Embed Size (px)
Citation preview

Gut, 1972, 13, 225-241
Progress reportThe synthesis and degradation ofliver-produced proteinsIn mammalian protein metabolism the term 'turnover' is used to convey theconcept of the combined synthesis and degradation of both intracellular andsecreted proteins. Although the synthetic and degradative processes areindependently sited and are probably controlled by separate regulatorymechanisms, there exists in steady state conditions a means for the main-tenance of a balance between the two. Only in these circumstances is theterm 'turnover rate' synonymous with both the rate of synthesis and degra-dation. In the case of liver-produced proteins it is more often the rate ofdegradation which is measured and the rate of synthesis inferred whenevidence is available that protein pool sizes are constant. With the develop-ment of methods for the direct measurement of hepatic protein syntheticrates in vivo1'2,3, there is little justification for the retention of impreciseterminology. The measured process should be clearly defined.For the purposes of this review, however, the concept of 'turnover' is
retained in order to emphasize the principle that proteins secreted by the cellfor export as well as proteins retained by the cell for home consumptionconsist of labile molecules which are continuously being catabolized andresynthesized. This phenomenon is not confined to situations in which there isnet transfer of amino acid nitrogen from one tissue to another, nor is itprevented by the supply of exogenous protein sufficient to keep the organismin nitrogen balance. One cannot do better than quote from Schoenheimer'sclassical work on the 'Dynamic state of body constituents' published in19424 which gives support to 'the generalization that all the reactions whichthe organism is capable of performing with protein constituents are carriedout continually, even when the total amount and constitution of the proteinsstay constant'. The amino acid products of protein degradation may undergoobligatory participation in a variety of metabolic cycles with the formationof new metabolites. These processes, like that of protein degradation, cannotbe prevented by the exogenous supply of identical substances.Although the basic concept of the dynamic interaction of body nitrogen
was first proposed by Borsook and Keighley5 in 1935, it was not until thestable isotope of nitrogen, 15N, became available for biological use that it waspossible to monitor accurately the interaction of food nitrogen with that ofplasma proteins6. These pioneer studies gave convincing experimental supportto the hypothesis of a cyclic process of release and uptake of nitrogen by thetissues to and from a circulating metabolic pool of amino acids. In the case ofthe liver, this process is not dependent upon the replacement of effete cells bynew cells. The life span of the rat liver cell is at least four to five months7'8 incontrast to the half life of total hepatic protein of two to four days9.The earliest measurement of protein turnover rates was based upon iso-
topic tracer techniques. Schoenheimer and his coworkers at ColumbiaUniversity obtained the first measurements of turnover rates of individual
225
on Decem
ber 3, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.13.3.225 on 1 March 1972. D
ownloaded from
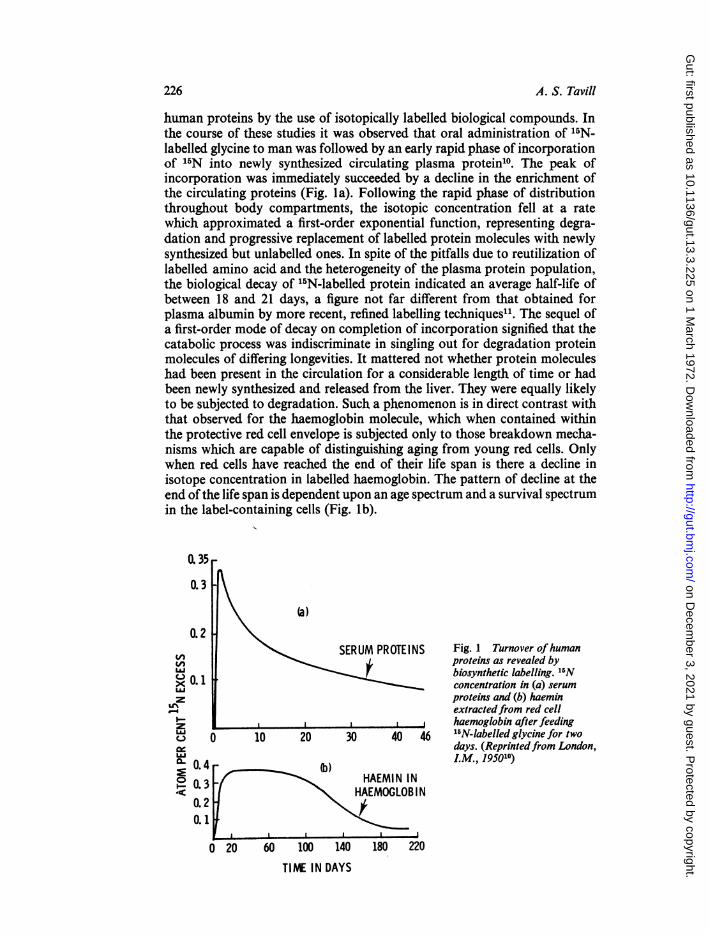
human proteins by the use of isotopically labelled biological compounds. Inthe course of these studies it was observed that oral administration of 15N-labelled glycine to man was followed by an early rapid phase of incorporationof 15N into newly synthesized circulating plasma protein10. The peak ofincorporation was immediately succeeded by a decline in the enrichment ofthe circulating proteins (Fig. la). Following the rapid phase of distributionthroughout body compartments, the isotopic concentration fell at a ratewhich approximated a first-order exponential function, representing degra-dation and progressive replacement of labelled protein molecules with newlysynthesized but unlabelled ones. In spite of the pitfalls due to reutilization oflabelled amino acid and the heterogeneity of the plasma protein population,the biological decay of '5N-labelled protein indicated an average half-life ofbetween 18 and 21 days, a figure not far different from that obtained forplasma albumin by more recent, refined labelling techniques". The sequel ofa first-order mode of decay on completion of incorporation signified that thecatabolic process was indiscriminate in singling out for degradation proteinmolecules of differing longevities. It mattered not whether protein moleculeshad been present in the circulation for a considerable length of time or hadbeen newly synthesized and released from the liver. They were equally likelyto be subjected to degradation. Such a phenomenon is in direct contrast withthat observed for the haemoglobin molecule, which when contained withinthe protective red cell envelope is subjected only to those breakdown mecha-nisms which are capable of distinguishing aging from young red cells. Onlywhen red cells have reached the end of their life span is there a decline inisotope concentration in labelled haemoglobin. The pattern of decline at theend ofthe life span is dependent upon an age spectrum and a survival spectrumin the label-containing cells (Fig. lb).
0.3
0.2(a)
5FRPUM PROTEINS
z
0.1
0 10 20 30 40 46
ff0.4 (b)HAEMIN IN
HAEMOGLOB I N0.2
0. 1
0 20 60 100 140 180 220
Fig. 1 Turnover ofhumanproteins as revealed bybiosynthetic labelling. 1"Nconcentration in (a) serumproteins and (b) haeminextractedfrom red cellhaemoglobin after feeding15N-labelled glycine for twodays. (Reprintedfrom London,LM., 195010)
TINE IN DAYS
226 A. S. Tavill
on Decem
ber 3, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.13.3.225 on 1 March 1972. D
ownloaded from

The synthesis and degradation of liver-produced proteins
Returning to the Schoenheimer quotation, it is necessary to emphasize hisstatement that turnover is carried on continually, even when the total amountand composition of the proteins remains constant. It is in these latter circum-stances that we talk of a 'steady-state' condition-a situation which mustimply a balance between anabolic processes on the one hand and catabolicprocesses on the other, and intrinsically therefore also implies the main-tenance of normal body pool sizes. Any perturbation in the balance betweensynthesis and catabolism will produce a change in pool masses. In theabsence of a compensatory response from the opposite side of the metabolicequation, the perturbation will ultimately lead to the destruction of the system.It is to the nature of these disturbing forces in the metabolism of liver-produced proteins and the compensatory respoinses induced by them that Ishould like to devote attention. Although they are finely balanced, the twosides of the turnover equation may for didactic purposes be consideredseparately. Physiologically they are very distinct biological processes. In termsof feedback control, however, they may have a common denominator,namely, the protein pool size and the concentration of the proteins in thesubdivisions of the body compartments.
Protein Catabolism
The half-life of a protein in the plasma, or for that matter an intrahepaticenzyme, is inversely proportional to the fractional catabolic rate expressed asa percentage of the relevant pool mass per unit time. This value can be theprimary determinant of the relative pool sizes of individual proteins. Thispoint may be illustrated more clearly by the following example. Two proteinsmay have identical turnover rates when expressed in absolute terms, ie, g perhour, but widely differing catabolic rates when expressed in terms of half-lifeor fractional rates of degradation. Their differing half lives reflect the relativeturnover times of differing pool masses. Since protein degradation appears tobe an obligatory physiological process obeying first-order kinetics, it followsthat it is the fractional rate of degradation rather than the rate of synthesisexpressed in absolute terms which may be responsible for the intrinsicdifferences in pool masses of various proteins and which determines thechange in pool mass in response to a change in synthesis.'2 Furthermore, incertain non-steady state conditions, modulation of catabolism alone ispotentially capable of restoring the status quo. This is illustrated in Figure 2.A protein pool size of 100 g may be maintained in balance by an input bysynthesis of 10 g and an output by degradation of a similar quantity. Areduction in the mass of protein in the pool by the removal of 50 g wouldrequire only that the fractional rate of breakdown of 10% of the pool per daybe maintained in order to restore the pool size to normal. Synthesis wouldexceed catabolism in absolute terms and the pool would enlarge (Fig. 2a).Likewise, expansion of the pool by the infusion of 50 g of protein would alsobe counterbalanced by maintenance of the normal fractional catabolic rate.In these circumstances the absolute catabolic rate would exceed the syntheticrate and the pool size would fall to normal (Fig. 2b). In contrast, a primarydefect in synthesis would result in a progressive fall in the pool mass with an
accompanying reduction in the mass of protein degraded. Only when the poolhas fallen to a level where the absolute rate of degradation is equal to the rate
227
on Decem
ber 3, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.13.3.225 on 1 March 1972. D
ownloaded from

A. S. Tavill
NORMAL STATE ABNORMAL STATE
509g(a)
10g 59g
S>C
50g
109 10010Sg
S -C
fb)10c
S<C
END RESULT
log0lg0gPool size restored
S -C
159 giOg9 g 10 g
Pool size restoredS -C
(C)5g9- 10_<10g
S<C
5 g 5 g
Pool size reducedS -C
Fig. 2 Metabolic responses mediated solely by the maintenance ofa constant fractionalrate of catabolism. The effects of(a) a reduction in pool size; (b) an increase in pool size;and (c) a reduction in the rate ofsynthesis. S = absolute rate ofsynthesis; C = absoluterate of catabolism.
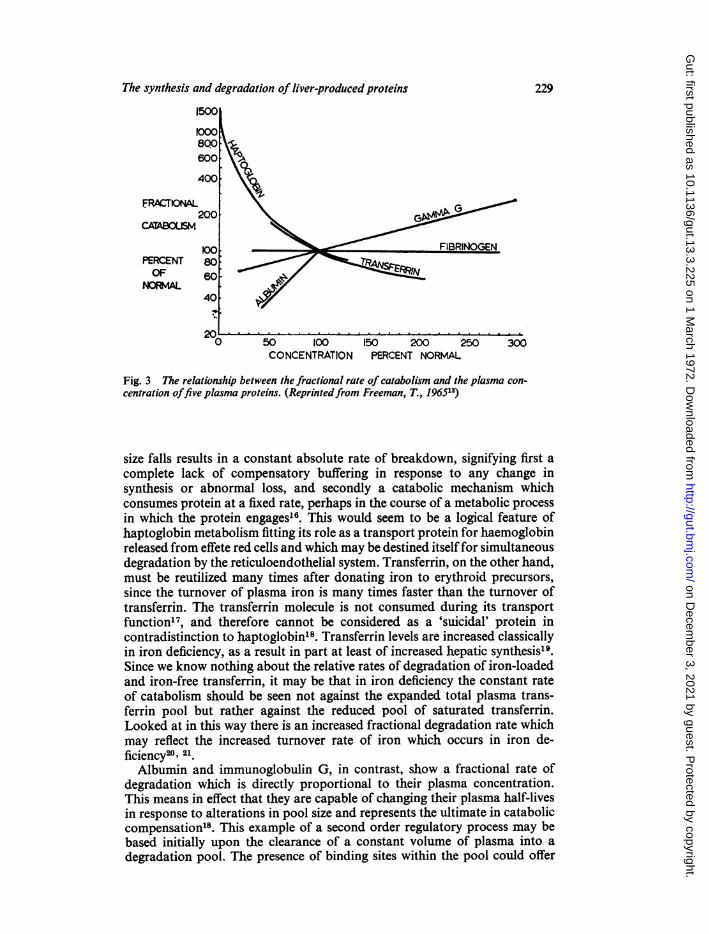
of synthesis will it stop shrinking (Fig. 2c). In the latter situation a responsewhich is geared to the maintenance of a constant fractional rate of degra-dation is capable only of re-establishing steady state conditions but is not initself able to restore the pool size to normal.Few plasma proteins only conform to this pattern of behaviour (Fig. 3).
One example is fibrinogen. Regardless of the plasma concentration or poolsize, the fractional rate of breakdown remains constant14. In this way, themass degraded falls as the pool size falls (assuming a constant plasma volume).This example of first-order control could be based on the clearance of aconstant volume ofplasma by the degradation process. Loss ofplasma proteininto the gastrointestinal tract is one means of catabolism which conforms tothe clearance principle. The fractional rate of degradation which is solely theresult of leakage of protein into the gut is equal for all plasma proteins,implying 'bulk loss' of proteins in the same proportions as they exist inplasma15. Regulation is intrinsically built into this mechanism of catabolism;a fall in plasma concentration is automatically accompanied by degradationof less protein.
In contrast, transferrin and haptoglobin are said to show an inverserelationship between fractional catabolic rate and plasma concentration13.An increase in fractional catabolic rate as the plasma concentration and pool
228
on Decem
ber 3, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.13.3.225 on 1 March 1972. D
ownloaded from
The synthesis and degradation of liver-produced proteins
PERCENTOF
NORMAL
50 100 150 200 2CONCENTRATION PERCENT NORMAL
Fig. 3 The relationship between the fractional rate of catabolism and the plasma con-centration offive plasma proteins. (Reprintedfrom Freeman, T., 196513)
size falls results in a constant absolute rate of breakdown, signifying first acomplete lack of compensatory buffering in response to any change insynthesis or abnormal loss, and secondly a catabolic mechanism whichconsumes protein at a fixed rate, perhaps in the course of a metabolic processin which the protein engages'6. This would seem to be a logical feature ofhaptoglobin metabolism fitting its role as a transport protein for haemoglobinreleased from effete red cells and which may be destined itself for simultaneousdegradation by the reticuloendothelial system. Transferrin, on the other hand,must be reutilized many times after donating iron to erythroid precursors,since the turnover of plasma iron is many times faster than the turnover oftransferrin. The transferrin molecule is not consumed during its transportfunction'7, and therefore cannot be considered as a 'suicidal' protein incontradistinction to haptoglobin'8. Transferrin levels are increased classicallyin iron deficiency, as a result in part at least of increased hepatic synthesis'9.Since we know nothing about the relative rates of degradation of iron-loadedand iron-free transferrin, it may be that in iron deficiency the constant rateof catabolism should be seen not against the expanded total plasma trans-ferrin pool but rather against the reduced pool of saturated transferrin.Looked at in this way there is an increased fractional degradation rate whichmay reflect the increased turnover rate of iron which occurs in iron de-ficiency20' 21.Albumin and immunoglobulin G, in contrast, show a fractional rate of
degradation which is directly proportional to their plasma concentration.This means in effect that they are capable of changing their plasma half-livesin response to alterations in pool size and represents the ultimate in cataboliccompensation'8. This example of a second order regulatory process may bebased initially upon the clearance of a constant volume of plasma into adegradation pool. The presence of binding sites within the pool could offer
229
on Decem
ber 3, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.13.3.225 on 1 March 1972. D
ownloaded from

protection against proteolytic activity up to a saturation limit22. A fallingconcentration of protein molecules in the cleared plasma would lead torelatively more complete binding and leave fewer available for degradation.A direct relationship between the plasma protein concentration and the frac-tional rate of degradation could thereby follow.Examples of the protective function of the albumin degradation process
are provided by the response to defects in synthesis. Patients with hepaticcirrhosis may have diminished synthetic rates, resulting in a low plasmaalbumin concentration and low intravascular pool sizes3. The survival of1311-labelled albumin in the circulation is greatly prolonged and both thefractional and absolute rates of degradation are much reduced3'2324 '25.Hypoalbuminaemia due to malnutrition is likewise accompanied by a re-duction in the fractional catabolic rate of albumin26'2728. In both theseexamples the primary disturbance is in the synthetic process. Secondarily,the concentration and pool size of the protein shrinks and the reduction indegradation follows. This sequence of metabolic alterations has been demon-strated by Kirsch and his coworkers29 in the rat subjected to experimentaldietary protein depletion. There was an early reduction in albumin synthesis(measured directly by the [14C] carbonate method) and a correspondingly earlyfall in plasma albumin concentration. A reduction in catabolism followedonly after a latent period. On repleting the animal with protein there was arapid synthetic response with an overshoot. The normal synthetic rate wasexceeded for a short time; the plasma albumin concentration increased andthe fractional catabolic rate rose in a more gradual fashion as the originalsteady state was restored.The liver-produced plasma proteins are distinctive in that they are for the
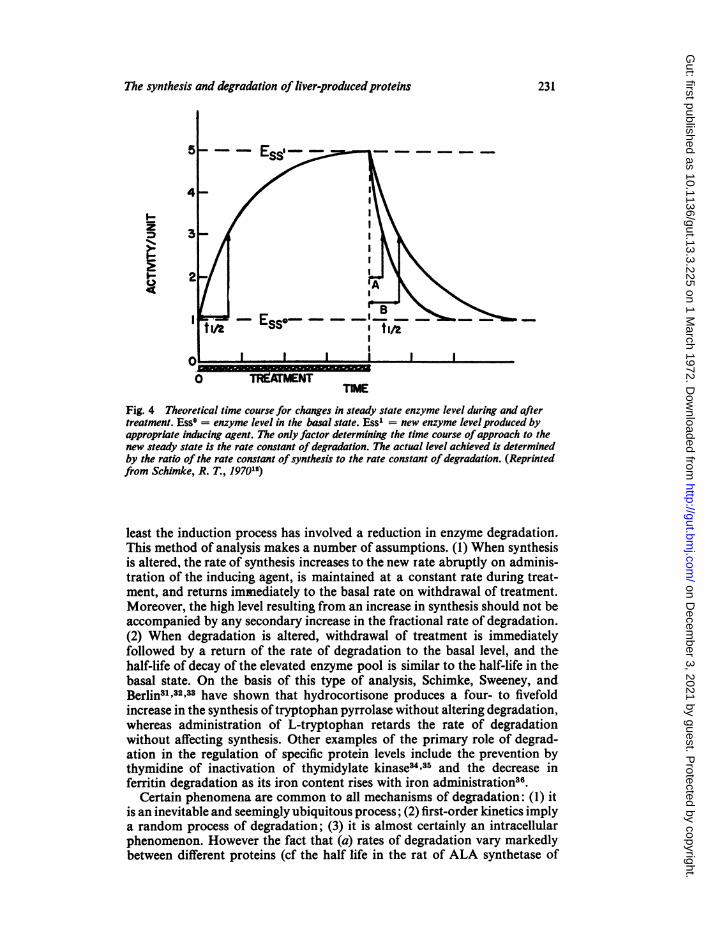
most part catabolized at extrahepatic sites. It is unlikely, for example, thatmore than 10% of the overall breakdown of albumin can be accounted for byintrahepatic degradation°0. Most hepatic enzymes on the other hand are bothsynthesized and degraded exclusively within the hepatocyte. However, asSchimkel2 has pointed out, both the proteins made for export and thoseretained within the cell demonstrate two features in common: first, the level ofthe protein (in the cell or plasma) may be modulated by changing either therate of synthesis or the rate of degradation. Secondly, there is a heterogeneityof degradation rates of specific proteins. It had initially been assumed that'induction' of enzyme activity by a suitable inducer, be it a factor in the diet,alterations of substrate level, or the administration of a drug or hormone, wassolely the result of a change in protein synthesis. More recent evidence hasdemonstrated that the levels of certain enzyme proteins may be controlled byalterations in degradation, depending upon the nature of the inducer. Thecharacteristics of turnover during and after the induction process which arenecessary to ascertain the mechanism of induction have been defined12. Theseare illustrated in Figure 4.The administration of an 'inducing' agent will produce an increase in the
enzyme level (from E8°0 to E88,) until a new steady state level is reached whereregardless of the mechanism of 'induction', synthesis again equals catabolismin absolute terms. Cessation of treatment is followed by a fall in enzymeconcentration to the initial resting level with restoration of the original steady-state conditions. If the rate of fall in enzyme concentration is equal to therate of rise during the induction process (B) then the increase was due to ahigher rate of synthesis. If the rate of fall is more rapid (A) then in part at
230 A. S. Tavill
on Decem
ber 3, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.13.3.225 on 1 March 1972. D
ownloaded from
The synthesis and degradation of liver-produced proteins
z I~~~~
I1-2 SS t,A0~~~~~~~
0 TATMENTTME
Fig. 4 Theoretical time course for changes in steady state enzyme level during and aftertreatment. Ess° = enzyme level in the basal state. Ess1 = new enzyme levelproduced byappropriate inducing agent. The only factor determining the time course ofapproach to thenew steady state is the rate constant of degradation. The actual level achieved is determinedby the ratio of the rate constant ofsynthesis to the rate constant of degradation. (Reprintedfrom Schimke, R. T., 197012)
least the induction process has involved a reduction in enzyme degradation.This method of analysis makes a number of assumptions. (1) When synthesisis altered, the rate of synthesis increases to the new rate abruptly on adminis-tration of the inducing agent, is maintained at a constant rate during treat-ment, and returns immediately to the basal rate on withdrawal of treatment.Moreover, the high level resulting from an increase in synthesis should not beaccompanied by any secondary increase in the fractional rate of degradation.(2) When degradation is altered, withdrawal of treatment is immediatelyfollowed by a return of the rate of degradation to the basal level, and thehalf-life of decay of the elevated enzyme pool is similar to the half-life in thebasal state. On the basis of this type of analysis, Schimke, Sweeney, andBerlin31 32'33 have shown that hydrocortisone produces a four- to fivefoldincrease in the synthesis oftryptophan pyrrolase without altering degradation,whereas administration of L-tryptophan retards the rate of degradationwithout affecting synthesis. Other examples of the primary role of degrad-ation in the regulation of specific protein levels include the prevention bythymidine of inactivation of thymidylate kinase34,35 and the decrease inferritin degradation as its iron content rises with iron administration36.
Certain phenomena are common to all mechanisms of degradation: (1) itis an inevitable and seemingly ubiquitous process; (2) first-order kinetics implya random process of degradation; (3) it is almost certainly an intracellularphenomenon. However the fact that (a) rates of degradation vary markedlybetween different proteins (cf the half life in the rat of ALA synthetase of
231
on Decem
ber 3, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.13.3.225 on 1 March 1972. D
ownloaded from

67 to 72 minutes37 with that of plasma albumin of three-and-a-half days38),and (b) the rate of degradation of individual proteins can be altered by specificagents, as illustrated in the foregoing examples, would seem to imply theexistence of a variety of mechanisms of degradation. Schimke12 has summar-ized the possible determinant factors which might decide when a givenprotein molecule is subjected to degradation. First the protein molecule itselfmay undergo conformational or other structural changes (possibly resultingfrom interactions with ligands) which render it more suitable or less suitableas a substrate for degradation. The stabilization of the apoenzyme tryptophanpyrrolase by conjugation with haematin in the presence of L-tryptophan isone such example39. Similarly, recent evidence has demonstrated that avariety ofplasma glycoproteins may undergo rapid degradation on removal ofsialic acid by the enzyme neuraminidase followed by binding to the hepato-cyte plasma membranes and subsequent intracellular uptake40 41'42. Thesurvival of such proteins is restored to normal either by restoration of thesialic acid residue or by further removal of the next residue of the carbohydratemoiety, namely, galactose43. Likewise, ferritin with a high iron content is lesssusceptible to the action of proteolytic enzymes44 which might explain thedecreased rate of degradation of ferritin synthesized under conditions of highiron load36. Although these are elegant demonstrations of the effects oflimited alterations in primary structure of a protein, no conclusive evidencehas yet been offered to suggest that this is the physiological mechanism ofdegradation in vivo. Secondly, the degrading system itself may be altered inactivity. This could take the form of a non-specific intracellular protease-peptidase system contained within lysosomal membranes. Control could beexercised by regulation of the level of the hydrolytic enzyme system, eg,inhibition of protein synthesis in rat liver inhibits the degradation of tyrosinetransaminase45 46 47. Such a model might be implicated in the changes incatabolism which occur in a response to protein deprivation. Alternatively,regulation of lysosomal activity might be mediated through stabilization orlabilization of the lysosome membrane, eg, the increased rate of hepaticprotein degradation which appears to occur following administration ofglucagon to the perfused rat liver48'49.
Hypoproteinaemic states in man may arise from a primary disorder ofcatabolism. When these situations are the consequence of an exaggeration ofa normal but usually minor degradative pathway, as occurs in the protein-losing enteropathies, methods are available for specific localization of theroute of loss50. Similarly, in the nephrotic syndrome it is possible to attributeto the kidney an exaggerated role in plasma protein degradation above andbeyond the protein overtly leaked in the urine. However, since no singleorgan or tissue has been identified as the primary site of degradation of theplasma proteins, other situations arise in which one is unable to pinpoint thebasic mechanism of hypercatabolism. Such is the case in infectious and toxicstates31 and in certain patients with malignant disease3. Experimental workhas suggested that pooling of plasma protein may develop in and aroundtumours52, which raises the possibility of subsequent catabolism of the proteinas a source of amino acids for growth of the cancer cell. Finally, evidence hasrecently been obtained to suggest that reticuloendothelial hyperplasia may beassociated with abnormal degradation ofplasma proteins53, although no proofcould be offered that this represented expansion of a normal catabolic path-way.
232 A. S. Tavill
on Decem
ber 3, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.13.3.225 on 1 March 1972. D
ownloaded from

The synthesis and degradation of liver-produced proteins
Protein Synthesis
I should like to discuss now the possible mechanisms ofthe synthetic response,either to the direct effect of hormones, drugs, or the level of enzyme substrate,or to the indirect effects of changes in catabolism.
In the simplified schema of protein synthesis depicted in Fig. 5, the twolevels at which synthesis may be controlled are distinguished, namely,transcription and translation. At the transcriptional level control ofthe rate ofprotein synthesis may be regulated by the rate of formation of mRNA fromthe DNA template by RNA polymerase, by the rate at which mRNA istransported from the nucleus to the cytoplasm or by the rate at which mRNAis degraded. Alternatively or additionally, the rate of protein synthesis couldbe regulated at a ribosomal level by the rate of translation of polypeptidechains, a process involving the activation of amino acids, their binding totRNA and the ribosome, the attachment of the ribosome-tRNA-activatedamino acid complex to the start of the mRNA strand, their relative movementalong the message with sequential propagation of the polypeptide chain andtheir ultimate release with completion of the polypeptide chain at the end ofthe mRNA.
Control at the level of transcription would involve a Jacob-Monod modelof gene expression56. In this model the primary effect of an inducer of proteinsynthesis would be to act as a derepressor of the operator gene upon whichthe structural gene coding for that protein relies for its activation. Con-versely, inhibitory agents could act as corepressors, undergoing allostericinteraction with the aporepressor to bind and inactivate the operator gene.While such a model would make an attractive hypothesis for the reg-ulation of protein synthesis in all types of cells, there is as yet no conclusiveproof that it is applicable to mammalian systems.
Protein end-product inhibition of its own synthesis would be a logicalsuggestion for both intracellular and secreted proteins. In the case of the
SCHEMA OF PROTEIN SYNTHESIS
rW tRNA --tRNAi.M. A
TRANSLATION RISOgOW
Fig. 5 Schematic representation of the processes involved in protein synthesis. Themessenger RNA molecule moves to the left so that there is relative movement of thepolyribosomes to the right resulting in the release of completedpolypeptide chains andsingle ribosomes. rRNA = ribosomal RNA; tRNA = transfer RNA; mRNA = messengerRNA; tRNAAA = transfer RNA-activated amino acid complex. (ReprintedfromTavill et al, 196714)
233
on Decem
ber 3, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.13.3.225 on 1 March 1972. D
ownloaded from

secreted, circulating plasma proteins, however, it is difficult to see how theconcentration of protein molecules in the intravascular or extravascularspace could affect the concentration within the cell16, except perhaps by reg-ulating release from the vesicles of the rough endoplasmic reticulum.For transcriptional control to provide a sensitive means of response to
.environmental stimuli, the specific mRNA for that protein must have arelatively rapid rate of turnover. It would be of limited value to shut off thesynthesis of new mRNA if preformed mRNA is stable enough to maintaintranslation for a significant period following the shut-off signal. It has beensuggested that proteins, viz, enzymes, which are rate-limiting in a biochemicalseries of reactions, are most likely to undergo rapid rates of degradation12. Itwould seem likely that the mRNA coding for such proteins would also have ashort turnover time. Conversely, those proteins not essential for rapidmodulation of biochemical reactions are likely to be slowly degraded, as aretheir specific messengers. The heterogeneity of degradation rates of hepaticmRNA has been elegantly demonstrated by Wilson and Hoagland56'57.Following a period of starvation the polyribosome population of rat liver isseverely reduced. If at the time of maximum reduction the animals wererefed, a new population of ribosomes was synthesized. Actinomycin D wasthen administered in order to block synthesis of further mRNA and thepolyribosome population allowed to turn over on the basis of mRNAdegradation. The capacity of these ribosomes to synthesize albumin wasstudied at various time intervals during this process. The newly formedribosomes decayed in a biphasic fashion; two-thirds had a half-life betweenthree and three-and-a-half hours; one third were much more stable with ahalf-life of at least 80 hours. Analysis of the protein synthesized by the morestable ribosomes showed it to be predominantly albumin. In other words,albumin synthesis by the liver is more refractory to conditions which reducemRNA synthesis than is the synthesis of most other liver proteins. While theseobservations preclude a transcriptional mechanism for rapid alterations ofhepatic albumin synthesis, it is still probable that more long-term effects mayhave such a basis.
Evidence to support a transcriptional means of control has for the mostpart relied upon the observed effect of actinomycin D on the activity of aknown inducer. Since actinomycin D inhibits DNA-dependent RNA syn-thesis, inhibition of induction implies that the increased rate of synthesis isdependent upon increased levels of mRNA. Possible examples of transcrip-tional control are the steroid induction of tryptophan pyrrolase58 and glutamicpyruvic transaminase59, the induction of hepatic microsomal enzymes andorganic anion-binding proteins by phenobarbitone and other drugs60 61, theincrease in acute-phase reactants in the plasma following injury62'63 and in thesynthesis of caeruloplasmin which follows the administration of copper torats64. Similarly, a repressor-operator mechanism has been suggested for theregulation of the structural gene coding for the mRNA of 8 aminolaevulinicacid (ALA) synthetase65. In this schema, haem serves as a corepressor of itsown synthesis66, while certain inducing chemicals, notably 53-H steroids,serve as derepressors by competing with haem for a binding site on therepressor, which is thereby inactivated. It has been suggested that derepres-sion precipitated by certain drugs may be responsible for exacerbations ofsymptoms in patients with porphyria and for the induction of cytochromeP450 and other enzymes involved in hepatic drug hydroxylation67'68.
234 A. S. Tavill
on Decem
ber 3, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.13.3.225 on 1 March 1972. D
ownloaded from

The synthesis and degradation of liver-produced proteins
Likewise, a good deal of conclusive evidence has been accumulated insupport of the translational control of protein synthesis at the level of thepolyribosome. Good correlation exists between the rate of protein synthesisand the proportion of the ribosomal population which exists in the form ofpolyribosomes, that is, messenger RNA molecules containing their fullcomplement of ribosomes (Fig. 6). An analogy may be drawn between a goodstrain travelling from A to B (the length of the mRNA strand). Clearly it willtransport more goods if it is drawing its maximum number of loaded wagons(polyribosomes carrying growing polypeptide chains) than if it is pullingfewer loaded wagons (functional oligosomes) and covering the journey in thesame time (translation time). This applies both to polyribosomes bound to therough endoplasmic reticulum and synthesizing proteins for export, eg, theplasma proteins and polyribosomes lying free in the cytoplasm which aresynthesizing proteins for home consumption, eg, intracellular enzymes andstorage, binding, transport, or membrane proteins69 70, 71. It is possible toanalyse the ribosomal population of the cell by means of differential sucrosegradient centrifugation in order to gain some quantitative idea of the degree ofaggregation of ribosomes. Baliga, Pronczuk, and Munro72 have shown in acell-free system that the response to amino acid deprivation is a rapiddisaggregation of polyribosomes into oligosomes. This process is reversibleon restoration of the essential amino acid content of the incubation medium,an effect which cannot be blocked by inhibition of RNA synthesis. This is aclear example of amino acid supply acting on protein synthesis within thelimits of preformed mRNA, ie, it denotes a translational mode of control.While similar acute effects have been noted in the intact liver73 74,75, trans-lational mechanisms cannot provide a complete answer to the defects inhepatic protein synthesis following upon chronic nutritional protein depri-vation. In rats subjected to a low protein diet for two to three weeks, albuminsynthesis, liver weight, and liver cell size fall markedly while the output ofalbumin per unit liver weight or body weight is reduced to a lesser de-
"Free" polyribosomie "Free po1yribosoMe Protein for home consumption"Free" oligosomes MRNA
Rough endoplasmic reticulum
"Membrane bound" oligosome - - //- Protein for export
"Membrane bound" polyribosomeFree ribosomes 0
Free ribosomal subunits
Fig. 6 Schematic representation of the liver cell to illustrate (1) the compartmentalizationof the machinery ofprotein synthesis and (2) the role ofpolyribosome aggregation in thequantitative control ofprotein synthesis at the translational level. (Modified from H. N.Munro. 1970104)
235
on Decem
ber 3, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.13.3.225 on 1 March 1972. D
ownloaded from

gree ''7778. However, polyribosome analyses of such livers have shown thataggregation may be preserved79. This would suggest that the output of proteinper cell is reduced and that this may be the result of an overall reduction inthe polyribosome population consequent upon a primary transcriptionaldefect in RNA synthesis80.The nature of the message to which the liver is capable of responding by
changing its rate of protein synthesis is reasonably clear in the case of aminoacid supply. It is probably based on a rate-limiting role for the amount ofcharged, ie, amino acid-loaded, transfer RNA. In spite of the fact that theliver maintains much of its amino acid pool by means of endogenous degra-dation of hepatic intracellular protein81, it is still dependent to a large extentupon exogenous dietary supply or amino acids released from extrahepaticprotein degradation for the replacement of obligatory losses from the pool.
While overall reduction in the supply of essential amino acids has the maxi-mum effect on protein synthesis, a special role for tryptophan has beensuggested82'83,84. This is supported by the effect of amino acids, and inparticular tryptophan, in preventing or reversing the reduction of hepaticprotein synthesis produced by alcohol85'86. It has been suggested that theunique rate-limiting role of tryptophan can be attributed to the very lowlevels of free tryptophan present in liver and plasma82'87.
Since we have already seen that a 'compensatory' fall in plasma proteindegradation may follow upon exogenous protein deprivation, it is possiblethat the fall in intracellular hepatic albumin catabolism88 and perhaps thecatabolism of other proteins may produce an abnormal dependence of syn-
thesis upon dietary amino acids. We have as yet no direct proof for thishypothesis. It does not conform with the sequence for circulating plasmaalbumin of, first, protein deprivation followed by reduced hepatic synthesis,and only then by reduced degradation29, and therefore must await simul-taneous measurements of hepatic protein synthesis and catabolism of bothplasma and intracellular proteins89.
In clinical practice hypoanabolic hypoproteinaemia resulting from inade-quate amino acid supply is seen in the kwashiorkor syndrome and severe
malabsorption states. A more subtle cause has been seen in the stagnant loopsyndrome90. Heavy bacterial contamination of the small intestine was
associated with low levels of many of the essential plasma amino acids and a
gross reduction in the hepatic synthesis (and degradation) of both albuminand fibrinogen. However, unlike defects in protein synthesis due to intrinsicliver disease, malabsorption or dietary deprivation, the associated rate ofurea synthesis measured directly was considerably increased. Much of thisurea was not recoverable in the urine. In this situation, the critical siting of alarge mass of bacterial flora could result in the degradation of dietaryaminoacids, principally by deamination. Absorption of released ammonia may haveresulted in the augmentation of urea synthesis by the induction of the po-
tentially inducible urea cycle enzyme system91. In addition, enterohepaticrecycling of urea could provide increased substrate for intestinal bacterialurease activity resulting in diminished urinary urea and further absorption ofammonia nitrogen, some of which may be utilized for protein synthesis92'93.The abnormalities in protein and urea synthesis and the levels of plasmaamino acids were corrected by antibiotic therapy.
In other situations, it appears that synthesis is able to respond to depletionof the protein pool resulting from extrinsic loss of protein from the plasma.
236 A. S. Tavill
on Decem
ber 3, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.13.3.225 on 1 March 1972. D
ownloaded from

The synthesis and degradatian of liver-produced proteins
This is the case in the nephrotic syndrome94 and in the protein-losing gastro-enteropathies3'96. In these examples, the nature of the feedback stimulus toprotein synthesis remains unsolved. It is difficult to imagine how a reductionin pool size alone could be detectable by the hepatocyte. The plasma con-centration of protein has been suggested as an alternative mediator. However,no consistent reduction in hepatic albumin synthesis was observed in patientsin whom low plasma albumin concentrations were corrected by intravenousalbumin infusions3. Nevertheless, Rothschild and coworkers96 have demon-strated that the isolated perfused liver is capable of responding to a reducedcirculating plasma albumin by increasing its rate of synthesis, and to anincreased concentration by reducing its rate of synthesis. However, they alsoshowed that isoosmolar replacement of albumin by sucrose had the sameeffect. It was suggested that the effect was exerted on the liver cell by changesin the oncotic pressure of the plasma via secondary but more profound directeffects of the hepatic interstitial fluid oncotic pressure97.Amino acid supply and plasma concentration effects are translational
regulatory mechanisms that may be common to many proteins of hepaticorigin. Hormonal effects may be more complex. Corticosteroids may have amarked anabolic effect on hepatic albumin synthesis in patients with activeliver disease, although no comparable effect was noted on fibrinogen synthe-sis98. This may in part have been due to an action at the level of the poly-ribosome99. It is hardly likely, however, that this anabolic role of corti-costeroids prevails throughout all tissues, since their effect on pool sizes andplasma concentrations of albumin were far less than that expected from theincrease in hepatic synthesis rate98 and their role in the whole organism ispredominantly catabolic. Clearly, there must be highly specific controlmechanisms which dictate which effect a hormone will produce in differenttissues and even within the same cell.
Other regulatory mechanisms may also exhibit a high degree of specificity.The role of iron in the control of synthesis of its storage protein, ferritin, 100,101and its carrier protein transferrin'9,or of lipid in the regulation of synthesis ofcarrier apolipoprotein102 may be examples of 'function-related' regulation. Inthis situation one may expect that eventually roles will be defined for pros-thetic groups, drugs, hormones, and anions in the regulation of the synthesisof their carrier apoproteins, which may have rate-limiting effects on themetabolism of these substances.
Since albumin makes the largest single contribution to plasma proteinturnover and the data on its metabolism are most complete, it would bevaluable to place this contribution quantitatively within the context of theprotein economy of the whole organism (Fig. 7). The evidence for some ofthese data has been reviewed by Fauconneau and Michel103. The averagealbumin synthesis of about 12 g per day (in a 70-kg man)"6 represents about25% of the total protein synthesized by the liver,7 and should be seen againstthe background of total body protein synthesis of about 300 g per day.Roughly 1 g per day of albumin is lost intact from the intravascular albuminpool into the gastrointestinal tract15. This represents a little less than 10% ofendogenous albumin catabolism under steady-state conditions, and is minutein relation to total exogenous and endogenous protein passing down the gut.Even if this were to increase by an order of magnitude it would produce nodrain on the total nitrogen economy since intestinal digestion and absorptionwould have little difficulty in dealing with it. It would, however, throw a
237
on Decem
ber 3, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.13.3.225 on 1 March 1972. D
ownloaded from

DIETARY INTAKE Fig. 7 The turnover of-g plasma albumin in a 70-kg
adult seen in the context of theHEPATIC 7 / t | daily protein economy of the
SYWrHESIS gastrointestinal tract andXYNJFHESI overall nitrogen balance. The121/day i s>)EN SECRETION total exchangeable albumin~ ENY?t~SECRTION pool of about 300 g is
INTRA- CELL TURNOVER distributed between theEXTRAVASCULA VASCULA DIGESTION intravascular and extravascularALBUMIN POOL ALBUMIN AND compartments in a ratio of
1809g= rPOOA PABSORPTION L approximately 2:3. In this1209g )iAMINOACID POOL simplified schema the balancel Jul sheet is expressed in terms of
g ofprotein (_ 6.25 x g ofCATABOLISM N). Losses do not include
relatively minor routes, eg, 2 gper day from the skin.
Ifl , { URINE N OUTPUTFAECAL OUTPUT 80 g protein
10g
considerable strain on the hepatic synthesizing capacity which would havedifficulty in doubling its total output of albumin. The effect would be anhypoalbuminaemic state without the consequences of overall nutritionalprotein deficiency.
Summary
In summary, the available evidence suggests that regulatory mechanisms inhepatic protein metabolism fall into two main categories. Specific control ofthe synthesis and degradation of individual proteins may be related to a vitaland unique metabolic role, while non-specific mechanisms such as amino acidsupply may range across the whole spectrum of liver-produced proteins.Other factors such as hormones may play a dual role. In the case of plasmaproteins of hepatic origin, homeostatic feedback mechanisms must logicallybe postulated to account for the remarkable stability of concentrations andpool sizes. Although the synthetic machinery for a specific protein may haveno direct control over the degradation of that protein it may compensate forthe effects of degradation on pool size by adjusting its own rate of activity.Likewise, catabolic processes can respond to alterations in pool size resultingfrom defective synthesis by a reduction in their activity. The exact mechanismsof these responses remain to be elucidated.
I should like to thank Dr R. Hoffenberg and Professor H. N. Munro for theirhelpful advice in the preparation of this manuscript.
Grateful acknowledgments are made to the authors and publishers con-cerned for permission to reprint Figures 1, 3, 4, 5, and 6.
A. S. TAVILLThe Division ofMetabolism, Clinical Research Centre (MRC), Northwick ParkHospital, Harrow, Middlesex
238 A. S. Tavill
on Decem
ber 3, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.13.3.225 on 1 March 1972. D
ownloaded from

The synthesis and degradation of liver-produced proteins 239
References
"Reeve, E. B., Pearson, J. R., and Martz, D. C. (1963). Plasma protein synthesis in the liver: Method for measure-ment of albumin formation in vivo. Science, 139, 914-916.
'McFarlane, A. S. (1963). Measurement of synthesis rates of liver-produced plasma proteins. Biochem. J., 89,277-289.
'Tavill, A. S., Craigie, A., and Rosenoer, V. M. (1968). The measurement ofthe synthetic rate ofalbumin in man.Clin. Sci., 34, 1-28.
4Schoenheimer, R. (1942). The Dynamic State of Body Constituents. Harvard University Press, Cambridge,Mass.
5Borsook, H., and Keighley, G. L. (1935). The 'continuing' metabolism of nitrogen in animals. Proc. roy. Soc.B., 118, 488-521.
'Schoenheimer, R., Ratner, S., Rittenberg, D., and Heidelberger, M. (1942). The interaction of the bloodproteins of the rat with dietary nitrogen. II. The interaction of antibody protein with dietary nitrogenin actively immunized animals. J. biol. Chem., 144, 541-554.
7Buchanan, D. L. (1961). Total carbon turnover measured by feeding a uniformly labeled diet. Arch. Biochem.,94, 500-511.
"Macdonald, R. A. (1961). 'Lifespan' of liver cells. Arch. intern. Med., 107, 335-343.9Swick, R. W. (1958). Measurement of protein turnover in rat liver. J. biol. Chem., 231, 751-764."°London, I. M. (1950). Studies on the rates offormation ofserum proteins in man. In Symposia on Nutrition.
Vol. II. Plasma Proteins, pp. 72-82. Thomas. Springfield, Ill."Cohen, S., Freeman, T., and McFarlane, A. S. (1961). Metabolism of '31I-labelled human albumin. Clin. Sci.,
20, 161-170.12Schimke, R. T. (1970). Regulation of protein degradation in mammalian tissues. In Mammalian Protein
Metabolism, vol IV, edited by H. N. Munro, pp. 178-228 Academic Press, New York.13Freeman, T. (1965). Gamma globulin metabolism in normal humans and patients. Ser. Haemat., 4, 76-86."'McFarlane, A. S., Todd, D., and Cromwell, S. (1964). Fibrinogen catabolism in humans. Clin. Sci., 26, 415-
420."Waldmann, T. A., Wochner, R. D., and Strober, W. (1969). The role of the gastrointestinal tract in plasma
protein metabolism. Studies with "'Cr-albumin. Amer. J. Med., 46, 275-285."Schultze, H. E., and Heremans, J. F. (1966). Molecular Biology ofHuman Proteins. Vol. 1. Nature and Metabo-
lism ofExtracellular Proteins. Elsevier, Amsterdam."Katz, J. H., and Jandl, J. H. (1964). The role of transferrin in the transport of iron into the developing red
cell. In Iron Metabolism (Ciba International Symposium), edited by F. Gross, p. 103. Springer, Berlin."Freeman, T. (1968). The function of plasma proteins. In Protides of the Biological Fluids, 15th Coll., Bruge
1967, edited by H. Peeters, pp. 1-14. Elsevier, Amsterdam."Tavill, A. S., and Kershenobich, D. (1971). Regulation of transferrin synthesis. In Protides of the Biological
Fluids, 19th Coll., Bruge, pp. 489-493. Pergamon, Oxford."Bohannon, R. A., Hutchinson, J. L., and Townsend, S. R. (1961). The use ofradioiron in the study ofanaemia.
Ann. intern. Med., 55, 975-988."'Finch, C. A., Deubebeiss, K., Cook, J. D., Eschbach, J. W., Harker, L. A., Funk, D. D., Marsaglia, G.,
Hillman, R. S., Slichter, S., Adamson, J. W., Ganzoni, A., and Giblett, E. R. (1970). Ferrokinetics inman. Medicine (Baltimore), 49, 17-53.
"Brambell, F. W., Hemmings, W. A., and Morris, I. G. (1964). A theoretical model of gamma-globulin cata-bolism. Nature (Lond.), 203, 1352-1354.
"3Sterling, K. (1951). Serum albumin turnover in Laennec's cirrhosis as measured by I'3-tagged albumin. J.clin. Invest., 30, 1238-1242.
"Berson, S. A., Yalow, R. S., Schreiber, S. S., and Post, J. (1953). Tracer experiments with I1"'-labelled humanserum albumin. Distribution and degradation studies. J. clin. Invest., 32, 746-768.
""Wilkinson, P., and Mendenhall, C. L. (1963). Serum albumin turnover in normal subjects and patients withcirrhosis measured by 113-labelled human albumin. Clin. Sci., 25, 281-292.
"Cohen, S., and Hansen, J. D. (1962). Metabolism of albumin and V-globulin in kwashiorkor. Clin. Sci., 23,351-359.
2"Hoffenberg, R., Black, E., and Brock, J. F. (1966). Albumin and y-globulin tracer studies in protein depletionstates. J. clin. Invest., 45, 143-152.
"James, W. P. J., and Hay, A. M. (1968). Albumin metabolism: effect of the nutritional state and the dietaryprotein intake. J. clin. Invest., 47, 1958-1972.
"Kirsch, R., Frith, L., Black, E., and Hoffenberg, R. (1968). Regulation ofalbumin synthesis and catabolism byalteration of dietary protein. Nature (Lond.), 217, 578-579.
3"Gordon, A. H. (I 957). The use of the isolated perfused liver to detect alterations to plasma proteins. Biochem.J., 66, 255-264.
"'Schimke, R. J., Sweeney, E. W., and Berlin, C. M. (1964). An analysis of the kinetics of rat tryptophan pyr-rolase induction: the significance of both enzyme synthesis and degradation. Biochem. biophys. Res.Commun., 15, 214-219.
"Schimke, R. T., Sweeney, E. W., and Berlin, C. M. (1965). The roles of synthesis and degradation in thecontrol of rat tryptophan pyrrolase. J. biol. Chem., 240, 322-331.
"3Schimke, R. T., Sweeney, E. W., and Berlin, C. M. (1965). Studies of the stability in vitro of rat tryptophanpyrrolase. J. biol. Chem., 240, 4609-4620.
"'Bojarski, T. B., and Hiatt, H. H. (1960). Stabilization of thymidylate kinase activity by thymidylate and bythymidine. Nature (Lond.), 188, 1112-1114.
'"Hiatt, H. H., and Bojarski, T. B. (1961). The effects of thymidine administration on thymidylate kinaseactivity and on DNA synthesis in mammalian tissues. Cold. Spr. Harb. Symp. quant. Biol., 26, 367-369.
3'Drysdale, J. W., and Munro, H. N. (1966). Regulation of synthesis and turnover of ferritin in rat liver. J.biol. Chem., 241, 3630-3637.
3"Marver, H. S., Collins, A., Tschudy, D. P., and Rechcigl, M. (1966). 8-aminolevulinic acid synthetase. II.Induction in rat liver. J. biol. Chem., 241, 4323-4329.
""Campbell, R. M., Cuthbertson, D. P., Matthews, C. M., and McFarlane, A. S. (1956). Behaviour of 1"C- and"I'I-labelled plasma proteins in the rat. Int. J. appl. Radiat., 1, 66-84.
"Knox, W. E., and Piras, M. M. (1967). Tryptophan pyrrolase of liver. 3. Conjugation in vivo during cofactorinduction by tryptophan analogues. J. biol. Chem., 242, 2959-2965.
on Decem
ber 3, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.13.3.225 on 1 March 1972. D
ownloaded from

240 A. S. Tavill
"Morell, A. G., Irvine, R. A., Sternlieb, I., and Scheinberg, I. H. (1968). Physical and chemical studies onceruloplasmin v. metabolic studies on sialic acid-free ceruloplasmin in vivo. J. biol. Chem., 243, 155-159.
"Hickman, J., and Ashwell, G. (1970). Physical and chemical studies on ceruloplasmin. IX. The role ofgalactosyl residues in the clearance ofceruloplasmin from the circulation. J. biol. Chem., 245,4397-4402.
"Pricer, W. E., and Ashwell, G. (1971). The binding of desialylated glycoproteins by plasma membranes ofrat liver. J. biol. Chem., 246, 4825-4833.
"Morell, A. G., Gregoriadis, G., Scheinberg, I. H., Hickman, J., and Ashwell, G. (1971). The role ofsialic acidin determining the survival of glycoproteins in the circulation. J. biol. Chem., 246, 1461-1467.
"Mazur, A., and Schorr, E. (1950). A quantitative immunochemical study of ferritin and its relation to thehepatic vasodepressor material. J. biol. Chem., 182, 607-627.
""Grossman, A., and Mavrides, C. (1967). Studies on the regulation of tyrosine aminotransferase in rats. J.biol. Chem., 242, 1398-1405.
"'Kenney, F. T. (1967). Turnover of rat liver tyrosine transaminase: stabilization after inhibition of proteinsynthesis. Science, 156, 525-528.
"Schimke, R. T. (1967). Protein turnover and the regulation of enzyme levels in rat liver. Nat. Cancer Inst.Monogr., 27, 301-314.
"'Green, M., and Miller, L. L. (1960). Protein catabolism and protein synthesis in perfused livers of normal andalloxan-diabetic rats. J. biol. Chem., 235, 3202-3208.
"Miller, L. L. (1961). Some direct actions of insulin, glucagon and hydrocortisone on the isolated perfused ratliver. Rec. Progr. Hormone Res., 17, 539-568.
"'Tavill, A. S. (1971). Protein-losing enteropathy. J. clin. Path., 24, Suppl. (Roy. Coll. Path.), 5, 45-54."'Jarnum, S., and Lassen, N. A. (1961). Albumin and transferrin metabolism in infectious and toxic diseases.
Scand. J. cdin. Lab. Invest., 13, 357-368."Wraight, E. P. (1971). Albumin metabolism in rabbits and rats with transplanted tumours. Brit. J. Cancer, 25,
365-376."Blaese, R. M., Strober, W., Levy, A. L., and Waldmann, T. A. (1971). Hypercatabolism of IgG, IgA, IgM and
albumin in the Wiskott-Aldrich syndrome. A unique disorder of serum protein metabolism. J. clin.Invest., 50, 2331-2338.
"ITavill, A. S., Grayzel, A. I., Vanderhoff, G. A., and London, I. M. (1967). The control of hemoglobinsynthesis. Trans. Ass. Amer. Phys., 80, 305-313.
'"Jacob, G., and Monod, J. (1961). Genetic regulatory mechanisms in the synthesis of proteins. J. molec. Biol.,3, 318-356.
"'Wilson, S. H., and Hoagland, M. B. (1967). Physiology of rat-liver polysomes. The stability of messengerribonucleic acid and ribosomes. Biochem. J., 103, 556-566.
7Wilson, S. H., Hill, H. Z.,and Hoagland, M. B. (1967). Physiology of rat-liver polysomes. Protein synthesisby stable'polysomes. Biochem. J., 103, 567-572.
5"Garren, L. D., Howell, R. R., Tomkins, G. M., and Crocco, R. M. (1964). A paradoxical effect of actino-mycin D: The mechanism of regulation of enzyme synthesis by hydrocortisone. Proc. nat. Acad. Sci.(Wash.), 52, 1121-1129.
"Segal, H. L., and Kim, Y. S. (1963). Glucocorticoid stimulation of the biosynthesis of glutamic-alaninetransaminase. Proc. nat. Acad. Sci. (Wash.), 50, 912-918.
"'Reyes, H., Levi, A. J., Gatmaitan, Z., and Arias, I. M. (1969). Organic anion-binding protein in rat liver: Druginduction and its physiologic consequence. Proc. nat. Acad. Sci. (Wash.), 64, 168-170.
"'Reyes, H., Levi, A. J., Gatmaitan, Z., and Arias, I. M. (1971). Studies of Y and Z, two hepatic cytoplasmicorganic anion-binding proteins: effects of drugs, chemicals, hormones and cholestasis. J. clin. Invest.50, 2242-2252.
'Koj, A. (1970). Synthesis and turnover of acute phase reactants. In Energy Metabolism in Trauma (CibaFoundation Symposium), edited by Ruth Porter and Julie Knight, pp. 79-92. Churchill, London.
"Sarcione, E. J. (1970). Synthesis and secretion of a,-(acute phase) globulin by the isolated perfused liver frominjured adult rats. Biochemistry, 9, 3059-3062.
"Evans, G. W., Majors, P. F., and Cornatzer, W. E. (1970). Induction of ceruloplasmin synthesis by copper.Biochem. biophys. Res. Commun., 41, 1120-1125.
"Kappas, A., and Granick, S. (1968). Steroid induction of porphyrin synthesis in liver cell culture. II. Theeffects ofheme, uridine diphosphate glucuronic acid, and inhibitors of nucleic acid and protein synthesison the induction process. J. biol. Chem., 243, 346-351.
"Marver, H. S., Schmid, R., and Schutzel, H. (1968). Heme and methemoglobin: naturally occurring repressorsof microsomal cytochrome. Biochem. biophys. Res. Commun., 33, 964-974.
"Marver, H. S. (1969). The role ofheme in the synthesis and repression of microsomal protein. In Microsomesand Drug Oxidations, edited by J. Gillette, A. H. Conney, G. J. Cosmides, R. W. Estabrook, J. R. Fouts,and G. J. Mannering, p. 495. Academic Press, New York.
"Tephly, T. R., Hasegawa, E., and Baron, J. (1971). Effect of drugs on heme synthesis in the liver. Metabolism,20, 200-214.
"Hicks, S. J., Drysdale, J. W., and Munro, H. N. (1969). Preferential synthesis of ferritin and albumin bydifferent populations of liver polysomes. Science, 164, 584-585.
"°Redman, C. M. (1969). Biosynthesis ofserum proteins and ferritin by free and attached ribosomes of rat liver.J. biol. Chem., 244, 4308-4315.
"'Takagi, M., Tanaka, J., and Ogata, K. (1969). Evidence for exclusive biosynthesis in vivo ofserum albumin bybound polysomes of rat liver. J. Biochem., 65, 651-653.
7'Baliga, B. S., Pronczuk, A. W., and Munro, H. N. (1968). Regulation of polysome aggregation in a cell-freesystem through amino acid supply. J. Molec. Biol., 34, 199-218.
7"Rothschild, M. A., Oratz, M., Mongelli, J., and Schreiber, S. S. (1968). Effects ofa short-term fast on albuminsynthesis studied in vivo, in the perfused liver, and on amino acid incorporation by hepatic microsomes.J. clin. Invest., 47, 2591-2599.
"Jefferson, L. S., and Korner, A. (1969). Influence of amino acid supply on ribosomes and protein synthesis ofperfused rat liver. Biochem. J., 111, 703-712.
7"John, D. W., and Miller, L. L. (1969). Regulation ofnet biosynthesis ofserum albumin and acute phase plasmaproteins. Induction of enhanced net synthesis of fibrinogen a, acid glycoprotein, a, (acute-phase)globulin and haptoglobin by amino acids and hormones during perfusion of the isolated normal ratliver. J. biol. Chem., 244, 6134-6142.
on Decem
ber 3, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.13.3.225 on 1 March 1972. D
ownloaded from

The synthesis and degradation of liver-produced proteins 241
"'Hoffenberg, R., Gordon, A. H., and Black, E. G. (1971). Albumin synthesis by the perfused rat liver. Acomparison of methods with special reference to the effect of dietary protein deprivation. Biochem. J.,122, 129-134.
"East, A., and Hoffenberg, R. Unpublished observations.78Tavill, A. S., Nadkarni, D., and Hoffenberg, R. Unpublished observations.7'Tavill, A. S., and Black, E. Unpublished observations."0Wannemacher, R. W., Wannemacher, C. F., and Yatvin, M. B. (1971). Amino acid regulation of synthesis of
ribonucleic acid and protein in the liver of rats. Biochem. J., 124, 385-392."'Gan, J. C., and Jeffay, H. (1967). Origins and metabolism of the intracellular amino acid pools in rat liver and
muscle. Biochim. biophys. Acta (Amst.), 148, 448-459.8'Sidransky, H., Sarma, D. S. R., Bongiorno, M., and Verney, E. (1968). Effect ofdietary tryptophan on hepatic
polyribosomes and protein synthesis in fasted mice. J. biol. Chem., 243, 1123-1132.83Rothschild, M. A., Oratz, M., Mongelli, J., Fishman, L., and Schreiber, S. S. (1969). Amino-acid regulation of
albumin synthesis. J. Nutr., 98, 395-403."'Sidransky, H., Verney, E., and Sarma, D. S. R. (1971). Effect of tryptophan on polyribosomes and protein
synthesis in liver. Amer. J. clin. Nutr., 24, 779-785."6Rothschild, M. A., Oratz, M., Mongelli, J., and Schreiber, S. S. (1971). Alcohol-induced depression of
albumin synthesis: Reversal by tryptophan. J. clin. Invest., 50, 1812-1818.""Jeejeebhoy, K. N., Phillips, M. J., Bruce-Robertson, A., Ho, J., and Sodtke, U. (1972). The acute effects of
ethanol on albumin fibrinogen and transferrin synthesis in the rat. Biochem. J., in press."'Munro, H. N. (1968). Role of amino-acid supply in regulating ribosome function. Fed. Proc., 27, 1231-1237.'8Hoffenberg, R., Gordon, A. H., Black, E. G., and Louis, L. N. (1970). Plasma protein catabolism by the
perfused rat liver. The effect of alteration of albumin concentration and dietary protein depletion.Biochem. J., 118, 401-404.
89Arias, I. M., Doyle, D., and Schimke, R. T. (1969). Studies on the synthesis and degradation of proteins of theendoplasmic reticulum of rat liver. J. biol. Chem., 244, 3303-3315.
9"Jones, E. A., Craigie, A., Tavill, A. S., Franglen, G., and Rosenoer, V. M. (1968). Protein metabolism in theintestinal stagnant loop syndrome. Gut, 9, 466-469.
'1Schimke, R. T. (1963). Studies on factors affecting the levels of urea cycle enzyme in rat liver. J. Biol. Chem.,238, 1012-1018.
92Richards, P., Metcalfe-Gibson, A., Ward, E. E., Wrong, O., and Houghton, B. J. (1967). Utilization ofammonia nitrogen for protein synthesis in man, and the effect ofprotein restriction and uraemia. Lancet,2, 845-849.
93Read, W. W. C., McLaren, D. S., Tchalian, M., and Nasser, S. (1969). Studies with "6N-labeled ammoniaand urea in the malnourished child. J. clin. Invest., 48, 1143-1149.
"4Katz, J., Bonorris, G., Okuyama, S., and Sellers, A. L. (1967). Albumin synthesis in perfused liver of normaland nephrotic rats. Amer. J. Physiol., 212, 1255-1260.
'5Wochner, R. D., Weissman, S. M., Waldmann, T. A., Houston, D., and Berlin, N. 1. (1968). Direct measure-ment of the rates of synthesis of plasma proteins in control subjects and patients with gastrointestinalprotein loss. J. clin. Invest., 47, 971-982.
9"Rothschild, M. A., Oratz, M., Mongelli, J., and Schreiber, S. S. (1969). Effect of albumin concentration onalbumin synthesis in the perfused liver. Amer. J. Physiol., 216, 1127-1130.
97Rothschild, M. A., Oratz, M., Evans, C. D., and Schreiber, S. S. (1966). Role of hepatic interstitial albuminin regulating albumin synthesis. Amer. J. Physiol., 210, 57-62.
98Cain, C. D., Mayer, G., and Jones, E. A. (1970). Augmentation of albumin but not fibrinogen synthesis bycorticosteroids in patients with hepatocellular disease. J. clin. Invest., 49, 2198-2204.
99Enwonwu, C. O., and Munro, H. N. (1971). Changes in liver polyribosome patterns following administrationof hydrocortisone and actinomycin D. Biochim. Biophys. Acta (Amst.), 238, 264-276.
1"°Yoshino, Y., Manis, J., and Schachter, D. (1968). Regulation of ferritin synthesis in rat liver. J. biol. Chem.,243, 291 1-2917.
"'OMunro, H. N., and Drysdale, J. W. (1970). Role of iron in the regulation of ferritin metabolism. Fed. Proc.,29, 1469-1473.
102Ruderman, N. B., Richards, K. C., Valles de Bourges, V., and Jones, A. L. (1968). Regulation of productionand release of lipoprotein by the perfused rat liver. J. Lipid Res., 9, 613-619.
1°Fauconneau, G., and Michel, M. C. (1970). The role of the gastrointestinal tract in the regulation of proteinmetabolism. In Mammalian Protein Metabolism. Vol. IV, edited by H. N. Munro, pp. 481-522. AcademicPress, New York.
104Munro, H. N. (Ed.) (1970). A general survey of mechanisms regulating protein metabolism in mammals. InMammalian Protein Metabolism, Vol. IV, pp. 3-130. Academic Press, New York.
on Decem
ber 3, 2021 by guest. Protected by copyright.
http://gut.bmj.com
/G
ut: first published as 10.1136/gut.13.3.225 on 1 March 1972. D
ownloaded from





























