Embed Size (px)
Citation preview

1
“PKC-mediated phosphorylation of BCL11B at Serine 2 negatively regulates its 1
interaction with NuRD complexes during CD4+ T cell activation” 2
3
Marion Dubuissez1, £, Ingrid Loison1, Sonia Paget1, Han Vorng2, Saliha Ait-Yahia2, 4
Olivier Rohr3,4, Anne Tsicopoulos2 and Dominique Leprince1,* 5
6
(1) Univ. Lille, CNRS, Institut Pasteur de Lille, UMR 8161 - M3T - Mechanisms of 7
Tumorigenesis and Targeted Therapies, F-59000 Lille 8
(2) Univ. Lille, CNRS, Inserm, CHU Lille, Institut Pasteur de Lille, U1019 - UMR8204 - 9
CIIL - Center for Infection and Immunity of Lille, F-59000 Lille, 10
(3) University of Strasbourg, IUT Louis Pasteur, EA7292, Dynamic of Host Pathogen 11
interactions, Institut of Parasitology and Tropical pathology, F-67000 Strasbourg France (4) 12
Institut Universitaire de France, Paris, France 13
* Corresponding author. Dominique Leprince, Institut de Biologie de Lille, CNRS UMR8161 14
1, rue du Pr. Calmette, CS 50447, 59021 Lille Cedex, France. 15
E mail : [email protected]; Phone : +33 3 20 87 11 19. Fax : +33 3 20 87 11 11 16
£ Present address: Maisonneuve-Rosemont Hospital Research Center, Maisonneuve-17
Rosemont Hospital, 5415 boulevard l'Assomption Montreal, QC H1T 3W5, Canada; 18
19
Key Words: BCL11B, NuRD chromatin remodeling complexes, PKC, CD4+ T cell 20
activation, Phosphorylation, SUMOylation, IL2, Id2 21
22
Running title: PKC switches BCL11B to an activator in CD4+ T cells 23
24
Number of characters (excluding spaces): 38878. 25
26
MCB Accepted Manuscript Posted Online 9 May 2016Mol. Cell. Biol. doi:10.1128/MCB.00062-16Copyright © 2016, American Society for Microbiology. All Rights Reserved.
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

2
Abstract 27
28
The transcription factor BCL11B/CTIP2 is a major regulatory protein implicated in various 29
aspects of development, function and survival of T cells. MAPK-mediated phosphorylation 30
and SUMOylation modulate BCL11B transcriptional activity, switching it from a repressor in 31
naïve murine thymocytes to a transcriptional activator in activated thymocytes. Here, we 32
show that BCL11B interacts via its conserved N-terminal MSRRKQ motif with endogenous 33
MTA1 and MTA3 proteins to recruit various NuRD complexes. Furthermore, we demonstrate 34
that PKC-mediated phosphorylation of BCL11B Ser2 does not significantly impact on 35
BCL11B SUMOylation but negatively regulates NuRD recruitment by dampening the 36
interaction with MTA1/3 and RbAp46 proteins. We detected increased phosphorylation of 37
BCL11B Ser2 upon in vivo activation of transformed and primary human CD4+ T cells. We 38
show that following activation of CD4+ T cells, BCL11B still binds to IL2 and Id2 promoters 39
but activates their transcription by recruiting P300 instead of MTA1. Prolonged stimulation 40
results in the direct transcriptional repression of BCL11B by KLF4. 41
Our results unveil Ser2 phosphorylation as a new BCL11B post-translational modification 42
linking PKC signalling pathway to TCR activation and define a simple model for the 43
functional switch of BCL11B from a transcriptional repressor to an activator during TCR 44
activation of human CD4+ T cells. 45
46
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

3
INTRODUCTION 47
48
Post translational modifications (PTMs) of transcription regulatory proteins allow integrating 49
various signalling and environmental cues into highly dynamic and controlled responses, 50
thereby achieving coordinated gene expression programs essential for cell proliferation or 51
differentiation. 52
The transcription factor BCL11B/CTIP2 was independently isolated as an interacting partner 53
of COUP-TF in neurons and as a tumour suppressor gene in mouse models of γ-ray induced 54
thymic lymphomas (1-3). Besides its expression in the CNS, BCL11B was shown to be widely 55
expressed in all T-cell subsets, starting from the DN2 stage and to be involved in various 56
aspects of development, function and survival of T-cells (4). Indeed, BCL11B is a focal point 57
essential for several checkpoints involved in T-cell commitment in early progenitors, 58
selection at the DN2 stage and differentiation of peripheral T cells (5-9). Furthermore, 59
monoallelic BCL11B deletions or missense mutations have been identified in the major 60
molecular subtypes of T-cell acute lymphoblastic leukaemia (10). Therefore, these 61
observations together with the occurrence of deletions and mutations in γ-ray induced 62
thymomas in mice identify BCL11B as a haploinsufficient tumor suppressor gene (11). 63
BCL11B is essential for T-cell development and is considered as a “guardian of T cell fate” 64
(12). Its closely related paralog BCL11A is essential for normal lymphopoiesis and 65
haemoglobin switching during erythroid differentiation (13-15). These two transcription 66
factors appear thus as key regulators of fundamental differentiation programs during normal 67
hematopoiesis. 68
BCL11B represses transcription of its targets genes through interaction with several 69
chromatin remodelling complexes and notably recruits NuRD complexes via interaction with 70
MTA1 and MTA2 (4, 11, 16-18). Albeit, originally characterized as a sequence-specific 71
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

4
transcriptional repressor, BCL11B also behaves as a context-dependent transcriptional 72
activator of the IL2 and Cot kinase genes in CD4+ T-cell activation (19, 20). 73
This dual behaviour of BCL11B as a transcriptional repressor and activator is not fully 74
understood but clearly rely on a dynamic cross-talk between BCL11B PTMs. Indeed, mass 75
spectrometry analyses of thymocytes isolated from 4-8 week-old mice and stimulated with a 76
mixture of phorbol ester and calcium ionophore used as an in vitro model mimicking TCR 77
activation identified several MAPK phosphorylation sites of Bcl11b and confirmed its 78
SUMOylation on Lysine 679 (21). These phosphorylation events then initiate a rapid and 79
complex cycle of Bcl11b PTMs including deSUMOylation, rephosphorylation and 80
reSUMOylation allowing recruitment of the transcriptional coactivator P300 to activate Id2 81
transcription (21, 22). 82
Here, we found that BCL11B interacts with the three MTA family members through its 83
conserved N-terminal MSRRKQ motif which is embedded in a potential PKC 84
phosphorylation consensus site. We demonstrated that a S2D phosphomimetic point mutation 85
is sufficient to abolish the interaction of BCL11B with all MTA corepressors and hence with a 86
wide range of NuRD complexes. Through generation of phosphospecific antibodies, we 87
identified in vivo Serine 2 phosphorylation of endogenous BCL11B proteins. We found that 88
activation of transformed Jurkat or primary human CD4+ T-cells results in a rapid and 89
transient PKC-induced phosphorylation of this BCL11B Ser2 culminating at 30 minutes of 90
treatment. In contrast with the MAPK-induced phosphorylations in late T-cell development, 91
this PKC phosphorylation peak precedes and does not impact on the SUMOylation peak 92
during activation of CD4+ T-cells. After prolonged activation (5 hours), the decrease of 93
BCL11B protein levels observed is due to the direct transcriptional repression of BCL11B by 94
KLF4. As shown by co-immunoprecipitation of endogenous proteins and chromatin 95
immunoprecipitation experiments, this BCL11B Ser2 phosphorylation through a decreased 96
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

5
interaction with MTA1 and a concomitant increased interaction with P300 participates to a 97
strong transcriptional up-regulation of Id2 and IL2, two BCL11B direct target genes, during 98
CD4+ T cell activation. A PKC inhibitor, Gö6983 abolishes this co-repressor/co-activator 99
switch in Jurkat cells. Thus, our findings identify BCL11B Ser2 phosphorylation as a new 100
mandatory step in the interconnected post-translational modifications converting BCL11B 101
from a transcriptional repressor to an activator and provide compelling evidence that together 102
with SUMOylation this PKC-mediated phosphorylation is essential for transcriptional 103
activation of IL2 during human CD4+ T-cell activation. 104
105
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

6
MATERIALS AND METHODS 106
107
Cell culture 108
HEK293T cells were maintained in DMEM (Gibco) supplemented with 10% FCS, non 109
essential amino acids and gentamycin. Jurkat and MOLT4 cells were maintained in RPMI 110
1640 (Gibco) supplemented with 10% FCS, non essential amino acids and 111
penicillin/streptomycin. 112
113
Isolation and activation of human T CD4+ lymphocytes. 114
Donor blood was provided by Etablissement Français du Sang - Nord de France (EFS), in 115
agreement with the official ethics statement between EFS and the Centre National de la 116
Recherche Scientifique (CNRS) - Délégation Nord Pas-de-Calais et Picardie. The study was 117
approved by the Institut de Biologie de Lille (CNRS) and EFS Institutional Review Board, 118
and informed consent was obtained in writing for each donor. Isolation of human CD4+ T 119
cells was performed as previously described (23). Briefly, human peripheral blood 120
mononuclear cells (PBMC) were isolated by Ficoll Hypaque density gradient centrifugation 121
(Amersham Pharmacia Biotech, Uppsala, Sweden) and CD4+ T cells were obtained by 122
negative selection using CD4+ T Cell Isolation Kit II according to the manufacturer’s 123
instructions (Miltenyi Biotech). The purity was determined by flow cytometry, and was 124
always ≥ 95%. CD4+ cells (106 cells/ml) were cultured in complete RPMI medium (RPMI 125
1640 (Gibco, Carlsbad, CA, USA), 10% heat-inactivated fetal calf serum (FCS) (Gibco), 126
2mM L-glutamine) and stimulated or not in 96-well flat-bottomed microculture plates coated 127
with anti-CD3 antibodies (10µg/ml, OKT3 clone) and soluble anti-CD28 (2µg/ml; BD 128
Biosciences) for 24 hr or with Phorbol 12-myristate 13-acetate (PMA 50ng/ml) and 129
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

7
Ionomycin (1μg/ml) for 5 hours. Cells were then recovered by centrifugation for further 130
analyses. 131
132
Plasmids and chemicals 133
The MTA1, MTA3, SENP2 and BCL11A expression vectors were kind gifts of Drs R. 134
Kumar, P. Wade, R. Hay and P. Tucker, respectively. The various BCL11B-Gal4 expression 135
constructs used in this study were generated as follows. Using as a template the wild-type 136
pcDNA3-FLAG-BCL11B plasmid (17, 24), we PCR amplified using relevant 137
oligonucleotides fragments corresponding to amino acids 1-20 and 1-58 of wild-type BCL11B 138
as well as 1-20 fragments with point mutation of Serine 2 (S2A, S2D, S2T). Next, these 139
fragments were cloned in frame with a C-terminal Gal-4 DNA-binding domain followed by a 140
Nuclear localization signal (NLS) and an HA epitope into the pSG5-Gal4-NLS-HA 141
eukaryotic vector to mimic their location in the full-length protein (25). The point (S2A, S2D) 142
and deletion (ΔMSRRKQ) mutants of BCL11B were PCR amplified from the wt BCL11B 143
plasmid with relevant oligonucleotides and subcloned in pcDNA3-FLAG-BCL11B using 144
standard procedures. All constructs were verified by sequencing. PMA (Phorbol-12-145
Myristate-13-Acetate) and Ionomycin both from Sigma-Aldrich were used at a final 146
concentration of 50 ng/ml and 1 μg/ml, respectively. Bisindolylmaleimide II (BIM II) (used at 147
a final concentration of 10μM for 30 minutes) and Gö6983 (used at a final concentration of 148
1μM for 15 minutes) which are general inhibitors of protein kinase C (PKC) subtypes were 149
purchased from Sigma-Aldrich and Selleckchem, respectively (26, 27, 28). The MEK 150
inhibitor U0126 was purchased from Merck Millipore and used at a final concentration of 10 151
μM for 30 minutes. Okadaic Acid (OA) and Calyculin A (Cal. A) which are potent inhibitors 152
of all three type 2A Ser/Thr phosphatases (PP1, PP2A and PP6) (29) were both obtained from 153
Santa Cruz. They were dissolved in DMSO and used at a final concentration of 1 μM and 154
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

8
50 nM, respectively. Calyculin A also inhibits arsenic-induced PML SUMOylation (30) and 155
BCL11B SUMOylation (21, 31). The SUMOylation inhibitors 2-DO8 which is specific and 156
blocks SUMO transfer from the Ubc9-thioester complex to the substrates (32) and Anacardic 157
Acid which blocks SUMOylation without affecting ubiquitination (33) were both obtained 158
from Sigma and used at a final concentration of 50 μM (34). Anarcadic acid is also a potent 159
inhibitor of several Histone Acetyl Transferase (HATs) such as PCAF, P300 and TIP60 (35). 160
161
Transfection, immunoprecipitation and co-immunoprecipitation 162
Cells were transfected in Opti-MEM (Invitrogen) by the PEI method using ExGen 500 163
(Euromedex), as previously described with 2.5 µg of DNA corresponding to the relevant 164
expression vectors or the empty vector used as control. Cells were transfected for 6h and then 165
incubated in fresh complete medium. After 48h transfection, cells were rinsed with ice PBS 166
and lysed in cold IPH buffer (50mM Tris, 150 mM NaCl, 5 mM EDTA, 0.5% NP40, protease 167
inhibitor cocktail (Roche)) for co-immunoprecipitation. Cell lysates were sonicated briefly 168
and cleared by centrifugation (14 000 rpm, 4°C, 15 min). The supernatants were pre-cleared 169
with 15 µl of protein A/G sepharose beads (Amersham Bioscience) incubating during 1 hour 170
on a rotator at 4°C. Then, lysates were incubated with 2 µg of antibody on a rotator at 4°C 171
overnight. Later, 20 µl of protein A/G beads were added and incubated 30 min at 4°C. Finally, 172
the beads were washed three times with IPH buffer. Bound proteins were eluted by boiling in 173
Laemmli buffer (36). 174
To detect SUMOylation of endogenous BCL11B proteins by immunoprecipitation studies, 175
Jurkat cells were lysed in a lysis buffer containing 1% SDS, 20 mM Tris-HCl pH 8, 10% 176
glycerol, 1 mM DTT and 15 mM NEM. These cells extracts were immediately boiled for 10 177
minutes and diluted in 9 volumes of RIPA buffer (20 mM Tris-HCl pH8, 0.5 mM EDTA, 150 178
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

9
mM NaCl, 0.5% NP40, 10% glycerol) supplemented with 10mM N-ethylmaleimide (NEM) 179
and protease inhibitor cocktail for immunoprecipitation with anti BCL11B antibodies (37). 180
To detect endogenous interactions between BCL11 (A or B) and MTA (1 or 3) proteins, we 181
prepared the combined cytoplasmic and nucleoplasmic fractions and the micrococcal 182
nuclease-solubilised chromatin fractions as previously described (36, 38, 39). Two chromatin 183
fractions prepared in parallel from the same number of cells were analyzed in co-184
immunoprecipitation assays as described above, with control rabbit IgG or MTA1 and MTA3 185
specific antibodies. 186
187
Transfection and luciferase transactivation test 188
HEK293T cells were plated in 12-well plates Cell Bind (Corning) and transfected with 500 ng 189
of DNA for 6h in triplicate with the luciferase reporter plasmid pGL3-5xGal4 RE-tk luc 190
(luciferase expression under the control of Gal4 responsive elements) or the pGL3-empty 191
vector in combination with various BCL11B-Gal4 constructs and then incubated in fresh 192
complete medium. 48 hours after transfection, cells were rinsed with ice PBS and lysed in the 193
Luc assay buffer. After normalization to the β-galactosidase activity, the data were expressed 194
as the Luc activity relative to the activity of pGl3-Luc with pSG5-Gal4-NLS-HA which 195
represents the basal condition and was given an arbitrary value of 1. The results represent the 196
mean values from three independent transfections in triplicate. Luciferase and β-galactosidase 197
activities were measured using respectively beetle luciferin (Promega) and the Galacto-Light 198
kit (Tropix) with a Berthold chemiluminometer (36). 199
200
Antibodies and Western Blot analyses 201
To generate polyclonal antibodies against Phosphorylated-Serine 2 of BCL11B, control and 202
phosphorylated peptides corresponding to AA 1-19 of human BCL11B (H2N-203
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

10
MS(PO3H2)RRKQGNPQHLC-CONH2) peptide were synthesized, coupled to KLH and used 204
to immunize rabbit (Eurogentec, Belgium). Control and phosphospecific antibodies were 205
purified by affinity chromatography using standard protocols. Commercial antibodies of the 206
following specificity were used: FLAG M2 from Sigma; HA from BABCO; BCL11B 207
(ab18465 for Western Blot, ab28448 for IP and ChIP and A300-383A for IP) from Abcam 208
and Bethyl Laboratories; Phospho-Erk1/2 (9106) from Cell Signaling; Actin (sc-1616R); 209
MTA1 (sc-9445 for WB, sc-10813 for IP and sc-10813 or ab50263 for ChIP) from Santa Cruz 210
and Abcam; MTA3 (ab87275) from Abcam; Erk2 (sc-154) from Santa Cruz; HDAC2 (sc-211
7899 H-54) from Santa Cruz; CHD4 (ab70469) from Abcam; RbAP46 (sc-8272) from Santa 212
Cruz; RbAP48 (C15200206 for ChIP) from Diagenode; Anti phospho-PKC substrate (2261) 213
from Cell Signaling; KLF4 (ab106629) and RanGAP1 (ab92360) from Abcam. 214
Western blots were performed as previously described (40). The secondary antibodies were 215
horseradish peroxidase-linked antibodies against rabbit, rat and mouse immunoglobulins 216
(Amersham Biosciences); goat immunoglobulins (Southern Biotech). 217
To analyze the SUMOylation of BCL11B proteins by Western blotting analyses, transfected 218
HEK293 cells or Jurkat and human CD4+ T cells pelleted by centrifugation were directly 219
lysed in Laemmli loading buffer, boiled for 10 minutes and processed for Western blotting as 220
described above. 221
222
Chromatin Immunoprecipitation 223
Control or activated Jurkat and primary human CD4+ T-cells were washed with PBS and 224
resuspended in 0.5 ml PBS for 5x106 cells. Then, cells were fixed by adding formaldehyde to 225
a final concentration of 1% for 8 min at room temperature. To stop fixation, glycine was 226
added to a final concentration of 0.125M. After 5 min at room temperature, cells were 227
collected by centrifugation (1500 rpm, at 4°C, 5 min). The supernatants were removed and 228
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

11
cells were lysed by resuspension in chilled cell lysis buffer for 10 min on a rotator at 4°C. 229
Then, the samples were pelleted, resuspended in 200 µl nuclei lysis buffer and sonicated to 230
chromatin with an average size of 250 bp using a cooling BioRuptor (Diagenode, Belgium). 231
20 µg of chromatin was immunoprecipitated with indicated antibodies and real-time PCR 232
analyses were performed as described (36). The primers used are summarized in 233
Supplementary Table 1. For the IL2 promoter, we designed primers around the BCL11B US1 234
binding sites (19). For the Id2 promoter, the Transcription Start Site (TSS) is derived from the 235
Id2 human locus found in the NCBI Nucleotide database (gi: 568815596). The region just 236
upstream of the TSS contains several BCL11B binding sites as shown by ChIP-Seq analyses 237
of BCL11B binding in double positive human thymocytes (9). Clustal alignments of the 238
corresponding human and murine regions highlighted a conserved potential BCL11B direct 239
binding site, TGGGC, which has been analyzed in ChIP-qPCR experiments using relevant 240
oligonucleotides. Similar analyses identified a potential KLF4 binding site in the BCL11B 241
promoter. For the ChIP in presence of the PKC inhibitors, Jurkat cells were first incubated 242
with Gö6983 (1μM for 15 minutes) before the relevant activation treatment. 243
244
RT and Quantitative PCR 245
Total RNAs were reverse transcribed using random primers and MultiScribeTM reverse 246
transcriptase (Applied Biosystems). Real-time PCR analyses were performed by Power SYBR 247
green (Applied Biosystems) in a MX3005P fluorescence temperature cycler (Stratagene) 248
according to the manufacturer’s instructions. Results were normalized to 18S RNA used as 249
internal control. The primers used are summarized in Supplementary Table 2. 250
251
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

12
Statistics 252
Experiments were realised at least twice independently in duplicates or triplicates. Statistical 253
analyses were performed by Student’s test. The asterisk (*) indicates p < 0.05, (**) indicates p 254
< 0.01 and (***) indicates p < 0.001. 255
256
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

13
RESULTS 257
258
BCL11B interacts with the three MTA proteins 259
MTA1, MTA2 or MTA3 are found in a mutually exclusive manner in different specialized 260
NuRD complexes (41, 42). BCL11B interacts with the closely related MTA1 and MTA2 261
proteins (18). We thus investigated if BCL11B also interacts with the functionally distinct 262
MTA3 protein. Co-immunoprecipitation experiments (Co-IPs) in transiently transfected 263
HEK293T (Fig. 1A-B) or between endogenous proteins in the human acute lymphoblastic 264
leukemia cell line, MOLT4 (Fig. 1C-D) and in the human CD4+ T-cell line Jurkat (Supp. Fig. 265
1A-B) (19), demonstrated that BCL11B interacts with MTA1 and MTA3. Contrary to Jurkat, 266
MOLT4 cells express both BCL11B and BCL11A but with lower levels of MTA1 and MTA3 267
(Supplemental Fig. 1C-F) (43, 44). Similarly, we unravelled an interaction between BCL11A 268
and MTA1 or MTA3 proteins either ectopically expressed in HEK293T (Supp. Fig. 1G-H) or 269
endogenous in MOLT4 cells (Fig. 1E-F). Taken together, these results demonstrate that 270
BCL11B and BCL11A can interact with the MTA1/MTA2 and MTA3 proteins and hence 271
with a wide variety of NuRD complexes. 272
273
Mimicked phosphorylation of the BCL11B N-terminal domain (S2D) disrupts its 274
interaction with MTA proteins and relieves its transcriptional repression activity 275
The 1-45 amino acids of BCL11B containing the MSRRKQ motif shared with FOG-1 and 276
SALL1 are sufficient for the interaction with MTA1 in GST pull-down experiments (18, 26, 277
45, 46). To assess this interaction in vivo, we fused the 1-20 or 1-58 amino acids of BCL11B, 278
in frame with a C-terminal Gal4 DNA-binding domain (Fig. 2A). Transient transfection 279
assays in HEK293T demonstrated that these 1-20- and 1-58-BCL11B-Gal4 chimeras are 280
nuclear, interact with MTA1 and MTA3 and mediate strong transcriptional repression in 281
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

14
luciferase reporter assays (Fig. 2B-C-D) (Supp. Fig. 2A-C). Serine 2 (Ser2) of the conserved 282
MSRRKQ motif is embedded in a potential consensus site (S/T-X2-0-R/K1-3) for 283
phosphorylation by protein kinase C (PKC), a family of Serine/Threonine kinases involved in 284
many cellular functions, including T-cell activation (47-48). To study the importance of Ser2 285
and the effect of its potential phosphorylation, we constructed several point mutants of Ser2 286
(S2X) in the 1-20 BCL11B-Gal4 chimera (Fig. 2A). First, the phosphomimetic S2D mutant is 287
unable to co-immunoprecipitate MTA1 or MTA3 proteins and in luciferase transactivation 288
assays, its repression activity is significantly relieved (Fig. 2B-C-E-F) (Supp. Fig. 2). In 289
addition, the repression activity of the phosphodeficient S2A mutant is also inhibited (49) 290
whereas a Serine to Threonine substitution (S2T) has no significant impact on the repression 291
potential of the BCL11B-Gal4 chimera (Fig. 2F-G) (Supp. Fig. 2). 292
Thus, the first 20 amino acids of BCL11B, including the conserved MSRRKQ motif are 293
sufficient to interact with MTA1 or MTA3 and have autonomous repression activity. Ser2 is 294
important for these two properties which are severely impaired by a phosphomimetic point 295
mutation, S2D. 296
297
A phophosmimetic mutation of BCL11B Ser2, S2D, impedes NuRD recruitment 298
We engineered a S2D and a S2A full-length BCL11B point mutant and a deletion mutant, 299
BCL11B-ΔMSRRKQ (Fig. 3A). They all display a nuclear localization (Supp Fig. 2G). In 300
Co-IPs, the phosphomimetic S2D mutant weakly interacts with MTA1 and MTA3 301
overexpressed in HEK293T (Fig. 3B-C). Similarly, co-IPs assays demonstrated that the S2A 302
and S2D point mutants ectopically expressed in HEK293T still weakly interact with 303
endogenous MTA1 proteins whereas the ΔMSRRKQ deletion mutant is unable to do so (Fig. 304
3D). The MSRRKQ motif of FOG1 also interacts with RbAp48 (49). We thus investigated the 305
interaction of these mutants with other endogenous components of NuRD. We demonstrated 306
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

15
that the S2A mutant still significantly interacts with MTA1, MTA3 and RbAp46 as compared 307
to wt BCL11B (Fig. 3E). By contrast, the S2D mutant is totally unable to interact with 308
endogenous RbAp46 whereas it weakly interacts with MTA1, MTA3, CHD4 and HDAC2 309
(Fig. 3E-F). These results are in close agreement with the crystallographic structure of the 310
FOG 1-15 peptide bound to RbAp48 since Ser2 is engaged in hydrogen bond interaction 311
whereas the adjacent Arg3, Arg4 and Lys5 participate to ion pair contacts with Glu residues 312
in RbAp48 (49). Thus, a S2D phosphomimetic mutation in the N-terminal MSRRKQ motif of 313
BCL11B negatively regulates the interactions with several endogenous NuRD components. 314
315
BCL11B is phosphorylated by PKC on Ser2 upon PMA activation of HEK293T cells 316
and is SUMOylated. 317
We next raised antibodies against Phospho-Ser2 BCL11B which were able to detect 318
phosphorylation of BCL11B on Ser2 residue in transfected HEK293T cells treated with PMA, 319
a potent PKC activator (48) (Fig. 4A) (Supp. Fig. 3). These antibodies detected a strong 320
induction of Ser2 phosphorylation after overexpression in PMA-treated HEK293T of wt 321
BCL11B but not of the S2A mutant, thereby demonstrating their specificity (Fig. 4B). Finally, 322
addition of BIMII, a PKC inhibitor decreased BCL11B phosphorylation on Ser2 whereas 323
Okadaic acid, a Ser/Thr phosphatase inhibitor, increased it (Fig. 4C-D). 324
In P/A-stimulated murine thymocytes, Bcl11b is SUMOylated (21). In PMA-activated-325
HEK293T cells transfected with BCL11B, we also observed a slowly migrating band (above 326
the 150KDa Marker) which corresponds to the SUMOylation of BCL11B since it totally 327
disappeared upon cotransfection of the deSUMOylase SENP2 (Fig. 5A) (Supp. Fig. 3). In 328
addition, the PKC inhibitor BIMII seems to have no significant effects on BCL11B 329
SUMOylation level while the ERK inhibitor U0126 seemed to slightly reduce it (Fig. 5B). 330
The PKC inhibitor also partially inhibits Erk activation as previously shown (51). Bcl11b 331
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

16
SUMOylation is important for its interaction with p300 (21). Interestingly, whereas the S2A 332
and S2D point mutants are severely affected in their interaction with MTA1 as compared with 333
wt BCL11B (Fig. 3), they display similar levels of SUMOylation and hence of interaction 334
with endogenous p300 proteins (Fig. 5C-D), correlating the lack of significant effects of the 335
PKC inhibitor on BCL11B SUMOylation. 336
Thus, the PKC-mediated phosphorylation of BCL11B in HEK293T impinges on its 337
interaction with MTA1 but not on its SUMOylation and interaction with P300. 338
339
PKC-mediated Ser2 phoshorylation of endogenous BCL11B proteins upon PMA-340
Ionomycin (P/I) activation of Jurkat cells has no impact on its SUMOylation. 341
We next addressed this PKC-mediated Ser2 phosphorylation on endogenous BCL11B 342
proteins during human CD4+ T-cell activation. Western blot analyses of Jurkat cell extracts 343
with the anti pSer2 BCL11B serum confirmed a peak of transient phosphorylation of BCL11B 344
Ser2 after 30 minutes of activation with PMA-Ionomycin (P/I) (Fig. 6A). 345
Immunoprecipitation of Jurkat cell extracts by a C-terminal BCL11B antibody followed by 346
Western blot analyses with the anti pSer2 BCL11B serum identified BCL11B proteins 347
phosphorylated on Serine 2 upon T cell activation, notably in presence of the phosphatase 348
inhibitor, Okadaic acid (Fig. 6B). Treatment with Okadaic acid induced a supershift of 349
BCL11B both in basal and activated conditions due to its phosphorylation by ERK and PKC 350
pathways as detected by the anti pSer2 (Fig. 6B, lane 3 and Fig. 6C, lanes 2 and 4). 351
Furthermore, Ser2 phosphorylation observed in presence of Okadaic acid is lost if, before 352
activation, cells are treated with the PKC inhibitor, Gö6983 whereas the supershift of 353
BCL11B is still observed (Fig. 6D, lanes 4 and 5). 354
In activated Jurkat cells, BCL11B phosphorylation is transient and precedes its SUMOylation 355
(Fig. 6A). As observed in HEK293T cells, the PKC inhibitor seems to have no significant 356
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

17
effects on BCL11B SUMOylation after 60 minutes of activation, whereas ERK inhibition by 357
U0126 seems to slightly decrease it (Fig. 6E). Another Ser/Thr phosphatase inhibitor, 358
Calyculin A, is also a SUMOylation inhibitor as initially shown for PML (30) and later for 359
BCL11B (21, 31). Upon activation and Calyculin A treatment of Jurkat cells, we observed an 360
electromobility shift for the major BCL11B proteins whereas the slowest migrating BCL11B-361
SUMO band totally disappeared (Fig. 6F), as shown in murine thymocytes (21). Similar 362
results were obtained with the specific SUMOylation inhibitor 2-D08 (32, 34) (Fig. 6G) and 363
with Anacardic Acid (AA), an inhibitor of SUMOylation (33) and of various Histone Acetyl 364
transferase (HATs) (35). This might explain through transcriptional inhibition, the decrease of 365
BCL11B protein levels as well as of several SUMOylated proteins observed with AA (Supp. 366
Fig. 5). Finally, Western blot analyses with anti-SUMO-2 after immunoprecipitation of 367
endogenous BCL11B proteins under denaturing conditions, allowed the detection of 368
SUMOylated BCL11B proteins in activated Jurkat cells (Fig. 6H) (37). As a whole, these data 369
identify phosphorylation of BCL11B Ser2 by PKC as a new post-translational modification of 370
BCL11B during human CD4+ T-cell activation, preceding and acting independently of its 371
SUMOylation. 372
373
KLF4 directly represses BCL11B transcription upon PMA-Ionomycin (P/I) activation of 374
Jurkat cells. 375
SUMOylation of Bcl11b in P/A-treated murine thymocytes ultimately results in its 376
ubiquitination and degradation (21). However, whereas prolonged stimulation of Jurkat cells 377
with P/I also resulted in a decrease of BCL11B protein levels after 5 hours (Fig. 6A), this 378
correlated with transcriptional repression of BCL11B rather than degradation after 379
ubiquitination (Supp. Fig. 5). Indeed, qRT-PCR analyses demonstrated a strong decrease of 380
BCL11B mRNAs starting at 2 hours of stimulation (Fig. 7A). Furthermore, BCL11B proteins 381
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

18
levels decreased after 5 hours of P/I stimulation, even in presence of three different 382
proteasome inhibitors, MG132, ALLN or Lactacystin (Fig. 7B). Inducible overexpression of 383
the transcriptional repressor KLF4 in Jurkat cells inhibits the expression of T-cell associated 384
factors including BCL11B and induces its degradation after 48 hours (31). These results 385
prompted us to further investigate the relationship between KLF4 and BCL11B. In Jurkat 386
cells, qRT-PCR analyses detected a strong induction of KLF4 expression peaking 30-60 387
minutes after activation (Fig. 7C) and thus preceding the repression of BCL11B expression 388
(Fig. 7A). Furthermore, in silico analyses of the BCL11B promoter identified a conserved 389
KLF4 binding site upstream of the TSS (Fig. 7D). ChIP analyses demonstrated the direct 390
binding of KLF4 on the BCL11B promoter and on CXCR4, a bona fide KLF4 direct target-391
gene (31) (Fig. 7E) after 5 hours of activation when BCL11B and CXCR4 expression are 392
severely repressed (Fig. 7A) (Supp. Fig. 6A-B). 393
Thus, BCL11B is a new direct target gene of the transcriptional repressor KLF4 during human 394
CD4+ T cell activation. 395
396
Endogenous BCL11B binds the IL2 and Id2 promoters but with the MTA1 co-repressor 397
or the P300 co-activator in resting versus PMA-Ionomycin (P/I) activated Jurkat T cells. 398
BCL11B participates through interaction with the P300 co-activator in the transcriptional 399
activation of IL2 and of Id2 (19, 21). Activation of Jurkat cells with P/I induced a huge up-400
regulation of IL2 gene expression starting as early as 30 minutes of treatment and a 401
significant, albeit slightly delayed up-regulation of Id2 around 120 minutes (Fig. 8A-B and 402
Supp. Fig. 6C), whereas BCL11B mRNA levels decreased after 2 hours of treatment (Fig. 403
7A). Co-immunoprecipitation of BCL11B with MTA1 or P300 endogenous proteins in Jurkat 404
cells treated for 30 minutes with P/I demonstrated that the peak of BCL11B Ser2 405
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

19
phosphorylation, we reproducibly observed (Fig. 6A), is concomitant with a slight decrease of 406
BCL11B-MTA1 interaction and a clear increase of BCL11B-P300 interaction (Fig. 8C). 407
To correlate these results with promoter occupancy, we first performed pilot ChIP 408
experiments with chromatin prepared from Jurkat cells and showed that BCL11B and MTA1 409
are bound on the Upstream Site 1 (US1) of the IL2 promoter (19) and on the Id2 promoter on 410
a newly identified and conserved BCL11B binding site located just upstream of the 411
transcription start site (Fig. 8D and Supp. Fig. 7). ChIP experiments conducted with Jurkat 412
cells treated with DMSO or with P/I for 30 minutes failed to detect any significant differences 413
in BCL11B binding to the IL2 and Id2 promoters (Fig. 8E-F). By contrast, in Jurkat cells 414
activated for 30 minutes to induce the peak of BCL11B Ser2 phosphorylation (Fig. 6), IL2 415
and Id2 promoter occupancy by MTA1 and MTA3 was markedly decreased as compared to 416
steady-state (Fig. 8E-H). This effect was not due to a down-regulation of MTA1 expression 417
since the levels of MTA1 RNAs (Supp. Fig. 6) are slightly increased by the treatment. 418
Conversely and in agreement with the Co-IP experiments, recruitment of the P300 coactivator 419
at the IL2 and Id2 promoters is significantly increased in activated Jurkat cells (Fig. 8I). As 420
control, none of these proteins were detected on the GAPDH promoter (Fig. 8G-I). 421
Ectopic expression of S2D or S2A BCL11B mutants in Jurkat cells to directly address the 422
impact of Ser2 phosphorylation on this MTA1/P300 switch is not feasible. Indeed, the prior 423
inactivation of the highly expressed endogenous BCL11B proteins will rapidly inhibit the 424
proliferation of Jurkat cells and induce apoptosis (51, 52). To obtain further evidence of the 425
specific involvement of PKC in regulating BCL11B interactions with MTA1 and P300, Jurkat 426
cells were activated in presence of a pan-PKC inhibitor, Gö6983. Pretreatment of Jurkat cells 427
with Gö6983 has no effects on BCL11B or MTA1 and RbAp48 expression levels but blocked 428
the P/I-induced activation of IL2 expression (Fig. 9A-B) (Supp. Fig. 6G-H). Furthermore, in 429
presence of the PKC inhibitor similar levels of BCL11B are bound to the IL2 promoter but 430
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

20
MTA1 and RbAp48 are not released from the promoter (Fig. 9C-D). Conversely, Gö6983 431
inhibits the P/I-induced increase of P300 binding on the IL2 US1 site (Fig. 9D). 432
Together, these data show that P/I treatment does not affect direct BCL11B binding to IL2 433
and Id2 promoters but induces its phosphorylation on Ser2 by PKC, to disrupt interaction with 434
NuRD repressive complexes and to mediate interaction with P300, thereby participating to its 435
transcriptional activation. 436
Activation of primary human CD4+ T cells switches BCL11B from a transcriptional 437
repressor to a transcriptional activator of IL2 438
Primary human CD4+ T cells were activated with P/I for 5 hours or with antiCD3/CD28 for 439
24 hours to induce TCR stimulation (23). In both conditions, we observed a 2-fold repression 440
of BCL11B mRNA levels whereas BCL11B protein levels were roughly identical suggesting 441
some post-translational stabilization mechanisms (Fig 10A-B). Interestingly, a concomitant 442
increase of BCL11B Ser2 phosphorylation and SUMOylation nicely correlating with 443
transcriptional activation of IL2 was observed, especially with the anti CD3/CD28 antibodies 444
(Fig. 10B). ChIP analyses were thus performed after 24h stimulation with anti CD3/CD28 445
antibodies, a treatment mimicking the canonical activation of T-cells through antigen 446
presenting cells, since IL2 induction was particularly efficient (ca. 4000-fold) in these 447
conditions (Fig. 10C). This activation had no effect on the binding of BCL11B on the IL2 448
promoter but led to a strongly increased recruitment of P300 to this region, concomitant with 449
the transcriptional activation of IL2 (Fig. 10D-E). 450
These results therefore show that upon physiological activation of primary human CD4+ T 451
cells, BCL11B through phosphorylation of Ser2 in its N-terminal repression domain is 452
switched from a transcriptional repressor to an activator of IL2 expression. 453
454
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

21
DISCUSSION 455
456
Multiple post-translational modifications (PTMs) of a given protein substrate can occur 457
simultaneously or sequentially to modulate its function, localization and stability through 458
cooperative or antagonizing effects on interacting partners. BCL11B is essential for T-cell 459
development. Recently, Bcl11b function was shown to be finely regulated during TCR 460
activation of resting murine thymocytes by a MAPK-initiated dynamic sequence of PTMs. 461
Indeed, treatment with phorbol ester and calcium ionophore resulted in rapid phosphorylation 462
on 23 S/TP sites, deSUMOylation, dephosphorylation, reSUMOylation, ubiquitination and 463
ultimately degradation of Bcl11b (21, 22, 53). Importantly, this complex pathway facilitates 464
derepression of repressed direct target-genes of Bcl11b, as shown for Id2, when immature 465
thymocytes need to initiate differentiation programs (21). 466
Here, we describe a different and much simpler PKC-mediated pathway which through 467
phosphorylation of Ser2 in the BCL11B conserved MSRKKQ motif participates to the up-468
regulation of IL2, another BCL11B direct target gene, during activation of human CD4+ T 469
cells by PMA/Ionomycin (P/I) or by anti CD3/CD28 antibodies as a surrogate for canonical 470
activation of T-cells through antigen-presenting cells (Fig. 11). Notably, this is the first 471
description at the endogenous level of a PTM targeting this important MSRRKQ repression 472
motif shared by several distinct nuclear proteins. The phosphorylation of Bcl11b Ser2 has not 473
been detected during differentiation of resting murine thymocytes (90% being DP cells) (21), 474
a population clearly distinct of the human CD4+ T cells analyzed here. However, the N-475
terminal region was not present in the coverage map of Bcl11b (21) since trypsin digests 476
which cut after Arg and Lys residues are predicted to generate hardly detectable MSR 477
peptides. 478
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

22
The Ser2 phosphorylation peak in Jurkat cells precedes the SUMOylation peak but, a specific 479
PKC inhibitor has no effect on the SUMOylation of BCL11B (Fig. 6). By contrast, the 480
multiple phosphorylation events induced by the MAPK pathway in murine thymocytes favor 481
the interaction of Bcl11b with the deSUMOylase SENP1 (21). Another major difference is the 482
lack of ubiquitination of human BCL11B after prolonged P/I treatment of Jurkat cells. In fact, 483
the decrease of BCL11B proteins levels observed after 5 hours of activation seems to mainly 484
rely on direct transcriptional repression of BCL11B by KLF4 rather than on BCL11B protein 485
degradation by the proteasome (Fig. 7). 486
These different MAPK- or PKC-mediated pathways however both orchestrate the 487
transcriptional activity of BCL11B to allow derepression of direct target genes, albeit through 488
strikingly different mechanisms. Indeed, through quantitative ChIP experiments in activated 489
Jurkat cells or in purified normal human CD4+ T cells, we observed similar binding of 490
BCL11B to the IL2 promoter as compared to basal conditions. However, in these activating 491
conditions, which result in BCL11B Ser2 phosphorylation, the levels of MTA1 are severely 492
reduced at the IL2 promoter whereas P300 levels reciprocally increase on line with the 493
induction of IL2 expression (Fig. 8-10). In the Co-IP experiments (Fig. 8C) whereas a net 494
increase of BCL11B-P300 interaction is clearly seen, the impact of T-cell activation on 495
BCL11B-MTA1 interaction is relatively low. However, in contrast with Co-IPs, the ChIP 496
experiments were performed on chromatin and are thus more likely to reflect the function of a 497
transcription factor and of its different partners in their natural context (54, 55). In developing 498
murine thymocytes, Bcl11b and MTA1 remained associated at the Id2 promoter both in basal 499
conditions and activating conditions, as shown by ChIP experiments (21). However, as stated 500
by these authors, the possibility that MTA1 and/or other NuRD component proteins became 501
post-translationally modified as a consequence of this treatment has not been investigated in 502
detail. This is an important issue since the C-terminal end of MTA proteins involved in the 503
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

23
interaction with MSRRKQ-proteins is subject to extensive post-translational modifications. 504
Notably, the methylation-demethylation of MTA1 Lysine 532 regulates its cyclical- and 505
signaling- dependent association with NuRD corepressor or NuRF co-activator complexes, 506
respectively (56). The transcriptional co-factor FOG-1 (Friend of GATA) also interacts with 507
NuRD through a N-terminal MSRRKQ motif and this interaction plays a key role for lineage 508
commitment during erythropoiesis and megakaryopoiesis (45, 57-58). Strikingly, ChIP 509
experiments revealed that NuRD components are present at both repressed and active 510
GATA1/FOG1 target genes (59). These results suggest two possible mechanisms for 511
interaction of MTA1 with different transcription factors depending on the cellular context: (i) 512
binding of MTA1 to the target genes both in active or repressive conditions but with PTMs 513
fine-tuning its interaction with repressive or activating complexes as reported for 514
GATA1/FOG1 or (ii) presence only in repressive conditions and loss of interaction in 515
activating conditions due to specific PTMs in short MTA1-interacting motifs, as shown here 516
for PKC-mediated phosphorylation of BCL11B in CD4+ T cells. 517
Here, we demonstrated that this short motif is necessary and sufficient to mediate binding of 518
BCL11A and BCL11B to the three members of MTA co-repressors family, thus enabling 519
BCL11A, BCL11B and more generally MSRRKQ-containing proteins to interact with the 520
complete range of NuRD complexes. This highlights their flexibility to participate to various 521
differentiation programs involved in the development of the immune and central nervous 522
(CNS) systems and to regulate tissue-specific gene expression in many distinct cellular types 523
including T and B cells, ameloblasts or keratinocytes for BCL11B (16) or lymphopoiesis and 524
erythroid precursors for BCL11A (15). Interestingly, the interaction of the BTB/POZ 525
transcriptional repressors HIC1 and BCL6 with MTA1 and MTA3, respectively, are inhibited 526
by acetylation of a lysine residue in different conserved peptidic interaction motifs. HIC1 527
acetylation on K314 in the MKHEP SUMOylation/acetylation motif negatively regulates its 528
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

24
interaction with MTA1 whereas its SUMOylation favors it, notably in the DNA-damage 529
response (40, 60). BCL6 acetylation on K379 in the KKYK motif abrogates its ability to 530
interact with MTA3 which is a major repression mechanism controlling Germinal Center B 531
cells differentiation (61-63) as well as CD4+ T-cell fate and function (64). 532
Although our studies have been focused on BCL11B, it is tempting to speculate that similar 533
PTMs could be identified in other MSRRKQ-containing nuclear proteins. Indeed, the 534
developmental regulator SALL1 is certainly regulated by PKC-phosphorylation on Ser2, as 535
suggested by in vitro studies (27). For FOG1, the role of Ser2 has been investigated only 536
through Alanine scanning experiments which demonstrated that this S2A mutation did not 537
affect the repression potential of a FOG1 1-12 Gal4 chimera (57). However, these in vitro 538
studies or in vivo Knock-in models in mice and the crystal structure of RbAp48 (another 539
NuRD component) bound to the 15 N-terminal amino-acids of FOG-1 have provided 540
compelling evidences for the key role played by the RKK residues in the conserved 541
MSRRKQ motif (45, 50, 57-59). Whereas Ser2 is only involved in Hydrogen bond 542
interaction, these RKK residues are engaged both in hydrogen bond interaction and in ion pair 543
contacts with acidic Glu residues in RbAp48 (49). These strong interactions could be 544
impaired by the negative charge brought by PKC-mediated phosphorylation of the adjacent 545
Ser2 residue as shown for the interaction between the phosphomimetic S2D BCL11B mutant 546
and the related RbAp46 protein (Fig. 3E). Recently, ZNF827 was shown to recruit NuRD to 547
telomeres in cells using ALT (Alternative Lengthening of Telomeres) to enable telomere 548
extension. This recruitment relies on RRK residues found in an N-terminal MPRRKQ motif 549
highly homologous to the motif found in FOG, SALL1 and BCL11A/B except for the 550
replacement of Ser2 by a Pro residue (65). Thus, the PKC-mediated phosphorylation of Ser2 551
that we have demonstrated here for BCL11B would provide to these MSRRKQ-containing 552
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

25
proteins a rapid and efficient mechanism to fine tune their interaction via a RRK motif with 553
NuRD. 554
In conclusion, we have identified a novel mechanism for how the essential regulator of T cell 555
development BCL11B is regulated by PKC-mediated phosphorylation of Ser2 in its conserved 556
N-terminal repression domain during activation of CD4+ T cells. In addition, this BCL11B N-557
terminal domain is essential for the transcriptional repression of HIV-1 LTR sequences which 558
thus affects both HIV-1 replication and virus production in CD4+ T lymphocytes (66). In 559
future studies, it would be interesting to explore if this switch between transcriptional 560
repression and activation by phosphorylation in their MSRRKQ motifs could be generalized 561
to other NuRD-interacting proteins, in particular to BCL11A or FOG-1, two important 562
regulators of differentiation programs during normal hematopoiesis. 563
564
ACKNOWLEDGMENTS 565
Marion Dubuissez and Sonia Paget are supported by predoctoral fellowships from the 566
University of Lille. We thank Dr Capucine Van Rechem for critical reading of the manuscript. 567
568
REFERENCES 569
570
[1] Avram D, Fields A, Pretty On Top K, Nevrivy DJ, Ishmael JE, Leid M. 2000. 571 Isolation of a novel family of C(2)H(2) zinc finger proteins implicated in 572 transcriptional repression mediated by chicken ovalbumin upstream promoter 573 transcription factor (COUP-TF) orphan nuclear receptors. J Biol Chem 275:10315-574 10322. 575
576 [2] Avram D, Fields A, Senawong T, Topark-Ngarm A, Leid M. 2002. COUP-TF (chicken 577
ovalbumin upstream promoter transcription factor)-interacting protein 1 (CTIP1) is a 578 sequence-specific DNA binding protein. Biochem J:368 555-563. 579
580 [3] Wakabayashi Y, Inoue J, Takahashi Y, Matsuki A, Kosugi-Okano H, Shinbo T, 581
Mishima Y, Niwa O, Kominami R. 2003. Homozygous deletions and point 582 mutations of the Rit1/Bcl11b gene in gamma-ray induced mouse thymic lymphomas. 583 Biochem Biophys Res Commun 301:598-603. 584
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

26
585 [4] Avram D.Califano D. 2014. The Multifaceted Roles of Bcl11b in Thymic and Peripheral 586
T Cells: Impact on Immune Diseases. J Immunol 193:2059-2065. 587 588 [5] Wakabayashi Y, Watanabe H, Inoue J, Takeda N, Sakata J, Mishima Y, Hitomi J, 589
Yamamoto T, Utsuyama M, Niwa O, Aizawa S, Kominami R. 2003. Bcl11b is 590 required for differentiation and survival of alphabeta T lymphocytes. Nat Immunol 591 4:533-539. 592
593 [6] Albu DI, Feng D, Bhattacharya D, Jenkins NA, Copeland NG, Liu P, Avram D. 2007 594
BCL11B is required for positive selection and survival of double-positive thymocytes. 595 J Exp Med 204:3003-3015. 596
597 [7] Li L, Leid M, Rothenberg EV. 2010. An early T cell lineage commitment checkpoint 598
dependent on the transcription factor Bcl11b. Science 329:89-93. 599 600 [8] Li P, Burke S, Wang J, Chen X, Ortiz M, Lee SC, Lu D, Campos L, Goulding D, Ng 601
BL, Dougan G, Huntly B, Gottgens B, Jenkins NA, Copeland NG, Colucci F, Liu 602 P 2010. Reprogramming of T cells to natural killer-like cells upon Bcl11b deletion. 603 Science 329:85-89. 604
605 [9] Kastner P, Chan S, Vogel WK, Zhang LJ, Topark-Ngarm A, Golonzhka O, Jost B, 606
Le Gras S, Gross MK, Leid M. 2010. Bcl11b represses a mature T-cell gene 607 expression program in immature CD4(+)CD8(+) thymocytes. Eur J Immunol 40:2143-608 2154. 609
610 [10] Gutierrez A, Kentsis A, Sanda T, Holmfeldt L, Chen SC, Zhang J, Protopopov A, 611
Chin L, Dahlberg SE, Neuberg DS, Silverman LB, Winter SS, Hunger SP, Sallan 612 SE, Zha S, Alt FW, Downing JR, Mullighan CG, Look AT. 2011. The BCL11B 613 tumor suppressor is mutated across the major molecular subtypes of T-cell acute 614 lymphoblastic leukemia. Blood 118:4169-4173. 615
616 [11] Kominami R. 2012. Role of the transcription factor Bcl11b in development and 617
lymphomagenesis. Proc Jpn Acad Ser B Phys Biol Sci 88:72-87. 618 619 [12] Di Santo JP. 2010. Immunology. A guardian of T cell fate. Science 329 44-5. 620 621 [13] Liu P, Keller JR, Ortiz M, Tessarollo L, Rachel RA, Nakamura T, Jenkins NA, 622
Copeland NG. 2003. Bcl11a is essential for normal lymphoid development. Nat 623 Immunol 4:525-532. 624
625 [14] Sankaran VG, Xu J, Ragoczy T, Ippolito GC, Walkley CR, Maika SD, Fujiwara Y, 626
Ito M, Groudine M, Bender MA, Tucker PW, Orkin SH. 2009. Developmental and 627 species-divergent globin switching are driven by BCL11A. Nature 460:1093-1097. 628
629 [15] Bauer DE, Kamran SC, Orkin SH. 2012. Reawakening fetal hemoglobin: prospects for 630
new therapies for the beta-globin disorders. Blood 120:2945-53. 631 632 [16] Le Douce V, Cherrier T, Riclet R, Rohr O, SchwartzC. 2014. The many lives of 633
CTIP2: from AIDS to cancer and cardiac hypertrophy. J Cell Physiol 229:533-537. 634
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

27
635 [17] Marban C, Suzanne S, Dequiedt F, de Walque S, Redel L, Van Lint C, Aunis D, 636
Rohr O. 2007 Recruitment of chromatin-modifying enzymes by CTIP2 promotes 637 HIV-1 transcriptional silencing. Embo J 26:412-423. 638
639 [18] Cismasiu VB, Adamo K, Gecewicz J, Duque J, Lin Q, Avram D. 2005 BCL11B 640
functionally associates with the NuRD complex in T lymphocytes to repress targeted 641 promoter. Oncogene 24:6753-6764. 642
643 [19] Cismasiu VB, Ghanta S, Duque J, Albu DI, Chen HM, Kasturi R, Avram D. 2006. 644
BCL11B participates in the activation of IL2 gene expression in CD4+ T 645 lymphocytes. Blood 108: 2695-2702. 646
647 [20] Cismasiu VB, Duque J, Paskaleva E, Califano D, Ghanta S, Young HA, Avram D. 648
2009.BCL11B enhances TCR/CD28-triggered NF-kappaB activation through up-649 regulation of Cot kinase gene expression in T-lymphocytes. Biochem J 417:457-466. 650
651 [21] Zhang LJ, Vogel WK, Liu X, Topark-Ngarm A, Arbogast BL, Maier CS, Filtz TM, 652
Leid M. 2012. Coordinated regulation of transcription factor Bcl11b activity in 653 thymocytes by the mitogen-activated protein kinase (MAPK) pathways and protein 654 sumoylation. J Biol Chem 287:26971-26988. 655
656 [22] Filtz TM, Vogel WK, Leid M. 2014. Regulation of transcription factor activity by 657
interconnected post-translational modifications. Trends Pharmacol Sci 35:76-85. 658 659 [23] Azzaoui I, Yahia SA, Chang Y, Vorng H, Morales O, Fan Y, Delhem N, Ple C, 660
Tonnel AB, Wallaert B, Tsicopoulos A. 2011. CCL18 differentiates dendritic cells 661 in tolerogenic cells able to prime regulatory T cells in healthy subjects. Blood 662 118:3549-3558. 663
664 [24] Cherrier T, Le Douce V, Eilebrecht S, Riclet R, Marban C, Dequiedt F, Goumon Y, 665
Paillart JC, Mericskay M, Parlakian A, Bausero P, Abbas W, Herbein G, 666 Kurdistani SK, Grana X, Van Driessche B, Schwartz C, Candolfi E, Benecke AG, 667 Van Lint C, Rohr O. 2013. CTIP2 is a negative regulator of P-TEFb. Proc Natl Acad 668 Sci U S A 110:12655-12660. 669
670 [25] Stankovic-Valentin N,Deltour S, Seeler J, Pinte S, Vergoten G, Guerardel C, Dejean 671
A, Leprince D. 2007. An acetylation/deacetylation-SUMOylation switch through a 672 phylogenetically conserved psiKXEP motif in the tumor suppressor HIC1 regulates 673 transcriptional repression activity. Mol Cell Biol 27:2661-2675. 674
675 [26] Lauberth SM, Rauchman M. 2006. A conserved 12-amino acid motif in Sall1 recruits 676
the nucleosome remodeling and deacetylase corepressor complex. J Biol Chem 677 281:23922-23931. 678
679 [27] Lauberth SM, Bilyeu AC, Firulli BA, Kroll KL, Rauchman M 2007. A 680
phosphomimetic mutation in the Sall1 repression motif disrupts recruitment of the 681 nucleosome remodeling and deacetylase complex and repression of Gbx2. J Biol 682 Chem 282:34858-34868. 683
684
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

28
[28] Chen L, Meng Q, Jing X, Xu P, Luo D. 2011. A role for protein kinase C in the 685 regulation of membrane fluidity and Ca(2)(+) flux at the endoplasmic reticulum and 686 plasma membranes of HEK293 and Jurkat cells. Cell Signal 23:497-505. 687
688 [29] Prickett TD, Brautigan DL. 2006. The alpha4 regulatory subunit exerts opposing 689
allosteric effects on protein phosphatases PP6 and PP2A. J Biol Chem 281:30503-690 30511. 691
692 [30] Muller S, Matunis MJ, Dejean A. 1998 Conjugation with the ubiquitin-related modifier 693
SUMO-1 regulates the partitioning of PML within the nucleus. Embo J 17:61-70. 694 695 [31] Li W, Jiang Z, Li T, Wei X, Zheng Y, Wu D, Yang Y, Chen S, Xu B, Zhong M, 696
Jiang J, Hu Y, Su H, Zhang M, Huang X, Geng S, Weng J, Du X, Liu P, Li Y, 697 Liu H, Yao Y, Li P. 2015. Genome-wide analyses identify KLF4 as an important 698 negative regulator in T-cell acute lymphoblastic leukemia through directly inhibiting 699 T-cell associated genes. Mol Cancer 14 26. 700
701 [32] Kim YS, Nagy K, Keyser S, and Schneekloth SJ Jr. 2013. An electrophoretic 702
mobility shift assay identifies a mechanistically unique inhibitor of protein 703 sumoylation. Chem Biol 20:604-613. 704
705 [33] Fukuda I, Ito A, Hirai G, Nishimura S, Kawasaki H, Saitoh H, Kimura K, Sodeoka 706
M, Yoshida M. 2009. Ginkgolic acid inhibits protein SUMOylation by blocking 707 formation of the E1-SUMO intermediate. Chem Biol 16:133-140. 708
709 [34] Hoellein A, Fallahi M, Schoeffmann S, Steidle S, Schaub FX, Rudelius M, Laitinen 710
I, Nilsson L, Goga A, Peschel C, Nilsson JA, Cleveland JL, Keller U. 2014. Myc-711 induced SUMOylation is a therapeutic vulnerability for B-cell lymphoma. Blood 712 124:2081-2090. 713
714 [35] Hemshekhar M, Sebastin Santhosh M, Kemparaju K, Girish KS. 2011. Emerging 715
roles of anacardic acid and its derivatives: a pharmacological overview. Basic Clin 716 Pharmacol Toxicol 110:122-132. 717
718 [36] Boulay G, Dubuissez M, Van Rechem C, Forget A, Helin K, Ayrault O, Leprince D. 719
2012. Hypermethylated in cancer 1 (HIC1) recruits polycomb repressive complex 2 720 (PRC2) to a subset of its target genes through interaction with human polycomb-like 721 (hPCL) proteins. J Biol Chem 287:10509-10524. 722
723 [37] Wu CS, Ouyang J, Mori E, Nguyen HD, Marechal A, Hallet A, Chen DJ, Zou L. 724
2014. SUMOylation of ATRIP potentiates DNA damage signaling by boosting 725 multiple protein interactions in the ATR pathway. Genes Dev 28:1472-1484. 726
727 [38] Van Dessel N, Beke L, Gornemann J, Minnebo N, Beullens M, Tanuma N, Shima H, 728
Van Eynde A, Bollen M. 2010. The phosphatase interactor NIPP1 regulates the 729 occupancy of the histone methyltransferase EZH2 at Polycomb targets. Nucleic Acids 730 Res 38 7500-12. 731
732
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

29
[39] Boulay G, Rosnoblet C, Guerardel C, Angrand PO, Leprince D. 2011. Functional 733 characterization of human Polycomb-like 3 isoforms identifies them as components of 734 distinct EZH2 protein complexes. Biochem J 434:333-342. 735
736 [40] Dehennaut V, Loison I, Dubuissez M, Nassour J, Abbadie C, Leprince D. 2013. 737
DNA double-strand breaks lead to activation of hypermethylated in cancer 1 (HIC1) 738 by SUMOylation to regulate DNA repair. J Biol Chem 288:10254-10264. 739
740 [41] Dege C, Hagman J. 2014. Mi-2/NuRD chromatin remodeling complexes regulate B and 741
T-lymphocyte development and function. Immunol Rev 261:126-140. 742 743 [42] Manavathi B, Singh K, Kumar R. 2007. MTA family of coregulators in nuclear 744
receptor biology and pathology. Nucl Recept Signal 5:e010. 745 746 [43] Satterwhite E, Sonoki T, Willis TG, Harder L, Nowak R, Arriola EL, Liu H, Price 747
HP, Gesk S, Steinemann D, Schlegelberger B, Oscier DG, Siebert R, Tucker PW, 748 Dyer MJ. 2001. The BCL11 gene family: involvement of BCL11A in lymphoid 749 malignancies. Blood 98:3413-3420. 750
751 [44] Durum SK. 2003. Bcl11: sibling rivalry in lymphoid development. Nat Immunol 4:512-752
514. 753 754 [45] Hong W, Nakazawa M, Chen YY, Kori R, Vakoc CR, Rakowski C, Blobel GA. 755
2005. FOG-1 recruits the NuRD repressor complex to mediate transcriptional 756 repression by GATA-1. Embo J 24:2367-2378. 757
758 [46] Alqarni SS, Murthy A, Zhang W, Przewloka MR, Silva AP, Watson AA, Lejon S, 759
Pei XY, Smits AH, Kloet SL, Wang H, Shepherd NE, Stokes PH, Blobel GA, 760 Vermeulen M, Glover DM, Mackay JP, Laue ED. 2014. Insight into the 761 architecture of the NuRD complex: structure of the RbAp48-MTA1 subcomplex. J 762 Biol Chem 289:21844-21855. 763
764 [47] Kennelly PJ, Krebs EG. 1991. Consensus sequences as substrate specificity 765
determinants for protein kinases and protein phosphatases. J Biol Chem 266:15555-766 15558. 767
768 [48] Kong KF, Altman A. 2013. In and out of the bull's eye: protein kinase Cs in the 769
immunological synapse. Trends Immunol 34:234-242. 770 771 49] Lejon S, Thong SY, Murthy A, AlQarni S, Murzina NV, Blobel GA, Laue ED, 772
Mackay JP. 2011. Insights into association of the NuRD complex with FOG-1 from 773 the crystal structure of an RbAp48.FOG-1 complex. J Biol Chem 286:1196-1203. 774
775 [50] Huang X, Chen S, Shen Q, Chen S, Yang L, Grabarczyk P, Przybylski GK, 776
Schmidt CA, Li Y. 2011. Down regulation of BCL11B expression inhibits 777 proliferation and induces apoptosis in malignant T cells by BCL11B-935-siRNA. 778 Hematology:16236-242. 779
780 [51] Puente LG, Stone JC, Ostergaard HL,Chen S, Yang L, Weng J, Du X, Grabarczyk 781
P, Przybylski GK, Schmidt CA, Li. 2000. Evidence for protein kinase C-dependent 782
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

30
and -independent activation of mitogen-activated protein kinase in T cells: potential 783 role of additional diacylglycerol binding protein. J Immunology 165:6865-71. 784
785 [52] Huang X, Shen Q, Chen S,Chen S, Yang L, Weng J, Du X, Grabarczyk P, 786
Przybylski GK, Schmidt CA, Li. Y. 2011. Gene expression profiles in BCL11B-787 siRNA treated malignant T cells. J Hematol Oncol 4:23. 788
789 [53] Vogel WK, Gafken PR, Leid M, Filtz TM. 2014. Kinetic analysis of BCL11B multisite 790
phosphorylation-dephosphorylation and coupled sumoylation in primary thymocytes 791 by multiple reaction monitoring mass spectroscopy. J Proteome Res 13:5860-5868. 792
793 [54] Li X, Wang W, Wang J, Malovannaya A, Xi Y, Li W, Guerra R, Hawke DH, Qin J, 794
Chen J. 2015. Proteomic analyses reveal distinct chromatin-associated and soluble 795 transcription factor complexes. Mol Syst Biol 11:775. 796
797 [55] Ji Z, Sharrocks AD. 2015 Changing partners: transcription factors form different 798
complexes on and off chromatin. Mol Syst Biol 11:782. 799 800 [56] Nair SS, Li DQ, Kumar R. 2013. A core chromatin remodeling factor instructs global 801
chromatin signaling through multivalent reading of nucleosome codes. Mol Cell 802 49:704-718. 803
804 [57] Lin AC, Roche AE, Wilk J, Svensson EC. 2004. The N termini of Friend of GATA 805
(FOG) proteins define a novel transcriptional repression motif and a superfamily of 806 transcriptional repressors. J Biol Chem 279:55017-55023. 807
808 [58] Gao Z, Huang Z, Olivey HE, Gurbuxani S, Crispino JD, Svensson EC. 2010. FOG-809
1-mediated recruitment of NuRD is required for cell lineage re-enforcement during 810 haematopoiesis. Embo J 29:457-468. 811
812 [59] Miccio A, Wang Y, Hong W, Gregory GD, Wang H, Yu X, Choi JK, Shelat S, Tong 813
W, Poncz M, Blobel GA. 2010. NuRD mediates activating and repressive functions 814 of GATA-1 and FOG-1 during blood development. Embo J 29:442-456. 815
816 [60] Van Rechem C, Boulay G, Pinte S, Stankovic-Valentin N, Guerardel C, Leprince D. 817
2010. Differential regulation of HIC1 target genes by CtBP and NuRD, via an 818 acetylation/SUMOylation switch, in quiescent versus proliferating cells. Mol Cell Biol 819 30:4045-4059. 820
821 [61] Fujita N, Jaye DL, Geigerman C, Akyildiz A, Mooney MR, Boss JM, Wade PA. 822
2004. MTA3 and the Mi-2/NuRD complex regulate cell fate during B lymphocyte 823 differentiation. Cell 119:75-86. 824
825 [62] Huang C, Gonzalez DG, Cote CM, Jiang Y, Hatzi K, Teater M, Dai K, Hla T, 826
Haberman AM, Melnick A. 2014. The BCL6 RD2 domain governs commitment of 827 activated B cells to form germinal centers. Cell Rep 8:1497-1508. 828
829 [63] Parekh S, Polo JM, Shaknovich R, Juszczynski P, Lev P, Ranuncolo SM, Yin Y, 830
Klein U, Cattoretti G, Dalla Favera R, Shipp MA, Melnick A. 2007. BCL6 831
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

31
programs lymphoma cells for survival and differentiation through distinct biochemical 832 mechanisms. Blood 110:2067-2074. 833
834 [64] Nance JP, Belanger S, Johnston RJ, Hu JK, Takemori T, Crotty S. 2015. Bcl6 835
middle domain repressor function is required for T follicular helper cell differentiation 836 and utilizes the corepressor MTA3. Proc Natl Acad Sci U S A 112:13324-13329. 837
838 [65] Conomos D, Reddel RR, Pickett HA. 2014. NuRD-ZNF827 recruitment to telomeres 839
creates a molecular scaffold for homologous recombination. Nat Struct Mol Biol 840 21:760-770. 841
842 [66] Cismasiu VB, Paskaleva E, Suman Daya S, Canki M, Duus K, Avram D. 2008 843
BCL11B is a general transcriptional repressor of the HIV-1 long terminal repeat in T 844 lymphocytes through recruitment of the NuRD complex. Virology 380:173-181. 845
846 TABLES 847
Two Tables listing the various oligonucleotides used in the RT-qPCR and ChIP-qPCR 848
experiments can be found as Supplemental Information on line. 849
850
FIGURE LEGENDS 851
852
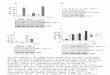
Figure 1. BCL11A and BCL11B interact with MTA1 and MTA3. (A) MTA1 interacts with 853
BCL11B. HEK293T were transfected with MTA1 and BCL11B expression vectors. Whole 854
cells extracts incubated with anti-FLAG antibodies (IP FLAG) and 1% of each lysate (Input) 855
were immunoblotted with the indicated antibodies. FLAG-BCL11B co-immunoprecipitate 856
endogenous MTA1 proteins (lane 3) visualized as a doublet (*). (B) BCL11B interacts with 857
MTA3. A similar experiment was conducted in HEK293T cells with BCL11B and MTA3 858
expression vectors. (C) Endogenous BCL11B and MTA1 proteins interact. Solubilized 859
chromatin fractions were prepared in duplicate from the same number of MOLT4 cells and 860
1% was retained for direct analyses by Western blot (Input). The interaction between 861
BCL11B and MTA1 was evaluated by co-immunoprecipitation assays. (D) Endogenous 862
BCL11B and MTA3; (E) BCL11A and MTA1 and (F) BCL11B and MTA3 proteins interact 863
in MOLT4 cells. The same procedure was used for panels C to E. 864
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

32
865
Figure 2. Ser 2 in the conserved N-terminal motif of BCL11B is essential for its interaction 866
with MTA1 and MTA3 and for its transcriptional repression activity (A) Schematic drawing 867
of the BCL11B-Gal4 (DNA-binding domain)-NLS (Nuclear localization signal)-HA fusion 868
proteins. (B) A phosphomimetic point mutation S2D in the BCL11B N-terminal domain 869
inhibits its interaction with MTA1. After transfection with the indicated expression vectors, 870
HEK293T cells lysates were immunoprecipitated with anti-c-myc antibodies. 871
Immunoprecipitated samples [IP c-myc (MTA1)] and 1% of whole cell extracts (Input) were 872
analyzed by immunoblotting with the indicated antibodies. (C) The BCL11B-MTA3 873
interaction is also negatively regulated by the S2D mutation. A similar experiment was 874
performed in HEK293T with a FLAG-MTA3 plasmid and the BCL11B-Gal4 chimeras. (D) 875
The conserved BCL11B N-terminal domain represses transcription. The transcriptional 876
activity of BCL11B-Gal4 constructs was tested by transient luciferase reporter assays in 877
HEK293T cells. The results represent the mean values from three independent transfections in 878
triplicate. (E) The S2D phosphomimetic mutation in the BCL11B N-terminal domain partially 879
inhibits its transcriptional repression potential. A similar luciferase reporter assay was 880
conducted in HEK293T with the 1-20 BCL11B wild-type (WT) and the S2D point mutant. (F-881
G) Repression potential of the various S2X point mutants BCL11B N-terminal domain. 882
Luciferase reporter assays were conducted with the S2A, S2D, S2T and wt BCL11B-Gal4 883
constructs as described above. 884
885
Figure 3. The phosphomimetic S2D mutation in BCL11B inhibits NuRD recruitment. (A) 886
Schematic drawing of the full-length BCL11B proteins tested. (B) The interaction with MTA1 887
is impaired by the S2D phosphomimetic mutation of BCL11B. HEK293T were transfected 888
with the indicated combination of expression vectors and Co-IP assays followed by 889
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

33
immunoblotting with the indicated antibodies were performed. (C) The BCL11B-MTA3 890
interaction is also strongly reduced by the S2D phosphomimetic mutation. A similar Co-IP 891
experiment was realized in HEK293T but with the FLAG-MTA3 expression vector. Relevant 892
pieces of the membranes were cut (around the 80KDa marker) to separate the MTA3 and 893
BCL11B proteins and probed with anti-FLAG antibodies. (D) Interaction of wt, S2A, S2D 894
and ΔMSRKKQ BCL11B with endogenous MTA1 proteins. Total extracts of HEK293T 895
transfected with the indicated plasmids were analysed by Co-IP with anti-FLAG antibodies 896
and imunoblotted with MTA1 and FLAG antibodies. (E-F) Interaction of the S2A and S2D 897
BCL11B point mutants with endogenous NuRD components. Co-IP experiments followed by 898
immunoblotting with the indicated antibodies were performed as in Panel D with the S2A and 899
S2D (Panel E) or S2D (Panel F) BCL11B mutants. 900
901
Figure 4. Phosphorylation of BCL11B Ser2 in HEK293T cells upon PKC activation. (A) 902
PMA-induced in vitro phosphorylation of BCL11B Ser2. HEK293T cells transfected with 903
FLAG-BCL11B were mock-treated (-), incubated with vehicle (DMSO) or with PMA (1μM) 904
for 20 minutes. Immunoprecipitated samples IP (FLAG) and Input lysates were analyzed by 905
immunoblotting with antibodies specific for phosphorylated Ser2 (pSer2 BCL11B) and FLAG 906
as control. (B) Specificity of the anti-pSer2 BCL11B antibodies. A similar experiment was 907
performed to compare in the three conditions the BCL11B S2A mutant and wt BCL11B. * 908
refers to a non-specific band. (C) PKC is implicated in BCL11B Ser2 phosphorylation. 909
HEK293T cells were transfected with BCL11B and activated exactly as in panel A. One plate 910
was pre-incubated with the pan-PKC inhibitor BIMII before PMA activation. The PKC 911
inhibitor also partially inhibits Erk activation as previously shown (51). (D) The phosphatase 912
inhibitor Okadaic Acid (OA) allows detection of BCL11B Ser2 phosphorylation in basal 913
conditions. HEK293T cells transfected with the indicated expression vectors were incubated 914
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

34
with DMSO or treated with OA. Total cell lysates were analyzed by immunoblotting with the 915
indicated antibodies. 916
917
Figure 5. BCL11B SUMOylation in HEK293T cells is independent of Ser2 Phosphorylation. 918
(A) BCL11B is SUMOylated. Total cells extracts of HEK293T transfected with the indicated 919
combinations of BCL11B, SUMO2 and the deSUMOylase SENP2 were prepared in 920
denaturing conditions and immunoblotted with indicated antibodies. The arrowhead 921
corresponds to SUMOylated forms of BCL11B (B) The PKC pathway does not impinge on 922
BCL11B SUMOylation in contrast with the ERK pathway. HEK293T cells were transfected 923
with BCL11B and pre-incubated with the PKC inhibitor BIMII or with the Erk1/2 inhibitor, 924
U0126 before PMA activation. The PKC inhibitor also partially inhibits Erk activation as 925
previously shown (51). Total cells extracts prepared in denaturing conditions were 926
immunoblotted as indicated. (C) The S2D and S2A point mutations have no significant 927
impact on BCL11B SUMOylation. HEK293T were transfected as indicated, lysed in 928
denaturing conditions and immunoblotted with anti FLAG antibodies to detect BCL11B and 929
its SUMOylated forms. Hsp60 was used to quantify the ratio of SUMOylated BCL11B to 930
total BCL11B using the Fujifilm MultiGauge software. The value obtained for BCL11B wt in 931
absence of SUMO2 (lane 5) was set up as 1. (D) The S2D and S2A point mutations do not 932
affect BCL11B interaction with endogenous P300 proteins. HEK293T transfected with the 933
indicated plasmids were subjected to Co-IPs analyses with anti-FLAG antibodies followed by 934
immunoblotting with P300 or FLAG antibodies. The ratio of interacting P300 relative to 935
BCL11B was measured as in panel C. 936
937
Figure 6. Endogenous BCL11B proteins are phosphorylated on Ser2 and SUMOylated in 938
PMA/Ionomycin-activated Jurkat cells. (A) Activation of Jurkat cells with P/I induces Ser2 939
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

35
Phosphorylation of endogenous BCL11B proteins. Jurkat cells were treated with P/I in a time-940
dependent manner and extracts prepared in denaturing conditions were analysed by 941
immunoblotting of two different SDS/PAGE gels. In gel A1, pSer2 BCL11B is shown as a 942
green arrow and a non-specific band as an *. In gel A2, slowly migrating BCL11B-943
SUMOylated species are shown as red arrows. Quantification of total BCL11B proteins in 944
DMSO versus P/I conditions (bottom panel) and of Ser2-phosphorylated and SUMO-945
BCL11B to BCL11B (right panel) were performed with the Fujifilm MultiGauge software. 946
(B) Detection of pSer2 BCL11B in activated Jurkat cells by IP/WB. Jurkat cells were treated 947
with DMSO or activated for 30 minutes and pre-treated or not with the phosphatase inhibitor 948
Okadaic acid. Total cell extracts were immunoprecipitated by BCL11B and immunoblotted 949
with the pSer2 BCL11B specific antibodies. The membrane was striped and reprobed with 950
anti-BCL11B antibodies. The green arrow corresponds to pSer2-BCL11B and the black 951
arrows to the supershift of phosphorylated BCL11B observed with OA. (C) The phosphatase 952
inhibitor Okadaic Acid (OA) favors detection of pSer2 BCL11B and induces a supershift of 953
BCL11B upon P/I activation of Jurkat cells. Cells were treated with DMSO (-) or activated 954
with P/I for 30 minutes and pre-treated or not with OA. Total cell extracts were prepared and 955
immunoblotted as indicated. Green arrows correspond to BCL11B and the black arrows to the 956
supershift of phosphorylated BCL11B observed with OA. (D) The PKC inhibitor Gö6983 957
abolishes Ser2 phosphorylation but not the supershift of BCL11B upon P/I activation. An 958
experiment was conducted essentially as described in panel C) but with pre-treatment with 959
Gö6983. (E) SUMOylation of endogenous BCL11B proteins is not affected by PKC 960
inhibition. Jurkat cells were treated with DMSO or activated for 60 minutes to favor 961
SUMOylation and treated or not with Gö6983 and U0126. Total cell extracts prepared in 962
denaturing conditions were immunoblotted as indicated. 963
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

36
(F) The SUMOylation inhibitor Calyculin A inhibits BCL11B SUMOylation. Jurkat cells 964
were treated with DMSO or activated for 60 minutes and pretreated or not with the 965
SUMOylation inhibitor Calyculin A (Cal A). Cal. A is also a phosphatase inhibitor and thus 966
induces a supershift for BCL11B. Red arrows correspond to SUMOylated forms of BCL11B. 967
(G) The specific SUMOylation inhibitor 2D-08 slightly inhibits BCL11B SUMOylation. A 968
similar experiment was performed with Jurkat cells pretreated or not with 2D-08 for the 969
indicated times. (H) Detection of endogenous BCL11B SUMOylation in Jurkat cells by 970
immunoprecipitation analyses. Jurkat cells were treated with DMSO or P/I for 1 hour to 971
favour SUMOylation (panel A). To detect SUMOylated proteins, the cells were lysed in 1 972
volume 1% SDS. The lysates were immediately boiled for 10 minutes, then diluted with 9 973
volumes of RIPA buffer without SDS and immunoprecipitated with anti-BCL11B antibodies. 974
The immunoprecipitates were then analysed by Western blotting with anti-SUMO2 (upper 975
panel). The SUMOylated BCL11B proteins are indicated by an arrowhead. The membranes 976
were striped and reprobed with anti-BCL11B antibodies. Please note that in contrast with the 977
other gels in this Figure, this gel is a 6% SDS/PAGE which might explain the “ladder-like” 978
aspect of the BCL11B bands. 979
980
Figure 7. Activation of Jurkat cells induces BCL11B down regulation by direct transcriptional 981
repression mediated by KLF4 (A) Prolonged activation of Jurkat cells induces down-982
regulation of BCL11B expression. Jurkat cells were treated with DMSO or activated with P/I 983
for the indicated times. BCL11B mRNAs levels were determined by RT-qPCR analyses. (B) 984
BCL11B proteins are not targeted for degradation by ubiquitination. Jurkat cells were 985
pretreated with different proteasome inhibitors for 1h before activation and activated for 5h as 986
indicated. Total cells extracts were immunoblotted with anti-BCL11B antibodies. (C) KLF4 987
expression is induced in activated Jurkat cells. KLF4 mRNA levels were analyzed by qRT-988
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

37
PCR in the samples used in panel A for BCL11B (D) Schematic drawing of the human 989
BCL11B proximal promoter region. The transcription Start Site (TSS), the conserved potential 990
KLF4 binding site (underlined) and the oligonucleotides used in ChIP-qPCR experiments are 991
shown. (E) KLF4 binds the BCL11B promoter. Chromatin was prepared from DMSO-treated 992
and P/I-activated Jurkat and ChIP analyses performed for KLF4 or IgG at KLF4 binding sites 993
in BCL11B and CXCR4 promoters. GAPDH was used as a nonbinding control. 994
995
Figure 8. BCL11B-mediated repression of IL2 expression through MTA1 or MTA3 996
recruitment in un-stimulated cells is lost during Jurkat cells activation by P/I treatment. (A, 997
B) Activation of human Jurkat CD4+ T cells by P/I induced an increase of IL2 and Id2 998
expression. Jurkat cells were treated with DMSO or activated with P/I for the indicated times. 999
Total RNAs were extracted and RT-qPCR experiments were performed to analyse the 1000
increase of IL2 (A) and Id2 (B) expression levels after activation of Jurkat cells. Due to the 1001
huge induction of IL2 expression after prolonged stimulation, the IL2 expression levels 1002
measured after 2 and 5 hours of treatment are shown with a different scale as Supplementary 1003
Figure 6C (C) Endogenous P300 preferentially interacts with BCL11B in activated Jurkat T 1004
cells. Cells were activated or not with P/I, lysed in IPH buffer and immunoprecipitated with 1005
anti BCL11B. As control, P/I-treated cells were immunoprecipitated with rabbit IgG. The 1006
immunoprecipitates were then analyzed by Western blot with the indicated antibodies. (D) 1007
Schematic drawing of the human Id2 proximal promoter region with the Transcription Start 1008
Site (TSS) and the conserved potential BCL11B direct binding site, TGGGC. The 1009
oligonucleotides used in ChIP-qPCR experiments are shown as arrowheads. (E- I) ChIP 1010
experiments were performed on Jurkat cells chromatin with antibodies against BCL11B, 1011
MTA1, MTA3 and rabbit IgG (panels E-H) or against P300 and mouse IgG (panel I). The 1012
bound material was eluted and analysed by quantitative PCR using primers flanking the US1 1013
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

38
BCL11B binding site in IL2 (19) and the newly identified BCL11B binding in Id2. For each 1014
promoter and antibody, GAPDH was used as an internal nonbinding control. 1015
1016
Figure 9. The PKC inhibitor Gö6983 abolishes the co-repressor-coactivator switch of 1017
BCL11B on the IL2 promoter. (A-B) Jurkat cells were treated with DMSO or activated with 1018
P/I for 30 minutes without or with pre-incubation with the PKC inhibitor Gö6983. RT-qPCR 1019
experiments were performed to analyse the expression levels of BCL11B and IL2. (C-D) 1020
Relative occupancy of BCL11B, MTA1, RbAp48 and P300 on IL2 promoter during Jurkat 1021
activation with or without treatment with the PKC inhibitor Gö6983. These bindings were 1022
assessed by ChIP-qPCR experiments in triplicate before and after activation and inhibitor 1023
treatment, as described in panels A-B. Error bars indicate standard deviation. The ChIP results 1024
for the US1 site in the IL2 promoter are shown on the left whereas those for the GAPDH 1025
promoter used as a nonbinding control are shown on the right. 1026
Figure10. Activation of primary human CD4+ T cells induces phosphorylation of BCL11B 1027
Ser2 and the corepressor-coactivator switch on the IL2 promoter (A) Activation of primary 1028
human CD4+ T cells by P/I or by anti CD3/CD28 antibodies induced down-regulation of 1029
BCL11B expression. RT-qPCR experiments were performed to analyse BCL11B expression. 1030
(B) Analyses of endogenous BCL11B phosphorylation on Ser2 and SUMOylation in 1031
activated human CD4+ T cells. CD4+ T cells were directly lysed in Laemmli buffer, boiled 1032
and analysed by Western blotting with the indicated antibodies, 5h or 24h after P/I- or anti-1033
CD3/CD28-treatment respectively. The red arrowhead corresponds to SUMOylated BCL11B 1034
proteins. (C) Increase of IL2 expression in activated human CD4+ T cells. RT-qPCR 1035
experiments were performed to analyse IL2 expression upon activation with P/I (left panel) or 1036
anti-CD3/CD28 antibodies (right panel). (D-E) Relative occupancy of BCL11B and P300 on 1037
the IL2 promoter during human CD4+ T-cell activation. BCL11B (D) and P300 (E) occupancy 1038
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

39
on the US1 site in the IL2 promoter (19) was analyzed by ChIP-qPCR experiments in human 1039
CD4+ T cells with and without activation. Error bars indicate standard deviation. Similar ChIP 1040
experiments were performed with control IgG. For each antibody, the ChIP results for the 1041
GAPDH promoter used as a nonbinding control are shown. 1042
1043
Figure 11. Working model on the co-repressor/activator switch on BCL11B target genes 1044
during activation of Jurkat or primary human CD4+ T cells. 1: In un-stimulated Jurkat or 1045
primary human CD4+ T cells isolated from PBMCs of healthy donors, BCL11B bound 1046
together with MTA1 and NuRD complex represses transcription of direct target genes such as 1047
IL2 and Id2. 2: Upon activation, phosphorylation of Ser2 in the conserved N-terminal 1048
repression domain allows disruption of the interaction with MTA1 and hence NuRD 1049
repressive complexes while BCL11B remains bound to the IL2 and Id2 promoters. 3: Then, 1050
SUMOylation on Lys679 in the C-terminal end of BCL11B allows the recruitment of P300 to 1051
activate transcription. 4: After 2-5 hours of treatment, a transcriptional down-regulation of 1052
BCL11B by KLF4 would stop and/or limit the IL2 and Id2 activation. 1053
1054
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

B
C D
Dubuissez et al., Figure 1
E F
A
Input 1%
IP MTA3 FLAG-BCL11B
FLAG-MTA3
FLAG-BCL11B
WB :
1 2 3 4
FLAG-MTA3
HEK293T
FLAG-BCL11B
MTA1
FLAG-BCL11B Input 1%
IP FLAG
(BCL11B)
MTA1
WB :
1 2 3 4
Short exp.
Long exp.
HEK293T
* *
* *
* *
IP
WB :
MTA1
BCL11A
Input 1%
1 2
MOLT4
1’ 2’
1 2
MOLT4
1’ 2’
Input 1%
IP
WB :
BCL11B
MTA3
1 2 1’ 2’
WB :
IP
Input 1%
MTA1
BCL11B
MOLT4
MOLT4
IP
WB :
1 2
Input 1%
1’ 2’
BCL11A
MTA3
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Figure 1. BCL11A and BCL11B interact with MTA1 and MTA3. (A) MTA1 interacts with BCL11B.
HEK293T were transfected with MTA1 and BCL11B expression vectors. Whole cells extracts
incubated with anti-FLAG antibodies (IP FLAG) and 1% of each lysate (Input) were
immunoblotted with the indicated antibodies. FLAG-BCL11B co-immunoprecipitate endogenous
MTA1 proteins (lane 3) visualized as a doublet (*). (B) BCL11B interacts with MTA3. A similar
experiment was conducted in HEK293T cells with BCL11B and MTA3 expression vectors. (C)
Endogenous BCL11B and MTA1 proteins interact. Solubilized chromatin fractions were prepared
in duplicate from the same number of MOLT4 cells and 1% was retained for direct analyses by
Western blot (Input). The interaction between BCL11B and MTA1 was evaluated by co-
immunoprecipitation assays. (D) Endogenous BCL11B and MTA3; (E) BCL11A and MTA1 and
(F) BCL11B and MTA3 proteins interact in MOLT4 cells. The same procedure was used for
panels C to E.
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Dubuissez et al., Figure 2
D
A
B C
WB :
MTA1
MTA1
HA
Input 1%
IP c-myc
(MTA1)
+ MTA1
HA
1 2 3 4 5 6 7 8 9 10
FLAG-MTA3
FLAG-MTA3
Input 1%
HA
HA
WB :
+ MTA3
1 2 3 4 5 6 7 8
0
0,2
0,4
0,6
0,8
1
1,2
1 2 3
*** ***
Gal4-NLS-HA 1-20 BCL11B
Gal4-NLS-HA
1-58 BCL11B
Gal4-NLS-HA
Fold
activation
0
0,2
0,4
0,6
0,8
1
1,2
1 2 3
**
**
Gal4-NLS-HA 1-20 S2D
BCL11B
Gal4-NLS-HA
1-20 WT
BCL11B
Gal4-NLS-HA
**
Fold
activation
***
BCL11B 1 894 ZF MSRRKQ
Gal4 1 58 1-58 BCL11B
NLS-HA
1 20 1-20 WT BCL11B
GAL4-NLS-HA 1 20 1-20 S2X BCL11B
MDRRKQ
Gal4
NLS-HA
Gal4
NLS-HA
Gal4
NLS-HA
IP FLAG
(MTA3)
MARRKQ MTRRKQ
S2D S2A S2T
E
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Dubuissez et al., Figure 2 continued
**
0
0,2
0,4
0,6
0,8
1
1,2
gal4 1-20 S2T 1-20 WT
Fold
activation
Gal4-NLS-HA 1-20 WT
BCL11B
Gal4-NLS-HA
1-20 S2T
BCL11B
Gal4-NLS-HA
NS **
G
0
0,2
0,4
0,6
0,8
1
1,2
gal4 1-20 S2A 1-20 S2D 1-20 WT
F F
old
activation
Gal4-NLS-HA 1-20 S2A
BCL11B
Gal4-NLS-HA
1-20 WT
BCL11B
Gal4-NLS-HA
***
** **
1-20 S2D
BCL11B
Gal4-NLS-HA
*** *** **
Figure 2. Ser 2 in the conserved N-terminal motif of BCL11B is essential for its interaction with
MTA1 and MTA3 and for its transcriptional repression activity (A) Schematic drawing of the
BCL11B-Gal4 (DNA-binding domain)-NLS (Nuclear localization signal)-HA fusion proteins. (B)
A phosphomimetic point mutation S2D in the BCL11B N-terminal domain inhibits its interaction
with MTA1. After transfection with the indicated expression vectors, HEK293T cells lysates
were immunoprecipitated with anti-c-myc antibodies. Immunoprecipitated samples [IP c-myc
(MTA1)] and 1% of whole cell extracts (Input) were analyzed by immunoblotting with the
indicated antibodies. (C) The BCL11B-MTA3 interaction is also negatively regulated by the
S2D mutation. A similar experiment was performed in HEK293T with a FLAG-MTA3 plasmid
and the BCL11B-Gal4 chimeras. (D) The conserved BCL11B N-terminal domain represses
transcription. The transcriptional activity of BCL11B-Gal4 constructs was tested by transient
luciferase reporter assays in HEK293T cells. The results represent the mean values from three
independent transfections in triplicate. (E) The S2D phosphomimetic mutation in the BCL11B
N-terminal domain partially inhibits its transcriptional repression potential. A similar luciferase
reporter assay was conducted in HEK293T with the 1-20 BCL11B wild-type (WT) and the S2D
point mutant. (F-G) Repression potential of the various S2X point mutants BCL11B N-terminal
domain. Luciferase reporter assays were conducted with the S2A, S2D, S2T and wt BCL11B-
Gal4 constructs as described above.
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

BCL11B WT
MDRRKQ
BCL11B S2D
MARRKQ
BCL11B S2A
DMSRRKQ
BCL11B DMSRRKQ
1
MSRRKQ
894 ZF
Dubuissez et al., Figure 3
B
Input 1%
IP c-myc
(MTA1) MTA1
FLAG-
BCL11B
MTA1
WB :
1 2 3 4 5 6
FLAG-
BCL11B
HEK293T
D
Input 1%
IP FLAG (BCL11B)
WB :
MTA1
MTA1
FLAG-
BCL11B
1 2 3 4 5 6
FLAG-
BCL11B
HEK293T
FLAG-BCL11B
FLAG-BCL11B
FLAG-MTA3
FLAG-MTA3
WB :
Input 1%
1 2 3 4 5 6
IP MTA3
C HEK293T
A
MTA1
RbAp46
MTA3
FLAG-
BCL11B
Input 1% IP FLAG (BCL11B
1 2 3 4
WB :
1’ 2’ 3’ 4’
HEK293T E
Input 1%
1 2 3
CHD4
HDAC2
RbAp46
MTA1
FLAG-
BCL11B
(BCL11B)
WB :
1’ 2’ 3’
IP FLAG
F HEK293T
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Figure 3. The phosphomimetic S2D mutation in BCL11B inhibits NuRD recruitment. (A) Schematic
drawing of the full-length BCL11B proteins tested. (B) The interaction with MTA1 is impaired by the
S2D phosphomimetic mutation of BCL11B. HEK293T were transfected with the indicated
combination of expression vectors and Co-IP assays followed by immunoblotting with the indicated
antibodies were performed. (C) The BCL11B-MTA3 interaction is also strongly reduced by the S2D
phosphomimetic mutation. A similar Co-IP experiment was realized in HEK293T but with the FLAG-
MTA3 expression vector. Relevant pieces of the membranes were cut (around the 80KDa marker)
to separate the MTA3 and BCL11B proteins and probed with anti-FLAG antibodies. (D) Interaction
of wt, S2A, S2D and DMSRKKQ BCL11B with endogenous MTA1 proteins. Total extracts of
HEK293T transfected with the indicated plasmids were analysed by Co-IP with anti-FLAG
antibodies and imunoblotted with MTA1 and FLAG antibodies. (E-F) Interaction of the S2A and
S2D BCL11B point mutants with endogenous NuRD components. Co-IP experiments followed by
immunoblotting with the indicated antibodies were performed as in Panel D with the S2A and S2D
(Panel E) or S2D (Panel F) BCL11B mutants.
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Dubuissez et al., Figure 4
C HEK293T + FLAG-BCL11B WT
IP FLAG
(BCL11B)
Input 1%
FLAG-BCL11B
pSer2 BCL11B
WB:
1 2 3
pErk1/2
A
IP FLAG
(BCL11B)
Input 1%
pSer2
BCL11B
FLAG-BCL11B
FLAG-BCL11B
pErk1/2
Short exp.
Long exp.
HEK293T + FLAG-BCL11B WT
1 2 3
WB:
B HEK293T +
-
IP FLAG
(BCL11B)
Input 1%
pSer2 BCL11B
FLAG-BCL11B
*
BCL11B S2A
*
BCL11B WT
- WB:
1 2 3 4 5 6
D
100
OA
DMSO - - -
- - -
+ + +
+ + +
HEK293T +
pSer2 BCL11B
FLAG-BCL11B
hsp60
100
1 2 3 4 5 6
Figure 4. Phosphorylation of BCL11B Ser2 in HEK293T cells upon PKC activation. (A) PMA-
induced in vitro phosphorylation of BCL11B Ser2. HEK293T cells transfected with FLAG-BCL11B
were mock-treated (-), incubated with vehicle (DMSO) or with PMA (1mM) for 20 minutes.
Immunoprecipitated samples IP (FLAG) and Input lysates were analyzed by immunoblotting with
antibodies specific for phosphorylated Ser2 (pSer2 BCL11B) and FLAG as control. (B) Specificity of
the anti-pSer2 BCL11B antibodies. A similar experiment was performed to compare in the three
conditions the BCL11B S2A mutant and wt BCL11B. * refers to a non-specific band. (C) PKC is
implicated in BCL11B Ser2 phosphorylation. HEK293T cells were transfected with BCL11B and
activated exactly as in panel A. One plate was pre-incubated with the pan-PKC inhibitor BIMII
before PMA activation. The PKC inhibitor also partially inhibits Erk activation as previously shown
(51). (D) The phosphatase inhibitor Okadaic Acid (OA) allows detection of BCL11B Ser2
phosphorylation in basal conditions. HEK293T cells transfected with the indicated expression
vectors were incubated with DMSO or treated with OA. Total cell lysates were analyzed by
immunoblotting with the indicated antibodies.
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

A
C D
Dubuissez et al., Figure 5
0
12
34
5
S2A S2D WT
- SUMO2
+ SUMO2
SU
MO
-BC
L11B
re
lative
to B
CL
11B
/hsp60
BCL11B
FLAG-BCL11B
FLAG-
BCL11B
0
0,5
1
1,5
2
1 2 3
HEK293T
Inte
racting P
300
rela
tive
to
IP
BC
L11B
2 3 4
Lanes
P300 Input 1%
P300
IP FLAG
(BCL11B)
WB :
1 2 3 4
Short exp.
Short exp.
Long exp.
Long exp.
FLAG-BCL11B
B
HEK293T +
150
100
150
100
SUMO2 - + - + - +
FLAG-
BCL11B
Short exp.
Long exp.
Hsp60
WB :
1 2 3 4 5 6
SUMO
100
150
HEK293T +
WB :
Actin
SUMO 2
SENP 2
FLAG-BCL11B WT
SENP2
- - +
1 2 3
SUMO
pErk 1/2
BCL11B SUMO BCL11B
FLAG
HEK293T +
FLAG-BCL11B WT
100
150
100
150
Erk 2 total
1 2 3 4
WB :
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Figure 5. BCL11B SUMOylation in HEK293T cells is independent of Ser2 Phosphorylation. (A)
BCL11B is SUMOylated. Total cells extracts of HEK293T transfected with the indicated
combinations of BCL11B, SUMO2 and the deSUMOylase SENP2 were prepared in denaturing
conditions and immunoblotted with indicated antibodies. The arrowhead corresponds to
SUMOylated forms of BCL11B (B) The PKC pathway does not impinge on BCL11B SUMOylation in
contrast with the ERK pathway. HEK293T cells were transfected with BCL11B and pre-incubated
with the PKC inhibitor BIMII or with the Erk1/2 inhibitor, U0126 before PMA activation. The PKC
inhibitor also partially inhibits Erk activation as previously shown (51). Total cells extracts prepared
in denaturing conditions were immunoblotted as indicated. (C) The S2D and S2A point mutations
have no significant impact on BCL11B SUMOylation. HEK293T were transfected as indicated,
lysed in denaturing conditions and immunoblotted with anti FLAG antibodies to detect BCL11B and
its SUMOylated forms. Hsp60 was used to quantify the ratio of SUMOylated BCL11B to total
BCL11B using the Fujifilm MultiGauge software. The value obtained for BCL11B wt in absence of
SUMO2 (lane 5) was set up as 1. (D) The S2D and S2A point mutations do not affect BCL11B
interaction with endogenous P300 proteins. HEK293T transfected with the indicated plasmids were
subjected to Co-IPs analyses with anti-FLAG antibodies followed by immunoblotting with P300 or
FLAG antibodies. The ratio of interacting P300 relative to BCL11B was measured as in panel C.
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

0
0,5
1
1,5
2
0 5' 30' 60' 120' 300'
Dubuissez et al., Figure 6
pSer2 BCL11B
BCL11B
BCL11B
Short
exp.
Long
exp.
pErk 1/2
Erk 2 total
DMSO P/I
Jurkat cells
(min) WB: 0 5 30 60 120 300 0 5 30 60 120 300
1 2 3 4 5 6 7 8 9 10 11 12
A
100
150
A1
A2 2 1
*
0
1
2
3
4
0 5' 30' 60' 120' 300'
1
2SUMO-BCL11B
0
50
100
150
0 5' 30' 60' 120' 300'
Rela
tive t
o B
CL11B
/Erk
2 a
nd
norm
alis
ed t
o D
MS
O
pSer2 BCL11B
Treatment (min)
BCL11B proteins level
Treatment (min)
BC
L1
1B
(S
ho
rt
exp
.) / E
rk2
to
tal
DMSO
P/I
D
B
C Pre-treatment 30’
Activation 30’
Jurkat cells
P/I
OA -
WB:
1 2 3 4
100
150
100
150
BCL11B
pErk 1/2
Short
exp.
Long
exp.
pSer2
BCL11B
Erk 2 total
+ + -
- - + +
100
100 Long exp.
Short exp.
100
100
150
100
1 2 3 4 5
100
150
pSer2 BCL11B
pErk 1/2
Erk 2 total
BCL11B
WB:
Short exp. Long exp.
Pre-treatment 30’
Activation 30’
Pre-treatment 15’
P / I
OA
Gö6983
-
-
- - -
- -
+ +
+ +
+ + + +
Jurkat cells
100
150 100
150
pSer2
BCL11B
Erk 2 total
BCL11B
pErk 1/2
WB:
*
Input 1%
100
100
100
1 2 3 4
Pre-treatment 30’
Activation 30’
Jurkat cells
P/I
OA - + + -
- - + +
BCL11B
IP:
BCL11B
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

E
pErk 1/2
BCL11B 100
150
1 2 3 4
Erk2 total
Jurkat cells
P/I
U0126
Gö6983
-
Pre-treatment 15’
Activation 60’ + + +
- - - +
+ - - -
WB: 2 1
F
H
WB :
Input 1.5%
P/I 60’ + - + + - +
BCL11B
SUMO2
Long
exp.
Short
exp.
pErk 1/2
Actin
100
150
100
150
100
150
Jurkat cells
IP
1 2 3 1’ 2’ 3’
G
Pre-treatment 30’
Activation 60’
100
150 BCL11B
pErk 1/2
Jurkat cells
P/I
Cal. A -
WB:
Erk 2 total
1 2 3
- +
- + +
2 1
Dubuissez et al., Figure 6 continued
pErk 1/2
Erk 2 total
1 2 3 4 5 6 7 8 9
100
150 2 1 BCL11B
WB:
1h 2h 4h
Jurkat cells
DMSO
1h 2h 4h
DMSO
1h 2h 4h
2D-08
DMSO P/I P/I Activation 60’
Pre-treatment
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Figure 6. Endogenous BCL11B proteins are phosphorylated on Ser2 and SUMOylated in
PMA/Ionomycin-activated Jurkat cells. (A) Activation of Jurkat cells with P/I induces Ser2
Phosphorylation of endogenous BCL11B proteins. Jurkat cells were treated with P/I in a time-
dependent manner and extracts prepared in denaturing conditions were analysed by
immunoblotting of two different SDS/PAGE gels. In gel A1, pSer2 BCL11B is shown as a green
arrow and a non-specific band as an *. In gel A2, slowly migrating BCL11B-SUMOylated species
are shown as red arrows. Quantification of total BCL11B proteins in DMSO versus P/I conditions
(bottom panel) and of Ser2-phosphorylated and SUMO-BCL11B to BCL11B (right panel) were
performed with the Fujifilm MultiGauge software. (B) Detection of pSer2 BCL11B in activated
Jurkat cells by IP/WB. Jurkat cells were treated with DMSO or activated for 30 minutes and pre-
treated or not with the phosphatase inhibitor Okadaic acid. Total cell extracts were
immunoprecipitated by BCL11B and immunoblotted with the pSer2 BCL11B specific antibodies.
The membrane was striped and reprobed with anti-BCL11B antibodies. The green arrow
corresponds to pSer2-BCL11B and the black arrows to the supershift of phosphorylated BCL11B
observed with OA. C) The phosphatase inhibitor Okadaic Acid (OA) favors detection of pSer2
BCL11B and induces a supershift of BCL11B upon P/I activation of Jurkat cells. Cells were treated
with DMSO (-) or activated with P/I for 30 minutes and pre-treated or not with OA. Total cell
extracts were prepared and immunoblotted as indicated. Green arrows correspond to BCL11B and
the black arrows to the supershift of phosphorylated BCL11B observed with OA. (D) The PKC
inhibitor Gö6983 abolishes Ser2 phosphorylation but not the supershift of BCL11B upon P/I
activation. An experiment was conducted essentially as described in panel C) but with pre-
treatment with Gö6983. (E) SUMOylation of endogenous BCL11B proteins is not affected by PKC
inhibition. Jurkat cells were treated with DMSO or activated for 60 minutes to favor SUMOylation
and treated or not with Gö6983 and U0126. Total cell extracts prepared in denaturing conditions
were immunoblotted as indicated.
(F) The SUMOylation inhibitor Calyculin A inhibits BCL11B SUMOylation. Jurkat cells were treated
with DMSO or activated for 60 minutes and pretreated or not with the SUMOylation inhibitor
Calyculin A (Cal A). Cal. A is also a phosphatase inhibitor and thus induces a supershift for
BCL11B. Red arrows correspond to SUMOylated forms of BCL11B. (G) The specific SUMOylation
inhibitor 2D-08 slightly inhibits BCL11B SUMOylation. A similar experiment was performed with
Jurkat cells pretreated or not with 2D-08 for the indicated times. (H) Detection of endogenous
BCL11B SUMOylation in Jurkat cells by immunoprecipitation analyses. Jurkat cells were treated
with DMSO or P/I for 1 hour to favour SUMOylation (panel A). To detect SUMOylated proteins, the
cells were lysed in 1 volume 1% SDS. The lysates were immediately boiled for 10 minutes, then
diluted with 9 volumes of RIPA buffer without SDS and immunoprecipitated with anti-BCL11B
antibodies. The immunoprecipitates were then analysed by Western blotting with anti-SUMO2
(upper panel). The SUMOylated BCL11B proteins are indicated by an arrowhead. The membranes
were striped and reprobed with anti-BCL11B antibodies. Please note that in contrast with the other
gels in this Figure, this gel is a 6% SDS/PAGE which might explain the “ladder-like” aspect of the
BCL11B bands.
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Dubuissez et al., Figure 7
B
0
0,2
0,4
0,6
0,8
1
1,2
0 5 30 60 120 300
A BCL11B mRNA levels in Jurkat cells
Re
lative
exp
ressio
n
Treatment (min)
NS NS
NS NS ** ***
DMSO
P/I
0
10
20
30
40
50
60
70
80
0 5 30 60 120 300
C
Treatment (min)
KLF4 mRNA levels in Jurkat cells
Re
lative
exp
ressio
n
NS NS
**
**
** ***
D
100 bp
ATG 5’ 3’ BCL11B
TSS
Mu:TTTGGGTTTTTGAGGGGGCGGGGTGAGGGGGTAC
Hu:TTTGGGTTTTTTGAGGGGGCGGGGTGAGGGGGTA
KLF4 binding site
Actin
P/I
BCL11B 100
150
WB :
Jurkat cells
MG132
ALLN
Lactacystin - - - + - - - - - +
+ - - - - -
Pre-treatment 60’
Activation 300’ - + + + +
DMSO
P/I
E CXCR4 promoter GAPDH promoter BCL11B promoter
0
0,2
0,4
0,6
0,8
1
1,2
KLF4 IgG
0
0,2
0,4
0,6
0,8
1
1,2
KLF4 IgG
0
0,2
0,4
0,6
0,8
1
1,2
KLF4 IgG
% In
pu
t
*
NS NS NS NS
% In
pu
t
% In
pu
t
ChIP KLF4 ChIP IgG ChIP KLF4 ChIP IgG ChIP KLF4 ChIP IgG
* DMSO 300’
P/I 300’
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Figure 7. Activation of Jurkat cells induces BCL11B down regulation by direct
transcriptional repression mediated by KLF4 (A) Prolonged activation of Jurkat cells
induces down-regulation of BCL11B expression. Jurkat cells were treated with DMSO or
activated with P/I for the indicated times. BCL11B mRNAs levels were determined by RT-
qPCR analyses. (B) BCL11B proteins are not targeted for degradation by ubiquitination.
Jurkat cells were pretreated with different proteasome inhibitors for 1h before activation
and activated for 5h as indicated. Total cells extracts were immunoblotted with anti-
BCL11B antibodies. (C) KLF4 expression is induced in activated Jurkat cells. KLF4 mRNA
levels were analyzed by qRT-PCR in the samples used in panel A for BCL11B (D)
Schematic drawing of the human BCL11B proximal promoter region. The transcription Start
Site (TSS), the conserved potential KLF4 binding site (underlined) and the oligonucleotides
used in ChIP-qPCR experiments are shown. (E) KLF4 binds the BCL11B promoter.
Chromatin was prepared from DMSO-treated and P/I-activated Jurkat and ChIP analyses
performed for KLF4 or IgG at KLF4 binding sites in BCL11B and CXCR4 promoters.
GAPDH was used as a nonbinding control.
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

0
5
10
15
0 5 30 60 120 300
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
ChIP BCL11B ChIP MTA1 ChIP IgG
D
100 bp
ATG 5’ 3’ Id2
TSS
Mu: ATTTGCCACCCCAGCTGGGCTCTCGCCCGCTCC
Hu: ATTTACCACCCCAGCTGGGCTCGCGCCC----C
BCL11 B binding site IL2 promoter
GAPDH promoter G
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
ChIP BCL11B ChIP MTA1 ChIP IgG
Id2 promoter F
ChIP BCL11B ChIP MTA1 ChIP IgG
% In
pu
t
NS NS
NS
D 30’
P/I 30’
ChIP IgG ChIP BCL11B ChIP MTA1
NS
NS
**
% In
pu
t
D 30’
P/I 30’
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
ChIP BCL11B ChIP MTA1 ChIP IgG
% In
pu
t
ChIP BCL11B ChIP MTA1 ChIP IgG
NS
NS
*** D 30’
P/I 30’
E
Dubuissez et al., Figure 8
**
P300
BCL11B
MTA1
P300
MTA1
BCL11B
IP
Input 2%
IgG
P/I 30’
BCL11B
WB : + - +
Jurkat cells
1 2 3
0
2
4
6
0 5 30
A IL2 mRNA levels in Jurkat cells R
ela
tive
exp
ressio
n
0 30 5
NS NS
Treatment (min)
DMSO
P/I
B Id2 mRNA levels in Jurkat cells
Re
lative
exp
ressio
n
Treatment (min)
DMSO
P/I ***
NS NS NS NS
0 30 60 5 120 300
**
C
0
0,5
1
1,5
2
1 2
0
0,5
1
1,5
2
1 2
D 30’
P/I 30’
H
% In
pu
t
ChIP MTA3 ChIP IgG
IL2 promoter
NS
*
ChIP MTA3 ChIP IgG
**
NS
Id2 promoter
ChIP MTA3 ChIP IgG
GAPDH promoter
NS % In
pu
t
% In
pu
t
NS
0
0,5
1
1,5
2
1 2
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Dubuissez et al., Figure 8 continued
D 30’
P/I 30’
I
0
1
2
3
4
5
6
7
P300 308 IgG 206
% In
pu
t
ChIP P300 ChIP IgG
IL2 promoter
NS
*
0
1
2
3
4
5
6
7
1 2ChIP P300 ChIP IgG
*
NS
Id2 promoter
0
1
2
3
4
5
6
7
1 2ChIP P300 ChIP IgG
GAPDH promoter
NS
*
% In
pu
t
% In
pu
t
Figure 8. BCL11B-mediated repression of IL2 expression through MTA1 or MTA3
recruitment in un-stimulated cells is lost during Jurkat cells activation by P/I treatment. (A,
B) Activation of human Jurkat CD4+ T cells by P/I induced an increase of IL2 and Id2
expression. Jurkat cells were treated with DMSO or activated with P/I for the indicated
times. Total RNAs were extracted and RT-qPCR experiments were performed to analyse
the increase of IL2 (A) and Id2 (B) expression levels after activation of Jurkat cells. Due to
the huge induction of IL2 expression after prolonged stimulation, the IL2 expression levels
measured after 2 and 5 hours of treatment are shown with a different scale as
Supplementary Figure 6C (C) Endogenous P300 preferentially interacts with BCL11B in
activated Jurkat T cells. Cells were activated or not with P/I, lysed in IPH buffer and
immunoprecipitated with anti BCL11B. As control, P/I-treated cells were
immunoprecipitated with rabbit IgG. The immunoprecipitates were then analyzed by
Western blot with the indicated antibodies. (D) Schematic drawing of the human Id2
proximal promoter region with the Transcription Start Site (TSS) and the conserved
potential BCL11B direct binding site, TGGGC. The oligonucleotides used in ChIP-qPCR
experiments are shown as arrowheads. (E- I) ChIP experiments were performed on Jurkat
cells chromatin with antibodies against BCL11B, MTA1, MTA3 and rabbit IgG (panels E-H)
or against P300 and mouse IgG (panel I). The bound material was eluted and analysed by
quantitative PCR using primers flanking the US1 BCL11B binding site in IL2 (19) and the
newly identified BCL11B binding in Id2. For each promoter and antibody, GAPDH was used
as an internal nonbinding control.
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

0
1
2
3
4
1 2
Dubuissez et al., Figure 9
0
0,5
1
1,5
2
DMSO PMA/Iono Gö6983+PMA/Iono
IL2 mRNA levels in Jurkat cells
Re
lative
exp
ressio
n
* NS
**
A B
0
0,2
0,4
0,6
0,8
1
1,2
DMSO PMA/Iono Gö6983+PMA/Iono
BCL11B mRNA levels in Jurkat cells R
ela
tive
exp
ressio
n NS NS
NS
IL2 promoter
% In
pu
t
NS NS
NS NS
DMSO 30’
P/I 30’
Gö6983 15’+ P/I 30’
GAPDH promoter C
ChIP BCL11B ChIP IgG
D
0
1
2
3
4
5
6
MTA1 RbAp48 P300 IgG
% In
pu
t
IL2 promoter
*
**
NS NS
** *
* **
0
1
2
3
4
5
6
MTA1 RbAp48 P300 IgG
NS NS
NS NS
NS NS
NS NS
GAPDH promoter
% In
pu
t
DMSO 30’
P/I 30’
Gö6983 15’+ P/I 30’
ChIP MTA1 ChIP IgG ChIP RbAp48 ChIP P300 ChIP MTA1 ChIP IgG ChIP RbAp48 ChIP P300
DMSO 30’ P/I 30’ Gö6983+P/I 30’ DMSO 30’ P/I 30’ Gö6983+P/I 30’
0
1
2
3
4
1 2 ChIP BCL11B ChIP IgG
% In
pu
t NS NS
NS NS
Figure 9. The PKC inhibitor Gö6983 abolishes the co-repressor-coactivator switch of
BCL11B on the IL2 promoter. (A-B) Jurkat cells were treated with DMSO or activated with
P/I for 30 minutes without or with pre-incubation with the PKC inhibitor Gö6983. RT-qPCR
experiments were performed to analyse the expression levels of BCL11B and IL2. (C-D)
Relative occupancy of BCL11B, MTA1, RbAp48 and P300 on IL2 promoter during Jurkat
activation with or without treatment with the PKC inhibitor Gö6983. These bindings were
assessed by ChIP-qPCR experiments in triplicate before and after activation and inhibitor
treatment, as described in panels A-B. Error bars indicate standard deviation. The ChIP
results for the US1 site in the IL2 promoter are shown on the left whereas those for the
GAPDH promoter used as a nonbinding control are shown on the right.
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

0
2
4
6
8
10
12
1 2
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
1 2
24h medium
24h CD3/CD28
ChIP BCL11B ChIP IgG 0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
1 2
% In
put
IL2 promoter
NS
NS
ChIP BCL11B ChIP IgG
NS
NS
GAPDH promoter
Dubuissez et al., Figure 10
D
C IL2 mRNA levels in LT CD4+ cells
0
1000
2000
3000
4000
5000
CD4 5h CD4 5h
PMA/iono
0
10
20
30
40
50
CD4 24h CD4 24h
CD3/CD28
Re
lative
exp
ressio
n
24h
medium
24h
CD3/CD28
5h
medium
5h P/I
** **
0
0,2
0,4
0,6
0,8
1
1,2
1,4
CD4 5h CD4 5h
PMA/iono
CD4 24h CD4 24h
CD3/CD28
Re
lative
exp
ressio
n
5h
medium
5h P/I 24h
medium
24h
CD3/CD28
A BCL11B mRNA levels in LT CD4+ cells
** *
B
24h medium
24h CD3/CD28
% In
put
0
2
4
6
8
10
12
1 2
GAPDH promoter IL2 promoter
ChIP P300 ChIP IgG
*
NS
ChIP P300 ChIP IgG
* NS
E
LT CD4+ cells
pSer2 BCL11B
WB:
P/I 5h
CD3/CD28 24h - - - +
Short exp.
Long exp.
- + - -
actin
BCL11B
0
1
2
1 2 3 4BC
L11B
/actin
80
80
80
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Figure 10. Activation of primary human CD4+ T cells induces phosphorylation of BCL11B
Ser2 and the corepressor-coactivator switch on the IL2 promoter (A) Activation of primary
human CD4+ T cells by P/I or by anti CD3/CD28 antibodies induced down-regulation of
BCL11B expression. RT-qPCR experiments were performed to analyse BCL11B expression.
(B) Analyses of endogenous BCL11B phosphorylation on Ser2 and SUMOylation in activated
human CD4+ T cells. CD4+ T cells were directly lysed in Laemmli buffer, boiled and analysed
by Western blotting with the indicated antibodies, 5h or 24h after P/I- or anti-CD3/CD28-
treatment respectively. The red arrowhead corresponds to SUMOylated BCL11B proteins.
(C) Increase of IL2 expression in activated human CD4+ T cells. RT-qPCR experiments were
performed to analyse IL2 expression upon activation with P/I (left panel) or anti-CD3/CD28
antibodies (right panel). (D-E) Relative occupancy of BCL11B and P300 on the IL2 promoter
during human CD4+ T-cell activation. BCL11B (D) and P300 (E) occupancy on the US1 site in
the IL2 promoter (19) was analyzed by ChIP-qPCR experiments in human CD4+ T cells with
and without activation. Error bars indicate standard deviation. Similar ChIP experiments were
performed with control IgG. For each antibody, the ChIP results for the GAPDH promoter
used as a nonbinding control are shown.
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

IL2 repression
NuRD
BCL11B
MTA 1
X
1
IL2, Id2
IL2, Id2 repression BCL11B IL2, Id2
NuRD
MTA 1 X
X
2 PKC
Stop IL2, Id2 repression
P
Ser2
30’
Time scale Inactivated LT CD4+
TCR
Activation
Activated LT CD4+
BCL11B
Downregulation BCL11B
by KLF4
Stop IL2, Id2 activation
4 2H
Gène cible IL2, Id2
IL2, Id2 activation
3 P
Ser2
30’ p300
BCL11B
Dubuissez et al., Figure 11
Figure 11. Working model on the co-repressor/activator switch on BCL11B target
genes during activation of Jurkat or primary human CD4+ T cells. 1: In un-stimulated
Jurkat or primary human CD4+ T cells isolated from PBMCs of healthy donors,
BCL11B bound together with MTA1 and NuRD complex represses transcription of
direct target genes such as IL2 and Id2. 2: Upon activation, phosphorylation of Ser2 in
the conserved N-terminal repression domain allows disruption of the interaction with
MTA1 and hence NuRD repressive complexes while BCL11B remains bound to the IL2
and Id2 promoters. 3: Then, SUMOylation on Lys679 in the C-terminal end of BCL11B
allows the recruitment of P300 to activate transcription. 4: After 2-5 hours of treatment,
a transcriptional down-regulation of BCL11B by KLF4 would stop and/or limit the IL2
and Id2 activation.
on April 2, 2018 by guest
http://mcb.asm
.org/D
ownloaded from