Embed Size (px)
Citation preview

PORTUGALA | 1 Fe ve re i ro 2 0 1 5 | S e m e s t ra l | Vo l u m e 1 6
PORTUGALA
BOLETIM DO INSTITUTO PORTUGUÊS DE MALACOLOGIA
Volume 22 | Semestral | Dezembro de 2018
Cefalópodes como vetores de biotoxinas

2 | PORTUGALA
Vol.22 | Dezembro de 2018 | Semestral
Caros leitores,É com grande satisfação que lançamos o segundo número da Portugala de 2018. Neste número abordamos o papel dos ce-falópodes como vetores de biotoxinas, o impacto da acidificação dos oceanos em cefalópodes (recém eclodidos) e bivalves, de espécies invasoras (não nativas) em estuários portugueses, assim como os re-sumos: i) das jornadas sobre conservação da lapa ferrugínea e ii) do congresso de cefalópodes que decorreu na Flórida (CIAC), entre vários outros assuntos. A não perder!
Boa leitura! Saudações malacológicas...
Nota do Editor
Rui RosaInvestigador MARE e Docente FCUL
NESTE VOLUME....DESTAQUE
Cefalópodes Como vetores de biotoxinas ____________________3
ARTIGOS
impaCto da aCidifiCação dos oCeanos no desenvolvimento e Comporta-mento de ChoCos reCém-eClodidos_________________________ 6
Qual será o futuro das populações naturais de pé-de-burrinho (Cha-melea gallina) e ConQuilha (Donax trunCulus) num oCeano aCidifiCado e aQueCido?_____________________________________ 10
Coleção de molusCos no museu da CiênCia da universidade de Coimbra: uma visão geral e perspetivas atuais________________12
Cephalopod international advisory CounCil - 2018 flórida ___14
invasão à vista nos estuários do tejo e do sado______________ 15
jornadas sobre a Conservação da lapa ferrugínea (Patella ferrugí-nea)_____________________________________________ 17
notíCias do reCtângulo_______________________________18
publiCações em destaQue_____________________________ 20
proposta de sóCio__________________________________ 22

PORTUGALA | 3
Por Vanessa Madeira Lopes1, Pedro R. Costa2, Rui Rosa1
Cefalópodes como vetores de biotoxinas
DESTAQUE
1MARE – Marine and Environmental Sciences Centre, Laboratório Marítimo da Guia, Faculdade de Ciências da Universidade de Lisboa, Av. Nossa Senhora do Cabo, 939, 2750-374 Cascais, Portugal.
2 IPMA - Insituto Português do Mar e da Atmosfera, Avenida Brasília, 1449-006 Lisboa, Portugal
As comunidades fitoplanctónicas são tipicamente benéficas para o ecossistema, uma vez que constituem fonte de alimento para inúmeros organismos. De todas as espécies de fitoplâncton conhecidas, apenas uma pequena percentagem produz toxinas (Hallegraeff, 2014; Smayda, 1997). No entanto, quando as condições do meio são consideradas ideais, estes organismos atingem elevadas taxas de crescimento e elevadas densidades, dando origem aos “blooms” de algas nocivas (Hallegraeff, 1993). De um ponto de vista ecológico, as toxinas provenientes de “blooms” de algas nocivas, podem causar efeitos devastadores nas comunidades
marinhas, uma vez que, dependendo da espécie produtora, as toxinas podem atuar no sistema nervoso central, a vários níveis, ou no trato gastrointestinal (Lopes et al., in press).
Uma das toxinas marinhas mais estudadas é o ácido domóico (AD), uma potente neurotoxina produzida por diatomáceas do género Nitzschia e Pseudo-nitzschia, que causa amnésia em vertebrados (Amnesic Shellfish Poisoning – ASP). Esta toxina atua ao nível das células neurais, competindo pelos mesmos recetores que o glutamato, um neurotransmissor excitatório. Assim, o glutamato é impedido de se ligar aos recetores,
despoletando a ativação de recetores de AMPA, cainato e NMDA, mantendo a membrana celular permanentemente aberta, que irá resultar num fluxo constante de Ca2+ para o interior das células (Berman and Murray, 1997), causando despolarização da membrana e subsequente degeneração das células neurais.
Outras neurotoxinas de ocorrência global são as toxinas paralisantes (Paralytic Shellfish Toxins – PSTs). Estes compostos são produzidos por algumas espécies de dinoflagelados dos géneros Alexandrium, Gymnodinium e Pyrodinium, sendo uma família de toxinas com mais de 50 derivados de saxitoxina
(STX) (Anderson et al., 2012). Estes compostos bloqueiam a condução de impulsos nervosos em células neurais através da inibição de canais de sódio mediados por voltagem nas membranas destas células, o que levará à hiperpolarização membranar e resultar na paralisia de células musculares (Kao and Nishiyama, 1965; Ritchie and Rogart, 1977).
Figura 1. Vista esquemática da transferência de biotoxinas provenientes de “blooms” de algas nocivas ao longo da teia trófica.

4 | PORTUGALA
Vol.22 | Dezembro de 2018 | Semestral
As biotoxinas, toxinas produzidas por microalgas, são principalmente acumuladas pela base da teia trófica, por organismos planctónicos filtradores, peixes planctívoros e bivalves. Estas toxinas produzidas por estas microalgas podem ser transferidas para níveis mais elevados da teia trófica marinha, como os cefalópodes (figura 1).
Os cefalópodes ocupam uma posição importante em todos os ecossistemas onde ocorrem, uma vez que são o alimento de predadores de topo, mamíferos e aves marinhas (Brock, 1985; Clarke and Goodall, 1994; Croxall and Prince, 1996; Daneri et al., 2000; Pauly et al., 1998; Stilwell and Kohler, 1982). Por outro lado, na dieta dos cefalópodes constam moluscos, crustáceos e pequenos
peixes, podendo assim constituir um elo de ligação entre a acumulação por consumidores primários (e.g. bivalves), os predadores de topo, e até mesmo o Homem (Hanlon and Messenger, 1996; Nixon, 1985; Rodhouse and Nigmatullin, 1996).
O presente trabalho tem como objetivo sintetizar a informação existente acerca de cefalópodes
Figura 2. Ilustração esquemática das concentrações máximas de ácido domóico (µg AD g-1) encontradas em tecidos de polvo comum (Octopus vulgaris).
como vetores de biotoxinas, nomeadamente AD e PSTs (Lopes et al., 2013).
É reportado, em estudos prévios, que os cefalópodes acumulam AD nos seus tecidos, tendo sido observadas concentrações de AD desde 1.1 a 166.2 μg AD g−1 na glândula digestiva (GD, Figura 2) do polvo comum Octopus vulgaris na costa portuguesa (Costa et al., 2004). As concentrações mais elevadas são verificadas na GD uma vez que este é o principal tecido envolvido na absorção de nutrientes e digestão
intracelular em cefalópodes. Adicionalmente, o AD é também acumulado em outras espécies de cefalópodes, como o choco comum (Sepia officinalis) e duas outras espécies de polvo (Eledone cirrhosa, E. moschata). Mais uma vez, esta toxina acumula preferencialmente na glândula digestiva. É de relevar que a presença de AD nos tecidos destes cefalópodes se prolonga além do período de “bloom” da microalga produtora, sugerindo uma retenção ou incapacidade de eliminar esta neurotoxina.
No que se refere à acumulação de AD em lulas, existe informação na literatura que indica que estes cefalópodes também acumulam esta toxina nos seus tecidos, principalmente no sistema digestivo de Doryteuthis opalescens (Bargu et al., 2008) e Dosidicus gigas (Braid et al., 2012). Aliás, há décadas que ocorrem periodicamente arrojamentos de D. gigas, e foi verificado que em espécimes arrojados existiam baixas concentrações de AD, sugerindo que a contaminação com esta toxina seja uma possível causa dos arrojamentos, devido à transferência

PORTUGALA | 5
ao longo da teia trófica.
Relativamente às outras neurotoxinas em destaque, a literatura revela que a acumulação de PSTs ocorre em apenas 3 espécies de cefalópodes, O. vulgaris (Costa et al., 2009; Monteiro e Costa, 2011), D. gigas (Braid et al., 2012) e Octopus (Abdopus) sp.5 (Robertson et al., 2004). Similarmente à acumulação de AD, as PSTs acumulam preferencialmente na GD, com a exceção do polvo australiano Octopus (Abdopus) sp.5, onde foram verificadas PSTs, nomeadamente STX, nos braços. Estes estudos mostram uma panóplia de derivados de STX bastante variada, uma vez
que o perfil toxicológico encontrado nos cefalópodes depende da dieta e do produtor de PSTs na região de captura.
Assim, os cefalópodes são bons candidatos para monitorizar a ocorrência destas biotoxinas, uma vez que, em alguns casos, estas são eliminadas de forma lenta, podendo ser transferidas ao longo da teia trófica e, em último caso, constituir uma ameaça para o consumo humano. É de relevar que, relativamente à segurança alimentar, as biotoxinas focadas apenas acumulam nas vísceras, não constituindo um problema para
a saúde pública, com a exceção do polvo australiano, onde foram verificadas PSTs na parte edível (braços).
Em estudos futuros, será crucial compreender os mecanismos de defesa que estes invertebrados excecionais poderão possuir, que os permite acumular concentrações elevadas destas neurotoxinas sem efeitos negativos aparentes, assim como investigar se estas toxinas serão acumuladas no sistema nervoso central e que potenciais efeitos terão ao nível da cognição e aprendizagem destes moluscos excecionais.
Referências Bibliográficas:
Anderson, DM, Alpermann, TJ, Cembella, AD, Collos, Y, Masseret, E, Montresor, M. (2012). The globally distributed genus Alexandrium: multifaceted roles in marine ecosystems and impacts on human health. Harmful Algae, 14, 10-35.
Bargu, S, Powell, CL, Wang, Z, Doucette, GJ, Silver, MW. (2008) Note on the occurrence of Pseudo-nitzschia australis and domoic acid in squid from Monterey Bay, CA (USA). Harmful Algae, 7, 45–51.
Braid, H; Deeds, J; DeGrasse, S; Wilson, J; Os-borne, J; Hanner, R. (2012) Preying on com-mercial fisheries and accumulating paralytic shellfish toxins: a dietary analysis of invasive Dosidicus gigas (Cephalopoda Ommastrephi-dae) stranded in Pacific Canada. Marine Biolo-gy, 159, 25-31.
Brock, RE. (1985) Preliminary study of the fee-ding habits of pelagic fish around Hawaiian fish aggregation devices or can fish aggrega-tion devices enhance local fisheries producti-vity? Bulletin of Marine Science, 37, 40–49.
Clarke, M, Goodall, N. (1994) Cephalopods in the diets of three odontocete cetacean spe-cies stranded at Tierra del Fuego, Globicepha-la melaena (Traill, 1809), Hyperoodon plani-frons Flower, 1882 and Cephalorhynchus com-mersonii (Lacepede, 1804). Antarctic Science, 6, 149–154.
Costa, PR, Rosa, R, Sampayo, MAM. (2004). Tis-sue distribution of the amnesic shellfish toxin,
domoic acid, in Octopus vulgaris from the Por-tuguese coast. Marine Biology, 144, 971–976.
Costa, PR, Botelho, MJ, Rodrigues, SM. (2009) Accumulation of paralytic shellfish toxins in digestive gland of Octopus vulgaris during bloom events including the dinoflagellate Gymnodinium catenatum. Marine Pollution Bulletin, 58, 1747-1750.
Croxall, JP, Prince, PA. (1996). Cephalopods as Prey I: Seabirds. Philosophical Transactions of the Royal Society B, 351, 1023–1043.
Daneri, GA, Carlini, AR, Rodhouse, PGK. (2000) Cephalopod diet of the southern elephant seal, Mirounga leonina, at King George Island, South Shetland Islands. Antarctic Science, 12, 16–19.
Berman, FW, Murray, TF. (1997) Domoic acid neurotoxicity in cultured cerebellar granule neurons is mediated predominantly by NMDA receptors that are activated as a consequen-ce of excitatory amino acid release. Journal of Neurochemistry, 69, 693–703.
Hallegraeff, GM. (1993). A review of harmful al-gal blooms and their apparent global increa-se. Phycologia 32, 79–99.
Hallegraeff, GM. (2014) Harmful Algae and their Toxins: Progress, Paradoxes and Para-digm Shifts, in: Toxins and Biologically Active Compounds from Microalgae. CRC Press, Ros-sini, G.P. (ed), pp. 3–20
Hanlon, RT, Messenger, JB. (1996) Cephalopod Behaviour; Cambridge University Press: Cam-
bridge, UK, p. 232.
Kao, CY, Nishiyama, A. (1965) Actions of saxi-toxin on peripheral neuromuscular systems. Journal of Physiology, 180, 50–66.
Lopes, VM, Costa, PR, Rosa, R. (In press) Effects of Harmful Algal Bloom Toxins on Marine Or-ganisms. In Duarte, B., Caçador, I. (Eds.), Eco-toxicology of Marine Organisms, CRC Press, ISBN-13: 978-1138035492.
Lopes, VM, Lopes, AR, Costa, PR, Rosa, R. (2013) Cephalopods as vectors of Harmful Al-gal Bloom toxins in marine food webs. Marine Drugs 11, 3381–3409.
Monteiro, A, Costa, PR. (2011) Distribution and selective elimination of paralytic shellfish to-xins in different tissues of Octopus vulgaris. Harmful Algae, 10, 732-737.
Nixon, M. (1985) Capture of prey, diet and fee-ding of Sepia officinalis and Octopus vulgaris (Mollusca: Cephalopoda) from hatchling to adult. Vie et Milieu. 35, 255–261.
Pauly, D, Trites, AW, Capuli, E, Christensen, V. (1998) Diet composition and trophic levels of marine mammals. ICES Journal of Marine Science, 55, 467–481.
Ritchie, JM, Rogart, RB. (1977) The binding of saxitoxin and tetrodotoxin to excitable tis-sue. Reviews of Physiology, Biochemistry and Pharmacology, 79, 1–50.
Robertson, A, Stirling, D, Robillot, C, Llewellyn, L, Negri, A. (2004) First report of saxitoxin in octopi. Toxicon, 44, 765-771.

6 | PORTUGALA
Vol.22 | Dezembro de 2018 | Semestral
Desde o início da Era industrial, a concentração de dióxido de carbono (CO2) dissolvido na atmosfera tem vindo a aumentar. Os valores observados atualmente são mais elevados do que aqueles que a Terra experienciou nos último 800.000 anos, sendo esperado que esta tendência se mantenha, dada a dependência humana da energia obtida através dos combustíveis fósseis (Rhein et al. 2013).
Da mesma forma que o CO2 atmosférico tem vindo a aumentar, também o pH dos oceanos tem vindo a diminuir, pois é intrinsecamente dependente das trocas gasosas com a atmosfera (Rhein et al. 2013). O dióxido de carbono de origem antropogénica é apontado como o principal agente da acidificação dos oceanos (AO), designação dada ao processo de redução do pH gerado pelo aumento excessivo da pressão parcial de CO2 (pCO2) na superfície dos oceanos (Ciais et al. 2013; Rhein et al. 2013). Nos últimos séculos, a absorção de CO2 pelos oceanos levou a um aumento de 26% na concentração de iões de hidrogénio (H+) e consequente redução de 0.1 unidades de pH. É esperado que, no final deste século, o valor médio de pH da superfície do oceano decaia 0.2 a 0.4 unidades (Rhein et al. 2013). Este fenómeno está a demonstrar sérias consequências para a saúde dos ecossistemas mundiais (Cubasch et al. 2013), interferindo com a biogénese de estruturas calcárias e induzindo alterações fisiológicas, neurológicas e comportamentais nos organismos marinhos (McNeil & Sasse 2016).
Os cefalópodes são moluscos altamente adaptados ao ecossistema aquático, apresentado uma elevada plasticidade a alterações ambientais (Fiorito et al. 2015; Doubleday et al. 2016). Estão considerados entre
os invertebrados mais “avançados”, dada a presença de cérebros multilobulares relativamente grandes e altamente diferenciados, um sofisticado conjunto de órgãos sensoriais e locomoção com propulsão a jato (Fiorito et al. 2015). Além disso, a maioria dos cefalópodes recém-eclodidos assemelham-se a indivíduos adultos e contam ainda com um sistema nervoso central altamente desenvolvido, como é o caso do choco-comum (Boyle 1987).
O choco-comum, Sepia officinalis, é uma espécie nectobentónica (Reid et al. 2005; Guerra 2006) que realiza migrações sazonais para zonas de baixa profundidade, como os estuários, em época de reprodução (Reid et
ARTIGO
Impacto da acidificação dos oceanos no desenvolvimento e comportamento de chocos recém-eclodidos
Por Érica Moura1, Catarina P. Santos1,2, Marta Pimentel1, Eduardo Sampaio1,
Cláudia Pereira1, Maria Rita Pegado1,2, Vanessa Madeira Lopes1, Rui Rosa1
1MARE – Marine and Environmental Sciences Centre, Laboratório Marítimo da Guia, Faculdade de Ciências da Universidade de Lisboa, Av. Nossa Senhora do Cabo, 939, 2750-374 Cascais, Portugal.
2IGC - Instituto Gulbenkian de Ciência, R. Quinta Grande, 6, 2780-156 Oeiras, Portugal
Rodhouse, PG, Nigmatullin, CM. (1996). Role as consumers. Philosophical Transactions of the Royal Society B, 351, 1003–1022.
Rosa, R, Calado, R, Andrade, AM, Narciso, L, Nunes, ML. (2005) Changes in amino acids and lipids during embryogenesis of Euro-pean lobster, Homarus gammarus (Crustacea : Decapoda). Comparative Biochemistry and Physiology B-Biochemistry & Molecular Bio-
logy, 140, 241-249.
Rosa, R, Marques, AM, Nunes, ML, Bandarra, N, Reis, CS. (2004) Spatial-temporal changes in dimethyl acetal (octadecanal) levels of Octopus vulgaris (Mollusca, Cephalopoda): relation to feeding ecology. Scientia Marina, 68, 227-236.
Smayda, TJ. (1997) Harmful algal blooms:
Their ecophysiology and general relevance to phytoplankton blooms in the sea. Limno-logy and Oceanography, 42, 1137–1153.
Stillwell, CE, Kohler, N.E. (1982) Food, feeding habits, and estimates of daily ration of the shortfin mako (Isurus oxyrinchus) in the Nor-thwest Atlantic. Canadian Journal of Fishe-ries and Aquatic Sciences, 39, 407–414.
Figura 1 – À esquerda a localização da área de recolha dos ovos de Sepia officinalis (assinalada a vermelho) e à direita um exemplar com 20 dias pós-eclosão.
PORTUGALA | 7
al. 2005). Trata-se de um animal com maior atividade durante a noite, passando o dia camuflado no substrato (Boyle 1987; Reid et al. 2005), e dotado de um campo visual bastante abrangente (Mäthger et al. 2013), estando dependente desta capacidade e deste modo de vida para maximizar a sua sobrevivência, quer pela evasão a predadores (Chichery & Chanelet 1976; Boyle 1987; Reid et al. 2005), quer pela procura de alimento (Wells 1958; Messenger 1968). Esta espécie tem um estilo de vida ativo com elevadas taxas metabólicas e naturalmente sujeito a hipercapnia (elevados níveis de pCO2) durante o desenvolvimento embrionário (Melzner et al. 2009). Estas características, associadas à sua elevada tolerância a diferentes ambientes (Reid et al. 2005; Guerra 2006), podem funcionar como uma pré-adaptação quando expostos a condições de acidificação (Melzner et al. 2009). Apesar desta possível pré-adaptação, a AO tem
sido apontada como um fator de stress nos sistemas bentónicos, amplificando as condições de hipercapnia e de hipoxia, naturais desses ambientes (Melzner et al. 2013).
Contudo, os estudos existentes sobre o efeito da AO nestes organismos focam-se, sobretudo, no impacto relativamente à concha interna (Gutowska et al. 2008; Gutowska et al. 2010; Dorey et al. 2013; Sigwart et al. 2015), ao desenvolvimento embrionário (Lacoue-Labarthe et al. 2009; Hu et al. 2011; Dorey et al. 2013; Rosa et al. 2013; Sigwart et al. 2015) e ao desenvolvimento de indivíduos recém-eclodidos/juvenis (Gutowska et al. 2008; Lacoue-Labarthe et al. 2009; Hu et al. 2011; Dorey et al. 2013; Sigwart et al. 2015), sendo que, o efeito da AO relativamente a questões comportamentais é praticamente desconhecido. Como tal, este estudo teve como objetivo descobrir e esclarecer as consequências do aumento de pCO2, previsto para o final do século (Δ
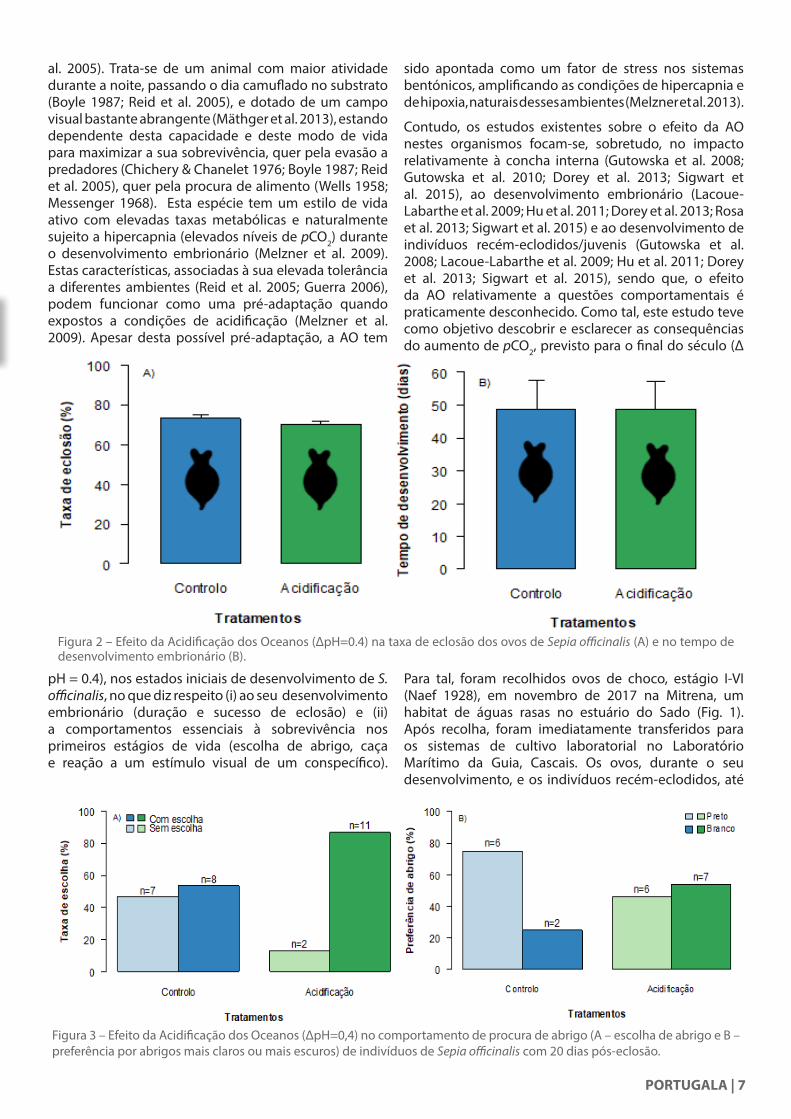
Figura 2 – Efeito da Acidificação dos Oceanos (∆pH=0.4) na taxa de eclosão dos ovos de Sepia officinalis (A) e no tempo de desenvolvimento embrionário (B).
pH = 0.4), nos estados iniciais de desenvolvimento de S. officinalis, no que diz respeito (i) ao seu desenvolvimento embrionário (duração e sucesso de eclosão) e (ii) a comportamentos essenciais à sobrevivência nos primeiros estágios de vida (escolha de abrigo, caça e reação a um estímulo visual de um conspecífico).
Para tal, foram recolhidos ovos de choco, estágio I-VI (Naef 1928), em novembro de 2017 na Mitrena, um habitat de águas rasas no estuário do Sado (Fig. 1). Após recolha, foram imediatamente transferidos para os sistemas de cultivo laboratorial no Laboratório Marítimo da Guia, Cascais. Os ovos, durante o seu desenvolvimento, e os indivíduos recém-eclodidos, até
Figura 3 – Efeito da Acidificação dos Oceanos (∆pH=0,4) no comportamento de procura de abrigo (A – escolha de abrigo e B – preferência por abrigos mais claros ou mais escuros) de indivíduos de Sepia officinalis com 20 dias pós-eclosão.
8 | PORTUGALA
Vol.22 | Dezembro de 2018 | Semestral
vinte dias pós-eclosão, foram mantidos em sistemas de suporte de vida independentes, com temperatura da água a 18 °C, salinidade a 36, fotoperíodo 12:12 h e sujeitos a um cenário de acidificação, obtido através da injeção de CO2, com valores estimados para o final do século (pH= 7,7, pCO2 ~ 1000 μatm), e a um cenário
de controlo, com valores atuais (pH = 8,1, pCO2 ~ 400 μatm). As condições físico-químicas da água foram monitorizadas diariamente e anotado o número de eclosões/mortalidades, para posterior cálculo da duração do desenvolvimento embrionário e sucesso de eclosão.
Os testes comportamentais foram realizados com
indivíduos entre quinze e vinte dias pós-eclosão e foi desenvolvido um desenho experimental apropriado às necessidades de cada teste. A recolha de dados foi feita através de gravações, sem interferência do observador, e os vídeos foram analisados a posteriori. Os testes comportamentais analisados focam-se em comportamentos essenciais à sobrevivência dos indivíduos, dada a dependência de alimento para sobreviverem e crescerem, abrigo para se poderem proteger dos predadores e reação a alarmes conspecíficos, como, por exemplo, a expulsão de tinta.
De acordo com os resultados obtidos em outros estudos com S. officinalis (Dorey et al. 2013; Rosa et al. 2013),
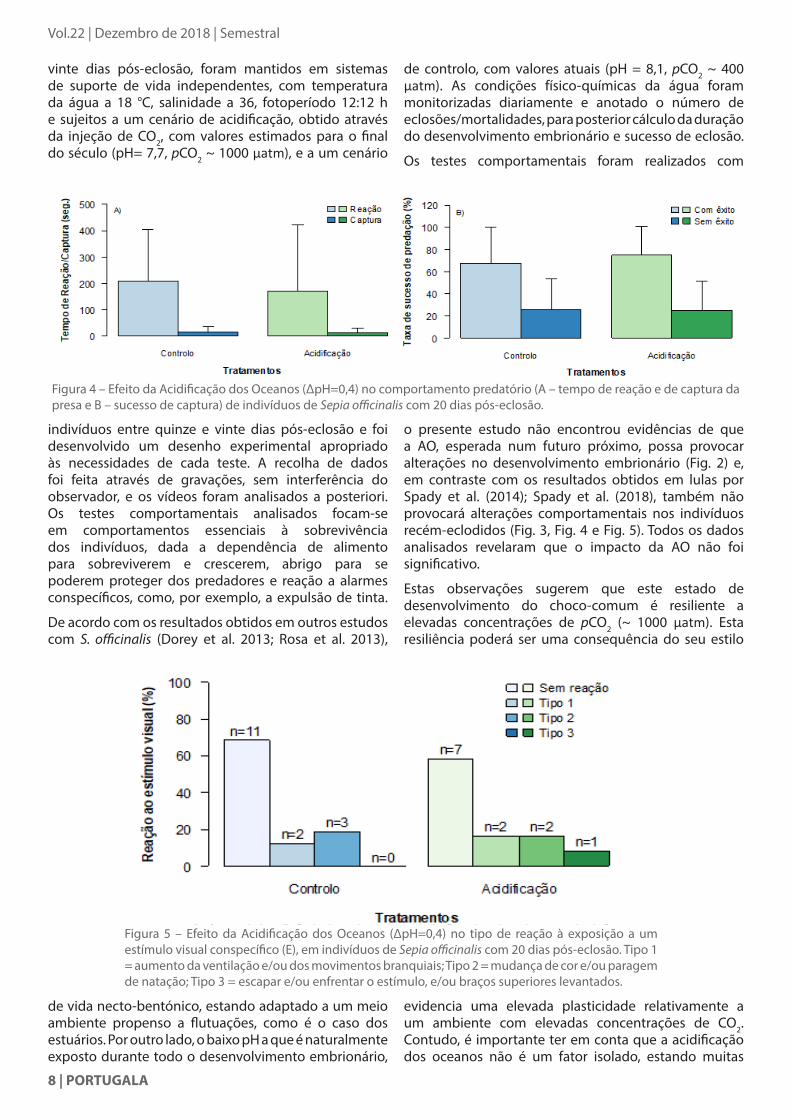
o presente estudo não encontrou evidências de que a AO, esperada num futuro próximo, possa provocar alterações no desenvolvimento embrionário (Fig. 2) e, em contraste com os resultados obtidos em lulas por Spady et al. (2014); Spady et al. (2018), também não provocará alterações comportamentais nos indivíduos recém-eclodidos (Fig. 3, Fig. 4 e Fig. 5). Todos os dados analisados revelaram que o impacto da AO não foi significativo.
Estas observações sugerem que este estado de desenvolvimento do choco-comum é resiliente a elevadas concentrações de pCO2 (~ 1000 μatm). Esta resiliência poderá ser uma consequência do seu estilo
Figura 4 – Efeito da Acidificação dos Oceanos (∆pH=0,4) no comportamento predatório (A – tempo de reação e de captura da presa e B – sucesso de captura) de indivíduos de Sepia officinalis com 20 dias pós-eclosão.
Figura 5 – Efeito da Acidificação dos Oceanos (∆pH=0,4) no tipo de reação à exposição a um estímulo visual conspecífico (E), em indivíduos de Sepia officinalis com 20 dias pós-eclosão. Tipo 1 = aumento da ventilação e/ou dos movimentos branquiais; Tipo 2 = mudança de cor e/ou paragem de natação; Tipo 3 = escapar e/ou enfrentar o estímulo, e/ou braços superiores levantados.
de vida necto-bentónico, estando adaptado a um meio ambiente propenso a flutuações, como é o caso dos estuários. Por outro lado, o baixo pH a que é naturalmente exposto durante todo o desenvolvimento embrionário,
evidencia uma elevada plasticidade relativamente a um ambiente com elevadas concentrações de CO2. Contudo, é importante ter em conta que a acidificação dos oceanos não é um fator isolado, estando muitas

PORTUGALA | 9
Referências Bibliográficas:
Boyle PR. (1987) Cephalopod life cycles. Volume II: Comparative reviews in Boyle PR, editor. Harcourt Brace Jovanovich, Pub-lishers, ACADEMIC PRESS.
Chichery R, Chanelet J. (1976) Motor and behavioural responses obtained by stimu-lation with chronic electrodes of the op-tic lobe of Sepia officinalis. Brain research, 105:525-532.
Ciais, P, Sabine, C, Bala, G, Bopp, L, Brovkin, V, Canadell, J, Chhabra, A, DeFries, R, Galloway, J, Heimann, M, Jones, C, Le Quéré, C, Myneni, RB, Piao, S, Thornton, P. (2013) Carbon and Other Biogeochemical Cycles in Stocker TF, Qin D, Plattner G-K, Tignor M, Allen SK, Bos-chung J, Nauels A, Xia Y, Bex V, and Midgley PM, (Eds.). Climate Change 2013: The Physi-cal Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cam-bridge, United Kingdom and New York, NY, USA.
Cubasch U, Wuebbles D, Chen D, Facchi-ni MC, Frame D, Mahowald N, Winther J-G. (2013) Introduction in Stocker TF, Qin D, Pla-ttner G-K, Tignor M, Allen SK, Boschung J, Nauels A, Xia Y, Bex V, and Midgley PM, (Eds.) Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergo-vernmental Panel on Climate Change. Cam-bridge University Press, Cambridge, United Kingdom and New York, NY, USA.
Dorey N, Melzner F, Martin S, Oberhänsli F, Teyssié J-L, Bustamante P, Gattuso J-P, La-coue-Labarthe T. (2013) Ocean acidification and temperature rise: effects on calcification during early development of the cuttlefish Sepia officinalis. Marine Biology, 160:2007-2022.
Doubleday ZA, Prowse TA, Arkhipkin A, Pier-ce GJ, Semmens J, Steer M, Leporati SC, Lou-renço S, Quetglas A, Sauer W. (2016) Global proliferation of cephalopods. Current Biolo-gy, 26:R406-R407.
Fiorito, G, Affuso, A, Basil, J, Cole, A, de Gi-rolamo, P, D’Angelo, L, Dickel, L, Gestal, C, Grasso, F, Kuba, M, Mark, F, Melillo, D, Osorio, D, Perkins, K, Ponte, G, Shashar, N, Smith, D, Smith, J, Andrews, PL. (2015) Guidelines for the Care and Welfare of Cephalopods in Re-search -A consensus based on an initiative
by CephRes, FELASA and the Boyd Group. Laboratory Animals, 49:1-90.
Guerra A. (2006) Ecology of Sepia officinalis. Vie et Milieu, 56:97-107.
Gutowska MA, Melzner F, Pörtner HO, Meier S. (2010) Cuttlebone calcification increases during exposure to elevated seawater pCO2 in the cephalopod Sepia officinalis. Marine Biology, 157:1653-1663.
Gutowska MA, Pörtner HO, Melzner F. (2008) Growth and calcification in the cephalo-pod Sepia officinalis under elevated seawa-ter pCO2. Marine Ecology Progress Series, 373:303-309.
Hu MY, Hwang P-P, Tseng Y-C. (2015) Recent advances in understanding trans-epithe-lial acid-base regulation and excretion me-chanisms in cephalopods. Tissue Barriers, 3:e1064196.
Hu MY, Tseng Y-C, Stumpp M, Gutowska MA, Kiko R, Lucassen M, Melzner F. (2011) Eleva-ted seawater PCO2 differentially affects bran-chial acid-base transporters over the course of development in the cephalopod Sepia officinalis. American Journal of Physiology--Regulatory, Integrative and Comparative Physiology, 300:R1100-R1114.
Lacoue-Labarthe T, Martin S, Oberhänsli F, Teyssié J-L, Markich S, Jeffree R, Bustamante P. (2009) Effects of increased pCO2 and tem-perature on trace element (Ag, Cd and Zn) bioaccumulation in the eggs of the common cuttlefish, Sepia officinalis. Biogeosciences, 6:2561-2573.
Mäthger LM, Hanlon RT, Håkansson J, Nilsson D-E. (2013) The W-shaped pupil in cuttlefish (Sepia officinalis): functions for improving horizontal vision. Vision Research, 83:19-24.
McNeil, B. I. and Sasse, T. P. (2016) Future ocean hypercapnia driven by anthropogenic amplification of the natural CO2 cycle. Natu-re, 529, 383.
Melzner F, Gutowska M, Langenbuch M, Du-pont S, Lucassen M, Thorndyke M, Bleich M, Pörtner H-O. (2009) Physiological basis for high CO2 tolerance in marine ectothermic animals: pre-adaptation through lifestyle and ontogeny? Biogeosciences, 6:2313-2331.
Melzner F, Thomsen J, Koeve W, Oschlies A, Gutowska MA, Bange HW, Hansen HP, Kört-zinger A. (2013) Future ocean acidification will be amplified by hypoxia in coastal habi-tats. Marine Biology, 160:1875-1888.
Messenger JB. (1968) The visual attack of the cuttlefish, Sepia officinalis. Animal Behaviour, 16:342-357.
Naef A. (1928) Die Cephalopoden. Embryo-logie. Die Fauna Flora Golf Neapel, 35:1-357.
Reid A, Jereb P, Roper CFE. (2005) Family sepiidae. Pages 57–152 in Jereb P, and Ro-per CFE, editors. Cephalopods of the world. An annotated and illustrated catalogue of species known to date. In: Chambered Nau-tiluses and Sepioids (Nautilidae, Sepiidae, Sepiolidae, Sepiadariidae, Idiosepiidae, and Spirulidae). FAO, Rome, Italy.
Rhein M, et al. (2013) Observations: Ocean In Stocker TF, Qin D, Plattner G-K, Tignor M, Allen SK, Boschung J, Nauels A, Xia Y, Bex V, and Midgley PM, (Eds). Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Re-port of the Intergovernmental Panel on Cli-mate Change. Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA.
Rosa, R, Trübenbach, K, Repolho, T, Pimentel, M, Faleiro, F, Boavida-Portugal, J, Baptista, M, Lopes, VM, Dionísio, G, Leal, MC, Calado, R, Pörtner, HO. (2013) Lower hypoxia thresholds of cuttlefish early life stages living in a warm acidified ocean. Proceedings of the Royal Society B: Biological Sciences, 280:20131695.
Sigwart JD, Lyons G, Fink A, Gutowska MA, Murray D, Melzner F, Houghton JD, Hu MY-a. (2015) Elevated pCO2 drives lower growth and yet increased calcification in the early life history of the cuttlefish Sepia officinalis (Mollusca: Cephalopoda). ICES Journal of Marine Science, 73:970-980.
Spady BL, Munday PL, Watson SA. (2018). Predatory strategies and behaviours in ce-phalopods are altered by elevated CO2. Glo-bal Change Biology, 24(6), 2585-2596.
Spady BL, Watson S-A, Chase TJ, Munday PL. (2014) Projected near-future CO2 levels in-crease activity and alter defensive behaviou-rs in the tropical squid Idiosepius pygmaeus. Biology open, 3:1063-1070.
Wells M. (1958) Factors affecting reactions to Mysis by newly hatched Sepia. Behaviour 13:96-111.
vezes associado a outros, como por exemplo o aumento da temperatura, diminuição da salinidade ou pressão antropogénica, sendo, por isso, necessária uma pesquisa
alargada, com base no efeito cumulativo de múltiplos fatores de stress, para prever, de modo mais preciso, as consequências do futuro das alterações climáticas nesta

10 | PORTUGALA
Vol.22 | Dezembro de 2018 | Semestral
Por Laura Sordo1, Domitília Matias1, Sandra Joaquim1, Margarete Matias1, and Miguel Gaspar1,2
ARTIGO
1 Instituto Português do Mar e da Atmosfera (IPMA), Avenida 5 de Outubro s/n, 8700-305 Olhão, Portugal
2 Centro de Ciências do Mar (CCMAR), Universidade do Algarve, Campus de Gambelas, 8005-139 Faro, Portugal
E-mail: [email protected] e [email protected]
Qual será o futuro das populações naturais de pé-de-burrinho (Chamelea gallina) e conquilha (Donax trunculus) num oceano
acidificado e aquecido?
A uma taxa sem precedentes na história geológica, a concentração de dióxido de carbono (CO2) atmosférico aumentou de 316 ppm em 1959 para mais de 400 ppm em 2016 (NOAA, 2018). Entre 2015 e 2016 registou-se o maior incremento da taxa de CO2 na história da humanidade (Betts et al., 2016) (Figura 1). O rápido aumento das emissões de CO2 de origem antropogénica tem contribuído para uma alteração do ambiente marinho a nível global. Como consequência da acidificação e aquecimento dos oceanos, processos críticos dos ecossistemas costeiros e oceânicos estão em risco de colapso, o que terá importantes repercussões ambientais e económicas (Talberth & Niemi, 2017).
Face ao aumento do CO2 atmosférico e consequente acidificação dos oceanos, os bancos naturais de bivalves poderão
Figura 1. Série temporal de CO2 atmosférico (linha vermelha), CO2 na água (linha verde) e pH da água (linha azul). Medições obtidas entre 1958 e 2018 na estação oceânica ALOHA no Oceano Pacífico Norte.
estar severamente ameaçados. A falta de consenso relativamente aos efeitos previsíveis da acidificação dos oceanos nos organismos calcificadores, tais como os bivalves, deve-se sobretudo à grande quantidade de factores envolvidos. No entanto, os ensaios levados a cabo até à presente data foram de curta duração, maioritariamente com indivíduos adultos, consideraram apenas um fator e grande parte foram dirigidos para o mexilhão e a ostra. Atualmente existe pouca informação sobre o efeito que as alterações climáticas podem exercer noutras espécies de bivalves com grande importância ecológica e económica, como sejam o pé-de-burrinho (Chamelea gallina) (e.g. Range et al., 2014; Matozzo et al., 2012 e 2013) e a conquilha (Donax trunculus) (e.g. Pereira et al., 2015).
A conquilha é uma espécie que habita praias arenosas, entre o nível inferior das marés e 10 metros de profundidade, distribuindo-se predominantemente no Mediterrâneo e no Atlântico. Por outro lado, o pé-de-burrinho habita fundos de areia e lodo entre 5 e 20 m de profundidade e distribui-se desde a Noruega, Ilhas Britânicas, Portugal e Marrocos, ocorrendo igualmente nos arquipélagos da Madeira e das Canárias, bem como no Mar Mediterrâneo e no Mar Adriático (Pereira et al., 2007).
Estudos realizados têm demonstrado que estas espécies são especialmente sensíveis ao aumento do CO2. Num ensaio de nove dias onde se testou o efeito de diferentes concentrações de CO2 em D. trunculus, Pereira et al. (2015) verificou que

PORTUGALA | 11
* Projecto VENUS. Estudo integrado dos bancos naturais de moluscos bivalves no Golfo de Cádis para a sua gestão sustentável e conservação dos habitats associados. Programa INTERREG V-A Espanha- Portugal, POCTEP 2014-2020 (Ref. 0139_VENUS_5_E - 2017-2019).
(https://www.project-venus.com/pt/?section=id1507193702434).
o aumento de CO2 tem um efeito negativo no desenvolvimento e sobrevivência larvar, em especial durante a fase de larva velígera D. Por outro lado, num ensaio mais longo (80 dias), onde se testou o efeito de diferentes concentrações de CO2, Range et al. (2014) constatou que a diminuição do pH provoca alterações na concha e afeta negativamente o crescimento em C. gallina. Também Matozzo et al. (2012) confirmou que após 7 dias de exposição a diferentes temperaturas e concentrações de CO2, C. gallina é mais vulnerável à acidificação do oceano que o mexilhão-do-mediterrâneo (Mytilus galloprovincialis).
O projecto de investigação “VENUS”* (“Estudo integral dos bancos naturais de moluscos bivalves no Golfo de Cádis para a sua gestão sustentável e conservação dos habitats associados” - Projecto INTERREG - Programa POCTEP - Cooperação Transfronteiriça Espanha-Portugal), tem como objetivo geral promover a gestão sustentável dos bancos de pé-de-burrinho e conquilha no Golfo de Cádis. Para tal, pretende-se contribuir para o melhoramento da conservação dos habitats associados através do estabelecimento de medidas de mitigação visando minimizar os potenciais impactes negativos dos fatores antropogénicos
Figura 2. Maternidade de moluscos bivalves nas instalações da Estação Experimental de Moluscicultura em Tavira (IPMA).
e ambientais que afetam a abundância daquelas espécies. De entre os trabalhos previstos, serão desenvolvidos ensaios em mesocosmos na Estação Experimental de Moluscicultura de Tavira do IPMA (Figura 2), com o objectivo de investigar os efeitos que a acidificação oceânica e o aquecimento global terão na reprodução, no desenvolvimento larvar e, consequentemente, no recrutamento de ambas as espécies (Figura 3). Para tal, será utilizado um sistema inovador de monitorização que permitirá controlar pequenas alterações de pH e temperatura, de modo a testar os cenários ambientais previstos pela IPCC
para os próximos 50 e 100 anos (IPCC IS92) (IPCC, 2000). Os resultados desta investigação permitirão prever e antecipar como as alterações climáticas afectarão a distribuição e abundância daqueles importantes recursos pesqueiros, permitindo avançar com medidas de mitigação perante os cenários previstos pela IPCC.
Figura 3. Juvenis de pé-de-burrinho (Chamelea gallina) produzidos na Estação Experimental de Moluscicultura de
Referências Bibliográficas:
Betts, RA, Jones, CD, Knight, JR, Keeling, RF; Kennedy, JJ. (2016) El Niño and a record CO2 rise. Nature Climate Change, 6: 806-810. DOI:
10.1038/nclimate3063
IPCC (2000). Emissions scenarios. Em: N. Nakićenović & R. Swart (Eds.), A Special Report of Working Group III of the Intergovernmental Pa-

12 | PORTUGALA
Vol.22 | Dezembro de 2018 | Semestral
ARTIGO
Coleção de Moluscos no Museu da Ciência da Universidade
de Coimbra: uma visão geral e perspetivas atuaisPor A. Breves1a, A.C. Rufino1, R. Paredes1,2,3, P.J.E. Casaleiro1
1Museu da Ciência, Universidade de Coimbra 2Grupo Processos Bióticos Mesozoicos, Universidade Complutense de Madrid, Espanha
3MARE-Marine and Environmental Sciences Centre, Universidade de CoimbraaE-mail: [email protected]
A coleção de moluscos sob a gestão do Museu da Ciência da Universidade de Coimbra (MCUC) é uma das mais importantes e significativas coleções do ponto de vista histórico e científico de Portugal. É uma das coleções de referência em Malacologia de Portugal e possivelmente é a maior coleção do género no país, com um acervo estimado em centenas de milhares de exemplares, oferecidos por naturalistas, investigadores e colaboradores desde o século XVIII até os dias atuais.
O início desta coleção remonta à criação do Gabinete de História Natural por Domingos Vandelli, a partir da reforma do ensino na Universidade de Coimbra pelo Marquês de Pombal em 1772 (Carreira et al, 2000). Ao longo dos anos, diversas coleções foram incorporadas, como as de Alexandre Rodrigues Ferreira, D. Pedro V, Barão de Castelo de Paiva, Jacinto da Silva Mengo, Augusto Pereira Nobre, António Augusto Carvalho Monteiro, Luis de Carvalho, Gumerzindo Henriques da Silva, entre outras (Carvalho, 1872; Ladeiro, 1936;
Carvalho, 1945). Mais recentemente, o MCUC recebeu uma volumosa coleção deixada em legado por Rolanda Maria Albuquerque de Matos, com dezenas de milhares de exemplares recolhidos pela própria, sendo esta, provavelmente, uma das mais significativas coleções de moluscos terrestres de Portugal Continental. Esta é a coleção de base utilizada para a publicação do Atlas dos Caracóis de Portugal Continental (Albuquerque de Matos, 2014).
A coleção de moluscos no MCUC inclui exemplares terrestres, marinhos e de água doce, sendo constituída por gastrópodes e bivalves, e também por grupos menos diversos, como poliplacóforos, cefalópodes e escafópodes. A maior parte da coleção encontra-se em reserva, enquanto uma pequena parte se encontra na área expositiva, nas diversas salas da Galeria de História Natural do MCUC. As conchas em reserva estão organizadas pelas seguintes coleções: Carvalho Monteiro, Geral e Portugal, constituídas por lotes
nel on Climate Change (pp. 570). Cambridge, UK and New York, NY: Cambridge University Press.
Matozzo, V, Chinellato, A, Munari, M, Finos, L, Bressan, M, Marin, MG. (2012) First evidence of immunomodulation in bivalves under seawa-ter acidification and increased temperature. PLoS ONE, 7:e33820. DOI:10.1371/journal.pone.0033820
Matozzo, V, Chinellato, A, Munari, M, Bressan, M, Marin, MG. (2013) Can the combination of decreased pH and increased temperature values induce oxidative stress in the clam Chamelea gallina and the mussel Mytilus galloprovincialis? Marine Pollution Bulletin, 72: 34-40. DOI: 10.1016/j.marpolbul.2013.05.004
NOAA (2018) Earth Systems Research laboratory, Hawaii (USA). Pági-na consultada o 23 de Outubro de 2018. ftp://aftp.cmdl.noaa.gov/products/trends/co2/co2_annmean_mlo.txt
Pereira, A, Palanco, I, Rufino, M, Moreno,O, Gaspar, MB. (2007) La pesca de bivalvos en el litoral oceánico de la costa sur-occidental de la Península Ibérica: Descripción de la pesquería y artes de pesca, medidas de gestión, biología de las especies comerciales y catálogo
de especies de bivalvos. Junta de Andalucía, Instituto de Investiga-ción y Formación agraria y Pesquera, Consejería de Innovación, Cien-cia y Empresa, Consejería de Agricultura y Pesca (141 pp.)
Pereira, AM, Range, P, Campoy, A, Oliveira, AP, Joaquim, S, Matias, D, Chícharo, L, Gaspar, MB. (2015) Larval hatching and development of the wedge shell (Donax trunculus L.) under increased CO2 in sou-thern Portugal. Regional Environmental Change, 16: 855–864. DOI: 10.1007/s10113-015-0803-4
Range, P, Chícharo, MA, Ben-Hamadou, R, Piló, D, Fernandez-Reiriz, MJ, Labarta, U, Marin, MG, Bressan, M, Matozzo, V, Chinellato, A, Mu-nari, M, El Menif, NT, Dellali, M, Chícharo, L. (2014) Impacts of CO2-in-duced seawater acidification on coastal Mediterranean bivalves and interactions with other climatic stressors. Regional Environmental Change, 14: 19-30. DOI: 10.1007/s10113-013-0478-7
Talberth, J, Niemi, E. (2017) Ocean acidification and warming: the economic toll and implications for the social cost of carbon. In: Elias SA et al., eds. Reference Module in Earth Systems and Environmental Sciences. Amsterdam: Elsevier, 1-19.

PORTUGALA | 13
distribuídos e organizados em armários metálicos (Figura 1). Predominantemente, os exemplares estão conservados a seco (conchas atuais e sub-fósseis), mas também existem vários exemplares conservados em meio líquido, em frascos de vidro.
Com o declínio da diversidade de organismos vivos em todo o mundo, a coleção de moluscos do MCUC tem uma grande importância pelos registos históricos de
ocorrências de espécies em locais onde neste momento já se encontram extintos ou em declínio acentuado, sendo esse um importante papel dos Museus de História Natural, de uma maneira geral (Shafter et al. 1998). Além disso, exemplares oriundos de expedições e coletas históricas feitas por naturalistas ou colecionadores de renome internacional fazem parte desta coleção, sendo estes de valor incalculável e insubstituíveis. Muito tem
Figura 1 - Coleção de moluscos no Museu da Ciência da Universidade de Coimbra em reserva e em exposição.
Figura 2 - Atividades didáticas realizadas com moluscos no Museu da Ciência da Universidade de Coimbra
sido também discutido sobre o papel social dos museus universitários, uma vez que as universidades detêm uma importante herança da humanidade (Lourenço, 2008). Assim, um outro considerável papel da coleção de Malacologia está relacionado ao seu caráter didático. Uma parte dos exemplares que são identificados para tal função, tem sido frequentemente utilizada em atividades educativas realizadas com crianças e adultos no MCUC (Figura 2).
Atualmente está a ser desenvolvido um trabalho essencial com as coleções, no âmbito dos projetos das
infraestruturas PORBIOTA - E-Infraestrutura Portuguesa de informação e investigação em Biodiversidade
e PRISC - Portuguese Research Infrastructure of Scientific Collections, visando a gestão de dados sobre a biodiversidade nacional e a conservação e acessibilidade das coleções científicas nos museus de Portugal, respetivamente. O acervo do MCUC está a ser inventariado e reacondicionado pelos elementos da equipa de curadoria e conservação do museu. Desde 2006 que está em curso o levantamento do número de exemplares, a obtenção dos dados de biometria, a

14 | PORTUGALA
Vol.22 | Dezembro de 2018 | Semestral
Por Rui Rosa11MARE – Marine and Environmental Sciences Centre, Laboratório Marítimo da Guia, Faculdade de Ciências da Universidade de Lisboa, Av. Nossa Senhora do Cabo, 939, 2750-374 Cascais, Portugal.
identificação do material tipo e toda a informatização do acervo, com a revisão taxonómica e o seu registo fotográfico. Isto permitirá aumentar a acessibilidade das coleções do MCUC a especialistas e ao público em geral,
através da possibilidade da sua consulta no museu digital (http://museudaciencia.inwebonline.net), assim como divulgar o importante património histórico-científico da Universidade de Coimbra e do país.
Referências Bibliográficas:
Albuquerque de Matos, RM. (2014). Atlas dos Caracóis de Portugal Continental. Edição da Autora, Lisboa. 258pp.
Carreira, I, Reis, JA, Baptista, MT, Ribeiro, Rui. (2000) Gabinete de história natural – revivências. Edição: Imprensa da Universidade de Coimbra 88pp.
Carvalho, JAS. (1872). Memória Histórica da Faculdade de Philosophia. Imprensa da Universidade. 344pp.
Carvalho, RN. (1945) Catálogo da Colecção de Invertebrados de Portugal existentes no Museu Zoológico de Coimbra. Mollusca
– Gastropoda. In: Memórias e Estudos do Museu Zoológico da Universidade de Coimbra. 167, 1-50.
Ladeiro, JM. (1936) Catálogo das Conchas Exóticas da Coleção António Augusto de Carvalho Monteiro. Memórias e Estudos do Museu Zoológico da Universidade de Coimbra. 88, 105-127.
Lourenço, MC. (2008) Where past, present and future knowledge meet: an overview of university museums and collections in Europe. Museologia Scientifica Memorie 2, 321-329.
Shafter, HB, Fisher, RN, Davidson, C. (1998) The role of natural history collections in documenting species declines. TREE 13 (1), 27-30.
Conferência Cephalopod International Advisory Council (CIAC) 2018, Florida, EUA - Breves notas
O Cephalopod International Advisory Council (CIAC) foi criado há 35 anos com o objetivo de facilitar a divulgação de informação e partilha de conhecimento sobre cefalópodes. Este ano, nomeadamente no passado mês de Novembro, o congresso trienal desde organismo realizou-se em S. Petersburgo, Florida, USA.
Este encontro iniciou-se com a realização de vários workshops, nomeadamente:
I. Workshop on genetic tools and live imaging in cephalopods
II. Paralarval and juvenile cephalopods: an updated identification guide
III. Hard structures of cephalopods and their application in your field of study
IV. The biogeochemical role of cephalopods in the world’s oceans
V. Cephalopod science: the direction of future research and the relevance of new policies
Cerca de 240 participantes (Figura 1) assistiram, durante 4 dias, a um total de 120 apresentações orais e mais de 100 comunicações em painel, sobre aspetos da biologia reprodutiva, fisiologia, taxonomia, pescas, genómica e ecologia destes moluscos. De especial relevo foram as apresentações orais feitas pelas investigadoras Alison Sweeney e Gretta Pecl (“keynote speakers”).
No último dia houve a atribuição de prémios para a melhor apresentação oral e comunicação em painel. O primeiro foi atribuído ao aluno de doutoramento Owen C. Nichols (título: “Offshore Influences on Inshore Squid: Linkages Between Water Mass Dynamics and
Figura 1 – Participantes da conferência CIAC 2018.

PORTUGALA | 15
Figura 2 - Foto durante o jantar do congresso CIAC 2018 – Aquário de St. Perterburgo, Florida, EUA (da esquerda para a direita: Angel González, Carlos Rosas, Roger Villanueva e Rui Rosa)
Doryteuthis pealeii Distribution)”, e o segundo ao aluno de mestrado Celso Cedillo-Robles (título: “Shallow water octopuses from off Mexico”). Ainda relativamente à atribuição de prémios, e com especial relevo, foram atribuídos os prémios de carreira ao Dr. Michael Vecchione, Dr. Marek Lipinski e Dr. B. Budelmann.
Este encontro foi sempre marcado pela boa disposição (Figura 2) e é com um grande prazer que informo que o próximo CIAC irá ser organizado por mim, em Lisboa, em 2021. Uma grande honra e um grande desafio pessoal!
Saudações malacológicas.
Rui Rosa
ARTIGO
Invasão à vista nos estuários do Tejo e Sado
Por Frederico B. Carvalho1, Sara Cabral1, José Lino Costa1,2, Miguel B. Gaspar3,4, Francisco Ruano3, Maria M. Angélico3, João Ramajal1 e Paula Chainho1,2
1 MARE – Centro de Ciências do Mar e do Ambiente2 Faculdade de Ciências da Universidade de Lisboa
3 IPMA – Instituto Português do Mar e da Atmosfera 4 CCMAR - Centro de Ciências do Mar
Uma espécie não indígena (ENI) pode ser definida como uma espécie que foi introduzida por ação humana, intencionalmente ou não, fora da sua área de distribuição natural (passada ou presente) e fora do seu potencial de dispersão natural (Chainho et al., 2015). As zonas costeiras e, mais particularmente, os estuários e as baías são dos ecossistemas marinhos mais vulneráveis ao risco de invasões biológicas, uma vez que são centros de atividade antropogénica (Ruiz et al., 1997). Consequentemente, nestas áreas ocorre a introdução de espécies através de vários vetores de introdução: (1) descarga das águas de lastro de embarcações, (2) dispersão através de canais de navegação, (3) incrustação nos cascos das embarcações, (4) repovoamento com espécies de interesse comercial ou para a pesca desportiva, (5) aquariofilia, (6) libertação de organismos vivos com fins alimentares, (7) libertação de isco vivo usado na pesca desportiva, (8) fuga de aquaculturas e (9) descarga de desperdícios vivos (Ruiz et al., 1997).
Apesar de Portugal ser, até muito recentemente, um dos países com menor número de ENI reportadas (Katsanevakis et al., 2013), isso deve-se sobretudo à ausência de estudos direcionados ao tema e de sistematização dos dados existentes numa listagem nacional (Chainho et al., 2015). Esse repositório de ENI tem vindo a ser feito ao longo da última década, o que
levou a um acréscimo exponencial de registos, que totalizou 166 ENI marinhas em Portugal em 2017 (ICES, 2018). Os estuários do Tejo e do Sado são especialmente suscetíveis à introdução de ENI, uma vez que, para além de serem os maiores estuários portugueses em termos de área total (França et al., 2009), estão localizados perto de grandes centros urbanos com portos comerciais, marinas de recreio e onde a pesca é praticada intensivamente e o setor da aquacultura está em crescimento (Duarte et al., 2014).
Apesar de haver diversos estudos sobre as ENI e os seus impactos, a informação existente acerca dos efeitos cumulativos das invasões biológicas na diversidade e abundância das comunidades é ainda escassa (Jimenez & Ruiz, 2016). Para além disso, a maioria dos estudos sobre a presença de ENI nos estuários foca-se principalmente em substratos duros, tais como estruturas artificiais em portos e marinas (Ruiz et al., 2009), mas as comunidades bentónicas de substratos móveis são raramente abordadas (Jimenez & Ruiz, 2016). Consequentemente, recentemente têm sido desenvolvidos projetos com o intuito de colmatar esta falta de conhecimento, nomeadamente nos estuários do Tejo (nos anos 2014 e 2015) e do Sado, (em 2015). Estes sistemas foram amostrados com recurso a uma ganchorra, tendo sido recolhidas amostras da comunidade bentónica subtidal
16 | PORTUGALA
Vol.22 | Dezembro de 2018 | Semestral
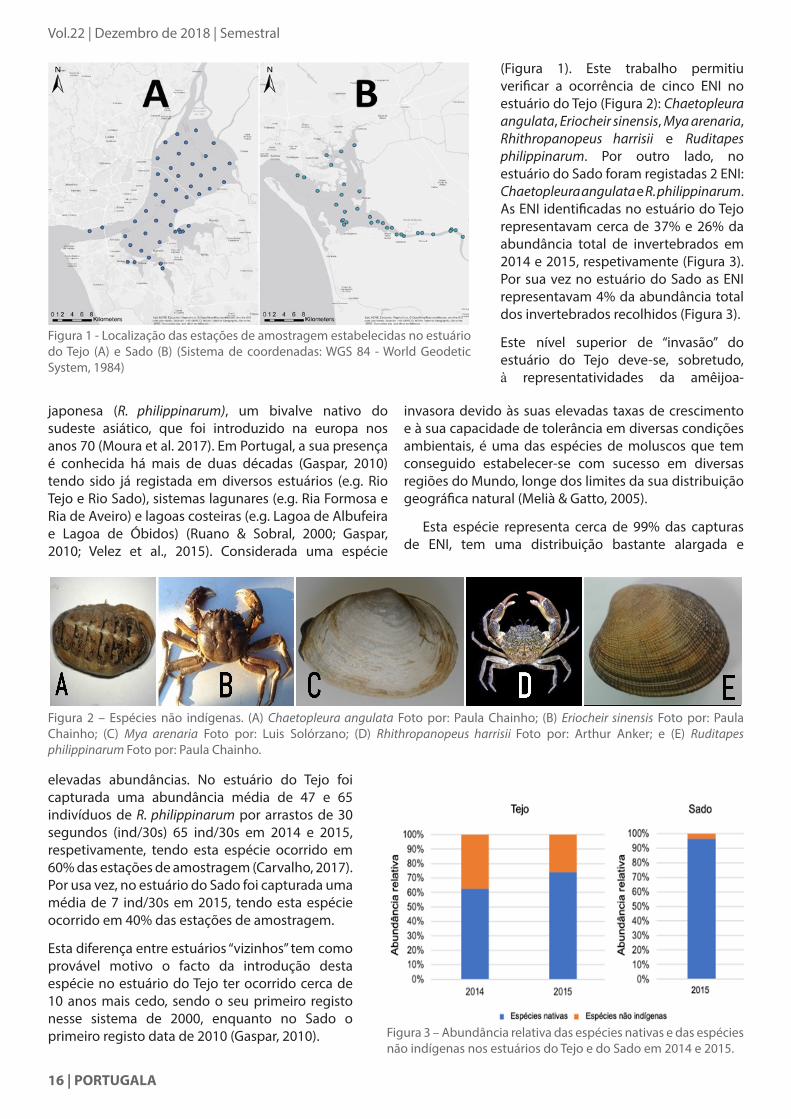
(Figura 1). Este trabalho permitiu verificar a ocorrência de cinco ENI no estuário do Tejo (Figura 2): Chaetopleura angulata, Eriocheir sinensis, Mya arenaria, Rhithropanopeus harrisii e Ruditapes philippinarum. Por outro lado, no estuário do Sado foram registadas 2 ENI: Chaetopleura angulata e R. philippinarum. As ENI identificadas no estuário do Tejo representavam cerca de 37% e 26% da abundância total de invertebrados em 2014 e 2015, respetivamente (Figura 3). Por sua vez no estuário do Sado as ENI representavam 4% da abundância total dos invertebrados recolhidos (Figura 3).
Este nível superior de “invasão” do estuário do Tejo deve-se, sobretudo, à representatividades da amêijoa-
Figura 1 - Localização das estações de amostragem estabelecidas no estuário do Tejo (A) e Sado (B) (Sistema de coordenadas: WGS 84 - World Geodetic System, 1984)
japonesa (R. philippinarum), um bivalve nativo do sudeste asiático, que foi introduzido na europa nos anos 70 (Moura et al. 2017). Em Portugal, a sua presença é conhecida há mais de duas décadas (Gaspar, 2010) tendo sido já registada em diversos estuários (e.g. Rio Tejo e Rio Sado), sistemas lagunares (e.g. Ria Formosa e Ria de Aveiro) e lagoas costeiras (e.g. Lagoa de Albufeira e Lagoa de Óbidos) (Ruano & Sobral, 2000; Gaspar, 2010; Velez et al., 2015). Considerada uma espécie
invasora devido às suas elevadas taxas de crescimento e à sua capacidade de tolerância em diversas condições ambientais, é uma das espécies de moluscos que tem conseguido estabelecer-se com sucesso em diversas regiões do Mundo, longe dos limites da sua distribuição geográfica natural (Melià & Gatto, 2005).
Esta espécie representa cerca de 99% das capturas de ENI, tem uma distribuição bastante alargada e
Figura 2 – Espécies não indígenas. (A) Chaetopleura angulata Foto por: Paula Chainho; (B) Eriocheir sinensis Foto por: Paula Chainho; (C) Mya arenaria Foto por: Luis Solórzano; (D) Rhithropanopeus harrisii Foto por: Arthur Anker; e (E) Ruditapes philippinarum Foto por: Paula Chainho.
elevadas abundâncias. No estuário do Tejo foi capturada uma abundância média de 47 e 65 indivíduos de R. philippinarum por arrastos de 30 segundos (ind/30s) 65 ind/30s em 2014 e 2015, respetivamente, tendo esta espécie ocorrido em 60% das estações de amostragem (Carvalho, 2017). Por usa vez, no estuário do Sado foi capturada uma média de 7 ind/30s em 2015, tendo esta espécie ocorrido em 40% das estações de amostragem.
Esta diferença entre estuários “vizinhos” tem como provável motivo o facto da introdução desta espécie no estuário do Tejo ter ocorrido cerca de 10 anos mais cedo, sendo o seu primeiro registo nesse sistema de 2000, enquanto no Sado o primeiro registo data de 2010 (Gaspar, 2010). Figura 3 – Abundância relativa das espécies nativas e das espécies
não indígenas nos estuários do Tejo e do Sado em 2014 e 2015.

PORTUGALA | 17
Referências Bibliográficas:
Carvalho, FB. (2017) Estado atual da população de amêijoa-japonesa (Ruditapes philippinarum) do estuário do Tejo e impactes da sua introdução. Dissertação de Mestrado, Faculdade de Lisboa de Universidade de Lisboa
Chainho, P, Fernandes, A, Amorim, A, Ávila, SP, Canning-Clode, J, Castro, JJ, Costa, AC, Costa, JL, Cruz, T, Gollasch, S, Grazziotin-Soares, C, Melo, R, Micael, J, Parente, MI, Semedo, J, Silva, T, Sobral, D, Sousa, M, Torres, P, Veloso, V, Costa, MJ. (2015) Non-indigenous species in Portuguese coastal areas, coastal lagoons, estuaries and islands. Estuarine Coastal Shelf Science 167, 199–211.
Duarte, B, Silva, G, Costa, JL, Medeiros, JP, Azeda, C, Sá, E, Metelo, I, Costa, MJ, Caçador, I. (2014) Heavy metal distribution and partitioning in the vicinity of the discharge areas of Lisbon drainage basins (Tagus Estuary, Portugal). Journal of Sea Research 93, 101–111.
França, S, Costa, MJ, Cabral, HN. (2009) Assessing habitat specific fish assemblages in estuaries along the Portuguese coast. Estuarine Coastal Shelf Science 83, 1–12.
Gaspar, M. (2010) Distribuição, abundância e estrutura demográfica da amêijoa-japonesa (Ruditapes philippinarum) no Rio Tejo. Relatório do IPIMAR, 1-6.
ICES (2018) Interim Report of the Working Group on Introductions and Transfers of Marine Organisms (WGITMO). Madeira, Portugal.
Jimenez, H, Ruiz, GM. (2016) Contribution of non-native species to soft-sediment marine community structure of San Francisco Bay, California. Biological Invasions 18, 2007–2016.
Katsanevakis, S, Gatto, F, Zenetos, A, Cardoso, AC. (2013) How many marine aliens in Europe? Manag. Biolological Invasions 4, 37–42.
Melià, P, Gatto, M. (2005) A stochastic bioeconomic model for the management of clam farming. Ecological Modelling 184, 163–174.
Moura, P, Garaulet, LL, Vasconcelos, P, Chainho, P, Costa, JL, Gaspar, MB. (2017) Age and growth of a highly successful invasive species: the Manila clam Ruditapes philippinarum (Adams & Reeve, 1850) in the Tagus Estuary (Portugal). Aquatic Invasions 12, 133-146.
Ruano, F, Sobral DV. (2000) Marine non-indigenous species current situation in Portugal Rodrigues, L, Reino, L, Godinho, LO, Freitas, H. (Eds.), Proceedings of the 1st Symposium on Non-indigenous Species: Introduction, Causes and Consequences. Liga Para a Protecção da Natureza, Lisbon, pp. 58-63.
Ruiz, GM, Carlton, JT, Grosholz, ED, Hines, AN. (1997) Global Invasions of Marine and Estuarine Habitats by Non-Indigenous Species: Mechanisms, Extent, and Consequences. American Zoology 37, 621–632.
Ruiz, GM, Freestone, AL, Fofonoff, PW, Simkanin, C. (2009) Habitat Distribution and Heterogeneity in Marine Invasion Dynamics: the Importance of Hard Substrate and Artificial Structure, in: Wahl, M. (Ed.), Marine Hard Bottom Communities. Springer-Verlag, Berlin, pp. 321–332.
Velez, C, Leandro, S, Figueira, E, Soares, AM, Freitas, R. (2015) Biochemical performance of native and introduced clam species living in sympatry: The role of elements accumulation and partitioning. Marine Environmental Research. 109, 81-94.
Contudo, é esperado que, não só o número de ENI aumente significativamente em ambos os estuários devido ao aumento da pressão associada aos diferentes vetores de introdução, mas também que haja um aumento da população de amêijoa-japonesa no estuário
do Sado, tendo em conta que esta espécie já se encontra plenamente estabelecida neste sistema, o que levará ao consecutivo aumento da representatividade de ENI no mesmo.
JORNADAS SOBRE A CONSERVAÇÃO DA LAPA FERRUGÍNEA (PATELLA
FERRUGINEA)
Por Joaquim Reis
Presidente do Instituto Português de Malacologia
Decorreram nos passados dias 4 a 6 de Outubro em Rodalquilar, Almería, Espanha, as jornadas sobre a conservação da “Lapa ferrugínea” (Patella ferrugínea), molusco marinho protegido pela União Europeia e em perigo de extinção em Espanha. O evento inclui-se no projeto Europeu “Life IP Intemares” para a gestão inovadora e participativa da Rede Natura 2000 no meio marinho, e organizado pela Consejería de Medio Ambiente y Ordenación del Territorio da Junta da Andaluzia, a Fundación Biodiversidad, o Ministerio para la Transición Ecológica, a Universidade Autónoma de Madrid e a Sociedade Espanhola de Malacologia.
O IPM foi convidado a participar nestas jornadas, que reuniram cerca de meia centena de participantes
dedicados à “lapa ferrugínea”, à malacologia em geral ou a questões de conservação e gestão do meio marinho. As comunicações versaram com maior destaque para questões da conservação desta espécie de molusco ameaçado, tendo também originado discussões muito relevantes sobre a conservação do meio marinho em geral.
No final das jornadas teve lugar a Assembleia geral da Sociedade Espanhola de Malacologia, onde entre os temas discutidos destacamos a atribuição do título de sócio honorário a Agustín Barrajón Mínguez, pelos seus contínuos esforços ao longo de 40 anos para a conservação do meio marinho e conhecimento da sua malacofauna, com especial destaque para as costas da

18 | PORTUGALA
Vol.22 | Dezembro de 2018 | Semestral
Andaluzia em Espanha, em colaboração com instituições como a Universidade Autónoma de Madrid, Universidade de Málaga, Junta da Andaluzia e Museo Nacional de Ciencias Naturales.
Por fim, os participantes tiveram oportunidade de visitar o Parque Natural de Cabo de Gata-Níjar, primeiro parque marítimo-terrestre em Espanha, designado reserva da biosfera e geoparque mundial pela UNESCO, e local onde ainda se pode encontrar a “lapa ferrugínea”, podendo apreciar a beleza paisagística e riqueza ecológica deste recanto.
Foi também discutido na Assembleia Geral a nomeação do molusco do
ano 2019, tendo sido proposto por Marcos Rubial o mexilhão Mytilus galloprovincialis, no contexto do projeto ECOS: “Novas ferramentas para avaliar o estado ecológico das costas rochosas e a sua relação com os serviços proporcionados pelos ecossistemas”, financiado pela FCT e desenvolvido em parceria entre o CIIMAR, a Universidade Autónoma de Madrid e a Universidade de Vigo. O projeto pretende usar o mexilhão como espécie modelo para a avaliação do estado ecológico do meio marinho, tendo como área geográfica de intervenção a Galiza e o norte de Portugal. Neste contexto, o IPM propôs associar-se à iniciativa, tornando o mexilhão Mytilus galloprovincialis molusco do ano Ibérico.
Wollastonaria, duas novas espécies recentes, W. jessicae e W. klausgrohi, e uma subespécie recente, W. jessicae monticola; são ainda descritas cinco novas espécies fósseis, H. microcarinata, W. beckmanni, W. falknerorum, W. ripkeni e W. inexpectata. São designados lectotipos ou neotipos para diversas espécies já conhecidas, há diversas mudanças de colocação de espécies em géneros e algumas subespécies são elevadas ao estatuto de espécies; e é ainda descrita a nova espécie fóssil Callina waldeni.
Em segundo lugar, aponta-se o artigo “Molecular phylogeny of Candidula (Geomitridae) land snails inferred from mitochondrial and nuclear markers reveals the polyphyly of the genus”, de Luis J. Chueca, Benjamín Juan Gómez-Moliner, Maria José Madeira & Markus Pfenninger, publicado em Molecular Phylogenetics and Evolution 118 (2018), 357-368. Neste trabalho, o grupo Candidula (Geomitridae) sofre uma realocação radical, sendo que muitas das espécies que ocorrem em Portugal passam para o género Xeroplexa.
Por fim, temos o artigo um pouco mais antigo “Species limits, interspecific hybridization and phylogeny in the cryptic land snail complex Pyramidula: the power of RADseq data”, da autoria de Oihana Razkin, Gontran Sonet, Karin Breugelmans, Maria José Madeira, Benjamín Juan Gómez-Moliner & Thierry Backeljau, publicado em Molecular Phylogenetics and Evolution 101 (2016), 267-278. Por este trabalho se verifica que enquanto até agora se considerava que a Pyramidula rupestris (Draparnaud, 1801) ocorria em Portugal, aparentemente a nossa
Foi recentemente publicada a descrição de duas novas espécies de Moluscos terrestres para a fauna portuguesa.
O artigo é da autoria de Geraldine & David Holyoak e de Rui Mendes. Os esforços desenvolvidos por estes nossos amigos para a inventariação da nossa fauna malacológica não marinha são já sobejamente conhecidos.
O artigo, da autoria de Geraldine & David Holyoak e de Rui Mendes, bem conhecidos pelos seus esforços de inventariação da nossa fauna malacológica terrestre, intitula-se “Descriptions of two new species of Belgrandia (Gastropoda: Hydrobiidae) from Central Portugal” e foi publicado no número 35(1) da conceituada revista Iberus. Nele se descrevem a Belgrandia alvaroi Holyoak, Holyoak & Mendes, 2017 e a B. jordaoi Holyoak, Holyoak & Mendes, 2017, ambas encontradas em cursos de água próximos de Alpedriz, no concelho de Alcobaça.
Outros trabalhos, relativos à fauna malacológica terrestre de Portugal, incluem os seguintes:
Em primeiro lugar o artigo “Revision of the genus-group Hystricella R. T. Lowe, 1855 from porto Santo (Madeira Archipelago), with descriptions of new recent and fossil taxa (Gastropoda, Helicoidea, Geomitridae)”, da autoria de Willy de Mattia, Marco T. Neiber & Klaus Groh, publicado em ZooKeys 732: 1-125 (24 Jan 2018). O trabalho pode ser consultado on line e utiliza caracteres conquiliológicos, anatómicos e genéticos para a proposta revisão. Nele é descrito o novo género
Por António Monteiro
NOTÍCIAS DO RECTÂNGULO

PORTUGALA | 19
espécie é mais exactamente a P. jaenensis (Clessin, 1882). Diz o povo que convém chamar os bois pelos nomes, claramente se aplicando o mesmo princípio às Pyramidula!
Quem visitar a magnífica vila de Sintra, a dois passos de Lisboa, terá oportunidade de admirar, perto da chamada Volta do Duche e do rio das Maçãs, pode ver-se uma estátua com o título “O Homem do Polvo”, da autoria do escultor Pedro Augusto Franco dos Anjos Teixeira (1908-1997).
Nascido em Paris, filho do também escultor Artur Gaspar dos Anjos Teixeira e de Ester de Oliveira Franco, Pedro Augusto veio para a zona Sintra com os pais quando contava seis anos, tendo a família ficado a residir em Mem Martins. Manteve sempre um íntimo contacto com a natureza, criando um grande apreço pela vida dos camponeses e pelos animais, reflectido em dois livros que escreveu, História dos Grilos, Amigos da Minha Infância e Memórias de um Grão de Trigo e também nas obras técnicas Anatomia Artística do Homem Comparada à dos Animais e Tecnologias da Escultura, assim como na temática escultórica “Saloios” e em muitas peças de temática humana e animal.
A partir dos dezasseis anos trabalhou num atelier em Lisboa, em colaboração com o seu pai, colaboração que durou até ao ano do falecimento deste, 1935. Nesse mesmo ano, Pedro dos Anjos Teixeira esculpe “Homem do Polvo”, obra considerada como uma das mais emblemáticas de sua autoria e com a qual dá início à sua carreira de escultor.
O jovem concluiu o curso superior de Escultura, na Escola de Belas Artes de Lisboa, em 1948. Expôs regularmente as suas obras, tendo recebido várias medalhas de honra e prémios. Frequentado o curso de Ciências Pedagógicas na Faculdade de Letras de Lisboa, na década de 1950, e
leccionou em escolas técnicas.
Aos 51 anos aceitou um lugar de docente na Academia de Belas-Artes da Madeira, mantendo-se por lá durante vinte e dois anos. No Funchal, foi fundador da Orquestra de Câmara da Madeira, onde também tocou; exerceu ainda actividade como jornalista no Jornal de Notícias da Madeira. Durante esse período contraiu matrimónio, pela terceira vez, com D. Maria Isabel Martins.
Foi no arquipélago da Madeira que produziu grande parte da sua obra escultórica, que se pode admirar em algumas das principais praças da região, como por exemplo as estátuas do Dr. Jaime Moniz (Funchal, 1961), de Tristão Vaz Teixeira (Machico, 1971), o busto de D. Filipa de Lencastre (Funchal, 1973), a Florista Madeirense (Funchal, 1980) e o Monumento às Bordadeiras (Funchal, 1986).
Regressou a Sintra na década de 1980, fixando residência na casa que é hoje em dia o Museu Anjos Teixeira; lá casou pela quarta vez, com D. Lídia das Neves Pacheco. Muitas das obras que produziu nesse período estão espalhadas pelo concelho, entre elas a estátua a D. Fernando II (Rotunda do Ramalhão), Monumento ao Trabalhador Rural (S. João das Lampas), bustos do Dr. Desidério Cambournac (Estefânea), do Dr. Nunes Claro, de Leal da Câmara (Rinchoa) e de Ferreira de Castro (Museu Ferreira de Castro) e, entre muitos outros trabalhos, o Monumento ao Professor Primário (Cacém).
Viria a falecer em Sintra, aos 88 anos, sem deixar descendentes. O seu legado inclui mais de 900 obras. Em 1981, a estátua “O Homem do Polvo” ou “Homem a Lutar com o Polvo” foi adquirida pela Câmara Municipal do Seixal e encontra-se hoje na Praça 1º de Maio, naquela localidade da margem sul do Tejo. A peça que se encontra em Sintra, junto ao Rio das Maçãs, é uma réplica ou um estudo, de menores dimensões.
O projecto The Cone Collector iniciou-se em meados do ano de 2006, por iniciativa do autor das presentes linhas, pretendendo contribuir para pôr em contacto todos aqueles – amadores ou profissionais – que se interessassem pela família Conidae de Moluscos Gastrópodes.
Com esse propósito, começou-se nesse ano a publicação de um boletim, em formato electrónico e distribuído gratuitamente a todos os assinantes. Rapidamente a

20 | PORTUGALA
Vol.22 | Dezembro de 2018 | Semestral
Por Vanessa Madeira Lopes
MARE – Marine and Environmental Sciences Centre, Laboratório Marítimo da Guia, Faculdade de Ciências da Universidade de Lisboa, Av. Nossa Senhora do Cabo, 939, 2750-374 Cascais, Portugal.
INTERNACIONAIS
BIVALVIA
Cao, R, Liu, Y, Wang, Q, Dong, Z, Yang, D, Liu, H, Ran, W, Qu, Y., Zhao, J. (2018). Seawater acidification aggravated cadmium toxicity in the oyster Crassostrea gigas: Metal bioaccumulation, subcellular distribution and multiple physiological responses. Science of the Total Environment, 642, 809–823.
Faggio, C, Tsarpali, V, Dailianis, S. (2018). Mussel digestive gland as a model tissue for assessing xenobiotics: An overview. Science of the Total Environment, 636, 220–229.
Ocharoen, Y, Boonphakdee, C, Boonphakdee, T, Shinn, AP, Moonmangmee, S. (2018). High levels of the endocrine disruptors bisphenol-A and 17β-estradiol detected in populations of green mussel, Perna viridis, cultured in the Gulf of Thailand. Aquaculture
497, 348–356.
Walker, DI, Younger, A, Stockley, L, Baker-Austin, C. (2018). Escherichia coli testing and enumeration in live bivalve shellfish – Present methods and future directions. Food Microbiology, 73, 29–38.
GASTROPODA
Fleming, AM, Dohner, MM, Phillips, NE, Ritchie, PA. (2018). Genetic connectivity among populations of two congeneric direct-developing whelks varies across spatial scales. New Zealand Journal of Marine and Freshwater Research , 52, 100–117.
Gérard, C, Hervé, M, Hechinger, RF. (2018). Long-term population fluctuations of the exotic New Zealand mudsnail Potamopyrgus antipodarum and its introduced aporocotylid trematode in northwestern France. Hydrobiologia 817, 253–266.
lista de distribuição atingiu perto de duas centenas de endereços, revelando-se assim a oportunidade da ideia, e o boletim passou a constituir um veículo para a troca de informações sobre o tema a que se destina, tendo sido publicados até agora trinta e quatro números.
Graças aos esforços de diversos colaboradores do projecto, rapidamente se alargou o seu âmbito, através da criação de um site na Internet, que começou a reunir um vasto leque de informações acerca da família Conidae e do seu estudo e colecção, até se converter numa fonte incontornável para todos os que se dedicam ao assunto. Presentemente, para além da colecção completa dos números do boletim. O site conta, entre outras, com as secções “What’s new” (onde são anualmente registados os novos taxa descritos para a família), “Checklist” (um importante registo organizado por Paul Kersten e onde se encontram informações sobre todas as espécies reconhecidas como válidas e também diversas suas variedades, todas adequadamente ilustradas), “Meetings” (onde se registam encontros entretanto realizados), “RKK” (onde está disponível o importante trabalho Manual of the Living Conidae, vol. 1 (1995), da autoria de Dieter Röckel, Werner Korn e Alan Kohn) e “Filmer” (que consiste no importante estudo de Mike Filmer sobre os exemplares tipo das diversas espécies descritas até 2013).
Interessa-nos sublinhar aqui os encontros internacionais
que, sob os auspícios de The Cone Collector, se têm realizado. O primeiro teve lugar em Estugarda (Alemanha) em 2010, seguindo-se-lhe La Rochelle (França, 2012), Madrid (Espanha, 2014) e Bruxelas (Bélgica, 2016).
O quinto encontro internacional está programado para Setembro de 2019 e terá lugar em Lisboa.
Para o efeito, contamos desde já com a colaboração do Museu Nacional de História Natural e da Ciência, que graciosamente aceitou colocar uma sala à nossa disposição para a realização do evento. Por esse motivo, agradecemos muito sinceramente à Direcção do Museu a generosidade com que acolheu o nosso pedido de participação e colaboração.
A exemplo dos anteriores, conta-se que este quinto encontro possa reunir um grande número de participantes, entre coleccionadores especializados na família Conidae e malacologistas profissionais que se dediquem ao estudo desta interessante família.
O programa está presentemente a ser preparado e será divulgado oportunamente. Tal como nos quatro encontros precedentes, esperamos que inclua uma panóplia de palestras, debates e workshops, versando os mais diversos aspectos do estudo dos Cones.
Como é natural, à medida que as informações forem surgindo, os leitores da Portugala não deixarão de ser postos a par de todos os desenvolvimentos.
Publicações em Destaque

PORTUGALA | 21
Rakauskas, V, Šidagytė, E, Butkus, R, Garbaras, A. (2018). Effect of the invasive New Zealand mud snail (Potamopyrgus antipodarum) on the littoral macroinvertebrate community in a temperate mesotrophic lake. Marine and Freshwater Research, 69, 155.
Wall-Palmer, D, Burridge, AK, Goetze, E, Stokvis, FR, Janssen, AW, Mekkes, L, Moreno-Alcántara, M, Bednaršek, N, Schiøtte, T, Sørensen, MV, Smart, CW, Peijnenburg, KTCA. (2018). Biogeography and genetic diversity of the atlantid heteropods. Progress in Oceanography, 160, 1–25.
CEPHALOPODA
Gong, Y, Chen, X, Li, Y, Fang, Z. (2017). Geographic variations of jumbo squid (Dosidicus gigas) based on gladius morphology. Fishery Bulletin, 116, 50–59.
Hoffmann, R, Lemanis, RE, Falkenberg, J, Schneider, S, Wesendonk, H, Zachow, S. (2018). Integrating 2D and 3D shell morphology to disentangle the palaeobiology of ammonoids: a virtual approach. Palaeontology 61, 89–104.
Laptikhovsky, V, Nikolaeva, S, Rogov, M. (2018). Cephalopod embryonic shells as a tool to reconstruct reproductive strategies in extinct taxa. Biological Reviews, 93, 270–283.
Spady, BL, Munday, PL, Watson, S-A. (2018). Predatory strategies and behaviours in cephalopods are altered by elevated CO2. Global Change Biology, 24, 2585–2596.
OUTROS
Armstrong, R, Riaz, S, Hasan, S, Iqbal, F, Rice, T., Syed, N. (2018). Mechanisms of Anesthetic Action and Neurotoxicity: Lessons from Molluscs. Frontiers in Physiology, 8, 1138.
Naji, A, Nuri, M, Vethaak, AD. (2018). Microplastics contamination in molluscs from the northern part of the Persian Gulf. Environmental Pollution, 235, 113–120.
NACIONAIS
BIVALVIA
Andrade, M, Soares, A, Figueira, E, Freitas, R. (2018). Biochemical changes in mussels submitted to different time periods of air exposure. Environmental Science and Pollution Research, 25, 8903–8913.
De Marchi, L, Neto, V, Pretti, C, Figueira, E, Chiellini, F, Morelli, A, Soares, AMVM, Freitas, R. (2018). Toxic effects of multi-walled carbon nanotubes on bivalves: Comparison between functionalized and nonfunctionalized nanoparticles. Science of The Total Environment, 622–623, 1532–1542.
Ilarri, MI, Amorim, L, Souza, AT, Sousa, R. (2018). Physical legacy of freshwater bivalves: Effects of habitat complexity on the taxonomical and functional diversity of invertebrates. Science of The Total Environment, 634, 1398–1405.
Vasconcelos, P, Moura, P, Pereira, F, Pereira, AM, Gaspar, MB. (2018). Morphometric relationships and relative growth of 20 uncommon bivalve species from the Algarve coast (southern Portugal). Journal of the Marine Biological Association of the United Kingdom, 98, 463–474.
GASTROPODA
Gomes, N.G., Fernandes, F., Carvalho, A., M Gomes, N.G., Madureira-Carvalho, Á., Valentão, P., Lobo-da-Cunha, A., Calado, G., Andrade, P.B., n.d. Profiling of Heterobranchia Sea Slugs from Portuguese Coastal Waters as Producers of Anti-Cancer and Anti-Inflammatory Agents. Molecules, 23(5), 1027.
Lobo-da-Cunha, A., Alves, Â., Oliveira, E., Malaquias, M.A.E., 2018. Histological, histochemical, and ultrastructural investigation of the male copulatory apparatus of Haminoea navicula (Gastropoda, Cephalaspidea). Journal of Morphology, 279, 554–565.
Madeira, C., Mendonça, V., Flores, A.A.V., Diniz, M.S., Vinagre, C., 2018. High thermal tolerance does not protect from chronic warming – A multiple end-point approach using a tropical gastropod, Stramonita haemastoma. Ecological Indicators, 91, 626–635.
Urushitani, H., Katsu, Y., Kagechika, H., Sousa, A.C.A., Barroso, C.M., Ohta, Y., Shiraishi, H., Iguchi, T., Horiguchi, T., 2018. Characterization and comparison of transcriptional activities of the retinoid X receptors by various organotin compounds in three prosobranch gastropods; Thais clavigera, Nucella lapillus and Babylonia japonica. Aquatic Toxicology, 199, 103–115.
CEPHALOPODA
Barroso-Barcenilla, F, Antunes, MT, Brandão, JM, Callapez, PM, Santos, VF, Segura, M. (2018). The d’Orbigny Palaeontological Collection of the National Museum of Natural History and Science, Lisbon, Portugal: Historical perspective and revision of Cretaceous Cephalopoda. Geodiversitas 40, 505–519.
Olmos-Pérez, L, Pierce, GJ, Roura, Á, González, ÁF. (2018). Barcoding and morphometry to identify and assess genetic population differentiation and size variability in loliginid squid paralarvae from NE Atlantic (Spain). Marine Biology, 165, 136.
Xavier, JC, Cherel, Y, Ceia, FR, Queirós, JP, Guimarães, B, Rosa, R, Cunningham, DM, Moors, PJ, Thompson, DR. (2018). Eastern rockhopper penguins Eudyptes filholi as biological samplers of juvenile and sub-adult cephalopods around Campbell Island, New Zealand. Polar Biology, 41, 1937–1949.
OUTROS
Cyrne, R, Rosa, IC, Faleiro, F, Dionísio, G, Baptista, M, Couto, A, Pola, M, Rosa, R., 2018. Nudibranchs out of water: long-term temporal variations in the abundance of two Dendrodoris species under emersion. Helgoland Marine Research, 72, 14.
Dell’Angelo, B, Landau, BM, Van Dingenen, F, Ceulemans, L., 2018. The upper Miocene chitons of northwest France (Mollusca: Polyplacophora). Zootaxa 4447, 1.
Veiga, P, Torres, AC, Besteiro, C, Rubal, M., 2018. Mollusc assemblages associated with invasive and native Sargassum species. Continental Shelf Research, 161, 12–19.

22 | PORTUGALA
Vol.22 | Dezembro de 2018 | Semestral
Instituto Português de MalacologiaZoomarine E.N. 125, Km 65 Guia 8201-864 Albufeira PORTUGAL Tel: 289 560 300 | Fax: 289 560 309 | Email: ipmalac @gmail
PROPOSTA DE SÓCIO
Nome Completo:
Data de Nascimento: (dd/mm/aa)
Nº BI/Cartão de Cidadão:
Morada:
Código Postal: - Localidade:
Correio electrónico: Telefone:
Habilitações: Profissão:
Os dados recolhidos destinam-se exclusivamente à gestão interna do I.P.M.. Poderão ser cedidos a outras instituições congéneres, nacionais ou internacionais para efeitos de divulgação de actividades, apenas com o consentimento expresso do proponente.
Autorizo/Não autorizo (riscar o que não interessa) a divulgação dos meus dados pessoais para os fins acima mencionados.
Assinatura do Proponente:
QUOTA 2015:
Sócio efectivo - 20 €
Sócio aderente individual - 20 €
Sócio aderente colectivo - 50 €
Sócio estudante - 10 € (anexar comprovativo de situação de estudante)
Formas de pagamento: Numerário; cheque ou vale postal em nome do Instituto Português de Malacologia; Transferência bancária para o NIB 003507720000248323096, do banco Caixa Geral de Depósitos com envio de comprovativo por email.
Agora também já o pode fazer através do preenchimento do formulário no nosso site:www.ipmalac.pt
/ /

PORTUGALA | 23
PORTUGAL ABoletim do Instituto Português de Malacologia
ISSN: 1645-9822Depósito Legal: 210446/04
Morada: Instituto Português de Malacologia (IPM), ZooMarine E.N. 125, KM65 Guia, 8201-864 AlbufeiraTelefone: 289 560 300Email: ipmalac @gmailSite IPM: http: www.ipmalac.pt
Corpo Editorial: Rui Rosa (Editor) MARE - Marine Environmental Sciences Centre [email protected]
Vanessa Madeira Lopes(Subeditora) MARE - Marine Environmental Sciences Centre [email protected]
Joaquim Reis (Presidente IPM) Presidente do Instituto Português de Malacologia [email protected]
António Frias Martins Departamento de Biologia da Universidade dos Açores [email protected]
António Marques Divisão de Aquacultura e Valorização, Instituto Português do Mar e da Atmosfera [email protected]
Diana Carvalho Museu Nacional de História Natural e da Ciência [email protected]
Dinarte Freitas Teixeira Secretaria Regional do Ambiente e dos Recursos Naturais da Madeira, Direcção Regional do Ambiente [email protected]
Domitília Matias Divisão de Aquacultura e Valorização, Instituto Português do Mar e da Atmosfera [email protected]
Miguel Nogueira Baptista MARE - Marine Environmental Sciences Centre [email protected]
Ronaldo Sousa CIIMAR/CIMAR e Departamento de Biologia da Universidade do Minho [email protected]
Autores neste volume:André Breves | António Monteiro | Catarina Santos | Cláudia Pereira | Cristina Rufino | Domitília Matias | Eduardo Sampaio | Érica Moura | Francisco Ruano | Frederico B. Carvalho | João Ramajal | Joaquim Reis | José Lino Costa
| Laura Sordo | Maragarete Matias | Maria M. Angélico | Maria Rita Pegado | Marta Pimentel | Miguel Gaspar | Paula Chainho | Pedro Casaleiro | Pedro R. Costa | Ricardo Paredes | Rui Rosa | Sandra Joaquim | Sara Cabral |
Vanessa Madeira
Foto Capa: edmondlafoto

24 | PORTUGALA
Vol.22 | Dezembro de 2018 | Semestral





























