Embed Size (px)
DESCRIPTION
Problem 1: Pakking av DNA. Hva er den maksimale pakkegrad for et DNA-stykke på 10 6 bp? 10 9 bp? Anta sylinder med 20Å diameter og lengde 3.4Å/bp. - PowerPoint PPT Presentation
Citation preview

Problem 1: Pakking av DNAHva er den maksimale pakkegrad for et DNA-stykke på 106 bp? 109 bp? Anta sylinder med 20Å diameter og lengde 3.4Å/bp
Den optimale pakkeform er en kule. Volumet av 106 bp DNA blir п·102·3.4·106 Å3 = 1.067·109 Å3. Den optimale kulen vil ha radius (3V/4 п)1/3 eller 630Å. Packing ratio: 3.4·106/1260=2698
For 109 pb blir volumet п·102·3.4·109 Å3 = 1.067·1012 Å3.
Radius 6300Å, packing ratio 3.4·109/6300 = 269000.
Diameter av en cellekjerne: 5 - 10 um = 50000 – 10 0000 Å

Problem 2: SupercoilingDersom et SV40-minikromosom relakseres slik at det danner en ikke oppvridd sirkel og så detproteiniseres fås et ccDNA med rundt –1 superheliksturn for hvert av de opprinnelige nukleosomene. Man skulle ventet –2 (DNA er opptvunnet med to omdreininger pr. nukleosom).
Forklaringen er at DNA kveilet rundt nukleosomer har høyere twist enn normalt, rundt 1 ekstra omdreining av dobbeltheliksen pr. nukleosom
L = T + W

To topologisk ekvivalente supercoilformer og en biologisk variant

Problem 3 Nukleosomoppbygging
Hvorfor vil sure polypeptider som polyGlu gjøre det lettere å sette sammen et nukleosom?
Histoner danner lett uspesifikke aggregater. Sure polypeptider vil kunne hindre slik aggregering og derved gjøre det lettere å få dannet spesifikke histonkomplekser

Problem 4. DNA-kompleksitet og cot-kurver
( ) 1500 400 400 000
oppkutting
Grønt DNA har kompleksitet 400 og rødt 400 000. Cot1/2-verdiene vil variere med en faktor på 1000

Reassosiasjonskinetikkd[A]/dt=-k2[A][B]
Integrering, [A]=[B]:
1/[A]=(1/[A0])+ k2 t
Fraksjonen uparrede tråder f=[A]/[A0]
f=1/(1+[A0] k2t)
Konsentrasjonene viser til konsentrasjonen av unike sekvenser, siden ikkekomplementære tråder ikke vil reassosiere. Dersom C0 er utgangskonsentrasjonen av basepar i løsningen:
[A0]= C0 /x hvor x er kompleksiteten av det DNA som analyseres. Dette gir
f=1/(1+ C0 t • k2 /x), og
C0 t1/2=x/ k2
xC0 t1/2=x/ k2

Reassosiasjonskinetikk for forskjellige typer DNA

Storfe sammenlignet med E. coli: Flere klasser av DNA og høyere kompleksitet

Cot-kurver
Cot-kurve for et hypotetisk DNA-molekyl som før fragmentering var 2 millioner bp langt og besto av en unik sekvens på 1 million bp og 1000 kopier av en sekvens på 1000 bp
Cot -kurve for Strongylocentrotus purpuratus
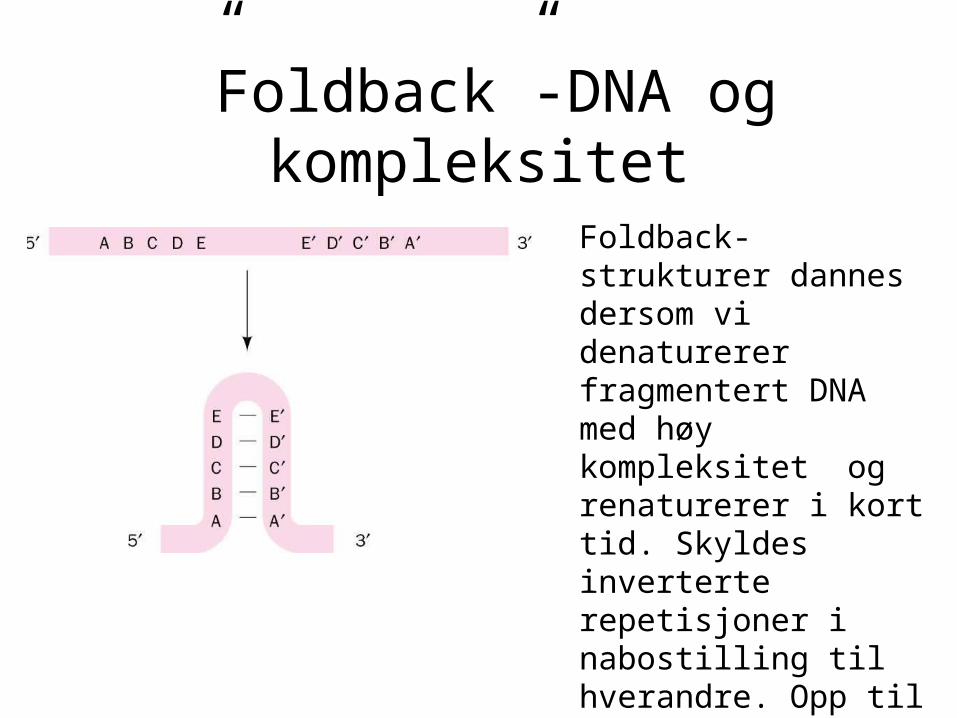
”Foldback”-DNA og kompleksitet
Foldback-strukturer dannes dersom vi denaturerer fragmentert DNA med høy kompleksitet og renaturerer i kort tid. Skyldes inverterte repetisjoner i nabostilling til hverandre. Opp til 10 % av noen genomer vil renaturere raskt med første ordens kinetikk, og utgjøres altså av slike inverterte repetisjoner

ss nuklease
Etter behandling med ss nuklease gir denaturering DNA med kompliserte Cot-kurver. Dette viser at noen slike tandemrepetisjoner forekommer flere ganger i genomet mens andre er unike

Problem 6: Mange rRNA-gener, ingen amplifisering av gener for ribosomale
proteiner
Xenopus lager 1012 ribosomer i løpet av en 2 mnd modningsperiode. Mulig fordi rRNA-genene foreligger i 1500 kopier.
a) Hvorfor er det tilstrekkelig med ett gen for hvert ribosomale protein?
Fordi hvert mRNA kan gi opphav til mange proteiner
b) Hvor lang modningstid uten amplifisering?
2 mnd x 1500 = 250 år

Problem 11: Retrovirus med oncogener


Hvorfor transformeres ikke vertsdyrcellene, mens immortaliserte
cellelinjer blir transformert?
Transformasjon i denne sammenheng: omdanning av en normal celle til en kreftcelle.
Transformasjon krever mange uavhengige genetiske endringer. Ett aktivert onkogen er ikke nok.
Immortaliserte celler har allerede gjennomgått flere av endringene på veien til å bli kreftceller. Her kan det ekstra onkogenet være alt som skal til for å fullføre transformasjonen

Problem 12: Hvorfor vil fusjoner mellom kreftceller og normale celler ofte miste
den tumorigene fenotypen?
Kreftaktivering av onkogener
Inaktivering av tumor su
ppressorgener
I den grad kreftcellens ukontrollerte proliferasjon skyldes mangel på genprodukter kan tilførsel av disse ved fusjon gi en mer kontrollert proliferasjon





























