Embed Size (px)
Citation preview

Copyright 2001 by the Genetics Society of America
Recombination Between Paralogues at the rp1 Rust Resistance Locus in Maize
Qing Sun,* Nicholas C. Collins,† Michael Ayliffe,‡ Shavannor M. Smith,*Jeff Drake,* Tony Pryor‡ and Scot H. Hulbert*
*Department of Plant Pathology, Kansas State University, Manhattan, Kansas 66506, †Sainsbury Laboratory, John Innes Centre,Norwich, Norfolk NR4 7UH, United Kingdom and ‡Division of Plant Industry, Commonwealth
Scientific and Industrial Research Organisation, Canberra, ACT 2601, Australia
Manuscript received November 16, 2000Accepted for publication February 13, 2001
ABSTRACTRp1 is a complex rust resistance locus of maize. The HRp1-D haplotype is composed of Rp1-D and eight
paralogues, seven of which also code for predicted nucleotide binding site-leucine rich repeat (NBS-LRR)proteins similar to the Rp1-D gene. The paralogues are polymorphic (DNA identities 91–97%), especiallyin the C-terminal LRR domain. The remaining family member encodes a truncated protein that has noLRR domain. Seven of the nine family members, including the truncated gene, are transcribed. Sequencecomparisons between paralogues provide evidence for past recombination events between paralogues anddiversifying selection, particularly in the C-terminal half of the LRR domain. Variants selected for completeor partial loss of Rp1-D resistance can be explained by unequal crossing over that occurred mostly withincoding regions. The Rp1-D gene is altered or lost in all variants, the recombination breakpoints occurthroughout the genes, and most recombinant events (9/14 examined) involved the same untranscribedparalogue with the Rp1-D gene. One recombinant with a complete LRR from Rp1-D, but the amino-terminal portion from another homologue, conferred the Rp1-D specificity but with a reduced level ofresistance.
DISEASE resistance (R) genes utilized by plant locus, flax M locus, the tomato Pto, I2, Cf2/5, and Cf4/9breeders arose by coevolution of pathogens with loci, the rice Xa21 locus (Martin et al. 1993; Whitham
ancestors of current crop species (Crute and Pink et al. 1994; Anderson et al. 1997; Song et al. 1997;1996). Disease resistance conferred by R genes is usually Thomas et al. 1997; Dixon et al. 1998; Simons et al.race specific, conditioning resistance reactions only to 1998), and the rp1 locus of maize. Many of the tightlypathogen biotypes that carry the corresponding aviru- linked gene families that have been characterized havelence gene. The effectiveness of a resistance gene in relatively simple structures, with small intergenic re-controlling disease depends on the frequency of the gions, which do not code for unrelated genes. The genecorresponding avirulence gene in the pathogen popula- cluster carrying the Dm3 gene of lettuce is notable intion. Plant pathogens, especially cereal rusts, are capa- that the distance between the homologous genes is esti-ble of altering their specific virulence through mutation mated at z100 kb yet no genes have been identified inor loss of Avr genes (Burdon and Silk 1997). Resistance the intergenic regions (Meyers et al. 1998a). A contrastloci that can match the genetic plasticity of pathogen is the Pto locus, which carries five homologous genespopulations would be an asset to the survival of plant including Pto that encode protein kinases and, imbed-species. The first indications that plants may carry such ded within this cluster, an unrelated gene Prf (Sal-loci were the multiple resistance specificities that have meron et al. 1996) that is also required for Pto-mediatedbeen mapped to loci like the L and M loci of flax (Shep- resistance. The barley Mla region carries three distinctherd and Mayo 1972), the Mla locus of barley (Jahoor nucleotide binding site-leucine rich repeat (NBS-LRR)et al. 1993), the Dm1 locus of lettuce (Hulbert and gene families, which are intermingled within a 240-kbMichelmore 1985), and the rp1 locus of maize (Saxena interval (Wei et al. 1999).and Hooker 1968). Molecular genetic studies of R gene loci have estab-
R genes may reside at single gene loci, like the flax L lished some trends in resistance gene evolution. Onelocus (Lawrence et al. 1995) and the Arabidopsis Rpm1 trend is the prevalence of diversifying selection in re-(Grant et al. 1995) and RPS2 (Mindrinos et al. 1994) gions encoding leucine rich repeats (LRRs). Sequenceloci, or they may belong to families of tightly linked comparisons of different members of resistance genegenes. Examples of the latter include the tobacco N families have found a significantly higher ratio of non-
synonymous substitutions to synonymous substitutions,particularly in codons encoding predicted solvent-
Corresponding author: Scot Hulbert, Department of Plant Pathology, exposed residues potentially involved in ligand binding.Throckmorton Hall, Kansas State University, Manhattan, KS 66506-5502. E-mail: [email protected] This has now been documented for a number of R
Genetics 158: 423–438 (May 2001)

424 Q. Sun et al.
gene families (Parniske et al. 1997; Botella et al. 1998; gene (Collins et al. 1999) has provided probes foranalysis of recombinant Rp1 haplotypes. This providedMcDowell et al. 1998; Meyers et al. 1998b; Wang et al.
1998; Noel et al. 1999; Ayliffe et al. 2000). molecular evidence that Rp1 genes are a family of tightlylinked genes that mispair in different arrangements inAnother trend is the involvement of recombination
in R gene evolution. The flax L locus is a simple locus meiosis, with subsequent recombination events creatingnew Rp1 haplotypes. The identification of new restric-with many alleles. Intragenic recombinants have been
identified in the progeny of L heterozygotes (Shepherd tion fragments in gel blot analysis of DNAs from someof the variants indicated that many of the recombinationand Mayo 1972; Ellis et al. 1997). For example, a cross-
over between L2 and L10 gave rise to suL10 with no events were occurring close to and possibly within theRp1 genes.detectable resistance specificity. A second crossover be-
tween suL10 and L9 resulted in RL10, a revertant ex- The HRp1-D haplotype carries the Rp1-D gene andeight homologous sequences (paralogues) as estimatedpressing the L10 specificity (Islam et al. 1989; Ellis et
al. 1997). Moreover, R genes like the L and M genes of by gel blot analysis. Here we characterize paraloguesfrom this haplotype and the HRp1-K haplotype to deter-flax have sequence duplications in the LRR region that
can mispair in meiosis. Three susceptible M gene vari- mine the nature of these genes. We also characterizethe haplotypes and recombinant genes in a collectionants presumably derived from unequal crossover events
were identified that carried only one direct repeat unit of spontaneous variants to identify the type of geneticevents that can generate variability at this locus.in their LRR instead of the two present in the parental
allele (Anderson et al. 1997). Variation in the numberof direct repeats in different alleles of the L locus is
MATERIALS AND METHODSprobably generated by similar processes (Ellis et al.1999). This type of intragenic recombination was also Genomic DNA libraries: Genomic DNA libraries of the
HRp1-D*21 and HRp1-D*24 lines were constructed by ligatingobserved at the Rpp5 locus of Arabidopsis where anSau3AI partially digested, glycerol gradient fractionated, geno-allele with a duplication of four LRRs was identified inmic DNA fragments to BamHI-digested EMBL3 l-vector (Pro-
a susceptible variant (Parker et al. 1997). Other gene mega, Madison, WI). The DNA library of the wild-type Rp1-Dfamilies, such as the Cf2/5 locus (Dixon et al. 1998), line was made by ligating Sau3AI partially digested, sucrose
gradient-fractionated, genomic DNA fragments to BamHI/also have duplicated stretches with sequence homologyXhoI double-digested l DASHII vector arms (Stratagene, Lain the LRR regions and also show variation in repeatJolla, CA). Ligation products were packaged using Goldennumber.II packaging kit (Stratagene). Library screening and l DNA
In families of closely linked R genes, genes at different preparations were performed as described (Sambrook et al.positions in the array may pair and recombine. A suscep- 1989).
Genomic clone characterization: Genomic clones from thetible variant derived from such an unequal crossing-HRp1-D haplotype were classified by amplifying the same re-over event was identified and molecularly characterizedgion from each clone using a pair of conserved rp1 primersat the tomato Cf-2/Cf-5 locus (Dixon et al. 1998). Fur-for the 59 ends of the rp1 gene family members. The forward
ther evidence is provided by comparing the patterns of primer sequence was P6, AGCTTCAGCTTACCTCAGTG, andsequence affinities between different family members. the reverse primer sequence was 1520R, CCAATCCAACAAT
GGCCAAAC. l-Clones were partially restriction mapped andThe mosaic pattern of DNA homologies of severaltested for their ability to hybridize to probes made from thecloned R genes and their family members suggests thatextreme 59 and 39 ends of the Rp1-D gene. Selected full-lengthmultiple steps of recombination between paraloguesl-clones from each sequence group were subcloned into
occurred during their evolution (Parniske et al. 1997; pUC19 for sequencing using the Big-Dye sequencing systemMcDowell et al. 1998; Ellis et al. 1999; Noel et al. (Applied Biosystems, Foster City, CA). Sequencing primers
were positioned throughout the gene sequence, and new prim-1999).ers were designed for specific genes where their sequencesThe rp1 locus of maize carries genes that conditiondiverged from the others.resistance to Puccinia sorghi, the causal agent of common
Sequencing data analysis: Sequence data were analyzed usingrust. Early recombination analyses, which showed that computer programs from the University of Wisconsin GeneticsRp1 genes could be recombined into coupling phase, Computer Group package Version 10. Sequence alignment was
done using the Clustal W program. The significance of differ-demonstrated that it was not a simple locus (Saxenaences in synonymous and nonsynonymous substitution rates wasand Hooker 1968). Additional recombination analysesexamined by using a 2 3 2 contingency table G-test (Zhangwith flanking markers indicated that the complex didet al. 1997). The sequences of the rp1 paralogues in the HRp1-D
not behave like a cluster of distinct loci. Instead, the haplotype were submitted to GenBank and given the followinggenes appeared to mispair in a number of ways as they accession numbers: rp1-dp1, AF342991; rp1-dp2, AF342992;
rp1-dp3, AF324993; rp1-dp5, AF342994; rp1-dp6, AF342995;recombined (Sudupak et al. 1993; Hu and Hulbertrp1-dp7, AF342996; rp1-dp8, AF342997; rp1-kp1, AF344308;1994). Recombination events in the rp1 complex haverp1-kp2, AF344311; and rp1-kp3, AF344309.also been associated with the generation of variants with
cDNA cloning and reverse transcriptase-PCR: The construc-new phenotypes, including novel race specificities tion of a cDNA library and cDNA cloning and sequencing(Richter et al. 1995) and lesion-mimic phenotypes (Hu were conducted as described previously (Collins et al. 1999).
RNA was isolated from fully expanded third leaves of 10-day-et al. 1996). Recent molecular identification of the Rp1-D

425Recombination at rp1
old seedlings. A primer pair was designed flanking the 39 28 bp downstream of the stop codon. Alignments of theintron and used in the reverse transcriptase (RT)-PCR to elimi- deduced amino acid sequences of the coding regionsnate the possibility of genomic DNA contamination (forward
of the nine paralogues are shown in Figure 1. Shortprimer, CTGTATTGCTCAACCACATGC; reverse primer,regions that were highly variable in both sequence andCCTGAACTCTGGAGCTTCAAG). RT-PCR was conducted us-
ing the RT-PCR kit (Stratagene) as recommended by the man- length were observed at two locations corresponding toufacturer. Following first strand synthesis using an oligo(dT) amino acids 505–512 and 671–677 in the Rp1-D codingprimer, several separate PCR reactions were conducted on region. These hypervariable regions were designatedeach sample and pooled before cloning.
regions A and B in Figure 1. Region A occurs shortlyPCR-based cloning and sequencing of recombinant genes:upstream of the LRR domain. The fourth paralogueFragments from most of the recombinant genes were PCR
amplified and cloned using gene-specific primers. Three Rp1-D- (rp1-dp4) encodes a truncated NBS-LRR protein thatspecific primers were used as reverse primers, to extend the corresponded to a previously identified cDNA from thenoncoding strand. The DR1 primer (TTTCCTCCGGAACCA Rp1-D haplotype (rp1-Cin4; GenBank accession no.GAACAC) was designed from sequences in a highly polymor-
AF107294; Collins et al. 1999). The 59 half of the genephic area corresponding to the C domain in the LRR of theis similar to the NBS encoding 59 half of the Rp1-DRp1-D gene. The DR4 (TAGCGGAGCAATACAAGCGGC) and
DR5 (GGCCACATGAATGATATAGC) primers correspond to gene but the 39 half of the gene is replaced by z480sequences in the 39-untranslated region (39-UTR). Primers nucleotides with homology to the Cin4 family of retro-from more conserved sequences were usually used as forward transposon-like elements (Schwarz-Sommer et al. 1987).primers. The P6 primer (AGCTTCAGCTTACCTCAGTG) cor-
The deduced amino acid sequence is 99% identical toresponds to conserved sequences starting 67 nucleotidesthat of Rp1-D until amino acid 462, after which thedownstream of the putative start codon and was the most
commonly used forward primer. Rp1-D-specific forward sequence diverges and the open reading frame ends.primer 1507F (TTGGTTTTTCATATCTATTGTGAT) was To determine if the truncated gene was caused by aused to amplify the recombinant gene in the HRp1-BD12 hap- simple insertion of a Cin4-like element, 13 kb of geno-lotype because gel blot analysis indicated it had the 59 end of
mic DNA downstream of the rp1-dp4 gene was subclonedthe Rp1-D gene. The primer corresponds to sequences in theand analyzed. A lack of hybridization to the 39 half of thebeginning of the LRR sequence. PCR products were cloned
by using the TOPO TA-cloning kit (Invitrogen, Carlsbad, CA). Rp1-D gene indicated that the gene was not truncated bya simple insertion of a small element. Sequence analysisof z5 kb 39 to the rp1-dp4 gene found no significant
RESULTSORFs. No significant homology to GenBank sequenceswas observed for the first 1.5 kb. The next 1.5 kb wasDNA sequence variation among rp1 genes from the
HRp1-D haplotype: Genomic DNA gel blot analysis indi- highly homologous (.87%) to the first 1.5 kb of a largeUTR region of the GapC2 gene (GenBank accessioncated that the HRp1-D haplotype contains approxi-
mately nine Rp1 paralogues, rp1-dp1 to rp1-dp9, with rp1- no. 312178) coding for glyceraldehyde-3-phosphate de-hydrogenase. Furthermore, hybridization of this se-dp9 corresponding to the Rp1-D gene (see below). Thirty
l-clones isolated from an Rp1-D genomic library by ho- quence to gel blots indicated that it occurs .50 timesin the maize genome and also occurs in the untranslatedmology with the cloned Rp1-D gene (Collins et al. 1999)
were resolved into nine classes by PCR amplifying and region of at least one other maize gene (accession no.1213278). Comparisons of PCR products from the l-clonesequencing clones containing a 600-bp segment of the
NBS encoding region. When possible, a single full- and genomic DNA from the Rp1-D line, using primers forRp1 and GapC2 sequences, indicated that the l-clone waslength l-clone was selected from each group by testing
for hybridization to probes from both the 59 and 39 ends not chimeric. Identically sized products were generatedfrom both genomic DNA and l-DNA templates.of the Rp1-D gene. Sequence analysis of these clones
indicated they represented the Rp1-D gene and eight The HRp1-K haplotype generally has more fragmentsin gel blot analysis than HRp1-D, although it is difficulthomologous family members.
Comparisons of the DNA sequences of coding regions to determine gene number because of the possibilityof comigrating fragments (data not shown). Three rp1of different rp1 family members reveals high levels of
sequence identity (91–97%) to the Rp1-D rust resistance family members from the HRp1-K haplotype werecloned and sequenced. In general, the rp1 genes fromgene (Table 1). The 39 halves of the genes, correspond-
ing to the LRR domain, were the most divergent. This the HRp1-K haplotype show high homology with thosefrom the HRp1-D haplotype. The rp1-kp1, rp1-kp2, andis especially true of the rp1-dp3 and rp1-dp1 genes, which
have only 87 and 88% identity with the Rp1-D gene rp1-kp3 genes have 96, 97, and 93% sequence identity,respectively, when compared with the Rp1-D gene. Inter-in their 39 halves. This trend can also be observed by
comparing the deduced amino acid sequences of the estingly, the rp1-kp2 gene had a point mutation thatresulted in a stop codon only 111 bp after the putativegenes (Table 1). Of the nine paralogues in the HRp1-D
haplotype, eight are similar in size, containing an open start codon. There were no other premature stop co-dons observed in the rest of the gene. The other tworeading frame (ORF) ranging from 3800 to 3862 bp in
length without introns. Small introns occur at positions HRp1-K genes have full-length ORFs similar to most ofthe HRp1-D genes.36 bp upstream of the proposed Rp1-D start codon and

426 Q. Sun et al.
TABLE 1
Similarities of rp1 family members to the Rp1-D gene
% nucleotide identity with Rp1-D % amino acid similarity with Rp1-D
Paralogue Whole ORF N terminus C terminus Whole ORF N terminus C terminus
rp1-dp6 97 98 96 94 98 92rp1-dp2 96 98 95 93 97 91rp1-dp7 95 98 94 93 97 90rp1-dp5 94 93 96 92 92 92rp1-dp3 91 98 87 88 97 82rp1-dp1 91 96 88 88 95 83rp1-dp8 91 93 90 88 92 86rp1-dp4 — 99 — — 99 —
Similarity scores between the ORF of Rp1-D and other rp1 family members were calculated by using theBestfit program in the GCG Wisconsin Package. The N terminus corresponds to the first 1485 nucleotides ofthe ORF of the Rp1-D gene and includes the NBS domain. The C terminus corresponds to the 39 2377nucleotides of the Rp1-D gene including the LRR domain. The ORF of the truncated rp1-dp4 gene ends 1396nucleotides after the beginning of the open reading frame.
Transcription of rp1 genes: Two different size tran- upstream of the putative translation start codons of therp1-dp2 and rp1-dp7 genes shows respective DNA se-scripts were previously observed on RNA gel blots with
Rp1-D probes. The larger transcript was z4 kb and corre- quence identities of 97 and 98% to the Rp1-D gene. Thehomologous regions include an z320-bp UTR with asponded to the size of the Rp1-D gene. The smaller one
was z1.5 kb and corresponded to the size of the rp1-dp4 142- to 144-bp intron. Upstream of this 1200-bp region,the sequences of all three genes show homology to the(rp1-Cin4) gene. Since most of the rp1 family members
would not be distinguishable from Rp1-D by RNA gel same retroelement sequence. Sequence homology ofthese three genes may extend beyond the determinedblot analysis, we used partial sequence analysis of cDNAs
to determine if other family members were transcribed. retroelement sequence point, but this was not exam-ined. The rp1-dp3 gene was also highly similar to Rp1-D forMost of the z80 cDNA clones (selected with an NBS
area probe) were derived from the rp1-dp4 gene. The the 420 bp preceding the start codon, but no additionalsequence was available from clones carrying this gene.sequence of 10 clones matched the Rp1-D sequence and
the remaining 2 clones matched the respective se- The 553 bp preceding the translation start codon of therp1-dp4 gene is also highly homologous to Rp1-D. Itquences of rp1-dp7 and rp1-dp8. RT-PCR was used to
identify additional transcribed members. Two PCR shows no noticeable homology upstream of this pointand shows clear retroelement homology at z730 nucleo-primers flanking the small intron in the 39-UTR of the
genes were designed from conserved sequences so that tides before the putative translation start. Since therp1-dp4 gene is transcribed, this may indicate that thethey would amplify all nontruncated rp1 genes. Cloning
and sequencing of 26 PCR products identified 6 identi- required 59 regulatory regions are included in these 553nucleotides. The rp1-dp6 gene is also homologous tocal to Rp1-D, 10 identical to rp1-dp7, and 4 identical to
rp1-dp8. The remaining 6 sequences did not perfectly Rp1-D beginning 937 nucleotides before the start codon.One exception to this homology is a 45-bp insertionmatch the sequence of any of the genomic clones and
differed from Rp1-D, rp1-dp7, or rp1-dp8 at one or more located 401 nucleotides 59 to the translation start codonin rp1-dp6, which may be responsible for the apparentnucleotides, probably due to errors introduced during
PCR amplification. Gene-specific primers were used in lack of transcription of this gene. The 59 noncodingregions of rp1-dp1 and rp1-dp8 show only low homologyRT-PCR experiments to determine if any of the other
genes are transcribed. Besides the previously detected to Rp1-D and the other homologues and are more simi-lar to each other, at least for the first 200 bp before themembers, 3 additional members, rp1-dp1, rp1-dp2, and
rp1-dp3, were found to be expressed. No RT-PCR prod- start codon. Analysis of a 59 RACE PCR-generated clonecorresponding to the rp1-dp1 gene indicated it had aucts could be identified for the rp1-dp5 and rp1-dp6
genes, indicating they may not be transcribed. Taken 513-bp UTR including a 95-bp intron. When comparedto the 59-UTR of the Rp1-D gene, rp1-dp1 and rp1-dp8together, these data indicate that there are at least 7
rp1 family members in the HRp1-D haplotype that are have numerous differences including insertions (up to81 bp) and deletions and a stretch of 67 bases withexpressed.
Comparison of the 59 and 39 sequence flanking para- no significant sequence similarity. Several stretches ofsequence homology (.80% identity) occur, however,logues in the HRp1-D haplotype: Some of the rp1 genes
in the HRp1-D haplotype have distinct promoter regions, including areas close to the transcription start sites andthe 59 intron splice site. There is no significant homol-as shown in Figure 2. The region consisting of 1200 bp

427Recombination at rp1
Figure 1.—Alignment of aminoacid sequences of the Rp1-D geneproduct with the seven other full-length genes from the Hrp1-D haplo-type and three genes from the HRp1-Khaplotype. Amino acids correspond-ing to long stretches of DNA sequenceidentity between different genes aresimilarly boxed or shaded. Hypervari-able regions (A and B), with frequentamino acid substitutions and small in-dels, are underlined. Deletions are in-dicated by dashes. Dots representamino acids identical to the Rp1-D se-quence.
ogy observed 20 bp upstream of the putative transcrip- until z340 bp after the translation stop codon of theRp1-D gene. While the 39-UTR regions in the differenttion start site of the rp1-dp1 gene and beyond this the
rp1-dp1 and rp1-dp8 genes showed no homology to the genes do not show the structural variation characteristicof the 59-UTRs, certain pairs of genes showed sequenceother paralogues or to each other. The 59-UTR se-
quence of the rp1-dp5 gene was similar to the rp1-dp1 affiliation. For example, the rp1-dp5 and Rp1-D are verysimilar (.98%) in this region. Some pairs of genesand rp1-dp8 genes but diverges from these genes 276
bases before the start codon and is missing the 59 splice retained their sequence affiliation after the others di-verged. For example, the rp1-dp7 and rp1-dp8 genes weresite of the intron in these two genes. Furthermore,
retroelement homology was observed in rp1-dp5 at a identical from 450 to 650 bases downstream of the trans-lation stop codon. Interestingly, these two genes werepoint corresponding to the transcription initiation site
in other gene family members, and this presumably among the most different for the 59-UTR region.Patterns of polymorphism among the coding regionsaccounts for the lack of expression of the rp1-dp5 gene.
Sequence comparisons of the 39 noncoding regions of the rp1 paralogues: Alignment of the DNA sequencesof the rp1 family members allowed the relationshipsrevealed that the rp1 genes retained sequence homology

428 Q. Sun et al.
Figure 1.—Continued.
of the genes to be examined in detail. No consistent haplotypes (Figure 1). Changes in affiliation are alsoapparent when comparing sequences flanking the cod-sequence affiliation between any pair of rp1 paralogues
was observed throughout the entire ORF. Instead, the ing regions as detailed above. This mosaic nature ofsequence identities implies that reassortment of se-coding regions of the genes are composed of mosaics,
or patchworks of stretches of similar or identical DNA quences has occurred through multiple genetic ex-change events during the evolution of the gene family.sequences, some of which are highlighted in Figure 1.
For example, shortly after the start codon, rp1-dp8 and Evidence for diversifying selection preferentially inthe C-terminal half of the LRR domain: Patterns ofrp1-dp5 share a 370-bp stretch of sequence identity. To-
ward the middle of the gene, rp1-dp8 and rp1-dp5 diverge nucleotide substitution can be informative in assessingthe type of selection pressure acting on the evolutionfrom each other and, instead, rp1-dp8 now has a stretch
of 240 bp that matches an identical stretch in the rp1-dp3 of gene family members. The rates of synonymous andnonsynonymous nucleotide substitutions were calcu-gene corresponding to amino acids 601–680 (Figure 1).
Near the 39 end of the gene, rp1-dp8 is again identical lated between all possible pairs of the rp1 genes from theHRp1-D haplotype, except the truncated rp1-dp4 gene.to rp1-dp3 for 242 nucleotides corresponding to amino
acids 1177–1248 except for one uninformative differ- These were calculated separately for different regionsof the gene (Table 2). Rates of synonymous (Ks) andence. The stretches of identical sequence and changes
in sequence affiliation are also observed between other nonsynonymous (Ka) substitutions were higher in theC-terminal region of the gene (LRR encoding domain)family members from the HRp1-D haplotype and when
comparing members from the HRp1-D and HRp1-K than those in the N-terminal region of the gene. Ratios

429Recombination at rp1
were ,1 in some comparisons and .1 in others, averag-ing z0.81. When the sites in the LRRs that match thexxLxLxx consensus sequence (Jones and Jones 1996)were calculated separately, all of the gene pairs hadKa:Ks ratios .1 (7 of 28 pairs significantly .1 at P 50.01) and the average Ka:Ks was z3.1. These regions,based on structural similarity to the porcine ribo-nuclease inhibitor protein, are predicted to be solventexposed and potentially involved in ligand binding(Kobe and Deisenhofer 1995). Like the functionalRP1-D protein (Collins et al. 1999), the Rp1 proteinscontain two LRR regions (domains B and D) that areseparated by a region (domain C) that shows little ho-mology to the LRR consensus. When the Ka:Ks ratio wascalculated for the regions with xxLxLxx consensus inLRR domain B and domain D separately, domain D hadan average Ka:Ks ratio of 3.5 (15 of 28 gene pairs withFigure 2.—Differences in the upstream regions of the rp1a Ka:Ks ratio significantly .1 at P 5 0.01 level) whilegenes from the HRp1-D haplotype. Regions with high levels
of sequence identity (.90%) are indicated by the same box domain B showed an average Ka:Ks ratio of only 1.4 (onlypatterns. Differently shaded regions show no noticeable se- 3 of 28 pairs significantly .1). This is consistent withquence homology. Transcription starts at z2320 in the Rp1-D the results obtained from analysis of Ka:Ks ratios of thegene, relative to the start of the putative coding region. The
Rp1 homologues in barley (Ayliffe et al. 2000). It im-site of an intron in the 59-UTR that corresponds to bases 2179plies that the carboxy-terminal half of the LRR region into 236 in the Rp1-D gene is also shown. Boxes made from
dotted lines indicate regions with retroelement homology. A this gene family is under stronger diversifying selectionsolid triangle in the rp1-dp6 sequence indicates an apparent pressure than that of the amino-terminal half. The con-45-bp insertion. served leucine sites likely serve a structural role and
are probably under selection for sequence conservation(Parniske et al. 1997).of nonsynonymous to synonymous substitutions (Ka/Ks)
Organization of rp1 paralogues in the HRp1-D haplo-were generally ,1 in the N-terminal region, averagingtype: Construction of a contig map of the HRp1-D haplo-only 0.46 for 28 pairwise comparisons. Ka/Ks ratios intype was not possible using the genomic clones carryingthe C-terminal region of the gene, including the LRR,the rp1 paralogues. The clones averaged z20 kb in size,but none contained more than a single rp1 paralogue.
TABLE 2 Instead, genetic recombinants from three different pop-ulations were used to determine the order and orienta-Evidence for diversifying selection in the evolution
of the rp1 gene family tion of the genes in the HRp1-D haplotype.Variants from an Rp1-D homozygote: The first set of vari-
Domain Ka Ks Ka/Ks ants consisted of a previously described (Collins et al.1999) panel of mutants derived from an Rp1-D homozy-Whole gene 6 (1–10) 10 (3–16) 0.74gote. All but two of the variants were susceptible to allN terminusrust isolates tested (Richter et al. 1995). Two others(including NBS) 3 (1–7) 9 (4–18) 0.46
C terminus had a reduced-resistance phenotype and a nonspecific(including LRR) 8 (1–13) 10 (1–19) 0.81 lesion-mimic phenotype (Hu et al. 1996). Gel blot analy-
Domain B 9 (2–16) 13 (10–27) 1.18 sis with an Rp1-D probe indicated that they representedxxLxLxx 16 (0–28) 17 (0–29) 1.48 spontaneous deletions of variable sizes, presumablyDomain C 8 (3–14) 9 (10–18) 1.16
caused by unequal recombination events. They could beDomain D 8 (3–10) 8 (2–12) 1.18grouped into nine different classes by restriction analysisxxLxLxx 19 (9–26) 8 (1–18) 3.5with AccI, BglI, BglII, DraI, NcoI, and NsiI (Table 3).C-terminal tail 10 (3–19) 28 (4–48) 0.47
Variants from Rp1-D/Rp1-A heterozygotes: The secondValues are calculated for synonymous (Ks) and nonsynony-
set of recombinants consisted of 26 susceptible variantsmous (Ka) substitutions in different domains of the proteinfrom Rp1-A/Rp1-D heterozygotes. All but 3 (DNCO1,coding regions of the genes. Ka values indicate nonsynony-
mous substitutions per 100 nonsynonymous sites. Ks values DNCO2, and DNCO3) appeared to be derived fromindicate synonymous substitutions per 100 synonymous sites. crossovers as indicated by flanking marker analysis. TenValues were calculated for all possible 28 pairwise comparisons of these recombinants had the Rp1-D parent allele atbetween the eight rp1 paralogues in the HRp1-D haplotype
the proximal marker (umc285) and the Rp1-A parent(excluding the truncated rp1-Cin4 gene) and averaged. Theallele at bln3.04, and the remaining 13 had the oppositeranges of values are given in parentheses to the right of the
Ka and Ks averages. flanking marker combination. These apparently con-

430 Q. Sun et al.
TABLE 3
Classification of deletion variants based on restriction enzyme digestion and Southern blot analysis
NcoI fragments from Rp1-D parentRecombinant classa
FlankingRp1-D homozygotes 1 2 3 4 (Cin4) 5 6 7 8 9 Rp1-D markers
D*1, 2, 3, 8, 20, 22, 23, 27 1 1 1 1 1 2 2 2 2 NAb
D*4 1 1 2 2 2 2 2 2 2 NAD*5 1 1 1 1 1 1 2 2 2 NAD*10, 11, 14, 16, 29 1 1 1 1 1 2 2 2 2 NAD*17 1 1 1 1 1 2 2 2 2 NAD*19 1 1 1 1 1 2 2 2 2 NAD*21 1 1 2 2 2 2 2 2 2 NAD*24 1 2 2 2 2 2 2 2 2 NAD*7, 12, 28 1 1 1 1 1 2 2 2 2 NARp1-A 3 Rp1-DAD7, AD1, AD18 2 NDc ND ND 2 2 2 2 2 285-A, 3.04-DAD22 2 ND ND ND 2 2 2 2 2 285-A, 3.04-DAD19 2 ND ND ND 2 2 2 2 2 285-A, 3.04-DAD20, AD4, AD10 1 ND ND ND 2 2 2 2 2 285-D, 3.04-AAD5 1 ND ND ND 1 1 1 2 2 285-D, 3.04-AAD9 1 ND ND ND 1 1 1 1 2 285-D, 3.04-AAD2 1 ND ND ND 1 2 2 2 2 285-D, 3.04-AAD15 1 ND ND ND 1 2 2 2 2 285-D, 3.04-AAD8 2 ND ND ND 2 2 2 2 2 285-D, 3.04-ADNCO1 1 ND ND ND 1 1 1 1 2 285-D, 3.04-DDNCO2 1 ND ND ND 1 1 1 1 1 285-D, 3.04-DDNCO3 1 ND ND ND 1 1 1 1 1 285-D, 3.04-DRp1-B 3 Rp1-DBD11 1 1 ND 1 2 2 2 2 2 285-D, 3.04-BBD13 1 1 ND 1 1 1 1 2 2 285-D, 3.04-BBD15 1 1 ND 1 1 2 2 2 2 285-D, 3.04-BBD1 1 1 ND 1 1 2 2 2 2 285-D, 3.04-BBD14 1 1 ND 1 1 2 2 2 2 285-D, 3.04-BBD3 1 1 ND 1 1 2 2 2 2 285-D, 3.04-BBD2 1 1 ND 1 1 2 2 2 2 285-D, 3.04-BBD4 1 1 ND 1 1 2 2 2 2 285-D, 3.04-BBD7 1 1 ND 1 1 1 1 2 2 285-D, 3.04-BBD8 1 1 ND 1 1 1 1 2 2 285-D, 3.04-BBD9 1 1 ND 1 1 2 2 2 2 285-D, 3.04-BBD12 1 1 ND 1 1 1 1 1 1 285-D, 3.04-BBNCO1 2 2 ND 2 2 2 2 2 2 285-B, 3.04-BDNCO4 1 ND ND 1 1 1 1 1 1 285-D, 3.04-D
DNAs were purified from individuals that were homozygous for mutant or recombinant haplotypes that were selected fromF1 crosses of Rp1-D homozygotes and test crosses of Rp1-A/Rp1-D and Rp1-B/Rp1-D heterozygotes. DNAs were digested with NcoIand probed with a NBS region probe. NcoI fragments are labeled according to their inferred position, from the centromere totelomere end of the array, and refer to the numbering of the HRp1-D haplotype fragments in Figure 3.
a Members of each class were indistinguishable based on gel blot analysis with Rp1 probes after digestion with NcoI and severalother restriction enzymes.
b NA, flanking RFLP markers could not be analyzed in the Rp1-D homozygote derivatives due to the absence of markersegregation. The designations 285-B and 285-D indicate whether the recombinant had the Rp1-B or Rp1-D parent allele at thecentromere-proximal RFLP marker npi285. The designations 3.04-A and 3.04-D indicate whether the recombinant had the Rp1-A orRp1-D parent allele at the distal marker bnl3.04.
c ND indicates we could not determine the presence of the HRp1-D haplotype fragment because a fragment of similar size waspresent in the other Rp1 parent.
flicting data have previously been used as evidence for NcoI (Table 3). The recombinants generally had restric-tion fragments from each of the two parental haplotypesthe occurrence of mispairing leading to unequal cross-
overs (Hulbert et al. 1997). Homozygous lines were and showed a great variety of different combinations offragments (data not shown).established from 16 of these variants, including the
three noncrossovers, and their rp1 haplotypes were ex- Variants from Rp1-D/Rp1-B heterozygotes:The third set con-sisted of 22 susceptible variants derived from Rp1-D/amined with multiple restriction enzymes, including
431Recombination at rp1
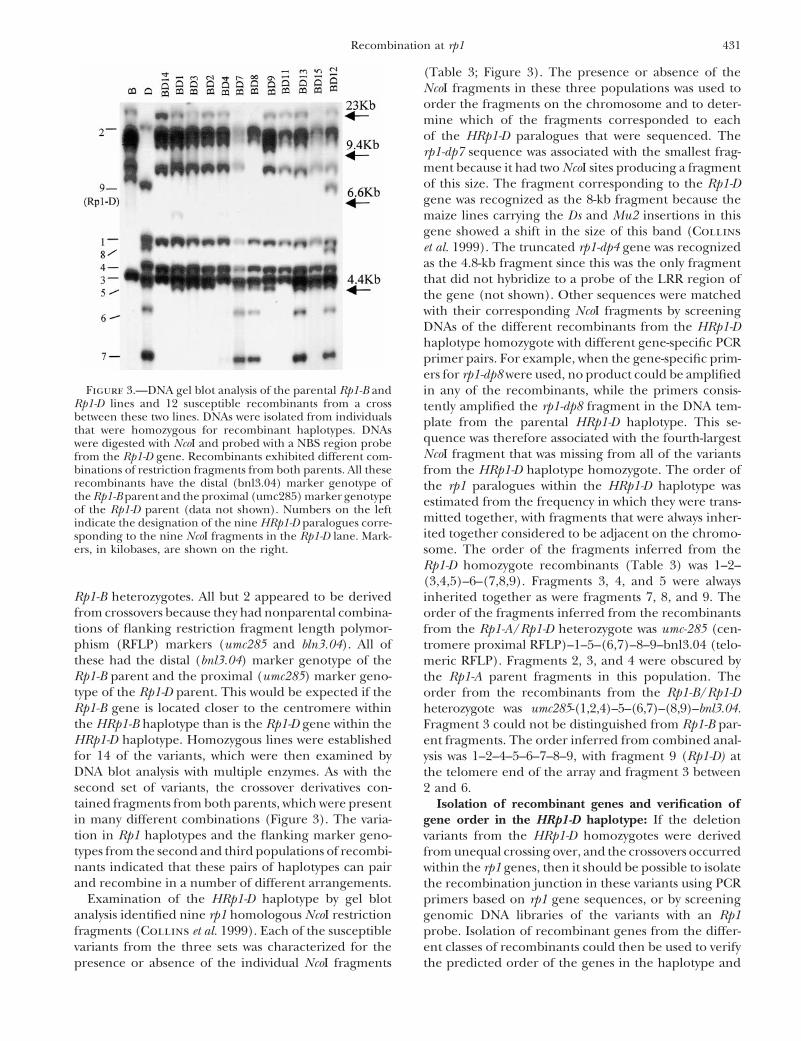
(Table 3; Figure 3). The presence or absence of theNcoI fragments in these three populations was used toorder the fragments on the chromosome and to deter-mine which of the fragments corresponded to eachof the HRp1-D paralogues that were sequenced. Therp1-dp7 sequence was associated with the smallest frag-ment because it had two NcoI sites producing a fragmentof this size. The fragment corresponding to the Rp1-Dgene was recognized as the 8-kb fragment because themaize lines carrying the Ds and Mu2 insertions in thisgene showed a shift in the size of this band (Collinset al. 1999). The truncated rp1-dp4 gene was recognizedas the 4.8-kb fragment since this was the only fragmentthat did not hybridize to a probe of the LRR region ofthe gene (not shown). Other sequences were matchedwith their corresponding NcoI fragments by screeningDNAs of the different recombinants from the HRp1-Dhaplotype homozygote with different gene-specific PCRprimer pairs. For example, when the gene-specific prim-ers for rp1-dp8 were used, no product could be amplified
Figure 3.—DNA gel blot analysis of the parental Rp1-B and in any of the recombinants, while the primers consis-Rp1-D lines and 12 susceptible recombinants from a cross tently amplified the rp1-dp8 fragment in the DNA tem-between these two lines. DNAs were isolated from individuals plate from the parental HRp1-D haplotype. This se-that were homozygous for recombinant haplotypes. DNAs
quence was therefore associated with the fourth-largestwere digested with NcoI and probed with a NBS region probeNcoI fragment that was missing from all of the variantsfrom the Rp1-D gene. Recombinants exhibited different com-
binations of restriction fragments from both parents. All these from the HRp1-D haplotype homozygote. The order ofrecombinants have the distal (bnl3.04) marker genotype of the rp1 paralogues within the HRp1-D haplotype wasthe Rp1-B parent and the proximal (umc285) marker genotype estimated from the frequency in which they were trans-of the Rp1-D parent (data not shown). Numbers on the left
mitted together, with fragments that were always inher-indicate the designation of the nine HRp1-D paralogues corre-ited together considered to be adjacent on the chromo-sponding to the nine NcoI fragments in the Rp1-D lane. Mark-
ers, in kilobases, are shown on the right. some. The order of the fragments inferred from theRp1-D homozygote recombinants (Table 3) was 1–2–(3,4,5)–6–(7,8,9). Fragments 3, 4, and 5 were always
Rp1-B heterozygotes. All but 2 appeared to be derived inherited together as were fragments 7, 8, and 9. Thefrom crossovers because they had nonparental combina- order of the fragments inferred from the recombinantstions of flanking restriction fragment length polymor- from the Rp1-A/Rp1-D heterozygote was umc-285 (cen-phism (RFLP) markers (umc285 and bln3.04). All of tromere proximal RFLP)–1–5–(6,7)–8–9–bnl3.04 (telo-these had the distal (bnl3.04) marker genotype of the meric RFLP). Fragments 2, 3, and 4 were obscured byRp1-B parent and the proximal (umc285) marker geno- the Rp1-A parent fragments in this population. Thetype of the Rp1-D parent. This would be expected if the order from the recombinants from the Rp1-B/Rp1-DRp1-B gene is located closer to the centromere within heterozygote was umc285-(1,2,4)–5–(6,7)–(8,9)–bnl3.04.the HRp1-B haplotype than is the Rp1-D gene within the Fragment 3 could not be distinguished from Rp1-B par-HRp1-D haplotype. Homozygous lines were established ent fragments. The order inferred from combined anal-for 14 of the variants, which were then examined by ysis was 1–2–4–5–6–7–8–9, with fragment 9 (Rp1-D) atDNA blot analysis with multiple enzymes. As with the the telomere end of the array and fragment 3 betweensecond set of variants, the crossover derivatives con- 2 and 6.tained fragments from both parents, which were present Isolation of recombinant genes and verification ofin many different combinations (Figure 3). The varia- gene order in the HRp1-D haplotype: If the deletiontion in Rp1 haplotypes and the flanking marker geno- variants from the HRp1-D homozygotes were derivedtypes from the second and third populations of recombi- from unequal crossing over, and the crossovers occurrednants indicated that these pairs of haplotypes can pair within the rp1 genes, then it should be possible to isolateand recombine in a number of different arrangements. the recombination junction in these variants using PCR
Examination of the HRp1-D haplotype by gel blot primers based on rp1 gene sequences, or by screeninganalysis identified nine rp1 homologous NcoI restriction genomic DNA libraries of the variants with an Rp1fragments (Collins et al. 1999). Each of the susceptible probe. Isolation of recombinant genes from the differ-variants from the three sets was characterized for the ent classes of recombinants could then be used to verify
the predicted order of the genes in the haplotype andpresence or absence of the individual NcoI fragments

432 Q. Sun et al.
their orientation on the chromosome. Among the vari-ants, the HRp1-D*24 haplotype has the largest deletionwith only a single rp1 family member remaining. A dele-tion causing a haplotype with only a single family mem-ber could be generated by recombination between thetwo genes located at opposite ends of the gene cluster.The recombinant gene from the HRp1-D*24 haplotypewas cloned from a genomic library constructed by usingthe genomic DNA from the recombinant. Comparisonof the sequences of the entire coding region of the clonefrom the HRp1-D*24 haplotype with that of the Rp1-Dand rp1-dp1 genes indicated that the recombinant geneconsists of the 59 end of rp1-dp1 and the 39 end of theRp1-D gene. This verifies that the two flanking mem-bers of the gene cluster are rp1-dp1 and Rp1-D. TheHRp1-D*21 haplotype has two NcoI fragments whenprobed with an Rp1 probe, indicating that it carriestwo family members. The recombinant gene fromHRp1-D*21 was cloned from a genomic library and wasfound to be a chimeric gene composed of the 59 halfof rp1-dp2 and 39 half of Rp1-D. This verifies that therp1-dp2 gene is the second gene from the centromereend of the array.
Other recombinant genes were cloned after PCR am- Figure 4.—Molecular characterization of unequal recombi-plification, utilizing the knowledge that the Rp1-D gene nation events from an HRp1-D haplotype homozygote. (A)is the distal gene in the array and that intragenic un- Structure of the HRp1-D haplotype and recombinant haplo-
types. Boxes represent the coding regions of the rp1 para-equal recombination events leading to its inactivationlogues corresponding to the gene designations at the top.should involve this gene. Like the recombinant genesGenes shown for each of the recombinant haplotypes arefrom the HRp1-D*21 and HRp1-D*24 haplotypes, other those that were not deleted by the unequal crossover events.
recombinant genes would be expected to be identical The two paralogues involved in the recombination event areto Rp1-D downstream of the crossover and identical to shown as partial genes flanking the deletion. Numbers in
parentheses with the gene designations indicate the percent-the other gene proximal to this. We therefore designedage sequence identity between the paralogue and the Rp1-Dprimer pairs to specifically amplify recombinant genesgene. Intergenic regions are not drawn to scale. (B) Recombi-from other deleted haplotypes. The upstream primers nant genes resulting from the unequal crossovers between
were generally non-gene specific, while the reverse prim- Rp1-D and the other gene family members in the HRp1-Ders were designed to be reasonably specific for Rp1-D haplotype. The points where the shading pattern changed
indicate the recombination breakpoints. Letters A, B, C, D,gene sequences on the basis of comparisons to otherand E represent the five domains of the Rp1-D gene (Collinsparalogues. Attempts were made to identify and se-et al. 1999) with A corresponding to the NBS domain and B–Equence the recombination junction from at least one corresponding to the LRR region.
member of each of nine classes (Table 2) of recombi-nants and multiple members of the larger classes. Therecombination junction amplified from the HRp1-D*5 termined by a lack of amplification with pairs of specific
primers, including those corresponding to sequenceshaplotype contained the 59 end of rp1-dp7 and the 39end of Rp1-D. This haplotype had the smallest deletion in the 39-UTR of Rp1-D. In addition, sequence analysis
of rp1-dp3 in the HRp1-D*4 haplotype and of rp1-dp5 inwith only two family members missing, as estimated byDNA gel blot analysis. The remaining nine recombina- HRp1-D*17 indicated that these genes were intact. These
are the genes that would have been expected to havetion junctions were isolated from haplotypes HRp1-D*2,D*3, D*7, D*8, D*10, D*19, D*20, D*23, and D*28 and recombined with Rp1-D on the basis of the number of
family members shown by DNA gel blot analysis to bewere each composed of a 59 region from rp1-dp5 and a39 region from Rp1-D. The exchange points could be left in the haplotypes. These two deletions may have
been caused by unequal recombination events that didlocated to intervals that were defined by polymorphismsbetween the recombined paralogues. As indicated in not involve the rp1 genes, or they may have been caused
by other types of events. Because they were isolated fromFigure 4, the recombination points were found to haveoccurred throughout the gene. a background with an active Ac element, it is possible
they were transposon-mediated events. In total, four ofNo recombination junctions were isolated from theHRp1-D*4 and HRp1-D*17 haplotypes. There were no the rp1 family members were observed to recombine
with Rp1-D (Figure 4). Presumably, these genes are inRp1-D sequences present in these two haplotypes as de-

433Recombination at rp1
the same orientation as Rp1-D, or the crossovers wouldhave caused chromosomal aberrations. Thus all fivegenes have their 39 LRR regions oriented toward thetelomere while the orientation of the other genes is notknown.
Effects of recombination events on Rp1 gene expres-sion and function: Most of the variant HRp1-D haplo-types, except HRp1-D*4 and HRp1-D*17, were derivedfrom unequal crossovers involving the Rp1-D gene andretained at least the 39 end of this gene. Therefore, theN-terminal sequences can be examined to provide in-sight into why the recombinant genes have null or modi-fied resistance phenotypes. Most of the recombinantgenes have the 59 regulatory sequences of the rp1-dp5gene. Since no transcript could be found for this gene,and its 59 regulatory sequences were truncated, the recom- Figure 5.—Phenotypes of the Rp1-D line and the HRp1-binant genes are probably not transcribed. For example, D*5 recombinant after inoculation with avirulent and virulentthe recombinant gene isolated from the HRp1-D*19 haplo- rust isolates. Rust isolate KS1 is virulent on the Rp1-D line
while isolates I-4, IN1, and KS2 are avirulent. The HRp1-D*5type was found to consist of mainly the Rp1-D sequencehaplotype confers resistance to the same rust biotypes as thebut with the first 354 bases of the coding region of theRp1-D gene, but has a reduced level of resistance, allowingrp1-dp5 gene. RT-PCR of RNA from this variant using some uredinia to form.
Rp1-D specific primers detected no transcript, as ex-pected if the promoter of rp1-dp5 is not functional. Re-combinants with the N terminus of rp1-dp5 are therefore the lack of resistance in the recombinant gene is not
apparent. Apparently, the 59 region of the rp1-dp1 geneuninformative in determining whether the novel codingregions code for functional proteins. either is nonfunctional or will not function with the
LRR of the Rp1-D gene to confer the Rp1-D resistanceThree of the recombinant genes involved Rp1-D andother transcribed family members. As shown in Figure specificity. Alternatively, the rp1-dp1 promoter may be
weaker than the Rp1-D promoter, thereby leading to an5, the D*5 haplotype is unique in that it is the onlyline that conferred the same resistance specificity as absence of resistance.
The recombinant gene in the HRp1-D*21 haplotypethe parental line but showed a lower level of resistance(partial resistance) to most rust biotypes to which the was derived from a crossover in the LRR encoding re-
gion, such that the predicted protein has the NBS do-parental line was resistant (Richter et al. 1995). Com-parison of the recombinant gene to the two parental main and the first 13 of the 15 repeats of the first LRR
region (domain B) from rp1-dp2. The remainder of thegenes located the crossover to a region spanning 202bases that are identical between the two genes. The LRR region comes from Rp1-D. The recombinant haplo-
type has lost the Rp1-D specificity, but still confers apromoter region and first 818–1020 bases of the codingregion come from the rp1-dp7 gene while the remainder highly necrotic reaction to rust inoculation and a lesion-
mimic phenotype (Hu et al. 1996). This nonspecificcomes from the Rp1-D gene. Therefore, the recombi-nant gene in the HRp1-D*5 haplotype has an identical phenotype may be caused by the recombinant gene. It
is also possible, however, that the phenotype is causedLRR to the functional Rp1-D allele, but its 59 regulatorysequences and part of the NBS region are different. by the unique combination of gene sequences that oc-
curs in this haplotype.This is consistent with the idea that the LRR domaincontrols specificity in this class of plant disease resistance All but one of the crossover derivatives of the Rp1-A/
Rp1-D and Rp1-B/Rp1-D heterozygotes lost the 8-kb NcoIgenes, but does not preclude other areas of the genefrom contributing to this function. fragment that represents the Rp1-D gene. The Rp1-BD12
haplotype from the Rp1-B/Rp1-D heterozygote had re-The coding region of the recombinant gene in theHRp1-D*24 haplotype has the whole LRR region of the combinant flanking RFLP markers and had rp1 homolo-
gous fragments from both parental haplotypes, indicat-Rp1-D gene, but the first 1385–1389 bases of the ex-pressed rp1-dp1 gene. The recombinant protein in- ing that it was derived from a crossover. Unlike the other
crossover variants, however, it had the 8-kb NcoI fragment.cludes the NBS domain of rp1-dp1 and adjacent aminoacids until 7 amino acids before the conserved amino To determine if this represents an intact Rp1-D gene, most
of the coding region was amplified with Rp1-D-specificacids MHD, which commonly occur before the LRR inNBS-LRR proteins. The predicted protein has only 28 primers, cloned, and sequenced. The recombinant gene
consisted of the 59 region of Rp1-D and a 39 sequence ofamino acid (aa) differences from the Rp1-D protein,and 32 differences from the product of the functional a gene that does not correspond to any of the HRp1-D
sequences and presumably came from an HRp1-B haplo-gene from the HRp1-D*5 haplotype. The reason for

434 Q. Sun et al.
type gene. The first 2107 bp of the coding region con-tains sequences from the Rp1-D gene encoding the NBSdomain and the first 10 LRR, while the remainder ofthe sequence is derived from a gene from the HRp1-Bhaplotype. Because this gene contains the functionalRp1-D promoter, its loss of function is likely due to therecombinant nature of the protein product.
Analysis of variants that are not associated with cross-overs: While most of the susceptible variants analyzedwere caused by crossovers, five variants from Rp1 hetero-zygotes did not show flanking marker exchange. Twononcrossover (NCO) variants were isolated from Rp1-B/Rp1-D heterozygotes. The first was a previously describedMu2 insertion mutant (Collins et al. 1999) containingRp1-D parent alleles for both flanking RFLP markers.The second was designated HRp1-BNCO1 and had theflanking marker alleles of the Rp1-B parent. Gel blotanalysis of HRp1-BNCO1 DNA with several enzymes(AccI, BamHI, BglII, EcoRI, EcoRV, HindIII, NcoI, SalI,XbaI, and XhoI) found the restriction patterns to be Figure 6.—Haplotypes of noncrossover (NCO) type mu-indistinguishable from the Rp1-B parent, indicating that tants from crosses with Rp1-A/Rp1-D and Rp1-B/Rp1-D hetero-
zygotes. DNAs of individuals that are homozygous for thethe mutation in the Rp1-B gene was not caused by avariant haplotypes were digested with NcoI and probed withcrossover or an insertion of a transposable element (Fig-the NBS region probe of the Rp1-D gene. Lanes 1 and 2 areure 6). The HRp1-DNCO1, HRp1-DNCO2, and HRp1- the Rp1-A and Rp1-D parents. Lanes 3–6 are noncrossover
DNCO3 haplotypes came from Rp1-A/Rp1-D heterozy- variants DNCO1, DNCO2, DNCO3, and DNCO4, which hadgotes and had the Rp1-D parent alleles at both flanking both flanking marker genotypes of the Rp1-D parent. Lane 7
is the Rp1-B parent and lane 8 is noncrossover variant BNCO1,marker loci. Gel blot analysis of the HRp1-DNCO1 haplo-which has the Rp1-B parent alleles at both flanking markers.type indicated it was probably derived from a crossoverDNA marker sizes, in kilobases, are shown on the left.
because it had fragments derived from both parents ineach of several digests tested. The parentally markedflanking markers indicated that there was a second DISCUSSION(double) crossover involved. In contrast, the restriction Characteristics of the HRp1-D haplotype: The HRp1-Dfragments of the HRp1-DNCO2 and HRp1-DNCO3 haplo- haplotype consists of Rp1-D and eight other homo-types were indistinguishable from the HRp1-D haplo- logues, seven of which have ORFs that could potentiallytype. These two mutants were further characterized by code for proteins similar to RP1-D, and one of whichcloning and sequencing the Rp1-D gene from these is truncated after the NBS encoding domain. All of thelines. Analysis of the coding region of the HRp1-DNCO2 22 susceptible variants analyzed from HRp1-D haplotypemutant allele identified only a single point mutation. homozygotes or 32 variants from HRp1-A/HRp1-D orThe mutation occurs in the 17th amino acid after the HRp1-B/HRp1-D heterozygotes had mutations or recom-putative translation start codon and results in a valine bination events that deleted or altered the Rp1-D gene.to alanine substitution (Figure 1). Sequencing of the In several of the variant HRp1-D haplotypes, includingmutant Rp1-D allele in HRp1-DNCO3 revealed a 33- two insertion mutants (Collins et al. 1999), only thenucleotide deletion in the region encoding the 10th Rp1-D gene was altered. Previous examination of theseLRR while the rest of the sequences are identical to the variants with a diverse collection of P. sorghi biotypesfunctional Rp1-D allele (Figure 1). Since in-frame indel indicated they were susceptible to all biotypes (Richterpolymorphisms are fairly common between the LRR et al. 1995). The Rp1-D gene is therefore the only generegions of rp1 paralogues, it is possible this change is in this haplotype with a detectable rust resistance pheno-not an actual deletion, but a gene conversion from an type. This is in contrast to several other clustered resis-allele from the HRp1-A haplotype. To find out if this tance gene families that have been examined. Loci, likepolymorphism was derived from the HRp1-A parent, a Cf4/9, Cf2/5, and Xa21 (Dixon et al. 1996; Parniske etpair of primers flanking the deletion was used in PCR al. 1997; Wang et al. 1998) have multiple genes in aamplification of HRp1-A genomic DNA. No amplified single haplotype that contribute to resistance. The intactfragments corresponding to this reduced size were ob- ORFs of most of the rp1 genes are also in contrast toserved, indicating that the gene is probably not present certain other resistance loci, like Xa21 (Song et al. 1997)in HRp1-A and that the 33-nucleotide deletion is proba- and Rpp5 (Noel et al. 1999), where many of the family
members have obvious mutations, such as insertions orbly not a consequence of gene conversion.

435Recombination at rp1
ORF-disrupting point mutations. The function of the 5000. It is possible that the overall rate of recombinationbetween Rp1-D and some of the other paralogues isrp1 paralogues is not clear; it is possible they could
provide resistance to rust biotypes that are obscure or similar to that observed with rp1-dp5, especially thosethat exhibit higher sequence homology. Since most ofno longer extant or even other nonrust pathogens.
No transcripts could be found for two of the HRp1-D the other genes were transcribed, some of these recom-bination events may have resulted in a functional Rp1-Dhaplotype genes with full-length ORFs. In addition, one
of the three HRp1-K haplotype genes characterized had specificity and would not have been selected. Intragenicrecombination events between paralogues in the Rp1a single stop codon located 111 bases after the putative
start codon in what would otherwise be a full-length complex may therefore be very common. This type ofrecombination will promote coevolution of the geneORF. Most of the susceptible recombinants from the
HRp1-D haplotype homozygotes resulted in various family as well as reshuffle them into new combinations.Recombination between paralogues at other clusteredamounts of the Rp1-D coding region being recombined
with a gene with an apparently nonfunctional promoter. resistance genes may be much less frequent. For example,genes in the Dm3 and Pto gene clusters appear to evolveFor example, the recombinant gene in the HRp1-D*19
haplotype had the LRR region from Rp1-D and differed more independently, since genes in orthologous posi-tions in different lines appear to be more similar thanfrom Rp1-D by only 13 amino acids, but no transcript
for the recombinant gene could be identified. The pres- paralogues (Michelmore and Meyers 1998). Sponta-neous susceptible variants have been characterized fromence of these unused, but largely intact, coding regions
in rp1 haplotypes raises the question of whether there other clustered resistance gene families. Recombinationevents have been examined at complex Cf loci of to-are some types of mechanisms that maintain them. Un-
equal recombination events probably play a role since mato. When five susceptible recombinants were isolatedfrom heterozygotes of the Cf4/9 locus, all five of thethey have a natural tendency to homogenize repeat units
(Smith 1976). Some of the genes may be useful after recombination breakpoints mapped to the intergenicregions (Parniske et al. 1997). However, a single suscep-they are activated by recombination events that donate
new 59 sequences, or useful as sequence donors in re- tible recombinant from a Cf2/Cf5 heterozygote wasfound to be intragenic (Dixon et al. 1998). It seemscombination events to make novel genes, but these se-
quences would not be expected to provide a large selec- that different complex R gene loci may vary in the extentto which paralogues recombine.tive advantage to the plant that carried them.
Recombination and generation of novel alleles: Previ- Analysis of some Arabidopsis Rpp5 and flax M mutantsrevealed unequal crossing over between duplications inous genetic studies have indicated that the rp1 complex
consists of sequence duplications that can mispair and the regions encoding the LRR sequences within a singlegene, resulting in changes in the number of duplica-recombine during meiosis, generating haplotypic varia-
tion (Hulbert 1997; Collins et al. 1999). It seemed tions (Anderson et al. 1997; Parker et al. 1997). OneRpp5 mutant recovered from a mutagenesis experimentlikely that at least some of these recombination events
occurred within the rp1 paralogues since they were occa- was found to carry a duplication corresponding to fourLRRs (154 amino acids) when compared to the parentalsionally associated with seedlings having novel pheno-
types (Richter et al. 1995; Hu et al. 1996). In this analy- Rpp5 gene. Similarly, susceptible variants at the flax Mlocus were identified with a deletion of one direct repeatsis, a collection of 22 variants from a HRp1-D haplotype
homozygote were examined. By completely or partially (corresponding to 45 amino acids) in the LRR domain(Anderson et al. 1997). Sequence comparison of differ-sequencing the recombinant rp1 gene from the individ-
ual variants, 13 of these were determined to be derived ent alleles at the tomato Cf2/Cf5 locus and flax L locus(Dixon et al. 1998; Ellis et al. 1999) revealed somefrom crossovers between paralogues. Another 7 also ap-
peared to be derived from very similar events on the alleles with variation in the number of LRRs, presumablyresulting from the unequal crossing over between thebasis of restriction fragment analysis. Only two of the
variants could not be explained by intragenic crossover internal duplications during the evolution of these genefamilies. No such unequal crossing-over events were ob-events. Seventeen of the 22 variants represented cross-
overs between the Rp1-D gene, at the distal end of the served in this study. Comparisons of all nine full-lengthrp1 genes from the HRp1-D haplotype and three fromarray, and the untranscribed rp1-dp5 gene in the middle
of the array. Since no clone carried more than one the HRp1-K haplotype provided no evidence of this typeof recombination.paralogue, the intergenic regions between the rp1 genes
appeared to be at least 15–20 kb (possibly much larger), Another feature of the recombination events at therp1 locus is that the recombination breakpoints are dis-indicating that the two paralogues are at least 80 kb
apart. The frequency of this class of susceptible recombi- tributed throughout the genes. Members of the riceXa21 gene family contain a conserved region shortlynants was z1/10,000. The reciprocal crossover, which
would result in a duplication and not a deletion, would after the putative translation start in which recombina-tion had most likely occurred during the evolution ofnot have been selected. The estimated rate of recombi-
nation between these two paralogues is therefore z1/ the Xa21 haplotype (Song et al. 1997). No obvious re-

436 Q. Sun et al.
combination hotspots were observed among the rp1 ing selection. Ayliffe et al. (2000) found a similar resultby comparing sequences of rp1 homologues from barley.genes. Studies of other maize loci, such as bronze and
waxy, also revealed that the recombination breakpoints This may be an indication that the carboxy terminus ofthe Rp1 family proteins is most directly involved in li-were well distributed across the genes (Dooner and
Martinez-Ferez 1997). However, alleles of these sim- gand binding. It is also clear, however, that LRR se-quences from domain B are functionally important. Theple loci usually exhibit levels of sequence identity higher
than those of different family members at resistance DNCO3 mutant allele is identical to the Rp1-D geneexcept for a 33-nucleotide deletion corresponding togene clusters like rp1 or Xa21. At other maize loci, like
al, bl, and rl, recombination breakpoints were more fre- 11 aa in the 10th LRR. Interestingly, this correspondsto a highly variable region of the gene.quently resolved in certain region of the genes (re-
viewed in Schnable et al. 1998). The factors controlling Most of the Rp1 variants selected are completely sus-ceptible to all the rust biotypes in our current collectionthe distribution of recombination breakpoints in plants
are not well characterized and are in need of additional and the majority of these were a consequence of anunequal exchange between Rp1-D and rp-dp5, a para-research.
Mutation, selection, recombination, and Rp1 gene logue for which there is no evidence of expression. Mostof these exchanges result in a gene with 39 sequencesfunction: Comparative analysis of the rp1 genes indicates
that mutation plays an important role in the evolution from Rp1-D and 59 sequences from rp1-dp5. Other un-equal exchanges may generate recombinant genes thatof the family. Compared with the functional Rp1-D al-
lele, all the other Rp1-D family members have numerous either code for nonfunctional proteins or for proteinsthat will not interact with any of the rust biotypes indifferences, with the 59 regions of the genes (including
the NBS domain) being more conserved than the 39 our current collection. Among the Rp1 variants ana-lyzed, only Rp1-D*5 retains race-specific resistance, con-regions (corresponding to the LRR). This is consistent
with the predicted functions of different domains of R ferring resistance to the same spectrum of rust isolatesas the parental Rp1-D, but at a reduced level. Character-genes. The NBS domain is thought to be the effector
domain involved in transducing a recognition signal ization of the recombinant allele from Rp1-D*5 revealedit had the LRR of the Rp1-D gene, having resulted fromand not the domain most likely to be directly involved
in ligand binding (Traut 1994; Baker et al. 1997). a crossover in the NBS domain. The lower level of resis-tance in Rp1-D*5 may be attributed to the differencesRecently, significant amino acid sequence similarity has
been found between the NBS domain of NBS-LRR resis- in the NBS region between the recombinant gene andthe wild-type Rp1-D or to promoter strength. Site-di-tance proteins and the nematode CED-4 and mamma-
lian Apaf-1 proteins, which are activators of apoptotic rected mutagenesis in the NBS region of the tobaccoN gene demonstrated that some mutations in the P loopproteases (Chinnaiyan et al. 1997; van der Biezen and
Jones 1998). The LRR domain is well documented as of the NBS can result in loss or partial loss of R genefunction (Baker et al. 1997). The identical race specific-a functional structure involved in protein-protein inter-
action (Kobe and Deisenhofer 1994, 1995). The LRR ity of the recombinant Rp1 gene and the Rp1-D geneand their identical LRRs are consistent with the ideaof plant disease resistance genes could participate in
the binding of pathogen-derived avr factors directly or that this domain is responsible for controlling race spec-ificity. However, the recombinant gene in the HRp1-indirectly (Ellis and Jones 1998) and thus variability
of this region will result in changes in recognition capa- D24 haplotype also has the LRR from the Rp1-D genebut shows no resistance to any of the rust isolates in ourbility. This hypothesis is supported by sequence compar-
isons of resistance gene alleles or gene family members, collection. The 59 sequences of the recombinant genewere donated by rp1-dp1, which is transcribed. These 59in which codons for predicted solvent-exposed residues
of the LRR regions show evidence for diversifying selec- sequences are probably either not functional in confer-ring a resistance reaction or not compatible with thetion (reviewed in Ellis et al. 2000). This pattern of
diversifying selection has been observed for several LRR domain of the Rp1-D gene. This latter phenome-non has been observed in domain swaps between allelesother types of genes involved in nonself recognition
(Hughes and Nei 1988; Endo et al. 1993). In some R of the L gene of flax (Ellis et al. 1999; Luck et al. 2000)and Mi gene family members in tomato (Hwang etgene families, the evidence for diversifying selection
may not be apparent through the whole LRR region. al. 2000). The phenotypes of these recombinant genesindicate that the 59 regions of NBS-LRR genes areFor example, in the lettuce RGC2 gene family, only
the putative solvent-exposed residues in the 39-encoded adapted to function with specific LRR regions in thegenes. Swapping different 59 regions with different LRRsLRR show good evidence of diversifying selection (Mey-
ers et al. 1998b). The Rp1-D gene product has 24 imper- can lead to loss of resistance, spontaneous necrosis(Hwang et al. 2000), or even unexpected race specificit-fect LRRs that can be divided into two parts (domains
B and D) that are separated by a region with no LRR ies (Luck et al. 2000).The recombinant gene in the HRp1-D*21 haplotypeconsensus (Collins et al. 1999). Only domain D, at the
39 end of the gene, shows strong evidence for diversify- was derived from a crossover in the LRR domain and

437Recombination at rp1
Dooner, H. K., and I. M. Martinez-Ferez, 1997 Recombinationlost its race-specific resistance. The recombinant LRRoccurs uniformly within the bronze gene, a meiotic recombination
may have lost the ability to interact with the fungal hotspot in the maize genome. Plant Cell 9: 1633–1646.Ellis, J., and D. Jones, 1998 Structure and function of proteinselicitor. It is also possible that the 59 sequences donated
controlling strain-specific pathogen resistance in plants. Curr.by the rp1-hd2 gene are not functional or will not func-Opin. Plant Biol. 1: 288–293.
tion with Rp1-D LRR sequences. The HRp1-D*21 haplo- Ellis, J. G., G. J. Lawrence, E. J. Finnegan, P. Anderson, N. Collinset al., 1997 Advances in the molecular genetic analysis of thetype confers an unusual nonspecific reaction to rust andflax-flax rust interaction. Annu. Rev. Phytopathol. 35: 271–291.a necrotic-spotting phenotype (Hu et al. 1996) indicat-
Ellis, J. G., G. J. Lawrence, J. E. Luck and P. N. Dodds, 1999 Identi-ing it can induce hypersensitive reactions, but we have fication of regions in alleles of the flax rust resistance gene L
that determine differences in gene-for-gene specificity. Plant Cellnot yet demonstrated that necrosis is caused by the11: 495–506.recombinant gene and not by some other aspect of the
Ellis, J. G., P. N. Dodds and T. Pryor, 2000 The generation ofnovel haplotype. plant disease resistance gene specificities. Trends Plant Sci. 9:
373–379.The analysis of the maize rp1 locus has shown thatEndo, T., K. Ikeo and T. Gojobori, 1993 Large scale search forboth recombination and mutation are active forces in
genes on which positive selection may operate. Mol. Biol. Evol.the evolution of this resistance gene family. Analysis of 13: 685–690.
Grant, M. R., L. Godiard, E. Straube, T. Ashfield, J. Lewald etspontaneous mutants (selection of susceptibles) demon-al., 1995 Structure of the Arabidopsis RPM1 gene enabling dualstrated that unequal crossing over occurs frequently atspecificity disease resistance. Science 269: 843–846.
the locus and that the crossovers are frequently intra- Hu, G., and S. H. Hulbert, 1994 Evidence for the involvement ofgene conversion in meiotic instability of the Rp1 rust resistancegenic. This was also indicated by the stretches of perfectgenes of maize. Genome 37: 742–746.sequence identity observed between paralogues in a sin-
Hu, G., T. Richter, S. Hulbert and A. Pryor, 1996 Disease lesiongle haplotype. The sequence analysis also indicates that mimicry caused by mutations at the rust resistance gene Rp1.
Plant Cell 8: 1367–1376.selection pressure favors diversifying selection in theHughes, A. L., and M. Nei, 1988 Pattern of nucleotide substitutioncarboxy-terminal half of the LRR of the protein. Recom-
at major histocompatibility complex class I loci reveals overdomi-bination and mutation therefore appear to be intimately nant selection. Nature 335: 167–170.
Hulbert, S. H., 1997 Structure and evolution of the Rp1 complexresponsible for generating new resistance specificitiesconferring rust resistance in maize. Annu. Rev. Phytopathol. 35:at the maize rp1 locus.293–310.
Hulbert, S. H., and R. W. Michelmore, 1985 Linkage analysis ofThis work was supported in part by grants MCB-9728490 andgene for resistance to downy mildew (Bremia lactucae) in lettuce9975971 from the National Science Foundation. This article is contri-(Lactuca sativa). Theor. Appl. Genet. 70: 520–528.bution 01-159-J from the Department of Plant Pathology, Kansas Ag-
Hulbert, S. H., G. Hu and J. A. Drake, 1997 Kansas rust-resistantricultural Experiment Station.sweet corn populations A and B. Hortscience 32: 1130–1131.
Hwang, C.-F., A. V. Bhakta, G. M. Truesdell, W. M. Pudlo andV. M. Williamson, 2000 Evidence for a role of the N terminusand leucine-rich repeat region of the Mi gene product in regula-LITERATURE CITEDtion of localized cell death. Plant Cell 12: 1319–1329.
Islam, M. R., K. W. Shepherd and G. M. E. Mayo, 1989 Recombina-Anderson, P. A., G. J. Lawrence, B. C. Morrish, M. A. Ayliffe, E. J.Finnegan et al., 1997 Inactivation of the flax rust resistance tion among genes at the L group in flax conferring resistance
to rust. Theor. Appl. Genet. 77: 540–546.gene M associated with loss of a repeated unit within the leucine-rich repeat coding region. Plant Cell 9: 641–651. Jahoor, A., A. Jacobi, M. E. Schuller and G. Fischbeck, 1993 Ge-
netical and RFLP studies at the Mla locus conferring powderyAyliffe, M. A., N. C. Collins, J. G. Ellis and A. Pryor, 2000 Themaize rp1 rust resistance gene identifies homologues in barley mildew resistance in barley. Theor. Appl. Genet. 85: 713–718.
Jones, D. A., and J. D. G. Jones, 1996 The role of leucine-rich repeatthat have been subjected to diversifying selection. Theor. Appl.Genet. 100: 1144–1154. proteins in plant defenses. Adv. Bot. Res. 24: 89–167.
Kobe, B., and J. Deisenhofer, 1994 The leucine-rich repeat: a versa-Baker, B., P. Zambryski, B. Staskawicz and S. P. Dinesh-Kumar,1997 Signaling in plant-microbe interactions. Science 276: 726– tile binding motif. Trends Biochem. 19: 415–421.
Kobe, B., and J. Deisenhofer, 1995 A structural basis of the interac-733.Botella, M. A., J. E. Parker, L. N. Frost, P. D. Bittner-Eddy, tions between leucine-rich repeats and protein ligands. Nature
374: 183–186.J. L. Beynon et al., 1998 Three genes of the Arabidopsis RPP1complex resistance locus recognize distinct Peronospora parasitica Lawrence, G. J., E. J. Finnegan, M. A. Ayliffe and J. G. Ellis,
1995 The L6 gene for flax rust resistance is related to the Arabi-avirulence determinants. Plant Cell 10: 1847–1860.Burdon, J. J., and J. Silk, 1997 Sources and patterns of diversity in dopsis bacterial resistance gene RPS2 and the tobacco viral resis-
tance gene N. Plant Cell 7: 1195–1206.plant pathogenic fungi. Phytopathology 87: 664–669.Chinnaiyan, A. M., D. Chaudhary, K. O’Rourke, E. V. Koonin and Luck, J. E., G. L. Lawrence, P. N. Dodds, K. W. Shepherd and J. G.
Ellis, 2000 Regions outside of the leucine-rich repeats of flaxV. M. Dixit, 1997 Role of CED-4 in the activation of CED-3.Nature 338: 728–729. rust resistance proteins play a role in specificity determination.
Plant Cell 12: 1367–1377.Collins, N., J. Drake, M. Ayliffe, Q. Sun, J. Ellis et al., 1999 Molec-ular characterization of the maize Rp1-D rust resistance haplotype Martin, G. B., S. H. Brommonschenkel, J. Chungwongse, A. Frary,
M. W. Ganal et al., 1993 Map-based cloning of a protein kinaseand its mutants. Plant Cell 11: 1365–1376.Crute, I. R., and D. A. C. Pink, 1996 Genetics and utilization of gene conferring disease resistance in tomato. Science 262: 1432–
1436.pathogen resistance in plants. Plant Cell 8: 1747–1755.Dixon, M. S., D. A. Jones, J. S. Keddie, C. M. Thomas, K. Harrison McDowell, J. M., M. Dhandaydham, T. A. Long, M. G. M. Aarts,
S. Goff et al., 1998 Intragenic recombination and diversifyinget al., 1996 The tomato Cf-2 disease resistance locus comprisestwo functional genes encoding leucine-rich repeat proteins. Cell selection contribute to the evolution of downy mildew resistance
at the RPP8 locus of Arabidopsis. Plant Cell 10: 1861–1874.84: 451–459.Dixon, M. S., K. Hatzixanthis, D. A. Jones, K. Harrison and J. D. Meyers, B. C., D. B. Chin, K. A. Shen, S. Sivaramakrishnan, D. O.
Lavelle et al., 1998a The major resistance gene cluster in let-Jones, 1998 The tomato Cf-5 disease resistance gene and sixhomologs show pronounced allelic variation in leucine rich re- tuce is highly duplicated and spans several megabases. Plant Cell
10: 1817–1832.peat copy number. Plant Cell 10: 1915–1925.

438 Q. Sun et al.
Meyers, B. C., K. A. Shen, P. Rohani, B. S. Gaut and R. W. Michel- Shepherd, K. W., and G. M. E. Mayo, 1972 Genes conferring specificplant disease resistance. Science 175: 375–380.more, 1998b Receptor-like genes in the major resistance locus
of lettuce are subject to divergent selection. Plant Cell 11: 1833– Simons, G., J. Groenendijk, J. Wijbrandi, M. Reijans, J. Groenenet al., 1998 Dissection of the Fusarium I2 gene cluster in tomato1846.
Michelmore, R. W., and B. C. Meyers, 1998 Clusters of resistance reveals six homologs and one active gene copy. Plant Cell 10:1055–1068.genes in plants evolve by divergent selection and a birth-and-
death process. Genome Res. 8: 1113–1130. Smith, G. P., 1976 Evolution of repeated DNA sequences by unequalcrossover. Science 191: 528–535.Mindrinos, M., F. Katagiri, G. L. Yu and F. M. Ausubel, 1994 The
Song, W. Y., L. Y. Pi, G. L. Wang, J. Gardner, T. Holsten et al.,A. thaliana disease resistance gene RPS2 encodes a protein con-1997 Evolution of the rice Xa21 disease resistance gene family.taining a nucleotide-binding site and leucine-rich repeats. CellPlant Cell 9: 1279–1287.78: 1089–1099.
Sudupak, M. A., J. L. Bennetzen and S. H. Hulbert, 1993 UnequalNoel, L., T. L. Moores, E. A. van der Biezen, M. Parniske, M. J.exchange and meiotic instability of Rp1 region disease resistanceDaniels et al., 1999 Pronounced intraspecific haplotype diver-genes. Genetics 133: 119–125.gence at the RPP5 complex disease resistance locus of Arabi-
Thomas, C. M., D. A. Jones, M. Parniske, K. Harrison, P. J. Balint-dopsis. Plant Cell 11: 2099–2112.Kurti et al., 1997 Characterization of the tomato Cf-4 geneParker, J. E., M. J. Coleman, V. Szabo, L. N. Frost, R. Schmidt etfor resistance to Cladosporium fulvum identifies sequences thatal., 1997 The Arabidopsis downy mildew resistance gene RPP5determine recognitional specificity in Cf-4 and Cf-9. Plant Cell 9:shares similarity to the toll and interleukin-1 receptors with N2209–2224.and L6. Plant Cell 9: 879–894.
Traut, T. W., 1994 The functions and consensus motifs of nineParniske, M., K. E. Hammond-Kosack, C. Golstein, C. M. Thomas,types of peptide segments that form different types of nucleotide-D. A. Jones et al., 1997 Novel disease resistance specificitiesbinding sites. Eur. J. Biochem. 222: 9–19.result from sequence exchange between tandemly repeated genes
van der Biezen, E. A., and J. D. Jones, 1998 The NB-ARC domain:at the Cf-4/9 locus of tomato. Cell 91: 821–832.a novel signaling motif shared by plant resistance gene productsRichter, T., T. Pryor, J. Bennetzen and S. Hulbert, 1995 Newand regulators of cell death in animals. Curr. Biol. 8: 226–227.rust resistance specificities associated with recombination in the
Wang, G. L., D. L. Ruan, W. Y. Song, S. Sideris, L. Chen et al.,Rp1 complex in maize. Genetics 141: 373–381.1998 Xa21D encodes a receptor-like molecule with a leucine-Salmeron, J. M., G. E. D. Oldroyd, C. M. T. Tommens, S. R. Scofield,rich repeat domain that determines race-specific recognition andH. Kim et al., 1996 Tomato Prf is a member of the leucine-rich is subject to adaptive evolution. Plant Cell 10: 765–780.repeat class of plant disease resistance genes and lies embedded Wei, F., K. Gobelman-Werner, S. M. Morroll, J. Kurth, L. Maowithin the Pto kinase gene cluster. Cell 86: 123–133. et al., 1999 The Mla (powdery mildew) resistance cluster is asso-
Sambrook, J., E. F. Fritsch and T. Maniatis, 1989 Molecular Clon- ciated with three NBS-LRR gene families and suppressed recombi-ing: A Laboratory Manual. Cold Spring Harbor Laboratory Press, nation within a 240-kb DNA interval on chromosome 5s (1HS)Cold Spring Harbor, NY. of barley. Genetics 153: 192.
Saxena, K. M. S., and A. L. Hooker, 1968 On the structure of a Whitham, S., S. P. Dinesh-Kumar, D. Choi, R. Hehl, C. Corr et al.,gene for disease resistance in maize. Proc. Natl. Acad. Sci. USA 1994 The product of the tobacco mosaic virus resistance gene61: 1300. N: similarity to toll and the interleukin-1 receptor. Cell 78: 1101–
Schnable, P. S., A.-N. Hsia and B. J. Nikolau, 1998 Genetic recom- 1115.bination in plants. Curr. Opin. Plant Biol. 1: 123–129. Zhang, Z., S. Khumar and M. Nei, 1997 Small-sample tests of epi-
Schwarz-Sommer, Z., L. Leclercq, X. Goebel and H. Saedler, sodic adaptive evolution: a case study of primate lysozymes. Mol.1987 Cin4, an insert altering the structure of the A1 gene of Biol. Evol. 14: 1335–1338.zea mays, exhibits properties of nonviral retrotransposons. EMBOJ. 6: 3873–3880. Communicating editor: J. A. Birchler





























