Embed Size (px)
Citation preview

49
5 REZULTATI
5.1 POTEK ODLOVA
Odlov je potekal brez težav. Vse živali so odlov in uspavanje preživele brez kakršnih koli nam opaznih
pomembnejših posledic.
5.1.1 Lokacije odlova in časovni potek odlova
Odlov medvedov smo izvajali v letu 2008 v obdobju od 13.10. do 24.11. ter v letu 2009 od 30.1. do 4.5.
Odlov je potekal v loviščih lovskih družin Begunje, Iga vas, Kozlek, Krka, Loka pri Črnomlju, Martin Krpan Nova vas, Rakitna, Ribnica, Senožeče, Stari trg Lož, Tabor Zagorje, Toško Čelo in Velike Lašče ter v loviščih s posebnim namenom Jelen, Ljubljanski vrh, Medved, Snežnik Kočevska Reka in Žitna gora. Aldricheve pasti
smo postavili na krmiščih na 11 lokacijah (prikazane na Slika 16). Čas aktiviranih pasti na posameznih odlovnih mestih je predstavljen v Preglednica 4. Časovni pregled odlovljenih medvedov je podan na Slika 17.
Preglednica 4: Časovni pregled aktiviranih Aldrichevih pasti za medveda na posameznih lokacijah.
LOKACIJA ČAS AKTIVIRANIH PASTI
SKUPAJ ŠT. NOČI ODLOVLJENI MEDVEDI 2008 2009
Hoberlog, Kočevski rog 13.10.-19.10. / 6 Hobi
Peči, Kočevski rog 13.10-18.10 / 5 Pečko
Mlaka, Kočevska Reka 21.10.-12.11. 9.4.-16.4. 30 Jani
Korita, Kočevska Reka 2.11.-13.11. / 11 /
Pirenpod, Rakitna 26.10.-13.11. / 18 Ana, Eva
Zagabrnice, Rakitna 26.10.-14.11. / 19 /
Zakotek, Rakitna 26.10.-13.11. / 18 Rožle*
Loka pri Črnomlju 2.11.-14.11. / 12 Katja
Mala gora nad Ribnico 31.10.-15.11. / 15 Zora
Laški gozd, Velike Lašče / 22.4.-3.5. 11 Nejc
Gor. Kališče, Velike Lašče / 25.4.-3.5. 8 /
* medved Rožle je bil mladič in ni bil opremljen z ovratnico

50
Slika 16: Lokacije odlovnih mest, kjer so bile postavljene Aldricheve pasti za medveda.
Slika 17: Časovni pregled uspešno odlovljenih rjavih medvedov, ki smo jih opremili s telemetrijskimi ovratnicami.
Načrtovani odlov je potekal od 13.10.2008 do 24.11.2008 in od 30.1.2009 do 4.5.2009. Po tem obdobju smo z
ovratnico opremili tudi medveda, ki se je 15.10.2009 po naključju ujel v past za risa.

51
5.1.2 Uspešnost odlova
Skupaj smo v okviru pričujočega projekta odlovili 26 medvedov na 20-ih lokacijah. Od tega smo jih 9 (35 %) odlovili s pomočjo Aldrichevih pasti, 11 medvedov (42 %) pa smo odlovili s prosto tehniko, od tega sta bila dva uspavana v okviru intervencije, ena medvedka pa ob zabojni pasti, v kateri so bili ujeti njeni mladiči.
Mladiči so se kot netarčna vrsta po naključju ujeli v zabojno past, v kateri je potekal odlov divjih prašičev za raziskovalne namene v okviru raziskav Oddelka za gozdarstvo. Poleg teh mladičev, sta se v isto zabojno past ujela še ena medvedka skupaj z enoletnim mladičem, en medved pa se je po naključju ujel v zabojno past, v
kateri je potekal odlov evrazijskega risa za raziskovalne namene.
Na Slika 18 so predstavljene lokacije, kjer smo odlovili medvede. Skupaj za obe metodi je uspešnost znašala v povprečju 1 medved na 12.8 noči.
Slika 18: Lokacije, kjer smo uspešno odlovili medvede z njihovimi imeni (za natančnejše podatke in način odlova glej
Preglednica 5 in PRILOGA 3). Opomba: na nekaterih mestih smo odlovili več kot enega medveda (Ana in Eva ter Goranka
in Senožečanka). Na karti ni označenih štirih mladičev, ki so bili ujeti skupaj z njihovima materama (Goranka in
Senožečanka) v zabojno past namenjeno odlovu divjih prašičev. Medved Rožle je bil mladič in ni bil opremljen z ovratnico.

52
5.1.2.1 Odlov z Aldrichevimi pastmi
Aldricheve pasti smo postavili na 11 lokacijah (Slika 16). Na posamezni lokaciji smo postavili od 1 do 5 pasti
glede značilnosti krmišča (število stečin, konfiguracija terena, primernost podlage in sidrišč za zanko itd.). Skupaj na vseh lokacijah so bile pasti aktivirane 153 noči. Ob odlovu 9 medvedov je bila torej uspešnost odlova 1 medved na 17 noč na posamezno lokacijo. V vseh primerih je bil medved ujet za prednjo taco.
Poleg uspešno ujetih medvedov so bile pasti sprožene še 20-krat. V dveh primerih je bil v pasti opažen ujet
medved, ki pa se je snel z zanke še preden nam ga je uspelo uspavati. Med preostalimi 18 primeri smo na podlagi sledi in dlake na pasti lahko ugotovili, da je 8-krat past po vsej verjetnosti sprožil medved, vendar se ni ujel. V 10-ih primerih je past sprožila katera druga (netarčna) vrsta ali pa vzroka sproženja nismo mogli
ugotoviti.
5.1.2.2 Odlov s prosto tehniko
S prosto tehniko ob čakanju na krmiščih smo tekom rednega odlova za potrebe projekta ujeli 8 medvedov. Na medveda se je čakalo 64 noči. Uspešnost odlova s prosto tehniko je znašala 1 medved na 8 noči. Po
omamljanju so bili s pomočjo psa ali brez najdeni vsi uspavani medvedi. Medvedi so bili najdeni 0 do 800 metrov (povprečno 217 m) od mesta omamljanja.
5.2 ODLOVLJENI MEDVEDI
5.2.1 Odlovljeni medvedi v okviru tega projekta
Od skupno odlovljenih 26 medvedov smo jih 21 uspavali in opremili s telemetričnimi ovratnicami (Preglednica 5). Pet medvedov je bilo premladih (starih manj kot dve leti) in bi jih zaradi hitre rasti ovratnica
lahko začela ovirati še preden bi odpadla. Med medvedi opremljenimi z ovratnicami je bilo 10 samcev in 11 samic. Glede na starost je bilo med njimi največ (11) mladih medvedov (starih od 1.5 do 4.5 leta). Srednje starih medvedov (5 do 10 let) je bilo 6, trije pa so bili starejši (več kot 10 let). Med njimi je bil tudi zelo star
samec z ocenjeno starostjo vsaj 20 let. Natančnejši podatki o posameznih medvedih opremljenih z ovratnicami so podani v PRILOGA 3. Fotografije nekaterih medvedov so v PRILOGA 4.
Spolna in starostna struktura medvedov opremljenih s telemetrijskimi ovratnicami (Preglednica 6)je primerljiva s strukturo celotne populacije spolno zrelih medvedov v Sloveniji in zato smatramo, da
predstavlja reprezentativen vzorec. Podatki o številki, barvi in VHF frekvenci ovratnice ter barvi in številki ušesne markice za posameznega medveda so podani v PRILOGA 3.
Med opremljenimi medvedi sta bila tudi dva medveda, ki ju je kot konfliktna medveda preselila intervencijska skupina ZGS. Medvedka »Neža«, ki je bila odlovljena v okolici Muljave, je bila preseljena na
Menišijo, medved »Rožnik« pa je bil iz okolice Ljubljane preseljen na Snežniško planoto (Slika 19). Spremljanje teh dveh medvedov nam nudi dodatne podatke o učinkovitosti preseljevanja kot upravljavskega ukrepa za zmanjševanje konfliktov.
53
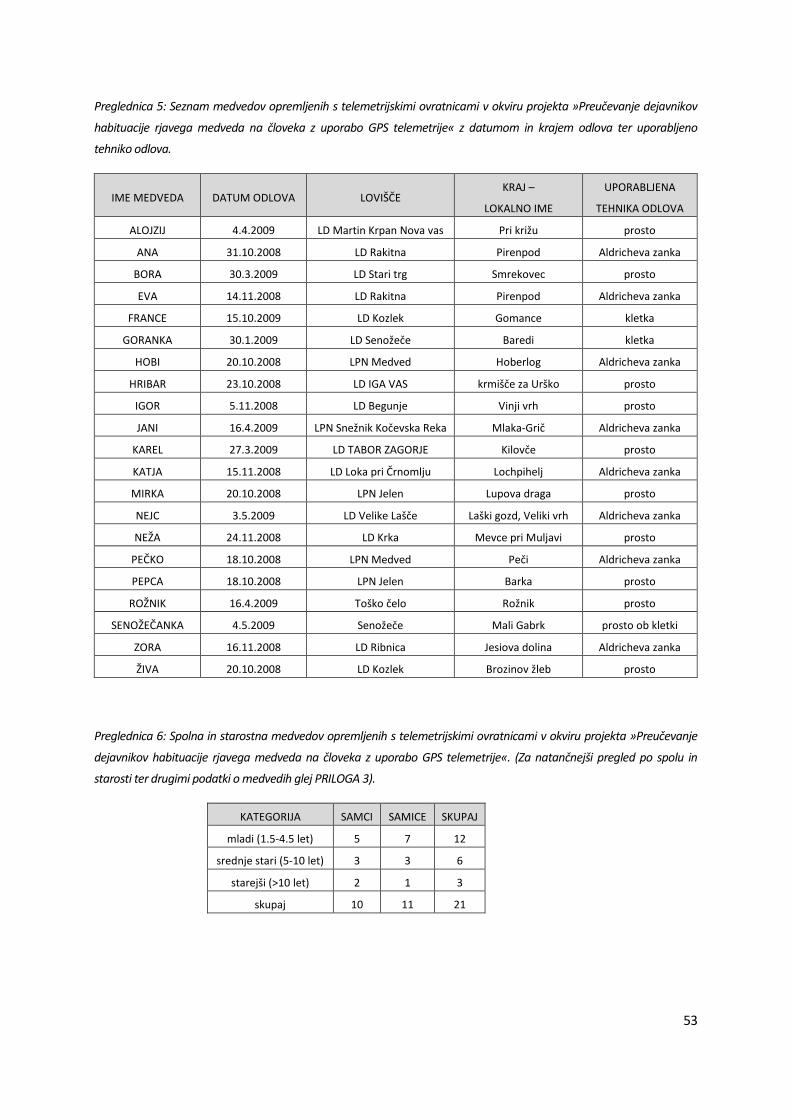
Preglednica 5: Seznam medvedov opremljenih s telemetrijskimi ovratnicami v okviru projekta »Preučevanje dejavnikov
habituacije rjavega medveda na človeka z uporabo GPS telemetrije« z datumom in krajem odlova ter uporabljeno
tehniko odlova.
IME MEDVEDA DATUM ODLOVA LOVIŠČE KRAJ –
LOKALNO IME
UPORABLJENA
TEHNIKA ODLOVA
ALOJZIJ 4.4.2009 LD Martin Krpan Nova vas Pri križu prosto
ANA 31.10.2008 LD Rakitna Pirenpod Aldricheva zanka
BORA 30.3.2009 LD Stari trg Smrekovec prosto
EVA 14.11.2008 LD Rakitna Pirenpod Aldricheva zanka
FRANCE 15.10.2009 LD Kozlek Gomance kletka
GORANKA 30.1.2009 LD Senožeče Baredi kletka
HOBI 20.10.2008 LPN Medved Hoberlog Aldricheva zanka
HRIBAR 23.10.2008 LD IGA VAS krmišče za Urško prosto
IGOR 5.11.2008 LD Begunje Vinji vrh prosto
JANI 16.4.2009 LPN Snežnik Kočevska Reka Mlaka-Grič Aldricheva zanka
KAREL 27.3.2009 LD TABOR ZAGORJE Kilovče prosto
KATJA 15.11.2008 LD Loka pri Črnomlju Lochpihelj Aldricheva zanka
MIRKA 20.10.2008 LPN Jelen Lupova draga prosto
NEJC 3.5.2009 LD Velike Lašče Laški gozd, Veliki vrh Aldricheva zanka
NEŽA 24.11.2008 LD Krka Mevce pri Muljavi prosto
PEČKO 18.10.2008 LPN Medved Peči Aldricheva zanka
PEPCA 18.10.2008 LPN Jelen Barka prosto
ROŽNIK 16.4.2009 Toško čelo Rožnik prosto
SENOŽEČANKA 4.5.2009 Senožeče Mali Gabrk prosto ob kletki
ZORA 16.11.2008 LD Ribnica Jesiova dolina Aldricheva zanka
ŽIVA 20.10.2008 LD Kozlek Brozinov žleb prosto
Preglednica 6: Spolna in starostna medvedov opremljenih s telemetrijskimi ovratnicami v okviru projekta »Preučevanje
dejavnikov habituacije rjavega medveda na človeka z uporabo GPS telemetrije«. (Za natančnejši pregled po spolu in
starosti ter drugimi podatki o medvedih glej PRILOGA 3).
KATEGORIJA SAMCI SAMICE SKUPAJ
mladi (1.5-4.5 let) 5 7 12
srednje stari (5-10 let) 3 3 6
starejši (>10 let) 2 1 3
skupaj 10 11 21

54
5.2.2 Pregled odlovljenih medvedov v okviru prejšnjih raziskav
Z namenom, da bi pridobili bolj zanesljive rezultate, smo v vse analize, kjer je bilo to mogoče in smiselno, vključili tudi podatke od medvedov, ki smo jih s pomočjo GPS telemetrije spremljali v okviru prejšnjih raziskav – projekt Life Nature III: LIFE02NAT/SLO/8585: Ohranitev velikih zveri v Sloveniji – Faza I (rjavi
medved), projekt Interreg III A/Phare CBC Italija-Slovenija 2000-2006: Trajnostno čezmejno upravljanje s favno ter v okviru rednih raziskav Raziskovalne skupine za varstvo gozdov in ekologijo prostoživečih živali na Oddelku za gozdarstvo in obnovljive gozdne vire Biotehniške fakultete Univerze v Ljubljani. V okviru prejšnjih
raziskav sicer nismo sistematično popisovali mikrohabitatne značilnosti lokacij medvedov, kljub vsemu pa bodo zbrani podatki iz GPS telemetrije, ki je potekala na enak način kot pri tem projektu, uporabni pri večini drugih analizah. V nadaljevanju zato na kratko podajamo pregled spremljanih medvedov v okviru teh
prejšnjih projektov (za podrobnosti glej poročila projektov).
Skupaj smo v okviru prejšnjih raziskav spremljali 12 medvedov, od tega 10 v Sloveniji in dva v Italiji (Slika 19). Podrobnosti o odlovljenih medvedih so podani v .
Preglednica 7. Spolna in starostna struktura medvedov je podana v Preglednica 8.
Preglednica 7: Seznam medvedov opremljenih s telemetrijskimi ovratnicami v okviru prejšnjih raziskav z datumom in
krajem odlova ter uporabljeno tehniko odlova.
IME MEDVEDA ODLOV LOVIŠČE KRAJ - LOKALNO IME UPORABLJENA TEHNIKA
ALJOŠA 29.4.2006 LPN Medved Hoberlog Aldricheva zanka
ANA SNEŽNIŠKA 2.4.2005 LPN Jelen Palež Prosto
ANDREA 2.4.2007 Italija Čedad Prosto
ANKA 4.8.2005 LPN Žitna gora Rjavi pesek Prosto
BEPI 4.3.2007 Italija Nadiške doline Prosto
BOHINJC 11.2.2008 LPN Prodi-Razor Bohinj Prosto
BORUT 18.5.2007 LD Nanos Požganina Prosto
BRANKO 8.5.2007 LD Kozja stena Kališki vrh Prosto
MARJETA 12.5.2005 LPN Jelen Žužtovec Prosto
PETER 24.4.2005 LPN Medved Bukova gora Aldricheva zanka
VALENTIN 23.3.2005 LPN Jelen Barka Prosto
ZORAN 23.3.2005 LPN Jelen Barka Prosto
Preglednica 8: Spolna in starostna medvedov opremljenih s telemetrijskimi ovratnicami v okviru prejšnjih raziskav.
KATEGORIJA SAMCI SAMICE SKUPAJ
mladi (1.5-4.5) 5 1 6
srednje stari (5-10) 2 1 3
starejši (>10) 2 1 3
skupaj 9 3 12

55
Slika 19: Lokacije, kjer smo uspešno odlovili medvede v okviru prejšnjih raziskav (rumene točke) in v okviru pričujočega
projekta (rdeče točke). Opomba: na nekaterih mestih smo odlovili več kot enega medveda. Na karti ni označenih štirih
mladičev, ki so bili ujeti skupaj z njihovima materama v zabojno past namenjeno odlovu divjih prašičev. Trije medvedi
(Bohinjc, Neža, Rožnik) so bili tekom raziskav preseljeni v okviru dela intervencijske skupine; modre puščice povezujejo
mesta, kjer je bil medved ujet in kasneje izpuščen.
5.3 GIBANJE IN AKTIVNOST MEDVEDOV
Skupaj smo v okviru telemetrijskega spremljanja medvedov zbrali 89 817 GPS lokacij medvedov (Slika 20 inSlika 21, Preglednica 9 inPreglednica 10). Od tega 74 211 v okviru pričujočega projekta in 15 606 v okviru
prejšnjih raziskav. Povprečen uspeh zajemanja GPS lokacij je bil 72,4 %. Poleg tega smo zbrali še 1 458 337 podatkov o aktivnosti medvedov (Preglednica 11 inPreglednica 12). Posameznega medveda smo spremljali v povprečju 249 dni (razpon 20-1040 dni).
Velikosti domačih okolišev in dolžine prehojenih poti posameznih medvedov so podani v Preglednica 13
Preglednica 14. Velikost domačih okolišev podajamo v obliki velikosti 100 % minimalnih konveksnih poligonov (MCP; Slika 22). Prehojene razdalje smo izmerili kot vsoto zračnih razdalj, ki povezujejo zaporedne GPS lokacije posameznega medveda. Mediana velikosti domačega okoliša spremljanih medvedov meri 317
km2. Kot je bilo opaženo že v okviru drugih raziskav rjavega medveda so povprečne velikosti domačih okolišev precej večje pri samcih (364 km2) kot pri samicah (129 km2). Skupaj smo zabeležili 37 205 km premikov medvedov. Karte z vrisanimi lokacijami, prehojenimi potmi in obrisom domačih okolišev za vse
spremljane medvede so prikazane v PRILOGA 8.

56
Slika 20: GPS lokacije vseh 33-ih spremljanih medvedov.

57
Preg
ledn
ica 9
: Pre
gled
zbr
anih
pod
atko
v o
zaje
man
ju G
PS lo
kaci
j, pr
ehaj
anju
med
drža
vnih
mej
a in
uso
dah
33 m
edve
dov
opre
mlje
nih
s te
lem
etrij
skim
i ovr
atni
cam
i. O
debe
ljena
črt
a
razm
ejuj
e m
edve
de s
prem
ljane
v o
kviru
pre
jšnjih
razis
kav
(1-1
2) in
med
vedo
v sp
rem
ljani
h v
okvi
ru p
roje
kta
»Pre
učev
anje
dej
avni
kov
habi
tuac
ije rj
aveg
a m
edve
da n
a čl
ovek
a z
upor
abo
GPS
tele
met
rije«
(13-
33).
AUT
– Av
strij
a, H
R –
Hrv
aška
, IT
– Ita
lija.
ZAP.
ŠT
. IM
E M
EDVE
DA
ZAČE
TEK
SPRE
M-L
JAN
JA
GIB
ANJA
KON
EC S
PREM
-LJ
ANJA
G
IBAN
JA
SKU
PAJ D
NI
BELE
ŽE-N
JA
GIB
ANJA
ŠT. P
OSK
USO
V ZA
JEM
A-N
JA
GPS
LO
KACI
JE
ŠT. Z
ABEL
E-ŽE
NIH
GPS
LO
KACI
J
USP
EH
ZAJE
MAN
JA G
PS
LOKA
CIJ [
%]
INTE
RVAL
ZA
JEM
ANJA
ZA
PORE
DN
IH
LOKA
CIJ [
h]
OBI
SKAN
E D
RUG
E D
RŽAV
E U
SOD
A
1 AL
JOŠA
30
.4.2
006
8.10
.200
6 16
1 19
47
545
28.0
2
HR
nezn
ano
- izg
ublje
n si
gnal
2 VA
LEN
TIN
24
.3.2
005
11.4
.200
6 38
3 45
98
1839
40
.0
2 H
R le
galn
o us
trel
jen
na
hrva
škem
3 PE
TER
25.4
.200
5 23
.11.
2005
21
2 25
52
711
27.9
2
ne
znan
o - i
zgub
ljen
sign
al
4 AN
A SN
EŽN
IŠKA
2.
4.20
05
9.9.
2006
52
5 55
65
1894
34
.0
2
nezn
ano
- izg
ublje
n si
gnal
5
ZORA
N
23.3
.200
5 1.
5.20
05
39
649
295
45.5
1.
5
lega
lno
ustr
elje
n
6 M
ARJE
TA
12.5
.200
5 18
.5.2
006
371
5397
14
64
27.1
1.
5
nezn
ano
- izg
ublje
n si
gnal
7
ANKA
4.
8.20
05
31.5
.200
6 30
0 35
57
959
27.0
2
ov
ratn
ica
odpa
dla
8 BO
RUT
19.5
.200
7 11
.5.2
008
358
4990
42
52
85.2
2
ne
znan
o - i
zgub
ljen
sign
al
9 BR
ANKO
8.
5.20
07
25.1
2.20
07
231
5171
23
51
45.5
1
ov
ratn
ica
pren
ehal
a z
delo
vanj
em
10
BOH
INJC
12
.2.2
008
9.3.
2008
26
63
8 46
7 73
.2
1
ovra
tnic
a od
padl
a
11
AND
REA
3.
4.20
07
29.8
.200
7 14
8 ni
pod
atko
v 49
3 ni
pod
atko
v 1
IT
ovra
tnic
a pr
eneh
ala
z de
lova
njem
12
BE
PI
5.3.
2007
27
.9.2
007
206
ni p
odat
kov
336
ni p
odat
kov
0.5
IT
lega
lno
ustr
elje
n
13
ALO
JZIJ
4.4.
2009
1.
10.2
009
180
4309
15
73
36.5
1
le
galn
o od
stre
ljen
14
ANA
RAKI
ŠKA
1.
11.2
008
21.1
0.20
09
354
6164
55
60
90.2
1
ov
ratn
ica
odpa
dla
15
PEPC
A 18
.10.
2008
31
.10.
2009
37
8 62
12
5124
82
.5
1
ovra
tnic
a od
padl
a
16
IGO
R 5.
11.2
008
12.4
.200
9 15
8 19
24
1601
83
.2
1 H
R sn
el o
vrat
nico
17
HRI
BAR
24.1
0.20
08
19.1
1.20
08
26
436
405
92.9
1
HR
snel
ovr
atni
co v
brlo
gu
(se
nada
ljuje
)

58
Preg
ledn
ica 9
(nad
alje
vanj
e)
ZAP.
ŠT
. IM
E M
EDVE
DA
ZAČE
TEK
SPRE
M-L
JAN
JA
GIB
ANJA
KON
EC
SPRE
M-L
JAN
JA
GIB
ANJA
SKU
PAJ D
NI
BELE
ŽE-N
JA
GIB
ANJA
ŠT. P
OSK
USO
V ZA
JEM
A-N
JA
GPS
LO
KACI
JE
ŠT. Z
ABEL
E-ŽE
NIH
GPS
LO
KACI
J
USP
EH
ZAJE
MA-
NJA
G
PS L
OKA
CIJ
[%]
INTE
RVAL
ZA
JEM
ANJA
ZA
PORE
DN
IH
LOKA
CIJ [
H]
OBI
SKAN
E D
RUG
E D
RŽAV
E U
SOD
A
18
ZORA
16
.11.
2008
29
.10.
2009
34
7 61
90
5427
87
.7
1
ovra
tnic
a od
padl
a
19
ROŽN
IK
17.4
.200
9 30
.5.2
009
43
1215
11
25
92.6
0.
25 d
o 1
HR
+ AU
T ile
galn
o od
stre
ljen
v Av
strij
i 20
ŽI
VA
20.1
0.20
08
12.1
.200
9 84
90
7 45
4 50
.1
1 H
R sn
ela
ovra
tnic
o v
brlo
gu
21
NEJ
C 3.
5.20
09
5.10
.200
9 15
5 37
21
3632
97
.6
1
povo
žen
na re
gion
alni
ce
sti L
J-Ko
čevj
e 22
KA
REL
27.3
.200
9 16
.4.2
009
20
482
462
95.9
1
le
galn
o od
stre
ljen
23
JAN
I 17
.4.2
009
10.6
.200
9 54
11
70
1123
96
.0
1
ovra
tnic
a od
padl
a (p
rehi
tro)
24
SE
NO
ŽEČA
NKA
4.
5.20
09
3.5.
2010
36
4 87
37
8250
94
.4
1
ovra
tnic
a od
padl
a
25
FRAN
CE
15.1
0.20
09
6.12
.200
9 52
12
45
844
67.8
1
HR
ovra
tnic
a pr
eneh
ala
z de
lova
njem
26
GO
RAN
KA*
30.1
.200
9 20
.9.2
011
943
6898
58
74
85.2
1
V
času
pis
anja
por
očila
ov
ratn
ica
še d
eluj
e
27
NEŽ
A
24.1
1.20
08
19.1
2.20
08
25
343
255
74.3
1
HR
izgu
blje
n si
gnal
; leg
alno
od
stre
ljena
(izr
edni
od
stre
l) br
ez o
vrat
nice
28
EVA*
14
.11.
2008
20
.9.2
011
1060
69
24
5995
86
.6
1
V ča
su p
isan
ja p
oroč
ila
ovra
tnic
a še
del
uje
29
KATJ
A 15
.11.
2008
17
.8.2
009
275
4300
34
68
80.7
1
HR
nezn
ano
- izg
ublje
n si
gnal
30
HO
BI
20.1
0.20
08
4.1.
2010
44
1 65
65
6290
95
.8
1
ovra
tnic
a pr
eneh
ala
z de
lova
njem
31
M
IRKA
20
.10.
2008
18
.4.2
010
545
7226
62
77
86.9
1
HR
ovra
tnic
a ve
rjet
no o
dpad
la
32
BORA
31
.3.2
009
14.1
1.20
09
228
4836
42
70
88.3
1
HR
ovra
tnic
a pr
eneh
ala
z de
lova
njem
33
PE
ČKO
19
.10.
2008
18
.2.2
010
487
6756
62
02
91.8
1
HR
ovra
tnic
a od
padl
a
OPO
MBA
: * v
čas
u pi
sanj
a po
roči
la o
vrat
nica
še d
eluj
e (s
hem
a: 1
loka
cija
na
23 u
r, VH
F se
vkl
aplja
na
dva
mes
eca)

Preglednica 10: Povzetek zbranih podatkov o zajemanju GPS lokacij 33 medvedov opremljenih s telemetrijskimi
ovratnicami.
SKUPINE OPISNA
STATISTIKA
SKUPAJ DNI
BELEŽENJA
GIBANJA
ŠT. POSKUSOV
ZAJEMANJA GPS
LOKACIJE
ŠT. ZABELEŽENIH GPS
LOKACIJ
USPEH ZAJEMANJA
GPS LOKACIJ [%]
skupaj
n = 33
minimum 20 343 255 27.0
mediana 228 4309 1601 83.2
maksimum 545 8737 8250 97.6
vsota 8167 121624 89817 73.9
povprečje 247 3923 2722 70.6
samci
n = 19
minimum 20 436 295 27.9
mediana 158 1947 1123 83.2
maksimum 487 6756 6290 97.6
vsota 3380 48368 34546 71.4
povprečje 178 2845 1818 70.3
samice
n = 14
minimum 25 343 255 27.0
mediana 359 5865 4697 83.8
maksimum 545 8737 8250 94.4
vsota 4787 73256 55271 75.5
povprečje 342 5233 3948 71.1
Slika 21: GPS lokacije 20-ih medvedov (brez Rožnika) spremljanih v okviru pričujočega projekta z obrobljenimi
domačimi okoliši.
59

60
Preglednica 11: Pregled zbranih podatkov o aktivnosti 33 medvedov opremljenih s telemetrijskimi ovratnicami.
Odebeljena črta razmejuje medvede spremljane v okviru prejšnjih raziskav (1-12) in medvedov spremljanih v okviru
projekta »Preučevanje dejavnikov habituacije rjavega medveda na človeka z uporabo GPS telemetrije« (13-33).
ZAP. ŠT. IME MEDVEDA
ZAČETEK
BELEŽENJA
AKTIVNOSTI
KONEC
BELEŽENJA
AKTIVNOSTI
SKUPAJ DNI
BELEŽENJA
AKTIVNOSTI
ŠTEVILO
PODATKOV O
AKTIVNOSTI
1 ALJOŠA 30.4.2006 15.10.2006 168 47764
2 VALENTIN 24.3.2005 11.4.2006 383 109071
3 PETER 25.4.2005 23.11.2005 212 60493
4 ANA SNEŽNIŠKA ni podatkov ni podatkov ni podatkov ni podatkov
5 ZORAN 23.3.2005 3.5.2005 41 11161
6 MARJETA ni podatkov ni podatkov ni podatkov ni podatkov
7 ANKA 4.8.2005 31.5.2006 300 85042
8 BORUT ni podatkov ni podatkov ni podatkov ni podatkov
9 BRANKO ni podatkov ni podatkov ni podatkov ni podatkov
10 BOHINJC 11.2.2008 12.3.2008 30 7738
11 ANDREA ni podatkov ni podatkov ni podatkov ni podatkov
12 BEPI ni podatkov ni podatkov ni podatkov ni podatkov
13 ALOJZIJ ni podatkov ni podatkov ni podatkov ni podatkov
14 ANA RAKIŠKA 1.11.2008 21.10.2009 354 100605
15 PEPCA 18.10.2008 31.10.2009 378 107346
16 IGOR 5.11.2008 12.4.2009 158 44705
17 HRIBAR 23.10.2008 19.11.2008 27 7619
18 ZORA 16.11.2008 29.10.2009 347 98594
19 ROŽNIK ni podatkov ni podatkov ni podatkov ni podatkov
20 ŽIVA 20.10.2008 12.1.2009 84 23678
21 NEJC ni podatkov ni podatkov ni podatkov ni podatkov
22 KAREL ni podatkov ni podatkov ni podatkov ni podatkov
23 JANI 17.4.2009 10.6.2009 54 15522
24 SENOŽEČANKA ni podatkov ni podatkov ni podatkov ni podatkov
25 FRANCE ni podatkov ni podatkov ni podatkov ni podatkov
26 GORANKA 30.1.2009 15.10.2009 258 73237
27 NEŽA ni podatkov ni podatkov ni podatkov ni podatkov
28 EVA 14.11.2008 29.4.2010 531 150901
29 KATJA 15.11.2008 28.7.2009 255 72306
30 HOBI 20.10.2008 6.11.2009 382 108500
31 MIRKA 20.10.2008 9.2.2010 477 135404
32 BORA 30.3.2009 12.11.2009 227 64345
33 PEČKO 20.10.2008 4.2.2010 472 134306

Preglednica 12: Povzetek zbranih podatkov o aktivnosti 33 medvedov opremljenih s telemetrijskimi ovratnicami.
SKUPINE OPISNA
STATISTIKA
SKUPAJ DNI
BELEŽENJA
AKTIVNOSTI
ŠTEVILO
PODATKOV O
AKTIVNOSTI
skupaj
n = 20
minimum 27 7619
mediana 256.5 72771.5
maksimum 531 150901
vsota 5138 1458337
povprečje 256.9 72916.85
samci
n = 10
minimum 27 7619
mediana 163 46234.5
maksimum 472 134306
vsota 1927 546879
povprečje 192.7 54687.9
samice
n = 10
minimum 84 23678
mediana 323.5 91818
maksimum 531 150901
vsota 3211 911458
povprečje 321.1 91145.8
S
lika 22: Domači okoliši (100 % MCP) vseh 33-ih spremljanih medvedov.61
62
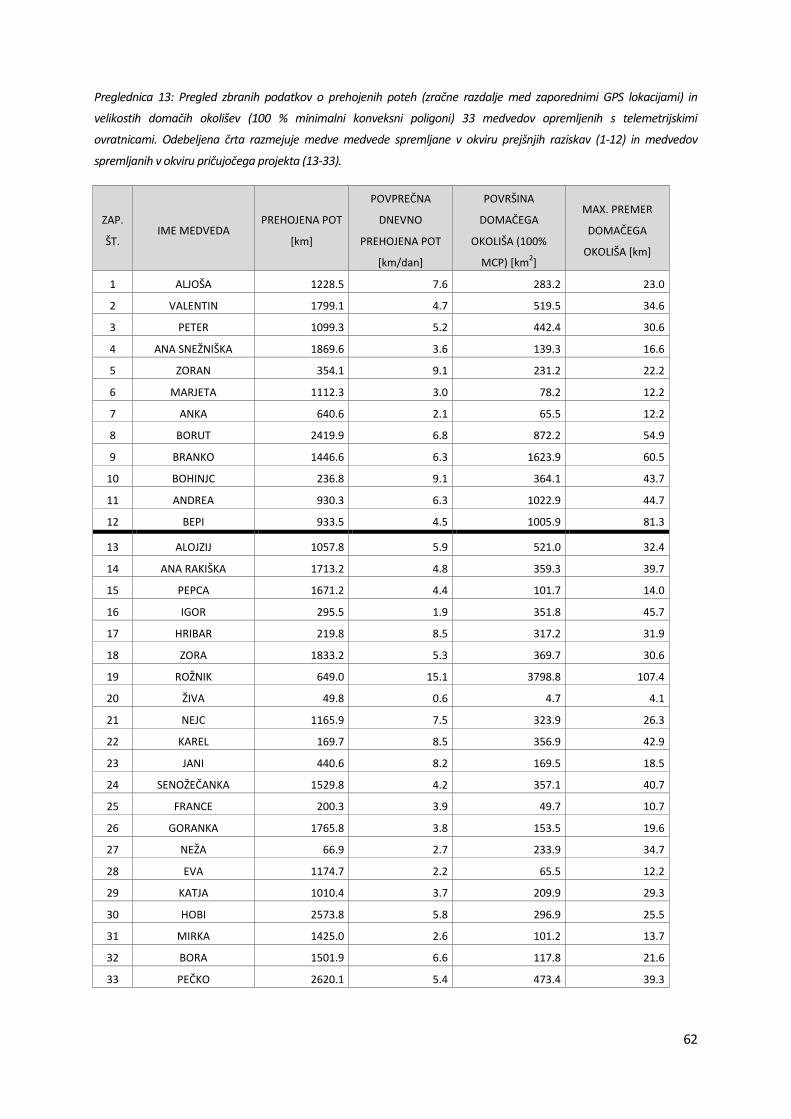
Preglednica 13: Pregled zbranih podatkov o prehojenih poteh (zračne razdalje med zaporednimi GPS lokacijami) in
velikostih domačih okolišev (100 % minimalni konveksni poligoni) 33 medvedov opremljenih s telemetrijskimi
ovratnicami. Odebeljena črta razmejuje medve medvede spremljane v okviru prejšnjih raziskav (1-12) in medvedov
spremljanih v okviru pričujočega projekta (13-33).
ZAP.
ŠT. IME MEDVEDA
PREHOJENA POT
[km]
POVPREČNA
DNEVNO
PREHOJENA POT
[km/dan]
POVRŠINA
DOMAČEGA
OKOLIŠA (100%
MCP) [km2]
MAX. PREMER
DOMAČEGA
OKOLIŠA [km]
1 ALJOŠA 1228.5 7.6 283.2 23.0
2 VALENTIN 1799.1 4.7 519.5 34.6
3 PETER 1099.3 5.2 442.4 30.6
4 ANA SNEŽNIŠKA 1869.6 3.6 139.3 16.6
5 ZORAN 354.1 9.1 231.2 22.2
6 MARJETA 1112.3 3.0 78.2 12.2
7 ANKA 640.6 2.1 65.5 12.2
8 BORUT 2419.9 6.8 872.2 54.9
9 BRANKO 1446.6 6.3 1623.9 60.5
10 BOHINJC 236.8 9.1 364.1 43.7
11 ANDREA 930.3 6.3 1022.9 44.7
12 BEPI 933.5 4.5 1005.9 81.3
13 ALOJZIJ 1057.8 5.9 521.0 32.4
14 ANA RAKIŠKA 1713.2 4.8 359.3 39.7
15 PEPCA 1671.2 4.4 101.7 14.0
16 IGOR 295.5 1.9 351.8 45.7
17 HRIBAR 219.8 8.5 317.2 31.9
18 ZORA 1833.2 5.3 369.7 30.6
19 ROŽNIK 649.0 15.1 3798.8 107.4
20 ŽIVA 49.8 0.6 4.7 4.1
21 NEJC 1165.9 7.5 323.9 26.3
22 KAREL 169.7 8.5 356.9 42.9
23 JANI 440.6 8.2 169.5 18.5
24 SENOŽEČANKA 1529.8 4.2 357.1 40.7
25 FRANCE 200.3 3.9 49.7 10.7
26 GORANKA 1765.8 3.8 153.5 19.6
27 NEŽA 66.9 2.7 233.9 34.7
28 EVA 1174.7 2.2 65.5 12.2
29 KATJA 1010.4 3.7 209.9 29.3
30 HOBI 2573.8 5.8 296.9 25.5
31 MIRKA 1425.0 2.6 101.2 13.7
32 BORA 1501.9 6.6 117.8 21.6
33 PEČKO 2620.1 5.4 473.4 39.3

63
Preglednica 14: Povzetek zbranih podatkov o prehojenih poteh (zračne razdalje, ki povezujejo zaporedne GPS lokacije) in
velikostih domačih okolišev (100 % minimalni konveksni poligoni) 33 medvedov opremljenih s telemetrijskimi
ovratnicami.
SKUPINE OPISNA
STATISTIKA
PREHOJENA POT
[km]
POVPREČNA
DNEVNO
PREHOJENA POT
[km/dan]
POVRŠINA
DOMAČEGA
OKOLIŠA (100%
MCP) [km2]
MAX. PREMER
DOMAČEGA
OKOLIŠA [km]
skupaj
n = 33
minimum 49.8 0.6 4.7 4.1
mediana 1112.3 5.2 317.2 30.6
maksimum 2620.1 15.1 3798.8 107.4
vsota 37205.0 15381.6 1077.3
povprečje 1127.4 5.4 466.1 32.6
samci
n = 19
minimum 169.7 1.9 49.7 10.7
mediana 933.5 6.3 364.1 34.6
maksimum 2620.1 15.1 3798.8 107.4
vsota 19840.7 13024.3 776.2
povprečje 1044.2 6.8 685.5 40.9
samice
n = 14
minimum 49.8 0.6 4.7 4.1
mediana 1463.4 3.6 128.5 18.1
maksimum 1869.6 6.6 369.7 40.7
vsota 17364.3 2357.3 301.0
povprečje 1240.3 3.5 168.4 21.5
14 od 33-ih spremljanih medvedov (42 %) so obiskali vsaj eno od tujih držav, največ (12 medvedov, 36 %) je
obiskalo Hrvaško (Preglednica 9). 6 medvedov je bilo legalno odstreljenih (eden od teh na Hrvaškem), 1 medved je bil nelegalno odstreljen v Avstriji (Rožnik), 1 medved je bil povožen na cesti (Nejc), 9 ovratnic je odpadlo s pomočjo »drop-off« mehanizma (1 prej kot načrtovano), 3 medvedi so si ovratnico sneli (2 med
zimskim dremežem v brlogu), 8 ovratnic pa je prenehalo z delovanjem medtem ko so bile še na medvedu (usoda medveda neznana, razen ene medvedke, ki je bila kasneje legalno odstreljena, vendar ni imela več ovratnice), od 6 medvedov pa smo signal izgubili v času, ko bi ovratnice še morale delovati (Preglednica 9).
Delovanje »drop-off« mehanizmov je bilo slabše od pričakovanih in so na nekateri celo popolnoma
odpovedali. Tega nismo opazili pri prejšnjih raziskavah na jelenjadi (Jerina in sod., neobjavljeni podatki), kjer smo uporabljali enak tip mehanizma. Vsi medvedi, ki so nosili ovratnice, pri katerih »drop-off« mehanizem ni deloval, so že popolnoma odrasli in samice, tako da ne pričakujemo, da bi imeli večje težava zaradi
neodpadlih ovratnic. Po dosedanjih izkušnjah z leti ovratnice kljub vsemu slej ali prej odpadejo, tako da pričakujemo, da bo tako tudi z medvedi opremljenimi z ovratnicami v okviru pričujočega projekta. Kljub vsemu pa priporočamo, da se v prihodnjih raziskavah poleg »drop-off« mehanizma uporabi tudi bombažni
trak, ki sčasoma preperi in omogoči, da ovratnica odpade, tudi ob odpovedi »drop-off« mehanizma. Takšen trak smo uspešno uporabili na Lotek-ovi ovratnici na medvedki »Senožečanki«, ki ni bila opremljena z »drop-off« mehanizmom.

64
5.3.1 Priprava GPS posnetih lokacij spremljanih medvedov za nadaljnje analize
Skoraj vse obstoječe statistične metode vsaj implicitno predpostavljajo, da so posnete lokacije in vsi podatki, ki jih iz njih ekstrahiramo (npr. razdalje med lokacijami), nepristranski vzorec spremljanih osebkov in ti
nepristranski vzorec preučevane populacije. Vendar predpostavka ni avtomatično zagotovljena. Za njeno zadostitev smo v pričujoči raziskavi odlov prilagajali lokalnim gostotam medvedov in z njim pokrili oba spola in starostne kategorije (razen mladičev) tako, da spremljan vzorec osebkov čim bolj reprezentativno
predstavlja populacijo, kot je opisano v poglavju 4.1.1. Poleg tega smo po postopkih, kot je opisano v nadaljevanju, posvetili veliko dela pripravi posnetih lokacij.
(i) Ovratnice, ki smo jih uporabili v pričujoči raziskavi, so bile nastavljene tako, da so nehale snemati lokacije, kadar je medved 4 ure miroval in se v normalno shemo delovanja vrnile takoj, ko je medved zopet postal
aktiven. To nastavitev smo uporabili, da bi preprečili veliko in nepotrebno uporabo toka baterij, ko bi medvedi šli v brlog na zimski počitek, in tako podaljšali življenjsko dobo delovanja ovratnic. Vendar so nekateri medvedi tudi med letom čez dan večkrat popolnoma mirovali več kot 4 ure skupaj (celo do 12 ur). V
takih primerih so vse lokacije, ki bi bile sicer posnete, po 4 urah mirovanja, dokler medved ni postal aktiven, izpadle. Take »lokacije« smo prepoznali po tem, da se ob predvidenem času meritve lokacije GPS sprejemnik sploh ni aktiviral, poleg tega je senzor za aktivnost prej vsaj 4 ure kazal na medvedovo mirovanje. Vse take
manjkajoče lokacije smo poiskali in jih nadomestili s podatki zadnjega snemanja lokacije, preden je medved začel mirovati. Za iskanje manjkajočih lokacij smo pripravili logične filtre v programu Excel. Na ta način smo število snemanj lokacij z 90 563 (podatek za uspešno posnete lokacije in tudi neuspele poskuse snemanj)
povečali za 10 950 na skupaj 101 513.
(ii) Pri GPS telemetriji se sprejemnik ne uspe vselej povezati z dovolj sateliti, da bi lahko posnel lokacijo. Na uspeh snemanj lahko vplivajo številni dejavniki, kot npr. topografija (v ozkih dolinah je npr. »vidnih« le malo satelitov), vegetacija (strnjene krošnje lahko zastrejo signal s satelitov), lega GPS antene v povezavi z
aktivnostjo živali (če je žival med počitkom npr. na hrbtu, snemanje lokacije ni uspešno, saj je sprejemna GPS antena narobe obrnjena). Zato je uspeh snemanj vselej manjši kot 100 %. Vsi dejavniki, ki vplivajo na uspeh snemanja in se v prostoru in času ne pojavljajo naključno, pa pomenijo bias v posnetih podatkih in
potencialno slabšajo rezultate raziskave. Za odstranitev biasa smo v pričujoči raziskavi za vse posnete lokacije in neuspešne poskuse snemanj pripravili vrsto neodvisnih spremenljivk, ki bi lahko vplivale na uspeh snemanja: topografija, vegetacijske značilnosti okoli lokacije, vedenje medveda (na osnovi meritve
aktivnosti), pričakovano število vidnih GPS satelitov ob času snemanja lokacije glede na njihovo lego in konfiguracijo terena, tip ovratnice, način prenosa podatkov iz ovratnice do uporabnika. Z logistično regresijo (odvisna spremenljivka: lokacija je ali ni posneta, neodvisne: vse prej naštete spremenljivke) smo za vsako
lokacijo ocenili posteriorno verjetnost uspešnega snemanja lokacije in inverzne vrednosti te verjetnosti uporabili kot utež pri naključnem izboru lokacij. Logistični regresijski model kaže, da je na uspeh snemanja najbolj vplival tip ovratnice (vsaka novejša generacija je boljša), vedenje medveda (ko so počivali, je bil uspeh
slabši), okoliška vegetacija in pričakovano število vidnih satelitov v času snemanja lokacije. Z odstranitvijo biasa smo osnovni set lokacij zmanjšali za približno petino, iz prek 100 000 na okoli 82 000 (Jerina, v pripravi).

65
5.3.2 Enourni premiki medvedov
5.3.2.1 Analiza podatkov
Da bi ugotovili, kateri dejavniki vplivajo na gibanje medvedov, smo izračunali evklidske razdalje med sosednjimi lokacijami v enournih intervalih. Kadar so bile lokacije zajete v daljših časovnih intervalih oz. v
primeru manjkajočih lokacij (ko se GPS sprejemnik ni uspel povezati s sateliti), smo upoštevali tudi razdalje v daljših intervalih, če ti niso bili daljši kot tri ure. Vsakemu takemu intervalu smo priredili enourno razdaljo, ki smo jo izračunali s pomočjo korekcijskih faktorjev. Te smo izračunali tako, da smo naredili regresijo med
povprečnimi enournimi razdaljami in pripadajočimi evklidskimi razdaljami v daljših časovnih intervalih (2h, 3h). Pri regresiji smo uporabili »loss funkcijo« z najmanjšo vsoto absolutnih odstopanj. Kadar je bil interval daljši kot tri ure (manjkale so vsaj tri zaporedne enourne lokacije), smo ga izpustili iz analize. Vsaki razdalji
smo priredili čas sredine pripadajočega časovnega intervala.
Za izračun cirkadianih (znotraj dneva) razdalj smo zaradi sezonskega nihanja dolžine dneva/noči čas relativizirali – izračunali smo, v katerem delu dneva oz. noči relativno glede na položaj sonca je bila enourna razdalja zabeležena, tako da smo upoštevali dolžino dneva/noči med sončnim vzhodom in zahodom. Pri tem
smo upoštevali podatke o civilnem sončnem vzhodu/zahodu1, to je čas, ko je center sonca 6° pod horizontom oz. ko običajno prižgemo oz. ugasnemo luči.
Za ugotavljanje razlik v povprečnih prehojenih enournih poteh med medvedi in med kategorijami medvedov, smo razdalje standardizirali glede na sezono (mesece), ko je bil osebek spremljan. Ker so bili
osebki spremljani različno dolga obdobja in v različnih sezonah, bi to namreč lahko vplivalo na rezultate. Za vsak mesec smo pri vsakem osebku izračunali povprečne urne prehojene razdalje in nato mesečna povprečja za vse osebke (izločili smo mesece pri osebkih, v katerih je bilo zabeleženih manj kot 40 lokacij).
Nato smo izračunali skupno povprečje mesečnih povprečij in glede na to povprečje vsakemu mesecu priredili ustrezen korekcijski faktor (mesece smo dali na »skupni imenovalec«). Vsako urno razdaljo smo nato korigirali glede na mesec, v katerem je bila razdalja zabeležena. Za testiranje značilnosti razlik enournih
razdalj med posameznimi skupinami medvedov smo uporabili neparametrične teste (Mann-Whitney U test in Kruskal-Wallis test).
5.3.2.2 Rezultati
Povprečna razdalja v enournem intervalu za vse osebke znaša 248 m (razpon od 0 m do 9783 m), kar pomeni povprečno dnevno razdaljo 5957 m (vsota razdalj v enournih intervalih). V cirkadianem ritmu
gibanja osebkov sta dva maksimuma, prvi je zjutraj, med 4:00 in 5:00, ko se medvedi povprečno vsako uro premaknejo za 382 m, najdaljši povprečni urni premik (462 m) pa je zvečer med 19:00 in 20:00. Najkrajše urne razdalje (68 m) so sredi dneva med 13:00 in 14:00 (Slika 23Error! Reference source not found.). Ob
upoštevanju relativnega časa sta maksimuma cirkadianih razdalj ob sončnem vzhodu (391 m) in sončnem zahodu (457 m; Slika 24).
1 http://www2.arnes.si/~gljsentvid10/ mrak_izracun_zahoda.html

66
Razdalje urnih premikov so odvisne tudi od sezone. Razlike v razdaljah med meseci so statistično značilne (p < 0,001). Največjo povprečno urno razdaljo smo zabeležili v mesecu juniju (356 m), najmanjšo pa januarja (71 m oz. 106 m, če izvzamemo vse lokacije, ko so osebki zimovali in se niso premikali; Slika 25). Glede na
sezono se spreminja tudi cirkadiano nihanje razdalj (t.j. razlika v gibanju v različnih delih dneva). Najbolj poudarjena nihanja smo zaznali v obdobju od sredine maja do sredine avgusta, najmanjše nihanje pa v obdobju od sredine novembra do sredine februarja (Slika 26).
Slika 23: Cirkadiano nihanje urnih razdalj med zaporednimi lokacijami medvedov.
Slika 24: Cirkadiano nihanje urnih razdalj med zaporednimi lokacijami medvedov v relativnem času glede na položaj
sonca.

67
Slika 25: Letna dinamika povprečnih urnih gibalnih razdalj.
Slika 26: Sezonske razlike v urnih razdaljah in njihovem cirkadianem nihanju.

68
Slika 27: Sezonska dinamika razdalj gibanja glede na spol in prisotnost ter starost mladiča (pri samicah). Za vsako
kategorijo je v oklepajih navedeno število spremljanih osebkov. Skupno število spremljanih osebkov je večje kot število
spremljanih medvedov, saj so nekatere samice v različnih obdobjih spremljanja pripadale različnim kategorijam.
Razlike v standardiziranih urnih razdaljah med posameznimi osebki so statistično značilne (p < 0,001).
Največja dejanska povprečna urna razdalja osebka ne glede na obdobje in trajanje spremljave osebka znaša 673 m (Rožnik), najmanjša pa 43 m (Živa). Pri medvedih, ki so bili spremljani vsaj 6 mesecev, je najdaljša povprečna urna razdalja 450 m (Borut), najkrajša pa 71 m (Anka).
Samci so se v urnih intervalih v povprečju premikali 300 m, samice 216 m, vendar razlike med spoloma niso
statistično značilne (p = 0,612). Če upoštevamo samo obdobje, ko osebki niso zimovali in so se dejansko premikali, pa so razlike (povprečje znaša 323 m za samce in 231 m za samice) blizu statistični značilnosti (p = 0,072).
Razlike v urnih razdaljah med konfliktnimi in nekonfliktnimi medvedi so statistično značilne (p < 0,001).
Povprečna urna razdalja znaša pri konfliktnih medvedih 480m, medtem ko se nekonfliktni medvedi v eni uri v povprečju premakanili 245 m. Večje ugotovljene razdalje pri konfliktnih medvedih so verjetno posledica njihove preselitve, saj so preseljeni medvedi nekaj časa po preselitvah pogosto premikajo dlje in intenzivneje
od ostalih.
Ugotavljali smo tudi razlike med standardiziranimi razdaljami samic glede na prisotnost in starostno kategorijo mladiča. Samice, za katere smo imeli informacijo o prisotnosti mladiča smo razdelili v 3 skupine: brez mladiča, z mladičem v 1. letu starosti, z mladičem v 2. letu starosti. Razlike med skupinami v splošnem
niso statistično značilne (p = 0,99). Za obdobje izven zimovanja pa so razlike med skupinami statistično značilne (p = 0,026), povprečne vrednosti razdalj pa znašajo: 266 m (nevodeče samice), 199 m (samice z mladičem 0+) in 203 m (samice z mladičem 1+). Razlike med razdaljami obeh skupin vodečih samic (z mladiči

69
0+ in 1+) tudi izven obdobja zimovanja niso statistično značilne (p = 0,102). Sezonsko dinamiko povprečnih urnih razdalj samic glede na prisotnost in starost mladiča ter samcev prikazujemo na Slika 27. Upoštevano je samo obdobje, ko medvedi niso zimovali. Meseci z manjkajočimi povprečji pomenijo, da v tistem mesecu
Testirali smo tudi razlike med urnimi premiki nevodečih samic, samic z mladičem 0+, samic z mladičem 1+ ter samcev za obdobje parjenja (1.5. do 15.6.). Razlike so statistično značilne (p < 0,001), vrednosti po padajočem vrstnem redu pa znašajo: 448 za samce, 313 za samice z mladičem 1+, 309 za samice brez
mladiča in 251 za samice z mladičem 0+. nismo spremljali osebka iz dotične skupine oz. da so vsi osebki te kategorije zimovali.
5.3.3 Aktivnost medveda
Podatke o aktivnosti smo dobili iz ovratnic 21-ih medvedov. Ovratnice so beležile nivo aktivnosti osebkov vsakih 5 minut s pomočjo senzorja gibanja, ki zaznava pospeške ovratnice v dveh smereh. Interval vrednosti je v obeh smereh od 0 do 255. Vsaki zabeleženi vrednosti smo po metodi, ki so jo opisali Gervasi in sod.
(2006), priredili binarni zapis: aktivno/neaktivno. Ta metoda upošteva individualne značilnosti osebkov, zato je klasifikacija za vsak osebek enolična. Vrednosti v 5-minutnih intervalih smo nato združili v urna povprečja (sredina urnega intervala je polna ura). Prikazane vrednosti aktivnosti torej pomenijo povprečne deleže
časovnega intervala (ure, meseca), ko je bil osebek aktiven. Podobno kot pri razdaljah smo dodatno računali še aktivnosti v relativnem času glede na položaj sonca. 5-minutne intervale smo relativizirali in nato izračunali povprečja za 20 časovno relativiziranih intervalov v dnevu. Za testiranje razlik med skupinami smo
na enak način kot pri razdaljah aktivnosti predhodno standardizirali po mesecih.
5.3.3.1 Rezultati
Povprečje vseh aktivnosti znaša 0,35, kar pomeni, da so bili medvedi aktivni 35 % časa. Upoštevaje samo obdobje, ko niso zimovali, so bili aktivni 37 % časa.
Cirkadiano nihanje povprečij aktivnosti se ujema s cirkadianim nihanjem pri urnih razdaljah premikov
medvedov. Dnevna maksimuma aktivnosti nastopita ob sončnem vzhodu (v povprečju ob 5:00) in ob sončnem zahodu (v povprečju ob 19:00), z večjimi vrednosti ob večernem višku. Dnevni minimum aktivnosti nastopi sredi oz. takoj na začetku druge polovice dneva (v povprečju ob 13:00), drugi minimum pa nastopi v
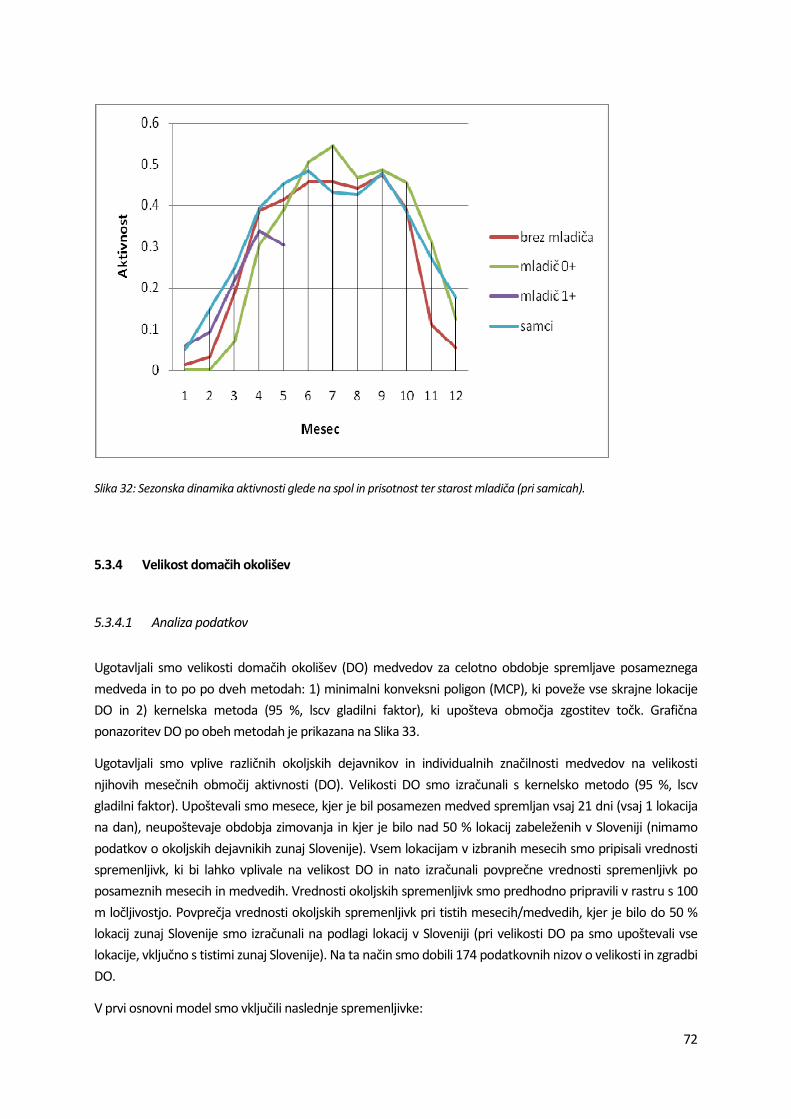
drugi polovici noči (Slika 28Slika 29). Vzorci cirkadianih aktivnosti se skozi leto spreminjajo. Najbolj aktivni so bili osebki v pomladno-poletnem obdobju, ko smo zaznali tudi največja cirkadiana nihanja aktivnosti. Najmanjši nivo in najmanjše cirkadiano nihanje aktivnosti je značilno za zimsko obdobje (Slika 30).
Natančnejše letno nihanje povprečnih aktivnosti prikazuje Slika 31.
Razlike med standardiziranimi vrednostmi aktivnosti pri 21 medvedih so statistično značilne (p < 0,001). Povprečna standardizirana vrednost aktivnosti je pri medvedu z največjo vrednostjo (Bohinjc) kar 19-krat večja kot pri najmanj aktivnem medvedu (Živa).

70
Slika 28: Cirkadiano nihanje aktivnosti.
Slika 29: Cirkadiano nihanje aktivnosti v relativnem času.

Slika 30: Sezonske razlike v vzorcih cirkadianih aktivnosti.
U
vam
ag
S
lika 31: Sezonsko nihanje v povprečni mesečni aktivnosti.71
poštevaje celotno obdobje spremljave so razlike med spoloma statistično značilne (p < 0,001), samci so bili
primerjavi s samicami 1,28-krat bolj (dlje časa) aktivni. V obdobju izven zimovanja nismo odkrili razlik v ktivnosti med spoloma (p = 0,449). Razlike med samicami glede na prisotnost in starostno kategorijo ladiča so značilne tako za celotno obdobje spremljave kot tudi za obdobje izven zimovanja; najmanj
ktivne so bile samice z mladičem v prvem letu starosti. Razlike v sezonski dinamiki aktivnosti med samicami lede na prisotnost in starost mladiča ter samci so razvidne iz Slika 32.
5
5
U
mDp
U
ngn
psp
mll
D
V
S
lika 32: Sezonska dinamika aktivnosti glede na spol in prisotnost ter starost mladiča (pri samicah).72
.3.4 Velikost domačih okolišev
.3.4.1 Analiza podatkov
gotavljali smo velikosti domačih okolišev (DO) medvedov za celotno obdobje spremljave posameznega
edveda in to po po dveh metodah: 1) minimalni konveksni poligon (MCP), ki poveže vse skrajne lokacije O in 2) kernelska metoda (95 %, lscv gladilni faktor), ki upošteva območja zgostitev točk. Grafična onazoritev DO po obeh metodah je prikazana na Slika 33.
gotavljali smo vplive različnih okoljskih dejavnikov in individualnih značilnosti medvedov na velikosti
jihovih mesečnih območij aktivnosti (DO). Velikosti DO smo izračunali s kernelsko metodo (95 %, lscv ladilni faktor). Upoštevali smo mesece, kjer je bil posamezen medved spremljan vsaj 21 dni (vsaj 1 lokacija a dan), neupoštevaje obdobja zimovanja in kjer je bilo nad 50 % lokacij zabeleženih v Sloveniji (nimamo
odatkov o okoljskih dejavnikih zunaj Slovenije). Vsem lokacijam v izbranih mesecih smo pripisali vrednosti premenljivk, ki bi lahko vplivale na velikost DO in nato izračunali povprečne vrednosti spremenljivk po osameznih mesecih in medvedih. Vrednosti okoljskih spremenljivk smo predhodno pripravili v rastru s 100
ločljivostjo. Povprečja vrednosti okoljskih spremenljivk pri tistih mesecih/medvedih, kjer je bilo do 50 % okacij zunaj Slovenije smo izračunali na podlagi lokacij v Sloveniji (pri velikosti DO pa smo upoštevali vse okacije, vključno s tistimi zunaj Slovenije). Na ta način smo dobili 174 podatkovnih nizov o velikosti in zgradbi
O.
prvi osnovni model smo vključili naslednje spremenljivke:

• okoljski dejavniki: populacijska gostota medvedov (GOST_MEDV), oddaljenost od gozdnega roba (navznoter in navzven; ODD_G_ROB), oddaljenost od najbližje hiše (ODD_HISE), oddaljenost od najbližje glavne ceste (ODD_CESTE), oddaljenost od najbližjega krmišča (ODD_KRM)
• ostale spremenljivke: dni spremljan (21 do 31 dni; DNI_SPREM), mesec spremljanja (MESEC),
kategorija medveda (6 kategorij: samice brez mladiča, samice z mladičem 0+, samice z mladičem 1+, samci do 4 leta, samci nad 4 leta, ostalo-ni podatka; KAT_MEDV), interakcija MESEC*KAT_MEDV
Poleg osnovnega modela (model 1) smo zgradili tudi model, kjer nismo vključili spremenljivke GOST_MEDV (ta korelira z ODD_KRM, zanimal nas je morebitni vpliv krmišč ob odstranitvi gostote medvedov; model 2).
Analize smo opravili z uporabo multivariatne statistične metode GRM (general regression model) in
algoritma forward stepwise v programu Ststistica 7.0. Zaradi zahtev po normalnosti porazdelitve smo odvisno spremenljivko – velikost mesečnega DO – logaritmirali.
5
Č
P2p
S
lika 33: Lokacije ter območje aktivnosti medveda Pečka po MCP in kernelski metodi.73
.3.4.2 Rezultati
as spremljave se pri medvedih močno razlikuje (min = 20 dni, max = 449 dni), kar vpliva tudi na velikost DO.
ovprečna velikost DO po MCP metodi znaša 35.038 ha (405 ha do 314.403 ha) in po kernelski metodi 6.788 ha (221 ha do 308.007 ha). Velikost DO posameznih medvedov in obdobje njihove spremljave rikazuje Preglednica 15.

74
Preglednica 15: Velikost območij aktivnosti medvedov v obdobju spremljave.
Ime medveda MCP (ha) Kernel 95% (ha) Čas spremljave (dni)
ŽIVA 405 221 69
PEPCA 1941 804 317
FRANCE 2624 1879 52
EVA 3186 1287 446
MIRKA 4009 1951 380
ANKA 4965 2020 229
MARJETA 5121 2222 240
BORA 5394 3388 229
JANI 8079 7751 55
ANA SNEŽNIŠKA 11131 6942 336
GORANKA 11673 4100 366
KATJA 13309 9081 183
ZORAN 14403 18305 39
NEŽA 15861 27354 20
HOBI 16079 4995 409
KAREL 17396 30684 21
ZORA 17442 1666 277
BOHINJC 20265 28929 27
ALJOŠA 20975 13509 154
NEJC 21184 7453 156
HRIBAR 23476 35622 27
IGOR 24770 33107 124
ANA RAKIŠKA 28791 11737 288
SENOŽEČANKA 30682 20574 388
PEČKO 34458 12973 449
(se nadaljuje)

75
Preglednica 15 (nadaljevanje)
PETER 37316 31451 206
ALOJZIJ 39724 21453 181
VALENTIN 42787 21101 354
BORUT 66705 26338 355
BEPI 85277 79557 144
ANDREA 90135 50389 126
BRANKO 122274 57149 227
ROŽNIK 314403 308007 44
Povprečje 35038 26788 210
5.3.4.3 Vplivi okolja in individualnih značilnosti/razlik na velikost mesečnih območij aktivnosti
Če izvzamemo mesece intenzivnega zimovanja (december, januar, februar), ko zaradi opisane metode
nismo zbrali dovolj podatkov o mesečnih DO, se velikosti povprečnih mesečnih DO gibljejo med 3721 ha meseca oktobra in 8353 ha meseca maja. Sezonsko variiranje povprečnih velikosti mesečnih DO prikazuje
Slika 35.
Slika 35: Sezonsko (marec do november) nihanje povprečnih velikosti mesečnih domačih okoliših.

76
Model 1 (vse spremenljivke) pojasni 57 % variabilnosti v velikosti mesečnih DO. Napoveduje, da je velikost mesečnega DO odvisna od naslednjih dejavnikov (navedeno po padajoči jakosti vpliva; glej tudi Preglednica 16):
• gostota medvedov
• kategorija medveda
• mesec spremljanja
• dni spremljave
Model 2 (brez populacijske gostote medvedov) pojasni 42 % variabilnosti v velikosti mesečnih DO. Napoveduje, da je velikost mesečnega DO odvisna od naslednjih dejavnikov (navedeno po padajoči jakosti
vpliva; glej tudi Preglednica 16):
• kategorija medveda
• mesec spremljanja
• oddaljenost od najbližjega krmišča
5.4 RABA KRMIŠČ
5.4.1 Analiza podatkov
Za vsako zabeleženo GPS lokacijo medvedov smo ugotavljali, ali je bila posneta na krmišču ali ne. GIS plast vseh zabeleženih lokacij medvedov znotraj meja Slovenije smo prekrili z GIS plastjo lokacij krmišč. Za vsako
lokacijo medvedov smo nato ugotavljali oddaljenost od najbližjega krmišča in oznako tega krmišča. Natančnost lokacij krmišč ima lahko določeno napako, razen tega pa se krma znotraj krmišča polaga na različnih mikrolokacijah. Ugotovili smo, da zgostitev lokacij medvedov z razdaljo od teoretičnih lokacij krmišč
nehomogeno upada oz. da je pri razdalji 140 m zaznati močan prelom. Za vse lokacije medvedov, ki od teoretične lokacije krmišča niso bile oddaljene več kot 140 m, smo zato privzeli, da so bile zabeležene na krmišču.
Ugotavljali smo ali na rabo krmišč vplivajo naslednji dejavniki:
• individualne razlike med osebki (n = 33)
• prisotnost in starost mladičev pri samicah in starostni razred pri samcih; 5 kategorij: nevodeče
samice, samice z mladičem 0+, samice z mladičem 1+, mlajši samci (do 4 leta), starejši samci (5 let ali več)
• ura v dnevu (12 kategorij)
• mesec (12 kategorij)
• status lune (zvezno)
Vplive vseh navedenih dejavnikov na rabo krmišč smo ugotavljali s pomočjo multivariatne analize. Uporabili smo binarno logistično regresijo, kjer je odvisno spremenljivko predstavljala pripadnost lokacije na krmišču

77
(je/ni na krmišču). Vse neodvisne spremenljivke smo v regresijo ponudili kot kategorialne, razen statusa lune, ki smo ga upoštevali kot zvezno kosinus funkcijo (interval med dvema mlajema). Ugotavljali smo vpliv dejavnikov na rabo krmišč enkrat za celotno obdobje spremljanja medvedov in enkrat samo za obdobje, ko
medvedi niso zimovali (niso bili v brlogu).
Za prikaz razlik med osebki v rabi krmišč smo vrednosti (delež lokacij na krmišču) standardizirali glede na obdobje spremljave osebka. Za vsak osebek smo izračunali kolikšen delež lokacij je bil na krmišču v posameznem mesecu, ko je bil osebek spremljan. Nato smo izračunali mesečna povprečja vseh vrednosti,
vendar pri tem nismo upoštevali osebkov, ki so bili spremljani manj kot 4 mesece in tistih mesecev pri posameznih osebkih, kjer je bilo zabeleženo 20 ali manj lokacij. Iz mesečnih povprečij smo nato izračunali skupno povprečje ter izračunali korekcijske faktorje za posamezne mesece (količnik med skupnim
povprečjem in povprečjem posameznega meseca). Nato smo pri vsakem osebku v posameznem mesecu spremljave vrednosti rabe krmišč korigirali s korekcijskimi faktorji. Skupno standardizirano povprečno rabo krmišč posameznega medveda smo izračunali kot ponderirano povprečje korigiranih mesečnih vrednosti.
Kot ponder smo uporabili število lokacij v posameznem mesecu, ko je bil osebek spremljan. Postopek smo izvedli enkrat za celotno obdobje spremljave, vključno z zimovanjem, in enkrat samo za obdobje izven zimovanja.
Izračunali smo tudi število različnih obiskanih krmišč na noč, mesec, obdobje spremljave za posamezne
medvede. Glede na to, da medvedi krmišča obiskujejo predvsem ponoči, smo dnevni interval zamaknili za 12 ur (od 12:00 prvega do 12:00 naslednjega dneva). Ponovno smo pri posameznem medvedu upoštevali samo tisto obdobje spremljave, ko je bil medved v Sloveniji, ker za druge države nismo imeli podatkov o lokacijah
krmišč. Pri ugotavljanju števila obiskanih krmišč v posameznih mesecih smo upoštevali samo tiste mesece pri posameznih medvedih, ko je bil medved spremljan vsaj 5 dni. Za boljše razumevanje podatkov iz spremljanja (štetja) medvedov na stalnih števnih mestih, smo posebej analizirali tudi rabo krmišč, ki se uporabljajo kot
stalna števna mesta (»števna krmišča«).
5.4.2 Dejavniki, ki vplivajo na rabo krmišč pri medvedih
Od 100.466 v analizo vključenih lokacij medvedov, jih je bilo 6.844 (6,8 %) na krmiščih (pozitivne lokacije),
medtem, ko jih je 93.622 (93,2 %) padlo izven krmišč (negativne lokacije). Za nadaljnjo analizo smo vzorec uravnotežili, tako da smo vsem negativnim lokacijam pripisali take ponderje, da je bila ponderirana vsota negativnih lokacij enaka številu pozitivnih lokacij (6.844).
Oba modela (kjer smo upoštevali celotno obdobje spremljave medvedov ali samo obdobje izven zimovanja),
napovedujeta, da na rabo krmišč vplivajo vsi vključeni dejavniki, razen statusa lune. Po padajoči jakosti vpliva (glede na Waldovo statistiko) si dejavniki sledijo:
• ura v dnevu
• osebek
• mesec
• status samic oz. starostni razred samcev
Oba modela pravilno razvrstita 70,5 % vseh lokacij (65 % oz. 63,3 % pozitivnih in 76 % oz. 77,2 % negativnih). R2 pri obeh modelih znaša 0,28.

Medvedi so bili na krmišču najpogosteje zvečer (višek okrog 21:00 ure z 16,5 % lokacij), preko noči pa delež lokacij postopoma upada. Najredkeje so se na krmiščih zadrževali med 7:00 uro in 17:00 uro, ko je bil delež lokacij na krmiščih med 1,4 % in 3,1 %. Dnevno dinamiko rabe krmišč prikazuje Slika 36.
D
zso
S
S
lika 36: Dnevna dinamika rabe krmišč.lika 37: Dnevna dinamika rabe krmišč pri posameznih kategorijah medvedov.
78
nevna dinamika se razlikuje glede na kategorijo medveda. Samice z mladičem so se najpogosteje
adrževale na krmišču okrog 20:00, samice brez mladiča in mlajši samci okrog 21:00, medtem ko so starejši amci v povprečju obiskovali krmišča najkasneje, okrog 22:00. V dnevnih urah so krmišča najpogosteje biskovale samice z mladičem 0+, najmanj pa starejši samci (Slika 37).

Razlike med osebki v rabi krmišč smo ugotavljali ločeno za celotno obdobje spremljave posameznega medveda ter za obdobje izven zimovanja. Ker sta standardizacija in ponderiranje mesečnih povprečij pri obeh načinih različna (glej poglavje 5.4.1), vrednosti ne moremo neposredno medsebojno primerjati in ju
zato prikazujemo ločeno (Slika 38 inSlika 39). Upoštevaje celotno obdobje spremljave je krmišča najpogosteje uporabljal samec Pečko (22,9 % lokacij). Če upoštevamo samo obdobje, ko osebki niso zimovali, pa je bila raba krmišč največja pri Živi (28,3 %). Vendar je potrebno omeniti, da je bila ta medvedka
izven zimovanja spremljana zgolj 3 dni in se je v tem obdobju razmeroma veliko časa zadrževala na krmiščih. Zaradi kratkega obdobja spremljave smo Živo izpustili iz prikazov individualnih razlik na Slika 38Slika 39.
Slika 38: Individualne razlike v rabi krmišč (standardizirano) v celotnem obdobju spremljave.
S
lika 39: Individualne razlike v rabi krmišč (standardizirano) samo za obdobje, ko medvedi niso zimovali.79

80
Najpogosteje so medvedi krmišča obiskovali aprila (9,3 % vseh lokacij), najmanj pa decembra (3,7 % lokacij). Če upoštevamo samo lokacije, ko medvedi niso zimovali, pa je največji delež lokacij na krmiščih januarja (12,1 %) in najmanjši še vedno decembra (5,3 %). Sezonsko dinamiko rabe krmišč prikazuje Slika 40.
S
Med obravnavanimi kategorijami medvedov so krmišča najpogosteje obiskovali mlajši samci (10,0 %) in
samice brez mladičev, najmanjši delež lokacij na krmišču pa je bil pri samicah z mladičem v 1. letu starosti (5,1 %). Raba krmišč po kategorijah medvedov je prikazana na Slika 41.
S
Slika 41: Raba krmišč za posamezne kategorije medvedov.
lika 15: Sezonska dinamika rabe krmišč lika 40: Sezonska dinamika rabe krmišč.

5.4.3 Število obiskanih krmišč
5.4.3.1 Število obiskanih krmišč na noč
Medvedi so na noč (od poldneva do poldneva) obiskali od 0 do 6 krmišč (od 6360 spremljanih noči je bilo sicer 6 krmišč obiskanih samo dvakrat). Večino vseh spremljanih noči (znotraj meja Slovenije) medvedi niso
obiskali nobenega krmišča (62,1 %), eno krmišče so obiskali v 31,1 % nočeh, dve krmišči v 5,7 % nočeh ter 3 ali več krmišč v 1,0 % vseh noči. Frekvenčno porazdelitev števila obiskanih krmišč/noč za posamezne medvede ter skupno prikazuje Slika 42.
P
sn
S
S
lika 43: Standardizirano povprečno število obiskanih krmišč na noč v obdobju spremljanja posameznega medveda.lika 42: Delež noči v obdobju spremljanja, ko je posamezen medved obiskal 0, 1, 2 oz. 3 ali več krmišč.
81
ovprečno število obiskanih krmišč na noč za posameznega medveda prikazuje Slika 43. Glede na to, da je
ezonska dinamika obiskovanj različnih krmišč (št. krmišč/noč) zelo poudarjena (največ aprila in junija, ajmanj januarja in decembra; Slika 44), smo povprečne vrednosti predhodno standardizirali glede na

82
sezono in trajanje spremljanja posameznega medveda. Povprečje vseh spremljanih noči (6360) znaša 0,46 obiskanih krmišč na noč.
5.4.3.2 Število obiskanih krmišč na mesec
Povprečno število obiskanih različnih krmišč na mesec za vse mesece in vse medvede znaša 4,1. Največ
različnih krmišč so v povprečju obiskali junija (5,9), najmanj pa januarja (0,5). Vrednosti pri posameznih medvedih v posameznih mesecih znašajo od 0 do 13 različnih obiskanih krmišč. Po 13 različnih krmišč so v enem mesecu obiskali Ana Rakiška, Rožnik, Nejc in Hobi. Mesečna povprečja obiskanosti različnih krmišč
prikazuje Slika 45. Za prikaz razlik med medvedi v mesečni obiskanosti različnih krmišč smo vrednosti pri posameznih medvedih standardizirali glede na obdobje njihovega spremljanja (Slika 46).
S
S
lika 45: Povprečno število različnih obiskanih krmišč po mesecih.lika 44: Povprečno število obiskanih krmišč na noč po mesecih.

5.4.3.3 Število obiskanih krmišč v celotnem obdobju spremljanja
Posamezni medvedi so v celotnem obdobju njihovega spremljanja obiskali zelo različno število krmišč, od 1 (Živa) do 35 (Pečko). Ker je število obiskanih krmišč odvisno tudi od trajanja spremljanja, na Slika 47 poleg števila različnih obiskanih krmišč pri posameznih medvedih prikazujemo tudi trajanje njihovega spremljanja
v mesecih.
Pkk
S
Slika 47: Število različnih obiskanih krmišč za obdobje spremljanja posameznega medveda v Sloveniji.
lika 46: Standardizirano povprečno število obiskanih krmišč na mesec v obdobju spremljanja posameznega medveda.
83
reverili smo tudi ali obstaja korelacija med rabo krmišč (delež lokacij na krmišču) in obiskanostjo različnih rmišč (število krmišč na mesec) pri posameznih medvedih. Povezavo smo potrdili s Spearmanovim orelacijskim koeficientom (α = 0,02).

84
5.4.4 Raba krmišč znotraj domačih okolišev posameznih medvedov
Preverjali smo kolikšen delež krmišč, ki so posameznemu medvedu bila na razpolago znotraj meja njegovega domačega okoliša (znotraj MCP) je medved dejansko uporabljal. Pri tem smo upoštevali samo lokacije
medvedov in krmišč v Sloveniji. Iz analize smo izločili medvede, ki so bili v Sloveniji spremljani manj kot 4 mesece.
V povprečju so medvedi uporabljali 39 % vseh krmišč znotraj njihovih domačih okolišev. Največji delež krmišč je uporabljala medvedka Marjeta (63 %), najmanj pa Bepi (8 %; Slika 48). Območja aktivnosti
posameznih medvedov ter rabo krmišč glede na njihovo razpoložljivost prikazuje Slika 49. Samice so v povprečju uporabljale 46 % vseh krmišč znotraj njihovih območij aktivnosti, medtem ko so samci uporabljali 28 % razpoložljivih krmišč.
Slika 48: Delež obiskanih krmišč od vseh razpoložljivih krmišč znotraj območja aktivnosti (MCP) posameznega medveda.

85
Slika 49: Območja aktivnosti medvedov v Sloveniji in lokacije krmišč, ki jih je vsaj enkrat obiskal vsaj en spremljan medved
(zelene pike) ter nikoli obiskanih krmišč (rdeče pike) znotraj območij.
5.4.4.1 Raba različnih števnih krmišč
Večino spremljanih noči v vseh sezonah medvedi niso obiskali nobenega števnega krmišča (88,2 % vseh
noči), eno števno krmišče so obiskali v 11,6 % noči in 2 števni krmišči v 0,3 % noči. Samo 5 medvedov je vsaj enkrat obiskalo 2 števni krmišči na noč. Noben medved ni nikoli obiskal več kot 2 števni krmišči na noč. Frekvenčno porazdelitev obiskanih števnih krmišč na noč za posameznega medveda prikazuje Slika 50.
Preverjali smo kakšna je bila obiskanost števnih krmišč med 18:00 in polnočjo (v tem času poteka štetje) na
dan štetja ter do 3 dni pred tem datumom in do 3 dni po njem (torej v 7 zaporednih dneh). Pri vsakem medvedu smo upoštevali vsa štetja, ki so se izvajala v času njegovega spremljanja. Med 33 medvedi je bilo 8 takih, ki nikoli niso bili spremljani v obdobju štetja oz. v teh dnevih med 18:00 in polnočjo nismo zabeležili
nobene lokacije. Rezultati se zato nanašajo na 25 medvedov.

86
Slika 50: Delež noči v obdobju spremljanja medveda, ko je ta obiskal 0,1,2 števnih krmišč.
Medvedi so med 18:00 in polnočjo obiskali števna krmišča v 9 % vseh števnih (+-3) dni. 9 medvedov ni v obravnavanem
obdobju in času dneva nikoli obiskalo števnega krmišča. Največji delež dni z obiskanimi krmišči je bil 28,6 % (Slika 51).
Slika 51: Delež števnih dni, ko so medvedi med 18:00 in 0:00 obiskali števno krmišče.
Glede na spol medvedov in status samic oz. starostno kategorijo samcev so deleži števnih dni (+- 3 dni), ko so
medvedi med 18:00 in 0:00 obiskali števno krmišče sledeči: samice brez mladiča: 14,7 %, samice z mladičem 0+: 8,6 %, samice z mladičem 1+: 4,2 %, samci do 4 leta starosti: 2,0 %, samci starejši od 4 leta: 10,3 %.

87
5.4.5 Raba prostora glede na oddaljenost od krmišč
5.4.5.1 Analiza podatkov
Z namenom vpogleda v vplive krmljenja na prostorsko razporeditev medveda smo analizirali, kako se raba prostora glede na oddaljenost od krmišča spreminja. Ker krmišča v prostoru niso naključno razporejena in z
njihovo oddaljenostjo lahko zato variirajo tudi drugi prostorski dejavniki, ki vlivajo na primernost nekega prostora v smislu potencialnega habitata za medvede, smo analizo izvedli le za gozdne površine. Rabo gozdnega prostora glede na njegovo razpoložljivost v odvisnosti od oddaljenosti do najbližjega krmišča smo
ugotavljali tako, da smo naključno žrebali 50000 medvedjih lokacij znotraj gozda ter v enakih gozdnih zaplatah, kjer ležijo te lokacije naključno izbrali 50000 (ne-medvedjih) lokacij. V različnih intervalih oddaljenosti od krmišč smo nato ugotavljali razmerje med medvedjimi in naključnimi lokacijami.
5.4.5.2 Rezultati
Raba prostora je v oddaljenosti do 200 m od krmišč 4,3-krat večja kot bi pričakovali glede na razpoložljivost
tega prostora, kar je posledica neposrednih obiskov krmišč. Količnik raba/razpoložljivo nato strmo pade in v oddaljenosti 201 – 400 m 1,7. Do intervala 801 – 1000 m je nato dokaj konstanten (pade na 1,5), nato pa začne linearno upadati. Na oddaljenosti večji od 1400 m medvedi prostor uporabljajo manj intenzivno kot bi
pričakovali, če bi bilo gibanje medvedov naključno (Slika 52).
Slika 52: Raba prostora (znotraj gozda) glede na njegovo razpoložljivost v gradientu oddaljenosti od krmišč.

88
5.4.6 Vplivi krmljenja na konfliktnost medveda
5.4.6.1 Analiza podatkov
Ena ključnih tem pričujoče raziskave je ugotavljanje vplivov krmljenja na konfliktnost medveda. Ugotavljali smo ali/kako raba krmišč vpliva na konfliktno obnašanje medvedov. Medvedjo lokacijo smo opredelili za
konfliktno, če bila zabeležena na oddaljenosti manj kot 150 m od najbližje hiše oz. če je bila v dnevnem času izven gozda manj kot 300 m od najbližje hiše. Z neparametrično korelacijo smo analizirali ali obstaja povezava med povprečno konfliktnostjo medvedov in njihovo pogostnostjo obiskovanja krmišč. Ker se med
letom spreminja tako stopnja konfliktnosti medveda kot tudi raba krmišč smo izvedli še dve podobni analizi in ugotavljali povezave med rabo krmišč in konfliktnostjo še za sumarne podatke za vsakega medveda po mesecih in tednih. Z enako statistično metodo smo preverjali povezavo med časom, ki je pretekel od
zadnjega obiska krmišča in deležem konfliktnih lokacij v zadnjih 24 urah (skupno za vse medvede).
5.4.6.2 Rezultati
Povezave med konfliktnostjo 33 medvedov in pogostnostjo obiskovanja krmišč nismo ugotovili za nobeno od opravljenih korelacijskih analiz ne glede na obdobje agregiranja podatkov (cela spremljava, mesec, teden) (Spearmanov r = -0.216, minimalni p = 0,227). Povezava med časom od zadnjega krmljenja in konfliktnostjo v
zadnjih 24 urah je pozitivna in statistično značilna (Spearmanov r = 0.029, p = 0,000; Slika 53). To pomeni, da verjetnost za nastop konfliktne lokacije narašča z daljšanjem obdobja od zadnjega krmljenja, vendar pa se je treba zavedati, da je zelo redko, da medved več dni zapored ne pride na nobeno krmišče, zato opisana
značilna povezava pojasnjuje le malo variabilnosti konfliktnosti medvedov.
Slika 53: Konfliktnost medvedov v zadnjih 24 urah glede na čas do zadnjega obiska krmišča.

89
5.5 PRIBLIŽEVANJE NASELJEM
Ugotavljali smo kako se medvedi izogibajo strnjenim naseljem in hišam. Odstranili smo lokacije, ki ležijo izven Slovenije in lokacije, ko so medvedi zimovali in jih tako v analizah obdržali 75 849. Tem smo priredili enako
število naključno izbranih lokacij. Te smo izbirali na površini, ki od medvedjih lokacij ni oddaljena več kot znaša povprečen dnevni premik medveda (2400 m). Za vse lokacije smo s pomočjo prekrivanja GIS-plasti izračunali njihovo oddaljenost od najbližjega strnjenega naselja in najbližje hiše.
5.5.1 Rezultati
Po posameznih razredih širine 20 m oddaljenosti od hiš oz. strnjenih naselij smo primerjali število medvedjih lokacij in naključno izbranih lokacij in tako dobili indekse rabe prostora glede na njegovo razpoložljivost. Slika
54 prikazuje rabo prostora glede na oddaljenost od najbližje hiše. Z naraščanjem oddaljenosti od hiš raba postopoma narašča in je vse do oddaljenosti 500 m manjša kot bi bila ob naključni rabi prostora. Pri interpretaciji teh podatkov pa je treba paziti, ker je taka raba lahko posledica več dejavnikov: odsotnosti
gozda, velike oddaljenosti od krmišč itd. in ne pa samo posledica izogibanja hišam. Slika 55 prikazuje rabo prostora glede na oddaljenost od strnjenih naselij. Tudi v tem primeru z oddaljevanjem od naselij raba prostora narašča vendar počasneje, saj je še na oddaljenosti 500 m od naselij komaj 35 % pričakovane
naključne rabe.
Slika 54: Razmerje med rabo prostora in njegovo razpoložljivostjo glede na oddaljenost od najbližje hiše.

90
Slika 55: Razmerje med rabo prostora in njegovo razpoložljivostjo glede na oddaljenost od najbližjega strnjenega naselja.
Posamezni medvedi so se različno intenzivno izogibali naseljem oz. hišam. Slika 56 prikazuje kolikšen delež
vseh lokacij posameznega medveda je bil zabeležen v intervalih oddaljenosti od najbližje hiše 0-50 m, 50-150 m in 150-250 m. Med medvedi izstopa Rožnik, pri katerem je kar 24 % lokacij na oddaljenosti od hiš manj kot 250 m. Med lokacijami vseh medvedov je takih lokacij sicer 2 %.
Slika 56: Delež vseh zabeleženih lokacij posameznega medveda v različnih intervalih oddaljenosti od najbližje hiše.
Približevanje hišam ni enako v vseh delih dneva. Slika 57 prikazuje dnevno dinamiko približevanja hišam (do 250 m oddaljenosti). Najpogosteje so se medvedi hišam približali med 22:00 in 2:00 uro. Najmanjkrat so bile v bližini hiš njihove lokacije zabeležene med 8:00 in 17:00 uro.

91
Slika 57: Delež lokacij na oddaljenosti do 250 od najbližje hiše glede na uro v dnevu.
5.6 HABITATNI MODEL RJAVEGA MEDVEDA V SLOVENIJI
5.6.1 Analiza podatkov
Vse medvedje lokacije smo najprej filtrirali oz iz njih izločili bias (glej poglavje 5.3.1). Iz analize smo izločili tudi vse lokacije, ko so medvedi zimovali, saj bi v nasprotnem primeru lahko model preveč poudaril pomen okoljskih dejavnikov, ki vplivajo na izbor brlogov, vendar na splošno niso toliko pomembni za celoletni
habitatni izbor medvedov. Na ta način smo dobili 75 849 medvedjih lokacij. Tem smo priredili enako število lokacij, kjer medvedi domnevno niso bili prisotni, oz. njihove prisotnosti tam nismo zaznali. T.i. (lažno) negativne lokacije smo izbirali na tak način, da smo naboru pozitivnih lokacij vsakega medveda priredili
enako število negativnih lokacij, ki od pozitivnih lokacij niso bile oddaljene več kot 2409 m, kar je povprečen dnevni premik medvedov. To pomeni, da so bile te lokacije medvedu vselej dosegljive. Poleg tega smo določili, da je vsaka negativna lokacija od najbližje pozitivne oddaljena vsaj 200 m (kar je tudi najslabša
resolucija kartiranja nekaterih vhodnih spremenljivk). S tem kriterijem smo povečali verjetnost, da se medved tam v resnici ni nahajal. Na ta način smo dobili 2 kategoriji prostora: prostor, ki, so ga spremljani medvedi z gotovostjo uporabljali in prostor, ki ga medvedi zelo verjetno niso uporabljali, kljub temu, da je bil
zanje razpoložljiv oz. dosegljiv. Pred analizo smo vse lokacije ponderirali tako, da je bila vsota ponderiranih pozitivnih oz. negativnih lokacij posameznega medveda enaka številu dni spremljave medveda. Medvedom, ki so bili spremljani več kot eno leto smo ponderje proporcionalno znižali kot da so bili spremljani 365 dni.
Z analizo smo ugotavljali vpliv različnih, potencialno relevantnih, okoljskih dejavnikov (npr. oddaljenost od
gozdnega roba, oddaljenost od najbližje hiše, oddaljenost od najbližjega krmišča, gozdnatost v različnih prostorskih oknih, itd.) na rabo prostora medvedov. Okoljske dejavnike v obliki vektorskih in rastrskih (100 m x 100 m ločljivost) GIS-plasti smo prekrili s pozitivnimi in negativnimi lokacijami in tako vsaki lokaciji priredili
vrednost posameznega okoljskega dejavnika. Vpliv okoljskih dejavnikov na rabo prostora oz. na prisotnost/odsotnost medvedov v prostoru smo analizirali z binarno logistično regresijo, algoritmom forward stepwise v programskem paketu SPSS 17.0.

92
Želeli smo dobiti tudi napovedi habitatne priljubljenosti prostora (celotne Slovenije) za medveda. Regresijsko enačbo, ki ponazarja smeri in jakosti vplivov pomembnih okoljskih dejavnikov, smo ekstrapolirali na celotno Slovenijo in tako dobili zvezno napoved priljubljenosti prostora v ločljivosti 100 m x 100 m. Ker pa verjetnost
prisotnosti medveda na neki lokaciji ni odvisna samo od habitatne primernosti tiste lokacije, temveč tudi od njene širše okolice, smo vsaki celici priredili povprečno verjetnost te celice in okoliških celic v oknu 5100 m x 5100 m. V naslednjem koraku smo s poligoni povezali območja, ki zajemajo prostor z enako verjetnostjo
rabe prostora. Poligone smo prekrili s (pozitivnimi) medvedjimi lokacijami in izločili poligone z najmanjšo verjetnostjo, ki zajemajo zgolj 5 % medvedjih lokacij. 95 % odstotkov medvedjih lokacij je zajetih v območjih, kjer verjetnost rabe ni manjša od 27 %. Ta območja smo privzeli kot neprečiščene habitatne krpe. Nadalje
smo izločili vse habitatne krpe, ki so premajhne da bi lahko predstavljale trajen habitat medveda. Tako smo odstranili vse krpe, ki so manjše od najmanjše habitatne krpe (21 km2), znotraj katere smo zabeležili medvedje lokacije ter vse krpe, ki so manjše od povprečnega območja aktivnosti medveda (350 km2) ter
hkrati od najbližje sosednje habitatne krpe oddaljene več kot znaša največja oddaljenost med habitatnimi krpami, znotraj katerih smo beležili medvedje lokacije (4,5 km).
5.6.2 Rezultati
Model logistične regresije pravilno napove 65,7 % vseh lokacij, od tega 67,9 pozitivnih in 54,5 negativnih. Med 10 ponujenimi okoljskimi dejavniki model 8 dejavnikov prepozna kot pomembnih. Statistično neznačilen je vpliv višine lesne zaloge plodonosnih listavcev in oddaljenosti od gozdnega roba navznoter (v
gozd). Habitatna primernost prostora za medveda narašča z (navedeno po padajoči jakosti vpliva; glej tudi Preglednica 16):
• deležem gozda v okolici osnovne celice (okno velikosti 900 m x 900 m)
• upadanjem oddaljenosti od najbližjega krmišča
• oddaljenostjo od najbližje hiše
• prisotnostjo gozda v osnovni celici (100 m x 100 m)
• manjšanjem oddaljenostjo od gozdnega roba (izven gozda)
• večanjem velikosti (najbližje) gozdne zaplate
• oddaljenostjo od najbližjega strnjenega naselja
• oddaljenostjo od najbližje glavne ceste
Pri tem je potrebno izpostaviti, da smo kot največjo oddaljenost od najbližje hiše, naselja, ceste postavili mejno vrednost 2000 m, to pomeni, da smo vsem vrednostim večjim od 2000 m priredili to vrednost. Na
podlagi preteklih raziskav (Güthlin et al. 2011) in na podlagi univariatne grafične primerjave med oddaljenostjo od omenjenih dejavnikov in rabo prostora medveda smo ocenili, da vpliv teh dejavnikov ne more segati dlje od 2000 m. Na enak način smo največjo oddaljenost od gozdnega roba navzven omejili na
300 m.
Slika 58 (zgoraj) prikazuje habitatni model medveda za Slovenijo. Celotno potencialno območje razširjenosti medveda smo razdelili na tri kategorije glede na potencialno gostoto medvedov. Med vsemi medvedjimi

93
lokacijami znotraj območja razširjenosti je bilo 95 % lokacij zabeleženih znotraj meja oranžno obarvanih območij (srednja gostota) oz. 65 % lokacij zabeleženih na rjavo obarvanih območjih (visoka gostota). Za primerjavo z današnjim stanjem prikazujemo tudi sliko današnjih populacijskih gostot medveda. Zaradi
različnega načina izdelave obeh slik je smiselna zgolj njuna grobo orientacijska primerjava iz katere je razvidno, da so največje razlike med potencialno in aktualno razširjenostjo medveda na območjih, ki so najdlje stran od meje s Hrvaško (npr. Pohorje), kar je verjetno posledica dejstva, da je bil medvd v Sloveniji
pred okoli 100 leti skoraj iztrebljen in je preživel le ob meji s Hrvaško, kasneje se je širil v druge dele države, vendar pa je pri njem širjenje počasno, saj je vrsta izrazito filopatrična, poleg tega pa se je njegovo širjenje zadrževalo z odstrelom prek omejevanja številčnosti in tudi ciljno – do leta 1993 je bila vrsta zunaj
osrednjega območja brez zaščite.
Preglednica 16: Rezultati habitatnega modela za rjavega medveda v Sloveniji s parametri spremenljivk, njihova
standardna napaka, pomembnost vpliva po Waldovi statistiki in stopnja značilnosti.
Dejavnik Parameter (B) Stand. napaka
Wald st. prost. p Exp(B)
gozd_900x900 0.0216 0.0025 72.29322 1 0.0000 1.0218 razd_krm -0.0003 0.0000 55.85077 1 0.0000 0.9997 hise_2000 0.0003 0.0001 31.23404 1 0.0000 1.0003 gozd_jeni(1) 0.6942 0.1758 15.59448 1 0.0001 2.0021 odd_gozd_ven_300 -0.0071 0.0021 10.98816 1 0.0009 0.9929 gozd_zaplata 9.345028 2 0.0093 gozd_zaplata(1) 0.2209 0.1428 2.392736 1 0.1219 1.2473 gozd_zaplata(2) 0.3071 0.1005 9.326788 1 0.0023 1.3594 nasel_2000 0.0002 0.0001 6.999509 1 0.0082 1.0002 ceste_2000 0.0001 0.0001 3.84667 1 0.0498 1.0001 konstanta -2.6605 0.2243 140.7396 1 0.0000 0.0699

94
Slika 58: Habitatni model oz. potencialne gostote medveda v Sloveniji (zgoraj) in aktualne dejanske gostote (spodaj).
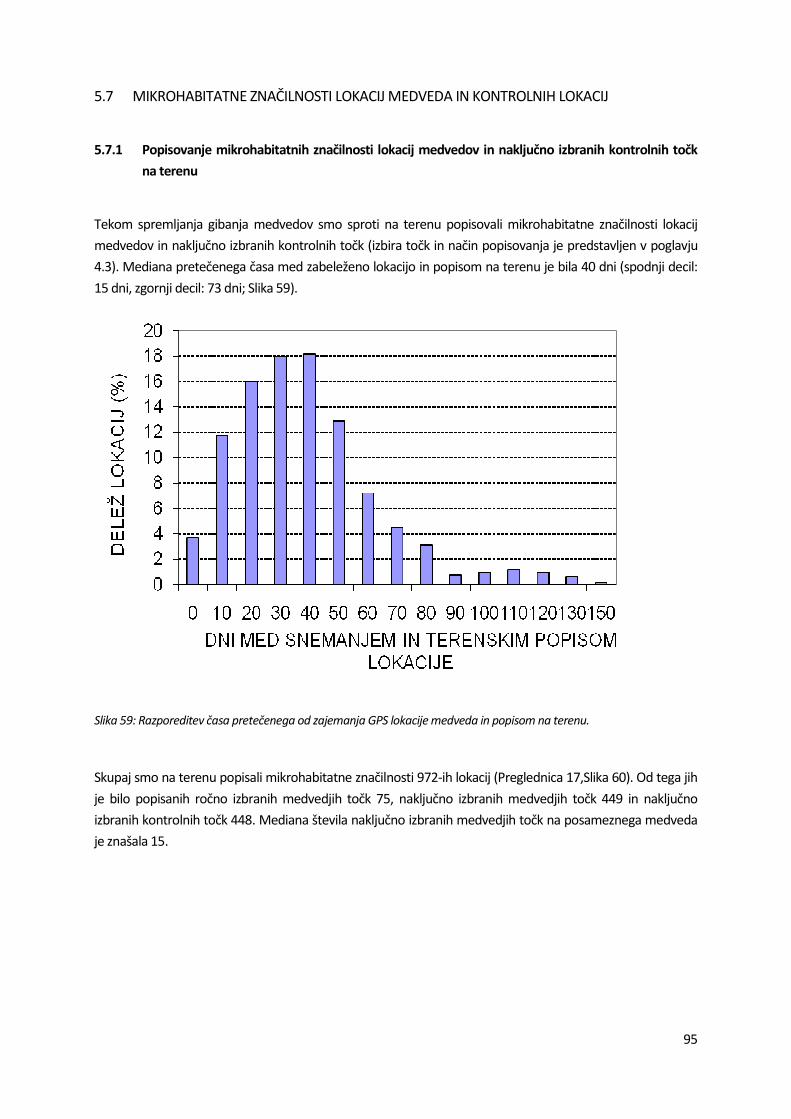
5.7 MIKROHABITATNE ZNAČILNOSTI LOKACIJ MEDVEDA IN KONTROLNIH LOKACIJ
5.7.1 Popisovanje mikrohabitatnih značilnosti lokacij medvedov in naključno izbranih kontrolnih točk na terenu
Tekom spremljanja gibanja medvedov smo sproti na terenu popisovali mikrohabitatne značilnosti lokacij medvedov in naključno izbranih kontrolnih točk (izbira točk in način popisovanja je predstavljen v poglavju 4.3). Mediana pretečenega časa med zabeleženo lokacijo in popisom na terenu je bila 40 dni (spodnji decil:
15 dni, zgornji decil: 73 dni; Slika 59).
S
jij
S
lika 59: Razporeditev časa pretečenega od zajemanja GPS lokacije medveda in popisom na terenu.95
kupaj smo na terenu popisali mikrohabitatne značilnosti 972-ih lokacij (Preglednica 17,Slika 60). Od tega jih
e bilo popisanih ročno izbranih medvedjih točk 75, naključno izbranih medvedjih točk 449 in naključno zbranih kontrolnih točk 448. Mediana števila naključno izbranih medvedjih točk na posameznega medveda e znašala 15.

96
Preglednica 17: Pregled števila točk s popisanimi mikrohabitatnimi značilnostmi za posamezne medvede.
IME MEDVEDA
NAKLJUČNO
IZBRANE
MEDVEDJE
TOČKE
NAKLJUČNO
IZBRANE
KONTROLNE
TOČKE
ROČNO
IZBRANE
MEDVEDJE
TOČKE
ALOJZIJ 12 12 5
ANA 62 62 14
BORA 96 96 3
EVA 20 20 5
FRANCE 1 1 1
GORANKA 15 15 7
HOBI 28 28 7
HRIBAR 1 1 0
IGOR 2 2 0
JANI 8 8 2
KATJA 14 14 1
MIRKA 37 36 6
NEJC 29 29 4
NEŽA 1 1 2
PEČKO 55 55 6
PEPCA 5 5 2
SENOŽEČANKA 40 40 6
ZORA 22 22 4
ŽIVA 1 1 0
SKUPAJ 449 448 75
SKUPAJ VSE 972
V okviru popisovanja mikrohabitatnih značilnosti točk na lokacijah smo zbirali tudi podatke o krmiščih, saj smo predvidevali, da bi lahko le-ta pomembno vplivala na gibanje medvedov. Za osnovo za popisovanje krmišč smo uporabili obstoječe baze krmišč in iz njih izbrali tista krmišča, na katera se polaga krma, ki je
primerna za medvede. Poleg tega smo na terenu obiskali lokacije, za katera smo na podlagi vzorcev gibanja medvedov sklepali, da bi lahko bila krmišča. Enako je veljalo tudi v primeru, da je bilo takšno mesto v bližini evidentiranega krmišča in smo tako potrdili ali ovrgli možnost, da je prišlo do napake pri določanju lokacije
evidentiranega krmišča.

97
.
Slika 60: Ročno izbrane medvedje točke (n = 75), naključno izbrane medvedje točke (n = 449) in naključno izbrane
kontrolne točke (n = 448) s popisanimi mikrohabitatnimi značilnostmi na terenu.
5.7.2 Vplivi antropogenih virov hrane na prisotnost medveda
5.7.2.1 Analiza podatkov
Ugotavljali smo ali prisotnost antropogene hrane pomembno vpliva kot atraktant za medvede. 445 naključno izbranim lokacijam medvedov (»medvedje točke«) smo priredili 445 parnih lokacij (»kontrolne
točke«), kjer ni bilo konkretnih podatkov o prisotnosti medveda. Kontrolne točke smo izbirali znotraj območij aktivnosti posameznega medveda obdanih s pasom, ki je ustrezal polovici razdalje premikov medveda v nočnem času, kar pomeni, da so bile te lokacije medvedu dejansko dosegljive. Pare smo izbirali
znotraj stratumov, ki so bili homogeni v naslednjih dejavnikih: prisotnost gozda, prisotnost ceste, oddaljenost od najbližje hiše. Točki v paru sta bili torej izbrani v območjih primerljive habitatne kakovosti za medveda. Na vsaki od 890 izbranih lokacij smo med drugim zabeležili morebitno prisotnost hrane
antropogenega izvora in vir hrane (krmišče, smetnjak, silažna bala…). Različne vire hrane smo kasneje združili v 3 skupine: »krmišča«, »kmetijstvo«, »odpadki«. Za podrobnosti zbiranja podatkov in izbire lokacije za popis glej poglavje 4.3).
Z McNemarjevim testom za testiranje razlik med dvema odvisnima vzorcema smo ugotavljali ali obstaja
razlika med medvedjimi in kontrolnimi točkami glede na prisotnost antropogene hrane (ne glede na njen vir). S hi-kvadrat testom smo ugotavljali ali obstaja razlika med medvedjimi in kontrolnimi točkami v deležu lokacij s prisotno antropogeno hrano posamezne skupine. Na enak način smo ugotavljali razlike v deležu

98
lokacij s prisotno antropogeno hrano (ne glede na vir. oz. skupino) po posameznih intervalih dejavnikov, ki so vplivali na vzorčenje točk (gozd, ceste, oddaljenost od hiš). Upoštevali smo tudi nekatere preseke intervalov različnih dejavnikov. Vpliv teh dejavnikov ter prisotnosti hrane in interakcij med dejavniki na
prisotnost/odsotnost medvedov smo ugotavljali tudi z logistično regresijo (medvedje vs. kontrolne točke kot odvisna spremenljivka). Z metodo odločitvenih dreves smo ugotavljali kako dejavniki gozd, ceste, oddaljenost od hiš vplivajo na prisotnost/odsotnost hrane na medvedjih točkah (upoštevali smo samo
medvedje točke in prisotnost antropogene hrane kot odvisno spremenljivko).
Preglednica 18: Število medvedjih in kontrolnih točk s prisotno antropogeno hrano glede na vir hrane oz. skupino virov hrane oz. skupino virov.
Vir hrane Medvedje Kontrolne Skupaj
Krmišča 29 2 31
Smetnjak 5 6 11
Smetišče 3 5 8
Smeti 15 9 24
Prostor za piknik 16 0 16
Klavniški odpadki 3 0 3
Kompost 9 5 14
Odpadki skupaj 41 22 63
Silažna bala 12 8 20
Čebelnjak 10 4 14
Domače živali 8 28 36
Pašnik 12 42 54
Hlev / kurnik 0 0 0
Živalska krma 0 3 3
Sadovnjak 79 39 118
Njiva/vrt 21 38 59
Travnik 7 11 18
Ostali viri hrane 12 5 17
Kmetijstvo skupaj 111 107 218
ANTROPOGENA HRANA SKUPAJ 166 118 284

5.7.2.2 Rezultati
Število medvedjih in kontrolnih točk s prisotno antropogeno hrano glede na njen izvor oz. skupino virov prikazujemo v Preglednica 18. Od skupno 890 popisanih točk je bila prisotnost antropogene hrane ugotovljena na 284 točkah (32 %), od tega na 166 medvedjih (37 % vseh medvedjih) in 118 kontrolnih (27 %
vseh kontrolnih). Z McNemarjevim testom smo ugotovili, da vzorca medvedjih in kontrolnih točk pripadata različnim skupinam glede prisotnosti antropogene hrane (α = 0,000), kar pomeni, da se antropogena hrana statistično značilno pogosteje nahaja na medvedjih točkah v primerjavi s kontrolnimi naključnimi točkami.
Antropogena hrana iz skupine »krmišča« je bila prisotna na 31 lokacijah (pozitivne lokacije), od tega na 29
medvedjih in 2 kontrolnih. S hi-kvadrat testom smo ugotovili (α = 0,000), da je porazdelitev pozitivnih lokacij preko obeh skupin (medvedje, kontrolne točke) značilno različna od pričakovane, kar pomeni, da je med medvedjimi lokacijami značilno več takšnih, kjer so prisotna krmišča. Na 218 točkah (111 medvedjih in 107
kontrolnih) smo ugotovili prisotnost hrane iz skupine »kmetijstvo«. Med skupinama ni statistično značilnih razlik v številu pozitivnih lokacij (α = 0,768). Hrano iz skupine »odpadki« smo našli na 63 točkah. Odpadki so bili pogosteje najdeni na medvedjih lokacijah (41 medvedjih, 22 kontrolnih), razlika je statistično značilna (α
= 0,017; Slika 61).
S
lika 61: Število medvedjih in kontrolnih točk, kjer je bila prisotna hrana iz posamezne skupine.99
Porazdelitev medvedjih in kontrolnih točk po intervalih posameznih spremenljivk (oddaljenost od najbližje
hiše, prisotnost gozda, prisotnost ceste) prikazujemo na Slika 62Slika 63Slika 64. V nadaljevanju podajamo rezultate o morebitnih razlikah med prisotnostjo pozitivnih medvedjih in pozitivnih kontrolnih točk v zanimivih intervalih posameznih spremenljivk oz. presekih intervalov različnih spremenljivk.
10
Slika 63: Število medvedjih in kontrolnih točk s prisotno antropogeno hrano glede na prisotnot/odsotnost gozda.
S
lika 62: Število medvedjih in kontrolnih točk s prisotno antropogeno hrano glede na oddaljenost od najbližje hiše.
0

101
Slika 64: Število medvedjih in kontrolnih točk s prisotno antropogeno hrano glede na prisotnot/odsotnost ceste.
V intervalu oddaljenosti od hiš do 50 m je bilo popisanih 182 lokacij, med temi je bila antropogena hrana prisotna na 95 (57 medvedjih in 38 kontrolnih). S hi-kvadrat testom smo ugotovili mejno statistično značilno
razliko (α = 0,051) med številom pozitivnih medvedjih in kontrolnih lokacij. V enakem intervalu oddaljenosti od hiš smo ugotavljali razliko med medvedjimi in kontrolnimi točkami glede na prisotnost gozda. 57 lokacij je bilo posnetih zunaj gozda, od tega 31 medvedjih in 26 kontrolnih. Razlika ni statistično značilna (α = 0,508).
38 lokacij je bilo v gozdu, od tega 26 medvedjih in 12 kontrolnih. V tem primeru je razlika statistično značilna (α = 0,023; Slika 65).
Slika 65: Število medvedjih in kontrolnih točk s prisotno antropogeno hrano v oddaljenosti od najbližje hiše do 50 m.
Na oddaljenosti od hiš nad 50 m je bilo v gozdu posnetih 396 lokacij. Če ne upoštevamo krmišč, je bila med temi na 33 ugotovljena prisotnost antropogene hrane, od tega na 17 medvedjih in 16 kontrolnih. Razlika ni
statistično značilna (α = 0,862). (Krmišča smo izključili, saj je samoumevno, da jih je več na medvedjih lokacijah kot na kontrolnih; v splošnem je 7 % vseh medvedjih lokacij na krmiščih, medtem ko je verjetnost,

102
da kontrolna točka pade na krmišče proporcionalna površini, ki jo znotraj posameznega stratuma pokrivajo krmišča, kar je zagotovo manj kot 7 %.)
Med vsemi posnetimi lokacijami jih je 378 zunaj gozda. Med temi je bila antropogena hrana prisotna na 199, od tega na 111 medvedjih in 88 kontrolnih. S hi-kvadrat testom nismo ugotovili statistično značilne razlike
med skupinama (α = 0,103). Med 512 lokacijami v gozdu je bila antropogena hrana prisotna na 85, od tega na 55 medvedjih in 30 kontrolnih. Razlika je statistično značilna (α = 0,007). Če iz 85 pozitivnih lokacij v gozdu izločimo krmišča, ostane 71 lokacij s prisotno antropogeno hrano, od tega 43 medvedjih in 28 kontrolnih. V
tem primeru je razlika blizu statistične značilnosti (α = 0,075).
V okolici cest je bilo od 102 popisanih točk antropogena hrana prisotna na 28, od tega na 17 medvedjih in 11 kontrolnih. Razlika ugotovljena s hi-kvadrat testom ni statistično značilna (α = 0,257), kar pa je lahko tudi posledica majhnosti vzorca. Od 788 lokacij posnetih vstran od cest je bila antropogena hrana prisotna na
256, od tega na 149 medvedjih in 107 kontrolnih. Razlika je statistično značilna (α = 0,009).
Z logistično regresijo smo preverjali ali interakcije prisotnosti antropogene hrane s prisotnostjo gozda, prisotnostjo cest oz. oddaljenostjo od hiš vplivajo na prisotnost medvedov. Zaradi zahtev metode smo v model ponudili tudi čiste učinke omenjenih okoljskih dejavnikov, vendar zaradi načina vzorčenja nismo
pričakovali značilnih čistih učinkov dejavnikov, temveč zgolj morebitne značilne interakcije z antropogeno hrano.
Model napoveduje, da je prisotnost medvedov pogojena z interakcijo med prisotnostjo antropogene hrane in oddaljenostjo od najbližje hiše. Prisotnost medvedov je večja, kjer je prisotna antropogena hrana in v
splošnem (neznačilno) večja, kjer je oddaljenost od hiš nad 50 m (kljub temu, da je spremenljivka pričakovano (zaradi načina vzorčenja) neznačilna, smo jo vsilili v model zaradi zahtev metode – kadar je značilna interakcija spremenljivk, mora model vsebovati tudi osnovne spremenljivke). Vendar pa je ob
prisotnosti hrane prisotnost medvedov značilno večja v bližini hiš kot če hrana ni prisotna. To pomeni, da se medvedi v splošnem izogibajo hišam, razen kadar je v bližini hiš prisotna hrana. Model pravilno razvrsti 56,3 % vseh točk (67,6 % kontrolnih in 44,9 % medvedjih).
Metoda odločitvenih dreves napoveduje, da je odd. od najbližje hiše najpomembnejši dejavnik, ki vpliva na
prisotnost/odsotnost hrane na medvedjih lokacijah. Če je pomembnost tega dejavnika 100, potem je pomembnost prisotnosti gozda 60 in pomembnost prisotnosti cest 2 (nepomemben dejavnik).
Slika 66 prikazuje kako se razvrščajo dejavniki in njihovi intervali, ki pomembno vplivajo na prisotnost antropogene hrane na medvedjih lokacijah. Odločitveno drevo napoveduje, da je v gozdu hrana v splošnem
prisotna v oddaljenosti od 0 do 50 m od hiš, medtem ko je na večji oddaljenosti v splošnem odsotna (leva veja). Zunaj gozda je hrana prav tako najbolj prisotna na oddaljenosti od hiš do 50 m, medtem ko na večji oddaljenosti (nad 50 m) prevladujejo lokacije z manj hrane. Oddaljenost nad 50 m od hiš se dalje deli na
lokacije nad 500 m, kjer prevladujejo lokacije brez hrane in lokacije od 50 do 500 m, kjer prevladujejo lokacije s hrano (desna veja).

Slika 67 doSlika 73 prikazujejo nekaj primerov antropogenih virov hrane, ki smo jih našli med popisi lokacij medvedov v bližini naselij in ki so očitno privabljali medvede v bližino človeka.
S
S
d
103
lika 67: Ilegalno odlagališče smeti s klavniškimi odpadki (ostanki mrtvega konja in divjega prašiča) v bližini Šmihela, ki je več
ni privabljalo medvedko Goranko v bližino vasi. (foto: Miha Krofel)
lika 66: Odločitveno drevo – razvrščanje dejavnikov, ki vplivajo na prisotnost antropogene hrane na medvedjih lokacijah.

104
Slika 69: Klavniški odpadki pri Oneku na Kočevskem. (foto: Miha Krofel)
Slika 68: Ilegalna odlagališče smeti z organskimi odpadki pri Ribnici (levo) in v Loški dolini (desno). (foto: Miha Krofel)

105
Slika 71: Obcestni smetnjaki pri Loški dolini (levo) in Blokah (desno). (foto: Miha Krofel)
Slika 70: Kompost s kuhinjskimi odpadki na Babnem polju. (foto: Miha Krofel)

106
Slika 73: Prostor za piknik z ostanki hrane pri Babnem polju. (foto: Miha Krofel)
Slika 72 Odvržene tropine pri Strugah. (foto: Miha Krofel)

107
Slika 74: Smetnjaka na Blokah, kjer redno prihaja do konfliktov z medvedi. (foto: Miha Krofel)

108

109
6 DISKUSIJA IN ZAKLJUČKI RAZISKAVE
Velika količina podatkov, ki smo jih pridobili s telemetrijskim spremljanjem medvedov in terenskimi snemanji njihovih lokacij omogoča podrobno analizo številnih vidikov biologije in ekologije rjavega medveda
v Sloveniji, kot so aktivnost in gibanje v različnih časovnih okvirih, dnevno-nočna, sezonska in celoletna raba prostora, raba antropogenih in naravnih virov hrane, razlike v teh parametrih med starostnimi kategorijami, spoloma in reprodukcijskim statusom skupin osebkov itd. Ti podatki so pomembna osnova za razumevanje
pojavljanja konfliktov med medvedom in človekom in s tem za pripravo smernic za učinkovito ohranitveno upravljanje vrste. V nadaljevanju glavne rezultate raziskave obravnavamo ločeno po vsebinskih sklopih pri čemer so izpostavljene predvsem navezave na razumevanje vzrokov nastanka in reševanje konfliktov med
medvedom in človekom in nekatera aktualna upravljavska vprašanja. Spoznanja raziskave, smo skupaj s tujimi izkušnjami o učinkovitosti različnih ukrepov za zmanjševanje konfliktov (pregled je podan v naslednjem poglavju) uporabili za pripravo upravljavskih priporočil za zmanjševanje konfliktov v poglavju 9.
6.1 VELIKOSTI OBMOČIJ AKTIVNOSTI MEDVEDOV
Povprečno območje aktivnosti rjavega medveda v Sloveniji znaša 350 km2 (ocena po metodi 100% MCP), kar je primerljivo z velikostjo pri drugih populacijah rjavega medveda po svetu, ki živijo v podobnih razmerah (za
pregled glej npr. Swenson et al. 2000 in Dahle & Swenson 2003). Glede na to, da medved uporablja tako velik prostor so tudi največji gozdni kompleksi z medvedje perspektive pri nas dokaj majhni. V Sloveniji je le nekaj neposeljenih območij pokritih s sklenjenim gozdom, ki imajo površino večjo od povprečnega območja
aktivnosti medveda. Zato ni presenetljivo, da so območja aktivnosti skoraj vseh spremljanih medvedov vključevali tudi negozdne površine in naselja (glej PRILOGA 8). To spoznanje predstavlja eno izmed ključnih osnov za upravljanje z medvedom, saj opozarja, da bo medved v Sloveniji preživel samo, če bo človek
pripravljen z njim sobivati. Za razliko od držav kot je npr. Kanada, Slovenija nima dovolj velikih neposeljenih površin, da bi lahko medvedi živeli ločeno od človeka.
Posledica velikih območij aktivnosti je tudi, da mnogi medvedi redno prehajajo državno mejo. To kaže na velik pomen čezmejnega sodelovanja pri upravljanju z vrsto v Sloveniji, še posebej na sodelovanje s Hrvaško.
Od medvedov, ki smo jih spremljali v raziskavi in so bili odlovljeni v Sloveniji, jih je skoraj polovica vsaj enkrat šla tudi na Hrvaško. Ker si dejasnko delimo velik del istih živali, je potrebno način upravljanja koordinirati s sosednimi državi in pri načrtovanju upravljanja upoštevati, kaj se s populacijo dogaja tudi na drugi strani
meje, saj lahko v nasprotnem primeru zgrešimo zastavljene upravljavske cilje.
Velika območja aktivnosti in velike razdalje, ki jih medved prepotuje v relativno kratkem času narekujejo previdnost pri odrejanju izrednega odstrela medvedov, ko se le-ta ne izvaja na samih mestih konflikta, ampak v širši okolici (npr. na krmiščih), kar je pogosta praksa. Medved se namreč lahko v kratkem času
premakne na zelo velike razdalje (glej poglavje 5.3), kar vodi do dveh problemov: 1.) Isti medved lahko povzroča konflikte na dokaj oddaljenih lokacijah, kar vodi do napačne ocene, da gre za več problematičnih medvedov. Dober tovrsten primer je bil samec Rožnik, ki je v roku enega meseca in pol povzročal konflikte
od Kočevske, do Blok, Rakitne, okolice Grosuplja, Zasavja, Vranskega in Koroške. Če ta medved ne bi bil opremljen s telemetrično ovratnico, bi verjetno ocenili, da v tistem času te konflikte povzroča večje število medvedov, za kar bi se izdalo več odločb za izredni odstrel. Poleg tega taki dogodki utrjujejo prepričanje o

110
velikem deležu problematičnih živali. Poleg tega bi to lahko vodilo so odstrela več medvedov, med katerimi večina v resnici ni problematičnih. 2.) Medvedi niso teritorialni, zato pogosto na istem območju živi večje število medvedov. Obenem medvedi v posameznem mesecu uporabljajo območja velika več tisoč hektarjev.
Zaradi tega je vprašljivo, ali se pri izvajanju izrednega odstrela, ko se le-ta ne izvaja na mestih konfliktov, res odstreli pravega medveda.
Domače raziskave kažejo, da imajo medvedi v alpskem delu še posebej velika območja aktivnosti (tudi preko 1000 km2), tako da se njihov življenjski prostor poleg alpskega območja v Sloveniji in sosednjih državah
(Avstrija, Italija) pokriva tudi območja znotraj ali v bližini osrednjega območja rjavega medveda v Sloveniji. Glede na trenutno majhno število medvedov v alpskem prostoru in dolgoročne ideje o spodbujanju širjenja vrste v Alpe, se je zato potrebno zavedati, da lahko tudi poseganje v populacijo izven vendar v bližini robnega
območja (npr. na Nanosu, Trnovskem gozdu, Hrušici) neposredno pomembno vpliva na dinamiko populacije v Alpah in uspeh morebitne rekolonizacije alpskega prostora v sosednjih državah.
Spremljanje medvedov, ki so bili preseljeni v okviru dela intervencijske skupine, je ponovno pokazalo, da ta ukrep na relativno majhnem območju, kot je Slovenija, verjetno ni učinkovit. Dva od treh preseljenih
medvedov sta se vrnila na mesto odlova in oba bila kasneje tudi odstreljena (Neža zakonito, Rožnik nezakonito). Za več informacij o učinkovitosti tega ukrepa glej poglavje 7.2.
Velikost območja aktivnosti je v splošnem obratnosorazzmerno odvisna od razpoložljivosti hrane in populacijske gostote, zato imajo medvedi v prehransko revnejših območjih na severu Evrope v povprečju
večja območja aktisnosti (tudi do 8 000 km2), kot to velja za območja zmernega pasu, kamor sodi Slovenija. Tudi znotraj istega klimata se velikosti območij aktivnosti precej razlikujejo med posameznimi medvedi in populacijami. V našem vzorcu medvedov, ki smo jih spremljali vsaj 4 mesece, se tako območja aktivnosti
gibljejo med 19 in 1 223 km2. Še večje pa je bilo območje, ki ga je v zgolj mesecu in pol prepotoval medved Rožnik, in je znašalo kar 3 144 km2. V splošnem smo opazili zelo velike razlike med obnašanjem različnih medvedov (poleg gibanja tudi pri aktivnosti, obiskovanju krmišč, približevanju naseljem itd.), kar je ponovno
potrdilo veliko individualno variabilnost pri vedenju te vrste.
Velikost območja aktivnosti je odvisna od več dejavnikov. V povprečju imajo samci za več kot 4-krat večje območja od samic. Pomembna je tudi starost medveda in reprodukcijska kategorija medvedk. Največja OA imajo navadno mladi samci med disperzijo, tudi do 12 000 km2 (Swenson et al. 2000). Takšen primer je bil v
naši raziskave mladi samec Rožnik. Veliko OA je bilo pri njem lahko tudi deloma posledica preselitve. Opazili smo tudi, da so vsaj pri samcih OA večja na robu populacije kot v osrednjem območju. To je najverjetneje posledica razlik v populacijskih gostotah medvedov, ki so na robu populacije bistvene manjše. Med ostalimi
dejavniki na velikost OA vpliva tudi: 1) fragmentacija prostora: npr. na območju Menišije, ki jo s treh strani omejujejo človeška naselja in prometnice, imajo medvedi manjša OA, 2) razpoložljivost hrane: večja območja aktivnosti medvedov v robnih delih populacije so lahko tudi posledica manjše prehranske nosilne
zmogljivosti prostora, po drugi strani pa prihaja do večjih premikov, ko se na določenih območjih pojavijo sezonsko omejeni bogati viri hrane (npr. žir v čistih bukovih sestojih, koruza v dolinah), 3) letni čas: npr. v času parjenja se povečajo OA pri odraslih samcih in zmanjšajo pri samicah z mladiči, pri nekaterih samicah pa
smo opazili tudi večje premike ob začetku in koncu zimskega mirovanja, ko so se odpravile v oddaljene brloge in 4) gostota krmišč: večja OA so imeli medvedi, ki so živeli na območjih z manj krmišči, kar pa ni nujno v neposredni povezavi s krmljenjem, ampak učinki krmljenja na populacijske gostote in specifične prostorske
razporeditve krmišč v Sloveniji.

111
Dobro razumevanje dejavnikov, ki vplivajo na velikost OA medvedov in njihove sezonske premike, je ključno za pravilno interpretacijo podatkov monitoringa medvedov. Še posebej je to pomembno, če se monitoring izvaja na majhnem območju in se nato želi na podlagi lokalnih podatkov ocenjevati populacijsko dinamika na
nivoju države ali ekstrapolirati lokalne gostote na širše območje. V Sloveniji, za katero velja velika pestrost v habitatih in lokalni razpoložljivosti hrane, se lahko lokalne gostote medvedov hitro spreminjajo, tako v geografskem kot časovnem merilu. Lokalno in trenutno razpoložljivi bogati viri hrane lahko na primer v
določenem obdobju privabijo medvede iz širšega območja na manjše območje, zaradi česar se začasno močno povečajo gostote medvedov na tem območju. Če ta lokalna opažanja nato posplošimo, lahko pridemo do napačnih zaključkov, da se je število medvedov v Sloveniji naenkrat močno povečalo. Do
podobnih sezonskih sprememb v razporeditvi medvedov in posledično lokalnih gostotah lahko pride tudi v času parjenja, še posebej na robu populacije, kjer je število samic omejeno in se lahko večje število samcev skoncentrira na območju z eno ali več medvedkami. Pazljivost je potrebna celo pri podatkih monitoringa, ki
se izvaja na celotnem območju Slovenije, saj si velik del populacije delimo s Hrvaško, pri čemer lahko prihaja do verletno velikih sezonsko pogojenih razlik v razporeditvi medvedov med obema državama.
6.2 AKTIVNOST IN GIBANJE MEDVEDOV
Dnevno-nočna aktivnost živali je v splošnem genetsko pogojena, pri čemer navadno svetloba nudi referenčno točko (Nielsen 1983). Kljub temu pa se lahko aktivnost spremeni zaradi različnih dejavnikov, kot so na primer dostopnost raznih dobrin, znotraj- in medvrstna kompeticija, plenjenje in različne motnje
(Kaczensky et al. 2006). V splošnem so medvedi bolj dnevno aktivni na območjih, kjer je manj motenj s strani človeka in so jih v preteklosti manj preganjali. Medvedi prilagajajo svojo aktivnost človeku na podoben način, kot se plen izogiba plenilcu (Ordiz et al. 2011).
Tekom stoletji sobivanja s človekom so se medvedi naučili ljudem izogibati, sicer jih je človek ubil. To se
posredno kaže tudi v podatkih zbranih v okviru tega projekta. Večina medvedov je bila aktivnih ponoči in v mraku, torej v času, ko je srečanje s človekom najmanj verjetno. Da gre pri opaženem vzorcu aktivnost najverjetneje res za izogibanje ljudem, kažejo podatki od medvedov, ki so izgubili strah pred človekom (npr.
Rožnik), saj so bili ti medvedi precej bolj aktivni prek dneva v primerjavi z ostalimi medvedi. Pri konfliktnih medvedih smo tudi opazili, da so se gibali več od ostalih.
Večjih razlik med samci in samicami oziroma med različnimi reprodukcijskimi kategorijami medvedk (z ali brez mladičev) v aktivnosti nismo opazili, so se pa samci nekoliko več premikali. V času parjenja smo opazili
največje premike pri samcih in najmanjše pri samicah z najmlajšimi mladiči. To je najverjetneje posledica aktivnega iskanja za parjenje godnih samic pri samcih in izogibanje potencialno infanticidnih samcev pri samicah z mladiči. Po pričakovanjih sta se aktivnost in gibanje medvedov zmanjšali v zimskem času, ki ga
mnogi medvedi preživijo v brlogih. Najdlje so v brlogih ostajale samice, ki so čez zimo povrgle mladiče, saj so ti ob rojstvu slabo razviti in potrebujejo več mesecev, preden lahko sledijo materam. Sezonski viški aktivnosti ustrezajo obdobju hiperfagije, t.j. čas intenzivnega prehranjevanja pred zimskim mirovanjem (jesen), in
obdobju parjenja (pozna pomlad). Največje razlike v aktivnosti in gibanju tekom dneva z najbolj izraženimi viški velike aktivnosti smo opazili poleti. Razlog bi lahko bil v dolžini dneva, zaradi česar morajo biti medvedi bolj aktivni, da v kratki noči najdejo dovolj hrane.

112
6.3 PRIBLIŽEVANJE NASELJEM
Podatki iz gibanja medvedov so pokazali, da se večina medvedov aktivno izogiba naseljem. Tudi kadar so prišli v bližino naselij, se je to največkrat dogajalo sredi noči. Na ta način so medvedi zmanjšali verjetnost
srečanja s človekom.
Pri približevanju medvedov človeškim bivališčem smo opazili velike individualne razlike med medvedi. Večina medvedov se je naseljem približevala zelo redko ali sploh nikoli, medtem ko so nekateri medvedi redno prihajali k vasem. Medvedi, za katere smo pri delu na terenu opazili, da so bili vsaj delno habituirani na
prisotnost človeka (npr. Rožnik in Bora), so tudi pogosteje zahajali v bližino naselij. Pri medvedu Rožniku smo npr. opazili, da se je naseljem približeval dvanajstkrat pogosteje, kot ostali medvedi. Ko smo primerjali podatke o zabeleženih konfliktih med medvedi in ljudmi ter gibanjem medveda Rožnika smo ugotovili, da je
bil v času spremljanja ta medved »kriv« za 40 % vseh prijavljenih konfliktov med medvedi in ljudmi v celotni Sloveniji (v aprilu 2009 je ta delež znašal celo 71 %). Ti podatki kažejo, da v populaciji obstajajo t.i. problematični medvedi in da majhno število takšnih medvedov povzroči velik del vseh konfliktov med
medvedi in ljudmi, medtem ko ostali medvedi v konflikte prihajajo zelo redko ali sploh nikoli. Ta ugotovitev razloži tudi opažanja, da razlike v gostotah medvedov ne vplivajo na pojavljanje konfliktov (Marenče et al. 2007; Kragelj et al., neobjavljeno). Precej bolj od števila medvedov na nekem območju je namreč
pomembno, ali so na območju prisotni problematični medvedi ali ne. Zaradi tega imajo tudi ukrepi, ki se nanašajo na splošno zmanjševanje števila medvedov precej omejen učinek na preprečevanje konfliktov, medtem ko je odstranitev problematičnih medvedov lahko vsaj kratkoročno zelo učinkovito.
Da bi ugotovili, kaj so razlogi za primere, ko se medvedi približajo naseljem, smo na terenu opravili popis
medvedjih lokacij in jih primerjali z naključno izbranimi kontrolnimi točkami. Podatki so pokazali, da je bila na lokacijah medvedov v bližini naselij pogosteje prisotna hrana antropogenega izvora (klavniški odpadki, smeti, ostanki hrane na prostorih za piknik itd.). To kaže, da so razpoložljivi antropogeni viri hrane v okolici naselij
pomemben dejavnik, ki vplivajo na zahajanje medvedov v bližino človeka, kar se sklada z izsledki raziskav iz tujine (za pregled glej poglavje 2.2.2). Medvedi se naseljem približujejo predvsem ponoči in se na ta način izogibajo ljudem. Vendar, če medved pogosto dobi hrano bližini naselij, verjetno postopoma začne
povezovati prisotnost človeka s hrano. Redne »nagrade« v obliki hrane, spodbujajo zahajanja v vasi, še posebej tam, kjer je mogoče redno najti hrano, obenem pa zaradi pomanjkanja negativnih izkušenj medved začne zgubljati strah pred človekom in se ga zato tudi vedno manj izogiba (postane habituiran na prisotnost
človeka). V končni fazi medved postane problematičen, saj se ljudi ne izogiba več in ob iskanju hrane prihaja v naselja tudi podnevi, kar razumljivo vzbuja strah med lokalnimi prebivalci in vodi v številne konflikte. V takšnem primeru je potrebno medveda odstraniti, kar pa dolgoročno ne reši težav, saj bo obilica lahko
dostopne hrane kmalu na isto lokacijo privabila novega medveda, ki bo postopoma prav tako postal problematičen. To kažejo tudi nekateri primeri iz Slovenije, kjer se na istih lokacijah vedno znova pojavljajo problematični medvedi in veliko število konfliktov z medvedi (npr. Bloke, Rakitna, Kurešček). Zato je
omejevanje dostopa do antropogene hrane v bližini ljudi na teh ključnih lokacijah verjetno eden izmed najpomembnejših ukrepov za preprečevanje habituacije medvedov na prisotnost človeka in posledično pojavljanja konfliktov.

113
6.4 RABA KRMIŠČ
Umetno krmljenje medvedov je v svetu zelo kontroverzen ukrep in marsikje prepovedan, ponekod pa se intenzivno uporablja za namene lova, ekoturizma, monitoringa populacije in/ali preprečevanja konfliktov
(Gray 2004, Dečak et al. 2005, Priestly 2009, Kojola et al. 2010, Jerina et al. 2011). Kljub dolgi tradiciji krmljenja medvedov v nekaterih državah pa so bili pozitivni in negativni učinki krmljenja do sedaj zelo slabo raziskani. Posledično so tudi mnenja med strokovnjaki o upravičenosti krmljenja medvedov zelo deljena
(Hererro 1985, Fersterer et al. 2001, Partridge et al. 2001, Gray et al. 2004, Rogers 2009).
Podatki so pokazali, da medvedi krmišča najpogosteje uporabljajo zvečer, preko noči se obiskovanje postopoma zmanjšuje, podnevi pa na krmišča zahajajo le izjemoma. Za razliko od aktivnosti pri uporabi krmišč ni opaženega viška ob zori. Te podatke lahko po eni strani razložimo z izogibanjem srečanj s
človekom, ki so manj verjetna ponoči, ko medvedi prihajajo na krmišča. Poleg tega preko noči medvedi in druge živali postopoma pojedo nastavljeno hrano, ki jo je proti jutru vse manj, zato je verjetno obiskovanje krmišč bolj intenzivno zvečer in v prvi polovici noči, medtem ko ob drugem višku aktivnosti ob zori medvedi
na krmišča zahajajo le redko. Razlike v obiskovanju krmišč med različno starimi samci ter med samicami z oziroma brez mladičem so najverjetneje posledica izogibanja človeka in drugih medvedov. Odrasli samci so dominantni osebki v populaciji medvedov. Ti se bojijo predvsem človeka, zato krmišča uporabljajo kasneje v
noči, ko so srečanja s človekom manjša. Tudi mladi samci in samice se izogibajo človeka, vendar pa se morajo obenem izogibati tudi velikim samcem, ki bi lahko bili nevarni za njih oziroma za njihove mladiče. Zato ti medvedi na krmišča prihajajo nekoliko bolj zgodaj, ko je sicer verjetnost srečanja s človekom nekoliko večja,
vendar pa na ta način zmanjšajo verjetnost srečanja z velikimi samci. Medvedji samci so še posebej nevarni za mladiče, kar bi lahko bil razlog, da na krmišča najbolj zgodaj prihajajo samice z mladiči. Samice z mladiči so tudi najredkeje obiskovale krmišča, kar je ponovno povezano z izogibanjem odraslim samcem. Najpogosteje
so krmišča uporabljali mlajši samci, kar bi lahko povezovali z njihovo neizkušenostjo pri iskanju hrane, zaradi česar so pogosteje uporabljali predvidljivo in lahko dostopno visokokalorično hrano na krmiščih. Medvedi so krmišča najpogosteje uporabljali zgodaj spomladi in pozno jeseni, kar ustreza času, ko je v naravi na voljo
najmanj naravne hrane. Anekdotična opažanja podobno kažejo, da je obiskovanje krmišč bolj intenzivno v letih z manjšo dostopnostjo naravne hrane.
Glede na podatke o številu obiskovanih krmišč na noč, posamezen medved pride na krmišče v povprečju približno vsako drugo do tretjo noč. Pri tem pa smo med različnimi medvedi opazili precejšnje razlike –
nekateri medvedi so prihajali na krmišča večino noči, nekateri pa so se hranili na krmiščih le sporadično. Medvedi, ki so pogosteje obiskovali krmišča, so v splošnem uporabljali tudi več različnih krmišč tekom leta. Navadno medvedi niso obiskali več kot eno krmišče na noč, pogosto pa so imeli nekatera priljubljena
krmišča, ki so jih redno obiskovali, del krmišč znotraj svojega območja aktivnosti pa niso nikoli obiskali. Dejavnikov, ki vplivajo na to, ali bo medved določeno krmišče obiskoval ali ne, v naši raziskavi nismo analizirali. Analiza podatkov iz obdobja, ko je še potekalo intenzivno krmljenje z mrhovino in po ukinitvi
mrhovišč, ni pokazala, da bi medvedi raje obiskovali krmišča, ko so založena z mrhovino v primerjavi s tistimi, kjer se je krmilo samo z rastlinsko krmo (Kavčič et al. 2011), akr se pogosto špekulira v praksi.
Naša analiza vplivov krmljenja medvedov v Sloveniji je pokazala, da imajo krmišča močan vpliv na rabo prostora pri medvedu. To se najbolj kaže v povečani rabi prostora v okolici krmišč, opazili pa smo tudi vpliv
na velikost območija aktivnosti. Prejšnje raziskave prehrane so pokazale tudi velik vpliv krmljenja na prehrano medvedov, saj po oceni krma predstavlja kar tretjino vse zaužite energije pri medvedih v Sloveniji

114
(Grosse et al. 2003, Kavčič et al. 2011). Po drugi strani pa telemetrični podatki niso pokazali, da bi krmljenje medvedov kakorkoli zmanjševalo zahajanje medvedov v naselja. Tudi število konfliktov je v Sloveniji kljub zelo intenzivnem krmljenju medvedov še vedno relativno visoko. Analiza podatkov iz obdobja, ko je še
potekalo intenzivno krmljenje z mrhovino, tudi ni pokazalo vpliva krmljenja z mrhovino na število napadov na domače živali (Kavčič et al. 2011). To kaže, da krmljenje medvedov verjetno ne daje želene učinkovitosti za preprečevanje konfliktov, obenem pa je povezano z dokaj močnimi stranskimi učinki. Zaradi tega bo v
prihodnje potrebno več pozornosti namenjati drugim ukrepom, ki bi lahko bili bolj učinkoviti za preprečevanje konfliktov (glej poglavje 7). Potrebne so tudi podrobnejše tarčne raziskave osredotočene na različne posledice krmljenja.
V Slovenijo pomemben del monitoringa medvedov predstavlja štetje na stalnih števnih mestih, ki se
nahajajo na krmiščih. Telemetrični podatki so pokazali, da v posamezni noči, velik del medvedov ne obišče števnih krmišč. Opazili smo tudi dokaj visoke razlike v obiskovanju krmišč med različnimi medvedi in različnimi kategorijami medvedov (npr. samci / samice ali samice z / brez mladičev). Ti rezultati kažejo na
previdnost, ki je potrebna pri interpretaciji podatkov pridobljenih iz monitoringa medvedov z opazovanji na krmiščih, še posebej če se želi sklepati na absolutno število medvedov v Sloveniji oziroma absolutne deleže posameznih kategorij medvedov v populaciji. Zato priporočamo, da se naj ti podatki še naprej, kot je bila v
zadnjih letih praksa, uporabljajo predvsem za ugotavljanje relativnih trendov v številu oziroma strukturi populacije medvedov, nikakor pa ne za ugotavljanje absolutne številčnosti.

115

116
7 PREGLED MOŽNIH UKREPOV ZA PREPREČEVANJE KONFLIKTOV IN NJIHOVA UČINKOVITOST
Tekom tisočletij sobivanja z medvedom je človek razvil vrsto ukrepov, s katerimi je bolj ali manj uspešno preprečeval konflikte. V nadaljevanju predstavljamo pregled teh ukrepov in izkušnje glede njihove učinkovitosti, tako iz tujine kot Slovenije. Za učinkovito zmanjševanje konfliktov je predvsem pomembno, da
se z ukrepi konflikte prepreči še preden do njih pride (Gunther et al. 2004, WSPA 2009). V splošnem velja, da je upravljanje z medvedom precej manj težavno in vanj je potrebno vlagati precej manj napora, če je prisotno dobro upravljanje s prostorom in ljudmi (Herrero et al. 2005). Kadar je mogoče, je zaželeno, da se v
preprečevanje konfliktov v čim večji meri vključuje tudi lokalno prebivalstvo in da se vpeljevanje novih ukrepov poveže z že obstoječimi (Primm & Wilson 2004). Nekateri avtorji so tudi predlagali, da se ukrepi spreminjajo glede na obrod za medvede pomembnih plodov v naravi (Mattson et al. 1992). Tako je na
primer priporočljivo z odstrelom konfliktnih osebkov, ki zahajajo v naselja, hitreje reagirati v obdobjih, ko je v naravi dosti druge hrane (Simonič 1994), preventivni ukrepi preprečevanja dostopa do antropogene hrane pa morajo biti intenzivnejši v času pomanjkanja hrane v naravi. V splošnem mednarodna ekspertna skupina,
ki se ukvarja s preučevanjem preprečevanjem konfliktov med medvedi in ljudmi (v okviru IBA) priporoča, da se pred odstrelom poskusi z vzpodbujanjem sprememb pri ravnanju ljudi (npr. kar se tiče odlaganja odpadkov v okolju) in uporabo drugih ukrepov, ki ne vsebujejo usmrtitve živali (WSPA 2009). Seveda je treba
pri sprejemanju posameznih ukrepov upoštevati predvsem njihovo učinkovitost.
7.1 ODSTREL
V preteklosti je upravljanje z medvedi in reševanje konfliktov temeljilo predvsem na odstrelu in drugih
načinih ubijanja medvedov (Witmer & Whittaker 2001). Zaradi nizkih populacijskih gostot in relativno počasnega razmnoževanja v primerjavi z drugimi vrstami je rjavi medved občutljiv na preveliko poseganje v populacije (Swenson et al. 2000, Bischof et al. 2009). Zato je na mnogih območjih prišlo do drastičnega
upada populacij medvedov in ponekod tudi do popolnega iztrebljenja. S časom je prišlo do sprememb v odnosu ljudi do narave ter v zavedanju pomena medvedov v naravnih ekosistemih in ljudski kulturi. Posledično je prišlo do pozivov za ohranitev medvedov in danes je eden izmed glavnih ciljev upravljanja z
medvedi tudi ta, da se zagotovi dolgoročno preživetje vrste. Obenem raziskave po vsem svetu kažejo, da postaja ubijanje karizmatičnih živali kot so velike zveri za javnost vedno manj sprejemljivo (Treves & Karanth 2003) in že v več primerih, so morali upravljavci zaradi močnih pritiskov javnosti spremeniti obseg in način
poseganja v populacije (Schwartz et al. 2005). Kljub temu pa odvzem medvedov iz narave še vedno predstavlja enega pomembnejših upravljavskih ukrepom in nujno sestavino dolgoročnega ohranitvenega ravnanja s temi vrstami na območju večje poseljenosti z ljudmi (Adamič 1997). Vendar pa je potrebno vsak
poseg v populacijo dobro načrtovati in odstrel čim bolj optimizirati, tako da se poveča učinek na zmanjšanje konfliktov. Odstrel je dosti bolj sprejemljiv ukrep v populacijah, ki so v ugodnem ohranitvenem stanju, kot pa v manjših, izoliranih populacijah, katerih številčnost stagnira ali celo upada. Zato je ključen del takšnega
ukrepa tudi sistematično spremljanje populacij in odvzema osebkov iz narave (McLellan et al. 1999, Swenson et al. 2000, Adamič & Jerina 2006).

117
Odstrel lahko v grobem razdelimo na dva dela: splošen odstrel z namenom zmanjšanja številčnosti celotne populacije in tarčni odstrel konfliktnih osebkov.
7.1.1 Zmanjševanje številčnosti populacije
Eno ključnih izhodišč dolgoročne ohranitve populacij medvedov, ki sobivajo z ljudmi, je, da se številčnosti medvedov vzdržuje na ravni, ki sprejemljiva za lokalno prebivalstvo (Huber 2010). V Sloveniji je trenutna velikost populacije 400-500 medvedov (Skrbinšek et al. 2008). Raziskave so pokazale, da ta številčnost še ni
blizu nosilne kapacitete okolja in da bi ob trenutnih razmerah v okolju v Sloveniji lahko živelo še več medvedov (Krofel et al., v recenziji). Če privzamemo, da je 400-500 medvedov tista številčnost, s katero so ljudje še pripravljeni sobivati, povečanje pa bi pripeljalo do občutnega upada te tolerance, je odstrel edini
ukrep, s katerim bomo populacijo lahko umetno zadržali na tej ravni in s tem posledično zagotovili obstoj vrste, ki je v prvi vrsti odvisen od tolerance ljudi.
Kljub vsemu pa se je potrebno zavedati omejitev takšnega splošnega odstrela za učinkovito preprečevanje konfliktov, saj je število konfliktov le v manjši meri odvisno od številčnosti medvedov in obsega odstrela (glej
poglavje 3.1). Dober primer zato je ravno Sloveniji, kjer imamo trenutno najmočnejše poseganje v populacijo z lovom v Evropi (20 % populacije letno; Krofel et al., v recenziji), vendar kljub temu še vedno prihaja do mnogih konfliktov. To se sklada z izkušnjami iz tujine, kjer se je povečevanje odstrela medvedov ni izkazalo
za učinkovit ukrep za namene zmanjševanja števila konfliktov (Kontio et al. 1998, Treves et al. 2010).
Pozitivna posledica splošnega odstrela medvedov bi lahko bil vpliv na vedenje medvedov. Vpliv lova na vedenje je sicer pri medvedih precej manjši kot pri vrstah, ki živijo v skupinah, saj so medvedi večinoma samotarski in zato je pri odstrelu le redko priložnost, da bi se kaj naučili (Herrero 2002). Kljub temu pa naj bi
bilo dolgotrajno preganjanje medvedov v Evropi eden glavnih razlogov, zakaj so evropski medvedi bolj plašni od rjavih medvedov v Aziji in Severni Ameriki (Herrero 2002). Tudi pregled raziskav po Evropi in Aziji je pokazal, da so v splošnem medvedi bolj plašni tam, kjer se jih lovi, vendar avtor hkrati opozarja, da na
plašnost medvedov precej bolj vpliva dostopnost do hrane antropogenega izvora ter da učinki lova pridejo do izraza šele, ko se omeji dostop do človeške hrane (Swenson 1999).
7.1.2 Odstrel konfliktnih osebkov
Glede na to, da je število konfliktov precej bolj odvisno od števila konfliktnih medvedov kot od številčnosti celotne populacije (Zedrosser et al. 1999, Witmer & Whittaker 2001, Huber 2010), je izredni odstrel konfliktnih osebkov vsekakor smiselna in pogosto učinkovita rešitev (Gunther et al. 2004). To še posebej
velja za območja kot je Slovenija, kjer populacija medvedov takšne posege prenese brez večjih vplivov na populacijsko dinamiko. Medveda je potrebno odvzeti iz populacije čim hitreje, ko se ugotovi, da gre za konflikten osebek, saj se s tem zmanjša število konfliktov in s tem vpliv na odnos javnosti do medveda
(Simonič 1994). Pri izvajanju takšnega odstrela je seveda nujno, da se odstreli dejansko medveda, ki je povzročal konflikte, kar je zaradi velikih razdalj, ki jih v kratkem času prepotujejo medvedi, lahko težavno, kadar se odstrel ne izvaja ob samih mestih s konflikti, ampak v bližnji okolici, npr. na krmiščih (glej poglavje
6.1).

118
Kljub učinkovitosti odstrela konfliktnih osebkov, pa ta ukrep ni vedno tudi dolgoročna rešitev (Sagor et al. 1997). To velja predvsem v primerih, ko se konfliktni medvedi redno pojavljajo na istih območjih. To kaže na to, da gre za posamezne konfliktne lokacije oziroma objekte (npr. določeni smetnjaki v naseljih, ki
medvedom ne preprečujejo dostopa do organskih odpadkov), ki prožijo razvoj konfliktnih osebkov. Odstranjevanje vedno novih medvedov na takšnih mestih ne predstavlja dolgoročne rešitve, saj kmalu na istem mestu pride do novih konfliktov z drugimi medvedi (Gunther et al. 2004). Odstranjevanje vedno novih
medvedov na takšnih mestih vodi do nastanka ponora populacije, kar lahko pomembno vpliva na širše območje (Knight et al. 1988), obenem pa ne reši nezadovoljstva lokalnega prebivalstva. V takšnih primerih bi konflikte z odstrelom rešili šele, ko bi na širšem območju odstrelili večino vseh medvedov. Zato je potrebno
hkrati ob odstrelu konfliktnega medveda nujno sprejeti tudi ukrepe, ki bodo na tem mestu preprečevali proženje novih konfliktov z drugimi medvedi. Pri tem se je kot najbolj učinkovito izkazalo preprečevanje dostopa do antropogene hrane (glej poglavje 7.5).
7.2 PRESELJEVANJE
Zaradi nasprotovanja javnosti proti ubijanju medvedov, se v zadnjih desetletjih vedno bolj pojavlja potreba po ukrepih za zmanjševanje konfliktov, ki ne obsega usmrtitve živali (Creachbaum et al. 1998). Možen ukrep,
s katerim se iz nekega območja medveda odstrani, vendar se ga pri tem ne ubije, predstavlja preseljevanje. Čeprav je preselitev rjavega medveda za javnost bolj sprejemljivo in v določenih situacijah lahko kratkoročno zmanjša konflikte, je po dosedanjih izkušnjah večinoma neučinkovito (Herrero 2002). V mnogih primerih se
medvedi vrnejo na mesto, od koder so bili preseljeni (tudi po več 100 km) ali pa se konflikti ob preselitvi samo prenesejo na drugo mesto (Knight et al. 1988, Vaughan et al. 1989), kar se je pokazalo tudi ob poskusih v Sloveniji (ZGS 2010), in je dokumentirala tudi pričujoča raziskava. Zato je preseljevanje smiselno uporabljati
bolj ali manj le, kadar so na voljo dovolj velika (več tisoč km2) neposeljena območja, kjer ne more prihajati do konfliktov z ljudmi in preseljenimi medvedi (Linnell et al. 1997). Poleg tega je ta ukrep zelo drag in preživetje medvedov po preselitvi je večinoma nizko (Linnell et al. 1997, Witmer & Whittaker 2001). Enako kot pri
odstrelu, preselitev tudi ni dolgoročna rešitev, saj se po preselitvi na istih mestih konflikti pogosto ponavljajo z drugimi medvedi (Gunther et al. 2004).
7.3 ODVRAČALNO KRMLJENJE
Po svetu je namerno krmljenje medvedov na večini območij prepovedano oziroma omejeno, ponekod (tudi v Sloveniji) pa se pogosto uporablja, med drugim tudi z namenom preprečevanja konfliktov med medvedi in ljudmi. Pri odvračalnem krmljenju se v predpisani oddaljenosti od najbližjega naselja v naravno okolje polaga
hrana za medveda, s čimer se želi vplivati na gibanje medvedov, ki bi zaradi tega redkeje zahajali v bližino naselij (Adamič 2005). Glede učinkovitosti te metode za preprečevanje konfliktov so si mnenja strokovnjakov deljena, kar je predvsem posledica pomanjkanja trdnih podatkov iz raziskav, ki bi se tarčno posvetila tej temi
(Sahlen 2007). Nekatere izkušnje iz krmljenja črnih medvedov kažejo, da se lahko s krmljenjem prepreči konflikte, ki jih povzročajo medvedi habituirani na človeka (Rogers 2009) in deloma zmanjšajo škodo na drevesih (Witmer & Whittaker 2001). Po drugi strani pa opažanja pri rjavih medvedih kažejo, da lahko
dopolnilno krmljenje poveča habituacijo medvedov na človeka in pogojevanje človeka z antropogeno hrano ter s tem ustvarja problematične medvede, še posebej pri mladih medvedih (Zedrosser et al. 1999, Budic

119
2010). Pri tem je verjetno pomembno, koliko stikov z ljudmi (oziroma njihovim vonjem) imajo medvedi na krmiščih. Zato bi bilo smiselno vzpodbujati uporabo avtomatskih krmilnic in omejevati prisotnost ljudi na krmiščih, s čimer se zmanjša verjetnost stikov medvedov in ljudi ob prisotnosti hrani. Ob takšni uporabi bi
krmljenje lahko prispevalo k zmanjševanju zahajanja medvedov v bližino človeka, še posebej, če bi se pri določanju minimalne razdalje od krmišča do naselja upoštevale medvedove gibalne razdalje in velikost domačih okolišev in če bi bil obenem preprečen dostop do hrane v bližini naselij.
Rezultati pričujoče raziskave kažejo, da krmljenje močno vpliva na prostorsko razporeditev medvedov, saj so
se krmišča izkazala kot ena ključnih spremenljivk, ki vplivajo na njegov celoletni habitatni izbor in mu prestavljajo nekake eko-centre, poleg tega imajo medvedi na območjih z intenzivnim krmljenjem manjša območja aktivnosti od medvedov, ki se več hranijo v prosti naravi. Zato bi lahko sklepali, da je mogoče s
krmljenjem odtegniti medvede stran od človeka, globlje v gozd, in s tem manjšati pogostnost njihovih konfliktov s človekom. Vendar pa naše analize tega ne potrjujejo. Obratno. Večina opravljenih analiz ni pokazala nobenih povezav med pogostnostjo zahajanja medveda v bližino naselij in pogostnostjo rabe
krmišč. Edina analiza ki to povezavo nakazuje, razkriva, da medvedi, ki dlje časa niso uporabljali krmišč pogosteje zahajajo v bližino človeka. Vendar pa so take situacije (ko medved več dni ne obišče nobenega krmišča) tako redke, da so v praksi – v prizmi zmanjševanja konfliktov – nepomembne, oz. ta povezava
pojasnjuje le izredno majhen del variabilnosti zahajanja medveda v bližino človeka. Pričujoča telemetrijska raziskava torej nakazuje, da je prostor v Sloveniji (isto velja ca celo Evropo) preveč fragmentiran in so odročni strnjeni bloki gozda premajhni, da bi s krmišči medvede lahko odtegnili dovolj daleč, da bi to preprečevalo
njihova zahajanja k človeku.
Vsekakor pa bi bilo raziskave vpliva krmljenja na konflikte poglobiti in jih razširiti s študijami vplivov različnih tipov hrane na gibanje medvedov, preden bo moč dajati zanesljiva priporočila. V Sloveniji se na primer v praksi pogosto veliko vlogo k zmanjševanju plenjenje drobnice pripisuje krmljenju z mrhovino. Le ta naj bi na
plenjenje drobnice vplivala prek več vzvodov. Po laičnih utemeljitvah, ki so anekdotične, medved kot vsejed potrebuje tudi beljakovinsko hrano in potrebe po njej zadosti bodisi na mrhoviščih, ali pa s plenjenjem drobnice. Dodatno naj bi bila mrhovina prehransko toliko bolj privlačna od koruze in žitaric, da naj bi
uspešno odtegnila medveda od naselij in naj bi na njih medved preživel več časa, kar naj bi oboje dodatno pripomogle k manj plenjenju drobnice. Vendar pa domača raziskava prehrane medveda (Kavčič et al. 2011) kaže, da je mrhovina pomembna prehranska komponenta medveda le spomladi, ne pa tudi poleti in jeseni,
ko je plenjenje drobnice pogosto, četudi je mrhovina dostopno večji del leta. Poleg tega so od beljakovinskih virov hrane pomembnejši od mrhovine domačih živali nevretenčarji in divji parkljarji (najdene poginule živali).
Pri priporočilih glede načina uporabe krmišč za medveda potrebno upoštevati tudi druge posledice, ki jih ima
takšno krmljenje in niso nujno povezane s konflikti. Z uporabo krmišč se na primer precej olajša spremljanje populacije medvedov, lažje se izvaja redni odstrel (možnost boljše presoje, manjša verjetnost zastreljenih medvedov), obenem pa lahko pričakujemo, da se zaradi intenzivnega krmljenja poveča nosilna kapaciteta
okolja, zviša reprodukcija in preživetje medvedov ter zmanjša pomen medvedov v ekosistemskih funkcijah, ki jih opravljajo (razširjenje plodonosnih rastlinskih vrst, pospeševanje dekompozicije živalskih trupel itd.).

120
7.4 POGOJEVANJE IZOGIBA
Pogojevanje izogiba dosežemo, če osebek ob določenih situacijah (npr. ob približevanju medveda naseljem) izpostavljamo negativnim dražljajem in s tem poskušamo doseči, da bi ta osebek začel povezovati takšne
situacije / lokacije s slabo izkušnjo in se jim zato v prihodnje izogibal. Ta način asociativnega učenja poskušajo izkoristiti upravljavci, tako da medvede, ki se približajo ljudem, streljajo z gumijastimi izstrelki, ki povzročajo bolečino, vendar medveda ne ubijejo. Na ta način poskušajo naučiti medvede, da je bližina človeka in naselij
nevarna (Herrero 2002). Namesto gumijastih izstrelkov se uporabljajo tudi razna razpršila, glasno pokanje, posebej izurjeni psi, ki agresivno preganjajo medveda ipd.
V splošnem so se takšni načini dolgoročno večinoma izkazali za neuspešne pri preprečevanju konfliktov med medvedi in ljudmi. Kljub kombiniranju več načinov pogojevanja izogiba se je večina črnih medvedov vrnila na
urbane površine v roku 40 dni po plašenju (Beckmann et al. 2004). Prav tako s pogojevanjem izogiba niso uspeli preprečiti konfliktov z medvedi v Avstriji (Zedrosser et al. 1999), le pri nekaterih medvedih pa so bili uspešni v Italiji (C. Groff, ustno sporočilo). V splošnem je pogojevanje izogiba manj uspešno pri medvedih, ki
so se že habituirali na človeka in so pogojeni na hrano človeškega izbora. Ta ukrep bo zato učinkovit le, če se reagira dovolj hitro in se medveda plaši že ob prvih konfliktnih situacijah, ko še ni prišlo do habituacije (Schirokauer & Boyd 1998) ter če se obenem preprečuje nadaljnji dostop do antropogene hrane (Herrero
2002).
7.5 OMEJEVANJE DOSTOPA DO HRANE ANTROPOGENEGA IZVORA
Hrana antropogenega izvora, ki je dostopna medvedom, je najpogostejši vzrok, ki privede do konfliktov med
medvedi in ljudmi, kar posredno nazorno nakazuje tudi pričujoča raziskava (glej poglavji 2.2.2 in 5.7.2). Zato je preprečevanje dostopa do takšne hrane eden najpomembnejših ukrepov za učinkovito zmanjševanje konfliktov med medvedi in ljudmi. Do tega spoznanja so prvi prišli v narodnih parkih v Severni Ameriki kot
posledica več napadov na ljudi (nekaterih s smrtnim izidom), ki so jih v 70-ih in 80-ih letih prejšnjega stoletja povzročili na človeka habituirani in pogojeni medvedi (Herrero 1994). Kot posledica teh tragičnih dogodkov so takrat začeli z uvajanjem strogega režima, kar se tiče upravljanja s človeškimi odpadki, shranjevanjem
hrane in neposrednega krmljenja medvedov (s kaznimi do $150 000 in pol leta zapora). Podobne ukrepe so začeli sprejemati tudi zunaj parkov, kjer se človeška naselja prekrivajo z območjem razširjenosti medvedov (Wilson et al. 2006). Zaradi preprečevanja dostopa do človeške hrane in smeti medved ni več nagrajen ob
stikih z ljudmi in zato ohranja strah pred človekom ter precej redkeje zahaja v bližino naselij (Knight et al. 1988, Herrero 2002, Herrero et al. 2005). Posledično so s takšnimi ukrepi marsikje uspeli močno zmanjšati število konfliktov med medvedi in potrebo po odstrelu medvedov (za konkretne primere glej poglavje 5).
Zato preprečevanje dostopa do antropogene hrane kot nujen upravljavski ukrep predpisuje tudi Akcijski načrt za upravljanje z rjavim medvedom v Evropi (Swenson et al. 2000).
Obstaja več načinov, kako medvedom uspešno preprečiti dostop do antropogene hrane (za podrobnosti glej navodila v prosto dostopni publikaciji Sowka 2009). Dostopnost do organskih odpadkov in drugih človeških
odpadkih v naseljih je možno zanesljivo preprečiti z uporabo t.i. ''medvedo-varnih'' (bear-proof) smetnjakov, ki omogočajo preprosto odlaganje odpadkov in praznjenje za ljudi, poseben pa mehanizem preprečuje, da bi medved odprl pokrov in prišel do hrane. Po svetu je danes že kar nekaj proizvajalcev takšnih specializiranih

121
smetnjakov. Možno pa je tudi dokaj poceni prirediti že obstoječe zabojnike, ki se nahajajo na lokacijah, kjer redno prihaja do konfliktov, kot so na primer storili v Italiji (Groff et al. 2010). Dostopnost do smeti je možno preprečiti tudi s skladiščenjem zabojnikov z odpadki v zaprtih ali zamreženih prostorih (podobno kot je že
praksa v mnogih mestih v Sloveniji, tam sicer iz drugačnih razlogov) ali z zavarovanjem zabojnikov z električnimi ograjami in rednim odvažanjem smeti. Izkušnje kažejo, da je najučinkoviteje, če so načini shranjevanja hrane in odpadkov čim bolj preprosti (Creachbaum et al. 1998) in če zahtevajo čim manj
dodatnega dela za ljudi, ki jih uporabljajo (C. Groff, osebno sporočilo).
Za preprečevanje škod na čebelnjakih, vrtovih in sadovnjakih se je tako v tujini kot pri nas kot učinkovit ukrep izkazalo preprečevanje dostopa z uporabo električnih ograj in električnih mrež (Zedrosser et al. 1999, Herrero 2002, Gunther et al. 2004). Premoženje je mogoče na ta način dokaj preprosto zaščititi, vendar je
potrebna dovolj visoka napetost – vsaj 10 000 V na izvoru in najmanj 6 000 V na najbolj oddaljeni točki (Wilson 2007). Pomembno je tudi, da so elektro-ograje delujoče ves čas, ko so postavljene in da se jih odstrani, ko jih ne potrebujemo več. Ob postavljenih ograjah, ki niso pod napetostjo se namreč lahko
medvedi naučijo, kako jih je mogoče prečkati (Wilson et al. 2006).
Pravilna uporaba električnih ograd tudi občutno zmanjša škode na domačih živalih (Sowka 2009). Kot dodaten ukrep lahko pomaga tudi zapiranje živine v zaprte prostore preko noči (pri tem je potrebno upoštevati moč, ki jo lahko uporabi medved za vdiranje v šibkejše ograde in hleve) in hitro odstranjevanje
trupel živali iz neposredne bližine pašnikov (Witmer & Whittaker 2001, Wilson 2007, Sowka 2009). Ker so za napade medvedov daleč najbolj dovzetne ovce (Projekt LIFE02NAT/SLO/8585 2005), mnogi avtorji v novejšem času kot ukrep za zmanjševanje konfliktov priporočajo zmanjšanje reje drobnice na območju
medveda (Sagor et al. 1997, Gunther et al. 2004, Swenson & Andre 2005, Wilson et al. 2006), kot so to z dobrimi rezultati že izvedli ponekod v Severni Ameriki (Mattson 1998). Kjer je to mogoče, se kot dobro alternativo ovčereji priporoča predvsem preusmeritev na govedorejo (MKGP 2002, Zimmermann et al.
2003) ali konjerejo, podobno kot to velja za zmanjševanje škod, ki jih povzročajo volkovi (Černe et al. 2010). Izkušnje namreč kažejo, da se ob prehodu iz ovčereje na govedorejo, napadi ne prenesejo na govedo.
Posebno pozornost je treba nameniti neposrednemu krmljenju medvedov ob stiku s človekom. Takšni primeri najhitreje prožijo pogojevanje človeka z antropogeno hrano in habituacijo na prisotnost človeka.
Takšno početje je potrebno zakonsko prepovedati in resno reagirati ob vsakih prestopkih. Obenem je potrebno tudi ozaveščanje javnosti, saj si drugače ljudje pogosto ne predstavljajo posledic takšnega ravnanja.
Poleg proženja konfliktov med medvedi in ljudmi je preprečevanje dostopa do hrane antropogenega izvora
pomembno tudi za zmanjševanje drugih stranskih učinkov, ki jih ima hranjenje s človeško hrano, kot je vpliv na velikost domačih okolišev, nenaravno povečanje telesne teže, lokalno povečanje gostot medvedov ob virih hrane in zmanjšanje v sosednjih območjih, spremembe v spolnem razmerju in spolnem razmerju itd.
(Beckmann & Berger 2003).
Preprečevanje dostopa do antropogene hrane je poleg same učinkovitosti priporočljivo tudi zato, ker se na ta način konflikte preprečuje v naprej in ne šele, ko do njih že prihaja. Potrebno pa je upoštevati, da je ukrep lahko manj učinkovit ob prisotnosti medvedov, ki so že močno habituirani na prisotnost človeka in pogojeni
na človeško hrano. V teh primerih je potrebno ta ukrep izvajati skupaj z izrednim odstrelom teh konfliktnih medvedov.

122
7.6 OZAVEŠČANJE JAVNOSTI
Raziskava napadov medvedov na človeka je pokazala, da se morajo ukrepi za zmanjševanje verjetnosti napada medvedov osredotočiti predvsem na obnašanje ljudi v okolju z medvedi, saj se lahko veliko napadov
medvedov zmanjša z uporabo znanja o obnašanju in ekologiji medvedov (Herrero & Higgins 1999, 2003). Podobno velja za tudi uspešno reševanje drugih konfliktov z medvedi, zato je ozaveščanje javnosti pomemben del upravljanja z medvedom (Witmer & Whittaker 2001, Beckmann et al. 2004, WSPA 2009).
Ljudje se bodo precej bolj držali določenih pravil, če bodo vedeli, kaj je njihov namen in kako preprečujejo nastajanje konfliktov.
Raziskava med lokalnim prebivalstvom v Sloveniji je pokazala, da le malo ljudi dobi informacije o pravilnem ravnanju na območju medveda (Sinquin et al. 2006). V tujini so eden glavnih virov informacij o pravilnem
ravnanju na območju medveda narodni parki (Herrero 1994). V Sloveniji naš edini narodni park, ki se nahaja na prehodnem življenjskem območju rjavega medveda, tej vrsti ne namenja posebne pozornosti. Zato bi bilo potrebno izvesti širšo izobraževalno akcijo med prebivalstvom na območju medvedov.
7.7 UPORABA RAZPRŠIL ZA ODVRAČANJA MEDVEDOV OD NAPADA NA LJUDI
Za namene preprečevanja napadov medveda na ljudi so raziskovalci poskušali z uporabo različnih sredstev. Kot najbolj učinkovito in z relativno preprosto uporabo se je izkazalo razpršilo narejeno na osnovi popra, ki
vsebuje kaspaicin (Herrero 2002). Testiranje v oborah in podatki iz dejanske uporabe v naravi ob napadih medvedov so pokazali, da uporaba takšnega razpršila v večini primerov (92 %) ustavi nezaželeno agresivno vedenje pri rjavem medvedu. Do poškodb ljudi ob napadu medveda je ob uporabi razpršila prišlo v 2 %
primerov (Smith et al. 2008). Čeprav je težko sklepati, kaj bi se v teh primerih zgodilo brez uporabe razpršila, je splošno sprejeto mnenje, da je uporaba tega sredstva priporočljiva (Herrero 2002, Gunther et al. 2005, Smith et al. 2008). Nošenje razpršila ob gibanju v naravnem okolju bi lahko bila priporočljiva tudi za
pomiritev ljudi z izrazito fobijo pred medvedi. Obenem pa se podobno kot ob nošenju orožja, ne smemo popolnoma zanašati na razpršilo in se zato obnašati nespametno ob srečanjih z medvedi (Herrero 2002). Omeniti velja še, da je uporaba razpršila učinkovita le ob neposrednih srečanjih z medvedom, ko se curek
usmeri direktno v medveda, medtem ko ima škropljenje določenih objektov ali oseb z namenom odganjanja medvedom nasproten učinek, saj vonj, ki ostane po uporabi razpršila medvede očitno celo privlači (Smith et al. 2008).
7.8 SPREMEMBA RABE PROSTORA
V določenih primerih lahko verjetnost konfliktov z medvedi zmanjšamo s spremembo v rabi prostora na območju medveda. Medvedi se počutijo bolj varne, če imajo na voljo kritje (npr. v obliki goste vegetacije).
Zato je verjetnost prisotnosti medvedov v bližini naselij (in s tem verjetnost postopne habituacije in odkritja potencialnih virov antropogene hrane v vaseh) večja, kjer je neposredna okolica naselij zaraščena z gosto vegetacijo. Tako bi bil eden izmed možnih ukrepov tudi vzdrževanje oziroma povečevanje odprte krajine v
okolici človeških bivališč. V drugih primerih lahko verjetnost konfliktov zmanjšamo tudi s prehodom na manj konfliktno rabo prostora, npr. iz ovčereje v govedorejo ali konjerejo, spremembe v izbiri poljščin,

123
spremembe v lokaciji posevkov oziroma živine in spremembe v rabi prostora v okoliških habitatih (Witmer & Whittaker 2001).
Pri načrtovanju s prostorom in poseganjem v naravno okolje je potrebno v čim večji možni meri zmanjšati učinke fragmentacije prostora. To je predvsem pomembno, ko se načrtuje poseg na lokacijah, ki služijo kot
prehod medvedov iz ene v drugo večjo habitatno krpo. Na ta način se bo ohranjala povezljivost habitatov, zmanjševali učinki izolacije, obenem pa tudi zmanjšala verjetnost konfliktov, do katerih bi lahko prišlo zaradi zahajanja medvedov na nove urbane površine, ki so bile postavljene na mesta tradicionalnih prehodov
medvedov.
7.9 POVEČANJE PROPUSTNOSTI PROMETNIC
Prometne nesreče, v katerih pride do trka z medvedom, se pogosto ponavljajo na istih lokacijah (poglavje
2.1.2). Tako gre v mnogih primerih za dokaj predvidljiv vir prometnih nesreč, ki se bodo zagotovo na teh mestih nadaljevale, če ne bo ukrepanj v obliki primerne ureditve cestišča. Ta vključuje izgradnjo primernega prehoda, ki živalim omogoča nenevarno prečkanje cest (t.i. »zeleni mostovi«, podhodi in mostovi),
predvsem pa vzporedno postavitev električne ograje na tem cestnem odseku. Sama elektrifikacija se je izkazala za učinkovit ukrep za preprečevanje prečkanja cest (npr. pri avtocestnem odseku Vrhnika – Logatec, malo nad Vrhniko), vendar tak ukrep brez vzporedne ureditve varnega prehoda ne poveča propustnosti
prometnic in lahko prometne nesreče samo preusmeri na drugo lokacijo.
7.10 IZPLAČEVANJE ODŠKODNIN
Ker smo se kot družba odločili, da želimo ohranjati rjavega medveda, ob zavedanju, da ta občasno ljudem
povzroča škodo, je pravično, da finančnega bremena ne prenašajo samo tisti prebivalci, ki živijo na območju medveda, ampak se to porazdeli med vso prebivalstvo. To je doseženo z ukrepom izplačevanje odškodnin oškodovancem iz proračunskega denarja, ki so utrpeli škodo zaradi aktivnosti medvedov. Raziskave so
pokazale, da je izplačevanje odškodnin lahko učinkovito za ohranjanje tolerance do velikih zveri med lokalnim prebivalstvom (čeprav ne vedno) in v določenih primerih za zmanjšanje nezakonitega odstrela (Naughton-Treves et al. 2003, Treves et al. 2009). Vendar pa je pri izplačevanju odškodnin potrebna
previdnost, da se ne doseže nasprotni učinek od želenega in preprečevati zlorabe. Kot prvo se je potrebno zavedati, da se z izplačevanjem odškodnin ne preprečuje konfliktov, zato je potrebno ta ukrep uporabljati v povezavi z drugimi ukrepi, ki vplivajo na verjetnost pojavljanja konfliktov (WSPA 2009). Dobro načrtovanje
sistema izplačevanja odškodnin je ključno, saj prevelike ugodnosti za oškodovance ne vzpodbujajo preprečevanja pojavljanja konfliktov (Zabel & Holm-Müller 2008, WSPA 2009). Odškodnine ne smejo predstavljati oblike dodatnih subvencij in prispevati k ohranjanju stalnega stanja konflikta, saj si oškodovanci
ne želijo izgubiti trenutnih ugodnosti (Cozza et al. 1996, Boitani & Ciucci 2009 – v Musiani et al. 2009). Potrebna je tudi dobra kontrola, da bi se preprečevale zlorabe in izkoriščanje sistema (Bulte & Rondeau 2005, Treves et al. 2009). Slabo načrtovanje odškodnin lahko zato prinese več negativnih kot pozitivnih
posledic za ohranjanje konfliktnih vrst živali ter privede do dodatnega zaostrovanja konfliktov (Bulte & Rondeau 2005) in nejevolje med oškodovanci.

124
Preglednica 19: Pregled glavnih tipov konfliktov med medvedi in ljudmi ter najpomembnejši ukrepi, s katerimi je možno
po dosedanjih izkušnjah učinkovito preprečevati te konflikte. Podčrtani so ukrepi, ki so preventivni in s katerimi je možno
preprečiti konflikte še preden do njih pride oziroma preprečiti njihovo ponavljanje. Z normalno pisavo so zapisani ukrepi, s
katerimi lahko preprečimo konflikt, do katerega prihaja v danem trenutku, običajno pa nimajo dolgoročnega učinka.
TIP KONFLIKTA: GLAVNI UKREPI ZA UČINKOVITO PREPREČEVANJE KONFLIKTA:
Škoda v živinoreji - zaščita živine z elektro-ograjami ali elektro-mrežami
- zapiranje živine v varne ograde/hleve preko noči
- odstrel konfliktnih medvedov
- izbira manj konfliktne vrste živine
Škoda na čebelnjakih, poljščinah, vrtovih, sadovnjakih in drugem človekovem premoženju
- zaščita premoženja z elektro-ograjami ali elektro-mrežami
- odstrel konfliktnih medvedov
Prometne nesreče - na ključnih prehodih ureditev cestišča, da omogoča prehod medvedov brez nevarnosti prometne nesreče
Zahajanja medvedov v naselja - preprečevanje dostopa medvedom do antropogene hrane
- odstrel konfliktnih medvedov
- izobraževanje javnosti
Napadi medvedov na človeka - izobraževanje javnosti
- preprečevanje dostopa medvedom do antropogene hrane in s tem zmanjševanje habituacije medvedov na človeka in pogojevanja prisotnosti človeka z antropogeno hrano
- uporaba razpršila za odvračanje napada medvedov
- odstrel medvedov, ki kažejo jasno agresivno vedenje ob prisotnosti človeka
Ogrožanje populacije medvedov - ohranjanje pozitivnega odnosa javnosti do medvedov preko učinkovitega zmanjševanja ostalih konfliktov in izobraževanja javnosti
- preprečevanje fragmentacije prostora

125

126
8 KONKRETNI PRIMERI USPEŠNEGA REŠEVANJA KONFLIKTOV V TUJINI
Do danes je znanih že kar nekaj primerov, ki so jasno pokazali, da je mogoče s pravo politiko upravljanja
učinkovito zmanjšati konflikte med medvedom in človekom. V tem poglavju predstavljamo nekaj takšnih konkretnih primerov iz tujine, tako iz zavarovanih območij namenjenih predvsem turistom kot iz zasebnih zemljišč s stalno naseljenim lokalnim prebivalstvom.
8.1 NARODNI PARKI
V narodnem parku Yellowstone so imeli do 80-ih let 20. stoletja veliko težav zaradi konfliktov med obiskovalci parka in medvedi, vključno z mnogimi napadi na ljudi, nekaj celo s smrtnimi žrtvami, obenem pa
so morali vsako leto odstreliti večje število medvedov, ki so postali konfliktni, kar je že ogrožalo populacijo medvedov v parku. Zaradi teh konfliktov so začeli z uvajanjem strogih pravil glede hranjenja medvedov, skladiščenja hrane in smeti, pri čemer so priredili smetnjake, da so postali odporni na medvede, zaprli
smetišča znotraj parka, redno odvažali odpadke ter uredili prostore za varno shranjevanje hrane preko noči na prostorih za taborjenje. Obenem so redno izobraževali ljudi o pravilnem skladiščenju smeti in hrane ter o možnih posledicah, če se ne upošteva teh pravil, še vedno pa so tudi odstranjevali konfliktne medvede. S
temi novimi ukrepi so uspeli močno zmanjšati konflikte med medvedi in ljudmi, saj se je število napadov ljudi zmanjšalo za 89 % (iz 19,5 na 0,3 napadov na milijon obiskovalcev na leto), zmanjšalo se približevanje medvedov k poslopjem in tabornim prostorom, obenem pa se je precej zmanjšala tudi potreba po odstrelu
in preseljevanju konfliktnih medvedov (Meagher & Phillips 1983, Gunther & Hoekstra 1998).
V narodnem parku Denali so z izobraževanje obiskovalcev, uvajanjem novih pravil shranjevanja hrane, zapiranjem nekaterih območij za ljudi in hitrim reagiranjem na konflikte s pogojevanjem izogiba pri medvedih še preden je prišlo do dokončne habituacije na človeka dosegli, da so se primeri hranjenja
medvedov na antropogenih virih hrane zmanjšali za 96 %, s čimer se je zmanjšalo število konfliktov med medvedi in ljudmi za 77 %, prav tako za 77 % pa se je zmanjšalo tudi število odstranjenih konfliktnih medvedov (Schirokauer & Boyd 1998).
S pomočjo omejevanja dostopa medvedom do človeških odpadkov so v narodnem parku Jasper zmanjšali
potrebo po odstrelu pri črnih medvedih za 94 % in pri rjavih medvedih za 86 % (Ralf 1995).
V narodnem parku Glacier so po spremembi načina upravljanja z medvedi, ko so iz odstrela in preseljevanja medvedov preusmerili pozornost predvsem v spreminjanje vedenja ljudi, s pomočjo omejevanjem dostopa do antropogene hrane (redno odvažanje smeti, stroga prepoved hranjenja medvedov, uporaba »medvedo-
varnih« smetnjakov) in izobraževanja ljudi močno zmanjšali konflikte, potreba po odstrelu pa se je zmanjšala za 75 % pri črnih in za 70% pri rjavih medvedih, čeprav se je številčnost medvedov v tem času povečala (Gniadek & Kendall 1998).

127
8.2 OBMOČJA S STALNO NASELJENIMI LOKALNIMI PREBIVALCI
Po spremembi strategije upravljanja z medvedi v Kennecott Valley (ZDA), ko so se preusmerili predvsem na
izobraževanje lokalnega prebivalstva in lokalne skupnosti opremili z električnimi ogradami in »medvedo-varnimi« smetnjaki, se je število konfliktov močno zmanjšalo in število medvedov, ki so jim morali odstreliti se je zmanjšalo iz 9 do 12 na leto pred uvedbo teh ukrepov na 0 po spremembi upravljanja (Wilder et al.
2007).
Na zasebnih zemljiščih v Zahodni Montani (ZDA) se je do leta 2003 število konfliktov med lokalnim prebivalstvom in medvedi povečevalo, predvsem zaradi naraščajočih škod na čebelnjakih, živini in zahajanjem medvedov v naselja. Leta 2003 so začeli z večjim projektom, v okviru katerega so začeli z
odvažanjem klavniških odpadkov in trupel živali iz okolice kmetij, ozaveščanjem javnosti o ravnanju s smetmi ter postavljanjem električnih ograj okoli pašnikov z živino, čebelnjakov in deponij odpadkov. S temi ukrepi so v treh letih uspeli zmanjšati število vseh konfliktnih dogodkov za 91 %, pri čemer ni bilo potrebno odstreliti
ali preseliti niti enega medveda (Wilson et al. 2006, Wilson 2007).

128

129
9 UPRAVLJAVSKA PRIPOROČILA ZA UČINKOVITO PREPREČEVANJE KONFLIKTOV MED MEDVEDI IN LJUDMI V SLOVENIJI
Uspešno upravljanje in preprečevanje konfliktov med rjavim medvedom in človekom je osnova za zagotavljanje čim lažjega soobstoja lokalnega prebivalstva s to vrsto. Dolgoletne izkušnje z upravljanjem z medvedi v Sloveniji so nam omogočile uspešno nadzorovanje velikosti populacije medvedov in strokovno
korektno poseganje vanjo glede na zastavljene upravljavske cilje tudi v obdobju, ko še ni bilo na voljo natančnih podatkov o absolutni številčnosti medvedov (Krofel et al. 2010). Odvzem osebkov iz narave bo nedvomno ostal pomemben upravljavski ukrep, še posebej ob pojavu problematičnih osebkov. Vendar sam
odstrel očitno ne bo dovolj, da nam bo uspelo zmanjšati sedanje visoko število konfliktov med medvedi in ljudmi na želeni nivo.
Drugi glavni ukrep poleg odstrela za preprečevanje konfliktov v Sloveniji je do sedaj predstavljalo odvračalno krmljenje, tudi z mrhovino. Čeprav raziskave glede učinkov krmljenja še niso povsem končane, dosedanji
podatki in rezultati pričujoče raziskave (Adamič et al. 2005, Kavčič et al. 2011, Jerina et al. 2011) kot tudi trendi pojavljanja konfliktov močno nakazujejo, da ta ukrep ni učinkovito sredstvo za preprečevanje konfliktov. Večina avtorjev sicer ne podpira drastične ukinitve krmljenja (Adamič 2005, Krofel et al. 2008), saj
je trenutno krma pomembna prehranska komponenta medveda in pokrije približno tretjino vseh njegovih energijskih potreb. Zato bi njegova nenadna ukinitev - v aktualni populaciji na ta vir hrane kondicioniranih osebkov - verjetno imela močne negativne posledice. Vendar pa bi bilo treba začeti izvajati tudi druge in bolj
učinkovite ukrepe. To se kaže tudi pri odnosu tistega dela prebivalstva, ki je najbolj izpostavljen, saj je raziskava pokazala, da se rejci živine v Sloveniji strinjajo z ohranjanjem medveda na njihovem območju, ne pa s trenutnim upravljanjem, ki ni dovolj učinkovito pri zmanjševanju konfliktov in škod v kmetijstvu zaradi
medveda (Sinquin et al. 2006). Očitno bo potrebno začeti z uvajanjem dodatnih ukrepov, pri čemer je smiselno uporabiti te, ki so se v tujini že izkazali za zelo učinkovite.
Ob pregledu konfliktov med medvedi in ljudmi po svetu, izkušenj z različnimi ukrepi za preprečevanje konfliktov ter na podlagi rezultatov naše raziskave, je očitno, da je povod večine konfliktov hrana
antropogenega izvora in da je zato preprečevanje dostopa medvedom do te hrane, še posebej v neposredni bližini človeških bivališč, ključen ukrep, s katerim je mogoče uspešno zmanjšati število konfliktov. To velja tako za območja znotraj parkov in drugih zavarovanih območij, kot za zasebna zemljišča s stalno naseljenim
lokalnim prebivalstvom, kakršna je situacija na večjem delu razširjenosti medveda v Sloveniji. V mnogih državah, kjer še živijo rjavi medvedi, je zato danes preprečevanje dostopa do antropogene hrane eden izmed glavnih upravljavskih ukrepov. To velja tudi za območja, kjer imajo z upravljanjem z medvedom precej manj
izkušenj kot pri nas – npr. v italijanskih Alpah (Groff et al. 2010). Nasprotno pa v Sloveniji še vedno nimamo niti enega samega ''medvedo-varnega'' zabojnika za smeti, kljub temu da organske smeti v zabojnikih pogosto privabljajo medvede leto za letom na ista mesta (npr. Bloke, Rakitna, Kurešček, Loški potok). Zaradi
tega se vsako leto znova pojavlja potreba po izrednem odstrelu medvedov, ljudje pa se zaradi ponavljajočega privabljanja in zadrževanja vedno novih medvedov v bližini svojih bivališč kljub odstrelu počutijo ogrožene. Ureditev zabojnikov bi zelo verjetno dolgoročno rešilo konflikte na teh lokacijah.
Podobno velja za klavniške odpadke, razna divja odlagališča organskih odpadkov v bližini naselij in prostore za piknik, ki privabljajo medvede k človeku. Posledično ti medvedi namreč postopoma izgubljajo strah pred človekom in postanejo problematični, kar vodi v številne konflikte.

130
Zaradi tega za Slovenijo priporočamo naslednje prioritetne ukrepe, ki bi po našem mnenju lahko občutno pripomogli k zmanjšanju konfliktov med medvedom in človekom pri nas:
1.) na lokacijah s ponavljajočimi konflikti ureditev zabojnikov za smeti, tako da človeški odpadki ne bodo dostopni za medvede
2.) resno ukrepanje inšpekcijskih služb ob nezakonitem odlaganju klavniških odpadkov v neposredni bližini
človeških bivališč
3.) zaščita kompostov, čebelnjakov, drobnice, sadovnjakov in drugih virov hrane, ki redno privabljajo medvede v bližino naselij, s pomočjo električnih ograd primerne višine in električne napetosti v času, ko so ti viri hrane atraktivni za medvede (npr. v času obroda sadežev)
4.) vzpodbujanje govedoreje in konjereje namesto reje drobnice na osrednjem območju razširjenosti
medveda, kjer okoljski dejavniki to dopuščajo
5.) prepoved neposrednega krmljenja medvedov (tudi mladičev!) ob prisotnosti človeka in resno ukrepanje ob kršenju te prepovedi
6.) izvedba širše izobraževalne kampanje na območju stalne prisotnosti medveda o pravilnem obnašanju ljudi za zmanjšanje konfliktov z medvedi s poudarkom na pomenu preprečevanje dostopa do hrane
antropogenega izvora
7.) ureditev problematike trkov medvedov z vozili
Obenem tudi še naprej priporočamo izvajanje izrednega odstrela konfliktnih medvedov, ki kažejo očitne znake habituacije na človeka.
Kot je zapisal dr. Miha Adamič: »Najpomembnejše orodje za preprečevanje in razreševanje konfliktov med rjavim medvedom in človekom je znanje.« Znanja o medvedih v Sloveniji je sedaj že precej. Zdaj je
pomembna še volja, da se to znanje tudi uporabi v praksi. V tujini so bili učinkoviti ukrepi glede omejevanje dostopnosti hrane medvedov običajno sprejeti šele po napadih na ljudi s smrtnimi žrtvami (Herrero & Higgins 1999). Upamo, da bomo v Sloveniji s takšnimi ukrepi začeli že prej.

131

132
10 KOMUNICIRANJE Z JAVNOSTJO
10.1 UVOD
Pomemben del projekta predstavlja tudi komuniciranje z javnostjo. Po eni strani je bilo to ključno za
motivacijo ljudi, ki so sodelovali pri odlovu medvedov (člani lovskih družin in poklicni lovci lovišč s posebnim namenom, ki so daljše obdobje redno zalagali krmišča, kjer se je kasneje izvajal odlov), po drugi pa je pomembno, da je javnost kar najbolje obveščena o problematiki konfliktov z medvedom, vzrokih, ki vodijo v
konflikte in kakšne so možne rešitve. Prepričani smo, da bi se z večjim znanjem in s spremembo obnašanja ljudi na območju medveda lahko izognili marsikateri konfliktni situaciji. Poleg tega smo z ozaveščanjem lovcev o poteku projekta želeli preprečiti, da bi bili medvedi opremljeni z ovratnicami odstreljeni pred
koncem delovanja ovratnic.
10.2 NOVINARSKE KONFERENCE
Dne 19.5.2009 smo v prostorih Agencije RS za okolje izvedli novinarsko konferenco, na kateri smo dr.
Klemen Jerina, Marko Jonozovič in Miha Krofel ob dia-projekciji javnosti predstavili namen in dosedanji potek projekta. Po oceni se je konference udeležilo okoli 40 ljudi. Mnogi mediji so nato o dogodku poročali v tiskani, elektronski in video obliki. Še eno novinarsko konferenco načrtujemo ob zaključenem projektu, ko
bomo predstavili končne rezultate projekta.
10.3 SPLETNA STRAN
Na spletni domeni http://www.medvedi.si/ (PRILOGA 7) smo pripravili več spletnih strani, na katerih smo
predstavili cilje in izvedbo projekta, izdelali »osebne izkaznice« za medvede opremljene z ovratnicami, redno objavljali vmesna poročila o napredovanju projekta in karte z lokacijami teh medvedov. Slednje podatke dajemo v javnost z nekajdnevnim zamikom, da bi preprečili morebitno zlorabo. Na spletni strani bomo tudi
objavili to končno poročilo projekta.
10.4 SODELOVANJE Z MEDIJI
Tekom projekta smo opravili že prek 20 intervjujev za različne medije: televizija, radio, časopisi in revije,
spletni mediji. V intervjujih smo predstavili projekt, njegove namene, orisali ozadja konfliktov medveda s človekom ipd. Večino intervjujev je dal vodja projekta dr. Klemen Jerina. Mnogo izjav smo dali tudi v zvezi z dogajanji ob preselitvi medveda »Rožnika«. Z različnimi mediji smo sodelovali pri pripravi več novic in objav
na temo sobivanja z medvedi, med drugim smo pripravili tudi več nasvetov za preprečevanje konfliktov z medvedi v okviru spletne strani Ministrstva za okolje in prostor »Pomagajte tudi vi« (http://www.biotskaraznovrstnost.si/) in te nasvete so kasneje povzemale tudi številne strani nevladnih

133
organizacij. Sodelovali smo tudi pri pripravi dokumentarnega filma o rjavem medvedu, ki ga je posnela RTV Slovenija in v katerem je predstavljeno tudi delo projekta:
VOKAČ, Aleksandra. Rjavi medved. Ljubljana: TV Slovenije, 2009, 80 min.
10.5 PRISPEVEK K ZNANOSTI
Na podlagi podatkov pridobljenih tekom raziskave bomo pripravili več znanstvenih člankov, ki bodo prispevali k razumevanju biologije rjavega medveda in konfliktov med medvedi in ljudmi v širši mednarodni znanstveni sferi. Dosedanje znanstvene objave:
- KROFEL, M., FILACORDA, S. & K. JERINA. 2010. Mating-related movements of male brown bears on the
periphery of an expanding population. – Ursus 21(1): 23-29.
- KACZENSKY, P., JERINA, K., JONOZOVIČ, M., KROFEL, M., SKRBINŠEK, T., RAUER, G., KOS, I. & B. GUTLEB. 2011. Illegal killings may hamper brown bear recovery in the Eastern Alps. – Ursus, 22(1): 37-46.
Vmesne rezultate projekte smo mednarodni strokovni javnosti predstavili tudi na dveh konferencah na temo raziskovanja in upravljanja z medvedi (International conference on bear research and management), ki
jih je organizirala organizacija International Bear Association, in sicer 19. IBA konferenca v Tbilisju, Gruzija leta 2010 ter 20. IBA konferenca v Ottawi, Kanada leta 2011. Na konferencah smo rezultate projekta predstavljali z naslednjimi prispevki:
- JERINA, Klemen, VIDEMŠEK, Uroš, JONOZOVIČ, Marko, KROFEL, Miha. 2010. Using GPS telemetry to study
human-bear conflicts in Slovenia (predavanje)
- KROFEL, Miha, POTOČNIK, Hubert, JONOZOVIČ, Marko, JERINA, Klemen. 2010 Brown bear demography in Slovenia: managing a part of a population (predavanje)
- JERINA, K., KROFEL, M., KAVČIČ, I. 2011. Effects of supplemental feeding on bear spatial behavior, habituation on people, and human-bear conflicts: studies of long-term intensive feeding of brown bears
in Slovenia, Europe (predavanje)
- KROFEL, M., KOS. I., JERINA, K.. 2011. The Noble Cat and the Big Bad Scavenger: the effects of kleptoparasitism by brown bears on Eurasian lynx. V: 20th International conference on bear research and management (predavanje)
- KAVČIČ, I. ADAMIČ, M., KROFEL, M., JERINA, K., KACZENSKY, P. 2011. Brown bear food habits in human
dominated landscapes of Slovenia: importance of intensive long-term supplemental feeding (poster)

134
10.6 POLJUDNI ČLANKI
Napisali smo več poljudnih člankov, v katerih smo širši slovenski javnosti in posameznim interesnim skupinam, kot so npr. lovci, predstavljali potek projekta, glavne rezultate in ozadje pojavljanja konfliktov med
medvedi in ljudmi:
- KROFEL, M. & K. JERINA. 2009. S pomočjo telemetrije do boljšega razumevanja konfliktov med človekom in medvedom. – Lovec, 92/1: 8-11.
- KROFEL, M. 2009. Mešana medvedja čustva. – Gea, 19/4: 58-63.
- KROFEL, M., JONOZOVIČ, M., MARINČIČ, A. & K. JERINA. 2009. Projekt telemetrije rjavih medvedov. – Lovec, 92/9: 434-436.
- KROFEL, M. 2009. Konflikti z medvedi: izkušnje iz tujine in Slovenije. – Naš časopis, 26. oktober 2009: 40.
- KROFEL, M. & K. JERINA. 2009. Dogodki povezani z medvedom Rožnikom. – Lovec, 92/7-8: 397-398.
- KROFEL, M., MARINČIČ A., JONOZOVIČ, M., JUG, D., FILACORDA, S. & K. JERINA. 2009. V dolino na punce,
potem pa nazaj v gore! Alpski medvedi. – Lovec, 92/12: 607-609.
- KROFEL, M. 2011. Življenje z medvedi. – National Geographic Junior, 78 (april '11): 28-31.
- KROFEL, M. 2011. Les grand carnivores de Slovénie. – La gazette des grand predateurs, n. 39: 21-25.
- KROFEL, M. & K. JERINA. 2011. Projekt telemetrije rjavih medvedov: Obiskovanje krmišč. – Lovec, 93/3: 139-140.
10.7 PREDAVANJA ZA JAVNOST
Po zaključku projekta nameravamo izvesti serijo predavanj za javnost, na katerih bomo lokalni javnosti predstavili rezultate projekte in načine, kako je mogoče zmanjšati konflikte z medvedi. Čim več predavanj bomo poskušali izvesti znotraj osrednjega območja medveda v Sloveniji. Že tekom projekta pa smo vmesne
rezultate projekta javnosti predstavljali v okviru naslednjih predavanj (večinoma v okviru društva Dinaricum):
- 20.11.2008 – Velike zveri v Sloveniji – Ljubljana, Prirodoslovno društvo Slovenije
- 19.2.2009 – Velike zveri v Sloveniji – Ljubljana, DOPPS
- 26.3.2009 – Velike zveri na območju Snežniške planote - Ljubljana, Državni svet
- 6.6.2009 – Velike zveri v Sloveniji – Kočevje, FD Grča
- 16.6.2009 – Velike zveri v Sloveniji – Vipava, gimnazija Vipava
- 18.9.2009 - Velike zveri v Sloveniji: stanje in upravljanje (predavanje za osnovnošolske učitelje biologije, organizacija: Prirodoslovno društvo) – Ljubljana, Tehnološki park Brdo

135
- 18.9.2009 - Osnove biologije velikih zveri (predavanje za osnovnošolske učitelje biologije, organizacija: Prirodoslovno društvo) – Ljubljana, Tehnološki park Brdo
- 10.11.2009 – Velike zveri v Sloveniji; Ljubljana, Ljubljansko geografsko društvo
- 25.11.2009 – Upravljanje in življenje z rjavim medvedom v Sloveniji – Okoljski center, Dinaricum & Grč Vrh
- 20.3.2010 – Pračenje medvjeda na sjevernim Dinaridima – Klana, Hrvaška
- 22.3.2010 – Sobivanje ljudi in velikih zveri – FDV, Ljubljana
- 22.4.2010 – Large carnivores in Slovenia – Mašun
- 28.5.2010 – Large carnivores in Slovenia – Mašun
- 5.10.2010 – Velike zveri v Sloveniji: vedenje in ekologija – Dinaricum, Ljubljana
- 6.10.2010 – Velike zveri v Sloveniji: raziskave in upravljanje – Dinaricum, Ljubljana
- 11.10.2010 – Velike zveri v Sloveniji: konflikti in škode v kmetijstvu– Dinaricum, Ljubljana
- 14.10.2010 – Velike zveri v Sloveniji: sobivanje in odnos ljudi – Dinaricum, Ljubljana
- 30.3.2011 – Velike zveri v Sloveniji – Gimnazija Poljane, Ljubljana
- 20.4.2011 – Large carnivores in Slovenia – FDV, Ljubljana
- 3.6.2011– Large carnivores in Slovenia – Mašun
- 19.6.2011 – Rezultati raziskav velikih zveri v Sloveniji – Črnomelj, Zbor lovcev Zveze lovskih družin Bele Krajine

136

137
11 ZAHVALA
Projekta ne bi bilo mogoče izpeljati brez sodelovanja in nesebične pomoči številnih posameznikov in inštitucij, katerim se na tem mestu iskreno zahvaljujemo. Ključen del projekta je bil odlov medvedov, ki smo
jih nato spremljali s pomočjo telemetrije. Pri tem gre največja zasluga obema vodjema odlovnih ekip: Marku Jonozoviču, dipl. inž. gozd. in Antonu Marinčiču, oba iz Zavoda za gozdove Slovenije. Zahvala gre tudi obema veterinarjema, ki sta bila prisotna ob uspavnanju medvedov: Aljoša Beljan, dr. vet. med. in mag. Tamara
Čehovin, dr. vet. med. Odlavljanje medvedov ne bi bilo tako uspešno, če ne bi krmišč pred odlovom redno zalagali številni člani lovskih družin ter poklicni lovci lovišč s posebnim namenom: LD Begunje, LD Iga vas, LD Kozlek, LD Krka, LD Loka pri Črnomlju, LD Martin Krpan Nova vas, LD Rakitna, LD Ribnica, LD Senožeče, LD
Stari trg Lož, LD Tabor Zagorje, LD Toško Čelo in LD Velike Lašče ter LPN Jelen, LPN Ljubljanski vrh, LPN Medved, LPN Snežnik Kočevska Reka in LPN Žitna gora. Pri kontroliranju pasti so nam pomagali še Josh Smith, Mario Theus in Anamarija Žagar. Zahvaljujemo se tudi vsem, ki so sodelovali pri odlovu medvedov v
okviru prejšnjih projektov, katerih podatke smo prav tako uporabili za analize v okviru te raziskave.
Veliko dela je bilo opravljenega tudi za popisovanje mikrohabitatnih značilnosti medvedjih in naključnih kontralnih lokacij na terenu. Za obežno bazo podatkov gre še posebno veliko zaslug študentom Univerze v Ljubljani in sodelavcem iz Oddelka za gozdarstvo Biotehniške fakultete Univerze v Ljubljani: Danijelu
Borkoviču, Klari Jarni, Ireni Kavčič, Janji Matičič, Nadji Medvešek, Aljažu Rijavcu in Matiji Špacapanu.
Tako obsežnega projekta seveda ne bi bilo možno izpeljati brez financiranja, ki nam jo je zagotovila Agencija Republike Slovenije za okolje v okviru Ministrstva za okolje in prostor (pogodba št.: 2523-08-100547).

138

139
12 VIRI
Adamič, M. 2003. The brown bear in Slovenia: a brief summary of the 20th century population dynamics and future conservation issues. Pages 157-171 in B. Kryštufek, B. Flajšman, and H. I. Griffiths, editors.
Living with bears: A large European carnivore in a shrinking world. Ecological Forum of the Liberal Democracy of Slovenia, Ljubljana.
Breck, S., C. Williams, J. Beckmann, S. Matthews, C. Lackey, and J. Beecham. 2008. Using genetic relatedness to investigate the development of conflict behavior in black bears. Journal of Mammalogy 89:428-434.
Breck, S. W., N. Lance, and V. Seher. 2009. Selective Foraging for Anthropogenic Resources by Black Bears:
Minivans in Yosemite National Park. Journal of Mammalogy 90:1041-1044.
Brosi, G., H. Jenny, R. Schnidrig, T. Briner, P. Molinari, and M. Theus. 2008. Protokoll und Beurteilung der Ereignisse rund um Bär JJ3 Sommer 2007 – Frühling 2008. Amt für Jagd und Fischerei Graubünden, Bundesamt für Umwelt.
Budic, L. 2010. Brown bears in northern and southern Europe: are they seeking food or avoiding dominant
bears? . Norvegian University of Life Sciences, Aas.
Chruszcz, B., A. P. Clevenger, K. E. Gunson, and M. L. Gibeau. 2003. Relationships among grizzly bears, highways, and habitat in the Banff-Bow Valley, Alberta, Canada. Canadian Journal of Zoology 81:1378-1390.
Creachbaum, M. S., C. Johnson, and R. H. Schmidt. 1998. Living on the edge: a process for redesigning
campgrounds in grizzly bear habitat. Landscape and Urban Planning 42:269-286.
Elgmork, K. 1994. The Decline of a Brown Bear Ursus-Arctos L Population in Central South Norway. Biological Conservation 69:123-129.
Floyd, T. 1999. Bear-inflicted human injury and fatality. Wilderness and Environmental Medicine 10:75-87.
Gibeau, M. and S. Stevens. 2005. Grizzly bear response to human use. Page 12 in S. Herrero, editor. Biology, demography, ecology and management of grizzly bears in and around Banff National Park and
Kananaskis Country: The final report of the Eastern Slopes Grizzly Bear Project. University of Calgary, Calgary.
Gniadek, S. J. and K. C. Kendall. 1998. A summary of bear management in Glacier National Park, Montana, 1960-1994. Ursus, Vol 10 - 1998 10:155-159.
Groff, C., D. Dalpiaz, C. Frapporti, R. Rizzoli, and P. Zanghellini. 2010. 2009 Bear Report of the Forestry and
Wildlife Department of the Autonomous Province of Trento. Autonomous Province of Trento, Trento.
Grosse, C., P. Kaczensky, and F. Knauer. 2003. Ants: A food source sought by Slovenian brown bears (Ursus arctos)? Canadian Journal of Zoology-Revue Canadienne De Zoologie 81:1996-2005.
Gunther, K. A., M. A. Haroldson, K. Frey, S. L. Cain, J. Copeland, and C. C. Schwartz. 2004. Grizzly bear-human conflicts in the Greater Yellowstone ecosystem, 1992-2000. Ursus 15:10-22.

140
Gunther, K. A. and H. E. Hoekstra. 1998. Bear-inflicted human injuries in Yellowstone National Park, 1970-1994. Ursus, Vol 10 - 1998 10:377-384.
Guthlin, D., F. Knauer, T. Kneib, H. Kuchenhoff, P. Kaczensky, G. Rauer, M. Jonozovic, A. Mustoni, and K. Jerina. 2011. Estimating habitat suitability and potential population size for brown bears in the
Eastern Alps. Biological Conservation 144:1733-1741.
Herrero, S. 2002. Bear attacks: Their causes and avoidance. 2 edition. Nick Lyons Books, New York.
Herrero, S. and A. Higgins. 1999. Human injuries inflicted by bears in British Columbia: 1960–97. Ursus 11:209-218.
Herrero, S. and A. Higgins. 2003. Human injuries inflicted by bears in Alberta: 1960-98. Ursus 14:44-54.
Herrero, S., T. Smith, T. D. DeBruyn, K. Gunther, and C. A. Matt. 2005. From the field: Brown bear habituation to people - safety, risks, and benefits. Wildlife Society Bulletin 33:362-373.
Horstman, L. P. and J. R. Gunson. 1982. Black bear predation on livestock in Alberta. Wildlife Society Bulletin
10:34-39.
Huber, D. 2010. Rehabilitation and reintroduction of captive-reared bears: feasibility and methodology for European brown bears Ursus arctos. International ZOO Yearbook 44:47-54.
Jope, K. L. 1985. Implications of Grizzly Bear Habituation to Hikers. Wildlife Society Bulletin 13:32-37.
Kaczensky, P. 1999. Large Carnivore Depredation on Livestock in Europe. Ursus 11:59-72.
Kaczensky, P., M. Blazic, and H. Gossow. 2004. Public attitudes towards brown bears (Ursus arctos) in Slovenia. Biological Conservation 118:661-674.
Kaczensky, P., K. Jerina, M. Jonozovic, M. Krofel, T. Skrbinsek, G. Rauer, I. Kos, and B. Gutleb. 2011. Illegal
killings may hamper brown bear recovery in the Eastern Alps. Ursus 22:37-46.
Knight, R. R., B. M. Blanchard, and L. L. Eberhardt. 1988. Mortality patterns and population sinks for Yellowstone grizzly bears, 1973-1985. Wildlife Society Bulletin 16:121-125.
Knight, R. R. and S. L. Judd. 1983. Grizzly Bears That Kill Livestock. International Conf. Bear Res. and Manage. 5:186-190.
Krofel, M., N. Pagon, P. Zor, and I. Kos. 2008. Analiza medvedov odvzetih iz narave in genetsko-molekularne
raziskave populacije medveda v Sloveniji. 2. del: Analiza vsebine prebavil medvedov (Ursus arctos L.) odvzetih iz narave v Sloveniji v letih 2006 – 2008. Oddelek za biologijio, Biotehniška fakulteta, Univerza v Ljubljani, Ljubljana, Ljubljana.
LIFE02NAT/SLO/8585, P. 2005. Nature project n° LIFE02NAT/SLO/8585 "Conservation of large carnivores in
Slovenia - Phase I (Ursus arctos)". Akcija A4. Načrt ukrepov za zmanjšanje konfliktov s kmetijstvom. Biotehniška fakulteta, Oddelek za gozdarstvo in obnovljive gozdne vire, Ljubljana.
Linnell, J. D. C., J. Odden, M. E. Smith, R. Aanes, and J. E. Swenson. 1999. Large carnivores that kill livestock: do "problem individuals" really exist? Wildlife Society Bulletin 27:698-705.

141
Marenče, M., M. Jonozovič, M. Adamič, and K. Jerina. 2007. Spatial analysis of damages caused by brown bear (Ursus arctos), bear harvest and bear population density in Slovenia: how efficient is the current policy of bear management? . International Symposium On Large Carnivores And Agriculture: Comparing
Experiences Across Italy And Europe, 9–10 March 2007, , Assisi, Italy.
Martin, J., M. Basille, B. van Moorter, J. Kindberg, D. Allainé, and J. E. Swenson. 2010. Coping with human disturbance: spatial and temporal tactics of brown bear. Canadian Journal of Zoology 88:875-883.
Mattson, D. J., B. M. Blanchard, and R. R. Knight. 1992. Yellowstone Grizzly Bear Mortality, Human Habituation, and Whitebark-Pine Seed Crops. Journal of Wildlife Management 56:432-442.
McLellan, B. N., F. W. Hovey, R. D. Mace, J. G. Woods, D. W. Carney, M. L. Gibeau, W. L. Wakkinen, and W. F.
Kasworm. 1999. Rates and causes of grizzly bear mortality in the interior mountains of British Columbia, Alberta, Montana, Washington, and Idaho. Journal of Wildlife Management 63:911-920.
Meagher, M. and J. R. Phillips. 1983. Restoration of natural populations of grizzly and black bears in Yellowstone National Park. Int. Conf. Bear Res. and Manage. 5:152-158.
Muhly, T., C. C. Gates, C. Callaghan, and M. Musiani. 2010. Livestock husbandry practices reduce wolf
depredation risk in Alberta, Canada. Pages 261-286 in M. Musiani, L. Boitani, and P. C. Paquet, editors. The world of wolves: new perspectives on ecology, behaviour and management. University of Calgary Press, Calgary.
Partridge, S. T., D. L. Nolte, G. J. Ziegltrum, and C. T. Robbins. 2001. Impacts of supplemental feeding on the
nutritional ecology of black bears. Journal of Wildlife Management 65:191-199.
Ralf, R. 1995. History of Bear\Human Conflict Management in Jasper National Park: 1907-1995. Jasper National Park.
Rodriguez, C., J. Naves, A. Fernandez-Gil, and M. Delibes. 2006. Brown bear food habits at the border of its range: A long-term study. Journal of Mammalogy 87:899-908.
Sato, Y., T. Mano, and S. Takatsuki. 2005. Stomach contents of brown bears Ursus arctos in Hokkaido, Japan.
Wildlife Biology 11:133-144.
Schirokauer, D. W. and H. M. Boyd. 1998. Bear-human conflict management in Denali National Park and Preserve, 1982-94. Ursus, Vol 10 - 1998 10:395-403.
Serban-Parau, N. 1999. Brown bear-man conflicts at the garbage deposits in Prahova valley and Brasov, Romania. Page 40 12th International Conference on Bear Research and Management, Poiana Brasov,
Romania.
Sertić, J. 2008. Štete od smeđeg medvjeda (Ursus arctos l.) u šumskim sastojinama na podrucju državnog lovišta broj viii/2 „Bjelolasica“. Diploma thesis. University of Zagreb, Zagreb.
Simonič, A. 1998. Srečanja z medvedom. Ministrstvo za kmetijstvo, gozdarstvo in prehrano, Ljubljana.
Smith, T. S., S. Herrero, and T. D. DeBruyn. 2005. Alaskan brown bears, humans, and habituation. Ursus 16:1-10.

142
Swenson, J. E., B. Dahle, H. Busk, O. Opseth, T. Johansen, A. Soderberg, K. Wallin, and G. Cederlund. 2007. Predation on moose calves by European brown bears. Journal of Wildlife Management 71:1993-1997.
Swenson, J. E., F. Sandegren, A. Söderberg, M. Heim, O. J. Sørensen, A. Bjärvall, R. Franzén, S. Wikan, and P. Wabakken. 1999. Interactions between brown bears and humans in Scandinavia. Biosphere
Conservation 2:1-9.
Team, A. B. E. 2006. JJ1 „Bruno“ in Austria and Germany 2006: Protocol and Risk Assessment. Austrian Bear Emergency Team, Vienna.
Treves, A. and K. U. Karanth. 2003. Human-carnivore conflict and perspectives on carnivore management worldwide. Conservation Biology 17:1491-1499.
Treves, A. and L. Naughton-Treves. 2005. Evaluating lethal control in the management of human-wildlife
conflict. . Pages 86-106 in R. Woodroffe, S. Thirgood, and A. Rabinowitz, editors. People and Wildlife, Conflict or Coexistence? Cambridge University Press, Cambridge, UK.
Ulamec, P. 2008. Analiza odškodninskih zahtevkov za škodo, ki so jo povzročile živali zavarovanih prosto živečih živalskih vrst v letu 2007. Agencija RS za okolje, Ljubljana.
Vaughan, M. R., P. F. Scanlon, S. E. Mersmann, and D. D. Martin. 1989. Black bear damage in Virginia.
Proceedings of the Eastern Wildlife Damage Control Conference 4:147-154.
Wilder, J. M., T. D. DeBruyn, T. S. Smith, and A. Southwould. 2007. Systematic collection of bear-human interaction information for Alaska's national parks. Ursus 18:209-216.
Wilson, S. M. 2007. Community-supported conservation of grizzly bears on private agricultural lands. Final close-out report for conservation innovation grant. U.S. Department of Agriculture – Natural Resources
Conservation Service, Portland, OR.
Wilson, S. M., M. J. Madel, D. J. Mattson, J. M. Graham, and T. Merrill. 2006. Landscape conditions predisposing grizzly bears to conflicts on private agricultural lands in the western USA. Biological Conservation 130:47-59.