribution of major vessels. These are confined to a horizontally oriented strip extending tem-
be demonstrated reliably with anti-GFAP antibodies. They were demonstrated much more reliably with MA1, and this was the major difference we noted between MA1 and the anti-GFAP antibodies.
Factors controlling the distribution of retinal astrocytes
The present observations lead us to make five related hypotheses concerning the distribution of retinal astrocytes:
1. They are not derived from the germinal epithelium of the retina but invade the retina from the optic disc, in company with the vessels of the retinal circulation.
Fig. 5. Astrocytes and Muller cells in rat retina. A. The optic disc region of the retina. The disc is the pale, fluorescent circle at centre, from which vessels radiate. The vessels are seen because they are outlined by the processes of astrocytes. The disc is seen because the lamina cribrosa region is rich in GFAP-positive processes. The intensely fluorescent structures a t the inner surface of the disc appear to be remnants of the hyaloid vessels. The arrow indicates the position of the region shown at higher power in B. B. In both pigmented and albino rats, the astrocytes did not strongly invest axon bundles. Only in occasional patches, such as this region, did astrocyte processes appear to stream along axon bundles, as they do in cat and rabbit. C. In sections, immnnoreactive processes running parallel to the surfaces of the retina are found only at its inner surface (at top). In favonrable circum- stances the radial process of Miiller cells could be discerned, spanning the retina. D. At the broken edge of a wholemount retina, the astrocytes spread- ing on the surface of the retina (upper left) and the radially oriented processes of Muller cells (lower right) could both be seen. The latter span the thickness of the retina. Antibody: A,B: MA1, C,D: monoclonal anti- GFAP, with Zamboni's fixative. Magnification: A-X87.5, B-X875, C,D- x350.
porally and nasally from the optic disc. The pattern of axon bundles over ;he vascular region is also shown. The dotted line represents the ridge of the horizontally oriented region of high ganglion cell density, the visual streak. The rectangle shows the region of retina shown in Figure 7A.
2. They are primarily attracted by blood vessels and sec- ondarily by the axons of retinal ganglion cells.
3. Their spread in the retina is inhibited by the somas of ganglion cells and perhaps by other neurone classes as well. 4. They are fibrous rather than protoplasmic astrocytes,
probably derived from fibrous astrocytes of the optic nerve. 5. They are not essential for retinal function.
The origin of retinal astrocytes There has been little discussion of this question in the
literature we have surveyed. There seem to be two possibil- ities. Astrocytes may form from the Muller cells of the retina, just as telencephalic astrocytes form from radial glia (Schmechel and Rakic, '79). Alternatively, astrocytes may invade the retina, perhaps following the disc-centred for- mation of blood vessels. We propose the latter hypothesis, for several reasons. First, astrocytes are not found in avas- cular retinas, or in avascular regions of partially vascular- ised retinas. The formation of retinal vessels is thus a prime factor in the distribution of astrocytes, and that formation is by way of invasion from the optic disc. Second, in a still- preliminary developmental study on the cat retina, we have noted that the appearance of astrocytzes matches the spread of vasculature, beginning in late embryonic life and spread- ing from the disc to peripheral retina (Halasz and Stone, '83). Third, astrocytes are tightly restricted to the inner- most layers of the retina. This suggests that the cells reach the retina at its inner surface and are inhibited from mov- ing out. Finally, if astrocytes are considered to be derived from the retina, it is necessary to postulate that the retina

Fig. 7. A. Region of a wholemount cresyl-violet-stained preparation of rabbit retina (indicated by rectangle in Fig. 6). The obliquely oriented structures at top are axon bundles, among which blood vessels are apparent. The most distal vessels are arrowed. Further from the disc, no vessels are present. Ganglion cell density is high at the bottom of the photomicrograph, which shows part of the visual streak. B,C. B shows the three main cellular layers of rabbit retina in a region peripheral to the vasculature. The gan- glion cell layer is at top, the pigment epithelium at bottom. Nearer the disc (C), a thick layer of axons (a) is present, with blood vessels (arrow) at the inner surface. Haematoxylin and eosin. D. Optic disc region of the possum retina, seen in a cresyl-violet-stained wholemount (kindly made available
by Elizabeth Tancred). Part of the circumference of the disc is apparent at the bottom of the photomicrograph. The radial pattern of axons converging on the disc is faintly apparent. The looping vessel may he a remnant of the hyaloid circulation, or a vessel from the choroid circulation extruded through the disc. Such loops are present only occasionally; usually no vessels extend from the disc into the retina. E. GFAP-positive fibres at the edge of the optic disc of the possum retina. Some fibres run circumferentially around the edge; others extend radially into the retina for as much as 100 pm. More than a few hundred microns from the edge of the disc no GFAP-positive processes were apparent. Magnifications: A,D- X 150, B,C-approximately x700, E-approximately x850.

RELATIONSHIPS OF RETINAL ASTROCYTES
Fig. 8. Astrocytes and Muller cells in the rabbit retina. A. Astrocytes in a region of rabbit retina comparable to that shown in Figure 7A. The astrocytes run along axon bundles and stop abruptly a t the edge of the vascular region. B. Individual processes of astrocytes terminate in small expansions. C. In sections, astrocyte processes appear as bright bundles at the inner surface of the retina; in this section the inner surface is a t left, the receptors a t right. This length of section spans the border of vascular (top) and avascular (bottom) areas of the retina. Thus the immunoreactive
47
processes prominent a t the inner surface at top are absent at bottom. The inner feet of Muller cells can be seen forming a regular, radially oriented array that extends unbroken across the astrocyte limit. D. Blood vessels were observed to be invested with GFAP-positive processes, although the contributions made by individual astrocytes could not be distinguished. Antibodies: Lower left-MAl, upper right-antiGFAP monoclonal, others- antiGFAP serum. Magnifications: A- x 175, B-approximately x 350, C- x 175, D-approximately ~ 8 5 0 .

48
generates a form of astrocyte that is incompatible with almost all its layers. The present suggestion is consonant with Bussow’s (’80) idea that the astrocytes of the retina and optic nerve head are a “special glia for ganglion cell axons.”
Role of Vasculature The primary importance of vasculature in the control of
astrocyte spread is suggested by the almost complete ab- sence of astrocytes from an avascular retina (possum), their restriction to the vascular region of the rabbit retina, and their lack of relation to axon bundles in the retina of the rat. Schnitzer (’85) and Stone and Dreher (‘85) have previ- ously commented on the close correlation between vascula- ture and astrocyte distribution in the rabbit retina.
Cellular inhibition of astrocyte spread This inhibition is suggested by three observations in the
cat retina. First, astrocytes are not found at the area cen- tralis. Second, astrocytes are found in the raphe at increas- ing frequency with distance from the area centralis; ganglion cell density decreases with distance. Third, astro- cytes are found in considerable numbers near the edge of the retina, where ganglion cells are very sparse. Appar- ently astrocytes spread freely in the axon layer but will not enter the ganglion cell layer except where ganglion cells are very sparse. In none of the species examined do astro- cytes enter or extend processes into deeper layers of the retina.
The “fibrous” nature of retinal astrocytes The morphology of retinal astrocytes resembles that of
fibrous rather than protoplasmic astrocytes (see for exam- ple Figs. 8 and 9 in Varon and Somjen ’79). The restriction of retinal astrocytes to the axon layer of the retina resem- bles the restriction of fibrous astrocytes to white matter of the brain; and their affinity for axons is obvious, except in the rat. The astrocytes of the optic nerve are mainly (but not exclusively) fibrous (Miller and Raff, ’84).
The function of astrocytes Several observations suggest that astrocytes are not es-
sential for retinal function. We do not suggest that, where they are present in the retina, astrocytes serve no func- tions. Rather we suggest that Miiller cells can perform all those functions and do so wherever astrocytes are absent. Thus (1) astrocytes are virtually absent from avascular retinas, and from the wide avascular regions of the rabbit retina, even though these regions contain the full neuronal machinery of the retina. (2) Astrocytes are absent from highly specialised regions of retina, such as the area cen- tralis of the cat retina and fovea centralis of the monkey retina (Ikui et al., ’76). (3) Astrocytes are absent from parts of large vessels that leave the axon layer (Fig. 1C). (4) Astrocytes are present in considerable numbers near the edge of the retina. (5) Miiller cells are present throughout all these retinas.
Perhaps the astrocytes of the retina are immigrants, fol- lowing vessels into the retina, and attracted by axon bun- dles, but essential for the function of neither.
ACKNOWLEDGMENTS We gratefully acknowledge the excellent technical assis-
tance of Mignon Wegner in the preparation and immuno- cytochemical processing of tissue and of Patrick de Permentier in the maintenance of the fluorescence micros-
J. STONE AND Z. DREHER
copy facility. M. Raff kindly made available the anti-GFAP serum. The work was supported by the National Health and Medical Research Council of Australia.
LITERATURE CITED Bignami, A. (1984) Glial fibrillary acidic (GFA) protein in Mueller glia.
Immunofluorescence study of the goldfish retina. Brain Res. 300:175- 178.
Bignami, A,, and D. Dahl(1979) The radial glia of Mueller in the rat retina and their response to injury-an immunofluorescence study with anti- bodies to the glial fibrillary acidic protein. Exp. Eye Res. 28t63-69.
Bishop, P.O., G.J. Vakkur, and W. Kozak (1962) Some quantitative aspects of the cat’s eye: Axis and plane of reference, visual field coordinates and optics. J. Physiol. (Lond.) 163.466-502.
Bjorklund, H., and D. Dahl(1985) Glial fibrillary acidic protein (GFAPI-like immunoreactivity in the rodent eye. Comparison between peripheral glia of the anterior uvea and the central glia of the retina. J. Neuro- immunol. 8~331-345.
Bjorklund, H., A. Bignami, and D. Dahl(1985) Immuno-histochemical dem- onstration of glial fibrillary acidic protein in normal ra t Mueller glia and retinal astrocytes. Neurosci. Lett. 54~363-368.
Bussow, H. (1980) The astrocytes in the retina and optic nerve head of mammals: A special glia for the ganglion cell axons. Cell Tissue Res. 206:367-378.
Davis, F.A. (1929) The anatomy and histology of the eye and orbit of the rabbit. Trans. Ophthalmol. SOC. UK 2i:401-441.
Eng, L.F. (1985) Glial fibrillary acidic protein (GFAP). The major protein of glial intermediate filaments in differentiated astrocytes. J. Neuroim- munol. 8.203-214.
Freeman, B.F., and E.J. Tancred (1978) The number and distribution of ganglion cells in the retina of the brush-tailed possum (Trichosurus uulpecula). J. Comp. Neurol. Iii:557-568.
Halasz, P., and J. Stone (1983) Development of retinal vasculature in the cat. Proc. Anat. Soc. Aust. N.Z. 35P.
Hogan, M.J., and L. Feeney (1963a) The ultrastructure of retinal vessels. I. The large vessels J. Ultrastruct. Res. 9~10-28.
Hogan, M.J., and L. Feeney (1963b) The ultrastructure of retinal vessels. 11. The small vessels. J. Ultrastruct. Res. 939-46.
Hogan, M.J., and L. Feeney (1963~) The ultrastructure of retinal vessels 111. Vascular-glial relationships. J. Ultrastruct. Res. 9t47-64.
Hughes, A. (1971) Topographical relationships between the anatomy and physiology of the rahhit visual system. Doc. Ophthalmol. 80:98-103.
Ikui, H., S. Uga, and T. Kohno (1976) Electron microscope study on astro- cytes in the human retina using ruthenium red. Opthalmol. Res. 8t100- 110.
Kondo, H., H. Takahashi, and Y. Takahashi (1984) Immunohistochemical study of S-100 protein in the postnatal development of Mueller cells and astrocytes in the rat retina. Cell Tissue Res. 238r503-508.
Korr, H. (1986) Proliferation and cell cycle parameters of astrocytes. In S. Fedoroff and A. Vernadakis (ed):Astrocytes. Orlando, Florida: Academic Press, (in press).
Michaelson, I.C. (1954) Retinal circulation in man and animals. Springfield, Illinois: Charles C. Thomas.
Miller, R.H., and M.C. Raff (1984) Fibrous and protoplasmic astrocytes are biochemically and developmentally distinct. J. Neurosci. 4~585-592.
Murakami, D., M.A. Sesma, and M.H. Rowe (1982) Characteristics of nasal and temporal retina in Siamese and normal cats: Ganglion cell compo- sition, axon trajectory and laterality of projections. Brain Behav. Evol. 21.67-113.
Ogden, T.E. (1978) Nerve fiber layer astrocytes of the primate retina: Mor- phology, distribution and density. Invest. Ophthalmol. Vis. Sci. 7~449- 510.
Provis, J.M. (1979) The distribution and size of ganglion cells in the retina of the pigmented rabbit: A quantitative study. J. Comp. Neurol. 185:121- 139.
Rakic, P. (1981) Neuronal-glial interaction during brain development. Trends Neurosci. 4:184-187.
Schmechel, D.E., and P. Rakic (1979) A Golgi study of radial glial cells in developing monkey telencephalon: Morphogenesis and transformatlon into astrocytes. Anat. Embryol. 156.115-152.
Schnitzer, J. (1985) Distribution and immunoreactivity of glia in the retina of the rabbit. J. Comp. Neurol. 240.128-142.
Shaw, G., and K. Weber (1984) The intermediate filament complement of the retina: A comparison between different mammalian species. Eur. J. Cell Biol. 33:95-104.

RELATIONSHIPS OF RETINAL ASTROCYTES 49
Stone J. (1965) A quantitative analysis of the distribution of ganglion cells
Stone, J. (1981) The Wholemount Handbook. Sydney: Maitland Publications.
Stone, J., and 2. Dreher (1985) Astrocytes in mammalian retina are related
to vascularisation. Proc. Aust. PhysioI. Pharmacol. SOC. 16:145P.
Res. Prog. Bull. 17. in the cat’s retina. J. Comp. Neurol. 148:l-32. Varon, S.S., and G.G. Somjen (1979) Neuron-glia interactions. Neurosci.





























