Embed Size (px)
Citation preview

PUV A (5-Methoxypsoralen Plus UV A) Enhances Melanogenesis and Modulates Expression of Melanogenic Proteins in Cultured Melanocytes
Valerie Mengeaud* and Jean-Paul OrtolU'le INSERM U.385 , Faculte de Mcdecine, 06107 Nice, France
PUV A (combination of psoralens and ultraviolet A radiation) is a potent inducer of melanogenesis in normal human skin. The molecular mechanisms underlying this effect are poorly characterized. This study was undertaken to investigate the action of PUV A on melanogenesis in S91 murine melanoma cells and in cultured normal human melanocytes. Tyrosinase and DOP Achrome tautomerase (DCT) activities as well as melanin neosynthesis were measured in PUV A-treated pigment cells. To determine whether a correlation exists between PUV A-induced melanogenesis and expression of n1.elanogenic enzymes, we analyzed the levels of tyrosinase, DCT, and tyrosinase-related protein-1 (TRP-1 or gp75) by western blotting in PUV A-treated cells. We demonstrate that UV A upregulates tyrosinase activity and melanin content with 5-methoxypsoralen at 1 /LM. This phenomenon depends on the energy delivered during phototreatment. In both human and mouse cells, stimulation of melanogenesis correlated with
Oral administration ofpsoraJen followed by sunl.ight or UV A radiation (PUV A) enhan ces human skin pigmentation (Fitzpatrick, 1989). This PUV Ainduced epidermal melanin pigmentation is a more e fi:ective filter than a tan induced by UVB
(Gschnait et ai , 1978) . In addition, PUV A indu ces re pigm entation of the hypo melanotic lesions of vitiligo (Nordlund and Lerner, 1982), by stimulating proliferation and! or migration of follicular melanocytes (OrtolUle et ai, 1983).
PUV A, like UW radiation, increases the number of epidermal L-3,4-dihydroxyphenylalanine (DOPA)-positive m elanocytes and induces melanin production ill ";"0 (Szabo et nl, 1982). In cultured
Manuscript received January 14 , 1994 ; revised February 8, 1996; accepted for publication March 5, 1996.
Reprint requests to: Valerie Mcngeaud ' and J. P. Ortonne, INSER.M U. 385, Faculte de Mcdccine, Av de Vnlombrose, 06107 Nice, Cedex 2, France.
Abbreviations: DOPA, L-3,4-dihydroxyphellylalanine; OHICA, 5,6-dihydroxyindole 2-carboxylic acid; DHl, dihydroxyindole; MSH, melanocyte-s timulating hormone; IBMX, isobutylmcthyl-xanthinc; PBS, phosphate-buffered saline; 5-MOP, 5-metho,:ypsoralen; OCT, OOPAchromc tautomerasc; TRP-1, tyrosinase-related protein-I.
• Current address: Institut Pierre Fabre, Ocpartement R.echerche Dcvcloppement Galcnique, R.ue Jean Rostand, B.P. 687, 31319, Labege Innopole, France.
an increase of the amount of tyrosinase. In PUV Atreated S91 cells, tyrosinase mRNA was increased, but no stimulation ofDCT activity occurred in these cells, in agreement with the unchanged amount of DCT protein in cell extracts. On the contrary, in melanocytes treated with PUV A, a decrease in DCT protein was observed. Finally, the amount of TRP-l protein was not affected by PUV A in either S91 cells or melanocytes.
These results show that melanogenesis induced by PUV A is related to an increase in expression of tyrosinase. In melanocytes, melanogenesis and DCT are negatively correlated, which suggests that PUV A favors the metabolic pathway of dark-eumelanins with high UV -protective properties. This study also suggests that PUV A regulates tyrosinase, DCT, and TRP-l expression in a noncoordinate manner. Kel' words: pso,'oleu/llitraJ,iolet A/PUVAlllormallum,.otl mela1l.ocytes/S91 me1a,wl1Ia cells/meiallogellesis/ty,'osillase. ] Itlvest D ermotol 107:57-62, 1996
human melanocytes, both PUV A and UVB radiation stimulate tyrosinase activity and decrease cell proliferation (Friedmann and Gilchrest, 1987; Kao and Y u, 1992; Aberdam e/ nl, 1993), Involvement of DNA alky lation by photoactivated psoralens and generation of reactive oxygen species have been suggested as melanogenic effects of PUVA (Chatteljee and Cantor, 1978; Cundari and Averbeck, 1988; Bordin et nl, 1994), but since high-affinity binding sites for 8-metho:\:ypsoralen have been identified in HeLa cells exposed to UV lig ht (Laskin et ai, 1985), it has also been suggested that PUV A exerts its action through hormone-receptor interactions.
T he effect of PUV A on the enzym es regulating melanogenesis is not und erstood. Burchill el nl (1990) showed that the amount of tyrosinase, the rate-limiting enzyme of melanogenesis, is increased in human skin biopsies after several weeks of PUV A therapy. Tyrosinase, the first enzym e described to regulate melanogenesis, initiates m elanogenesis by catalyzing the convers ion of tyrosine to L-3,4-dihydroxyphenylalnnine (DOPA) a.nd then to dopaquinone. Synthesized as a precursor form of 55 kDa, tyrosinase undergoes extensive post-translation al modifications in the Golgi apparatus where, after glycosylatio n , it is found as a mature form of about 70 kDa (Hearing cl nl, 1978). Recently, two other enzymes, DOPAchrome tautom erase (DCT) and tyrosinase-related-protein 1 (TRP-1 , analogous to human gp75) have also been sh ov\1l1 to regulate m elanogenesis. DCT catalyzes the transformation of DOPA chrome
0022-202X/96/$10.50 • Copyright 1996 by The Society for Investigative Dermatology, Illc.
57

58 MENGEAUD AND ORTONNE
to th e ca rboxylated d e riva tive 5,6-dihydroxyindole 2-carboxylic acid (OHlCA) (Korner and Getting, 1985; Aroca et ai, 1990a;
Tsukamoto et ai, 1992) and regulates the ratio of OHlCA and dihydro,-:yindole (OHI) . Althoug h TRP-1 h as b een prove d to be respon sible for the produ ction of e umelanins, littl e is known about its catalytic function Oackson, 1988; Vijayasaradhi el ai, 1990). Among the activities postulated for TlUl-1 are tyrosinase with tyrosin e hydroxylase activity Oimenez el ai, 1991; Jimenez- Cerventes et ai, 1.994), OOPAchrome tautomerase (Winder et ai, 1993), m elanosomal ca talase (Halaban and Moellman, 1990), and OHICA converting factor (Kobayashi el ai, 1994). Regulation of tyrosinase, OCT, and TRP-1 have been studie d particularly in pigment cell s stimulate d by m e lanocyte-stimuJating hormone (MSH), chole ra toxin, or isobutyl methyl-xanthine (IBMX) and a fter UVB radiation (FuIJ e r e/ ai, 1987; Aroca et ai, 1993 ; Orlow e/ ai, 1 993; Romero el ai, 1994). Whether the regulation of these m e lanogenic e n zym es occurs by modulation of synthetic rate of prote in s o r mRNA levels is not cl early establishe d. In S91 melanoma cell s, MSH-, IBMX-, or UVB-indu ced melanogenesis involves an inc re ase in th e synthesis of tyrosin ase (Halaban e/ ai, 1983; Aberdam el ai, 1993). In B16FI0 melanoma ce ll s, however, MSH stimulates m e lanogen esis by inc reasing tyrosinase, TRP-l , and OCT activities w ith no c h ange in the rate of synth es is of these en zymes (Kameyama el ai , 1993). Therefore, the biochemical processes respon sible for PUV A-induced skin tanning are poorly understood. T his study was d es ig ned to analyze th e modula tion of
m e lanogenic proteins (tyrosinase, OCT, and TRP-l) in culture d S91 mouse m e lanoma cells and cul tured m e lanocytes a fter PUVA.
MATEIUALS AND METHODS
Cells and Cultures Techniques Murine melanoma cell s S91-M3 (Flow Laboratori es, Irvine, Scotland) , were cultured accord ing to standard proccdures. Human melanocytes were cultured using a modification of thc proccdurc of Eisinger and Marko (1982) . BrieRy, cpidcrma l ce ll suspensions were obta in cd from foreskins of caucasoid childrcn by ovcrnjght digestion in t rypsi .~/eth y l cnediamil1c terraacctic acid solution [0.05')10 /0.02% in phosphate-buffered saline (PBS)) at 37· C. They werc grown in MCDU 153 medium supplemented with 5 fLg insulin pe r ml , 0 .5 fLg hydrocorti sone per ml , 16 nM PMA, 10 fLM forsko lin , 30 fLg of prote in 1m I of bovine pituitary extract [bPE; preparcd according to Wilkins cl al (1985)] and penicillinl streptomycin (100 IU /5 0 fLg/m l). Pure melanocyte subcultu res were trypsinized every 3 weeks. Second-passagc melanocytes were used for experiments. Viable cell numbers were dctermined by the trypan blue dye exclusion assay.
PUVA Treatment The source of ultraviolet radiation was a Vilbe r Lourmat stimu lator (Marne la V"llcc, Francc) witll a UVA irradi, .• tion source (Vi lbe r Lourmat T-20.L, 365 nm , no UVU, no UVC emission). The energy was measured with the Vilbcr Lourmat radiometer, RMX-365/312, a microprocessor programmable in energy (milliJ o ules) with measurement of 6 irrad iations per second.
One mg of 5-mcthoxypsoralen per ml (kindly provided by Bergaderm Laboratory, Rungis, France) in 70'X, ethano l was diluted in growth medium to the appropriate concentrations to ob tain a fina l concentration ofless than 0.01 "I., e tl"'nol. Psoralen so lu tions and treated cell s werc protected from light.
Two days befo re photo treatment of ce ll culturcs , growth mcdium was removed and rep laced by MCDU 153 supplemcnted with 30 fLg/ml bPE and 1 % feta l c:Il f sc rum (basa l medium) for thc phototreatmcnt period in order to avoid poss ible nonspecific effects of the mitogens 12-0-tetradecanoylphorbol- '13-acetate and forskolin . T hrce hours before the first irradiation, 5-MOP (10 - 0 M) w as added to the medium :lI1d was prescnt during the whole phototreatmellt period. The ce ll s were irradiated in PBS to avoid formation of mcdium-derived toxic photoproducts induced by UV cxposure. All of the culture di shes (with the plas ti c lids) were placed into the UVA irradiation cham ber. Each dose was corrcctcd by a factor set given by thc manufacturer to takc into account the refractive index of the plastic cul ture dishes and the PJ3S fi lm on the cell s. Immediately after irradiation, the PBS was replaced with basa l medium containing 1.0 - 0 M 5-MOP. C ultures were irradiated dail y, and responses were measured 24 h after the last exposure. Unirradiated control cells were maintained in PBS at room temperature during the time of phototrea tlllent. S91 ce ll s wcre irradiated according to the same procedurc.
THE JO URNAL OF INVESTIGATIVE DER.MATOLOGY
DNA Synthesis Assay Immediately after PUV A irradiation, medium of contro l or irradiated cell s was replaced by 1. ml of basa l medium containjng 5 fLC i / ml [llIcI"yl-' H]thymidine. After 24 h of incubation, cells were washed three times with PBS, and 5% cold trichloroacetic acid was added for 10 min at 4°C . Cell s were then washed twice with 90% ethano l and lysed in 0.1 N NaOH. R adioactivity was counted by liquid scintillation and norm:.lized to the amount of protein as determined by the method o f Lowry cl al (1951) . The ce ll numbers were determined by the trypan blue dye exclusion assay in parallel.
Melanogenic Activity Assays and Melanin Synthesis T hc tyrosine hydroA"lase activity of tyrosinase (O-diphenyl: O 2 oxidoreductase, EC 1.10.3.1) in li ving cell s was estimated from the amount of ' H 20 released into thc mcdium in 24 h based on the conversion of L-3' ,5' -[' H]tyrosine to DOPA according to an adaptation of Lotan and Lo tan (1980) of the methods of Pomerantz (1966) and Oikawa cl al (1972). Immediately aftcr thc last irradiation, 1 X 10" cpm/ ml of ['l-l]tyrosinc were add cd to the mediulll . Twenty-fouTs ho u rs la ter, tritiated water rel ease in the nlcdiulTl was mc:.sured by liquid scintillation counting. All data were corrected by subtracting thc amount Of'1-l20 formcd spontancously fi'om L-['H]tyros inc in growth medium without ce ll s during 24 h. R esults were normalized to cell number for each de termination in triplicate.
DOPAchrol11c tautonlerasc ac tivity assay was pcrfornled accord ing to Aroca cl al (1 990b). One lnut of OCT was defined as the amount o f enzyme that cata lyzes the transformation of 1 fLmo l of DOPAchrome per min at 30· C as measured by spectrophotometry at 308 nm. Each determination was done in tripl.i cate.
N ewly synthcs ized mela.uns wcre de termined after the last irradiation of the phototre"tmcnt as desc ribed by Hearing and Ekel (1976). 3T3 fibroblasts were used as negative control. Results were normaljzed to the cell nun,bcr for each assay , and dctcrnlinations were Illude in triplicate.
Expression of Melanogenic Proteins Fo r immunoblot analysis, cells wcre scraped fi'om the di sh and homogenized in 1% Triton Xl OO/phosphate buffer (0.1 M, pH 7.2) containing 1 mM phenylmethylsu lfonyl RUOl'ide , 10 fLg/m l aprotinin, and 50 mM NaF. 50 fLg o f the rcsulting sample proteins as detcrmined by the method of Lo wry cl " I (1.951) werc scparated under reducing condition(s) by sodium dodccyl sulfate-polyacrylamide gel electrophoresis (7.5% "crylamide) (Lacmmli . 1970) and elec troblottcd (Towbin el ai, 1979). The immunorcactio llS were pe rformed with antibodies directcd aga inst DOPAchrome tautomcrasc (PEP8) (Tsukamoto ct ",. 1992), T IU>-l (PEP1) Uimcnez el ai, 1991) (gifts of Dr. Hcaring), and tyrosinase (specific antise rum kindly providcd by Dr. Bouchard) (Bouchard cl nl , 1994).
Total ccllular RNAs were purified using the single stcp isolation method described by C homczynski and Sacchj (1 987). For northe rn blot analysis. total RNA (70 fLg) was fi'ac tionatcd on a 1% agarose gel and transferred to Nytran nylon fi lters as described by Maniatis cl " I ("1 982). A cDNA probe against human tyrosinasc, pIU-10HT2 (Taked:. cl "I, 1989) , was gcnero usly provided by Dr. Shibahara (Tohoku Univcrs ity School of Medicine, Scndai, Japan).
RESULTS
PUVA Enhances Melanogenesis in S91 and NHM Cells To d etermine 5-MOP conce ntration and UV A doses by whi c h melan ogene sis is maximally stimulated with min.imal cytotoxic efFect, w e exposed S91 cell s over periods rangin g from 1 to 10 d to 5-MOP con centrations ranging fi'om 10- . 0 M to 10- 5 M and to dail y UVA irradiation (0.1 to 2 jlcm2
) . Maximal stimulation of tyros in ase ac tivity with minimal toxicity (less than 20'1.. of cells death) occurred after 6 daiJ y irrad iations of 0.1 jlc m 2 in the presen ce of 10 - 6 M 5-MOP. A t im e-dependent increase of tyrosinase activity was observed after a la g phase of 2 d (Fig 1A). After 6 irradiations, a lO-fold stimulation of tyrosinase ac tivity and a similar in crease in melanin conte nt was found (data not sh own) . T his effect was n ot observed when S91 cells w e re tre ated with either UVA or 5-MOP (10 - r. M) a lone. Determination of the numbe r of cell per dish after PUV A treatment sh owed a 75'Yo d ecrease in cell numbe r afte r the 6th irradiation as compare d to untrcate d control c ultures (Fig IB) .
In addition to tyrosinase activity, we also examine d the efFect of PUV A on S91 OCT, a distal e n zym e in m e lanoge nic pathway . OCT activi ty was measured 24 h after the 6th irradiation ofPUVA treatment and showed no c hanges in basal activity (Table I).
Melanoc ytes w e re then exa mined for their capacity to respond to
PUV A treatment ;11 11;/1'0. PUVA parameters established for S91
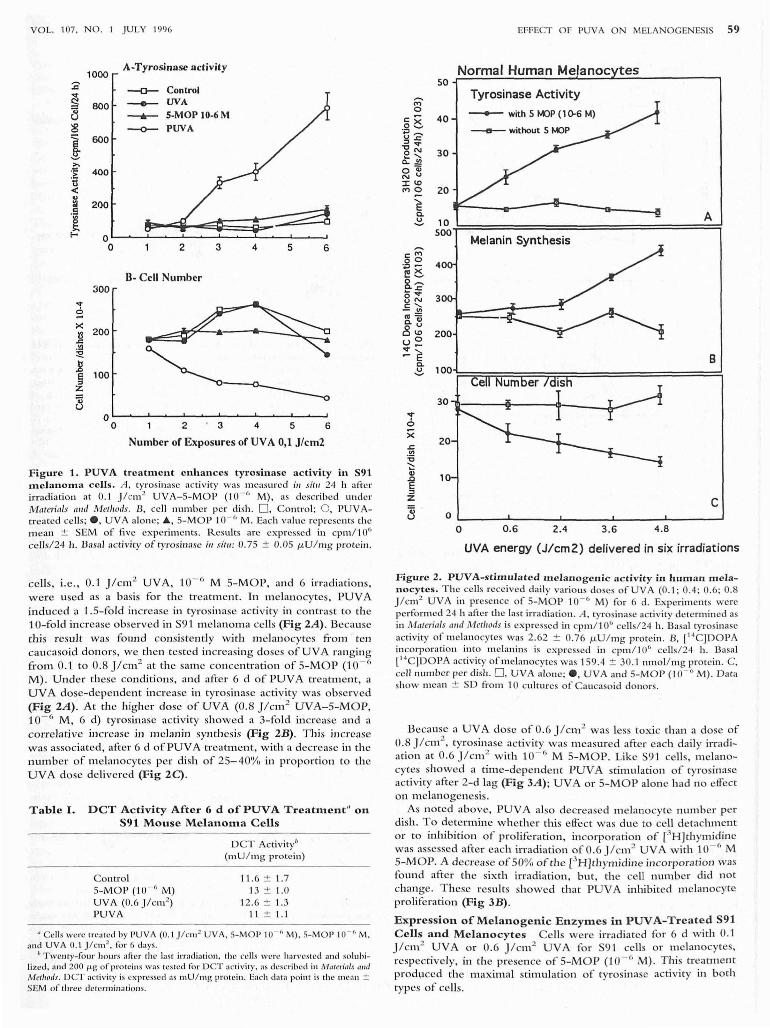
VOL. 107. NO.1 J ULY 1996
1000 A-Tyrosinase activity
:;:;---0-- Control
~ 800 - UVA "il ---.- S-MOP IO-6M u ~ ~ PUVA
I 600
>.
:~ 400 tl < .. :ri 200 .S [ f- 0
0 2 3 4 5 6
n- Cell Number 300
"'i :=! x
200 !3 .c .!!l :!:!. ]
100 § z 'ii u
0 0 2 3 4 5 6
Number of Exposures of UVA 0,1 J/cm2
Figure 1. PUV A treatment enhances tyrosinase a c tivity in 591 melanoma cells. A. tyrosinase activity was measured ;11 s;tll 24 h afte r irradiation at 0. 1 J /cm 2 UVA-S-MO P (10- (' M) , as described under Materials a",1 Methods. B , cell number per di sh. D, Contro l; O . PUV At reated ce ll s; e, UVA alone; ., S-MO P 10- 6 M. Each value represen ts the m ean ± SEM of fi ve experiments. R.csults are expressed in cpm / 10" ce/J sl24 h . Dasa l acti vity of tyrosinase ill Jilll : 0 .75 ± 0.05 f.LU / l11g protein.
celis, i.e ., 0 .1 jlcm2 UV A, 10- 6 M S-MOP, and 6 irradiations, were used as a basis fo r the treatment. In melanocytes, PUV A induced a 1 .S- fo ld increase in tyrosinase activity in contrast to the l O- fo ld increase o bserved in S9 1 melanom a cells (F ig 2A ). Beca use this resul t W ,IS found consistently with m elan ocytes £i'om ten cau casoid donors, we th en tested increasing doses o f UV A rang ing from 0.1 to 0 .8 j /cm 2 at the sam e concentration of S-MOP (1 0- 6
M) . Under th ese conditions, and afte r 6 d of PUV A treatment, a UV A dose-dependent increase in ty rosinase activity w as observed (Fig 2A ). At the higher dose o f U VA (0.8 jlcm 2 UV A- 5-MO P, 10- 6 M , 6 d) tyrosinase activ ity show ed a 3-fo ld increase and a correlati ve in crease in melanin synth esis (Fig 2B). T lus in crease was associated , after 6 d ofPUV A trea tm ent, with a decrease in th e number of melanocytes per dish o f 25- 40% in propo rtion to th e UV A dose delivered (Fig 2C) .
Table I. DCT Activity After 6 d of PUV A Treatment" on S91 Mouse Melanoma Cells
ontrol S-MO P (1 0 · " M) UVA (0.6 J /cm 2
)
PUVA
DCT Activity" (I1l U / mg protein)
11 .6 ± 1.7 13 ± 1.0
12.6 ± 1.3 11 ± 1.1
" Ce lls were treated by PU VA (0 .1 J /CIII2 UV A. 5-MOP 10- (' M) . S-MOI' 10 - (' M. and UVA 0. 1 Jlcm2
• fo r 6 days. IJ Twenty-four ho urs after the 1.!st j rr;ldi:l ti o ll . th e cells were harvested and solu bi
lized. and 200 J..I..S of procci ll s was tested fo r DeT activilY. as dcscl'ibcd in Nlntcn'nls IIIU/
t\lldl/Ods . DCT ;Ic ti v it)' is ex pressed as IIIU / m g prote in . Each datn point is the IIl Cr\ l1 :t SEM of three dctcfl1liJwtio IJ ....
..r 6 X .s:. '" :c ~ Q)
~ :l Z
"ii u
EI: FECT OF PUVA ON MELANOGENESIS 59
Normal Human Melanoc tes 50~~~~~~~~~~~~~-----------'
40
30
200
Tyrosinase Activity
--- with 5 ~p (1 ()'6 M)
Melanin Synthesis
A
B 100~~~~T=~~r=================~
30 1r-----t----~~----~-
20
10
c 0
0 0 .6 2.4 3 .6 4.8
UVA energy (J/cm2) delivered in six irradiations
Figurc 2 . PUVA-s timula t c d m cla nogcnic activity in hum" n mela nocytcs . T he cells received daily va rio us doses of UV A (0 .1 ; 0.4 ; 0.6; 0.8 j /cm 2 UVA ill presence of S- MOP 10. 6 M) fo r 6 d. Experiments were perfo rmed 24 h after the last irradiation . A, tyrosinase activity determined as in Materia ls alld M ethods is expressed in cpm/10" cells/24 h . Basal tyrosinase activity of melanocytes was 2.62 ± 0.76 f.LU / mg protein . B, r '4C)DOPA incorpora tion in to melanins is expressed in cpm/ l06 cells/24 h. Basal [" 'C]DOPA activity of mclanocytes was 159.4 ± 30.1 nmol/ mg protein. C, ceU number per d ish. D, UVA alo lle; e . UVA and S- MOP (10 · " M). Dar" show mean ± SD fro m 10 cul tures of Caucasoid donors.
Because a UVA dose of 0.6 j /cm 2 was less toxic than a dose of 0.8 j /cm 2
, tyrosinase activity was m easured after each dai ly irradiation at 0.6 j1cm 2 w ith 10 · /; M 5-MOP. Like S91 celi s, melanocytes showed a time-dependent PUVA stimulation of tyrosinase activity afte r 2-d lag (Fig 3A ); UVA or 5- MOP alone had no effect on melanogen esis.
As noted above, PUVA also decreased m elanocyte nu m ber per dish . To de termine whether tlus effect was due to cell detachment o r to inhi bi tio n of pro li feration , incorpo ration of [3H ]thymidine was assessed after each irradiation ofO.6j/cm 2 UVA with 10 . 6 M 5-MOP. A decl'ease of 50% of th e eH]thymid ine incorporation was fo und after the sixth irradiation , bu t, th e cell number d id not change. T hese results showed that PU VA inhibi ted melanocyte proli feration (Fig 3B) .
Expression of Melanogenic Enzyme s in PUV A-T r eated S91 Cells a nd M el anocy tes Cells were irradiated for 6 d with 0.1 j /cm 2 UV A o r 0 .6 j1cm 2 UVA for S9 1 cells o r melanocytes, respectively, in the presence of 5-M OP (10 - 6 M). T his treatm ent produced th e max;mal stimulation of tyros inase activ ity in both types of cells.
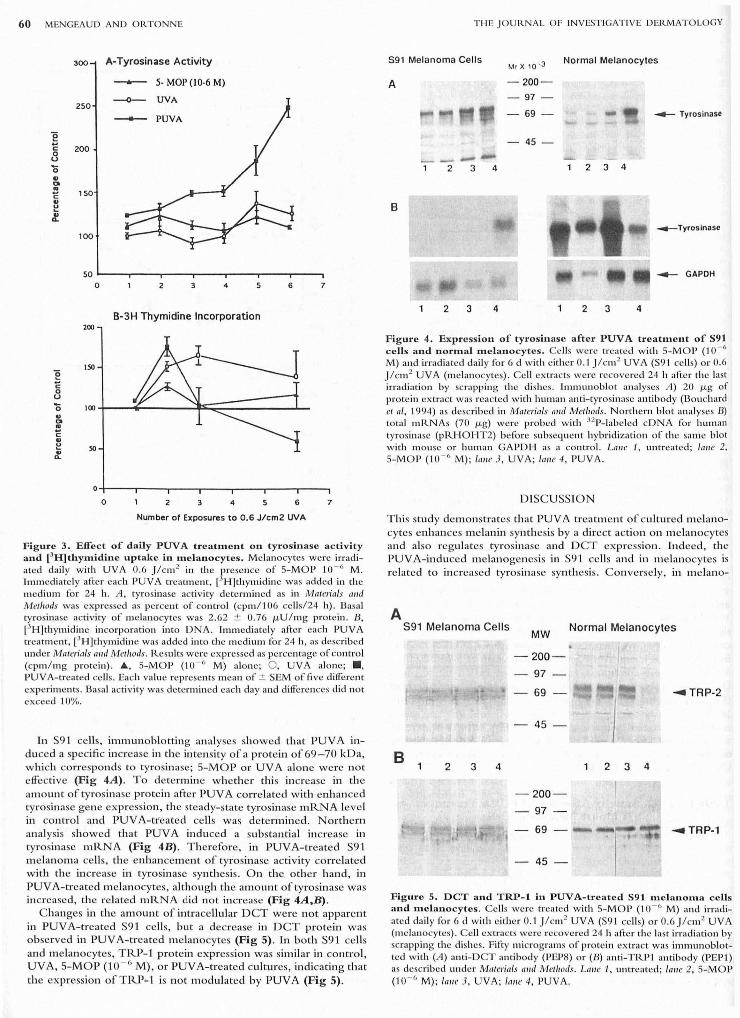
60 MENGEAUD AND OR.TONNE
g c 8 '0 ., CI !! c ., e .f
e c o u '0 ., CI
! c ., e ~
300
250
200
150
100
50
A-Tyrosinase Activity - 5- MOP (10-6 M)
0 4 6
B-3H Thymidine Incorporation
o 2 3 4 6
Number of Exposures to 0.6 J/cm2 UVA
7
F igure 3. Effect of daily PtJvA treatment 011 tyrosinase actIvIty and [JH]thymidine uptake in melanocytes . Melanocytes were in·adiated daily with UVA 0.6 Jlcm 2 in the presence of S-MOP 10- 6 M. Immediately after each pUVA treatment, l'H]th ymidine was added in the medium for 24 h. A , tyrosinase activity determined as in Materials (///(1 Methods was expressed as percent of contro l (cpm / l06 ce ll s124 h). Basa l tyrosinase activity of melanocytes was 2.62 :±: 0.76 J.LU / mg protein . B, [JH]thymidine incorporation into DNA . Immediately afcc r each pUVA treatment, ['H]thymidine was added in to the medium for 24 h, as described under Materials alld MetllOds. R.esults were expressed as percentage of control (cpm/ mg protein). A, S-MO P ('10 - " M) alone; 0 , UVA alone; ., PUVA-treated ce ll s. Each value represents mean of:±: SEM of five dilfereJlt experiments. Basal activity was determined each day and dilferell ces did not exceed '10%,
In S9 1 cells, immuno blottin g an alyses sh owed that PUVA induced a specific in crease in the intensity of a protein of 69 -70 kDa, w hich corresponds to tyrosinase; S-MOP or UV A alone were n ot e ffective (Fig 4A). To determine w heth er this in crease in the amoun t of tyrosin ase protein after PUV A correlated with enhanced tyros in ase gen e expression, the steady-state tyrosinase mRNA leve l ill control and PUVA-tr'eated ceUs was d etermin ed. Northe rn ana lysis showed that PUV A induced a substantial increase in tyrosillase mRNA (Fig 4B) . T h erefore, in PUV A-treated S91 melanoma ceUs, the e nhancem ent of tyrosinase ac tivi ty correlated with th e increase in tyrosinase synthesis. O n th e oth er hand , in PUV A-treated melanocytes, although the am o unt of tyrosinase was increased, the re lated mRNA did not increase (Fig 4A,B) .
C hanges in th e am o un t of intracellul ar DCT were not appare nt in PUYA-treated S91 ceUs, but a decrease in DCT prote in was o bserved in PUV A-trea ted m e lan ocytes (Fig 5). In both S91 cells and melanocytes, TRP-1 protein express ion was simil ar ill contro l, UV A, 5-M OP (10 - 0 M) , or PUYA-treated cultures, indicating t hat the expression of TRP~ 1 is not m o dulated by PUY A (Fig 5) .
T H E JOURNAL OF INVESTIGATIVE DERMATOLOGY
591 Me lanoma Cells M, x '0 .3 Normal Melanocytes
A - 200-
- 97
69 ..- Tyrosinase
- 45 -
'--1 2 3 4 2 3 4
B
~Tyrosinase
..- GAPDH
2 3 4 2 3 4
F igure 4. Expression of tyrosinase after PUVA treatment of S91 cells and normal melanocytes . Cells were treated with S-MO P (10 - 6
M) and inadiated daily for 6 d with either 0.1 J/c m2 UVA (S9 '1 ce lls) or 0.6 J/cm2 UVA (melanocytes). Cell extracts were recovered 24 h afte r the last irradiation by scrapping the dishes, Immunoblot analyses 11) 20 J.Lg of pro tein extract was reacted with human anti-tyrosinase antibody (Bouchard et ai, 1994) as described in Materials m,d Met/r ods. Northern blo t analyses B) tota l mR.NAs (70 J.Lg) were probed with 32P-labeled cDNA fo r human tyrosinase (p lU-I O HT2) before subsequent hybridization of the same blot w ith 111 0 ll SC or hunlan GAPD H. as a contro l. Lalle I, untreated; lalle 2. S-MOp (10 - 6 M); lalle J, UVA; larr e 4, PUVA.
DISCUSSION
T his study d e m o n strates that PUY A treatment of c ulture d melanocytes e nh ances melanin synthes is by a direct action o n m elanocytes and a lso regulates tyrosinase and DCT expressio n . Indee d , the PUY A-induced m e lan ogenesis in S91 cell s and in m e lanocytes is re lated to in creased tyrosinase synthes is. Conve rsely, in m e lano-
A S91 Melanoma Cells
B 2 3 4
MW
- 200 -
97
69
- 45-
- 200-
97
69
- 45-
Normal Melanocytes
~TRP-2
2 3 4
~ TRp·1
F igure 5. DCT and TRP-l in PUVA-treated S91 melanoma cells and melanocytes. Cells were treated with S-MOP (10- (' M) and irradiated dail y for (j d w ith either 0.'1 J/cm2 UVA (S91 cell s) Or 0.6 J/c 11l 2 UVA (melanocytes). Cell extracts were recovered 2'1 h aftcr the last irradiation by scrapping the di shes. Fifty micrograms of protein extract was iml11unoblotted with (A) anti-DCT antibody (PEP8) or (B) anti-TRPl antibody (PEP1) as described under Materia ls ami Metlrods. Lalle ./, untreated; larre 2. S-MOP (10 - 6 M) ; lalle 3, UVA; larre 4, PUVA .

VOL. 107. NO. J J ULY 1996
c ytes but not in S91 , OCT protein expression is substantially decreased by PUVA. Finally, Tl~-l expressio n is not m od ifi ed in e ither cell type.
As ha s been described for UVB (Abe rdam cl ai, 1993 ) , PUVA inhibited prolife ration of cu ltured melanocytes . T IllS result, in view of the incl'ease in the number of epid ermalmelanocytes observed ;11 11 ;110 with PUVA (Zaynoun cl ai, 1977; B10g and Sza bo, 1979; Zelickson el ai, 1979) , suggests that f.1cto rs extel'llal to m elanocytes may mediate this effect. Indeed, 11 0 nirrad iated and UV-irradiated keratinocytes in culture release growth facto rs such as basic fibroblast growth fa ctor (Halaban el ai, 1988) and cndothc lins (Yada el
aI, 1991) , which are known to stimulate me lanocyte proliferation. T he refore, the possibility exists that ;11 11 ;110 UV irradiation of e pide rmal keratinocytes induces a similar effect, resulting subsequently in melanocyte proliferation (Jmokawa and Motegi, 1993; Archambault and Gilchrest, 1995) . T hu s, the Inck of UV-induced proliferation of melanocyte ill lIilm may be the result of the absence of keratinocytes in the culture.
It is well established that cuJtured melanocytes isolated from their natural environment respond to UVB irradiation by in creasing melanin produc tion (Fri cd m an n and Gilchres t, 1987; Aberdam cl ai, 1993). Here, we report that PUV A treatment enhan ces melanin synthesis by a synergistic e ffect of psoralen andUV A. O ur model sys tem is based o n repeated dail y UVA irradiatio n of m elanocytes and S91 cells. A significant UV A dose-dependen t increase in tyros ina se activity and m elanin synthesis occur afte r 6 d of PUVA treatment. Compared to S91, melanocytes require higher doses of UV A irradiation to stimulate m elanogenesis, and the in crease in their tyrosinase activity remains proportionally lower. A similar relative refr,lctorin ess of m eJanocytes has also been observed after UVB stimulation (Friedmann and G ilchrest, 1987) and m ay result from a hig her basal tyrosinase activity in m elanocytes, m aking these cells less sensitive to PUV A than S91 cells. Diff:erel1ces in regulatory mechanisms be tween the two ceU types may also account for this observation . Pre vious studies of ;11 1I ;lm PUVA treatment with a single low dose of UVA (ran gin g fro m 31 to 124 mJ /cm 2
) in cultured m elanocytes grown in th e presence of mitogens and melanogenic inducers (CT, IBMX) (Kao and Yu , 1992) , showed a moderate but non-UVA-dependen t increase in tyrosinase activity. T he absence of melanogenic inducers and m e lanocyte mitogens in our culture system during PUV A treatment m ay account for the fact that higher UV A doses and repeated daily irradiation we re required to obtain a significant stimulation of m elanogenesis.
Several hypotheses have been put forward to understand the sp ecifi c tyrosinase-rela ted m echanism s responsibl e for the stimulation of m elanogenesis: (i) an increase ill the rate of de 110110 synthesis of the enzyme (Halaban el ai , 1984; B urchill el ai, 1988; Burchill el ai, 1989); (ii) activatio n of immature pre-existing enzyme (Wong and Pawelek, 1975; FuUer el ai, 1987) and lor (iii) the removal of a tyrosinase inhibitor (Halaban el ai, 1983) . O ur results show that in PUV A-treated S91 melanoma cells, these m echanisms are not mutually e xclusive. Indeed, analysis of tyrosinase mRNA and protein 24 h after the last PUVA irmdiation revea led an increase of the de ilOIlO syn thesis of tyrosinase . Furthermore, th e fact that tyrosinase activity increased afte r PUVA treatment, proportionally more (10-fold) than the total amount of the enzym e (2-3-fold) indicates that, in S91 cells, activation of a pool of pre-existing tyrosinase or rem oval of a melanogenic inhibitor m ay account for the enhanced tyrosinase activity. III melanocytes, the increase in th e amount of tyrosinase was not associated with any detectable increase in the corresponding mRNA level. An explanation for tlus apparent discrepan cy may be til e instabi li ty of tyrosinase mRNA . A lso, an increase in tyrosinase mRNA may occur early after PUV A treatment, but, beca use of rapid turnover of the tran script, may no lo nger be detectable 24 h later when o ur analysis W,IS performed. To unravel these hypoth eses, further studies such as de termination of mRNA stabi.lity are required . Finally, the finding of an increased amount of tyros inase in PUV A-treated m elanocytes is in agreem ent with previous studies, w hich dem onstrated that PUV A therapy increases the amount of tyrosina se in human skin (Burc1ull c( ai,
EFFECT OF PUVA ON MELANOGENESIS 61
1990) . Our results show sp ecifically that PUVA, like UVB irradiation (Aberdam el ai, 1993) or other m elanogenic inducers such as MSH o r IBMX (BurchiJl e( aI, 1988; Hoganson e/ aI, 1989), in volves, at least in part, in creased de 11 0110 synthesis of tyrosinase to enh ance m elanogenesis in S91 cells and m elanocytes.
R ecent advan ces in the fi eld of m elanogenesis have sh own that, in addition to tyrosinase, TRP-1 and D CT are involved U1 tl1e biosynthesis of m elanin . Altho ugh TRP-1 was the fIrst m ember of the tyrosinase fa mily to be cloned , its ca talyti c fun ction rem ains elu sive and controversial. Tn tlus study, we examined its expression after 6 d of PUV A treatment and did not observe any variation in the am o un t ofTI~-1 in me hmocytes or 591 cells. This finding is in keepin g w ith the observation that UVB irradiation does not modulate Tl~-1. expression (Abdel-M alek el ai, 1994) in cultured m elanocytes. On the contrary, a correlation b etween TRP-1 expression and m elanin synthesis was found (Orlow cl ai, 1993) in melanoma cells (B16F10 , S91) stimul ated by a-MSH. Interestingly, we found that PUV A treatm en t m arkedly decreased tile amount of DCT Ul m elanocytes, but in S91 cell s, the level of this protein was not m odifi ed by PUV A .
A survey of repol'ts shows that regulation of DCT has been studi ed in detail after cx-MSH stimulation, but the results are conflicting. It ha s been demonstrated both that MSH Ulcreases D CT activity ofB16Fl 0 and S91 m elanoma cells with no change in the syn the tic rate of the enzym e (Orlow el ai, 1990; Kameyama el
ai, 1993) and that MSH decreases DCT activity (Martinez-Liarte e(
ai, 1993) . In normal ceUs, recen t reports claimed dlat a decrease of DCT protein occurred in m elan ocytes afte r UVB irradiation (Romero el ai, 1.994). Taken together, o ur findings support a negative corre lation between DCT expression and m elanogenic activity Ul pigment cells. The reduced expression of DCT by PUV A is also Ul agreement w ith this hypothesi s, suggesting that, during m etabolic stresses such as UV irradiation or prolonged hormo nal or phamncologic treatment, the e umelanogenic pathway is preferentially stim ulated, lea ding to increased synthesis of dark mehmins. Indeed, a redu ction Ul DCT would result in a decreased tau tomeriza tion of DOPA chrome UltO DHICA. H ence, DOPAchwme would be spontaneously converted in to DHI, which is involved in the formation of dark melanin polymers, the more photopro tective o nes (Aroca et ai, 1992). N evertheless , to confiml tlus hypothesis, complementary studies an alyzing the chemical structure of m elanins produced il 1 PUV A- treated pigment cells are needed.
Taken togeth er, these results show that, Ul m elanocytes as well as in murine me lano ma cells, the regulation of PUV A-induced m elanogenesis is directly re lated to an increase of tyrosinase synthesis. They also support the idea that DCT regulation after PUV A trea tmen t differs accordulg to the cell type. In melanocytes, the DCT protein is down-regulated after PUV A, whereas Ul neoplasti~ S91 cells such regulation is apparently lost. On the other hand , TRP-l expression is n ot reguiated by PUV A treatment in bo tll m e lanocytes and S91 cell s. These findings demonstrate that a coordinated regulation of syn th etic rates of tyrosinase, DCT, and TIU)-1 observed ul vario us pig me n ted cell lines (B16FI O, JB / MS) (Kam eyama ef ai, 1. 993 ) does not occur after PUVA. In this situation , the three enzym es are regulated indep endently in S91 cells as well as in m elan ocytes.
vVc Ihallk Dr. Ii. J-lclIrillgji"'prollidillg rhcpolyr/oll al lI lIlibodies PEP1 , PEP8; D r. B. BO llchard jill' Ih e 1IIIII,all rYfOsillasc flI, ribody; flI,d Dr. S l,ilJalr ara jar r)'fOsi rrase
cDNA probe pRHOHT2. '~/e Ilrfll,k D,:,. G. Mellcgllzzi, E. Abana"" arrd C. Ro", crojorcrirical reanillg ojrlris ", allrrscripl as ",cll as C. M;rrglrelli jorfll1,"ork. Ii. Merrgealld lIIas sllppo,1ed hy Bergader", Laharalo,}, (Rllrrg is, Frallce) . We Ifwrrk also uLa lig ll l' l\Jatiollnle Frrl11(aisc CO l1fre Lc Ca llcer," ((Associa tion Vn iucrc Ie Melarr,, ", c, " olld GEFLUC.
R EFER ENCES
Abdcl- Molck Z. Swope V. Smolol'o D . l3abcock G. Dawes S. Nordlund] : Analysis of the UV indu ced m e lanoge nes is :lnd g row th arrest of hulIlrm I11c )anocytcs. P(CI1l CIIf
Cel l Res 7:326 - 332, 1994

62 MENGEAUD AND ORTONNE
Abcrdillll E. R..omcro C. Ortonne JP: Repeated UVB irradia tions do not have the same potential to pro m ote stimulation of mc l:'Hlogcllcs is ill c ultu red 110rmal 11\1111 :111 melanocytes. ) Cdl Sci 106:10 15-1022. 1993
Archambault M. Gi lchrest BA: Kcrati nocytcs and fibrob lasts in a human skin cqui v:llent model enhance mchll1 0cytc survi val :lI1d me l.min synthesis after ul traviolet irradia tion.) [,/IIeSI De"'lIIlol '104:859 -867 , '1995
A roca p. Garcia-Dorron Je. Solano F. Loz:\I1 o JA: l'tCgUliltiOIl of m:llllmaHan m elanogenesis. I: Partial purificati on .md charll ctcriz atioll of:l D O PA chro l11c convertin g fa ctor: DO PAchrome tautomerase. /3iochilll BioIJI,)'s Aan 1035 :266-275 , 1990a
Aroell P. Sol:1 l'1 o F. Garcia-Borron JC. LOZOl Il OJA: A new spcctroplloto11lctr ic assay fo r DO PAchro rnc tautOI1lCTn sc. J BioclwlII B;ol'/I)IS iVlcllwds 2 1 :35-46. 1990b
Aroca P, Solan o F, Salin as C, Garcia-Bo rron J C, Lozano J A: Regu lation o f the fonal phase o f mammalian mclan ogcncsis: thc ro lc of DO PA chro mc tautOll1crasc and the ra tio hetwccn 5,6-dih ydro:x),indo lc-2-carboxylic acid and S.6-dillydroxyindole . EliI') /3iorhell/ 208:155-163 . 1992
Aroea P, Urabe K, Kobayashi T, T suka moto K. Hearing VJ: Melanin biosynth esis patterns fo llow in g hormonal stimulation . ) /3iol Chell/ 268:25650-25655, 1993
Blog FB, Szabo G: T he effect of psora len and UVA (PUV A) o n ep ide l111il1 melanocytes of the tail in C57 BL mi ce. ) /II"esl De,."lIIlol 73:5 13-533 . 1979
Dardin F. Marz:lno C . Gatto C . C"I'i:lssarc F. R o dighicro P. B:l ccichc tti F: 4 .6 . .11'Trimcthylangelic in induccs intc rstrand cross- links in mammalian cc ll DNA. J P/lOloch"", P/lOlobi,,1 /3 26: 197-20 1. '1994
Uo uch:lrd 13. Vijayasaradhi S, Ho ug hto n AN: Pro du ction and c1wf:lc tcrizatioll of antibo dies against hlllllnn tyfOSi11:lSC. J IlI l/CS t DeYIIUlto/ 102:291-295, 1994
Burchill SA. Mark s JM , T hody AJ: T yrosin ase synth esis in diRerent skin types and the clfccts of a-mclanocytc-stimula til lg ho rmo nc and cycl ic AMP.) fll" est DCI'II/ fllol
95 :558-56 1, 1990 Burchill SA. Virden R. Full er BE, Thody AJ : R egul ation of tyrosinase synthesis by
a -melanocyte-stimulating ho rmo nc in hair fo ll icul nr I1lc lanocytcs o f thc I'nOllsc. ) Elldocr;,,,,1 116:17-23 ,1 988
Burchill SA, Virden R., Thod )' AJ : R egul ation of tyrosin ase synthesis and its processing in the hair fo llicular IIIc lanocytes of the mOllse during cumc l:lIlogcncsis and phaeomelanogenesis. ) 1""csl De,.,,,nlol 93:236-240, 1989
C hatterjee PK, C antor C R.: Photochemical production o f psora len-DNA mo noadducts capablc of subsequcnt photocross lin king. N llcldc Acids Res 5:3619-3633, 1978
C ho mczynski P, Sacchi N : Single-step mcthod o f I"tN A iso latio n by acid gU ~lIlidinium
thjocya na tc-pheno l-chloroform ex traction . A II/lill /3iorlt cIII 162: 156-159. 1987 C undari E. Averbeck 0: 8-l1lct11oxypsora lcn- pho toindu ccd DNA crosslin ki'l g as
determin cd in YC:lst w hen diffe rent rei rradiation conditio ns. R elation w ith gcne ti c effects. PItOlocl,elll pltoleM,,1 48:3 15-320, 1988
Eisin ger M, Mnrko 0: Selective pro li feratio n of normal human II1c ):l l1 ocytes;II IIil1'o in thc prcscnce of phrobol ester und cho le ra toxin . Proc N(/ tf Acnd Sci USIi 79:2018-2022, 1982
Pitz)Jntrick TD: Thc psornle n story: photochc l'llothcrapy and pll otocl lcrno pro tec ti o n . In : Fitzpa tri ck TB, Forl ot P. Pathak MA , Urbach F: (cds.). Psornle"s: Pnsl, PreS""1 ami Fu/llre oj PllOWcll clll opnJlectioll alld a tIlt:,. J3i"h~<: ;caf ACli"i/;cS. Jo hn Libbey Euro'tcx t, Frnncc, 1989 , pp 5-'10
Friedmann PS, Gilchrcst B: U ltr:lv io le t radi:ltio n d irectl y induccs pig mcll t productio n b)' cultured hUlllan mel anocytes. ) Cd l PIt),si,,1 133:88-94, 1987
Fullcr BB , Lunsford jB. Jman OS: a-Melanocyte-stimul atin g ho rmo nc rcgulntio n of tyrosinase in C io udlllan S9 '1 mo use m elano ma cell culturcs. J Bioi CllcIIJ 262:4024-4033 . 1987
Gschn ait F. Brc nner W, Wolff' K: Ph otoprotective CUCCI' of" psor:1l cn-UVA-induced tan. Arclt Dermnlol Re .• 263 :1 81-1 88.1 978
Halaban R , Langdon R, Birchall N , C uono C. Baurd A, Scott G, Mocllmann G , McG uire j : Basic fi brobl ast growth r.1c lo r from human kcratinocytcs is " nHtura l mitogen for melanoc), tes. ) Cell /3iol 107:161 1- 16 19, 1988
Halaban fl. Mocllman G: Murinc and human b locus pig mentatio n gcnes cncodc :.1
glycoprotein (gp75) with cata lase activity. i'mc N nll Arnd Sd USA 87:4809-4813, 1990
Halaban R. Pomerantz SH, Marshall S. Lambert DT, Lerne r AB: R egulation of tyrosin:tsc in human mclanocytcs g rov,rrl in culturc.) C dl Bioi 97:480- 488. '19H3
J-Ialaban fl, Porncrantz SH , Marshall S. Lcrncr AD : Tyrosinase acti vity and abundan ce in C10udman melanoma cells. Arrlt l3iorhcm /3i"ph)'s 230:383-387. 1984
Hearing Vj , Ekcl T M : M ammalian tyrosinasc. A comparison of tyros in c hydroxylatio n ,md melanin formation. Dioc/wIIJ 157:549-557, 1976
Hearing VJ , Nicholson JM . Mon ta gue PM , Ekel T M , Tomecki KJ: Mammalian tyrosin:1 sc stru ctural and fun ctio nal intere lationship of isocllzy.-nes. Di(Jritilll /3ioph),s Ac,n 522:327-339. 1978
Hoganson GE, Ledwitz-Rigby F. Davidson RL, Full er BB: R egulation of tyrosinase mR.NA levcls in mo usc me lanoma ccll s by Ill c l:1 nocytc-st imul ating horlllo ne and cyclic AMP. Somn,;c Cell M"I Cellcl 15:255-263, 1989
Imokawa G, Motegi I: Skin o rg;lI1 culturc mo del fo r examining ep idc rtlli11 melan ization.) I" "cs l DcrmnlOl 100:47-54. 1993
jackson IJ : A cDNA cncoding tyrosina sc-re lated protc in maps to thc brown locus in the mo use . Proc Nail Aend Sci USA 85:4392- 4396, 1988
Jimenez M. Tsuk:l llloto K. Hearing Vj : Tyrosil1:lScs from two dHfc rent lod arc ex pressed by normal and by tra nsformed melanocytes . ) /3iol C it"", 266: J 147-11 56, 199 1
jilllc ll cz-Ccrvcntes C . Soli1n o F, Ko ba yashi T. U rabc K. Hearin g Vj. Loz;lI1o jA.
T H E J O URNAL OF INVESTIGAT IVE DERMATO LOGY
Ga rcin-Bo rro n JC : A ncw cnzymatic fUB ction ill th e melanogenic piltltway. The 5.G- dihyd roxyi ndo lc-2-ctlrboxylj c acid ox idasc ilc tivity o f tyorsinasc-rela lcd protein- 'I (TR.pl}.) /3iol C ltelll 269: 17993- 18000 , 1994
Kamc yama K. Takcrtlur:1 T . Hamada Y , Sak:li C. Ko ndo h S. Ni shiya m:1 S. Urabc K, Hearing Vj : Pig lllc nt pro du ctio n in murine Ill cl:mo ma cclls is rcgulatcd by tyrosin ase, tyrosinase-re lated protein 'I (TIU'I), DO PAch rome ta lltornerasc (TR.P2), and a melanogeni c inhi bitor.) I" "CSI Der",nl"/IOO:126-'I3 1, 1993
Kao C H , Yu I-I S: Comparison of the effect o f 8-methoxypsoralen (8-MO P) plus UVA (PUV A) o n hum,lII Illc lanocytcs in v itiligo vul g:1ris and ill I/i(l"O. J 11"J(~s t Den"nlol 98:734-740. 1992
Ko bayashi T , Urabe K. Windcr A. jirncllcz- Cervcntes C . Imo knwa G. Brcwington T . Solano F, Garcia-Borron J C, I-Iearing VJ: Tyrosinase related pro tein I (TIU'-1) fUll ctions as a DH ICA ox idase in rncl"nin biosynth csis. EMBO J 24:58 18-5825, 1994
Korner A, Getting P: Synthesis i" "ilm o f 5.6-dih),droxyindo le-2-carboxylic acid by dopacliro tTl c convcrsio n fa cto r fi'o lll C io udlll;m 51) '1 mclanoma cc lls. J IIl11cst Dcr",nrol 85:229-231, 1985
Laeml11li UK: C leavage of stru ctural IJroteins durin g the assernbl), of the head of ba cteri ophage T4 . N nr"re 227:680-685. 1970
Lask illJD , Lec E. Yurko w EJ, Laskin DL. G:lll o MA: A possibl c mechani sm o fpsoralc ll phototoxicit), not in volvin g direct in teraction with DNA . i'm( N nll A((/(I Sri USA 82:6 158-0 '162. 1985
Lotan !'t, Lo tan 0: Stimulatio n of melanogcnesis in a human mclanoma cell lin e by re tino ids. Callrer Res 40:3345-3350, 1980
Lowry 0 1-1 , P....osc boug h Nj , F:lrr AL: Pro tein IllC:lSUl'e ll1Cllt with folin phcllolrc i'l gc llr. ) /3i,,1 C I/CII/ 193:265-275, '1951
Maniatis T, Frish EF. Sa11lbrook J: Analysis o f I~A . In : Irw in N (cd .) . M"/cCIIlnr C lolli"S: A LnIJoralcJI1' IVlallllal. Cold Spring I-IOIrbo r Labo ratory. Cold Spring Harbor. NY , Vol 2. 1982 , PI' 7.37-7.52
Martin ez-Liane JH , Solano F, Garcia-Borron J C, J ara JR, Lozano JA: lY- MSJ-I and othe r melall ogenic :lCti vators mcdiMc o pposite c lfects 0 11 tyros ina se and DO PAchr01llc t:tu tolllcrasc in ll16/ FI 0 mo use mclano ma cc lls. ) IlIl/cst D el'll/awl
99:435-439, 1993 N o rdlllndji . Le rn er AB: Vi tiligo. It is importa nt. A rch Demwlo/1 '18:5- 8 . 1982 O ikawa A, Nakayasll M , No hara M , T chell T: Fate of L-(3 ,5-' H}tyrosine in ce ll-free
extra cts and tissllc cultures of melanoma cell s: a ncw assay mcthod for. tyrosin ase in li vin g cells. A rch Bi"cJli' /I1 /Jjophrs '148:548-557, '1972
O rlow SJ . C ha krabort), AI<, Pawelek JM: R e tin oic acid is a IJo tellt inhibitor of induciblc pi gmentatio n ill murine and hamster m clall o ma ce ll lines. J llll/f'st
D,.,.,, /iliol 94:4 61 - '104, 1990 , O rlow SJ , Eoissy RE, Mo ran OJ, Pifko-Hirst S: Su bcellul ar distribution of tyrosinase
and tyrosinasc-relatcd protcin- l : implic;Il'io ns fo r Illc lanosomal biogenesis. J /" ,,"s, Der",nl,,1 100:55-64, 1993
O rtonll c JP, Moshcr DB. Fitzpa trick TB: Vilil(~o lllld O tJ, er H)'IWllldmlOsis of Hair alld S kill . Plenull1 Press, New York , 1983 , PI' 257-258
Po mer:mtz SJ-I : Tll c tyrosinc hydroxyl:l sc acti vity of man 1 III a Ii an tyros inase .) iJiol C hew 24 1:16 1- 1()8, I%6
lto mc ro C, Aberd:IIH E, larnic r C . Onol1l1 c jP: R c tin o ic ;Ic id as 11lo dulator of thc UVB-induccd IlI c l:mocytc dHlc rellti:1tio ll : ill volvcment of the melan ogenic cnzYll1es expression . ) Cell Sci 107 :1 095- 11 03, 1994
Szabo G, Dlog rn. Ko rnhausc r A: Toxic e fi'cct o r ultravio let: light 0 11 11lc liln ocytes: usc of animal mo dels in pig mc nt rcsearch. J N fl lf Cfll/eel' f /lS l 69:245-250. 1982
T ;lk cci:1 A, Tomit:l Y. O kinag;1 S. T :lga mi H. Shib:l hara S: Function:!1 analys is of the c DNA c ncod in g hUIll:1n tyrosin ase precursor. 13iorhcIII B;opll}IS R('s C Ol/1II11111
162:984 -990, 1989 Towbin 1-1 , Stachclill T, GordonJ : Electrophorctic transfcr of proteins fro l11 po lyacryl
:Imidc gels to nitroccllulose shec ts: proccdure and SOme appli c:ltioll s, Proc Natl Awl Sci USA 76:4350-4354. 1979
T suka ll1 oto K, J ackson IJ , Urabe K, Montague PM , Hearin g VJ: A second tyrosinasere lated protc in . TRJl2 , is :l melan ogenic enzymc tcrmcd D O PAchromc tall tOl1l erase. F.M/JO) 11 :5 '19-526, 1992
VU:1yasaradhi S, Bouchard D, Ho ughton AN : The mclanoma antige n gp75 is thc huma . homo logue o f the mo use b (brown) locus gene product. ) Exp Med 17'1: 1375-1380, 1990
Wilkin s M, Gilchrcst 13A . Szabo G. Weinstein n.... MacingT: T hc stimulation ofno nnal human melan o cy tc prolifcration ill "i/m by mc lanocytcs g rowth f:lctO I' (rol1l bovine I>ra in . ) Cell Plt),siol 122:350-36 1. 1985
W inder AJ. W ittbjer A, R osengren E, \1..o rsll1an H : T he m ouse brown (b) loclls protein has do p:1chrOll1 c t:llIto lllerasc activity and is IOCMCd in lyso so l11cs in tr:msrected fibrob lasts. ) Cell Sci 106:153-166, 1993
Wong G, P:1 welck j : Mel ano cyte stimulating: hormone promotes ac tiv<1 tio n of preex isti llg tyrosin asc mo lecules in C lo udmall S9 1 melano ma cclls. Nature (Lo"d" ,,) 255:644 -646 . 1975
Yada Y , Hig uchi K. Imo k:1 wa G: Thc clfcct of c ndothclin o n sig nal transduction and prolife ratio n of human Ille l:mocytes.) /3iol C ite", 266:18352-18357, 199 1
Zayno ull S, Konrad K, Gscl lllait F. WoHr 0 : T he pi gmc ntary rcsponsc to phorochemoth erapy. I lrln 0",.,,, Vellere,,1 57:43 1-440, 1977
Zelickson AS. Mottaz JH, Muller SA : Melanocytes changes following PUVA the rapy. ) A ll, Acnd Der",nlol 1 :422-430, 1979



![Comparison of the Effects of 8-Methoxypsoralen (8-MOP) plus … · 2017. 2. 1. · cutaneous disorders such as psoriasis and vitiligo vulgaris 11], and a great deal of effort has](https://img.dokumen.tips/doc/110x75/5ffa9e0278612f6e0174e1f1/comparison-of-the-effects-of-8-methoxypsoralen-8-mop-plus-2017-2-1-cutaneous.jpg)

























