Embed Size (px)
Citation preview

Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Copyright © 2010 American Association for Cancer Research
1
Preclinical Evaluation of AMG 900, a Novel Potent and Highly Selective Pan-Aurora Kinase Inhibitor with Activity in Taxane-Resistant Tumor Cell Lines Marc Payton1, 5, Tammy L. Bush1, Grace Chung1, Beth Ziegler1, Patrick Eden1, Patricia McElroy2,
Sandra Ross2, Victor J. Cee3, Holly L. Deak3, Brian L. Hodous3, Hanh Nho Nguyen3, Philip R.
Olivieri3, Karina Romero3, Laurie B. Schenkel3, Annette Bak4, Mary Stanton4, Isabelle Dussault1,
Vinod F. Patel3, Stephanie Geuns-Meyer3, Robert Radinsky1, and Richard L. Kendall1
Departments of 1Oncology Research, 2Hematology Research, 3Medicinal Chemistry, and
4Pharmaceutics, Amgen Inc., Thousand Oaks, California 91320 or Cambridge, Massachusetts 02142
5To whom correspondence should be addressed:
Amgen Inc., One Amgen Center Drive, Mailstop 14-1-B, Thousand Oaks, CA 91320-1789.
Phone: (805) 447-6697, Fax: (805) 375-8368
Email: [email protected]
Running title: AMG 900, a Novel Pan-Aurora Kinase Inhibitor
Keywords: AMG 900, aurora kinase, multidrug resistance, P-gp, antimitotic
Published OnlineFirst on October 8, 2010 as 10.1158/0008-5472.CAN-10-3001
Research. on February 3, 2019. © 2010 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 8, 2010; DOI: 10.1158/0008-5472.CAN-10-3001

Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Copyright © 2010 American Association for Cancer Research
2
Abstract
In mammalian cells, the aurora kinases (aurora-A, -B, and -C) play essential roles in regulating cell
division. Aurora-A and -B expression is elevated in a variety of human cancers and is associated with
high proliferation rates and poor prognosis, making them attractive targets for anticancer therapy. AMG
900 is an orally bioavailable, potent and highly selective pan-aurora kinase inhibitor that is active in
taxane-resistant tumor cell lines. In tumor cells, AMG 900 inhibited autophosphorylation of aurora-A and -
B as well as phosphorylation of histone H3 on Ser10, a proximal substrate of aurora-B. The predominant
cellular response of tumor cells to AMG 900 treatment was aborted cell division without a prolonged
mitotic arrest, which ultimately resulted in cell death. AMG 900 inhibited the proliferation of 26 tumor cell
lines at low nanomolar concentrations, including cell lines resistant to the antimitotic drug paclitaxel and to
other aurora kinase inhibitors (AZD1152, MK-0457, and PHA-739358). Furthermore, AMG 900 was
active in an AZD1152-resistant HCT116 variant cell line that harbors an aurora-B mutation (W221L). Oral
administration of AMG 900 blocked the phosphorylation of histone H3 in a dose-dependent manner and
significantly inhibited the growth of HCT116 tumor xenografts. Importantly, AMG 900 was broadly active
in multiple xenograft models representing five tumor types, including three multidrug-resistant xenograft
models. AMG 900 has entered clinical evaluation in adult patients with advanced cancers and has the
potential to treat tumors refractory to anticancer drugs such as the taxanes.
Research. on February 3, 2019. © 2010 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 8, 2010; DOI: 10.1158/0008-5472.CAN-10-3001

Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Copyright © 2010 American Association for Cancer Research
3
Introduction
Somatic cell division is a complex and highly coordinated process that ensures faithful
segregation of duplicated chromosomes into two daughter cells. Deregulation of the cell cycle is a
hallmark of cancer, characterized by uncontrolled proliferation and defects in chromosome segregation.
Antimitotic drugs that block tumor cell division are a proven intervention strategy in the treatment of
cancer. However, the clinical benefits of classical antimitotic drugs may be hampered by development of
multidrug resistance (MDR) and collateral damage to non-dividing cells including for example, peripheral
neuropathy (1). Aurora kinases are essential mitotic regulators and their potential role in tumorigenesis
makes them attractive targets for anticancer therapy (2-4).
In mammalian cells, the aurora family of serine/threonine protein kinases is comprised of three
paralogous genes (aurora-A, -B, and -C). Aurora-A and -B are essential regulators of mitotic entry and
progression, whereas aurora-C function is primarily restricted to male meiosis during spermatogenesis (5-
10). Aurora-A can function as an oncogene and is amplified in a subset of human tumors. Aurora-A and
-B expression is frequently elevated in human cancers and is associated with advanced clinical staging
(11).
The mitotic checkpoint, also referred to as the spindle assembly checkpoint (SAC), is a
surveillance mechanism responsible for controlling proper alignment, microtubule-kinetochore
attachments, and segregation of duplicated chromosomes (12). In tumor cells, genetic depletion or
pharmacological inhibition of aurora-A results in abnormal spindle formation and SAC activation. In
constrast, depletion or inhibition of aurora-B inactivates the SAC, resulting in aborted cell division without
a mitotic arrest. Importantly, dual suppression of aurora-A and -B phenocopies the effects of inhibiting
aurora-B alone (13, 14). The silencing of the SAC leads to an accumulation of tumor cells that contain 4N
DNA content in the G1-phase of the cell cycle. Continued suppression of aurora-B activity leads to further
rounds of genome replication without division, a process referred to as endoreduplication, which
ultimately results in tumor cell death (15, 16). This mechanism of action is distinct from that of
microtubule-binding antimitotic drugs (e.g. taxanes, vinca alkaloids, and epothilones) in that death of
Research. on February 3, 2019. © 2010 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 8, 2010; DOI: 10.1158/0008-5472.CAN-10-3001

Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Copyright © 2010 American Association for Cancer Research
4
tumor cells is primarily driven by continued cell-cycle progression rather than by SAC activation and
prolonged cell arrest in mitosis (16, 17).
Drug resistance is a major problem limiting the efficacy of many current anticancer therapies.
The underlying mechanisms of clinical resistance to microtubule-binding agents are multifactorial and not
fully understood (18). In cultured tumor cells, two prominent mechanisms of resistance to the taxanes are
overexpression of drug efflux pumps and tubulin modifications (19-21). One strategy to overcome
susceptibility to the effects of MDR would be to design a novel antimitotic drug candidate whose activity is
not influenced by drug efflux, mediated by ATP-binding cassette (ABC) transporters such as P-
glycoprotein/P-gp (ABCB1) and BCRP (ABCG2) (22). Moreover, a small molecule inhibitor that is
equipotent against two essential mitotic kinases may reduce the potential for resistance driven by target-
modifying mutations (23, 24).
Currently, a variety of ATP-competitive inhibitors that target one or more of the aurora kinases
and that have varying degrees of kinase specificity are in early clinical development (25). This report
describes the preclinical activities of AMG 900, an orally bioavailable, potent and selective pan-aurora
kinase inhibitor with activity in MDR tumor cell lines. In contrast to paclitaxel and three well-characterized
aurora kinase inhibitors (AZD1152, MK-0457, and PHA-739358), AMG 900 showed uniform potency
across tumor cell lines, including P-gp and BCRP expressing cell lines. Furthermore, AMG 900 was
active in an HCT116 cell line adapted to grow in the presence of AZD1152. This HCT116 variant cell line
carries a missense mutation in one allele of the aurora-B gene resulting in an amino acid substitution
(W221L) in its activation loop. In vivo, AMG 900 blocks the phosphorylation of histone H3, a proximal
substrate of aurora-B (26) and inhibits the growth of multiple tumor xenografts, including three MDR
xenograft models resistant to paclitaxel or docetaxel. Our data provides compelling evidence that AMG
900 may be active in tumors resistant to taxanes and three other well-characterized inhibitors that target
aurora-B. AMG 900 is presently under clinical evaluation in adult patients with advanced cancers.
Materials and Methods
Research. on February 3, 2019. © 2010 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 8, 2010; DOI: 10.1158/0008-5472.CAN-10-3001

Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Copyright © 2010 American Association for Cancer Research
5
Chemistry. AMG 900 N-(4-((3-(2-amino-4-pyrimidinyl)-2-pyridinyl)oxy)phenyl)-4-(4-methyl-2-thienyl)-1-phthalazinamine) was
synthesized at Amgen (WO 2007087276). The molecular structures have been disclosed for the following compounds: paclitaxel
and docetaxel (18), MLN8054 (25), MK-0457 (4), AZD1152 (25), and PHA-739358 (25).
Cell lines. Tumor cell lines were obtained from the American Type Culture Collection (ATCC) unless otherwise specified. Cell lines
were grown under recommended conditions. The HCT116 p21+ and p21- cell lines were obtained from Johns Hopkins University
under a licensing agreement. The CAL51 cell line was obtained from Deutsche Sammlung von Mikroorganismen und Zellkulturen
(DSMZ, Germany). The MCF-7 p53+ and p53- cell lines were generated as previously described (27). The MDA-MB-231-PTX and
NCI-H460-PTX cell lines were established by growing the cells in the presence of increasing concentrations of paclitaxel over a
period of 6 months. The HCT116 AZD1152-resistant cell line was established by growing the cells in the presence of AZD1152 at
80 nmol/L. The histone H2B-GFP (BD Biosciences) HeLa cell line was established at Amgen.
Animals. All experimental procedures were conducted in accordance with Institutional Animal Care and Use Committee and U.S.
Department of Agriculture regulations. Four to six-week-old female athymic nude mice (Harlan Sprague Dawley) were housed in
sterilized cages and maintained under aseptic conditions. Eight-week-old female BDF1 mice (Charles River Laboratories) were
housed five per filter-capped cage under pathogen-free conditions. The laboratory housing the animals provided alternating light
and dark cycles (12 hours each) and met the standards of the Association for Assessment and Accreditation of Laboratory Animal
Care specifications. Food, water, and nutritional supplements were offered ad libitum. All drugs were administered based on the
individual body weight of each mouse.
Western blot analysis. Cell lysates were processed for Western blot analysis as previously described (27). Immunoblots were
probed with the following antibodies: anti-aurora-A, anti-aurora-B (BD Biosciences), anti-p-aurora-A Thr288, anti-p-aurora-B Thr232,
anti-cleaved caspase-7 Asp198, and anti-total p53 (Cell Signaling), anti-β-actin (Sigma), and anti-p21Cip1 (Santa Cruz). Next, the
immunoblots were probed with either anti-rabbit or anti-mouse IgG (Vectastain kit, Vector Laboratories) and the protein bands were
detected using the Lightning-Enhanced Chemiluminescence Kit from PerkinElmer.
Details regarding enzyme assays, nocodazole block, cell imaging assays, colony formation assays, flow cytometry assays, aurora
kinase gene analysis, pharmacodynamic assays, tumor xenograft models, neutrophil assessment study, and statistical analyses are
described in the Supplementary Materials and Methods.
Results
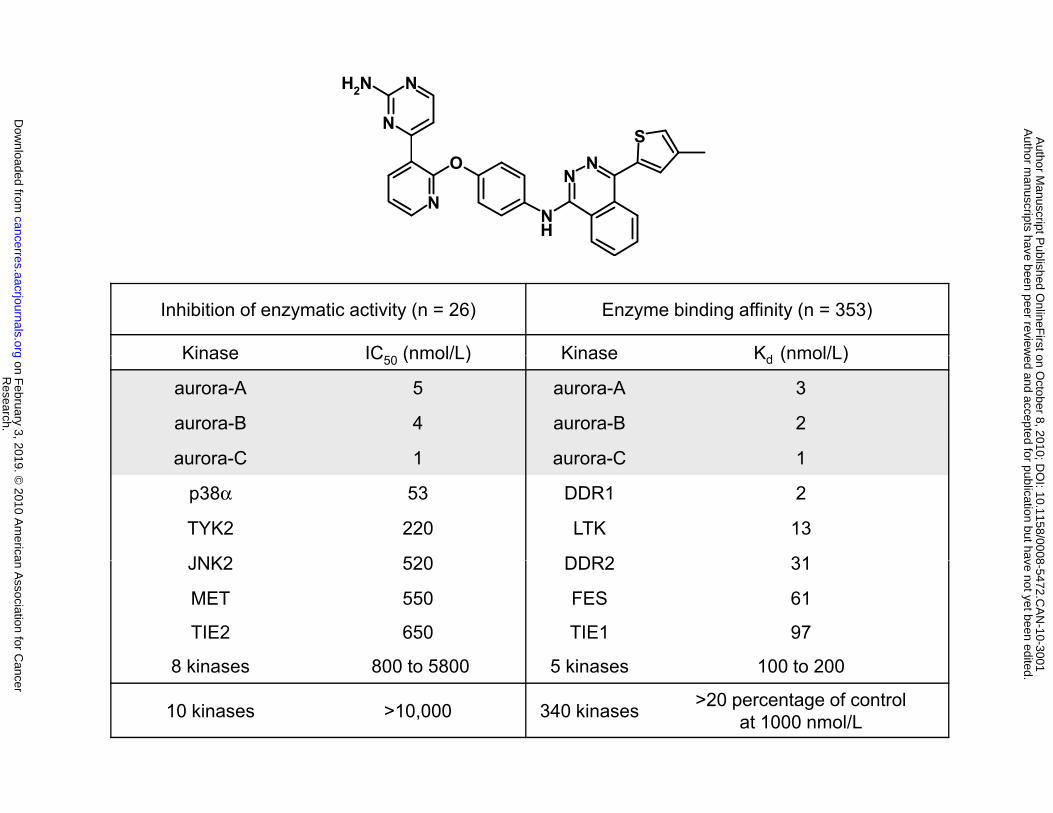
AMG 900 is a potent and highly selective pan-aurora kinase inhibitor. The discovery of a
novel class of ATP-competitive phthalazinamine small molecule inhibitors of aurora kinases led to the
identification of AMG 900 (S. Geuns-Meyer, manuscript in preparation). AMG 900 inhibits the enzyme
activity of all three aurora kinase family members with IC50 values ≤ 5 nmol/L (Fig. 1A). To determine the
Research. on February 3, 2019. © 2010 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 8, 2010; DOI: 10.1158/0008-5472.CAN-10-3001

Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Copyright © 2010 American Association for Cancer Research
6
specificity of AMG 900 across the kinome, a panel of 26 kinases was screened, and only p38α and TYK2
enzymes were inhibited (> 50%) at concentrations less than 500 nmol/L (Fig. 1A). Further screening was
performed against a panel of 353 distinct kinases using an ATP-site dependent competition binding assay
(28). AMG 900 exhibited low nanomolar binding affinity for aurora kinases as well as interactions (Kd < 50
nmol/L) with DDR1, DDR2, and LTK receptor tyrosine kinases (Fig. 1A).
To investigate the inhibition of aurora kinase activity in cells by AMG 900, the levels of
phosphorylated aurora-A Thr288, aurora-B Thr232, and histone H3 Ser10 (p-histone H3) were determined
using Western blotting and quantitative cell imaging techniques. In HeLa cells, AMG 900 inhibited
autophosphorylation of aurora-A and -B in a concentration-dependent manner (Fig. 1B). MLN8054
(aurora-A inhibitor) and AZD1152 (aurora-B inhibitor) selectively blocked the autophosphorylation of
either aurora-A Thr288 or aurora-B Thr232, respectively. In the cell imaging assay, HeLa cells treated with
AMG 900 for 6 hours showed a concentration-dependent decrease in p-aurora-A Thr288 and p-histone H3
immunofluorescence staining with IC50 values of 6.5 and 8.2 nmol/L, respectively (Fig. 1C). Next, the
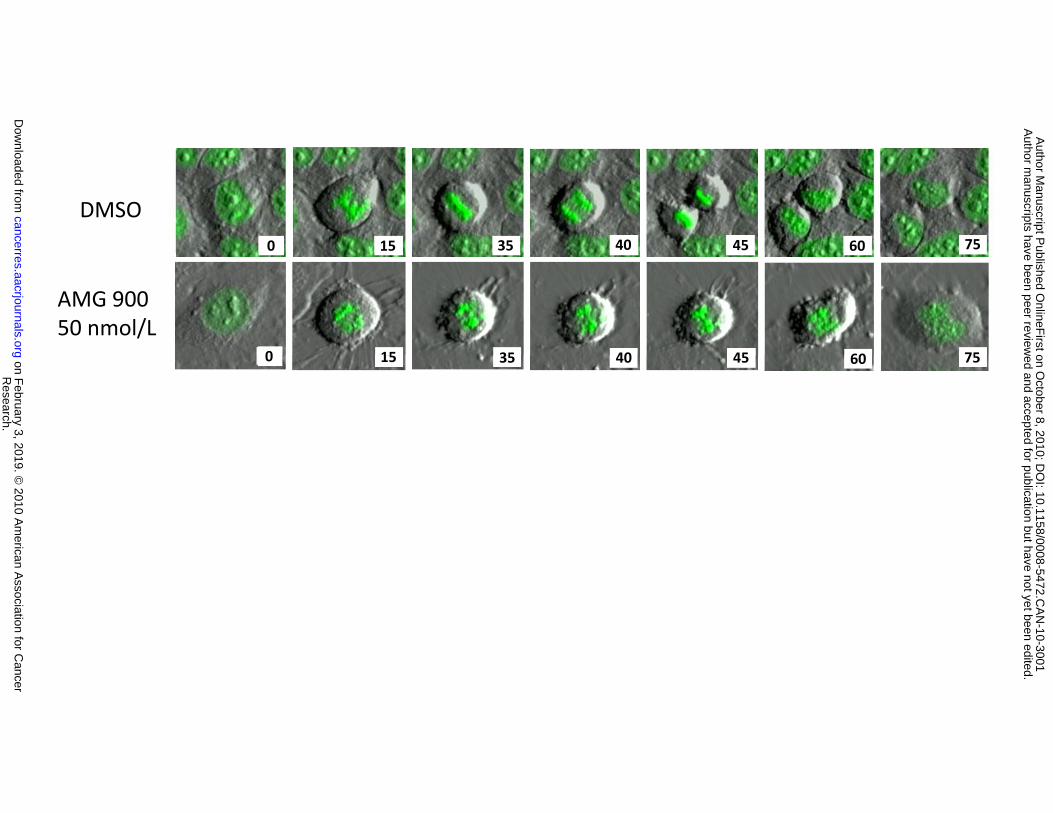
effects of AMG 900 on chromosome dynamics and cell-cycle progression were examined by live-cell
imaging of HeLa cells that express a histone H2B-GFP fusion protein. The top image panel of Figure 1D
shows a representative DMSO-treated cell entering and exiting mitosis to yield two daughter cells. In
contrast, a representative AMG 900-treated cell entering mitosis failed to properly congress
chromosomes to the metaphase plate, which subsequently resulted in aborted cell division (bottom panel
of Fig. 1D). These data demonstrate that AMG 900 does not induce a prolonged cell arrest in mitosis,
consistent with inhibition of aurora-B and SAC silencing.
Effects of AMG 900 on proliferating cells are consistent with inhibition of aurora-B activity.
Suppression of aurora-B activity in mitosis is known to induce polyploidy in tumor cells as a consequence
of cytokinesis failure. To confirm this, we first determined the concentration of AMG 900 that completely
suppressed p-histone H3 in HCT116 cells (Fig. 2A). Treatment of HCT116 cells with 50 nmol/L of AMG
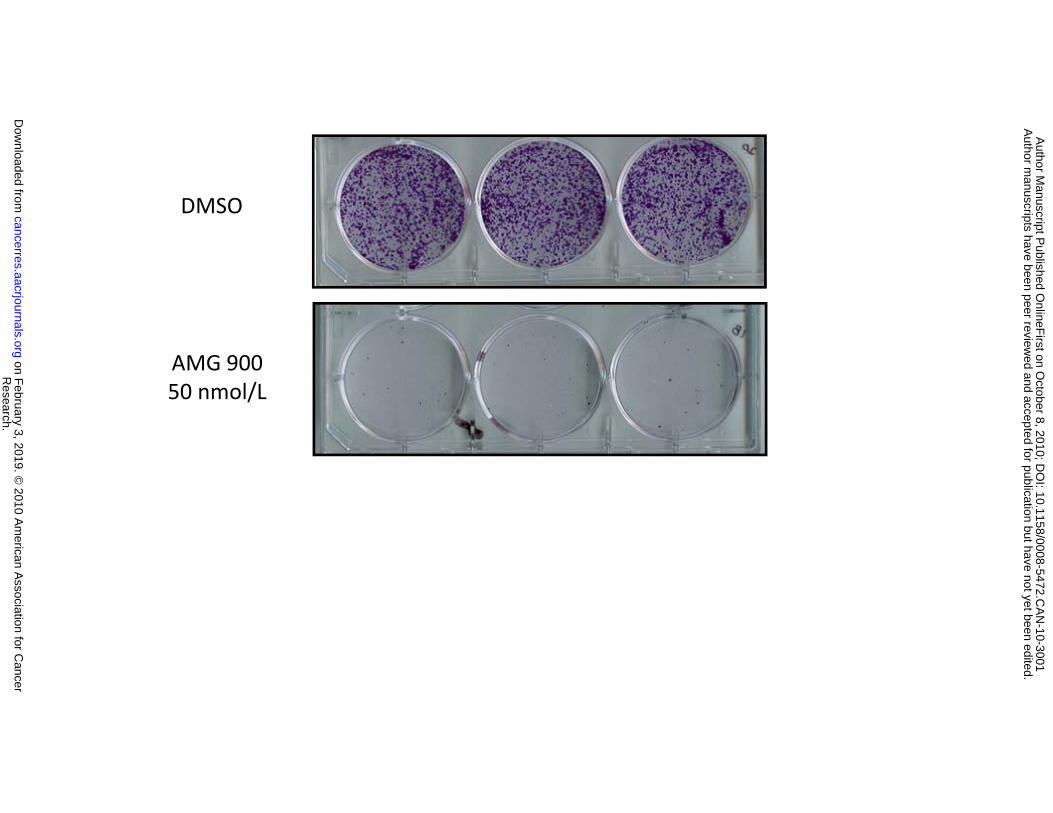
900 for 48 hours resulted in polyploidy and suppressed the formation of colonies after cell replating (Fig.
2B and 2C). Several reports have established that inhibition of aurora-B activates the post-mitotic G1-
checkpoint mediated by the p53 tumor suppressor protein (29, 30). Consistent with these findings, AMG
Research. on February 3, 2019. © 2010 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 8, 2010; DOI: 10.1158/0008-5472.CAN-10-3001

Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Copyright © 2010 American Association for Cancer Research
7
900 induced a significant increase in p53 and p21Cip1 proteins that correlated with a corresponding
decrease in 5-bromo-2'-deoxyuridine (BrdUrd) incorporation, a direct measure of DNA synthesis
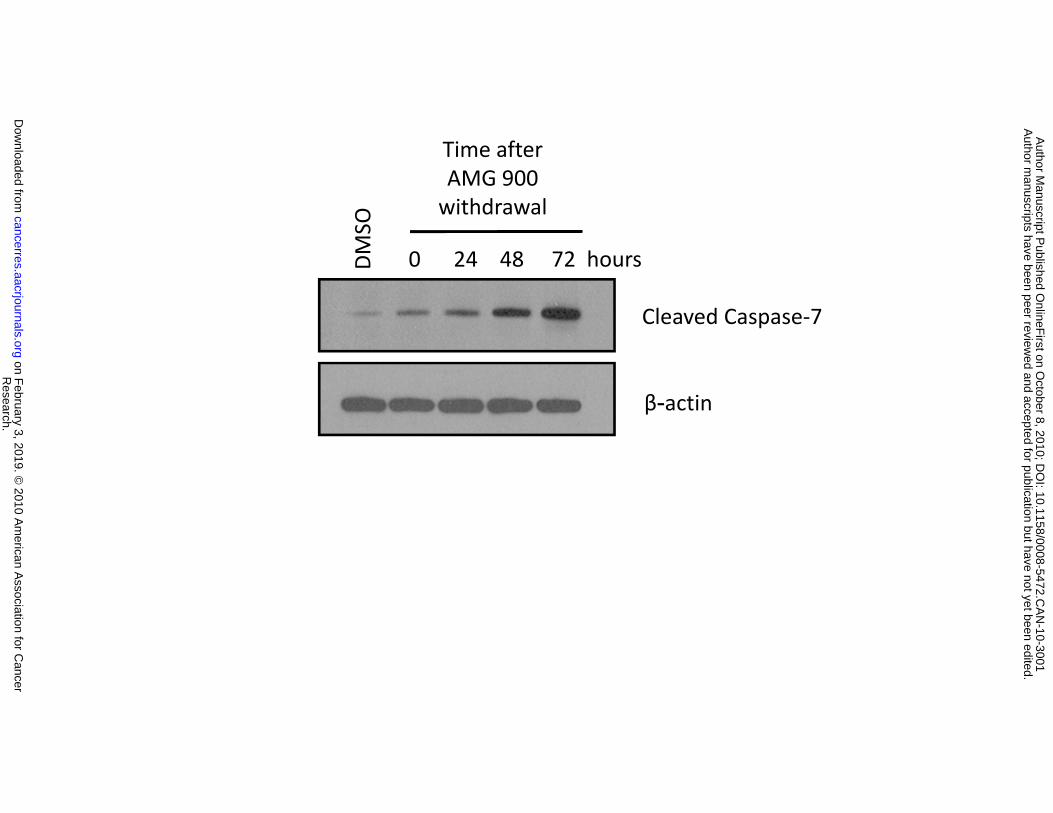
(Supplementary Fig. S1). To assess the kinetics of cell death induction by AMG 900, HCT116 cells were
treated with compound for 48 hours, washed twice, and then cultured in complete media lacking
compound for 0, 24, 48, and 72 hours. Treatment with AMG 900 induced a time-dependent increase in
apoptosis measured by the induction of cleaved caspase-7 (Fig. 2D). Other mechanisms of cell death
(e.g. mitotic catastrophe, multipolar cell division, and giant-cell necrosis) may also contribute to the loss of
tumor cell viability after treatment with AMG 900 (unpublished observations, 31). Consistent with the
highly selective profile of AMG 900, Jurkat cells treated with compound for 48 hours over a 5000-fold
concentration range (0.0012 to 5 μmol/L) maintained a stable polyploidy phenotype, indicating that the
dominant phenotype of AMG 900 inhibition is through inhibition of aurora-B (data not shown). AMG 900
had no obvious toxic effects on non-cycling human foreskin fibroblast cells up to a concentration of 25
μmol/L; however it did inhibit cell-cycle progression (without endoreduplication) and induced cell death in
proliferating human bone marrow mononuclear cells at low nanomolar concentrations as expected with
an on-mechanism effect of this agent (data not shown).
AMG 900 blocks the proliferation of multiple human tumor cell lines, including cell lines
resistant to paclitaxel, AZD1152, MK-0457, and PHA-739358. To investigate the antiproliferative
effects of AMG 900, a panel of 26 cell lines from diverse tumor types were treated with compound for 24
hours, washed twice in complete media lacking AMG 900, and then cultured for an additional 48 hours.
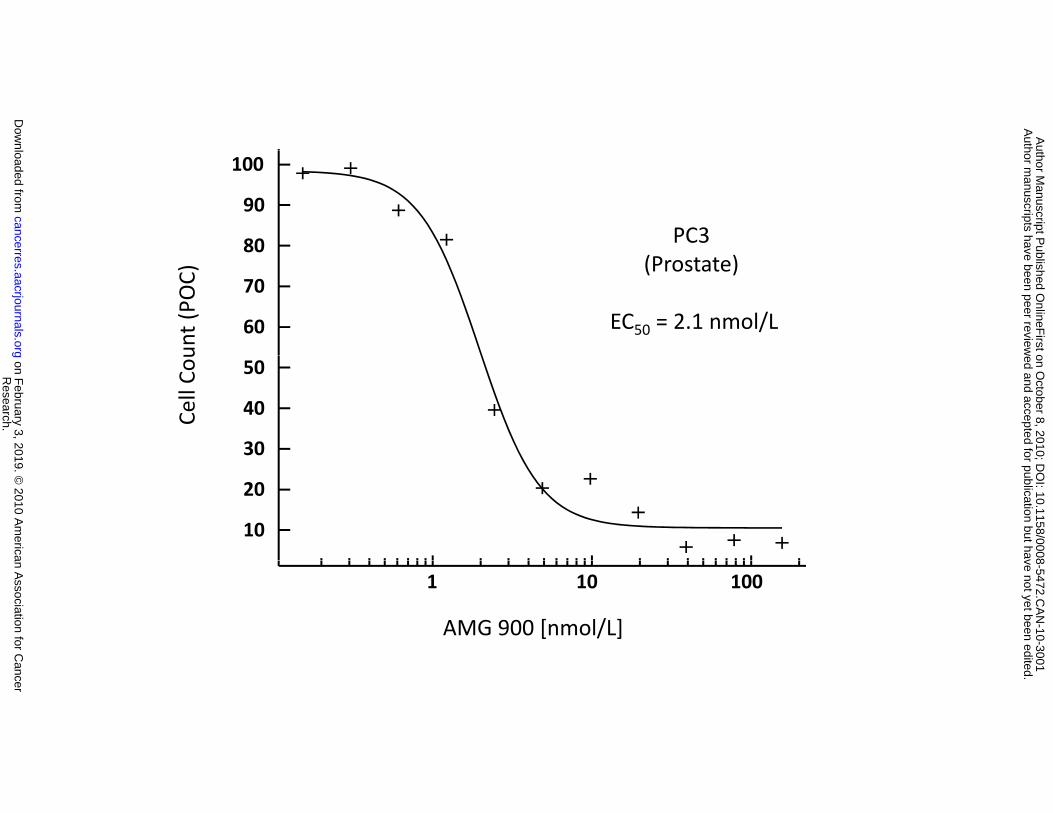
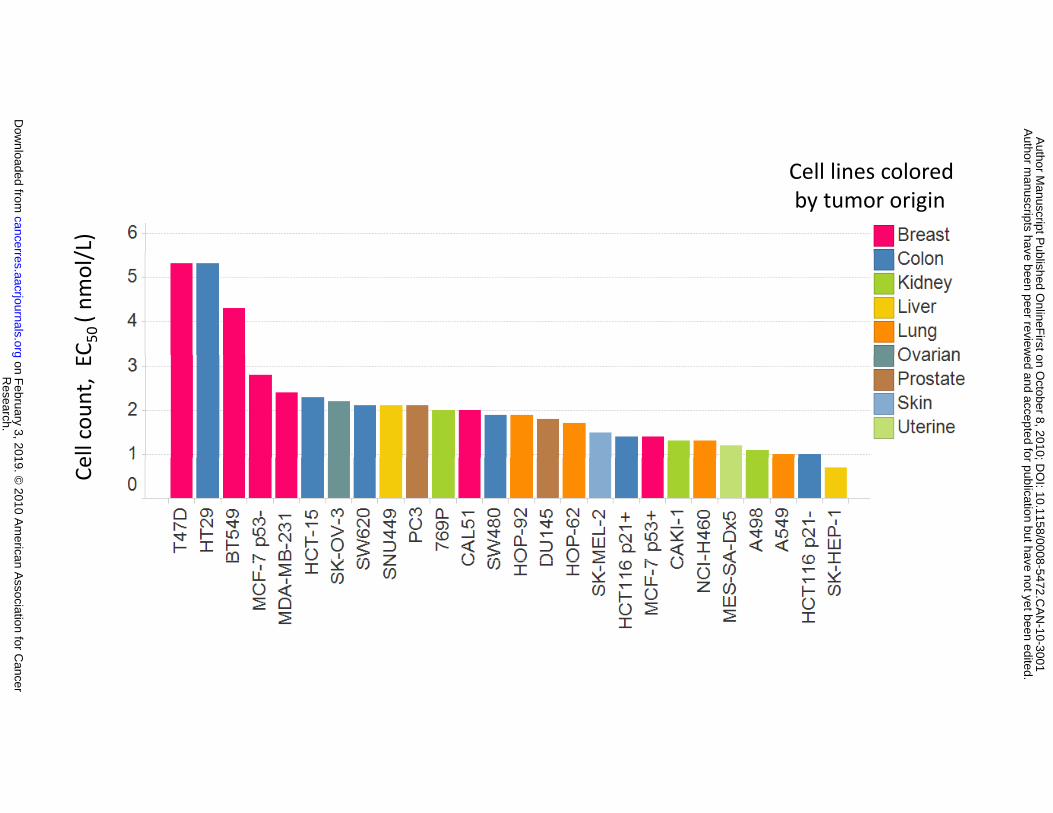
Concentration-response curves and corresponding cell count EC50 values were established for each cell
line (Fig. 3A and 3B). The results show that AMG 900 inhibited cell proliferation with EC50 values ranging
from 0.7 to 5.3 nmol/L. Importantly, four of these AMG 900-sensitive cell lines (HCT-15, MES-SA-Dx5,
769P, and SNU449) are resistant to paclitaxel and other anticancer agents (32-34). To further investigate
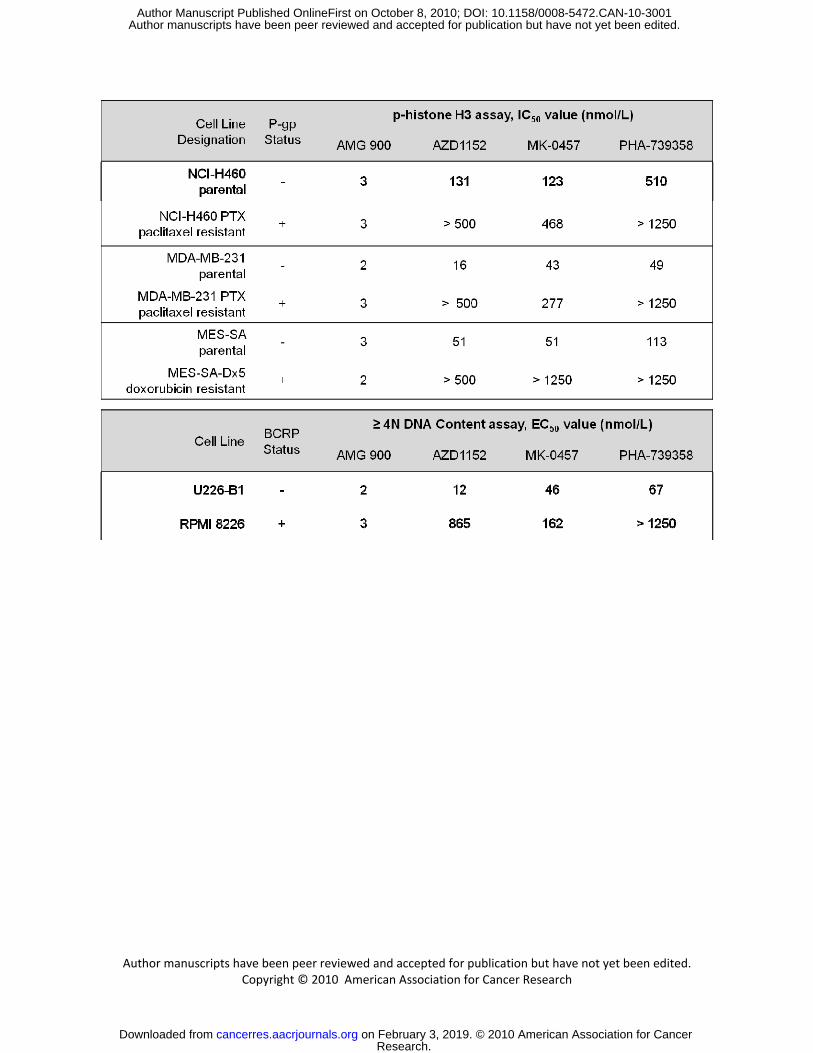
the activity of AMG 900 in MDR cells, four tumor cell lines expressing P-gp, were compared with three
non-P-gp expressing parental cell lines. As shown in Figure 3C, in a colony formation assay, there was a
profound decrease in the number of detectable colonies in all of the cell lines treated with AMG 900 at 5
or 50 nmol/L. By contrast, paclitaxel failed to inhibit colony formation in all four of the P-gp expressing
Research. on February 3, 2019. © 2010 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 8, 2010; DOI: 10.1158/0008-5472.CAN-10-3001

Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Copyright © 2010 American Association for Cancer Research
8
tumor cell lines at concentrations effective in the parental cell lines. Intrigued by this potentially important
finding, we sought to understand if this was a common property of aurora kinase inhibitors. We therefore
evaluated three well-characterized aurora kinase inhibitors (AZD1152, MK-0457, and PHA-739358) in a
subset of MDR tumor cell lines expressing either P-gp or BCRP drug efflux transporters. Interestingly,
AMG 900 inhibited p-histone H3 or induced polyploidy across all the cell lines tested irrespective of P-gp
or BCRP status with uniform potency (IC50 or EC50 values ranging from 2 to 3 nmol/L). By contrast, the
other aurora kinase inhibitors were less potent in one or more of the MDR cell lines compared with the
matched sensitive tumor cell lines (Table 1). Inhibition of P-gp and BCRP drug pumps with GF120918
(21) sensitized HCT-15 and RPMI-8226 cells to AZD1152, suggesting that the loss of potency in the MDR
cell lines was due to drug efflux (data not shown).
To investigate alternative mechanisms of resistance to aurora kinase inhibitors (35), HCT116
cells were adapted to grow in the presence of AZD1152. The activity of AMG 900 was then evaluated in
the HCT116 parental and AZD1152-resistant cell lines. The cellular ≥ 4N DNA content EC50 values for
AMG 900 were 2 and 5 nmol/L compared to 34 and 672 nmol/L for AZD1152, respectively
(Supplementary Fig. S2A). AMG 900 inhibited the colony formation in both HCT116 cell lines at
concentrations ≥ 5 nmol/L, whereas the variant subline was insensitive to AZD1152 at 50 nmol/L
(Supplementary Fig. S2B). Both of the HCT116 cell lines were equally sensitive to paclitaxel and were
negative for P-gp and BCRP expression (Supplementary Fig. S2C). Interestingly, the HCT116 variant
subline harbors a missense mutation in one allele of the aurora-B gene (TGG TTG; W221L), whereas
no mutations were detected in the aurora-A and -C genes. These results suggest that AMG 900
maintains activity in tumor cells carrying a heterozygous mutation in aurora-B that may be responsible for
resistance to AZD1152. Somatic point mutations at the analogous residue adjacent to the invariant DFG
motif have been found in epidermal growth factor receptor and BRAF genes (L858R and L596R,
respectively) in primary human tumors (36, 37). Studies are underway to more fully characterize the
biological effects of this aurora-B mutation.
AMG 900 inhibits the phosphorylation of histone H3 and suppresses the growth of human
tumor xenografts in vivo. To confirm that AMG 900 inhibits aurora-B activity in vivo, mice bearing
Research. on February 3, 2019. © 2010 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 8, 2010; DOI: 10.1158/0008-5472.CAN-10-3001

Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Copyright © 2010 American Association for Cancer Research
9
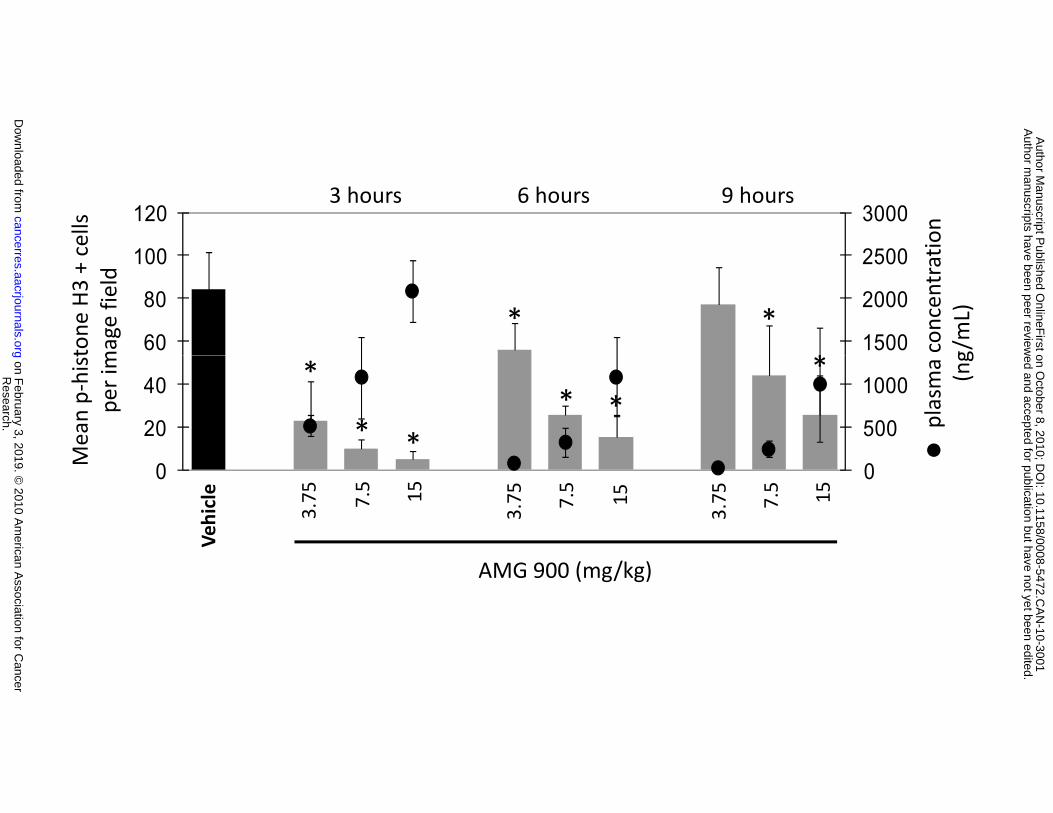
established HCT116 tumors were administered a single oral dose of vehicle alone or AMG 900 at 3.75,
7.5, or 15 mg/kg. As shown in Figure 4A and 4B, administration of AMG 900 resulted in significant dose-
dependent inhibition of p-histone H3 in tumors compared with the vehicle-treated control group (P ≤
0.008). Furthermore, the degree of p-histone H3 suppression in tumors correlated with plasma drug
concentrations. AMG 900 also inhibited p-histone H3 in mouse bone marrow cells at comparable doses
and plasma drug concentrations (data not shown). In a separate multiple dose study using the HCT116
tumor xenograft model, AMG 900 induced a marked increase in the percentage of tumor cells with ≥ 4N
DNA content (Supplementary Fig. S3).
Next, we compared the inhibition of aurora-B activity, as measured by the degree and duration of
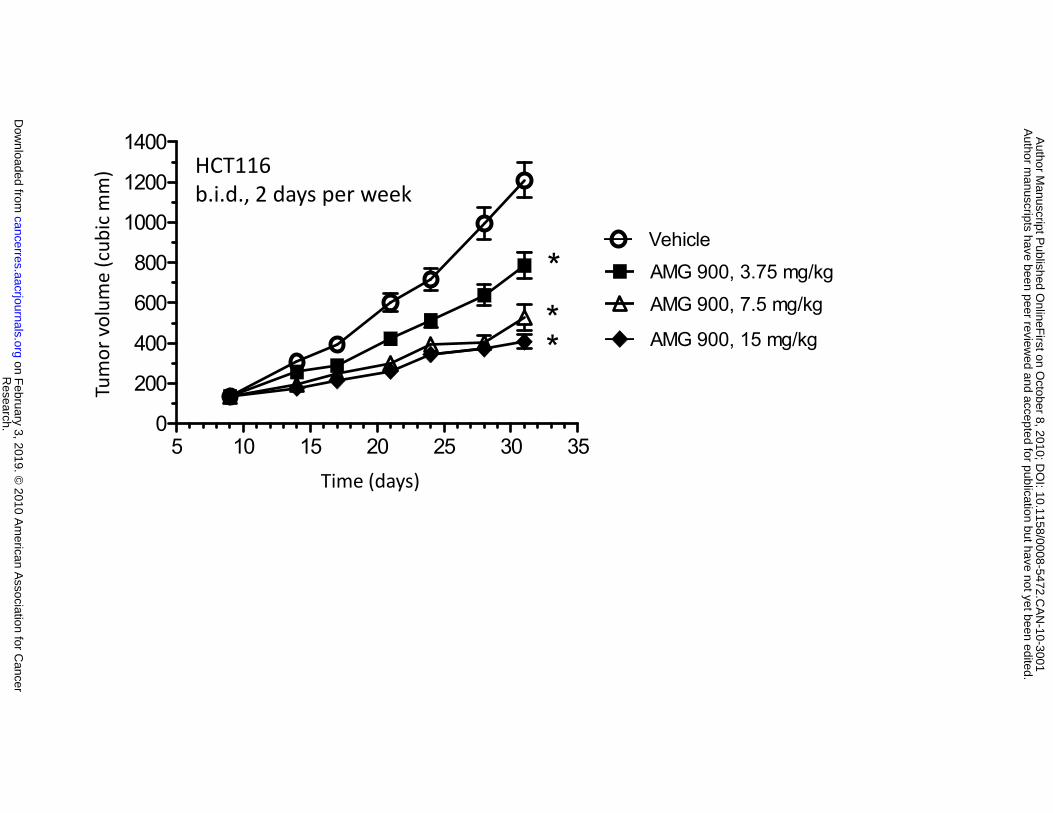
p-histone H3 inhibition, with suppression of tumor growth in vivo. Mice bearing established HCT116
tumors were orally administered vehicle alone or AMG 900 at 3.75, 7.5, or 15 mg/kg twice daily (b.i.d.) for
2 consecutive days per week for 3 weeks. As shown in Figure 4C, intermittent administration of AMG 900
resulted in a dose-dependent inhibition of tumor growth compared with the vehicle-treated control group
(3.75 mg/kg (40%), 7.5 mg/kg (64%), 15 mg/kg (75%), P ≤ 0.0001). The main adverse effects observed
with AMG 900 at the highest dose were moderate body weight loss (average of 7%, Supplementary Fig.
S4A) with transient and reversible myelosuppression (data not shown). An alternative schedule based on
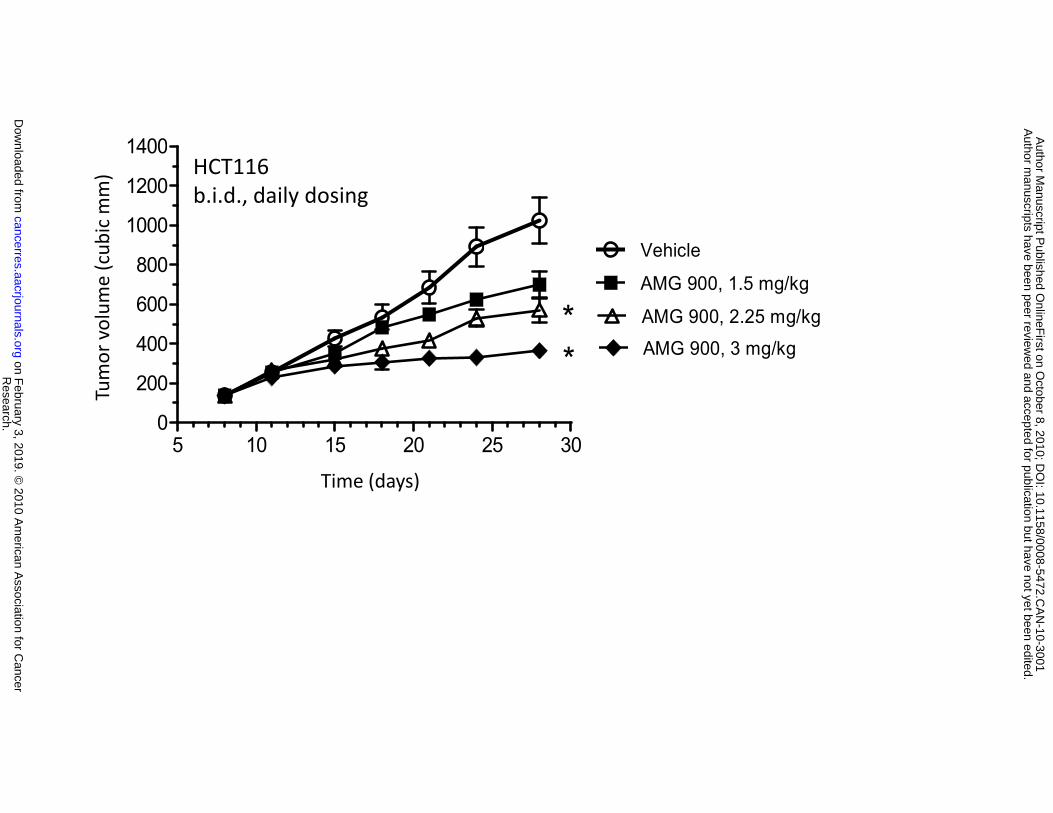
daily dosing was also tested. Mice with established HCT116 tumors were orally administered vehicle
alone or AMG 900 at 1.5, 2.25, or 3 mg/kg b.i.d. for 3 weeks (Fig. 4D). This continuous schedule with
lower doses of AMG 900 resulted in significant tumor growth inhibition when compared with the vehicle-
treated control group (2.25 mg/kg; 51% inhibition, P = 0.0019) and 3 mg/kg; 74% inhibition, P < 0.0001).
Daily dosing with AMG 900 was well-tolerated with no adverse effect on body weight (Supplementary Fig.
S4B), although a decrease in neutrophil counts was observed at the end of the study (data not shown).
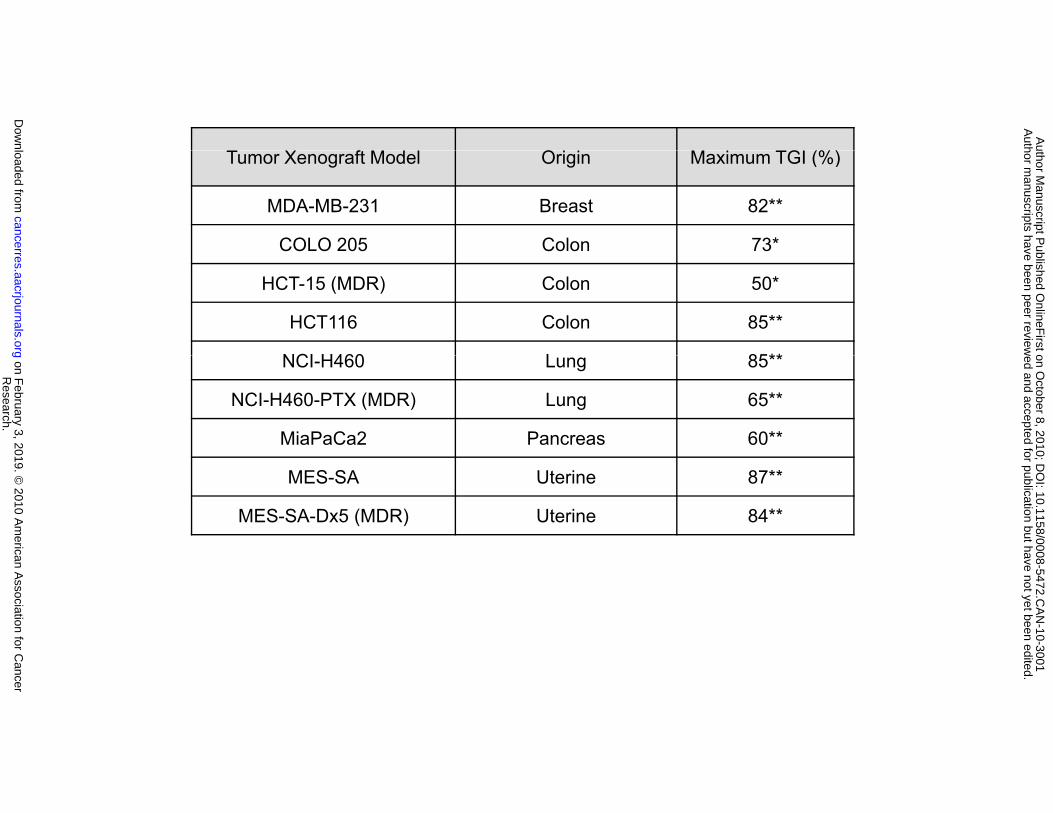
The effect of AMG 900 on tumor growth was further evaluated in a panel of human xenografts
from five different tumor types (breast, colon, lung, pancreatic, and uterine), including three MDR
xenograft models. Mice bearing established tumors were orally administered AMG 900 at 15 mg/kg b.i.d.
for two consecutive days per week or at 3 mg/kg b.i.d. every day for the duration of the study. As shown
in Figure 5A, AMG 900 exhibited significant antitumor activity in all nine xenograft models tested (50 to
Research. on February 3, 2019. © 2010 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 8, 2010; DOI: 10.1158/0008-5472.CAN-10-3001

Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Copyright © 2010 American Association for Cancer Research
10
97% tumor growth inhibition (TGI) compared with the vehicle-treated control group, *P < 0.005, **P <
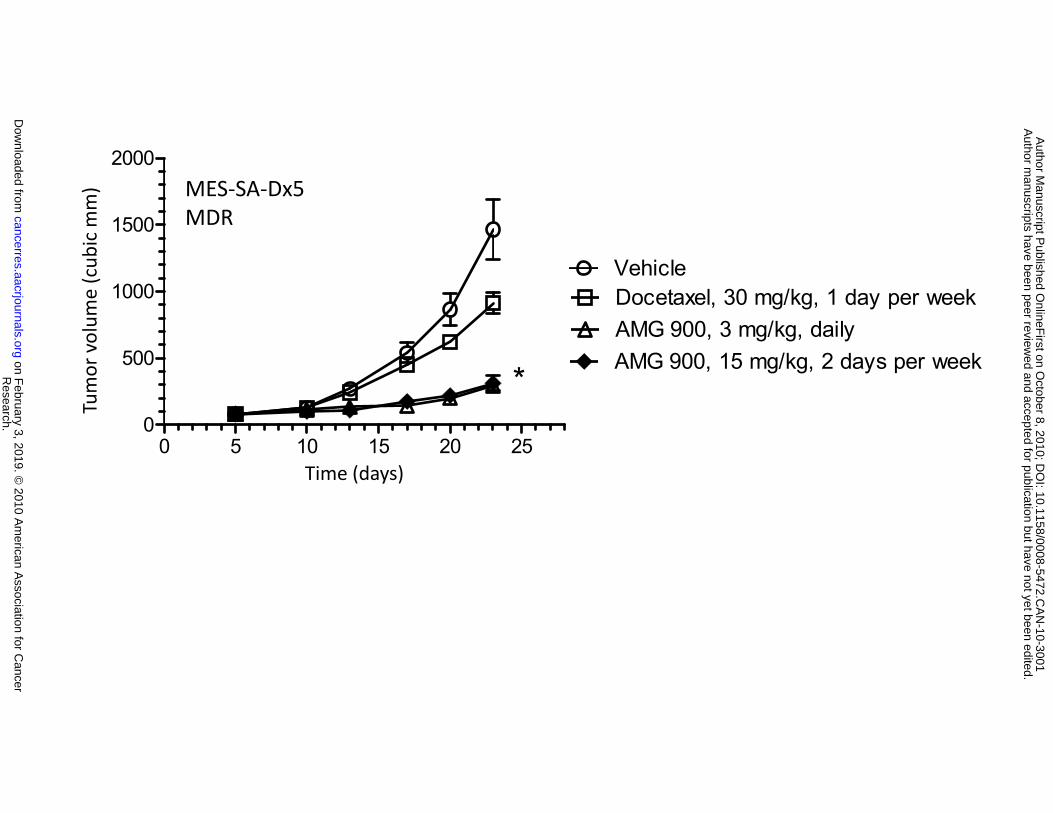
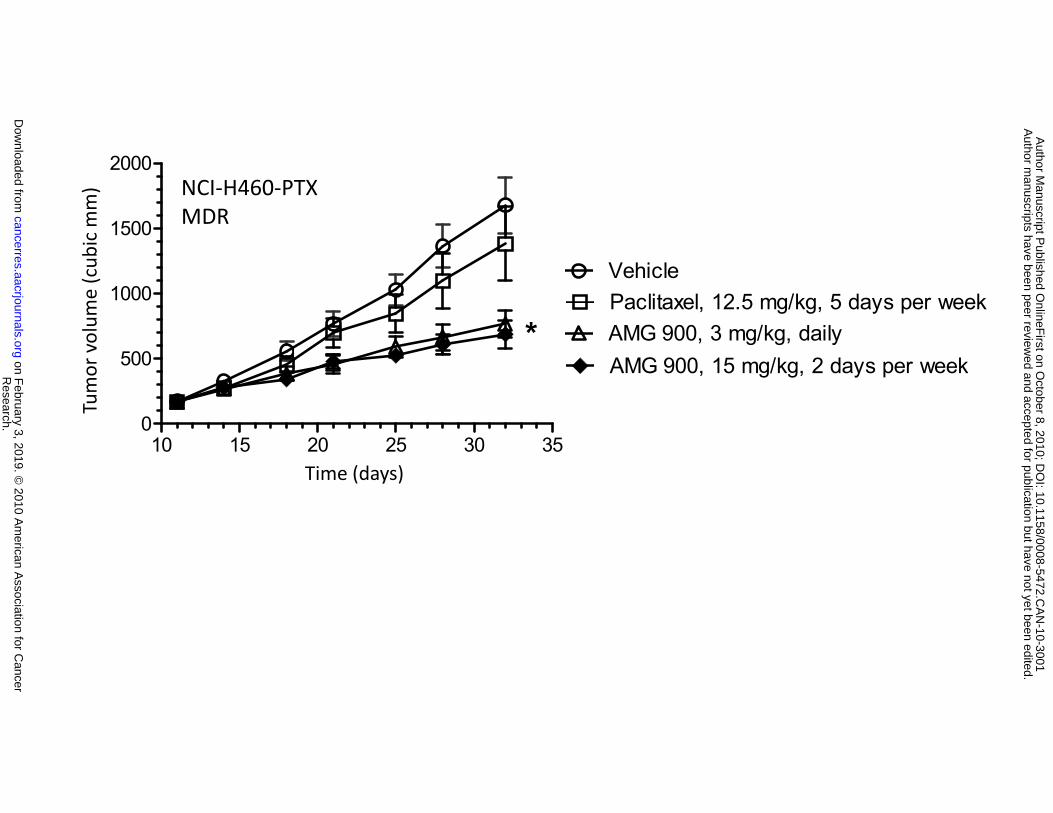
0.0005). Importantly, AMG 900 was active in the MES-SA-Dx5 (84% TGI, P < 0.0001) and NCI-H460-
PTX (66% TGI, P < 0.0001) xenograft models that were resistant to either docetaxel or paclitaxel
administered at their respective maximum tolerated doses (Fig. 5B and 5C). Together, these in vivo data
provide evidence that AMG 900 inhibits the activity of aurora-B in HCT116 tumors and suppresses the
growth of multiple xenografts that represent diverse tumor types. Notably, our data indicate that
AMG 900 has the potential to treat tumors resistant to standard-of-care antimitotic drugs.
Since inhibition of aurora-B activity is known to cause myelosuppression (11, 25), studies were
performed to examine the effects of PEGylated-granulocyte colony-stimulating factor (SD/02)
administration on AMG 900-induced neutropenia. Mice were treated with AMG 900 at 15 mg/kg b.i.d for
two consecutive days alone or with SD/02 at 1 mg/kg one day subsequent to the first day of AMG 900
treatment. Administration of AMG 900 alone showed a marked reduction in neutrophils by day 4 after
treatment followed by a full recovery of neutrophils by day 8. Mice administered SD/02 one day after the
initiation of AMG 900 treatment showed a significant decrease in the duration of neutropenia compared to
AMG 900 treatment alone (days 6, 7, and 8 (P < 0.0001) and day 11 (P < 0.0018), Supplementary Fig.
S5).
Discussion
Targeting the structural components of the mitotic machinery is a proven intervention strategy in the
treatment of cancer. Microtubule-binding agents such as the taxanes and vinca alkaloids are highly
active against a variety of human cancer types, although multidrug resistance and peripheral neuropathy
remain persistent problems. Consequently, there is a need for novel antimitotic drugs that target
nonmicrotubule proteins such as the mitotic kinases (38, 39). In the past decade, aurora-A and -B have
gained prominence as essential regulators of somatic cell division and represent promising mitotic targets
for anticancer therapy.
In this report, we have described the preclinical activities of AMG 900, a novel potent and highly
selective pan-aurora kinase inhibitor. The predominant response of tumor cells to AMG 900 treatment
Research. on February 3, 2019. © 2010 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 8, 2010; DOI: 10.1158/0008-5472.CAN-10-3001

Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Copyright © 2010 American Association for Cancer Research
11
was aborted cell division without a protracted mitotic arrest resulting in polyploidy-specific lethality. AMG
900 induces p53 and p21Cip1 proteins, which may act to regulate the rate of endoreduplication.
Remarkably, AMG 900 is active in all of the tested tumor cell lines at low nanomolar concentrations,
suggesting that it can inhibit the proliferation of tumor cells irrespective of genomic alterations or tumor
origin. It is possible that the underlying profile of genetic and epigenetic modifications in tumor cells may
play an important role in determining the fate of remnant cells that survive AMG 900 treatment. Because
cells must pass through G2-M phase of the cell cycle in order for AMG 900 to inhibit aurora kinase activity,
other factors such as mitotic and proliferation indices may impact tumor cell response. Further efforts will
be required to define potential susceptibility markers to help identify the most AMG 900-vulnerable tumors
as well as determining if combinatorial approaches lead to durable tumor regression.
Despite the ongoing debate regarding the relevance of drug efflux-mediated clinical resistance,
the consistent potency of AMG 900 against tumor cells irrespective of P-gp or BCRP status may
circumvent a mechanism by which tumor cells could otherwise escape the activity of this experimental
agent. Furthermore, we found that AMG 900 is active in an AZD1152-resistant cell line harboring a
missense mutation in aurora-B resulting in an amino acid substitution (W221L) in its kinase domain
activation loop. A recent report has shown that other aurora-B mutant alleles (Y156H, G160V) located in
the ATP-binding pocket confer resistance to multiple aurora inhibitors (35). It will be of interest to test
whether AMG 900 inhibits aurora-B activity in these catalytic domain mutants.
Consistent with our in vitro observations, AMG 900 inhibits the phosphorylation of histone H3 and
growth of multiple human tumor xenograft models using either intermittent or continuous dosing
schedules. Most relevant to our goal of developing an antimitotic agent that is effective against drug
resistant tumors, AMG 900 shows promising antitumor activity in tumor xenografts resistant to taxanes.
As anticipated, treatment of mice at efficacious doses of AMG 900 leads to a transient loss of body weight
and reversible myelosuppression.
In summary, AMG 900 is an orally bioavailable, potent and highly selective pan-aurora kinase
inhibitor with activity in taxane-resistant tumor cell lines. Together these features distinguish AMG 900
from other antimitotic drugs as well as three other aurora kinase inhibitors (AZD1152, MK-0457, and
Research. on February 3, 2019. © 2010 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 8, 2010; DOI: 10.1158/0008-5472.CAN-10-3001

Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Copyright © 2010 American Association for Cancer Research
12
PHA-739358). These key attributes contribute to the profile of this attractive clinical candidate; AMG 900
has entered phase 1 evaluation in adult patients with advanced cancers.
Acknowledgements
The authors would like to thank our Aurora team colleagues in the Departments of Lead Discovery,
Oncology and Hematology Research, Pathology, Toxicology, Medicinal Chemistry, Pharmacokinetics and
Drug Metabolism, and Pharmaceutics for their contributions and to Julie Bailis, Greg Friberg, Robert
Loberg, and Glenn Begley for their critical reading of this manuscript. We also gratefully acknowledge the
excellent technical assistance by Brian Belmontes, Jacob Corcoran, Tibor Gyuris, Scott Heller,
Raheemuddin Khaja, and Ji-Rong Sun.
Figure 1. AMG 900 is a potent and selective inhibitor of aurora-A, -B and -C. A, chemical structure
and enzyme selectivity profile of AMG 900. B, Western blotting of cell lysates from HeLa cells
synchronized in mitosis and treated with DMSO, AMG 900 at the indicated concentrations, MLN8054
(aurora-A inhibitor) at 500 nmol/L, or AZD1152 (aurora-B inhibitor) at 1000 nmol/L for 3 hours. C,
immunofluorescence imaging assay (ArrayScan VTi, Cellomics) was used to determined the inhibition of
aurora kinase activity in HeLa cells. The concentration-response curves were calculated based on the
decrease in the fluorescence intensity of p-histone H3 Ser10 () and p-aurora-A Thr288 () as a
percentage of the DMSO control (POC). IC50 values represent two independent experiments run in
duplicate (error bars, ±SD). D, representative live-cell imaging analysis of histone H2B-GFP HeLa cells
treated with DMSO or AMG 900 at 50 nmol/L. Cells were imaged over a period of 75 minutes (lower-right
hand corner of each image denotes the time in minutes).
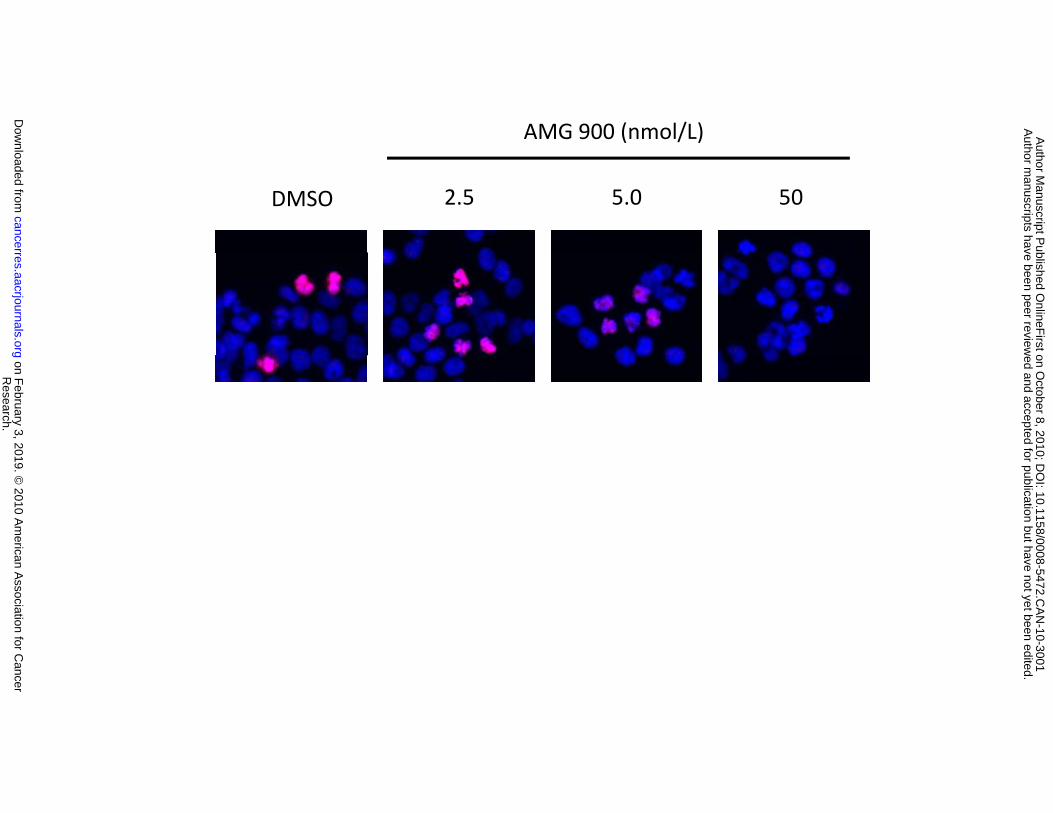
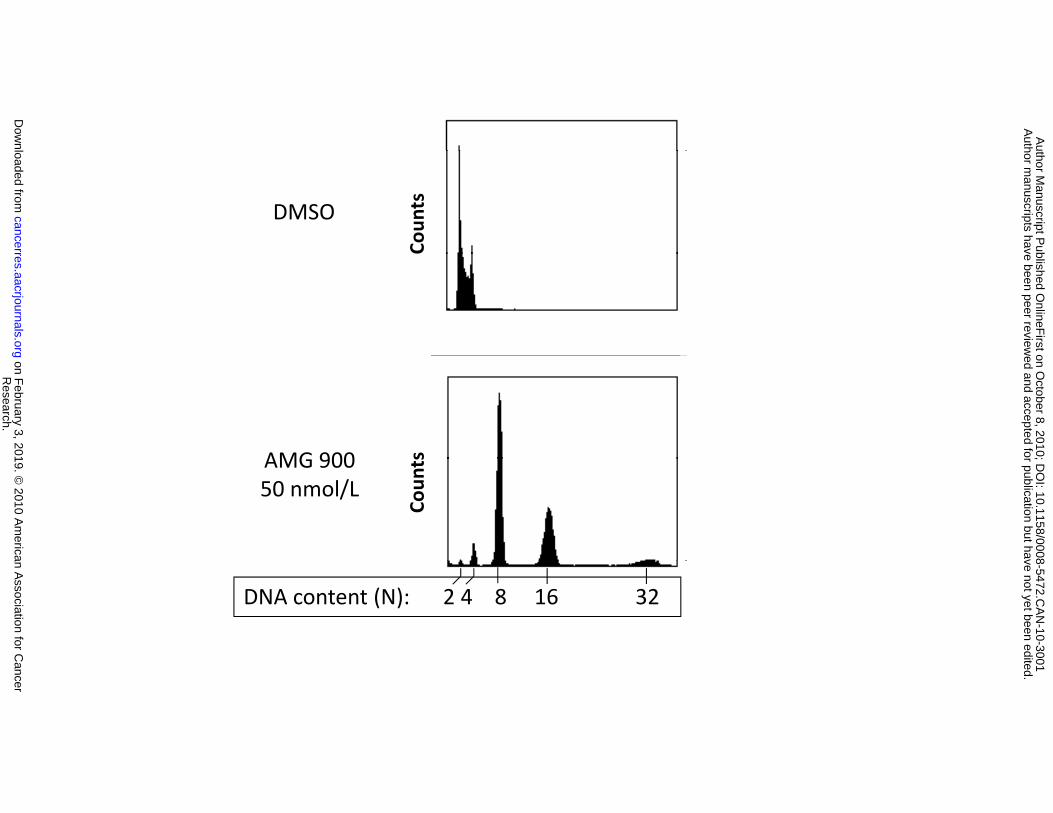
Figure 2. AMG 900 induces polyploidy and apoptosis in vitro in the human colon carcinoma
HCT116 cell line. A, representative images of cells treated with DMSO or AMG 900 at the indicated
concentrations for 6 hours. Cells were stained with p-histone H3 antibody (red) and DAPI (blue). B,
Research. on February 3, 2019. © 2010 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 8, 2010; DOI: 10.1158/0008-5472.CAN-10-3001

Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Copyright © 2010 American Association for Cancer Research
13
DNA content profiles of cells treated with DMSO or AMG 900 at 50 nmol/L for 48 hours. Cells were
stained with propidium iodide and evaluated by flow cytometry. C, cells were treated with DMSO or AMG
900 at 50 nmol/L for 48 hours and replated in complete media lacking inhibitor. After 10 days of growth,
cells were stained with crystal violet and imaged (triplicate wells). D, cells were treated with DMSO or
AMG 900 at 50 nmol/L for 48 hours, washed in complete media, and then cultured in complete media
lacking inhibitor for the indicated time intervals. Western blotting documenting the protein levels of
cleaved caspase-7 and β-actin loading control.
Figure 3. AMG 900 inhibits cell proliferation and colony formation in human tumor cell lines at low
nanomolar concentrations, including MDR cell lines resistant to paclitaxel. A and B, fluorescence-
based cell count imaging assay (ArrayScan VTi) was performed on a panel of 26 tumor cell lines treated
with AMG 900 (concentration range 0.3 to 156 nmol/L). Cells were treated for 24 hours, washed twice,
and incubated in complete media lacking inhibitor for 48 hours. A, representative concentration-
response curve of PC3 cells treated with AMG 900. B, cell lines rank ordered by sensitivity to AMG 900
and color coded by tumor origin. C, colony formation assay was performed in three paclitaxel-sensitive
cell lines (parental) and in four MDR cell lines (P-gp expressing). Cells were treated with paclitaxel or
AMG 900 at the indicated concentrations for 48 hours and replated in complete media lacking inhibitor.
After the DMSO-treated cells reached confluence, the cells were stained with crystal violet and imaged
(duplicate wells).
Table 1. AMG 900 exhibits uniform potency in human tumor cell lines expressing P-gp and BCRP
resistant to one or more aurora kinase inhibitors
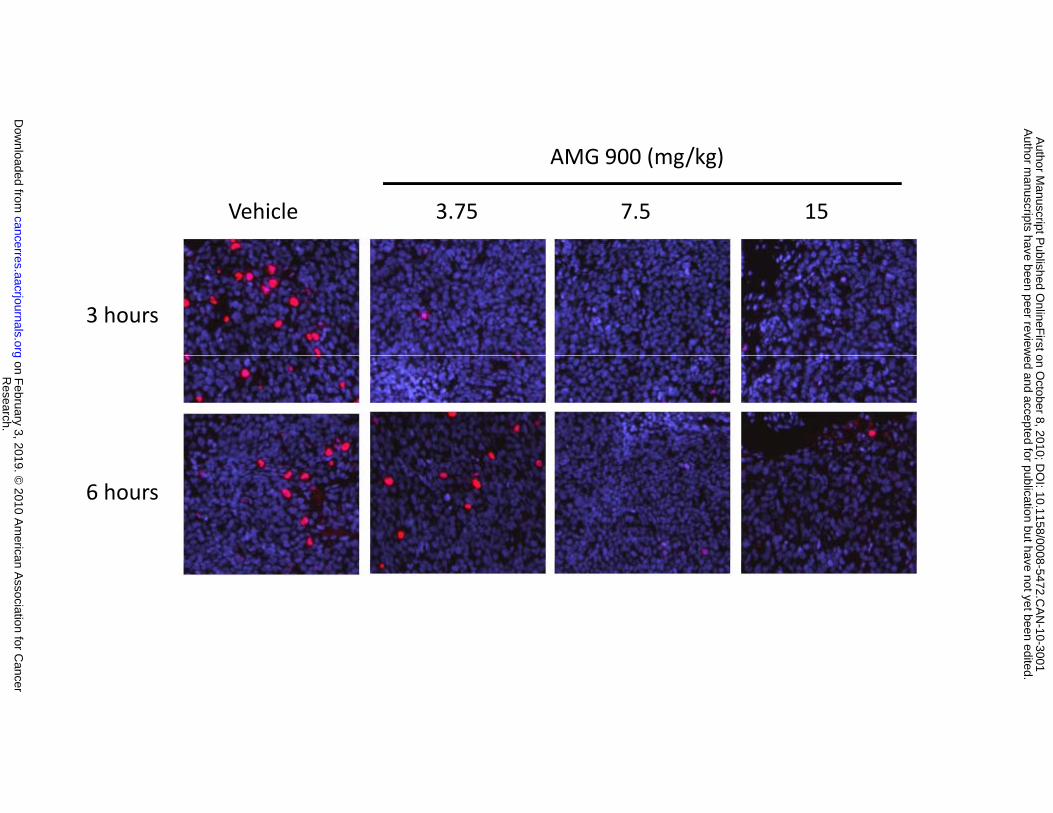
Figure 4. AMG 900 inhibits phosphorylation of histone H3 and the growth of HCT116 tumor
xenografts in vivo. A and B, mice bearing established tumors were orally administered a single dose of
vehicle alone or AMG 900 at 3.75, 7.5, or 15 mg/kg. Tumors were collected at 3, 6, or 9 hours after
treatment (n = 4 per time point per dose) and the level of p-histone H3 was determined by fluorescence-
based microscopy. Representative images of sectioned tumors stained with p-histone H3 antibody (red)
Research. on February 3, 2019. © 2010 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 8, 2010; DOI: 10.1158/0008-5472.CAN-10-3001

Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Copyright © 2010 American Association for Cancer Research
14
and DAPI (blue). B, bar graph representing the pharmacodynamic-pharmacokinetic profile of AMG 900
in mice. The number of p-histone H3 positive cells per tumor image field (8 image fields per column,
mean +SD) and concentration of AMG 900 in mouse plasma (, mean ±SD). The control represents the
combined 3 and 6 hour vehicle-treated tumors (16 image fields per column, mean +SD). Statistically
significant inhibition of p-histone H3 compared with the vehicle-treated control is denoted by an asterisk
(*P ≤ 0.008). C and D, mice bearing established HCT116 tumors were orally administered vehicle alone
() and AMG 900 b.i.d. either intermittently at 3.75 mg/kg (), 7.5 mg/kg (), or 15 mg/kg () for 2
consecutive days per week for 3 weeks (C) or every day at 1.5 mg/kg (), 2.25 mg/kg (), or 3 mg/kg
() every day for 3 weeks (D). Tumor volumes (cubic mm) are represented as mean ±SE (n = 10).
Statistically significant tumor growth inhibition compared with the vehicle-treated control is denoted by an
asterisk (*P ≤ 0.0001).
Figure 5. AMG 900 inhibits the growth of multiple human tumor xenografts in vivo, including
multidrug resistant models. A, mice bearing established tumor xenografts were orally administered
AMG 900 at 15 mg/kg b.i.d. for 2 consecutive days per week for 3 weeks or at 3 mg/kg b.i.d. every day for
3 weeks. The maximum percentage of tumor growth inhibition (TGI) is reported for each xenograft model.
Statistically significant tumor growth inhibition compared with the vehicle-treated control is denoted by
asterisks (*P < 0.005, **P < 0.0005). B and C, mice bearing established multidrug resistant MES-SA-Dx5
or NCI-H460-PTX tumor xenografts were orally administered vehicle alone (), AMG 900 at 15 mg/kg
b.i.d. () for 2 consecutive days per week or at 3 mg/kg b.i.d. () every day. As a control, mice were
administered docetaxel intraperitoneal (i.p.) at 30 mg/kg () once per week or paclitaxel i.p. at 12.5
mg/kg () 5 days per week. Tumor volumes (cubic mm) are represented as mean ±SE (n = 10).
Statistically significant tumor growth inhibition compared with the vehicle-treated control is denoted by an
asterisk (*P ≤ 0.0019).
References
1. Quasthoff S, Hartung HP. Chemotherapy-induced peripheral neuropathy. J Neurol 2002;249:9-17.
2. Keen N, Taylor S. Aurora-kinase inhibitors as anticancer agents. Nat Rev Cancer 2004;4:927-36.
Research. on February 3, 2019. © 2010 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 8, 2010; DOI: 10.1158/0008-5472.CAN-10-3001

Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Copyright © 2010 American Association for Cancer Research
15
3. Keen N, Taylor S. Mitotic drivers - Inhibitors of the aurora B kinase. Cancer Met Rev 2009;28:185-95.
4. Carvajal RD, Tse A, Schwartz GK. Aurora kinases: new targets for cancer therapy. Clin Cancer Res 2006;12: 6869-75.
5. Barr AR, Gergeley, F. Aurora-A: The maker and breaker of spindle poles. J Cell Science 2007;120:2987-96.
6. Macurek L, Lindqvist A, Lim D, et al. Polo-like kinase-1 is activated by aurora A to promote checkpoint recovery. Nature
2008;455:119-23.
7. Ditchfield C, Johnson VL, Tighe A, et al. Aurora B couples chromosome alignment with anaphase by targeting BubR1,
Mad2, and Cenp-E to kinetochores. J Cell Biol 2003;161:267-80.
8. Steigemann P, Wurzenberger C, Schmitz MHA, et al. Aurora B-mediated abscission checkpoint protects against
tetraploidization. Cell 2009;136:473-84.
9. Liu D, Vader G, Vromans MJM, et al. Sensing chromosome bi-orientation by spatial separation of aurora B kinase from
kinetochore substrates. Science 2009;323:1350-53.
10. Kimmins S, Crosio C, Kotaja N, et al. Differential functions of the aurora-B and aurora-C kinases in mammalian
spermatogenesis. Mol Endocrinol 2007;21:726-39.
11. Gautschi O, Heighway J, Mack PC, Purnell PR, Lara PN, Gandara DR. Aurora kinases as anticancer drug targets. Clin
Cancer Res 2008;14:1639-48.
12. Musacchio A, Salmon ED. The spindle-assembly checkpoint in space in time. Nat Rev Mol Cell Biol 2007;8: 379-93.
13. Hauf S, Cole RW, LaTerra S, et al. The small molecule hesperadin reveals a role for aurora B in correcting kinetochore–
microtubule attachment and in maintaining the spindle assembly checkpoint. J Cell Biol 2003;161: 281-94.
14. Yang H, Burke T, Dempsey J, et al. Mitotic requirement for aurora A kinase is bypassed in the absence of aurora B
kinase. FEBS Letters 2005;579:3385-91.
15. Girdler F, Gascoigne KE, Eyers PA, et al. Validating aurora B as an anti-cancer drug target. J Cell Sci 2006; 119:3664-
75.
16. Weaver BA, Cleveland DW. Decoding the links between mitosis, cancer, and chemotherapy: The mitotic checkpoint,
adaptation, and cell death. Cancer Cell 2005;8:7-12.
17. Gascoigne KE, Taylor SS. Cancer cells display profound intra- and interline variation following prolonged exposure to
antimitotic drugs. Cancer Cell 2008;14:111-22.
18. Perez EA. Microtubule inhibitors: Differentiating tubulin-inhibiting agents based on mechanism of action, clinical activity,
and resistance. Mol Cancer Ther 2009;8:2086-95.
19. Yin S, Bhattacharya R, Cabral F. Human mutations that confer paclitaxel resistance. Mol Cancer Ther 2010; 9:327-35.
20. McGrogan BT, Gilmartin B, Carney DN, McCann A. Taxanes, microtubules, and chemoresistant breast cancer. Biochim et
Biophys Acta 2008;1785:96-132.
21. Szakács G, Paterson JK, Ludwig JA, Booth-Genthe C, Gottesman MM. Targeting multidrug resistance in cancer. Nat Rev
Drug Disc 2006 5:219-34.
Research. on February 3, 2019. © 2010 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 8, 2010; DOI: 10.1158/0008-5472.CAN-10-3001

Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Copyright © 2010 American Association for Cancer Research
16
22. Guo J, Anderson MG, Tapang P, et al. Identification of genes that confer tumor cell resistance to the aurora B kinase
inhibitor, AZD1152. Pharmacogenomics J 2009;9:90-102.
23. Carter TA, Wodicka LM, Shah NP, et al. Inhibition of drug-resistant mutants of ABL, KIT, and EGF receptor kinases.
PNAS 2005;102:11011-16.
24. Yun C-H, Boggon TJ, Li Y, et al. Structures of lung cancer-derived EGFR mutants and inhibitor complexes: mechanism of
activation and insights into differential inhibitor sensitivity. Cancer Cell 2007;11:217–27.
25. Lapenna S, Giordano A. Cell cycle kinases as therapeutic targets for cancer. Nat Rev Drug Disc 2009;8:547-66.
26. Crosio C, Fimia GM, Loury R, et al. Mitotic phosphorylation of histone H3: spatio-temporal regulation by mammalian
aurora kinases. Mol Cell Bio 2002;22:874-85.
27. Payton M, Chung G, Yakowec P, et al. Discovery and evaluation of dual CDK1 and CDK2 inhibitors. Cancer Res
2006;66:4299-4308.
28. Karaman MW, Herrgard S, Treiber DK, et al. A quantitative analysis of kinase inhibitor selectivity. Nature Biotech
2008;26:127-32.
29. Gizatullin F, Yao Y, Kung V, Harding MW, Loda M, Shapiro GI. The Aurora kinase inhibitor VX-680 induces
endoreduplication and apoptosis preferentially in cells with compromised p53-dependent post-mitotic checkpoint function.
Cancer Res 2006;66:7668-77.
30. Nair JS, Ho AL, Tse AN, et al. Aurora B kinase regulates the postmitotic endoreduplication checkpoint via
phosphorylation of the retinoblastoma protein at serine 780. Mol Biol Cell 2009;20:2218-28.
31. Vakifahmetoglu H, Olsson M, Zhivotovsky B. Death through a tragedy: mitotic catastrophe. Cell Death and Diff
2008;15:1153-62.
32. Harker WG, Sikic BI. Multidrug (pleiotropic) resistance in doxorubicin-selected variants of the human sarcoma cell line
MES-SA. Cancer Res 1985;45:4091-96.
33. Szakacs G, Annereau J-P, Lababidi S, et al. Drug sensitivity and resistance: Profiling ABC transporter genes in cancer
cells. Cancer Cell 2004;6:129-37.
34. Gyorffy B, Surowiak P, Kiesslich O, et al. Gene expression profiling of 30 cancer cell lines predicts resistance towards 11
anticancer drugs at clinically achieved concentrations. Int J Cancer 2006;118:1699-1712.
35. Girdler F, Sessa F, Patercoli S, et al. Molecular basis of drug resistance in aurora kinases. Chemistry & Biology
2008;15:552-62.
36. Davies H, Bignell GR, Cox C, et al. Mutations of the BRAF gene in human cancer. Nature 2002;417:949-54.
37. Paez JG, Janne PA, Lee JC, et al. EGRF mutations in lung cancer: Correlation with clinical response to Gefitinib therapy.
Science 2004;304:1497-1500.
38. Jackson JR, Patrick DR, Dar MM, Huang PS. Targeted anti-mitotic therapies: Can we improve on tubulin agents? Nat Rev
Cancer 2007;7:107-17.
Research. on February 3, 2019. © 2010 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 8, 2010; DOI: 10.1158/0008-5472.CAN-10-3001

Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Copyright © 2010 American Association for Cancer Research
17
39. Luo J, Solimini NL, Elledge SJ. Principles of cancer therapy: oncogene and non-oncogene addication. Cell 2009;136:823-
37.
40. Freeman D, Bush T, Ogbagabriel S, et al. Activity of panitumumab alone or with chemotherapy in non-small cell lung
cancinoma cell lines expressing mutant epidermal growth factor receptor. Mol Cancer Ther 2009;9:1536-46.
Research. on February 3, 2019. © 2010 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 8, 2010; DOI: 10.1158/0008-5472.CAN-10-3001

Figure 1Figure 1(A, B, C, D)
ColorColor
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001
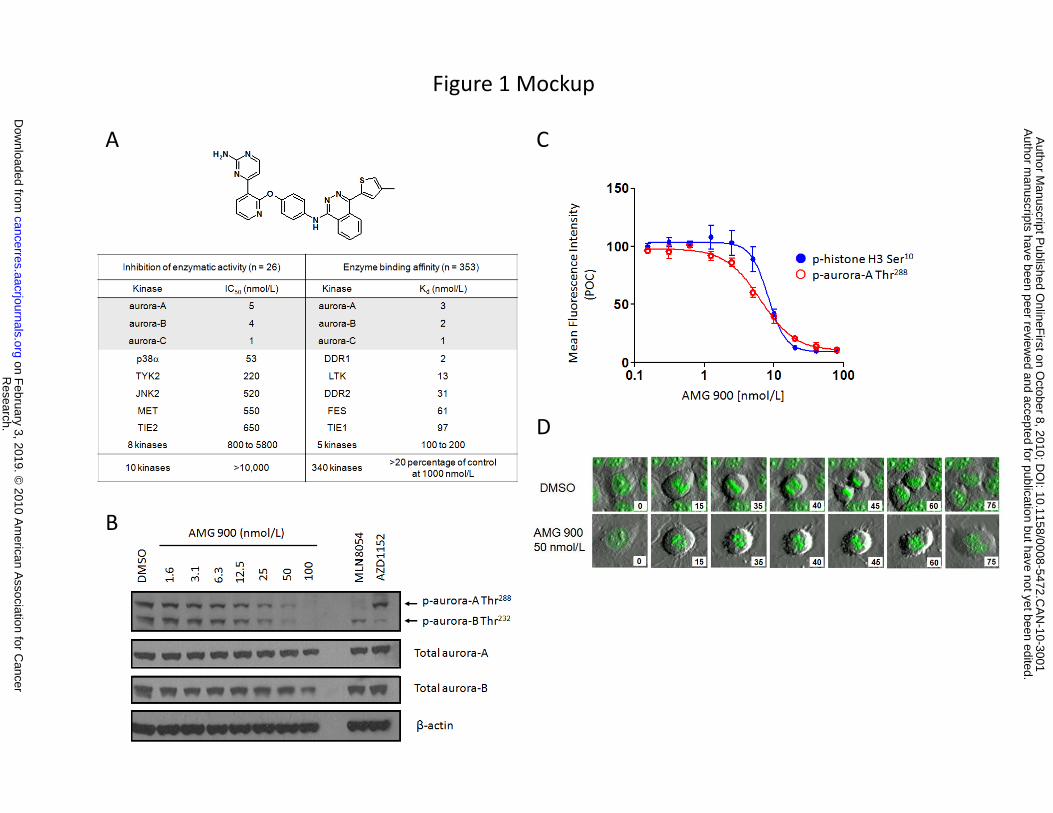
Figure 1 Mockup
A C
D
B
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001
N
NNH2
S
N
O
NH
NN
Inhibition of enzymatic activity (n = 26) Enzyme binding affinity (n = 353)
Kinase IC (nmol/L) Kinase Kd (nmol/L)Kinase IC50 (nmol/L) Kinase Kd (nmol/L)
aurora-A 5 aurora-A 3
aurora-B 4 aurora-B 2
aurora C 1 aurora C 1aurora-C 1 aurora-C 1
p38α 53 DDR1 2
TYK2 220 LTK 13
JNK2 520 DDR2 31JNK2 520 DDR2 31
MET 550 FES 61
TIE2 650 TIE1 97
8 kinases 800 to 5800 5 kinases 100 to 2008 kinases 800 to 5800 5 kinases 100 to 200
10 kinases >10,000 340 kinases >20 percentage of control at 1000 nmol/L
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001

O 054
152AMG 900 (nmol/L)
p-aurora-A Thr288
DM
SO
1.6
3.1
6.3
12.5
25 50 100
MLN
80
AZD
11
p
p-aurora-B Thr232
Total aurora-A
Total aurora-B
β-actin
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001

150
sity
100 p-histone H3 Ser10
p-aurora-A Thr288
cenc
e In
ten
OC)
50
p
an F
luor
esc
(PO
0.1 1 10 1000M
ea
AMG 900 [nmol/L]AMG 900 [nmol/L]
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001
DMSO0 15 35 40 45 60 75
AMG 90050 nmol/L
0 15 35 40 45 60 750 15 35 40 45 60 75
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001

Figure 2Figure 2(A, B, C, D)
ColorColor
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001
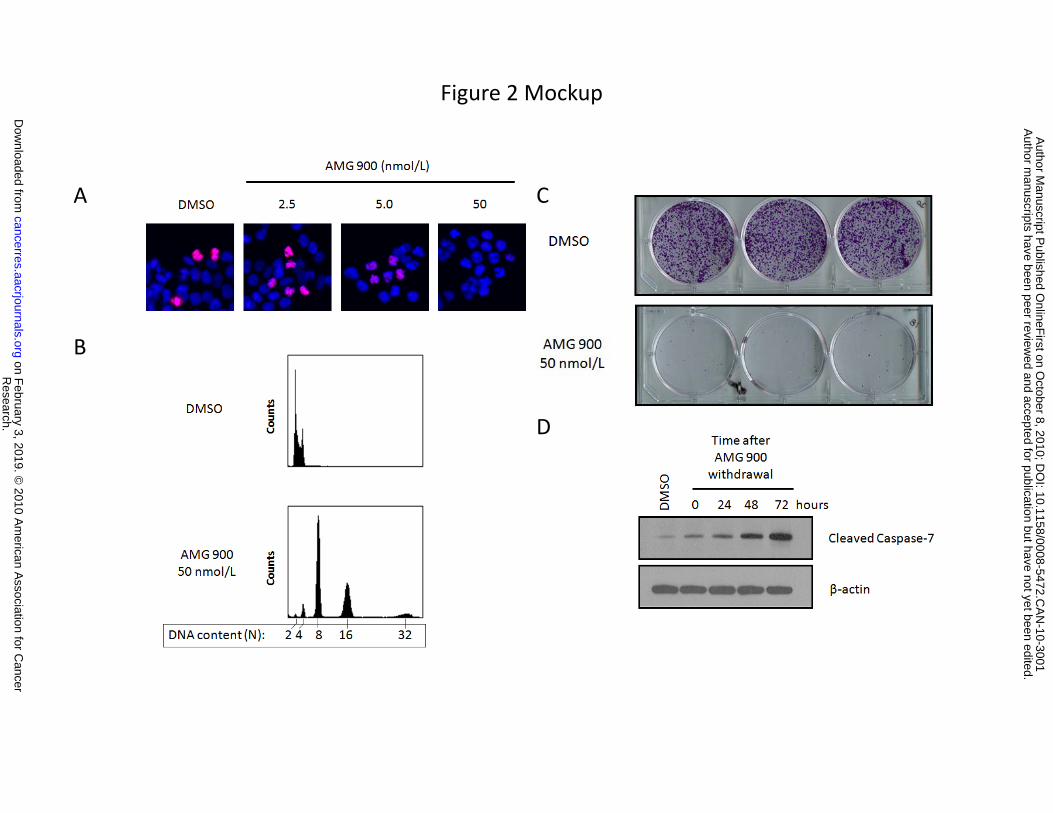
Figure 2 Mockup
A C
B
D
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001
AMG 900 (nmol/L)
DMSO 2.5 5.0 50
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001
Coun
tsDMSO
C
AMG 900 sAMG 90050 nmol/L
Coun
ts
DNA content (N): 2 4 8 16 32
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001
DMSO
AMG 900AMG 90050 nmol/L
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001
Time after
0 24 48 72 hMSO
Time after AMG 900
withdrawal
Cleaved Caspase-7
0 24 48 72 hoursD
β-actin
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001

Figure 3Figure 3(A, B,C)ColorColor
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001
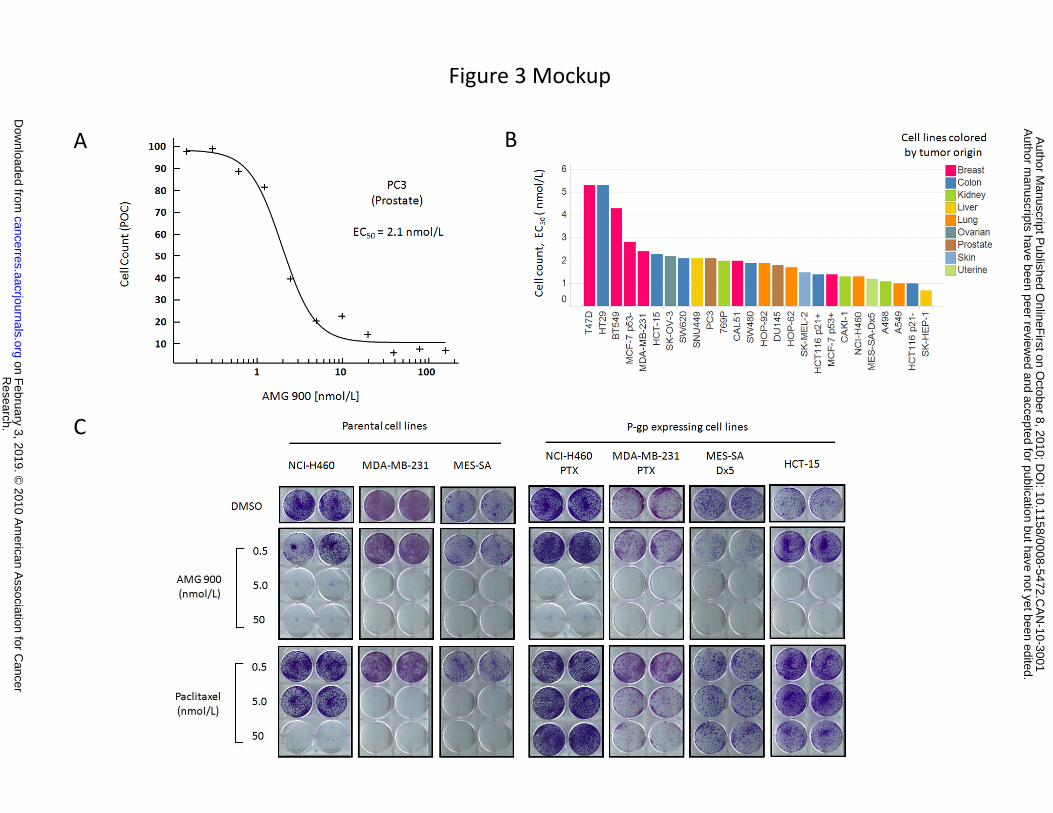
Figure 3 Mockup
A B
C
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001
PC3 80
90
100
(Prostate)
EC50 = 2.1 nmol/L
unt (
POC)
60
70
Cell
Cou
30
40
50
10
20
AMG 900 [nmol/L]
1 10 100
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001
/L)
Cell lines colored
by tumor origin
C 50 (
nmol
/el
l cou
nt,
ECe
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001

NCI H460 MDA MB 231 MES SA
Parental cell lines
DMSO
NCI-H460 MDA-MB-231 MES-SA
0.5
5.0
0
AMG 900(nmol/L)
50
0 5
Paclitaxel(nmol/L)
0.5
5.0
50
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001

NCI-H460 MDA-MB-231 MES-SA HCT 15
P-gp expressing cell lines
PTX PTX Dx5 HCT-15
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Copyright © 2010 American Association for Cancer Research
Research. on February 3, 2019. © 2010 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 8, 2010; DOI: 10.1158/0008-5472.CAN-10-3001

Figure 4Figure 4(A, B, C, D)
ColorColor
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001

A B
Figure 4 Mockup
C D
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001
AMG 900 ( /k )
Vehicle 3.75 7.5 15
AMG 900 (mg/kg)
3 hours
6 hours
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001
2500
3000
100
1203 hours 6 hours 9 hours
tion
cells
1500
2000
2500
60
80
100
conc
entr
atg/
mL)* *
*
tone
H3
+ c
mag
e fie
ld
500
1000
20
40
pl
asm
a (ng
*
* ** *
*
Mea
n p-
hist
per
im
00
Vehicle
3.75 7.
5 15
3.75 7.
5
15
3.75 7.
5 15
M
AMG 900 (mg/kg)
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001
1400
1000
1200
Vehicle*
HCT116b.i.d., 2 days per week
cubi
c m
m)
400
600
800
AMG 900, 15 mg/kg
AMG 900, 7.5 mg/kgAMG 900, 3.75 mg/kg*
**or
vol
ume
(c
5 10 15 20 25 30 350
200Tum
o
Time (days)
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001
1400
1000
1200
1400
Vehicle
HCT116b.i.d., daily dosing
ubic
mm
)
400
600
800Vehicle
AMG 900, 3 mg/kg
AMG 900, 2.25 mg/kg
AMG 900, 1.5 mg/kg
**or
vol
ume
(cu
5 10 15 20 25 300
200
, g g*
Tum
o
Time (days)
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001

Figure 5Figure 5(A, B,C)
Black and WhiteBlack and White
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001
Figure 5 Mockup
A B
C
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001
f O G (%)Tumor Xenograft Model Origin Maximum TGI (%)
MDA-MB-231 Breast 82**
COLO 205 Colon 73*
HCT-15 (MDR) Colon 50*
HCT116 Colon 85**
NCI H460 L 85**NCI-H460 Lung 85**
NCI-H460-PTX (MDR) Lung 65**
MiaPaCa2 Pancreas 60**
MES-SA Uterine 87**
MES-SA-Dx5 (MDR) Uterine 84**
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001
2000MES-SA-Dx5MDR1500
2000
bic
mm
)
500
1000Vehicle
AMG 900, 3 mg/kg, dailyAMG 900 15 /k 2 d k
Docetaxel, 30 mg/kg, 1 day per week
volu
me
(cub
0 5 10 15 20 250
500 AMG 900, 15 mg/kg, 2 days per week*
Tum
or
Time (days)
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001
NCI-H460-PTXMDR
bic
mm
)
1500
2000
volu
me
(cub
500
1000Vehicle
AMG 900, 3 mg/kg, dailyPaclitaxel, 12.5 mg/kg, 5 days per week
*
Tum
or
10 15 20 25 30 350
500 AMG 900, 15 mg/kg, 2 days per week
Time (days)
Research.
on February 3, 2019. ©
2010 Am
erican Association for C
ancercancerres.aacrjournals.org
Dow
nloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on O
ctober 8, 2010; DO
I: 10.1158/0008-5472.CA
N-10-3001

Published OnlineFirst October 8, 2010.Cancer Res Marc N. Payton, Tammy L. Bush, Grace Chung, et al. Taxane-Resistant Tumor Cell LinesSelective Pan-Aurora Kinase Inhibitor with Activity in Preclinical Evaluation of AMG 900, a Novel Potent and Highly
Updated version
10.1158/0008-5472.CAN-10-3001doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2010/10/08/0008-5472.CAN-10-3001.DC1
Access the most recent supplemental material at:
Manuscript
Authoredited. Author manuscripts have been peer reviewed and accepted for publication but have not yet been
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/early/2010/10/06/0008-5472.CAN-10-3001To request permission to re-use all or part of this article, use this link
Research. on February 3, 2019. © 2010 American Association for Cancercancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 8, 2010; DOI: 10.1158/0008-5472.CAN-10-3001









![Furine and Pyrimidine Ribonucleoside Monophosphate ...cancerres.aacrjournals.org/content/canres/42/4/1326.full.pdf · [CANCER RESEARCH 42, 1326-1330, April1982] 0008-5472/82/0042-0000$02.00](https://img.dokumen.tips/doc/110x75/5a7e72ec7f8b9ae9398e75ca/furine-and-pyrimidine-ribonucleoside-monophosphate-cancer-research-42-1326-1330.jpg)










![Characteristics of Mitochondria Isolated by Rate Zonal ... · [CANCER RESEARCH 40, 1486-1492. May 1980] 0008-5472/80/0040-OOOOS02.00 Characteristics of Mitochondria Isolated by Rate](https://img.dokumen.tips/doc/110x75/5faea77da5997b3e8b24fc87/characteristics-of-mitochondria-isolated-by-rate-zonal-cancer-research-40.jpg)








