Embed Size (px)
Citation preview

PURIFICATION AND CHARACTERIZATION OF STORAGE PROTEINS

3. PURIFICATION AND CHARACTERIZATION OF STORAGE PROTEINS
3.1 INTRODUCTION
3.1.1 Characteristic features of storage proteins
The major proteins of seeds display a number of
characteristic features which help distinguish them as
storage proteins.
1. They are synthesized at a particular stage during seed
development to be utilized during germination. These
proteins are therefore called storage proteins.
2. They tend to be rich in certain amino acids .like
aspargine, glutamine, arginine and proline.
3. They accumulate inside the cells in special vacuoles
termed protein bodies.
3.1.2 Classification of storage proteins
on the basis of solubility in different solvents, storage
proteins have been divided into four major classes
(Derbyshire et al., 1976). They are albumins (water
soluble), globulins (salt soluble), prolamins (alcohol
soluble), and glutelins (acid or alkali soluble). Globulins
are the major storage proteins of dicots while prolamins and
glutelins that of monocots. Albumins have been reported to
be present in varying amounts in both monocots and dicots.
68

3.1.2.1 Albumins
Albumins are water soluble proteins which have been
considered as the metabolic proteins. It has now become
evident that the albumins may have storage functions as
well. Furthermore, it has been shown that the albumins of
pea cotyledons are used as a reserve during and after
germination. (Murray, 1979). Two major albumin fractions
with molecular weights of 23,000 and 25,000, have been
reported from pea {Croy et al., 1984; Higgins et al.,1987).
Albumins frequently account for 30% to 40% of
extractable seed protein, as seen in pea and soybean.
Albumin fraction contains a higher percentage of methionine
and cysteine, which makes it nutritionally more important
(Murray, 1984). In addition, some low molecular weight
albumins, which have high methionine content, have also been
purified from various plants. Recently, a gene encoding
methionine rich albumin protein has been used for raising
transgenic Brassica plants (Guerche et al.,1990).
3.1.1.2 Globulins
Globulins are soluble in high salt buffers and they are the
major proteins of most legumes (Higgins, 1984) and Oat
(Brinegar et al., 1982; Higgins, 1984). They are separated
into different classes by cryoprecipitation, differential
69

salt solubility and ultracentrifugation. Most storage
globulins fall into two major groups with sedimentation
coefficients of 11S and 7S. Seeds of many plant species
contain both 11S and 7S storage proteins, although in most
cases, one or the other may predominate (Derbyshire et al.,
1976) .
To a large extent, the molecular structure of these
proteins are conserved among species. Soybean globulins are
the most extensively analysed storage proteins. They are
composed of both 7S and 11S globulins. 7S fraction contains
a group of heterogenous storage proteins called conglycinins
(Ladin et al., 1984) . 11S fraction contains, glycinin, the
major storage protein. It has a molecular weight of about
300 kDa and is composed of 6 to 7 subunits (Shotwell and
Larkins, 1989). Similarly in pea, the globulin fraction is
composed of legumin and vicilin proteins. Legumin has a
sedimentation coefficient of 12S and a molecular weight of
330-400 kDa whereas vicilin sediments at 7S and has a
molecular weight of 186 kDa (Derbyshire et al., 1976). They
are deposited in the seed in an insoluble form in protein
bodies.
3.1.2.3 Prolamins
constitute the major protein fraction in the Prolamins
monocots. They are rich in proline and glutamine but
70

contain less amounts of lysine, methionine and tryptophan.
They are hydrophobic proteins that are soluble in alcoholic
solutions. They exist as families of proteins and are
coordinately synthesized during seed development. They are
secreted into protein bodies.
3.1.2.4 Glutelins
The glutelin protein fraction is soluble in acid and alkali
solutions. Except in rice and sorghum, where glutelins
occur in protein bodies (Larkins, 1981), they are normally
present as an insoluble matrix containing starch grains
and protein bodies. In rice, glutelins occur as major
storage proteins and it constitutes 8 O% of the total
protein. Glutelins are similar to prolamins in their
biochemical properties.
3.1.3
plants
Similarities between storage protiens of different
The storage proteins of evolutionarily diverse groups of
plants show a lot of homology. Globulins occur in both
monocots and dicots. They exist as oligomers of six
subunits. Each subunit is composed of a disulfide bonded
acidic and basic polypeptides (Higgins, 1984). Common
antigenic determinants have been found among the globulins
71

of many members of the leguminosae (Dudman et al., 1975) and
the prolamins of wheat,barley and maize (Dierks-Ventling and
Cozens, 1982).
storage proteins exist as families of proteins rather
than as a single protein species. They are coordinately
synthesized during seed development. They may differ from
each other in their isoelectric points or molecular weight
but they are similar in their primary structure (Messing et
al., 1987).
3.1.4 composition of Amaranth protein
The major cereal crops are all grasses; Amaranth is one of
the few nongrasses to produce significant amounts of grain.
Amaranth grain contains about 16 to 19% protein, of which
albumins and globulins together constitute some 50% of the
total protein. Next major fraction is.the glutelins which is
about 30-33%. Prolamins, the major protein of cereals are
present in very low amounts (less than 3%). (Paredes-Lopez
et al., 1988).
3.2 RESULTS AND DISCUSSION
3.2.1 Sedimentation value of the major protein fractions
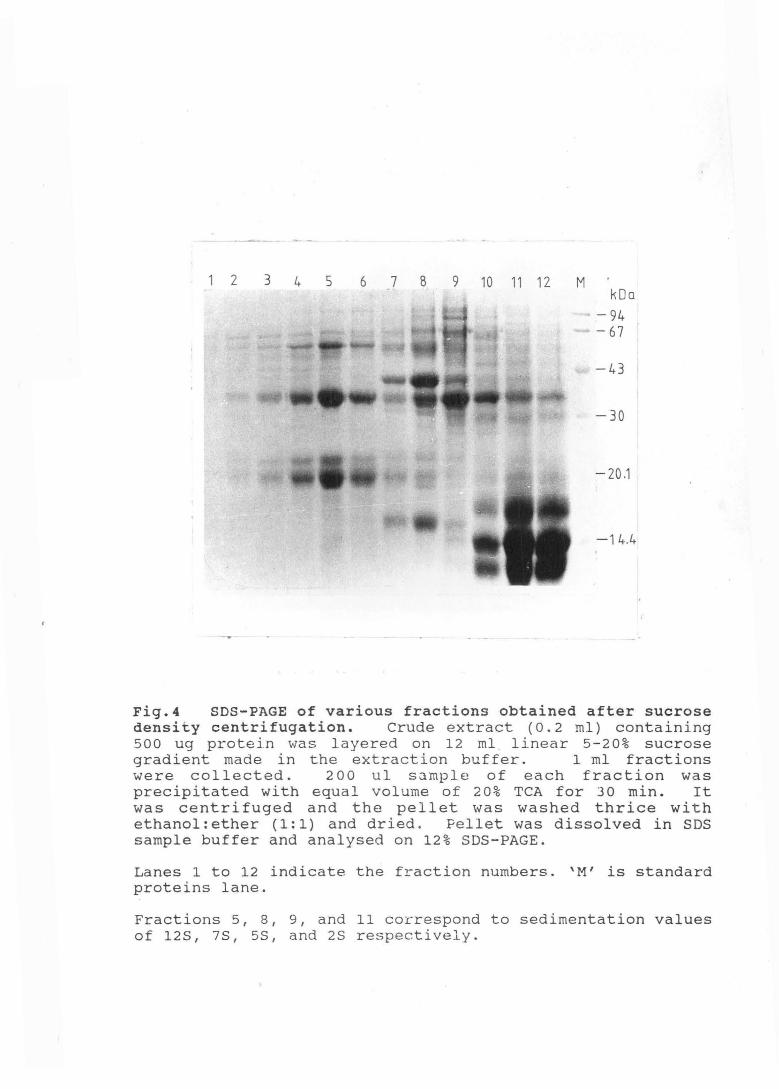
Sedimentation value of the major proteins present in
Amaranth seeds was checked on a 5-20% sucrose density
72

gradient. Four major protein fractions corresponding to
sedimentation values of 12S, 7S, 5S, and 2S were obtained.
2S fraction was most abundant (Fig. 4) . Globulins, th.e
major seed proteins of legumes,
7S and 11S ( Derbyshire et al.,
exist in two size classes,
197 6) • In addition some
small molecular weight proteins have also been found in most
of the plants tested (Youle et al., 1978, Murray, 1979, Croy
et al., 1984). When Amaranth albumin and globulin fractions
were separately checked on sucrose-gradient, it was seen
that the major proteins in the 11S and 7S fractions were
globulins, while those of 5S fraction were albumins. Both
the classes of proteins, albumins as well as globulins, were
found in the 2S fraction (data not shown). It may
therefore, represent a heterogeneous population of proteins.
3.2.2 Identification of storage proteins
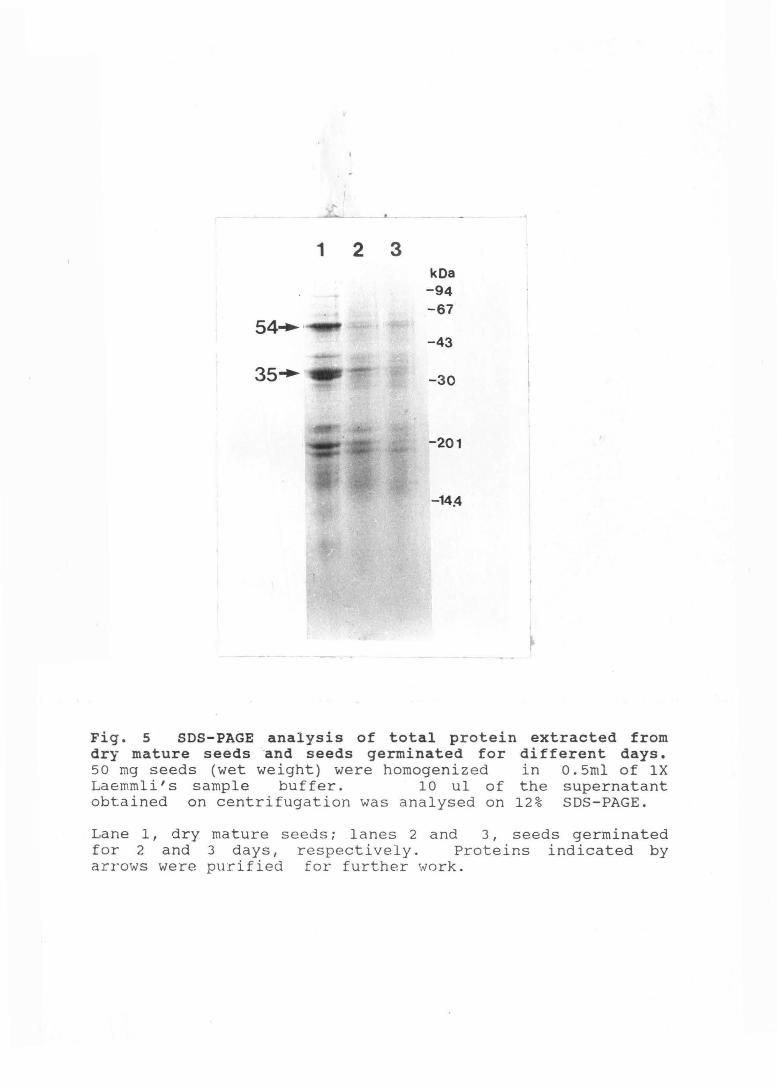
Amaranth seed extracts contain atleast four polypeptides in
large amounts which may be considered as storage proteins
(Fig. 5). Three of these polypeptides, 54, 34, and smaller
than 12 kDa disappear within a day of germination; however,
the 35 kDa polypeptide disappears only by the third day of
germination (Fig. 5). To further analyze their functions,
two of these proteins, 54 kDa and 35 kDa (indicated by an
arrow in Fig. 5) were purified.
73
2 3 4 5 6 7 8 9 10 11 12 M kDa
-94 -67
-43
-30
-20.1
-14.4
Fig.4 SDS-PAGE of various fractions obtained after sucrose density centrifugation. Crude extract (0.2 ml) containing 500 ug protein vJas layered on 12 ml . linear 5-20% sucrose gradient made in the extraction buffer. 1 ml fractions were collected. 2 0 0 u1 sample of each fraction was precipitated with equal volume of 20% TCA for 30 min. It was centrifuged and the pellet was washed thrice with ethanol:ether (1:1) and dried. Pellet was dissolved in SDS sample buffer and analysed on 12% SDS-PAGE.
Lanes 1 to 12 indicate the fraction numbers. 'M' is standard proteins lane.
Fractions 5, 8, 9, and 11 correspond to sedimentation values of 128, 7S, 5S, and 2S respectively.
1 2 3
54 ... ·
kOa -94 -67
-43
-30
-201
-14.4
Fig. 5 SDS-PAGE analysis of total protein extracted from dry mature seeds -and seeds germinated for different days. 50 mg seeds (wet weight) were homogenized in 0.5ml of lX Laemmli' s sample buffer. 10 ul of the supernatant obtained on centrifugation was analysed on 12% SDS-PAGE.
Lane 1, dry mature seeds; lanes 2 and 3, seeds germinated for 2 and 3 days, respectively. Proteins indicated by arrows were purified for further work.

3.2.3 Purification and Characterization of the 35 kDa
Albumin Protein
3.2.3.1 separation of albumins from globulins
seeds were extracted in high salt buffer and the crude
extract obtained was extensively dialysed against Tris
acetate buffer pH 8. 5. Globulins (insoluble in low salt
buffers) that precipitated during dialysis, were removed by
centrifugation. Supernatant having the albumin protein was
the source of 35 kDa protein.
3.2.3.2 Chromatofocussing
Chromatofocussing separates ·proteins on the basis of their
isoelectric points. Albumin fraction was bound to a DEAE
sepharose column previously equilibrated with Tris-acetate
buffer, pH 8.5. The column was washed with the same buffer
and the bound protein was eluted with a multicomponent
buffer system (Table 1), adjusted to a pH of 5. Proteins get
eluted on the basis of their pi values due to a pH gradient
generated in the column. Fractions were analysed on SDS
PAGE and their pH was determined (Figs. 6A and 6B).
The 35 kDa albumin protein eluted in four
different peaks at pH values of 7.4, 7.1, 6.8, 6.7 and also
in the salt wash (Figs. 6A and 6B). Protein peak, at pH
74

Seed meal (lg)
Two extractions with 20 ml cold acetone
Two extractions with 10 ml 0.5 M NaCl in 25 mM Tris-acetate buffer, pH 8.5
Dialysis against 25 mM Tris-acetate,pH8.5
Supernatant (Source of albumin protein)
Loaded on the Chromatofocussing column
35 kDa albumin protein, referred as AmAl,passed through gel filteration column
AmAl concentrated by centricon
... Lipids
precipitate(source of globulin protein)
Dissolved in Laemmeli buffer and separated on SDS-PAGE.
54 kDa band cut and electroeluted
SDS removed by KCl precipitation
Flow sheet depicting the purification of two proteins from Amaranth seeds.

Fig.6 Fractionation chromatofocussing column.
of soluble proteins on
A, Elution profile of the column eluate. ( - ) 1 Absorbance at 280 nm; ( ....... ) 1 pH of various fractions.
B, SDS-PAGE analysis of different column fractions. L, load; W, wash; M, standarQ.. proteins. Numbers indicate the fractions analysed. ·; Arrow indicates the 35 kDa protein. Equal volume from each fraction was analysed.

A - -
9
8
7
6
J 5 a.
4
I io I I I I I 10 30 40 50 60 70 alo FRACTION NUMBER
B
n
r 0·5 NaCl
91
0 ' 120
~ 0·5
-25
I 0 CXl N <i
130 140
.- 66
-53
-45
-36 - 29
-24
-20.1
-14.2
L W 28 29 30 42 48 67 69 85 88 130 M

7. 4, had the highest yield of 35 kDa protein and so was
taken for further work.
(Amaranth albumin 1).
It is being referred to as AmAl
3.2.3.3 Gel filtration
AmAl, from the chromatofocussing column, was concentrated to
a small volume using centricon-10 (Amicon) and applied to a
gel filtration column. Some small molecular weight
contaminants were retained in the column and the eluted
protein was checked for its purity by one- and two
dimensional PAGE. About 10 ug protein on one dimensional
and 5 ug on two dimensional-PAGE gave a single band (Fig. 7A
and 7B), so the protein is considered pure.
3.2.3.4 Isoelectric point of AmAl
Isoelectric point of AmA1 calculated from NEPHGE was about
7.8 (Fig. 7B). Proteins are not focussed to equilibrium in
NEPHGE. From the chromatofocussing data, isoelectric
point of AmA1 came to be about 7.4. Proteins are eluted from
the chromatofocussing column on the basis of their
isoelectric points. pi of AmA1 may therefore be around
7.4-7.8.
75

A
1 2 3 '· • ikOa
1--'94 .1 ... 167
! i• 43
........ ! ! .. 30
..... 20·1
8
· + ;kDa: -!94 \
~67
"-.• 43 l
Fig. 7 Purity of AntAl by one-dimensional (A) and two-dimensional (B) gel electrop~resis.
A, 10 ug (lane 1) and 5 ug (lane 2) of the purified protein were analysed on 12% SDS PAGE and stained with Coomassie Blue. Lane 3 shows molecular weight standards.
B, 5 ug protein was analysed on 2-D PAGE (NEPHGE in the first dimension and 10% SDS PAGE in the second dimension) and stained with Coomassie Blue. Dark patch at the bottom of the gel is due to ampholytes.

3.2.3.5 Molecular weight determination
Native molecular weight of AmAl determined by Sephadex G-200
was about 55 kDa (Fig. 8). However, on SDS-PAGE only a 35
kDa protein was observed. To see, if it was a dimer, the
protein was checked on SDS-PAGE without mercaptoethanol
treatment and also after treating the sample with 10 mM
iodoacetate.
the protein.
monomer, with
No difference was observed in the mobility of
It was, therefore, concluded that AmAl is a
a molecular weight of 3 5 kDa. The high
molecular weight observed with sephadex G-200 may be due to
some non specific protein aggregation within the column.
3.2.3.6 Antibody preparation and Immunodiffusion
Antibodies were raised against AmAl by immunization of
rabbits. Titre was checked by Ouchterlony immunodiffusion
technique as described by Garvey et al. ( 1977) . 10 ug of
crude extract and 1 ug of purified protein was titred with
different dilutions of antiserum. Precipitin line could be
observed upto a dilution of 1:16 (data not shown).
3.2.3.7 Specificity of antibodies as checked by Western blot
analysis
Western blot analysis was performed according to the
procedure of Towbin et al. ( 1979) . A single band was
76

20 -------'-:t 0
I . 0 .-->< ..,._____
-+--'
...c Ol
t>l
~ 4
L._
0
::J u
2 t>l
0 2
1 0 0.4 0.5 0.6
K ov
Fig. 8 Estimation of molecular weight of filtration chromatography. Purified chromatographed on Sephadex G-200 column. curve was constructed by plotting Kav values proteins against their molecular wei~hts.
AmAl by gel AmAl vias
Calibration of standard
( o ) indicates standard proteins; ( • ) indicates the size of AmAl.

observed on the blot when checked with crude extract. At
times, some faint high molecular weight bands were observed
when purified protein was used. So antibodies were affinity
purified (Section 2.13) and then used for immunodetection.
Affinity purified antibodies, at a titer of 1:500 could
detect upto 0.1 ng protein (data not shown). High molecular
weight bands were still visible. It may simply be due to
incomplete denaturation of the protein.
3.2.3.8 Isoe1ectric forms of AmA1
35 kDa albumin protein, as reported earlier, eluted from
the chromatofocussing column in four different peaks (Fig.
6) . To see if these proteins were related, the peak
fractions were checked for their reactivity against AmAl
antibodies. They were all seen to react (Fig.9), indicating
that they are the different isoelectric forms of 3 5 kDa
albumin protein. Storage proteins from other systems are
known to exist in families of proteins where the different
proteins may differ on the basis of charge (Higgins, 1984).
'!'he exact mechanism by which different charge variants of a
protein originate is not known, but they may be the products
of a single gene (Beachy et al., 1985).
77

1 2 3 kDa
35----.--
4- 5 6
I .I
Fig. 9 Western blot analysis of different peak fractions of chromatofocussing column probed with AmAl antibodies. Equal volume of su.rnple from different fractions obtained after chromatofocussing vlas resolved on 12% SDS-PAGE and transferred to nitrocellulose membrane. Immunoreactive polypeptide was detected with AmAl antibodies.
Lane 1, load; lanes 2-6, indicate fraction numbers 28, 42, 48, 67 and 130, respectively, of the elution profile shown in Fig.6A.

3.2.3.9 Glycosylation and Lectin activity
Lectins often accumulate in seeds in large amounts and are
known to agglutinate erythrocytes (Elzler, 1985). They are
mostly glycosylated. To see if AmAl has lectin activity, it
was incubated with rabbit erythrocytes as described
(Section 2 .15). No lectin activity was seen to be
associated with it. When stained for glycoproteins by PAS
staining, AmA1 was found not to be glycosylated (data not
shown).
3.2.4 Purification of 54 kDa polypeptides
3.2.4.1 Separation of globulin proteins on SDS-PAGE
Globulin fraction was precipitated from the crude
extract by extensive dialysis of crude extract against 25
mM Tris-acetate, pH 8.5 or water.
in SDS-sample buffer of Laemmli
Precipitate was dissolved
(1970) and separated on a
preparative SDS-PAGE. Protein standards were run
simultaneously.
54 kDa was cut
2. 8) •
Band corresponding to a molecular weight of
after staining the gel with KCl (Section
Electroelution
Proteins were electrophoretically eluted from the gel slice
in 50 mM Ammonium bicarbonate containing 0.1% SDS. SDS
78

confers a negative charge on the proteins and disaggregates
them. As a result quantitative recovery is possible. Eluted
protein was concentrated by acetone precipitation. Some SDS
also precipitated along with the protein.
3.2.4.3 Removal of residual SDS
The residual SDS was removed by precipitation with KCl
(Suzuki and Terada,l988). Potassium salts, at a final
concentration of 20 mM have been found to precipitate SDS
from protein solutions with no protein loss (Suzuki and
Terada, 1988). About 50% of the protein was recovered after
precipitation.
3.2.4.4 Purity of the protein
To see if some contaminating protein is copurified, the
eluted 54 kDa polypeptide was rechecked on SDS-PAGE. It
gave a single band, indicating that it is pure (Fig.lO).
3.2.4.5 Antibody preparation and specificity
Antibodies raised against this polypeptide gave a titer of
1: 16 by Ouchterlony immunodiffusion technique. When
specificity of the antibody was checked by Western blot
analysis, it was found to cross react with many bands in the
crude extract. Since the antibodies were raised against gel
79

1 2 kDa
-94 ,.,....., -67 ............... ·-·· ~,.
~ •• ' •
-43
-30
;_20.1
-14.4
Fig 10 Purity of 54 kDa globulin protein. was analysed for purity on 12% SDS-PAGE.
54 kDa protein
Lane 1, crude extract; Lane 2, purified protein.

purified protein, crossreactivity indicates that it shares
some common antigenic determinants with many other proteins
in the crude extract. It remains to be seen, if these
proteins have some common properties, like glycosylation, or
if they are related in some other way.
3.2.5 Amino acid composition
Purified storage proteins were analysed for their amino acid
composition. As seen in Table 2 , 54 kDa polypeptide is rich
in aspartate, glutamate, arginine and proline as compared to
AmAl which has high content of aspartate only. In general
seed storage proteins are known to contain high level of
these amino acids and they are know·n to be the chief
nitrogen transport compounds in plants (Goodwill and Mercer,
1983). In addition in legume seeds, it has been shown that
the amide nitrogen of asparagine is the chief available
source of nitrogen during development (Goodwill and Mercer,
1983). Both these proteins may therefore have storage
functions. AmAl also has high · lysine content ( 6. 6%) and
fairly good methionine level (1.6%) and so it may be more
important, as far as human nutrition goes. It was therefore
chosen for further work.
80
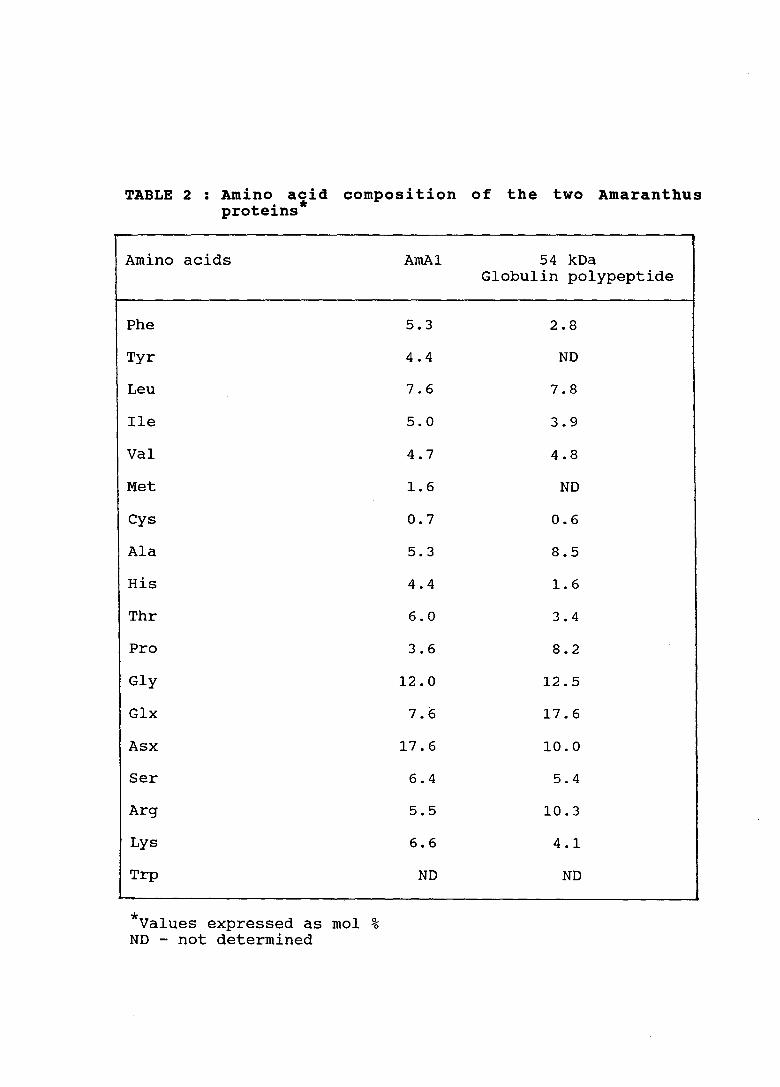
TABLE 2 : Amino acid composition of the two Amaranthus t . * pro e1ns
Amino acids
Phe
Tyr
Leu
Ile
Val
Met
Cys
Ala
His
Thr
Pro
Gly
Glx
Asx
Ser
Arg
Lys
Trp
* mol ~ Values expressed as 0
ND - not determined
AmA1
5.3
4.4
7.6
5.0
4.7
1.6
0.7
5.3
4.4
6.0
3.6
12.0
7.6
17.6
6.4
5.5
6.6
ND
54 kDa Globulin polypeptide
2.8
ND
7.8
3.9
4.8
ND
0.6
8.5
1.6
3.4
8.2
12.5
17.6
10.0
5.4
10.3
4.1
ND

3.2.6 Protein accumulation during development
seeds were classified into seven different developmental
stages on the basis of their weight (Table 3) . Protein
extracts of all the different stages, when analysed by
immunoblotting indicated that the synthesis of 54 kDa
polypeptide begins later than AmAl (Figs. 11A and 11B). AmAl
synthesis (monitored using anti-AmA1 antibodies) starts
soon after fertilization when the seed weight is only about
0.1 mg and reaches a maximum when the weight is about 0.3 mg
(Fig.11B). Accumulation of the 54 kDa polypeptide (studied
using anti-54 kDa antibodies) begins much later. It was
seen to be present in the initial stages of development as
well, though at a very low level (Fig.11A).
This kind of differential accumulation of different
proteins during seed development has been reported in other
plants, as well (Meinke et al., 1981; Goldberg et al., 1981;
Higgins, 1984). For example, in soybean seeds, 2S proteins
predominate during early stages of development but their
proportion decreases later when the 7S and 11S proteins are
synthesized (Hill and Briedenbach, 1974).
Before the accumulation of 54 kDa polypeptide begins, a
35 kDa polypeptide, which cross reacts with the 54 kDa
antibodies, is seen to appear, but disappears at the stage
81

TABLE 3 : Average wei~ht of seeds at different stages of development .
STAGE SEED WEIGHT(in mg)
I 0.05 ~
II 0.1
III 0.2
IV 0.3
v 0.4
VI o.a
VII 1
* Seeds were collected at different days after fertilization and separated into seven stages on the basis of their weight.

A 8
123456 71234567 kDa kOa
~35
Fig 11 Analysis of stage specific synthesis of the two proteins by Western blot. Crude extracts (20 ug protein) of seeds at different stages of development, as given in Table 2, were reso l ved on 12% SDS-PAGE,transferred to nitrocellulose membrane and probe d with A 54kDa antibodies and B, AmAl a nt ibodies .

when the 54 kDa polypeptide gets accumulated. We do not
know as yet, if these two polypeptides are related, since
the antibody raised against 54 kDa polypeptide was seen to
cross react with many other polypeptides.
3.2.7 Presence of translatable AmAl mRNA in mature seeds
Levels of seed protein mRNA is known to increase during seed
development, and then decrease as the seed matures (Goldberg
et al., 1981} . To see if AmAl mRNA is present in mature
seeds, poly(A)+ mRNA was isolated from seeds at stage V,
Table 3, (when this protein was not getting accumulated any
further) and dry mature seeds. Immunoprecipitation of the
in vitro translated p~oduct (Section 2.19} indicated that,
AmAl mRNA was present in mature seeds and was translatable
(Fig.12}.
3.2.8 seed specificity and function
On the basis of the germination-dependent disappearance of
the 54 kDa polypeptide (Fig.5), and also on its aminoacid
composition (Table 2), one can say that it is a storage
protein. But the role of AmAl as a storage protein is not
very clear, since it does not disappear immed~ately on
germination. Albumins have normally been considered as
metabolic proteins, but there are reports to show that some
82

kDa .. A., B 't
. ~.; .
. ' -~~~:,::· .( ·,.:
....
Fig 12 Imrnunoprecipi ta tion of in vitro translated products. Poly (A)+ RNA isolated from seeds, at stage V (Table 2) , (A} , and dry mature seeds, (B) , was translated in vitro in reticulocyte lysate system. Translated product was precipitated with AmAl antibodies bound to Protein ASepharose beads and analysed on 12% SDS-PAGE. 35s labeled and immunoprecipi tated product was then detected by autoradiography.

albumins also have storage functions. ·;,·:·~.Croy et al. (1984)
have reported an albumin protein from pea which does not
disappear immediately on germination but acts as a storage
protein. The delayed breakdown of this protein, according
to Higgins et al. ( 198 7) , is due to its cytoplasmic
localization. According to them, the proteolytic enzymes
are not released into the cytosol until the final stages of
cellular disorganisation. It is not known as yet, whether
AmAl is localized within protein bodies, or not, but a
• similar mechanism is possible for AmAl as well.
w~stern blot analysis of crude extracts from different
regions of the plant e.g. root, stem and leaves, did not
show any reactivity when probed with both the antibodies
(data not shown) . On the basis of above results and ·the
fact that they accumulate in large amounts during seed
development, it can be said that they are storage proteins.
3.2.9 Summary
1. Amaranth grain is rich in certain essential amino
acids like lysine and methionine which are normally
deficient in tradi tiona! crops. As a first step in the
isolation of a gene encoding a protein of high nutritional
value two proteins with molecular weights of 54 kDa and 35
kDa, have been purified from the seeds of Amaranthus
hypochondriacus.
83

2. The 35 kDa protein is soluble in low salt buffer
while the 54 kDa polypeptide requires high salt buffer for
solubilization.
3. On germination 54 kDa polypeptide disappears within a
day while the 35 kDa polypeptide is seen to disappear only
by the third day of germination.
4. 35 kDa protein was purified by chromatofocussing and
gel filtration columns. Purity of the protein was checked by
one dimensional and two dime.nsional PAGE.
5. 54 kDa polypeptide was purified by electroelution from
an 80S-polyacrylamide gel.
6. Antibodies, raised against both the proteins were
found to be of high titre.
7. Aminoacid composition of the two proteins indicated
that the 35 kDa protein has higher content of lysine.
8. For further work, the 35 kDa (AmAl) protein was chosen
because of its high lysine content. It was further
characterized to show that
a. It is a monomer of 35 kDa molecular weight.
b. It exists in four different isoelectric forms
c. It is not glycosylated.
84

d. Its accumulation in the seed starts soon after
fertilization when seed weight is only about 0.1 mg as
compared to 54 kDa polypeptide which is synthesized
much later.
e. mRNA encoding AmAl is present in dry mature seeds
as well.
85