Embed Size (px)
Citation preview

J. Anat. (1980), 130, 1, pp. 55-61 55With 5 figuresPrinted in Great Britain
Specialized contacts of astrocytes with astrocytes and withother cell types in the hypothalamus of the hamster
I. SUAREZ NAJERA, B. FERNANDEZ RUIZAND L. M. GARCIA SEGURA
Cdtedra de Citologia e Histologia, Universidad de Alcald de Henares,Madrid, Spain
(Accepted 24 January 1979)
INTRODUCTION
Electron microscopy of nervous tissue has made it possible to see and describethe contacts existing between its cellular components. Intercellular junctions involvea localized modification of the plasma membrane. Revel & Karnovsky (1967)reported that the junctions between astrocytic processes were of 'gap junction'type. Inter-astrocytic gaD junctions have been found very frequently in the peri-vascular and subpial areas (Reese & Karnovsky, 1967; Brightman & Reese, 1967,1969).
Specialized contacts between astrocytes and neuronal processes have also beendescribed, namely by Palacios-Pru & Mendoza-Brinceino (1972), and Guldner &Wolff (1973). These authors refer to such contacts as 'synaptoides'. Morales &Duncan (1975) confirmed the presence of specialized astrocytic-neuronal contactsin the medulla of the cat, and pointed out the existence of gap junctions betweenastrocytes and oligodendroglial cells.
Sipe & More (1976) described inter-astrocytic junctions in the lateral hypo-thalamic area of the rat and their relationship to synaptic endings.
In this study we describe the various types of intercellular junction observed inthe hypothalamus of the hamster.
MATERIALS AND METHODS
Brains of hamsters, ofboth sexes, were fixed by perfusion with 2-5 % glutaraldehyde(Millonig buffer, at pH 7 3). After isolating the hypothalamus and dicing it into1 mm square blocks, the tissue was kept in cold fixative for 3-4 hours. Blocks werepost-fixed in 1 % osmium tetroxide, dehydrated in acetone and embedded in Araldite.Thin sections were stained with lead citrate and examined in a Jeol 100-B electronmicroscope.
OBSERVATIONS
Most junctions between astrocytes occur in the perivascular and subpial regions.They are characterized by parallel plasma membranes separated by a space varyingfrom region to region and usually occupied by material of slight electron density.These junctions are structurally equivalent to those between ependymal cells(SuArez & Fernandez Ruiz, 1978).Some astrocyte processes at the ventral surface of the hypothalamus have terminal
0021-8782/80/2828-6990 $02.00 C 1980 Anat. Soc. G.B. & L.
56
APW . ,,
I. SUAREZ NAJERA AND OTHERS
w "'''.~~~~~~~~~~~~~~~~~~~~ 44oe
- E. ;^|; v o.;s o*:_.4
- ..
. .
AP
-AP.'t, $--t' -;; g ?92@'§;>IS S
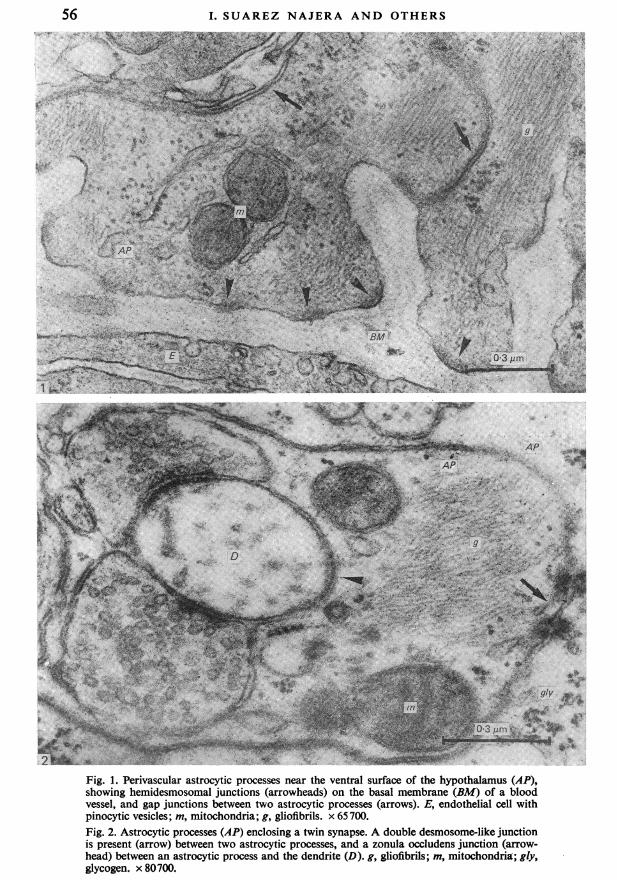
Fig. 1. Perivascular astrocytic processes near the ventral surface of the hypothalamus (AP),showing hemidesmosomal junctions (arrowheads) on the basal membrane (BM) of a bloodvessel, and gap junctions between two astrocytic processes (arrows). E, endothelial cell withpinocytic vesicles; m, mitochondria; g, gliofibrils. x 65700.Fig. 2. Astrocytic processes (AP) enclosing a twin synapse. A double desmosome-like junctionis present (arrow) between two astrocytic processes, and a zonula occludens junction (arrow-head) between an astrocytic process and the dendrite (D). g, gliofibrils; m, mitochondria; gly,glycogen. x 80700.
.Ip
t
1. ..1
I
go..sjk.
I
.1, ,-z
,l.*
3L , ,bc
11
-4.1-4ft, 1.0 -.e,
,e4.r.` " 91t I It..,A."i rl.

Specialized contacts of astrocytesexpansions where their plasma membranes are applied to the basal membranes ofblood vessels, forming perivascular feet. These plasma membranes have a somewhathigher electron density in some areas, forming junctions of hemidesmosomal type.But at the same time, such astrocytes also form junctions with each other in whichthe plasma membranes of two astrocytic processes participate (Fig. 1), both mem-branes being slightly more electron-dense than elsewhere. Sometimes the junctionsare of great length, but the local cytoplasm is not specialized; these are generallytermed 'gap junctions'.Desmosome-like junctions between two astrocytic processes are also frequent.
Figure 2 shows a double desmosome-like junction between astrocytic processesin the hypothalamic supraoptic nucleus, enclosing and isolating a twin synapse.Within this astrocytic process can be seen mitochondria. a cistema of rough endo-plasmic reticulum, a bundle of gliofibrils cut obliquely, and glycogen granules. Thesame micrograph shows a structural specialization of zonula occludens type betweenthe plasma membranes of the astrocytic processes sheathing the synapse and thedendrite.We have also observed another type of junction in which three or more processes
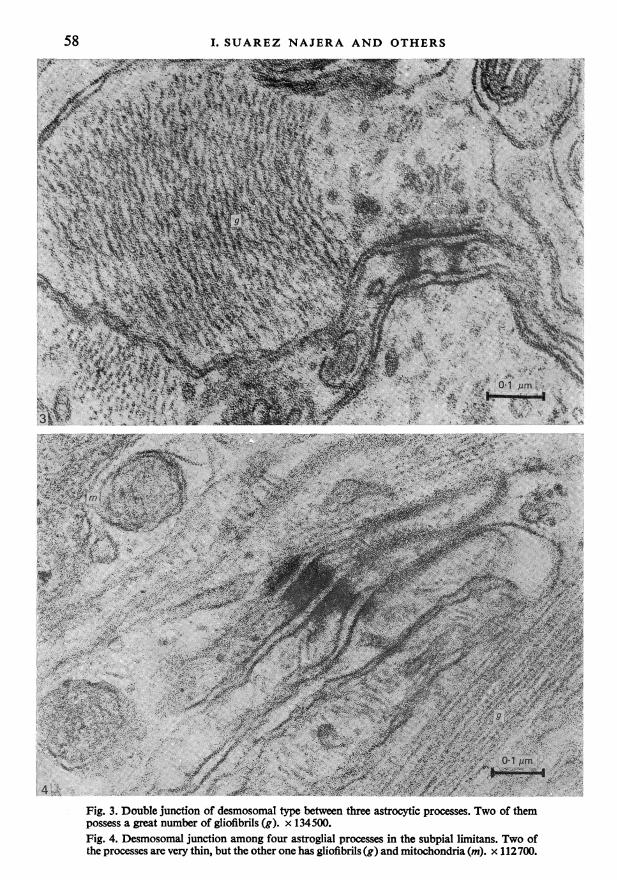
are related to each other. One of these was located in the upper zone of the diffusesupraoptic nucleus (Fig. 3). In this junction three glial processes make contact bymeans of a double membrane structure exhibiting high electron density, speciali-zation of the adjacent cytoplasm, and accumulation of dense material in theintercellular space. In this type of junction the cytoplasm shows greatly increasedelectron density compared to that of normal astrocytic cytoplasm, and forms abridge linking both astrocytic process extremities. The two processes participatingin the junction possess a great number of gliofibrils, and some transversely sectionedmicrotubules, and are situated amongst numerous non-myelinated fibres.
Astrocytic processes form a complete layer of subpial feet beneath the pia matercovering the ventral surface of the hypothalamus. These processes are laterallylinked to each other by many desmosomal junctions showing not only participationof plasma membranes and specialization of the two adjacent cytoplasms (Fig. 4).Since the two central astrocytic processes are very thin, the whole cytoplasm partici-pates in the desmosomal junction. Bundles of gliofibrils extend along the thickerperipheral processes, and in these glycogen granules, and mitochondria containinga dense matrix and numerous osmiophilic granules are also observed.The type of junction described above is very similar to the 'autodesmosome'
described by Alvarez-Uria (1972) in the infraorbital nerve of the rat in the Schwanncell at the level of the node of Ranvier. In this type ofjunction the terminal lamellaeof the myelin adjacent to the Schwann cell participate, and it is named for thisreason an 'autodesmosome'. Because of the difficulty of determining if all thesmall laminae observed by us belong to the same astrocyte, we cannot apply theterm autodesmosome to this type of junction without reservation. although itsmorphology resembles this type.
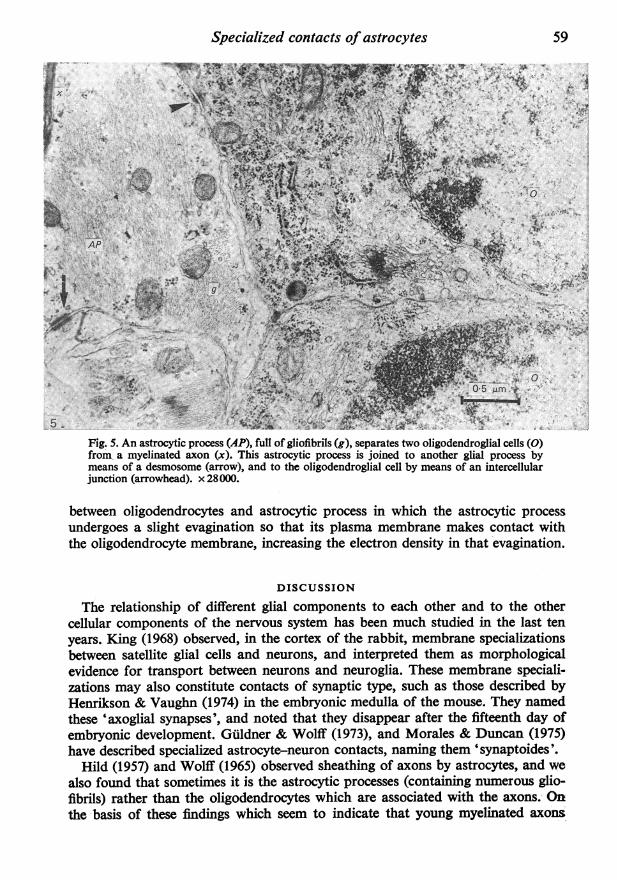
Morales & Duncan (1975) have described contacts between astrocytes and oligo-dendrocytes in the medulla of the cat. Our Figure 5 shows an intercellular junctionbetween an astrocyte process (containing numerous gliofibrils and mitochondria)and an oligodendrocyte. In this junction the plasma membranes of both structuresand the adjacent cytoplasms participate, but there are no accumulations of electron-dense material in the intercellular space as in the inter-astrocytic junction in theleft part of the same micrograph. We have also frequently observed contacts
57
58 I. SUAREZ NAJERA AND OTHERS
$,~~~~~~~' ''S!r .9- f e
'.'
4.
Fig. 3. Double junction of desmosomal type between three astrocytic processes. Two of thempossess a great number of gliofibrils (g). x 134500.Fig. 4. Desmosomal junction among four astroglial processes in the subpial limitans. Two ofthe processes are very thin, but the other one has gliofibrils (g) and mitochondria (m). x 112700.
Specialized contacts of astrocytesIX ; ~~~~~~~~~~~~~~~~~-,. , .iii 7*, , ..',,iX'-Q.'.' '.
VAA
N- t.44.--
AP
_-~ ~ ~ ~ f~;44Lz,, X,l-uF1t.sO+~ ~~,te 't o*0 & 1 1S': & / '' tti 4f4A, -,>>,, . ,~~~~~~~~~..v Aw
4' -~g'
Fig. 5. An astrocytic process (AP), full of gliofibrils (g), separates two oligodendroglial cells (0)from, a myelinated axon (x). This astrocytic process is joined to another glial process bymeans of a desmosome (arrow), and to the oligodendroglial cell by means of an intercellularjunction (arrowhead). x28000.
between oligodendrocytes and astrocytic process in which the astrocytic processundergoes a slight evagination so that its plasma membrane makes contact withthe oligodendrocyte membrane, increasing the electron density in that evagination.
DISCUSSION
The relationship of different glial components to each other and to the othercellular components of the nervous system has been much studied in the last tenyears. King (1968) observed, in the cortex of the rabbit, membrane specializationsbetween satellite glial cells and neurons, and interpreted them as morphologicalevidence for transport between neurons and neuroglia. These membrane speciali-zations may also constitute contacts of synaptic type, such as those described byHenrikcson & Vaughn (1974) in the embryonic medulla of the mouse. They namedthese 'axoglial synapses', and noted that they disappear after the fifteenth day ofembryonic development. Guldner & Wolff (1973), and Morales & Duncan (1975)have described specialized astrocyte-neuron contacts, naming them 'synaptoides'.
Hild (1957) and Wolff (1965) observed sheathing of axons by astrocytes, and wealso foud that sometimes it is the astrocytic processes (containinlg numerous glio-~fibrils) rather than the oligodendrocytes which are associated with the axons. Onthe basis of these findings which seem to indicate that young myelinated axons
59

I. SUAREZ NAJERA AND OTHERSare surrounded by astrocytic processes, we suggest that in certain circumstancesastroglia are in some way related to the myelinization process.
Certain junctions between astrocytes and oligodendroglial cells, interpreted as
'gap junctions' by Morales & Duncan (1975), have also been found by us in thehypothalamus of the hamster. However, we interpret them not as gap junctions,but as desmosome-like ones, since we observed accumulations of dense material bothin the astrocytic area and in that corresponding to the oligodendrocyte cytoplasm.The existence of tight junctions around the bouton-like synaptic endings was
reported by Gonzalez Aguilar & De Robertis (1963). These authors consideredthat the glial process around the synaptic complex would constitute a glial barrier,delaying and blocking the diffusion of transmitter released into the synaptic cleftto the intercellular spaces. By means of such 'synaptic barriers', the glial cellswere in a position to influence the physiology of synaptic transmission (Gray, 1961;De Robertis, 1965). We did not observe such tight junctions between glial processes
and bouton-like synaptic endings, but between glial processes and dendrite in thetwin synapse (Fig. 2). Such a type of junction should, besides influencing synaptictransmission by delaying diffusion of the transmitter, protect the synapse frombeing affected by any element or substance coming from the extracellular space.
Such protection could best be achieved by linking the plasma membranes so closelythat the extracellular space is wholly obliterated.Where the astrocytic processes extend in parallel towards the ventral surface of
the hypothalamus they are linked to each other by typical desmosomal junctions;but when these processes are of laminar form, they link to each other by severaldesmosomes at the same level (Fig. 4). We have already noted the similarity of thistype of junction to the autodesmosomes described by Alvarez Uria (1972). Auto-desmosomes are desmosomes situated between membranes of the same cell: in
such junctions the two parts of the desmosome are related to each other, butinour observations the relation is double since there are not only two hemidesmosomesof the same process in contact but also one of these hemidesmosomes is related tothe hemidesmosome of the parallel process. Probably these desmosomal formationsdescribed by us represent not only a stronger anchoring to maintain links with theastrocytic processes that run in parallel to form the subpial layer, but also in thisway isolate the nerve cells from the external environment, viz the cerebrospinalfluid. The fact that the processes of subpial astrocytes link to each other by suchstrong junctions reminds us, in spite of the morphological differences, of thejunctions between the ependymal cells that line the third ventricle (Suarez Najera,Fernandez Ruiz & Carrato-Ibainez, 1978), and which by means of their close contactprevent substances in the ventricular fluid coming into contact with the nerve cells.
SUMMARY
Adult hamsters were used for this electron microscopic study of the hypothalamicregion. Specialized contacts between astrocytes and astrocytes, and between astro-cytes and other cellular elements, are described and illustrated. The specializedinter-astrocytic junctions occur primarily in perivascular and subpial regions, butalso in areas of high synaptic density. The junctions between astrocytic processesare of hemidesmosomal type. Astrocytes are connected to oligodendroglial cells bymeans of desmosomes, and to neuronal processes by means of zonulae occludens.The functional significance of these arrangements is discussed.
60

Specialized contacts of astrocytes 61
REFERENCES
ALVAREZ-URIA, M. (1972). Ultrastructura de la degeneracion walleriana en el nervio infraorbitario.Trabajos del instituto Cajal de investigaciones biol6gicas 64, 159-222.
BRIGHTMAN, M. W. & REESE, T. S. (1967). Astrocytic and ependymal junctions in the mouse brain.Journal of Cell Biology 35, 16A-17A.
BRIGHTMAN, M. W. & REESE, T. S. (1969). Junction between intimately apposed cell membranes in thevertebrate brain. Journal of Cell Biology 40, 648-677.
DE ROBERTIS, E. (1965). Some new electron microscopical constitution in the biology of neuroglia. InBiology ofNeuroglia. Progress in Brain Research, 15, 1-11. Amsterdam, London, New York: Elsevier.
GONZALEZ AGUILAR, F. & DE ROBERTIS, E. (1963). A formalin-perfusion fixation method for histo-physiological study of the central nervous system with the electron microscope. Neurology 13, 758-777.
GRAY, E. G. (1961). Ultrastructure of synapses of cerebral cortex and certain specializations of neuro-glial membranes. In Electron Microscopy in Anatomy (ed. J. D. Boyd, F. R. Johnson & J. D. Lever),pp. 54-66. London: Edward Arnold.
GULDNER, F. H. & WOLFF, J. R. (1973). Neurono-glial synaptoid contacts in the median eminence ofthe rat: Ultrastructure, staining properties and distribution on tanycytes. Brain Research 61, 217-234.
HENRIKSON, C. K. & VAUGHN, J. E. (1974). Fine structural relationships between neurites and radialglial processes in developing mouse spinal cord. Journal of Neurocytology 3, 659-675.
HILD, W. (1957). Myelogenesis in cultures of mammalian central nervous tissue. Zeitschrift fdr Zell-forschung und mikroskopische Anatomie 46, 71-95.
KING, J. S. (1968). A light and electron microscopic study of perineuronal glial cells and processes inthe rabbit neocortex. Anatomical Record 161, 111-124.
MORALES, R. & DUNCAN, D. (1975). Specialized contacts of astrocytes with astrocytes and with othercell types in the spinal cord of the cat. Anatomical Record 182, 255-266.
PALAcIOs-PRU, E. L. & MENDOZA-BR1NZE&O, R. V. (1972). An unusual relationship between glial cellsand neuronal dendrites in olfactory bulbs of Desmodus rotundus. Brain Research 36, 404-408.
REESE, T. S. & KARNOVSKY, M. J. (1967). Fine structural localization of a blood-brain barrier toexogenous peroxidase. Journal of Cell Biology 34, 207-217.
REVEL, J. P. & KARNOVSKY, M. J. (1967). Hexagonal array of subunits in intercellular junctions of themouse heart and liver. Journal of Cell Biology 33, C7.
SIPE, J. C. & MORE, R. Y. (1976). Astrocytic gap junctions in the rat lateral hypothalamic area. AnatomicalRecord 185, 247-252.
SUAREZ NAJERA, I., FERNANDEZ Ruiz, B. & CARRATO IBAf-EZ, A. (1978). The fine structure of ependymain the hypothalamus of the hamster. Anatomischer Anzeiger 143, 466-477.
WOLFF, J. R. (1965). Elektronenmikropische Untersuchungen uber Struktur und Gestalt von Astro-zytenforsatzen. Zeitschrift far Zellforschung und mikroskopische Anatomie 66, 811-828.





























