Embed Size (px)
Citation preview

Recovery of red coral Corallium rubrum colonies
after fishing events in the Marseilles area
(France, NW Mediterranean)
Master’s thesis by Marina García Pizá
Master in Fundamental and Applied Ecology
20 September 2011
Project directors:
Dra. Cristina Linares Prats
Universitat de Barcelona
Dr. Joaquim Garrabou Vancells
Institut de Ciències del Mar,
CSIC

Presentation
This master’s thesis consists of an analysis of experimental data from a long-term
monitoring photographic study of red coral populations in the coast of Marseille. A
total of 351 red coral colonies have been monitored throughout seven years by
analyzing digital photographs in the video lab at the ICM, Institut de Ciències del
Mar, CSIC, Barcelona.
The project has been coordinated by MedRecover: Marine Biodiversity Conservation
Group, recognized as a consolidated research group by the Government of Catalonia’s
Agency for the Administration of University and Research Grants (Agència de Gestió
d’Ajuts Universitaris i de Recerca -AGAUR-). MedRecover studies the direct and
potential combined effects of global change on the conservation of marine
biodiversity in rocky coastal ecosystems, mainly in the Mediterranean Sea.
The master’s thesis contributes to the knowledge of the recuperation capacity of a
longevous species after a fishing event and the results will help in the improvement of
the management strategies valid until now. It forms part of the project: “Marine
conservation biology in coastal ecosystems (2009-2013)”, funded by AGAUR, and
coordinated by Joaquim Garrabou and Cristina Linares.
Marina García Pizá
Institut de Ciències del Mar, CSIC, 20 September 2011

Recovery of Red Coral Corallium rubrum colonies after fishing events in the Marseilles area (France, NW Mediterranean)
Summary
The Mediterranean red coral, Corallium rubrum, is a slow-growing longevous
gorgonian that produces a red calcium carbonate skeleton, which is being used by the
jewellery industry. Even though intensive fishing has caused a shift in size structure
for most of the populations in the Mediterranean, this species seems to persist. The
hypothesis that allows the persistence of C. rubrum despite the fishing pressure is
based on: 1) Small size at first reproduction and 2) Capacity of colonies to re-grow
new branches after being fished. From long-term photographic series monitoring two
populations in the Marseilles area, we studied the impact and recovery dynamics of
red coral after two fishing events. The results showed that >90% of affected colonies
have the capacity to recover by re-growing new branches in less than 7 years, but
colony size is far from the intact colony size before being fished. On the other hand,
scuba diving acts as an additional disturbance as it slows down this recovery rate.
Fishing practices and subsequent damage caused by diving, contribute to the
reduction of mean colony size with its corresponding effects such as decrease in larval
productivity and therefore an increase in risk of local extinction. Dramatic long-
lasting shifts in population demography highlight the need of improved management
regulations.
!
!

! "!
Table of contents
"#! $%&'()*+&,(% ################################################################################################################################# -!
"#"#! ./)!0('12!3,45,%6################################################################################################################ 7!
"#-#! 8,45,%6!/33/+&4!1%)!+(%4/'91&,(%!(%!'/)!+('12!:(:*21&,(%4 ############################ 7!
"#7#! ./)!+('12!:/'4,4&/%+/ ###################################################################################################### ;!
-#! <1&/',124!1%)!</&5()4 ############################################################################################################ =!
-#"#! >&*)?!4:/+,/4@!./:'()*+&,(%A!6'(B&5!1%)!C('&12,&?!'1&/4############################## =!
-#-#! >&*)?!1'/1###########################################################################################################################"D!
-#7#! E5(&(6'1:5,+!4*'9/?!(3!:/'C1%/%&!&'1%4/+&4@!./)!+('12!C(%,&(',%6######""!
-#F#! E5(&(6'1:5,+!1%12?4,4@!>*'9,912!1%)!'/G6'(B&5!4&*)?####################################"-!
-#H#! >&1&,4&,+12!1%12?4,4###########################################################################################################"I!
7#! ./4*2&4 ##########################################################################################################################################"I!
7#"#! 8,45,%6!,C:1+& ##################################################################################################################"I!
7#-#! J(%6G&/'C!C('&12,&?!'1&/4!(3!133/+&/)!1%)!*%133/+&/)!+(2(%,/4##################"K!
7#-#"#! <('&12,&?!'1&/4!(3!+(2(%,/4!133/+&/)!L?!1!3,45,%6!/9/%& ###########################"K!
7#-#-#! <('&12,&?!'1&/4!(3!+(2(%,/4!*%133/+&/)!L?!3,45,%6######################################"K!
7#-#7#! 0(C:1',4(%!L/&B//%!133/+&/)!1%)!*%133/+&/)!+(2(%,/4 ########################"K!
7#7#! 0(2(%?!'/+(9/'?!3'(C!3,45,%6!,C:1+& #####################################################################-"!
F#! M,4+*44,(% ###################################################################################################################################-F!
H#! 0(%+2*4,(%###################################################################################################################################-I!
;#! ./3/'/%+/4###################################################################################################################################-=!
!

! -!
1. Introduction
The Mediterranean red coral, Corallium rubrum (Linaeus 1758, Gorgonacea,
Octocorallia) is a colonial sessile cnidarian with an arborescent growth form. Red
coral is an important species of the rocky bottoms of the Mediterranean and is
essentially distributed in the Western basin even though it is also present in some
areas of the Eastern basin and the neighbouring African Atlantic coast (Zibrowius
1984; Garrabou & Harmelin 2002).
Red coral is a sciaphilous species, typically associated with animal-dominated
communities growing in dimly lit habitats such as rocky cliffs, overhangs and caves
from 10 to more than 600m in depth (Laborel & Vacelet 1960; Chintiroglou et al.
1989). Water flow, food supply, temperature and light regimes are the key factors that
explain its distribution (Weinberg 1979). Its main habitat is the hard substratum in the
coralligenous where it is a characteristic species of high importance (Ballesteros
2006).
C. rubrum is an asymbiotic anthozoan species and a passive benthic suspensivore that
feeds on detritic organic particles in suspension, copepods, invertebrate’s eggs,
phytoplankton, nano- and pico-plancton (Picciano et al. 2007). Like other suspension
feeders, red coral depends on water circulation and trophic content for its energy
inputs and thus, plays an important role in pelago-benthic energy transfer processes
(Arntz et al. 1999).
Long-lived gorgonians such as red coral, play an essential role in marine ecosystems
because they structure and stabilize the ecosystem (Mitchell et al. 1993) by providing
three-dimensional complexity and increasing diversity (Jones et al. 1994). For these
reasons, these kind of species are often called ecosystem engineers. Beyond its
ecological importance, red coral is an emblematic species of the Mediterranean
because it has been harvested since antiquity for the high value of its skeleton in
jewellery and ornamentation (Santangelo et al. 1993; Morel et al. 2000).

! 7!
1.1. Red Coral fishing
The Mediterranean red coral is considered the precious coral par excellence
(Figure 1). In addition to its ornamental use in the Neolithic (3000-5000 years ago),
red coral also developed a tremendous cultural importance, as it appeared in
decorative arts of the Minoan and Mycenean civilization documents in the Bronze
Age. Red coral fishing became over-efficient when the Greeks and Arabs developed a
wooden dredging device known as "Saint Andrew's Cross" (Galasso 1998, 2001)
(Figure 2). It consisted of a wooden cross with attached nets to it that was used to
entangle red coral. Most fisheries depleted their stocks rapidly and moved on to new
ones afterwards. This "boom and bust" exploitation, more similar to coal mining than
to fishery, created unstable yields, with various peaks and drops (Santangelo et al.
1993, 2001). Today, these non-selective methods such as the "Saint Andrew's Cross"
are prohibited in all the Mediterranean countries because they cause serious damage
to benthic communities. After the 1950's, scuba diving found its application in red
coral harvesting as it allowed harvesting populations dwelling in protected crevices
and cave entrances that were inaccessible by dredging. Today, scuba diving remains
as the exclusive exploitation method of C. rubrum (Figure 3).
Almost all known C. rubrum stocks are overexploited (Santangelo et al. 2001;
Tsounis et al. 2007). The overall Mediterranean yield has decreased from ~100 t. yr-1
before the 1980's to an estimated ~25-30 t. yr-1 during recent years (Tsounis et al.
2007). Today, C. rubrum fisheries (legal and illegal) continue to impede the total
recovery of exploited populations and most populations are considered overfished
(Linares et al. 2010).
1.2. Fishing effects and conservation on red coral populations
Despite the fishing pressure, red coral is still found in the NW Mediterranean
sublittoral habitats. However, intensive harvesting has resulted in significant shifts in
the size structure of current C. rubrum populations (Santangelo et al. 1993; FAO
1998), causing a decrease in biomass and mean and maximum colony size (Garrabou
& Harmelin 2002; Linares et al. in press). Fishermen continue to harvest on already
decimated populations, resulting in a further reduction of abundance and mean size.

! F!
Due to the history of intense harvesting, mankind has not had the chance to document
pristine red coral populations (Jackson et al. 2001). Private collections and museum
specimens have provided precious information regarding the maximum size this
species can potentially reach: up to 50cm and 4cm in height and diameter respectively
(Garrabou & Harmelin 2002). The oldest Mediterranean marine reserves harbouring
red coral are around 20 to 30 yr old (Francour et al. 2001) offering an excellent
opportunity to analyze the effects of protection on red coral populations. The limited
data available indicate an increase in maximum size and a shift towards more mature
populations in the oldest marine reserves (Tsounis et al. 2007; Linares et al. 2010).
Despite these positive effects, colony sizes do not reach characteristic values of
pristine populations (Garrabou & Harmelin 2002), most likely as a result of other
impacts such as poaching and diving frequentation, which do not allow their total
recovery (Linares et al. in press). Overall, despite the actual regulation efforts (see
below), protected and unprotected red coral populations are submitted to other
disturbances such as mass mortality events linked to climate change, sedimentation
increase or increased diver frequentation. This scenario could be driving C. rubrum
populations, especially in shallow water habitats, to situations of risk of local
extinction in the near future.
Therefore, management and conservation measures based on sound ecological data
are urgently needed. Until now, C. rubrum has been included in three international
conventions for the protection and conservation of species (Specifically in the 3rd
Annex of the Bern Convention in 1979, in the 5th Annex of the European "Habitats,
Fauna, Flora" in 1994, and in the 3rd Annex of the Barcelona Convention in 1995).
Moreover, during recent years, some attempts have been made to include the genus
Corallium in the Appendix II of CITES (Convention on the international trade in
endangered species of wild fauna and flora).

! H!
Figure 1: Red coral colony and small pieces of jewellery in a souvenir at L’Estartit, Costa
Brava, Spain. (Photo: Marina García Pizá).
Figure 2: Non-selective fishing of red coral under a rock using the “St Andrews cross”
(Illustration: L.F Marsigli 1725).

! ;!
Figure 3 Selective fishing by scuba diving remains the exclusive exploitation method of
C.rubrum today (Photo: J.G. Harmelin).
1.3. Red coral persistence
Red coral displays high longevity (>200 years), very low growth rates (0.20 mm.
year-1 for basal diameter) and low mortality rates (<1% in absence of fishing)
(Torrents 2007; Garrabou & Harmelin 2002; Linares et al. in press). Bearing in mind
these life history traits, it seems a paradox that this slow-growing species has not
undergone extinction yet despite being exposed to intensive harvesting since ancient
times. Therefore, the question on how do these shallow populations persist and
replenish arises. A study analyzing the size and age at first reproduction of red coral,
partially contributed to answering this question. This study showed that the size of the
smallest fertile colonies is surprisingly small, only 2.4 cm in average height, which
corresponds to an age of 7-10 years (Torrents et al. 2004). This small size would have
allowed the colonies to avoid from being harvested, while they could contribute to the
reproductive output of the population and reveal at least partially, a potential
persistence mechanism.
On the other hand, red coral as other clonal organisms, has the capacity to recover
after events of partial mortality (e.g. breakage of a branch). In fact, red coral colonies
with signs of breakage and recent re-growth of new branches have been observed
(Hereu et al. 1999; Rossi et al. 2008).

! I!
Taking into account that harvesting methods do not always harvest the whole colony,
re-growth of the parts of the colony still attached to the substrate could be a potential
source of recovery for harvested populations. However, this process has never been
studied in red coral populations and it is unclear up to what extent this mechanism
may contribute to the recovery after a fishing event. Under the framework of the study
of red coral population dynamics, two fishing events were observed, which offered
the opportunity to analyze for the first time, the recovering capacity of two
populations of the coast of Marseille.
Given this lack of knowledge over the response of this species to fishing events, the
aim of this study was to asses the capacity to recover of two C. rubrum populations of
the coast of Marseille, by analyzing long-term photographic series and quantifying
mortality and branch re-growth rates during five and seven years after each fishing
event.

! =!
2. Materials and Methods
2.1. Study species: Reproduction, growth and mortality rates
Red coral is a gonochoric species (Lacaze-Duthiers 1864) that reproduces annually
during late spring-early summer (Santangelo et al. 2003; Tsounis et al. 2007).
C. rubrum is an internal brooder and planulae have limited dispersal capabilities
(Weinberg 1979; Bramanti et al. 2005). Studies on the genetic structure of red coral
populations have uncovered genetic differentiation at small scales (~10m),
corroborating the short dispersal of this species (Ledoux et al. 2010). The dispersal of
red coral planulae is restricted to short distances because their free life is brief,
reaching 4-12 days under laboratory conditions (Weinberg 1979; Grillo et al. 1992),
but probably much less in the field (Ledoux et al. 2010). Low natatory capacity and
geonegative behaviour tend to confine larvae to the vicinity of parental colonies
within cavities (Weinberg 1979). These philopatric dispersal features are reflected in
the frequent aggregative distribution of red coral colonies (Harmelin 1985) and by the
limited gene flow between populations (Ledoux et al. 2010).
Size at first reproduction in C. rubrum is much smaller compared to other gorgonian
species. As commented above, the colony height is 2-3 cm at first reproduction
corresponding to an age of 7-10 years (Torrents et al. 2004). Despite the small size at
first reproduction, it would be large enough to reduce most mortality risks by
overgrowth (specially by sponges) regardless the age at first reproduction (Torrents et
al. 2007).
Recruitment rates of red coral vary form one site to another (different colony
abundance, density and size, crevice size and shape and exposure to currents)
(Bramanti et al. 2007; Linares et al. in press) and high recruitment pulses seem to
combine with many years of near zero recruits (Garrabou & Harmelin 2002; Linares
et al, in press) or be very regular from 5-10 colonies. year-1. dm-2 (Bramanti et al.
2005). Nevertheless, the unique two long-term studies on this species: Garrabou &
Harmelin 2002 (22-year study) and Linares et al. in press (13-year study), suggest that
in most cases, recruitment of this species is very low (<1 recruit/300 cm2), especially

! K!
over periods longer than 5 years. This is consistent with the life-history traits, which
state that adult survival in long-lived species such as red coral, is often associated
with infrequent and low recruitment (Stearns 1992). Annual mortality rates for these
recruits are high (4% in colonies less than 5 years old) resulting form the competition
for space with other sessile organisms. Mortality rates then decrease progressively up
to a value of about 1% for colonies over 15 years old (Garrabou & Harmelin 2002).
Growth ring analysis and experimental studies have determined very slow growth
rates of this long-lived organism that reaches 50 cm of height and reaches more than
100 years of age (Garrabou & Harmelin 2002; Marschal et al. 2004). This species has
a growth rate of about 0.20 mm.year-1 in diameter and 1.78 ± 0.7 mm.year-1 in height
(Garrabou & Harmelin 2002; Marschal et al. 2004; Torrents 2007; Linares et al.
2010).
Red coral annual mortality rate is usually very low (less than 1%) when populations
are not subjected to human-induced disturbances. The principal sources of natural
mortality include competition for space with sponges and other sessile biota, removal
from the substrate due the action of boring species (such as the clionaidae family,
which perforate corals by secreting acid over the calcified skeletons), storms,
increased sedimentation of fine particles (Harmelin 1985) and predation of the
coenchyme by the small gastropod Pseudosmnia carnea and the crustacean Balssia
gasti (Abbiatti et al. 1992). However, predators of red coral are scarce (Santangelo et
al. 1993) and as in other sessile invertebrate dominated communities, predation
appears to have a minor impact on the red coral community (Garrabou & Harmelin
2002). The principal sources of anthropogenic mortality are due to professional
fishing or poaching (Hereu et al. 1999; Tsounis et al. 2007) and massive mortality
events linked to positive temperature anomalies (Garrabou et al. 2009). Another
source of anthropogenic mortality is the increase of pollutants and water from
desalination plants (Harmelin 1985; Garrabou et al. 2001).
Besides overfishing, during the last decade, climate-induced mortalities have caused
the most important impacts on red coral populations. In the NW Mediterranean area,
at the end of the summers of 1999 and 2003, two mass mortality events affected
marine rocky benthic communities on large spatial scales (more than 500 km)
(Garrabou et al. 2001). These two large-scale mortality outbreaks have been related to

! "D!
positive temperature anomalies (Garrabou et al. 2001), severely affecting red coral
populations. In the Marseilles region during the summer of 2003, 40% of the
populations of red coral were affected with a medium degree of impact (Garrabou et
al. 2009). As over-fishing could now be coupled with mass mortality events (i.e. due
to thermal anomalies), then the co-occurrence of these two mortality sources could
dramatically depress the recovery of red coral populations (Santangelo 2006).
2.2. Study area
The study area is the limestone rocky coast of Massif des Calanques in the SE of
Marseilles, France (Figure 4). Massif des Calanques is located in a eutrophic area near
a urbanized coast, influenced by wind-induced local upwelling (Millot et al. 1980)
and is affected by the Ligurian current. Due to the specific habitat characteristics of
the area: like submerged cavities and overhangs (Laborel & Vacelet 1960), along with
instability of the water column during summer, the development of red coral at
shallow depths (20-22m) is favoured.
The two shallow red coral populations studied are located in two different islands;
Riou Sud located in Riou Island is characterized by a vertical wall and Grotte Corail,
located in Maire Island, which consists of a cave-like tunnel. At these depths, both
sites are mainly characterized by the presence of perennial animals associated to
coralligenous assemblages (Coma et al. 1998; Garrabou & Harmelin 2002). In the
study area, red coral populations have been exploited since the medieval ages using
different kinds of techniques (Morel et al. 2000). At present, harvesting is permitted
to 10 authorised red coral fishermen and 2 of them are present in the Calanques area,
even though poaching is likely to be common. Harvesting by divers threatens mainly
the largest and most attractive colonies.
Lastly, scuba divers frequentation is almost null in Riou Sud, while Grotte Corail is
one of the most commonly frequented diving sites in the Marseille’s area, with more
than 5000 dives per year. At this site, the near presence of detached branch fragments
on the sea bottom seems to be the evidence of high diving pressure.

! ""!
Figure 4: Map of the Calanques region in the coast of Marseille. The two dots in red
represent the study sites: Riou Sud in Riou Island and Grotte Corail in Maire Island.
2.3. Photographic survey of permanent transects: Red coral
monitoring
When studying a slow-growing species, long-term photographic series are useful to
detect and quantify changes over spatial and temporal scales. During this long-term
study on the population dynamics of red coral in the coast of Marseille, two fishing
events took place, affecting the populations of Riou Sud and Grotte Corail. We
analyzed the photographic series obtained yearly during seven years.

! "-!
A permanent plot was set up in each population to monitor the colonies
photographically over time. Each plot was 2-4m long and PVC screws were attached
to the wall every 40-50cm. An elastic band was placed along each transect and around
each screw in order to guide the photographer through the transect. The pictures were
taken above and below the band following the transect. A fixed frame was used to
ensure a same study area in each photograph of 20x20cm. This method allowed a
monitoring of each colony individually throughout the years.
In Riou Sud, two permanent plots were established at 20 and 22m in 2005.
Photographs were analyzed from 2005 to 2011. Throughout the monitoring of the
photographs, a fishing event was detected between April and June 2006, therefore a
comparison could be done before and after the fishing event. Moreover, the fished
colonies could be identified.
In the Grotte Corail population, two permanent transects were established at 15 m just
after a fishing event was observed during June 2002. Photographs were analyzed from
2002 to 2009, but due to logistic constraints, pictures were not taken during 2007 and
2008. Among the colonies observed in 2002, some were half broken with hanging
broken branches, as well as recent wounds with tissue coming out. This gives us the
evidence that the colonies had just been fished. However, we have no before-after
information on this site because the plots were set up just after the fishing event.
2.4. Photographic analysis: Survival and re-growth study
All the analyzed colonies were individually identified and followed annually
throughout the study period. Mortality and branch re-growth was surveyed on
photographs corresponding to 1-year intervals whenever it was possible. A total of
351 colonies were monitored during the 7-year study: 211 colonies in Riou Sud and
140 in Grotte Corail. After the fishing event in each site, the studied colonies were
divided into two groups; affected and unaffected by fishing. This allowed us to
explore potential differential mortality traits among groups.! For each colony, we
determined the type of mortality suffered or not and the presence and number of new
branches. After the fishing event, the affected colonies were monitored not just to

! "7!
study new branch re-growth but also to see if being affected by fishing in previous
years, could influence into a future mortality.
The following parameters were defined from the photographs:
Re-growth of a branch was defined as the first bump observed on the base of the
affected colony. Figure 5 shows the evolution on new branches.
Partial mortality was defined when a part of the colony was lost between the time
intervals considered. After the fishing event, the colony base could either have a
smooth surface if no branches were present (e.g. after the fishing event) or a rough
surface when a branch was beginning to grow or some branches had been left (e.g.
due to a more conscious fishing extraction). Figure 6 shows an example of partial
mortality.
Total mortality was defined when the colony disappeared and was not seen again in
the following years. Total mortality could either be due to type of extraction when the
whole colony was removed with the substratum still attached to the base, or to the
death of the colony some time after the extraction (e.g. the colony base is epiphyted
by sponges and eventually disappears). Figure 7 shows an example of total mortality.
Figure 5: Branch re-growth. The colony on the left shows two small bumps and a clear
branch, while the colony on the right shows a group of higher developed branches.

! "F!
Figure 6: Partial mortality: The circles indicate a smooth base for each harvested colony.

! "H!
Figure 7: Total mortality: The circles show the three missing colonies on the left, which
disappear after the fishing event in 2006.

! ";!
Figure 8: Evolution of fished colonies in Riou Sud from 2005 to 2007 (top to bottom). The
fishing event is clearly observed in 2006, where partial mortality is shown for all the colonies.

! "I!
2.5. Statistical analysis
The differences between affected and unaffected colonies and the differences between
sites were analyzed using non-parametric analysis, since our data violated normality
and homocedasticity.
A Kolmogorov-Smirnov two-sample test was used to compare affected and
unaffected colonies between the two sites, and to compare the average number of
branches in each site. The data representing the number of branches in affected
colonies contained a large number of zero values, especially during the first two years
of monitoring, when many colonies still had no branches. Even when the data were
log transformed, the data did not follow a normal distribution. Thus, an ANOVA
repeated measures test could not be applied in order to compare differences between
sites and years. Finally, a non-parametric Kolmogorov-Smirnov two-sample test was
applied in order to compare the number of branches between sites during the last year
of monitoring (five and seven years after the fishing event). Meanwhile, to compare
affected and unaffected colonies as well as the type of mortality suffered by the
colonies within the same site, a Mann-Whitney U test was applied.
3. Results
3.1. Fishing impact
Figure 9 shows the percentage of affected and unaffected colonies by a fishing event
in each site. The fishing event that took place in Riou Sud during 2006 affected 38%
of the colonies within the transect. Meanwhile the fishing event in Grotte Corail
during 2002, affected 57% of the colonies. The differences among affected and
unaffected colonies between sites were not significant (Kolmogorov-Smirnov test,
p=0.10). The type of mortality caused by fishing was mainly partial mortality.
Between 90-100% of the colonies were affected by this type of mortality in both sites
(Figure 10). Total mortality represented only 7% of the affected colonies in Riou Sud,
meanwhile, it was not present in Grotte Corail.

! "=!
Figure 9: Percentage of red coral colonies affected and unaffected by fishing in Riou Sud
during 2006 and in Grotte Corail during 2002. N is the number of colonies studied in each
site.
Figure 10: Percentage of affected colonies by fishing that suffer partial mortality and total
mortality. N is the number of affected colonies in each site.

! "K!
3.2. Long-term mortality rates of affected and unaffected colonies
3.2.1. Mortality rates of colonies affected by a fishing event
Colonies affected by fishing in Riou Sud showed very low values of new mortality
“post-affection” from 2007-2011: only 1.2% of the affected colonies suffered total
mortality (Figure 11 a). In contrast, the monitoring from 2003-2009 in Grotte Corail
showed a higher percentage of affected colonies suffering from mortality “post-
affection”: approximately 15% of the affected colonies showed partial mortality and
1.2% total mortality (Figure 11b).
3.2.2. Mortality rates of colonies unaffected by fishing
In Riou Sud, a small percentage (5%) of unaffected colonies showed partial mortality
and no total mortality was observed during 2007-2011 (Fig 11a). However, in Grotte
Corail, up to 60% of the unaffected colonies suffered partial mortality, while total
mortality was recorded on 4% in these colonies (Figure 11b).
3.2.3. Comparison between affected and unaffected colonies
In Riou Sud, partial mortality was significantly higher in unaffected colonies (Mann-
Whitney U test, p<0.05). Meanwhile, differences in total mortality between affected
and unaffected colonies were not significantly different (Mann-Whitney U test, p
0.86). However, in Grotte Corail, both differences in partial and total mortality
between affected and unaffected colonies showed significant differences (Mann-
Whitney U test, p<0.05).

! -D!
Figure 11 a) New mortality of affected and unaffected colonies by fishing in Riou Sud after
the fishing event from 2007-2011. b) New mortality of the affected and unaffected colonies
by fishing in Grotte Corail after the fishing event from 2003-2009.

! -"!
3.3. Colony recovery from fishing impact !
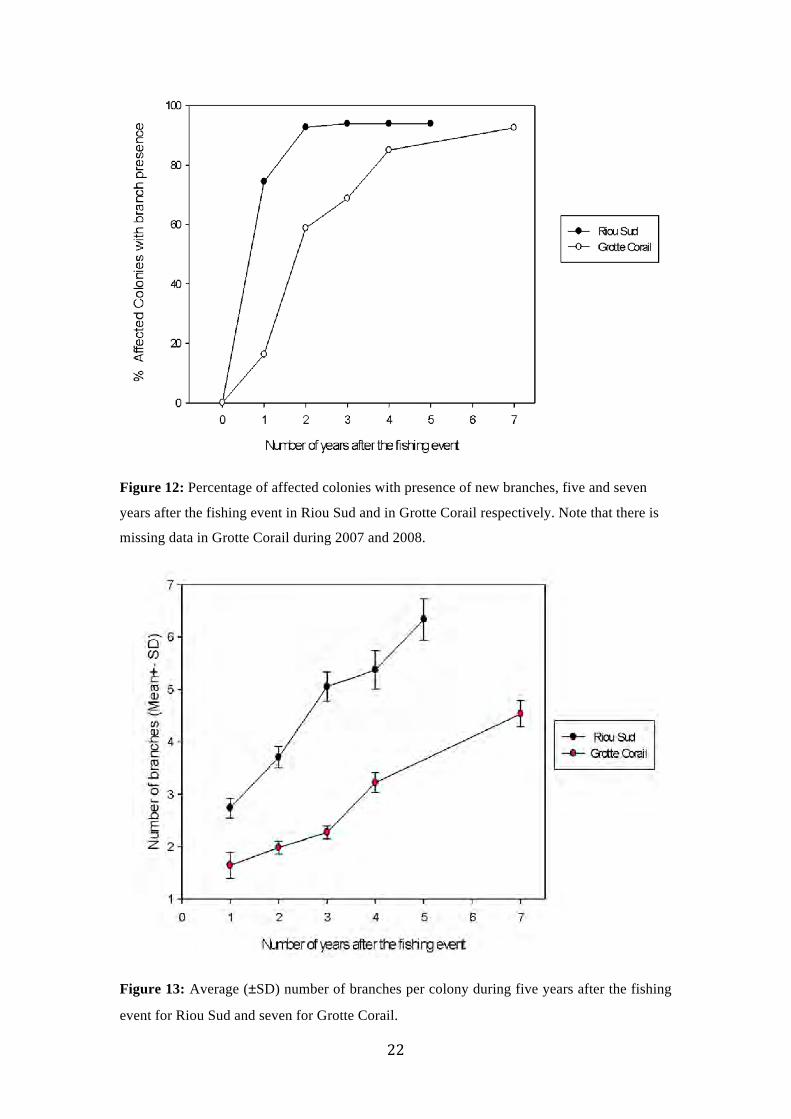
All the colonies that survived the fishing event show branch re-growth during the
seven-year monitoring. Figure 12 shows the percentage of affected colonies with
presence of new branches over the study time. A small percentage of affected
colonies died in the following years after the fishing event and therefore, branch
presence was not reached in 100% of the affected colonies.
During the first year after the fishing event, colonies in both sites started growing new
branches. The greatest difference in branch presence was during the first year after the
fishing event, in which Riou Sud had 75% of its colonies with presence of new
branches, while Grotte Corail showed new branch presence in 15% of its colonies
(Figure 12). During this first year, colonies in Riou Sud had from one to four branches
as well as a small percentage (15%) of colonies with more than four branches (Figure
14 a). Meanwhile, Grotte Corail had colonies ranging form one to three branches
(Figure 14 b).
The overall number of branches marked another difference between sites. In Grotte
Corail, colonies with 4 branches were less abundant and were not observed until 4
years after the fishing event, whereas in Riou Sud, these colonies were already present
since the first year after the event. Five years after the event in Riou Sud, 55% of its
colonies grew more than four branches compared to seven years after the event in
Grotte Corail, where only 35% of its colonies grew more than four branches. Both
sites reached a similar degree of colonies with branch presence, even though there
were differences between the number of branches per colony.
Figure 13 shows the evolution of the average number of branches per colony during
the consecutive years after the fishing event for each site. Riou Sud displayed a higher
mean number of branches throughout the study compared to Grotte Corail, reaching
an average of six branches per colony, while Grotte Corail reached an average of four
branches per colony. The average number of branches between sites; five years after
the event for Riou Sud and seven for Grotte Corail, was significantly different
(Kolmogorov-Smirnov test, p<0.05).
! --!
Figure 12: Percentage of affected colonies with presence of new branches, five and seven
years after the fishing event in Riou Sud and in Grotte Corail respectively. Note that there is
missing data in Grotte Corail during 2007 and 2008.
Figure 13: Average (±SD) number of branches per colony during five years after the fishing
event for Riou Sud and seven for Grotte Corail.

! -7!
Figure 14: Number of branches in affected colonies in Riou Sud (a) and Grotte Corail (b)
during the consecutive years after each fishing event.

! -F!
4. Discussion
This study showed that most colonies (>90%) could recover from a fishing event
when fishermen did not harvest the whole colony. Between five and seven years were
necessary to observe at least one new branch in colonies affected by the fishing
events. However, recuperation of the affected colonies at the end of the study was not
complete. This could be observed in Riou Sud where colony size after five years was
far away from the size of intact colonies before the event. Nevertheless, quantitative
measurements would be necessary to establish a more precise recuperation status.
Previous studies have shown the slow growth rates of red coral colonies being around
0.20 mm.year-1 for basal diameter and 1.78 mm.year-1 for colony height (Garrabou &
Harmelin 2002, Torrents 2007, Linares et al. 2010). Bearing in mind these growth
rates and the size of the larger colonies found in studied populations (97.2 mm in
height, Garrabou & Harmelin 2002), it is clear that a full recovery will require at least
60 years of effective absence of harvesting.
Going back to the paradox commented in the introduction, concerning the persistence
of a slow dynamic species such as red coral despite being exposed to intensive
harvesting since ancient times, we could consider the re-growth of new branches as a
meaningful potential mechanism to explain today’s abundance of populations. This is
clearly illustrated by comparing the growth dynamics of colonies either through re-
growth of new branches in adults or by growth patterns observed in sexual recruits,
which is the alternative mechanism available for the recovery of populations. With
reference to the mean number of branches, our results showed that five and seven
years after fishing, colonies had a mean number of 5 branches. Meanwhile, the
monitoring of colonies recruited on experimental panels showed about 3 branches
after 22 years of growth (Garrabou & Harmelin 2002). Branch growth rates are 8
times higher in affected adult colonies than in recruits. Moreover, mortality rates in
recruits are remarkably high (4% in colonies less than 5 years old, Garrabou &
Harmelin 2002) compared to mortality rates observed in affected adult colonies
quantified in this study (0.30%). This contrast between branch growth and mortality
in recruits and adult colonies should be taken into account when studying
recuperation capacity.

! -H!
Differences in partial mortality rates found in this study between Riou Sud and
Grotte Corail could be attributed to the difference in diver frequentation in both sites
(see Material and methods). A recent study based on 13 years of red coral monitoring,
showed a significant negative correlation between the annual number of recreational
dives and the basal diameter of red coral colonies, suggesting that divers may increase
the mortality rates of the largest red coral colonies (Linares et al. in press) and
therefore impede the total recovery of the affected colonies. In our study, despite the
high scuba diving pressure over Grotte Corail, the affected colonies seem to be
“protected” due to their small size and are thus, less susceptible to breakage by divers
in contrast with the unaffected colonies which are larger in size. Overall, we
demonstrated differences between sites by quantifying partial mortality rates due to
mechanical breakage of branches, which occurred at higher rates in frequented diving
sites and slowed down recovery rates. In fact, red coral colonies in Riou Sud have a
higher mean height (37.8mm) than the colonies in Grotte Corail (26.5mm) (Linares et
al. 2010).
The conservation of Mediterranean red coral is a worldwide concern as it has been
proved by the recent struggle to include the Genus Corallium in the Appendix II of
CITES (Bruckner 2009, Tsounis et al. 2009). This attempt failed because
Mediterranean red coral cannot be considered an endangered species due to the
extensive distribution of dense populations in the Mediterranean Basin. It should be
taken into account that using density and abundance seems an incorrect assessment of
the state of this species, because changes in these parameters are inadequate measures
of the decline for colonial organisms such as C. rubrum, as they exhibit a size-based
exponential increase in reproductive output (Bruckner 2009).
New management and conservation plans should focus on enhancing the persistence
of populations. In the case of red coral, it seems urgent to promote the increase of the
abundance of large colonies, as these colonies have enhanced survival likelihood and
thus increased future reproductive success (Szmant-Froelich 1985). For example, in a
C. rubrum population, a young colony produces only tens of planulae, while an old
and large colony, produces more than 2000 planulae (Santangelo et al. 2003).

! -;!
Red coral populations have been historically submitted to harvesting and have coped
with the effects of this strong disturbance. However, disturbances such as the increase
in diver frequentation, mass mortality events linked to climate change and
sedimentation increase (Ballesteros 2006; Garrabou et al. 2009; Linares et al. in
press), may act synergically jeopardizing the persistence of red coral populations.
Overall, this new disturbance regime could reduce the capacity of colonies to recover.
From this study, it is clear that new management regulations which pursue a
sustainable fishery should contemplate measures such as; at least, leaving the colony
base intact and if possible, leaving some branches attached to the base, in order to
help the recovery of the affected colonies. Another point to consider is to limit the
access of scuba divers to sites were red coral dwells, such as submarine caves, which
represent the most vulnerable habitat to scuba diving activity. Combining both
recommendations and applying a reasonable rotational harvest (over periods of
several decades) to allow the stocks to recover before re-harvesting, would be a
precautionary approach for the protection of red coral and would facilitate the
recovery of the populations.

! -I!
5. Conclusion
Fishing practices and subsequent damage caused by diving, contribute to the
reduction of mean colony size with its corresponding effects such as decrease in larval
productivity and therefore an increase in risk of local extinction.
Re-growth of new branches acts as an important mechanism in the recovery of
affected colonies, allowing the persistence of red coral despite the harvesting
pressure. Scuba diving activities can affect branch re-growth rate, and thus slow down
the recovering capacity of the colonies. Future management plans should recommend
fishermen to keep the colony base intact and if possible leave at least half of the
branches, which will ensure a faster and more robust recovery of the colonies. On the
other hand, imposing a strict regulation plan over diving activities, especially in
vulnerable sites, will ensure more efficient protection plans.
Acknowledgements
I would specially like to thank Cristina Linares and Joaquim Garrabou for giving me
the opportunity to be part of their research group and for their support during these
months. I would also like to thank my video lab mates for sharing such a pleasant
atmosphere of work and for their advice while writing this project. And finally, thank
Luis for his help with my difficulties with the computer and for his infinite patience.

! -=!
6. References
Abbiati, M., G.Buffoni, G.Caforio, G.Dicola, G.Santangelo. 1992. Harvesting, predation and
competition effects on a red coral population. Netherlands Journal of Sea Research 30: 219–
228.
Arntz W.E., J.M.Gili, K.Reise. 1999. Unjustifiably ignored: reflections on the role of benthos
in marine ecosystems. Biogeochemical cycling and sediment ecology. Kluwer Academic
Publishers, Netherlands pp 105–124.
Ballesteros, E. 2006. Mediterranean Coralligenous Assemblages: A synthesis of Present
Knowledge. Oceanography and Marine Biology 44: 123-195.
Bone, E.K., M.J.Keough. 2005. Responses to damage in an arborescent bryozoan: Effects of
injury location. Journal of Experimental Marine Biology and Ecology 324: 127-140.
Bramanti, L., G.Magagnini, L.Maio, G.Santangelo. 2005. Recruitment, early survival and
growth of the Mediterranean red coral Corallium rubrum, a 4-year study. Journal of
Experimental Marine Biology and Ecology 314: 69-78.
Bramanti, L., S.Rossi, G.Tsounis, J.M.Gili and G.Santangelo. 2007. Settlement and early
survival of red coral artificial substrates in different geographic areas: some clues for
demography and restoration. Hydrobiología 580:219-224.
Bruckner, A.W. 2009. Rate and extent of decline in Corallium (pink and red coral)
populations: existing data meet the requirements for a CITES Appendix II listing. Marine
Ecology Progress Series 397: 319-332.
Bruckner, A.W. 2010. Quantifying the decline in Corallium rubrum populations: Reply to
Santangelo & Bramanti (2010). Marine Ecology Progress Series 418: 299-303.
Coma, R., M.Ribes, J.M.Gili, M.Zabala. 1998. Growth in a modular colonial marine
invertebrate. Estuarine and Coastal Shelf Science 47:459-470.
Chintiroglou, H., C.Dounas, A.Koukouras. 1989. The presence of Corallium rubrum in the
Eastern Mediterranean. Mitteilungen aus dem Zoologischen Museum in Berlin 65:145–149.
FAO yearbook .1998. Fishery Statistics: Capture-Production 1996. FAO Fisheries no. 50.

! -K!
Francour, P., J.G.Harmelin, D.Pollard & S.Sartoretto. 2001. A review of marine protected
areas in the northwestern Mediterranean region: siting, usage, zonation and management.
Aquatic Conservation: Marine and Freshwater Ecosystems 11: 155–188.
Galasso, M. 2001. La pesca del corallo in Sardegna: evoluzione, persistenze e innovazioni
tecniche. Centro Studi sul Corallo (Coralium rubrum L.) Convegno di Studi Sassari 7-10.
Gallmetzer, I., A.Haselmair, B.Velimirov. 2010. Slow growth and early sexual maturity:
Bane and boon for the red coral Corallium rubrum. Estuarine, Coastal and Shelf Science 90:
1-10.
Garrabou J., T.Pérez, S. Saroretto, J.D.Harmelin. 2001. Mass mortality event in red coral
Corallium rubrum populations in Provence region, France, NW Mediterranean. Marine
Ecology Progress Series 217: 263- 272.
Garrabou, J., J.G.Harmelin. 2002. A 20-year study on life-history traits of a harvested long-
lived temperate coral in the NW Mediterranean: insights into conservation and management
needs. Journal of Animal Ecology 71:966-978.
Garrabou, J. Informe en relación el comís de corall vermell per part del cos d’agents rurals.
Setembre 2009. Expedient Número 01026-2009.
Garrabou, J., R.Coma, N.Bensoussan, M.Bally, P.Chevaldonné, M.Cigliano, D.Diaz,
G.Harmelin, M.Gambis, D.Kersting, J.B.Ledoux, C.Lejeusne, C.Linares, C.Marshal, T.Pérez,
M.Ribes, J.Romano, E.Serrano, N.Teixidó, O.Torrents, M.Zabala, F.Zuberer and C.Cerrano.
2009. Mass mortality events in Northwestern Mediterranean rocky benthic communities:
effects of the 2003 heat wave. Global Change Biology doi:10.1111/j.1365-2486.2008.01823.x
Grillo, M.C., D.Allemand. 1992. Ca uptake and deposition by the Mediterranean red coral.
Journal of Experimental Zoology 262: 237-246.
Harmelin, J.G. 1985. Organisation spatiale des communautés sessiles des grottes sous-
marines de Méditerranée. Rapports Commission Internationale pour l’Exploration
Scientifique de la Mer Méditerranée 29: 149–153.

! 7D!
Hereu. B., C. Linares, M.Zabala .1999. Avaluació de l’impacte de l’episodi de corall vermell
(Corallium rubrum) de la zona protegida de les Illes Medes detectat durant l’hivern del 2000.
In: Seguiment temporal de les Illes Medes (1999) Informe técnic per al Departament de Medi
Ambient, Generalitat de Catalunya.
Jackson, J.B.C. 2001. What was natural in the coastal oceans? Proceedings of the National
Academy of Sciences USA, 98, 5411– 5418.
Jones, C.G., J.H.Lawton, M.Shachak. 1994. Organisms as ecosystem engineers. Oikos
69:373–386.
Laborel, J., J.Vacelet. 1960. Répartition bionomique du Corallium rubrum dans les grottes et
falaises sous-marines. Rapports et Procès-Verbaux des Réunions de la Commission
Internationale pour l’Exploration Scientifique de la Mer Méditerranée 16:465–469.
Lacaze-Duthiers, H. 1864. Histoire Naturelle du Corail. J. B. Bailière et fils, Paris.
Ledoux, J.B. 2010. Biologie de la conservation du corail rouge, Corallium rubrum (Linnaeus,
1758): Impact du changement global sur l’évolution des populations infralittorales en
Méditerranée Nord-Occidentale. Thèse de Doctorat de L’Université de la Méditerranée.
Centre d’Océanologie de Marseille.
Linares, C., O.Bianchimani, O.Torrents, C.Marshal, P.Drap, J.Garrabou. 2010. Marine
Protected Areas and the conservation of long-lived invertebrates: The Mediterranean red
coral. Marine Ecology Progress Series 402: 69-79.
Linares, C., J.Garrabou, B.Hereu, D.Díaz, C. Marschal, E.Sala, M.Zabala. In press.
Conservation Biology.
Marshal, C., J.Garrabou, J.G.Harmelin, M.Pichon. 2004. A new method for measuring growth
and age in the precious coral Corallium rubrum. Coral Reefs 23: 423-432.
Millot, C., L.Wald. 1980. The effect of the Mistral wind on the Ligurian current near
Provence. Oceanol Acta 3:399–402
Mitchell, N., M.R.Dardeu, W.Schroeder. 1993. Colony morphology, age structure and
relative growth of two gorgonian corals, Leptogorgia bebes (Verrill) and Leptogorgia
virgulata (Lamarck) from the northern Gulf of Mexico. Coral Reefs 12:6-70.
Morel, J.P., C.Rondi-Costanzo, D.Ugolini, 2000. Corallo di Ieri & Corallo di Oggi. Atti Del
Convegno Ravello, Villa Rufolo, 13–15 Dicembre 1996. Centro Universitario Europeo Per I
Beni Culturali, Ravello. Edipuglia, Bari.

! 7"!
Picciano, M., C.Ferrier-Pagès. 2007. Ingestion of pico- and nano-plankton in the
Mediterranean red coral Corallium rubrum. Marine Biology 150:773–782.
Proceedings of the First International Workshop on Corallium Science, Management and
Trade. March 16-20, 2009. Hong Kong, China. A. Bruckner and G. Roberts.
Proceedings of the International Red Coral Science, Management and Trade: Lessons from
the Mediterranean. September 23-26, 2009. Naples, Italy. E.Bussoletti, A. Bruckner, D.
Cottingham, G. Roberts, R.Sandulli.
Rossi, S., G.Tsounis, C.Orejas, T.Padrón, J.M. Gili, L.Bramanti, N.Teixidó, J.Gutt. 2008.
Survey of deep-dwelling red coral (Corallium rubrum) populations at Cap de Creus (NW
Mediterranean). Marine Biology 154: 533-545.
Sala, E., J.Garrabou, M.Zabala. 1996. Effects of diver frequentation on Mediterranean
sublittoral populations of the bryozoan Pentapora fascialis. Marine Biology 126:451-459.
Santangelo, G., M.Abbiati, F.Giannini, F.Cicogna, F. (1993a) Red coral fishing trends in the
western Mediterranean Sea. Scientia Marina 57: 139–143.
Santangelo G., M.Abbiati. 2001. Red coral: conservation and management of an over-
exploited Mediterranean species. Aquatic Conservation: Marine and Freshwater Ecosystems
11:253-259.
Santangelo, G., E.Carletti, E.Maggi, L.Bramanti. 2003. Reproduction and population sexual
structure of the overexploited Mediterranean red coral Corallium rubrum. Marine Ecology
Progress Series 248: 99–108.
Santangelo, G., L.Bramanti, M.Iannelli. 2006. Population dynamics and conservation biology
of over-exploited Mediterranean red coral. Journal of Theoretical Biology 244:416-423.
Santangelo, G., L.Bramanti. 2010. Quantifying the decline in Corallium rubrum populations.
Marine Ecology Progress Series 418: 295-297.
Szmant-Froelich, A.M. 1985. The effect of colony size on the reproductive ability of the
Caribbean coral Montastrea annularis (Ellis and Solander). Proceedings of the 5th
International Coral Reef Congress, Tahiti. Pages: 295–300.
Stearns, S.C. 1992. The evolution of life histories. Oxford University Press.

! 7-!
Torrents, O., J.Garrabou, C.Marschal, J.G. Harmelin. 2004. Age and size at first reproduction
in the commercially exploited red coral Corallium rubrum (L.) in the Marseilles area (France,
NW Mediterranean). Biological Conservation 121: 391-397.
Torrents, O., E.Tambutté, N.Caminiti, J. Garrabou. 2006. Upper thermal thresholds of
shallow vs. deep populations of the precious coral Corallium rubrum (L.): Assessing the
potential effects of warming in the NW Mediterranean. Journal of Experimental Marine
Biology and Ecology 357: 7-19.
Torrents, O., 2007. Biologie des populations du corail rouge Corallium rubrum (L. 1758) de
Méditerranée nord–occidentale. Thèse de Doctorat de L’Université de la Méditerranée.
Centre d’Océanologie de Marseille.
Torrents, O., J.Garrabou. 2011. Fecundity of red coral Corallium rubrum (L.) populations in
contrasting environmental conditions in the NW Mediterranean. Marine Biology 158: 1019-
1028.
Tsounis, G., S.Rossi, J.Laudien, L.Bramanti, N.Fernández, J.M. Gili, W.Arntz. 2005. Diet
and seasonal prey capture in the Mediterranean red coral (Corallium rubrum L.). Marine
Biology 149: 313-325.
Tsounis, G., S.Rossi, J.M. Gili, W.Arntz. 2007. Red Coral Fishery at the Costa Brava (NW
Mediterranean): Case Study of an Overharvested Precious Coral. Ecosystems 10: 975-986.
Tsounis, G., S.Rossi, R.Grigg, G.Santangelo, L.Bramanti, J.M. Gili. 2009. The Exploitation
and Conservation of Precious Corals. 2009. Oceanography and Marine Biology 48: 161-212.
Weinberg, S. 1979. The light-dependent behaviour of planulae larvae of Eunicella singularis
and Corallium rubrum and its implication for octocorallian ecology. Bijdragen tot de
Dierkunde 49: 16–30.
Zabala, M., E.Ballesteros. 1989. Surface-dependent strategies and energy flux in benthic
marine communities or why corals don't exist in the Mediterranean. Scientia Marina 53:3–17.
Zakai, D., N.E.Chadwick-Furman. 2001. Impacts of intensive recreational diving on reef
corals at Eliat, northen Red Sea. Biological Conservation 105: 179-187.
Zibrowius, H., V.Monteiro-Marques, M.Grasshoff. 1984. La répartition du Corallium
rubrum dans l’Atlantique (Cnidaria: Anthozoa: Gorgonaria). Téthys 11:163–170.





























