Embed Size (px)
Citation preview

Recent developments in drug resistance mechanism inchronic myeloid leukemia: a reviewShantashri Vaidya, Kanjaksha Ghosh, Babu Rao Vundinti
National Institute of Immunohaematology (ICMR), Parel, Mumbai, India
Chronic myeloid leukemia (CML) is a clonal hematopoi-
etic stem cell disorder. It is characterized by excess pro-
liferation of myeloid progenitor that retains the capacity
for differentiation during the stable or chronic phase of
the disease. Patients with CML usually present in the
chronic phase of the disease during which there is a grad-
ual expansion of mature myeloid cells in the bone mar-
row and peripheral blood. Without treatment, patients
inevitably progress through an accelerated phase of dis-
ease to a terminal acute phase known as blast crisis,
characterized by a massive increase in undifferentiated
blast that can be either myeloid or lymphoid in nature.
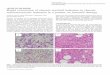
Hallmark of this disease is a unique chromosome, known
as the Philadelphia chromosome (Ph), which results from
the reciprocal translocation between the long arms of
chromosomes 9 and 22 [t(9;22) (q34;q11)] (Figs 1 and 2
)and leads to the fusion of the breakpoints cluster (BCR)
and human ABL1 genes (Fig. 3). The root cause of this
disease is still unknown. The majority of patients with
CML have no history of excessive exposure to ionizing
radiations or carcinogens, but the incidence does increase
progressively with exposure to increasing doses of radia-
tion. This has been noted following chronic exposure in
radiologists who practiced without adequate shielding, in
patients who received radiation treatment, and in sub-
jects exposed to a single massive dose of radiation, as in
the atomic bomb explosions in Japan in 1945 (1). CML
is not hereditary. The incidence of CML in United States
and most Western countries is about 1.5 per 100 000
population per year and accounts for about 15% of all
cases of leukemia; about 4500 new cases are diagnosed in
the United States each year. CML is slightly more fre-
quent in men than in women (incidence ratio: 1.4 to
2.2 : 1), but the course of the disease is the same. The
median age is about 50; it is rare in children, and the
incidence increases progressively with age.
Molecular biology and pathophysiology ofchronic myeloid leukemia
Chronic myeloid leukemia is probably the most exten-
sively studied human malignancy. The discovery of Phil-
adelphia chromosome (Ph) in 1960 as the first consistent
chromosomal abnormality associated with a specific type
of leukemia was a breakthrough in cancer biology. It
took 13 yrs before it was appreciated that the Ph chro-
mosome is the result of a t(9;22) reciprocal chromosomal
translocation and another 10 yrs before the translocation
Abstract
A revolution in medical science was marked with the advent of imatinib, a site-specific drug for the man-
agement of patients with chronic myeloid leukemia (CML). Imatinib mesylate (also known as Glivec, Glee-
vec, STI-571, CGP57148), an orally administered 2-phenylaminopyrimidine derivative approved by FDA in
2001 for the treatment for CML, is highly effective in treating the early stages of CML, but remission
induced in advanced phase was observed to be relatively short-lived. The primary cause of resistance in
patients with CML is the mutations in the BCR–ABL kinase domain. This review discusses the different
mechanisms leading to imatinib resistance and various treatment options to over-ride imatinib resistance.
Key words chronic myeloid leukemia; BCR–ABL fusion; drug resistance; imatinib; chronic myeloid leukemia therapy
Correspondence Babu Rao Vundinti, Department of Cytogenetics, National Institute of Immunohaematology (ICMR), 13th Floor,
New Multistoried Building, K.E.M Hospital campus, Parel, Mumbai-400012, India. Tel: +91 22 24138518, 24138519; Fax: +91 22
24138518; e-mail: [email protected]
Accepted for publication 30 July 2011 doi:10.1111/j.1600-0609.2011.01689.x
REVIEW ARTICLE
European Journal of Haematology 87 (381–393)
ª 2011 John Wiley & Sons A/S 381
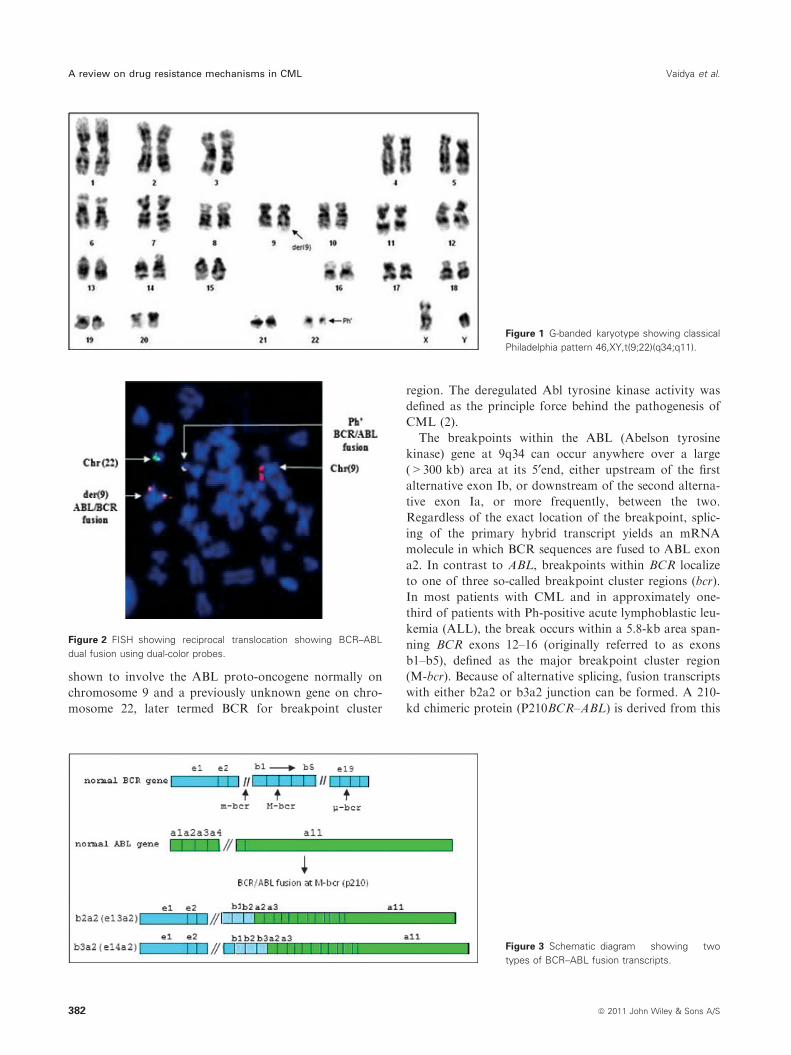
shown to involve the ABL proto-oncogene normally on
chromosome 9 and a previously unknown gene on chro-
mosome 22, later termed BCR for breakpoint cluster
region. The deregulated Abl tyrosine kinase activity was
defined as the principle force behind the pathogenesis of
CML (2).
The breakpoints within the ABL (Abelson tyrosine
kinase) gene at 9q34 can occur anywhere over a large
(>300 kb) area at its 5¢end, either upstream of the first
alternative exon Ib, or downstream of the second alterna-
tive exon Ia, or more frequently, between the two.
Regardless of the exact location of the breakpoint, splic-
ing of the primary hybrid transcript yields an mRNA
molecule in which BCR sequences are fused to ABL exon
a2. In contrast to ABL, breakpoints within BCR localize
to one of three so-called breakpoint cluster regions (bcr).
In most patients with CML and in approximately one-
third of patients with Ph-positive acute lymphoblastic leu-
kemia (ALL), the break occurs within a 5.8-kb area span-
ning BCR exons 12–16 (originally referred to as exons
b1–b5), defined as the major breakpoint cluster region
(M-bcr). Because of alternative splicing, fusion transcripts
with either b2a2 or b3a2 junction can be formed. A 210-
kd chimeric protein (P210BCR–ABL) is derived from this
Figure 1 G-banded karyotype showing classical
Philadelphia pattern 46,XY,t(9;22)(q34;q11).
Figure 3 Schematic diagram showing two
types of BCR–ABL fusion transcripts.
Figure 2 FISH showing reciprocal translocation showing BCR–ABL
dual fusion using dual-color probes.
A review on drug resistance mechanisms in CML Vaidya et al.
382 ª 2011 John Wiley & Sons A/S

mRNA. In the remaining patients with ALL and rarely in
patients with CML, characterized clinically by prominent
monocytosis, the breakpoints are further upstream in the
54.4-kb region between the alternative BCR exons e2¢ ande2, termed the minor breakpoint cluster region (m-bcr).
The resultant e1a2 mRNA is translated into a 190-kd pro-
tein (P190BCR–ABL). Recently, a third breakpoint clus-
ter region (l-bcr) was identified downstream of exon 19,
giving rise to a 230-kd fusion protein (P230BCR–ABL)
associated with the rare Ph-positive chronic neutrophilic
leukemia (2).
BCR–ABL plays a central role in the pathogenesis of
CML, which was unraveled by two independent studies
(3, 4). The activity of this fusion protein is no longer
under the regulatory control mechanisms for ABL and
thus induces malignant disease by activating multiple
cytoplasmic and nuclear signal transduction pathways
that influence the growth and survival of haematopoietic
cells. The targets for Bcr–abl include members of the
Ras, phosphotidylinositol-3 kinase (PI3K) ⁄Akt, and
Jak ⁄Stat signaling pathways, which regulate cell prolifer-
ation and apoptosis. Bcr–abl abrogates cell dependence
on external growth factors by upregulating interleukin-3
production and alters the cell adhesion properties by
modulating expression and activation of focal adhesion
kinase and associated proteins. The kinase also has
diverse effects on the DNA repair response, which may
promote additional chromosomal alterations and muta-
tions involved in the progressive nature of late-stage
CML (5).
Additional insights into the pathogenesis of CML
came from the study of transforming retroviruses, specifi-
cally the Abelson murine leukemia virus. The transform-
ing protein of this virus, v-ABL, was shown to be a
tyrosine kinase with its transforming ability dependent
on this activity. Subsequently, the Bcr–abl fusion product
proved to display similar tyrosine kinase activity. The
constitutive tyrosine kinase activity of Bcr–abl causes
activation of a variety of intracellular signaling path-
ways, leading to alterations in the proliferative, adhesive,
and survival properties of CML cells, with all of these
events dependent on the tyrosine kinase activity of the
Bcr–abl protein. Transduction of p210 bcr–abl into mur-
ine hematopoietic stem cells, followed by transplantation
into syngeneic mice, causes a CML-like syndrome; mice
transgenic for p190 bcr–abl develop acute leukemia.
These findings provide convincing evidence that Bcr–abl
is a leukemic oncogene (6).
The disease characteristically develops in three phases.
Left untreated, the initial chronic phase lasts approxi-
mately 3–6 yrs; the disease then progresses, often
through an accelerated phase, to a terminal blastic phase.
Most patients are diagnosed in chronic phase, which is
characterized by an increased number of leukocytes
and ⁄or platelets and a bone marrow blast count <10%.
The accelerated phase may be marked by one or more of
the following: increasing splenomegaly and leukocytosis,
an increase in basophils to 20% or greater, thrombocyto-
penia, and clonal evolution. In the blastic phase, for
which the median survival is 2–4 months, 30% or more
of blood and bone marrow cells are blasts, and myeloid
precursors may also form tumors in the lymph nodes,
skin, and bone. (5).
Since the description of BCR–ABL oncogene, a myr-
iad of treatment options have been explored in CML,
including arsenic trioxide, spleenic irradiation, busul-
phan, hydroxycarbamide, stem cell transplantation, and
interferon a. Among these, interferon a showed superior
activity compared with other chemotherapeutic agents. It
can promote long-term survival, but the proportion of
such patients is small. Stem cell transplantation can cure
the disease; however, as the average age of onset is
>50 yrs of age, this factor, combined with the inability
to identify suitably matched donors in all cases, limits
this option to a minority of patients (6).
Imatinib therapy in patients with CML
Imatinib was developed in the late 1990s by biochemist
Nicholas Lydon, a former researcher for Ciba Geigy
(now Novartis, Basel, Switerland), oncologist Brian
Druker of Oregon Health and Science University, and
Charles Sawyers of Memorial Sloan-Kettering Cancer
Centre, who led the clinical trials confirming its efficacy
in CML.
Imatinib was developed by rational drug design. After
the Philadelphia chromosome mutations and defective
bcr–abl protein were discovered, the investigators
screened chemical libraries to find a drug that would
inhibit that protein. With high-throughput screening,
they identified 2-phenylaminopyrimidine, which showed
low potency and poor specificity. This lead compound
was then tested and modified by the introduction of
methyl and benzamide groups to give it enhanced bind-
ing properties, resulting in imatinib (6).
Structural analyses have revealed that imatinib is sand-
wich between the N-lobe and C-lobe of the Abl kinase
domain and penetrates through the central region of the
kinase. The pyrimidine and pyridine rings of imatinib
occupy the region where an adenine ring of ATP nor-
mally binds. The rest of the molecule wedges itself into a
hydrophobic cage between the activation loop and the
C-helix of the N-lobe, specifically recognizing the inactive
and unphosphorylated conformation of Abl. Much of
the remarkable efficacy of imatinib appears to derive
from its ability to stabilize the inactive conformation
through an ‘induced fit’ mechanism. The potency of
imatinib against the activated form of bcr–abl
Vaidya et al. A review on drug resistance mechanisms in CML
ª 2011 John Wiley & Sons A/S 383

presumably arises from the dynamic nature of kinase
molecules, which transiently switch between inactive and
active forms, allowing imatinib to trap the kinase in its
inactive conformation (7).
Imatinib as a single agent has shown impressive activity
in the chronic phase of CML, with >95% complete hema-
tologic response and >73% complete cytogenetic remis-
sion in patients (8). However, patients treated in the later
stages of the disease do not respond as well, with only
about 65% displaying hematologic responses (9).
Resistance to imatinib
The emergence of resistance to imatinib has dampened the
enthusiasm for this drug. The rate of relapse and resis-
tance appear to correlate with disease stage, and the inci-
dence increases as CML progresses. Resistance to imatinib
can be divided into primary resistance, in which patients
show lack of efficacy to this tyrosine kinase inhibitor
(TKI) from the start of the therapy, and secondary resis-
tance, also known as acquired resistance, which is defined
as a loss of hematologic, cytogenetic, or molecular
response, as well as progression to advanced phases of
CML. Resistance can further be divided into hematologic
(lack of normalization of peripheral blood counts), cyto-
genetic (persistence of Ph chromosome), and molecular
[persistence of BCR–ABL1 transcripts by reverse trans-
criptase polymerase chain reaction (RT-PCR)].
Patients can be classified according to their response
to treatment, which can be considered optimal, subopti-
mal, or failure. Failure indicates that primary resistance
patients in this category should be recommended a
change in therapy for second-generation TKI. Patients
with suboptimal response need close monitoring, and a
dose escalation from 400 to 800 mg is justified (10).
However, resistance to imatinib can be overcome with
various strategies at different levels (Fig. 4).
Mechanisms of imatinib resistance
Imatinib has become the standard first-line therapy for
CML. Indeed, imatinib induces complete remission in
virtually all patients with the indolent chronic phase of
CML who are treated immediately upon diagnosis (9).
When treated during the more aggressive stage of blast
crisis, most patients ultimately evolve drug resistance dis-
ease. Various mechanisms were postulated for imatinib
resistance, which can be broadly divided into BCR–
ABL-dependent and BCR–ABL-independent mechanisms
of imatinib resistance.
BCR–ABL-independent mechanisms of resistance
Quiescent CML stem cell: Quiescent CML stem cells
account for approximately 0.5% of the CD34+ popula-
tion and are characterized by intrinsic resistance to
Figure 4 Strategies to overcome imatinib resis-
tance at various levels.
A review on drug resistance mechanisms in CML Vaidya et al.
384 ª 2011 John Wiley & Sons A/S

imatinib. Jorgensen and Holyoake (11) noticed that
although CD34+38- HSCs carried a single copy of
BCR–ABL, they still exhibited a significant (10-fold)
increase in Bcr–abl kinase level compared with total
mature mononuclear cells. This study was in consensus
with the study carried out by Copland et al. (12).
While rationally designed TKIs have proven effective
in disease management, they do not offer a cure owing
to the persistence of insensitive HSCs. One possible
explanation for this molecular persistence is oncogene
overexpression at the message, protein and kinase activ-
ity levels that is not overcome by possible subtherapeutic
levels of TKI within HSCs (11). Nevertheless, introduc-
tion of novel agents such as the putative FTI BMS-
214662 will make significant advances towards the goal
of elimination of the diseased stem cell population, pref-
erentially targeting quiescent leukemic cells over normal.
Further elucidation of its true mechanism of action will
potentially inform drug development for use in other leu-
kemias and stem cell–derived cancers.
Pharmacokinetics: Pharmacokinetic data indicates that
patients receiving 400 mg imatinib per day have plasma
concentrations constantly higher than 1 lmol ⁄L (13). In
patients receiving higher doses (600–800 mg ⁄d), increasedplasma concentrations have been observed, with some
patients having concentrations higher than 20 lmol ⁄L. As
imatinib is metabolized largely by the cytochrome p450
isoenzymes P3A4 (CYP3A4) and P3A5 (CYP3A5), differ-
ences in the concentrations of CYP3A4 ⁄A5 or drugs that
can inhibit or induce said enzymes have the potential to
greatly affect the levels of imatinib in plasma (10).
It has been proposed that binding of a-1-acid glyco-
protein (AGP), an acute phase plasma protein, can inhi-
bit the activity of imatinib (10, 14). Therefore, imatinib
present in the plasma could be mostly bound to AGP
and thus biologically inactive. To test this hypothesis,
blood samples from two patients who were resistant to
imatinib were incubated with erythromycin (which
competes with imatinib for binding of AGP) for 1 h
before separation of mononuclear cells and lysis. The
activity of imatinib on Bcr–abl autophosphorylation was
restored, indicating that most plasma imatinib is bound
to AGP so not active (13).
Furthermore, when patients who were being treated
with imatinib received a simultaneous infusion of clinda-
mycin, another molecule known to bind AGP, large
decrease in total plasma concentrations of imatinib was
observed in minutes. This finding also shows that most
imatinib present in plasma is not in equilibrium with
tissues and that there is a gradient between plasma and
tissues (13).
Although the exact concentration of imatinib inside
cells – the final target of the drug – is not known, these
findings show that plasma concentrations of the drug are
not a reliable indicator of the concentration present
inside leukemic cells.
Drug efflux mechanism: Several other cellular mecha-
nisms of resistance to imatinib have also been identified,
but the possible importance of drug-transporter proteins
that affect drug concentration, such as permeability glyco-
protein (Pgp), human organic cationic transporter
(hOCT), and ATP-binding cassette G2, has been only
recently appreciated. The generally accepted action of
MDR1 (or adenosine triphosphate–binding cassette trans-
porter ABCB1) is to reduce intracellular drug accumula-
tion through Pgp-mediated efflux, thus hampering the
achievement of effective drug levels at the target site.
Galimberti et al. (15) and Mahon et al. (16) separately
showed that those patients who failed to attain a major
cytogenetic response or progressed exhibited MRD1 gene
overexpression. In contrast to this study, Ferrao et al. (17)
have shown that overexpression of MDR1 in K562 cells
does not confer resistance to imatinib in vitro. Association
between single-nucleotide polymorphisms in MDR1 gene
and response to imatinib has also been observed. Dulucq
et al. (18) have demonstrated the usefulness of three sin-
gle-nucleotide polymorphisms (1236C>T, 2677G>T ⁄A,
3435C>T) in the identification of CML in patients who
may or may not respond optimally to imatinib.
Inhibition of imatinib influx through the hOCT1 has
also been proposed as an important factor regulating
intracellular imatinib availability. The above-mentioned
transporters regulate in concert the active transport of
imatinib in and out of the cell and potentially play
an important role in pharmacogenetics of imatinib. Modi-
fication of imatinib regimen by increasing the standard
dose from 400 to 600–1000 mg ⁄d is suggested in some
patients showing an altered expression of transporter pro-
teins.
Cellular signaling: It is hypothesized that pathways
affected by Bcr–abl, which also contribute to the malig-
nant transformation, are involved in drug resistance
(Fig. 4). Yamada reported involvement of STAT 5, as a
critical transcription factor, confers imatinib resistance
on leukemic cells through the transcription of TERT and
MDR1 (19).
BCR–ABL-dependent mechanisms of resistance
Mutations within the ABL kinase domain (point mutations
and frame shift mutations): The mutations in the ABL
kinase domain and other domains regulating the confor-
mation of ABL kinase domain result in imatinib resis-
tance.
To date, mutations have been discovered that sterically
hinder drug occupancy of the active site, alter the
deformability of the highly conserved phosphate-binding
Vaidya et al. A review on drug resistance mechanisms in CML
ª 2011 John Wiley & Sons A/S 385

P-loop, and influence the conformation of the activation
loop surrounding the active site (20).
Studies carried out all over the world for screening of
mutations in BCR–ABL kinase domain have reported
many novel mutations with different frequencies occur-
ring in the population. The first mutation linked to
imatinib resistance in a cohort of relapsed patients,
in American population, discovered by Gorre et al. (21),
was T315I. Soon after the report of T315I, several other
mutations were identified in closely juxtaposed residues.
Studies carried out on German population, by von Bub-
noff et al., revealed five distinct mutations in the BCR–
ABL domain (sample size 7). All five mutations resulted
in exchange of amino acids within the ATP-binding site
(in six patients) or in the activation loop (one patient) of
Bcr–abl kinase domain. Their study strongly suggests
that a patient could be resistant to STI-571 by acquisi-
tion of different individual point mutations within the
ATP-binding pocket or activation loop of Bcr–abl (22).
Timothy Hughes, Australia, reported that among few
mutations in BCR–ABL kinase domain, there are 9 that
account for more than 85% of all mutations: M244Vg,
G250E, Y253F ⁄H, E255K ⁄V, T315I, M351T, and
F359V. The greatest degree of resistance has been associ-
ated with the T315I mutation and point mutations in the
P-loop domain, i.e., G250E, Q252H, Y253F, and
E255K ⁄V (23).
To date, 17 distinct point mutations in the ABL kinase
domain conferring resistance to imatinib have been iden-
tified (24), which mainly cluster into four main groups
(13): imatinib-binding site, nucleotide-binding site for
ATP, activation loop, and hydrophobic patch between
helices E,F, and I in the terminal lobe of the enzyme.
Despite the differences in location, several of these
mutations decrease the binding affinity of Bcr–abl for
imatinib (25) although with different indices of resistance.
Mechanisms by which mutations in the imatinib-bind-
ing site lead to reduced binding affinity are as follows:
T315: Thr315 also known as gatekeeper residue located
at the periphery of the imatinib-binding site of Abl.
Mutations of threonine 315 to isoleucine result in disrup-
tion of H-bond (10). The absence of oxygen atom nor-
mally provided by the side chain of Thr315 precludes
formation of a hydrogen bond with the secondary amino
group of STI-571. In addition, isoleucine contains an
extra hydrocarbon group in the side chain, which results
in steric clash with STI-571 and presumably inhibits
binding (25). F317L and V289A mutations cause a loss
of van der Waal’s contacts with the pyridine and piper-
azyl moiety of the inhibitor, respectively. These amino
acids are located at the two extremes of the binding site
and shield it from water; thus, the reduction in the size
of the amino acid side chain increases the accessibility of
water to the binding site, and the strength of the interac-
tion within the binding site is reduced because of the
increased competition by water (10).
In the nucleotide-binding loop, the mutation Y253H is
predominantly found. Tyrosine 253 forms a hydrogen
bond with asparagine 322, which holds in place a folded
loop between two b-strands, increasing the surface com-
plementarity with STI-571 (22). Also, a small reduction
in hydrophobic interactions is observed, because the his-
tidine side chain is smaller than the tyrosine side chain.
The loss of a hydrogen bond is probably the major con-
tributing factor to resistance, because it causes a change
in the equilibrium between the active and inactive con-
formation, forcing the balance toward the active confor-
mation (13). Mutation at G250E reduces the space
available at the ‘entrance’ of the imatinib-binding pocket;
thus, the access to the imatinb-binding pocket is hin-
dered. Mutations at Q252 and E255K probably change
the protein dynamic towards activation by modifying the
intrinsic flexibility of the nucleotide-binding loop; thus,
the ability of the inhibitor to fit in the binding pocket is
hindered (13).
The third group of mutations is located in the activa-
tion loop and close to the hinge region of the flexible
sequence. So far, a single mutation at position 396
(H396P) has been described in this region. This region of
the kinase does not interact with STI-571, except for the
anchor region, which is located at the amino-terminal of
histidine 396. However, the activation loop has been
shown to be in closed (inactive) conformation to allow
STI-571 binding to take place; stabilization of the open
(active) conformation by phosphorylation of tyrosine 393
results in diminished susceptibility of ABL to inhibition
by STI-571. Thus, H396P mutations inhibits binding of
STI-571 by stabilization of the activation loop in an
open conformation or by destabilization of the closed
conformation, both of which can result in an enzyme
that remains in the active form for longer (22).
Introduction of polar residues into the hydrophobic
patch, as in the mutations M351T and F486S, decreases
the strength of the hydrophobic interactions and thus
decreases the stability of the structure, shifting the equi-
librium in favor of the active conformation resulting in a
decreased binding affinity for imatinib.
Rare cases of splicing events inducing deletions or
insertions of multiple nucleotides in the ABL kinase
domain leading to imatinib resistance have been
described. Hayette et al. (26) reported a novel frame shift
mutation acquired at the moment of imatinib resistance,
consisting an insertion of 12 nts and leading to the
conservation of open reading frame.
Mutations outside the kinase domain: It has been
shown in vitro that mutations outside the kinase domain
in the neighboring linker, SH2, SH3, and Cap domains
can confer imatinib resistance (27). In the context of
A review on drug resistance mechanisms in CML Vaidya et al.
386 ª 2011 John Wiley & Sons A/S

ABL, these domains have an autoinhibitory effect on
kinase activity, and mutations in this region can activate
the enzyme. In an attempt to determine the frequency
and relevance to resistance of regulatory domain muta-
tions, Sherbenou et al. screened for such mutations in a
cohort of consecutive patients with CML with various
levels of response. They found mutation T212R con-
ferred resistance to tyrosine kinase inhibitor and was
associated with relapse, whereas most other mutations
did not affect drug sensitivity (28).
BCR–ABL amplification: Overexpression of the Bcr–
abl protein owing to amplification of the BCR–ABL
gene was first observed in vitro when resistant CML cell
lines were generated by exposure to gradually increasing
doses of imatinib (29, 30). In one study, three of 11
patients with CML in blast crisis who relapsed after ini-
tially responding to imatinib were shown to have multi-
ple copies of the BCR–ABL gene by fluorescence in situ
hybridization (FISH) (25). In another study, seven of 55
patients showed a more than 10-fold increase in
BCR–ABL transcript levels, and two of the 32 patients
evaluated were found to have genomic amplification of
BCR–ABL by FISH (31). In the latter two patients, resis-
tance was primary and not acquired. Overexpression of
Bcr–abl leads to resistance by increasing the amount of
target protein needed to be inhibited by the therapeutic
dose of the drug. It is also possible that a transient over-
expression of BCR–ABL may be an early phenomenon in
the establishment of imatinib resistance, preceding the
emergence of a dominant clone with a mutant kinase
domain, as suggested by kinetic studies in cell lines (30).
Gene amplification as a mechanism of drug resistance
is also commonly observed where the drug inhibits an
enzyme within a critical biosynthetic pathway. For exam-
ple, methotrexate resistance is associated with amplifica-
tion of the dihydrofolate reductase (DHFR) gene 22.
Thus, amplification of genes encoding drug targets or
enzymes that can bypass a drug effect is a frequent
mechanism of drug resistance. However, amplification of
a tyrosine kinase in response to a tyrosine kinase inhibi-
tor has not previously been reported. The mechanisms of
gene amplification are not well understood. Genes may
be amplified in situ at intrachromosomal locations
through a mechanism that may involve repeated cycles
of breakage–fusion–bridge cycles, where chromosome
breakage is initiated at fragile sites (32).
Strategies to overcome imatinib resistance
Targeting tyrosine kinase activity using second-gener-ation and third-generation tyrosine kinase inhibitors
Second-generation TKIs: Several clinical observations
have shown impressing results against second-generation
Abl inhibitors nilotinib (AMN107, developed by Novar-
tis Pharmaceuticals, Basel, Switzerland) and dasatinib
(BMS-354825, developed by Bristol-Myers Squibb,
Princeton, USA). Nilotinib, like imatinib, requires the
Abl protein to be in the inactive conformation for opti-
mal binding. Nilotinib was found to be 10- to 25-fold
more potent as compared to imatinib in the reduction in
both autophosphorylation and proliferation (33) and sig-
nificantly active against 32 ⁄ 33 imatinib-resistant BCR–
Abl mutants (25). Dasatinib is a dual-specific SRC and
ABL inhibitor, structurally unrelated to imatinib that is
able to bind and inhibit both the active and inactive con-
formations of Abl, resulting in 100- to 300-fold higher
activity than imatinib. Shah et al. (34) recently demon-
strated that dasatinib has up to 100-fold increased activ-
ity against the Abl kinase compared to imatinib and
retains activity against 14 of 15 imatinib-resistant BCR–
ABL mutants in vitro.
Third-generation TKIs: The second-generation TKI, da-
satinib, holds great promise for the management of practi-
cally all imatinib-resistant mutations except for the T315I
mutation. T315I currently remains the most troublesome
mutant, and there are no effective treatment options for
patients harboring this mutation. Preclinical research has
demonstrated that AP24534 (ponatinib), an orally active
pan inhibitor of Bcr–abl, can inhibit the entire spectrum
of mutations that cause resistance to other Bcr–abl inhibi-
tors including the most resistant T315I (35). Ponatinib is
currently in phase II trials and showing impressing results
against T315I. Carbon–carbon triple bond linker is the
key structural feature of ponatinib that makes hydropho-
bic contact with the side chain of I315, allowing inhibition
of the T315I mutant. It also acts as an inflexible connector
that makes obligatory correct positioning of the two bind-
ing segments into their binding pockets. Its inhibitor pro-
file incorporates multiple contact points to confer very
high potency and balances the overall binding affinity.
The extensive network of optimized molecular contacts
leads to high potency and renders binding less susceptible
to disruption by single-point mutations (36). Another
third-generation TKI, bosutinib (SKI-606), is a dual Src–
abl inhibitor that is active in the low nanomolar range
against Bcr–abl (37). Bosutinib is now in phase III clinical
trials and has shown good activity in patients resistant to
imatinib or other TKIs in phase II (38). At nanomolar
concentrations, it inhibits the autophosphorylation of
both Abl and Src kinase, resulting in inhibition of cell
growth and apoptosis. Because of dual mechanism of
action, this agent shows activity in resistant CML disease,
other myeloid malignancies, and solid tumors. It seems to
cause fewer side effects because it more selectively inhibits
the faulty proteins in the leukemic cells and does not
affect similar proteins in normal cells as much as the ear-
lier drugs do.
Vaidya et al. A review on drug resistance mechanisms in CML
ª 2011 John Wiley & Sons A/S 387

Recently, two novel compounds were reported,
ONO12380 (39) and MK-0457 (40), to inhibit Bcr–abl
kinase activity through a distinct mechanism in cell lines
expressing the T315I mutation. Table 1 describes the
response of patients to second- and third-generation
TKIs.
Combination therapy
TKIs alone can induce remission in CML but do not elim-
inate leukemia stem cells, which remains a potential source
of relapse. Monotherapy leads to the generation of
mutants. Combination therapy involves the combination
of two or more anticancer drugs of which one is mostly a
TKI. Treatment with histone deacetylase inhibitor (HDA-
Cis) combined with imatinib effectively induced apoptosis
in quiescent CML progenitors resistant to elimination by
imatinib alone and eliminated CML stem cells capable of
engrafting immunodeficient mice. The interaction between
imatinib and HDACis inhibited genes regulating hemato-
poietic stem cell maintenance and survival (41). Combina-
tion of interferon-a and imatinib has also showed
impressing results. At 12 months, the rate of superior
molecular response was significantly higher among
patients receiving imatinib and peginterferon alfa-2a
(30%) than among those receiving 400 mg of imatinib
alone (14%) (42). Burchert et al. demonstrated that treat-
ment with IFN enables discontinuation of imatinib in
most patients after prior imatinib–IFN combination ther-
apy and may result in improved molecular response.
Induction of proteinase-3-specific CTL response by IFN
was supposed to contribute to this effect (43).
Targeting pathways downstream of BCR–ABL
The Ras ⁄Raf pathway is intimately linked to BCR–ABL
through the adaptor molecules Grb2 (growth factor
receptor–bound protein 2) and Crkl, inhibition of which
with a farnesyl transferase inhibitor (FTI), such as lona-
farnib, would theoretically reduce nuclear transcription
(11).
PI3K and its downstream targets, including the ser-
ine ⁄ threonine kinases AKT, mTOR, and p70S6 kinase,
play a key role in the regulation of cell survival and pro-
liferation. Recently, it was shown that activation of the
PI3K ⁄mTOR pathway by Bcr–abl contributes to
increased production of reactive oxygen species (44), thus
linking PI3K ⁄mTOR to mechanisms that have been
implicated in genomic instability and imatinib resistance.
Signal transduction through this pathway can be blocked
by inhibitors such as LY294002 (11) or wortmannin,
which specifically target PI3K. Cellular response to PI3K
inhibition includes induction of apoptosis and inhibition
of proliferation.
Modulating the levels of imatinib in Plasma
Imatinib is metabolized largely by the cytochrome p450
isoenzymes CYP3A4 and CYP3A5. Differences in the
concentrations of CYP3A4 ⁄CYP3A5 or drugs that can
inhibit or induce said enzymes have the potential to
greatly affect the levels of imatinib in plasma (10). Also,
in this regard, dose escalation of imatinib (600–
1000 mg ⁄d) can overcome resistance to standard-dose
imatinib (400 mg ⁄d) in some patients with CML. In
case of P-loop mutations, escalation of the dose of
imatinib is not recommended owing to highly resistant
nature of these mutations (38). Additionally, it is possi-
ble that this treatment will benefit only patients who
have suboptimal intracellular drug level with standard
dose of imatinib.
CML vaccines
In recent years, a series of DNA vaccines have been
developed based on different aspects to target the CML
cells. Lucansky et al. (45) demonstrated the use of a plas-
mid carrying either the complete BCR–ABLl fusion gene
or fragment thereof coding for a 25-amino-acid-
sequence-long junction zone (bcr–abl 25 amino acids)
linked with genes coding for a variety of immunostimula-
tory factors, as the DNA vaccines in mice. Peptide vac-
cines derived from amino acid sequences crossing the
b3a2 fusion breakpoint have proved their efficacy in
patients with chronic-phase CML (46). These vaccines
elicit class I restricted cytotoxic T lymphocytes and class
II responses. Investigators from Johns Hopkins Sidney
Kimmel Comprehensive Cancer Center have developed a
vaccine made from CML cells irradiated to halt their
cancerous potential and genetically altered to produce an
immune system stimulator called GM-CSF. The tumor
cells also carried antigens specific to CML cells, which
primed the immune system to recognize and kill circulat-
ing CML cells.
Destabilizing BCR–ABL protein
Stability of Bcr–abl is dependent upon its ability to form
complex with HSP90 and co-chaperone protein p23.
HSP90 antagonist such as 17-allylamino-17-demethoxy-
geldanamycin (17-AAG) interfere with Bcr–abl chaperon-
ing such that the mature protein would be unable to fold
correctly into its quaternary structure, leaving it suscepti-
ble to proteasome degradation (11).
Targeting mRNA of BCR–ABL
RNA Interference Technology: RNA interference is a
powerful tool that has been used to inhibit gene function
A review on drug resistance mechanisms in CML Vaidya et al.
388 ª 2011 John Wiley & Sons A/S

Tab
le1
Responses
tofirs
t-,
second-,
and
third-g
enera
tion
tyro
sin
ekin
ase
inhib
itors
(TK
Is)
Dru
gs
Mechanis
mof
action
Chem
ical
str
uctu
re
Response
Advanta
ges
Dis
advanta
ges
Chro
nic
phase
Refe
rences
Accele
rate
dphase
and
bla
st
phase
Refe
rences
HR
CR
MR
HR
CR
MR
Imatinib
Imatinib
functions
by
bin
din
gin
the
ATP
bin
din
gsite
and
sta
bili
zes
the
inactive
confo
rmation
of
BC
R-A
BL,
in
whic
hth
ew
ell
know
n‘D
FG
tria
d’
isin
the
‘out’
confo
rmation
93%
CH
R
45%
CC
yR
,
11%
MC
yR
,
9%
mC
yR
93%
MM
R,
7%
CM
R
54
43%
CH
R
14%
CC
yR
,
14%
MC
yR
,
14%
mC
yR
54
First-
line
treatm
ent
option
for
chro
nic
myelo
idle
ukem
ia
(CM
L).
First
site-
directe
danti-c
ancer
dru
gw
ith
few
er
sid
e
eff
ects
.S
afe
for
pedia
tric
patients
with
CM
L
Sto
mach
upset,
muscle
cra
mps,
poundin
gheart
beat,
extr
em
etire
dness,
unexpla
ined
weig
ht
gain
.R
are
:
bla
ck
⁄blo
ody
sto
ols
,
dark
urine,
yello
win
g
eyes
⁄skin
,fe
ver,
chill
s,
pers
iste
nt
sore
thro
at,
serious
alle
rgic
reaction,
itchin
g,
trouble
bre
ath
ing
Nilo
tinib
(firs
t-lin
e)
Mechanis
mof
action
of
nilo
tinib
issim
ilar
toim
atinb.
Only
diffe
rence
is,
incorp
ora
tion
of
altern
ative
bin
din
ggro
ups
to
N-m
eth
ylp
ipera
zine,
while
am
ide
pharm
acophore
is
pre
serv
ed
tore
tain
H-b
ond
inte
ractions
toG
lu286
and
Asp381
96%
CC
yR
85%
MM
R
55
Nilo
tinib
was
specifi
cally
desig
ned
toim
pro
ve
upon
the
mole
cula
rstr
uctu
re
of
GLIV
EC
as
aw
ay
of
pre
venting
Ph+
CM
Lre
sis
tance
to
the
imatinib
mole
cule
Unexpla
ined
ble
edin
g
or
bru
isin
g,
blo
od
in
urine
or
sto
ol,
fever,
sore
thro
at,
chill
s,
or
oth
er
sig
ns
of
infe
ction,
sw
elli
ng
of
hands,
ankle
s,
feet,
or
face
Nilo
tinib
(second-lin
e)
67%
CH
R
22%
mC
yR
,
11%
PC
yR
,
11%
CC
yR
33%
MM
R
56
100%
CH
R
0%
mC
yR
,
0%
CC
yR
,
50%
PC
yR
0% M
MR
56
Dasatinib
(firs
t-lin
e)
Dasatinib
bin
ds
inth
e
ATP
-bin
din
gsite
of
the
AB
L
kin
ase.
Itbin
ds
toth
eactive
or
‘opened’
confo
rmation
of
the
BC
R-A
BL
kin
ase
dom
ain
and
ispre
dic
ted,
based
on
modelin
gstu
die
s,
toals
obin
d
toth
ein
active
or
‘clo
sed’
confo
rmation
98%
CC
yR
82%
MM
R,
10%
CM
R
55
77%
CC
yR
,46%
MM
R
57
Patients
with
new
ly
dia
gnosed
CM
L
show
superior
com
ple
te
cyto
genetic
response
and
majo
r
mole
cula
rre
sponse
in1
yr
when
treate
d
with
once
daily
dasatinib
com
pare
d
with
once
daily
imatinib
.A
lso,
active
again
st
muta
tions
whic
hchange
the
confo
rmation
of
kin
ase
from
clo
sed
toopen
Severe
alle
rgic
reactions,
bla
ck,
tarr
y,
or
bright
red
sto
ols
,blo
od
inth
e
vom
it,
depre
ssio
n,
unusualsw
elli
ng
or
weig
ht
gain
,unusual
tire
dness
or
weakness,
vom
it
that
looks
like
coff
ee
gro
unds
Dasatinib
(second-lin
e)
81%
CH
R
25%
mC
yR
,
31%
CC
yR
,
6%
PC
yR
13%
MM
R56
88%
CH
R
13%
mC
yR
,
25%
CC
yR
,
13%
PC
yR
13%
MM
R
56
Vaidya et al. A review on drug resistance mechanisms in CML
ª 2011 John Wiley & Sons A/S 389

either by increasing the destruction of mRNA corre-
sponding to the gene or, in some cases, by inhibiting the
transcription of the gene or the translation of mRNA to
the corresponding protein. The use of RNA interference,
using a breakpoint-specific short interfering RNA, has
been shown to selectively inhibit BCR–ABL-dependent
proliferation, as well as to enhance the effect of imatinib
against cells expressing an imatinib-resistant point
mutant and cells overexpressing Bcr–abl (47).
Antisense oligonucleotide
Bcr–abl is an ideal molecular target for therapy based
on antisense (AS) strategies. By designing an AS oligo-
nucleotide that is complementary to both the BCR and
ABL sequences on either side of the BCR–ABL tran-
script junction, it should be possible to generate a spe-
cies that would hybridize to BCR–ABL mRNA, thus
forming mRNA ⁄oligonucleotide heteroduplexes and thus
preventing BCR–ABL mRNA from associating with
ribosomes, leaving the transcript open for degradation
by RNase H.
Antisense strategies include antisense molecules such
as 2¢-O-methoxyethyl RNA, locked nucleic acids, and
peptide nucleic acids, as well as more exotic chemical
species such as morpholinos (48). Despite its encouraging
results with murine models, this technique has failed to
fulfill its promise as a treatment for CML. Lack of speci-
ficity of the junction AS oligos, the need to synthesize
them with special chemical backbones and to introduce
them into the cells via stringent permeabilization proce-
dures, and the extremely long life of the Bcr–abl protein
are some of the reasons for the failure (49). Another dis-
advantage of this method includes high quantities of
antisense oligonucleotides required to reach an effective
dose and thus the potential toxicity as a result of non-
specific binding (50).
Ribozymes
Ribozymes are RNA molecules that are capable of
associating with other RNA molecules by base-pairing
and catalyzing the hydrolysis of specific phosphodiester
bonds within the target RNA sequence. To date, vari-
ous types of ribozyme have been constructed to cleave
other RNAs and such trans-acting ribozymes include
hammerhead, hairpin, and HDV ribozymes. Ribozymes
can discriminate mutant sequences from their wild-type
counterparts, even when they differ by single nucleotide
base. Lance H. Leoplod demonstrated 3 log reduction
in the level of bcr–abl1 mRNA when multiunit ribo-
zyme was transfected via folate receptor–mediated
uptake into transformed 32Dcells (expressing bcr–abl
protein) (51).Tab
le1
(Continued)
Dru
gs
Mechanis
mof
action
Chem
ical
str
uctu
re
Response
Advanta
ges
Dis
advanta
ges
Chro
nic
phase
Refe
rences
Accele
rate
dphase
and
bla
st
phase
Refe
rences
HR
CR
MR
HR
CR
MR
Bosutinib
Bosutinib
isa
dualkin
ase
inhib
itor.
Itin
hib
its
the
auto
phosphory
lation
of
both
Abland
Src
kin
ases,
resultin
gin
inhib
itio
nof
cell
gro
wth
and
apopto
sis
86%
CH
R37%
MC
yR
,
22%
CC
yR
27%
MM
R,
5%
CM
R
58
61%
CH
R48%
MC
yR
,
33%
CC
yR
15%
MM
R,
4%
CM
R
58
Superior
rate
sof
majo
rm
ole
cula
r
response
(MM
R)
and
cum
ula
tive
com
ple
tem
ole
cula
r
response,
com
pare
d
with
the
sta
ndard
thera
py,
imatinib
Dia
rrhoea,
rash,
incre
ased
ala
nin
e
transam
inase,
thro
mbocyto
penia
,
neutr
openia
,
anaem
ia
Ponatinib
Carb
on-c
arb
on
trip
lebond
linker
makes
hydro
phobic
conta
ct
with
the
sid
echain
of
I315,
allo
win
g
inhib
itio
nof
the
T315I
muta
nt.
It
als
oacts
as
an
inflexib
le
connecto
rth
at
makes
oblig
ato
ry
corr
ect
positio
nin
gof
the
two
bin
din
gsegm
ents
into
their
bin
din
gpockets
.It
sin
hib
itor
pro
file
incorp
ora
tes
multip
le
conta
ct
poin
tsto
confe
rvery
hig
h
pote
ncy
and
bala
nce
the
overa
ll
bin
din
gaffi
nity
95%
CH
R66%
MC
yR
,
53%
CC
yR
No
CH
R,
35%
MH
R.
InT315I,
20%
MH
R
24%
MC
yR
,
12%
CC
yR
.
InT315I
muta
tions,
20%
MC
yR
Active
again
st
T315I
and
oth
er
TK
I
resis
tant
muta
nts
Sid
eeff
ects
:
Thro
mbocyto
penis
,
headache,
nausea,
art
hra
lgia
,fa
tigue,
anem
ia
HR
,haem
ato
logic
response;
CyR
,cyto
genetic
response;
MR
,m
ole
cula
rre
sponse;
CH
R,
com
ple
tehaem
ato
logic
response
(Norm
aliz
ation
of
blo
od
count)
;C
CyR
,com
ple
tecyto
genetic
response
(Ph+
0);
PC
yR
,part
ialcyto
genetic
response
(Ph+
1–35%
);M
CyR
,
min
or
cyto
genetic
response
(Ph+
36–65%
);m
CyR
,m
inim
alcyto
genetic
response
(66–95%
);C
MR
,com
ple
tem
ole
cula
rre
sponse
(BC
R–A
BL
transcripts
non
quantifiable
and
non
dete
cta
ble
).
A review on drug resistance mechanisms in CML Vaidya et al.
390 ª 2011 John Wiley & Sons A/S

Blocking the oligomerization of BCR–ABL
Monomeric Bcr–abl is non-transforming, whereas Bcr–
abl tetramers are oncogenic, transforming species. Thus,
Bcr–abl must undergo transformation to become acti-
vated. Beissert et al. (52) demonstrated that targeting the
N-terminal coiled coil oligomerization interface of bcr
interferes with the transformation potential of Bcr–abl
and increases sensitivity to STI571. Although currently
none of the available therapies for CML target the oligo-
merization domain of Bcr–abl, this approach could be
exploited for the therapeutic gain as a combined therapy
with imatinib.
Entrapment of BCR–ABL in the nucleus
BCR–ABL has a cytoplasmic localization, in contrast to
ABL, mostly nuclear. The shuttling of ABL between the
nucleus and the cytoplasm is accomplished by the presence
of import and export signal sequences in the COOH- ter-
minal region of ABL. Interestingly, these sequences are
conserved in BCR–ABL. It has been shown that the inhi-
bition of BCR–ABL export from the nucleus has been
shown to induce apoptosis in Bcr–abl-expressing cells.
This inhibition is accomplished by the use of export inhibi-
tors such as leptomycin. Vigneri et al. showed that com-
bining STI571 with leptomycin B blocks the nuclear
export of BCR–ABL, which in turn activates apoptosis.
This combined therapy causes the irreversible and com-
plete killing of BCR–ABL-transformed cells, whereas the
effect of either drug alone is fully reversible (53). These
results indicate that nuclear entrapment of BCR–ABL can
be used as a therapeutic strategy to selectively kill chronic
myelogenous leukemia cells.
Conclusion
A remarkable breakthrough in cancer therapy came with
the introduction of imatinib to the treatment for CML.
Like other anticancer drugs, clinical resistance to imati-
nib therapy emerged. This picture gave rise to invention
of new second-generation and thereafter third-generation
TKI.
With so much advancement in CML therapy, the fact
that there is no permanent cure for CML and relapses is
a common scenario in patients with CML on treatment,
which still remains unavoidable. Bearing in mind that
CML is a pluripotent stem cell disorder, newer methods
targeting the CML stem cells should be designed.
References
1. Clarkson B. Chronic myelogenous leukemia: etiology,
incidence, and clinical features. Encycl Cancer
2002;1:505–18.
2. Deininger MWN, Goldman JM, Melo JV. The molecu-
lar biology of chronic myeloid leukemia. Blood
2000;96:3343–55.
3. Daley GQ, Van Etten RA, Baltimore D. Induction of
chronic myelogenous leukemia in mice by the P210bcr ⁄ ablgene of the Philadelphia chromosome. Science 1990;
247:824–30.
4. Lugo TG, Pendergast AM, Muller AJ, Witte ON. Tyro-
sine kinase activity and transformation potency of bcr ⁄ abloncogene products. Science 1990;247:1079–82.
5. Kantarjian HM, Talpaz M, Giles F, O’Brien S, Cortes J.
New insights into the pathophysiolody of chronic myeloid
leukemia and imatinib resistance. Ann Intern Med
2006;145:913–23.
6. Druker BJ. STI571 (Gleevec) as a paradigm for cancer
therapy. Trends Mol Med 2002;8(4 Suppl.):S14–8.
7. Azam M, Daley GQ. Anticipating clinical resistance to
target-directed agents: the BCR-ABL paradigm. Mol
Diagn Ther 2006;10:67–76.
8. Weisberg E, Griffin J. Resistance to imatinib (Glivec):
update on clinical mechanisms. Drug Resist Updat
2003;6:231–8.
9. Kantarjian H, Sawyers C, Hochhaus A, et al. Hematologic
and cytogenetic responses to imatinib mesylate in chronic
myelogenous leukaemia. N Engl J Med 2002;346:645–52.
10. Quintas-Cardama A, Kantarjian H, Cortes J. Mechanisms
of primary and secondary resistance to imatinib in chronic
myeloid leukemia. Cancer Control 2009;16:122–31.
11. Jorgensen HG, Holyoake TL. Characterization of cancer
stem cells in chronic myeloid leukaemia. Biochem Soc
Trans 2007;35:1347–51.
12. Copland M, Hamilton A, Elrick LJ, Baird JW, Allan EK,
Jordanides N, Barow M, Mountford JC, Holyoake TL.
Dasatinib targets an earlier progenitor population than
imatinib in primary CML but does not eliminate the
quiescent fraction. Blood 2006;107:4532–9.
13. Gambacorti-Passerini CB, Gundy RH, Piazza R, Galietta
A, Rostagno R, Scapozza L. Molecular mechanisms of
resistance to imatinib in Philadelphia-chromosome-posi-
tive leukaemias. Lancet Oncol 2003;4:75–85.
14. Larghero J, Mahon FX, Madeleine-Chambrin I, et al.
Elevated levels of the plasma protein alpha 1 acid glyco-
protein in chronic myelogenous leukemia in blast crisis
mediate pharmacological resistance to Gleevac (STI571,
imatinib) in vitro and are associated with primary resis-
tance in vivo. Presented at the 43rd annual meeting of the
American Society of Hematology 2001, Orlando.
15. Galimberti S, Cervetti G, Guerrini F, Testi R, Pacini S,
Fazzi R, Simi P, Petrini M. Quantitative molecular moni-
toring of BCR-ABL and MDR1 transcripts in patients
with chronic myeloid myeloid leukaemia during imatinib
treatment. Cancer Genet Cytogenet 2005;162:57–62.
16. Mahon FX, Belloc F, Lagarde V, Chollet C, Moreau-
Gaudry F, Reiffers J, Goldman J, Melo J. MDR1 gene
overexpression confers resistance to imatinib mesylate in
leukaemia cell line models. Blood 2003;101:2368–73.
Vaidya et al. A review on drug resistance mechanisms in CML
ª 2011 John Wiley & Sons A/S 391

17. Ferrao P, Frost MJ, Siah SP, Ashman L. Overexpression
of P-glycoprotein in K562 cells does not confer resistance
to the growth inhibitory effects of imatinib (STI571) in
vitro. Blood 2003;102:4499–503.
18. Dulucq S, Bouchet S, Turcq B, Lippert E, Etienne G,
Reiffers G, Molimard M, Krajinovic M, Mahon FX.
Multidrug resistance (MDR1) polymorphisms are associ-
ated with major molecular responses to standard-dose
imatinib in chronic myeloid leukemia. Blood
2008;112:2024–7.
19. Yamada O, Ozaki K, Furukawa T. Activation of STAT 5
confers imatinib resistance on leukemic cells through the
transcription of TERT and MDR1. Cell Signal
2011;23:1119–27.
20. Shah NP, Nicoll JM, Nagar B, Gorre ME, Paquette RL,
Kuriyan J, Sawyers CL. Multiple BCR ⁄ABL kinase
domain mutations confer polyclonal resistance to the tyro-
sine kinase inhibitor imatinib (STI571) in chronic myeloid
leukaemia. Cancer Cell 2002;2:117–25.
21. Nardi V, Azam M, Daley G. Mechanisms and implica-
tions of imatinib resistance mutations in BCR-ABL. Curr
Opin Hematol 2004;11:35–43.
22. von Bubnoff N, Schneller F, Peschel C, Duyster J. BCR-
ABL gene mutations in relation to clinical resistance of
Philadelphia-chromosome-positive leukemia to STI-571; a
prospective study. Lancet 2002;359:487–91.
23. Hughes T. Mechanisms of resistance, common BCR-ABL
mutations, and monitoring response to treatment in
CML. Medscape 2008. http://www.medscape.org/viewarticle/
575913. Accessed August 17, 2011.
24. Branford S, Rudzki Z, Walsh S, et al. Detection of
BCR ⁄ABL mutations in patients with CML treated with
imatinib is virtually always accompanied by clinical resis-
tance, and mutations in the ATP phosphate-binding loop
(P-loop) are associated with a poor prognosis. Blood
2003;102:276–83.
25. Gorre ME, Mohammed M, Ellwood K, Hsu N, Paquette
R, Rao PN, Sawyers CL. Clinical resistance to STI-571
cancer therapy caused by BCR-ABL gene mutation or
amplification. Science 2001;293:876–80.
26. Hayette S, Chabane K, Tchirkov A, Berger M, Nicolini
F, Tournilhac O. Detection of twelve nucleotide insertions
in the BCR ⁄ABL kinase domain in an imatinib resistant
but dasatinib sensitive patient with bi-phenotypic acute
leukemia. Haematologica 2009;94:1324–6.
27. Azam M, Latek R, Daley G. Mechanisms of autoinhibi-
tion and STI-571 ⁄ imatinib resistance revealed by muta-
genesis of BCR-ABL. Cell 2003;112:831–43.
28. Sherbenou D, Hantschel O, Kaupe I. BCR-ABL SH3-
SH2 domain mutations in chronic myeloid leukemia
patients on imatinib. Blood 2010;116:3278–85.
29. Mahon FX, Deininger MW, Schultheis B, Chabrol J,
Reiffers J, Goldman JM, Melo JV. Selection and charac-
terization of BCR-ABL positive cell lines with differential
sensitivity to the tyrosine kinase inhibitor STI571: diverse
mechanisms of resistance. Blood 2000;96:1070–9.
30. Barnes D, Schultheis B, Adedeji S, Melo J. Dose-depen-
dent effects of Bcr-Abl in cell line models of different
stages of chronic myeloid leukemia. Oncogene
2005;24:6432–40.
31. Hochhaus A, Kreil S, Corbin AS. Molecular and chromo-
somal mechanisms of resistance to imatinib (STI571)
therapy. Leukemia 2002;16:2190–6.
32. Weisberg E, Griffin J. Mechanism of resistance to theABL
tyrosine kinase inhibitor STI571 in BCR ⁄ABL-trans-
formed hematopoietic cell lines. Blood 2000;95:3498–505.
33. Weisberg E, Manley PW, Breitenstein W. Characterization
of AMN107, a selective inhibitor of native and mutant
BCR ⁄ABL. Cancer Cell 2005;7:129–41.
34. Shah NP, Tran C, Lee FY, Chen P, Norris D, Sawyers
CL. Overriding imatinib resistance with a novel ABL
kinase inhibitor. Science 2004;305:399–401.
35. O’Hare T, Shakespeare W, Zhu X, et al. AP24534, a
Pan-BCR-ABL inhibitor for chronic myeloid leukemia,
potently inhibits the T315I mutant and overcome
mutation-based resistance. Cancer Cell 2009;16:401–12.
36. Zhou T, Commodore L, Huang WS, et al. Structural
mechanism of the Pan-BCR-ABL inhibitor ponatinib
(AP24534): lessons for overcoming kinase inhibitor resis-
tance. Chem Biol Drug Des 2011;77:1.
37. Golas JM, Arndt K, Etienne C, Lucas J, Nardin D,
Gibbons J, Frost P, Ye F, Boschelli DH, Boschelli F.
SKI-606, a 4-anilino-3-quinolinecarbonitrile dual inhibitor
of Src and Abl kinases, is a potent antiproliferative agent
against chronic myelogenous leukemia cells in culture and
causes regression of K562 xenografts in nude mice. Cancer
Res 2003;63:375–81.
38. Cortes-Franco J, Kantarjian H, Kim DW, Khoury HJ,
Turkina A, Shen ZX, Brummendorf TH, Chandy M,
Arkin S, Gambacorti-Passerini C. Efficacy and safety of
bosutinib (SKI-606) among patients with chronic phase
Ph+ chronic myelogenous leukemia (CML). Blood
2007;110:11.
39. Bartholomeusz G, Talpaz M, Kapuria V, Kong LY,
Wang S, Estrov Z, Priebe W, Wu J, Donato N. Activation
of a novel Bcr ⁄Abl destruction pathway by WP1130
induces apoptosis of chronic myelogenous leukemia cells.
Blood 2007;109:3470–8.
40. Giles FJ, Cortes J, Jones D, Bergstrom D, Kantarjian H,
Freedman SJ. MK-0457, a novel kinase inhibitor, is active
in patients with chronic myeloid leukemia or acute lym-
phocytic leukemia with the T315I BCR-ABL mutation.
Blood 2007;109:500–2.
41. Zhang B, Guilhot J, Nicolini FE. Effective targeting of
quiescent chronic myelogenous leukemic stem cells by his-
tone deacetylase inhibitors in combination with imatinib
mesylate cancer. Cell 2010;17:427–42.
42. Preudhomme C, Guilhot J, Nicolini FE, et al. Imatinib
plus peginterferon alfa-2a in chronic myeloid leukemia.
N Engl J Med 2010;363:2511–21.
43. Burchert A, Muller MC, Kostrewa P, Erben P, Bostel T,
Liebler S, Hehlmann R, Neubauer A, Hochhaus A, et al.
A review on drug resistance mechanisms in CML Vaidya et al.
392 ª 2011 John Wiley & Sons A/S

Sustained molecular response with interferon alfa mainte-
nance after induction therapy with imatinib plus inter-
feron alfa in patients with chronic myeloid leukemia.
J Clin Oncol 2010;28:1429–35.
44. Kim J Chu S, Gramlich J, Pride Y, Babendreier E,
Chauhan D, Salgia R, Podar K, Griffin J, Sattler M, et al.
Activation of the PI3K ⁄mTOR pathway by BCR ⁄ABL
contributes to increased production of reactive oxygen
species. Blood 2005;105:1717–23.
45. Lucansky V, Sobotkova E, Tachezy R, Duskova M,
Vonka V. DNA vaccination against bcr-abl positive cells
in mice. Int J Oncol 2009;35:941–51.
46. Pinilla-Ibarz J, Cathcart K, Korontsvit T, Soignet S,
Bocchia M, Caggiano J, Lai L, Jimenez J, Kolitz J,
Scheinberg DA. Vaccination of patients with chronic
myelogenous leukemia with bcr-abl oncogene breakpoint-
fusion peptides generates specific immune responses. Blood
2000;95:1781–7.
47. Wohlbold L, van der Kuip H, Miething C, Vornlocher H,
Knabbe C, Duyster J, Aulitzky W. Inhibition of
BCR ⁄ABL gene expression by small interfering RNA sen-
sitizes for imatinib mesylate (STI-571). Blood 2003;2:2236–
9.
48. Melo J, Hughes T, Apperley J. Chronic myeloid leukemia.
Am Soc Hematol 2003;1:132–52.
49. Clark RE. Antisense therapeutics in chronic myeloid
leukaemia: the promise, the progress and the problems.
Leukemia 2000;14:347–55.
50. Dhut S, Chaplin T, Young BD. BCR ⁄ABL and BCR
proteins: biochemical characterization and localization.
Leukemia 1990;4:745–50.
51. Leopold L, Shore S, Newkirk TA, Reddy R, Reddy EP.
Multi-Unit Ribozyme-Mediated Cleavage of bcr-abl
mRNA in Myeloid Leukemias. Blood 1995;85:2162–70.
52. Beissert T, Puccetti E, Bianchini A, Guller S, Boehrer S,
Hoelzer D, Ottmann OG, Nervi C, Ruthardt M. Target-
ing of the N-terminal coiled coil oligomerization interface
of BCR interferes with the transformation potential of
BCR-ABL and increases sensitivity to STI571. Blood
2003;102:2985–93.
53. Vigneri P, Wang J. Induction of apoptosis in chronic mye-
logenous leukemia cells through nuclear entrapment of
BCR–ABL tyrosine kinase. Nat Med 2001;7:228–34.
54. Parikh P, Saikia T, Tripathi A, Shah S, Sheth A, Shah P.
Role of Imatinib Mesylate in Chronic Myeloid Leukemia.
In: Agarwal MB, ed. Chronic Myeloid Leukemia, 1st edn.
Mumbai: Ranbaxy Super Specialities Ranbaxy House,
2007:89–105.
55. Rosti G, Castagnetti F, Gugliotta G, Palandri F,
Baccarani M. Second-generation BCR-ABL inhibitors
for frontline treatment of chronic myeloid leukemia in
chronic phase. Crit Rev Oncol Hematol 2011. doi: 10.1016/
j.critrevonc.2011.04.002. Epub ahead of print.
56. Garg R, Kantarjian H, O’Brien S, Quinta¢s-Cardama A,
Faderl S, Estrov Z, Cortes J. The use of nilotinib or
dasatinib after failure to 2 prior tyrosine kinase inhibitors:
long-term follow up. Blood 2009;114:4361–8.
57. DiBella N. First-line treatment of chronic myeloid leuke-
mia: imatinib versus nilotinib and dasatinib. Commun
Oncol 2011;8:65–72.
58. Nelson R. Bosutinib effective as second- and third-line
therapy in CML. ASCO 2010 Annual Meeting.
Vaidya et al. A review on drug resistance mechanisms in CML
ª 2011 John Wiley & Sons A/S 393