Embed Size (px)
Citation preview

he genus Lepiota (Pers.: Fr.) S.F. Gray, with its sa-tellite genera, Cystolepiota Singer, PulverolepiotaBon, and Melanophyllum Velen., forms a monophy-
letic group within the family Agaricaceae (VELLINGA 2003).This group also includes Coprinus comatus and its allies.However, Chamaemyces fracidus, often assigned to the sametribus as Lepiota, viz. tribus Lepioteae, is excluded from it andappears to be basal to the family Agaricaceae.
The family as a whole is a widely distributed monophy-letic group of saprotrophic fungi, which display a huge di-versity in spore colour, and structure of the pileus covering2.Recently it has been shown that secotioid (e.g. Endoptychumagaricoides, Podaxis pistillaris, and Longula texensis) andgasteroid taxa (Lycoperdaceae and Tulostomataceae) alsobelong to this family (VELLINGA, DE KOK & BRUNS 2003;KRÜGER et al. 2001; MONCALVO et al. 2002).
Morphologically, the clade to which Lepiota s.l. belongs,comprises species with a regular lamella trama, and in mostcases with clamp-connections. Some authors, e.g. BON (1993),included Melanophyllum with coloured spores in the same tri-bus as Lepiota, whereas other authors placed Melanophyllumclose to Agaricus L.: Fr., e.g. KERRIGAN (1986), SINGER (1986)and WASSER (2002). Generally speaking, the other lineages inthe family either have pale spores and a trabecular lamella
trama, or brown spores and a regular lamella trama, or aresecotioid or gasteroid. Clamp-connections are rare in thosegroups.
Genus delimitations and the number of genera within tribusLepioteae have been hotly disputed over the last decades. Avery wide concept of the genus Lepiota, close to the originalFriesian concept (FRIES 1821) is still used by many Americanauthors of popular guide books (e.g. ARORA 1986; LINCOFF
1981); Lepiota in their sense includes the genera Cystolepiota,Lepiota, Leucoagaricus, Leucocoprinus, and Macrolepiota ofother classifications. A much more restricted concept of thegenus Lepiota has been applied by recent European authors(e.g. BON 1993; CANDUSSO & LANZONI 1990; VELLINGA
2001a), and by SINGER (1986). BON (1993) applied the nar-rowest genus concept of all, excluding members of sect. Echi-natae and transferring them to a separate genus, Echinoderma(Locq. ex Bon) Bon. The genus Lepiota, in the rest of this pa-per, is taken in the sense of VELLINGA (2001a), i.e. includingsect. Echinatae, but excluding Chamaemyces, Cystolepiota,Melanophyllum, and Pulverolepiota.
JOHNSON (1999) and JOHNSON & VILGALYS (1998) eluci-dated the phylogenetic relationships of Lepiota s.l. but usedonly seven species to represent Lepiota s. str. and Cystolepiota.On this limited basis they concluded that Lepiota consists ofan assemblage of paraphyletic lineages, with the two Cysto-lepiota species and L. aspera (as L. acutesquamosa) forminga monophyletic clade.
Coprinus comatus (O.F. Müll.: Fr.) Pers. and C. sterqui-linus (Fr.: Fr.) Fr., and Montagnea arenarius (DC.) Zellerwere included in their studies, and were placed either in themiddle of Lepiota (based on ITS-sequences), or paraphyleti-cally between Lepiota and the Leucoagaricus/Leucocoprinus
Phylogeny of Lepiota (Agaricaceae) – Evidence from nrITS andnrLSU sequences
Else C. VELLINGA1
The phylogeny of Lepiota, Cystolepiota, Melanophyllum, Coprinus comatus, and Chamaemyces fracidus was examinedwith molecular techniques, and the consequences for morphological classification were considered. This group (excludingChamaemyces fracidus and Coprinus comatus and allies) can be split up into several clades, which do not correspond tocurrently recognized genera, subgenera and sections. Structure of the pileus covering appears to be a key morphologicalcharacter; colour and shape of the spores do not reflect the new phylogeny. Lepiota sect. Ovisporae is not monophyletic,and a separate genus for Cystolepiota, Lepiota sect. Echinatae, Melanophyllum and Pulverolepiota combined seems war-ranted. More research, especially on developmental morphology and on more tropical taxa, will be necessary to solidly erecta new classification.
Mycological Progress 2(4): 305–322, November 2003 305
1 Department of Plant and Microbial Biology, University of Californiaat Berkeley, 111 Koshland Hall, Berkeley CA 94720-3102, U.S.A.,[email protected]
2 The neutral term pileus covering is used as it is not known whetherthe covering layers are developmentally of velar origin or a real pi-leipellis.
© DGfM 2003
T

306 Phylogeny of Lepiota
© DGfM 2003
assemblage (JOHNSON 1999; JOHNSON & VILGALYS 1998) (seealso REDHEAD et al. (2001) on the position of Coprinus co-matus and its allied species). These taxa are morphologicallyquite different from the rest of the group; the spores are black,with a germ pore, the basidia are of different length, and se-parated from each other by pavement cells, and the lamellaeare autolytic. The hymenium characters and the presence of agerm pore in the spores hint at a relationship with Leucoco-prinus Pat., but that is not supported by the molecular data.
The genus Lepiota embraces species with quite variedtypes of pileus covering, ranging from a hymeniderm, a cutis,a trichoderm to a special type of epithelium (rounded cells inchains which are agglutinated to form acute pyramidal warts).A second group of very variable characters involves the spores,viz. their shape and size, chemical reactions and the numberof nuclei per spore. Most striking is the diversity in sporeshape, from broadly ellipsoid and ellipsoid to spurred, to fusi-form or ‘penguin-shaped’, resembling typical bolete spores.Combinations of spore shape and the structure of the pileuscovering, have been used in the past to distinguish at least fivegroups within the genus, on subgenus and genus levels (e.g.BON 1993) or on section level (e.g. SINGER 1986; VELLINGA
2001a). Older infrageneric classifications, like KÜHNER’s(1936) are more difficult to compare with present day ones,as tribus Leucocoprineae and the genus Cystolepiota were stillincluded. BON’s classification is slightly deviant, as he reco-gnized the groups at different ranks than other systematists,and he used less known names for most of them. It should bekept in mind that the classifications especially below genuslevel, were all based on European and other temperate mate-rial. The systematic position of the taxa used in the presentstudy according to BON’s classification (1993) is displayed inTable 1, as his ideas are widely followed by European authors.
VELLINGA & HUIJSER (1999) used ITS-sequences to showthat Lepiota cristata (Bolt.: Fr.) Kumm., a species characte-rized by spurred spores and a hymenidermal pileus covering,is more closely related to species with ellipsoid spores and ahymenidermal pileus covering, than to spurred-spored taxawhich have a trichodermal pileus covering. This is an indica-tion that the spurred spore might have evolved several times,but the hymenidermal pileus covering only once.
ITS sequences have also proven to be a powerful tool toreveal specific and infraspecific variation that was hitherto un-recognized (VELLINGA 2001b on L. magnispora and L.spheniscispora; VELLINGA 2001c on L. cristata and L. casta-neidisca).
The present study sets out to explore the circumscriptionof the genus Lepiota itself and its satellite genera, and to eva-luate and test the existing morphological infrageneric classi-fications, in which so much emphasis is placed on spore mor-phology and colour. For these purposes an ITS data set, and acombined ITS-LSU data set were used, encompassing a widesample of species. The ingroup comprised 81 species, withfour other Agaricaceae for comparison, viz. Agaricus subru-
tilescens (Kauffman) Hotson & Stuntz, Macrolepiota procera(Scop.: Fr.) Singer, Chlorophyllum rachodes (Vittad.) Vel-linga, and Leucoagaricus leucothites (Vittad.) Wasser. Theoutgroup comprised two species from outside the Agarica-ceae: Limacella glioderma (Fr.: Fr.) Maire and Pseudobaeo-spora pyrifera Bas & L.G. Krieglst.
The new data may help us understand the evolution of se-veral characters, suggest hypotheses concerning speciationand distribution of the taxa, and indicate some fruitful direc-tions for further morphological and molecular studies.
Background information – genera involved
Lepiota (Pers.: Fr.) Gray, Natural Arrangement of BritishPlants 1: 601. 1821.
FRIES (1821) gave a wide definition of the genus, which in-cluded squamose and annulate white-spored species: Cysto-derma, Limacella, and all the more recent split-offs from thegenus, like Leucocoprinus, Leucoagaricus, Macrolepiota, andCystolepiota. The present definition (VELLINGA 2001a) is muchnarrower, while still allowing for variation, and is mainlybased on microscopical characters: pileus and stipe contextnot confluent, universal and partial veils present, lamellae free,spore print white to cream, microscopically with (rarely with-out) clamp connections, dextrinoid (rarely non-dextrinoid),non-metachromatic (rarely metachromatic), bi-nucleate (rare-ly uni-nucleate) generally non-ornamented spores, without agerm pore, with pileus covering made up of either long elements(trichodermally arranged or in chains and radially adnate orascending), or narrowly clavate elements (hymeniderm-like),regular lamella trama.
The following infrageneric groups are generally recognized:i. species with fusiform to penguin-shaped spores, and a
trichoderm as pileus covering – Section Lepiota.ii. species with spurred spores, and a trichoderm – Section
Stenosporae.iii. species with ellipsoid spores and a trichoderm – Section
Ovisporae (with two subsections: subsect. Helveolinaewith a trichoderm made up only of long, cylindrical ele-ments, and subsect. Felinae with a mixed trichoderm madeup of both long, cylindrical and short, clavate elements).
iv. species with a hymeniderm and ellipsoid or spurred spo-res – Several names are available for this group, on sectionlevel sect. Lilaceae is the oldest. BON (1993) recognizedtwo sections, sect. Integrellae with a pileus covering whichdoes not break open into squames, and mainly uni-nucleate spores, and sect. Lilaceae with a pileus coveringwhich does break open into squames, and mainly bi-nucleate spores. Species with spurred spores are sometimesaccommodated into a section of their own.
v. species with acute warts on the pileus, made up of roundelements – Section Echinatae / genus Echinoderma (seebelow).

Mycological Progress 2(4) / 2003 307
© DGfM 2003
Lepiota fuscovinacea is placed in its own section Fuscovinaceaeby all authors, as it has a unique combination of characters:no clamp-connections, uni-nucleate spores, and a cutis-likepileus covering.
Lepiota parvannulata and L. nigrescentipes are assignedto sect. Helveolae subsect. Parvannulatae in BON’s classifi-cation (1993), and were tentatively placed in sect. Ovisporaesubsect. Helveolinae by VELLINGA (2001a).
SINGER’s classification (1986) is different in that he reco-gnized sect. Amyloideae for L. lignicola P. Karst., which is nowaccommodated in Leucopholiota (Romagn.) O.K. Mill., T.J.Volk & Bessette. Furthermore, SINGER (1986) recognized sect.Anomalae, for species without clamp-connections – a very ar-tificial group, with species belonging to Leucoagaricus as wellas to other sections within Lepiota – and sect. Amylosporae forspecies with amyloid (non-dextrinoid) spores.
Melanophyllum Velen., Ceské Houby: 569. 1921.
This small genus of currently three species was erected to ac-commodate species with a pileus covering made up of globosecells, and spores which change from green when fresh to blackwhen dry. Nowadays, species with persistent green spores areincluded, and the spores of all species are small and rough (war-ted when observed with Scanning Electron Microscope).
Lepiotula (Maire) Locq. ex Horak, Synopsis Generum Aga-ricalium: 337. 1968.
Lepiota species with spurred spores, disregarding the type ofpileus covering, were combined in this genus.
Cystolepiota Singer in Singer & Digilio in Lilloa 25: 281.('1951') 1952.
Cystolepiota is a genus for species with generally small ba-sidiocarps which have a veil composed of loosely arrangedglobose cells. The spores are uni-nucleate, and in most spe-cies non-dextrinoid. Clamp-connections are present in mostspecies. This concept was amended by KNUDSEN (1978) to ac-commodate Lepiota sect. Echinatae, and amended again toremove that section (KNUDSEN 1980). VELLINGA (1992) in-cluded L. pulverulenta, with inflated, non-globose, looselyarranged velar cells, bi-nucleate spores, and without clamp-connections, in Cystolepiota.
Echinoderma (Locq. ex Bon) Bon in Documents Mycolo-giques 21 (82): 61. 1991.
This genus was split off from Lepiota for the species withrounded to ellipsoid elements in chains, arranged to formacute to pyramidal warts on the pileus. The main differencesfrom Cystolepiota are the bi-nucleate, dextrinoid spores, andthe presence of so-called intermediate cells in the pileus co-vering (intermediate between globose and cylindrical) (KNUD-SEN 1980). Developmentally, a distinct ‘metablematogenic’layer stands out between the veil and the pileus trama (GREIS
1937; terminology according to CLÉMENÇON (1997)), a layerwhich is absent in Cystolepiota.
Pulverolepiota Bon in Documents Mycologiques 22 (88):30. 1993.
This genus was erected for L. pulverulenta, a species resemb-ling Cystolepiota, but differing in the elongate inflated elementsof the pileus covering, the absence of clamp connections, andthe bi-nucleate, rough spores (versus uni-nucleate and in mostcases smooth spores in Cystolepiota). The American speciesL. petasiformis Murrill belongs here as well, though BON
(1993) put it together with L. pseudogranulosa (Berk. & Br.)Sacc. in section Fuscovinaceae.
Leucopholiota (Romagn.) O.K. Mill., T.J. Volk & Bessettein Mycologia 88: 138. 1996.
This monotypic genus harbours L. decorosa (Peck) O.K.Mill., T.J. Volk & Bessette, a species treated by KNUDSEN
(1980) and SINGER (1986) (as Lepiota lignicola) in Lepiotasect. Amyloideae. HARMAJA (2002) erected the genus Amy-lolepiota Harmaja for L. lignicola, disputing the synonymyof L. decorosa and L. lignicola. He considered it closely re-lated to Lepiota.
It differs from Lepiota species in the gelatinized hyme-nophoral trama, and the amyloid thin-walled spores (VEL-LINGA 2003). MILLER, VOLK & BESSETTE (1996) placed thegenus in the Tricholomataceae, and a position outside the fa-mily Agaricaceae was confirmed by molecular evidence(VELLINGA 2003).
Chamaemyces Battarra ex Earle in Bulletin of the New YorkBotanical Garden 5: 446. 1909.
Chamaemyces was originally distinguished from Lepiota be-cause of the non-dextrinoid spores. It differs from all othermembers of tribus Lepioteae in a number of characters: theconfluent stipe and pileus context, presence of pleurocystidia,the (ixo)hymenidermal pileus covering, and the presence ofclusters of caulocystidia, which are different from the pileuscovering. The development is monovelangiocarpic and sti-pitocarpic, whereas all other Agaricaceae are bivelangiocarpicand not stipitocarpic.
Coprinus (Pers.: Fr.) Gray, Natural Arrangement of BritishPlants 1: 632. 1821.
It has been shown repeatedly that Coprinus is a polyphyleticgroup (see REDHEAD et al. 2001 for an overview), and that thetype of the genus, C. comatus, together with a few other spe-cies, belongs to the Agaricaceae, close to Lepiota. Morpho-logically this genus (in its restricted sense) is quite differentfrom Lepiota, because of the structure of the lamellae with thepavement cells between the basidia, the heteromorphic basi-dia themselves, the autolytic lamellae, and the black sporeswith a germ pore. Currently it is debated whether the genusCoprinus should not be typified by C. atramentarius, in whichcase the genus name Annularius would be used for the pre-sent group (JØRGENSEN et al. 2001).

308 Phylogeny of Lepiota
© DGfM 2003
Montagnea Fr., Flora Scanica: 339. 1836.
Montagnea is a sister taxon of Coprinus (in the restricted sen-se), and harbours coprinoid taxa with a secotioid habit.
Three tropical genera are not included in the present study:Smithiomyces Singer (see VELLINGA (1999) for a descriptionof S. mexicanus (Murrill) Singer, the type species); JanauariaSinger from the Amazon region (SINGER 1986), and Hiatu-lopsis Singer & Grinling, described from tropical Africa (SIN-GER & GRINLING 1967).
Morobia Horak is regarded by some authors as a synonymof Lepiota (e.g. SINGER 1986), but the adnate (not free) lamel-lae, the structure of the pileipellis and the presence of rhizo-morphs, connecting the basidiocarps (HORAK 1979), suggesta closer relationship with Marasmius and allies.
Material and methods
In total 81 species and varieties of the ingroup, two outgroupspecies not belonging to the family Agaricaceae, and four spe-cies from other clades within the family Agaricaceae havebeen sampled. Most collections originated from Europe andNorth America; Asia, tropical regions and the Southern Hemi-sphere are strongly under-represented.
An overview of all taxa studied is given in Tab. 1, whichalso shows to which infrageneric taxon the Lepiota specieswere assigned. GenBank accession numbers are given thereas well.
Molecular analysis
DNA was extracted from fresh and herbarium material; themethods described by GARDES & BRUNS (1993 and 1996)were either followed completely, or instead of the alcoholprecipitation the GeneClean kit with Glassmilk® (Bio101Systems, Carlsbad, California, U.S.A.) was used. The inter-nal transcribed spacer (ITS), and part of the Large SubUnit(LSU) of the nuclear ribosomal repeat were amplified by thepolymerase chain reaction (PCR) with the fungal specific pri-mers ITS1F and ITS4 (GARDES & BRUNS 1993) for the ITS-region, and primers LR0R, LR3R, LR7 and LR16 for LSU(see Large subunit RNA (25-28S) primer sequences at http://www.biology.duke.edu/fungi/mycolab/primers.htm). Sequenc-ing of both strands was performed with an ABI model 377 se-quencer or an ABI model 3100 automated sequencer (AppliedBiosystems, Foster City, California, U.S.A.) using a ThermoSequenase™ Dye terminator Cycle Sequencing Pre-Mix Kit(Amersham Pharmacia Biotech, Piscataway, New Jersey,U.S.A.) or a BigDye™ Terminator Cycle Sequencing ReadyReaction Kit (Applied Biosystems); the primers ITS1 or ITS5,ITS2, ITS3, and ITS4 were used for the ITS-region, and pri-mers LR0R, LR3R, LR7, and LR16 for the LSU-region. Theraw data were processed with the use of DNA SequencingAnalysis v.2.1.2 and Sequence Navigator v.1.0.1 (also of Ap-plied Biosystems).
Alignment
The ITS sequences were aligned with the program PartialOrder Alignment (POA) (LEE, GRASSOW & SHARLOW 2002)with minimal visual alignment. CLUSTAL X 1.81 (THOMPSON
et al. 1997), with default settings, needed such heavy visualalignment that objectivity of the data could not be guaranteed.The LSU sequences were visually aligned only. The alignmentshave been deposited in TreeBase under number SN1461.
Phylogenetic analysis
Maximum parsimony analyses were performed using PAUP*version 4.0b10 (SWOFFORD 2002). One hundred heuristicsearches were conducted with random sequence addition andtree bisection-reconnection (TBR) branch-swapping algo-rithms, collapsing zero-length branches and saving all minimallength trees (MulTrees), with MaxTrees set at 5000. Limacel-la glioderma was chosen as outgroup. Non-informative cha-racters and a part of c. 150 base pairs (the number depends onthe actual alignment) of the ITS1-region that was not unambi-guously alignable, were excluded from all the analyses. Gapswere treated as missing data. To measure relative support forthe resulting clades, 3000 bootstrap replications (FELSENSTEIN
1985) were performed using the “fast” stepwise addition mode;groups with a support of ≥ 50% were retained.
To test alternative phylogenetic relationships, the Kishi-no-Hasegawa maximum likelihood ratio test (KISHINO & HA-SEGAWA 1989) with default parameters as implemented inPAUP* was performed. Several hypotheses, mainly based onthe morphologically based existing classifications, were for-mulated and tested.
Results
Alignment
The total length of the ITS fragment studied (including the 5.8Sgene) ranged from 610 base pairs in L. clypeolaria to 711 inL. luteophylla.
The ITS data set comprises 81 taxa and 87 collections, andis made up of 855 characters in total, of which 316 are parsi-mony informative. The aligned combined ITS-LSU data set,comprising 42 taxa, consists of a total of 1776 characters, ofwhich 415 are parsimony informative. The LSU data setconsists of 967 characters, of which 154 are parsimony in-formative. Part of the ITS1 segment was excluded from allanalyses, as it was not unambiguously alignable.
Phylogenetic analyses
Three distinct clades were recovered from all data sets, Clade1, comprising sect. Lepiota and sect. Ovisporae pro parte; Cla-des 2 and 3 (sometimes combined, sometimes separate), com-prising the species with trichodermal and hymenidermal pileuscoverings, and ellipsoid to spurred spores (sect. Ovisporae proparte, sect. Stenosporae, and sect. Lilaceae) (Figs. 1-4).

Fig. 1. One of 448 most parsimonious trees, based on all 314 parsimony informative characters of the ITS data set. Threemorphological characters are indicated for each species; sp sh indicates the spore shape, and p cov stands for pileus covering.L = 2768, CI = 0.2337, RC = 0.1174
© DGfM 2003
Mycological Progress 2(4) / 2003 309
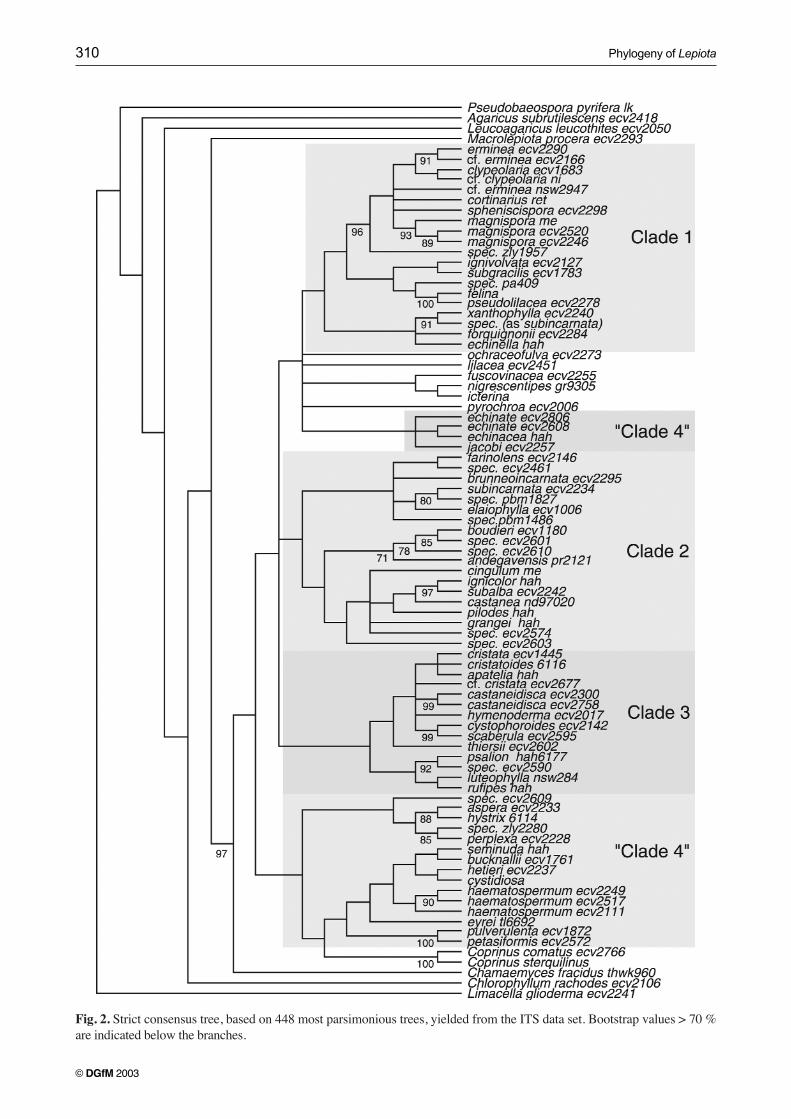
Fig. 2. Strict consensus tree, based on 448 most parsimonious trees, yielded from the ITS data set. Bootstrap values > 70 %are indicated below the branches.
© DGfM 2003
310 Phylogeny of Lepiota

Fig. 3. One of seven most parsimonious trees, with best –lnL, based on ITS and LSU data together. Bootstrap values ≥ 50 %are indicated below the branches. 414 Characters are parsimony informative. L = 1987, CI = 0.3531, RC = 0.1698
© DGfM 2003
Mycological Progress 2(4) / 2003 311

Fig. 4. One of 290 most parsimonious trees, based on the LSU data set. Bootstrap values > 60 % are indicated below the bran-ches. 154 Characters are parsimony informative. L = 490, CI = 0.4429, RC = 0.2410
© DGfM 2003
312 Phylogeny of Lepiota

Mycological Progress 2(4) / 2003 313
© DGfM 2003
Clade 4, the combination of Cystolepiota, Melanophyllum,and sect. Echinatae, can be recovered from a CLUSTAL Xaligned ITS data set (after visual alignment; data not shown),but it is paraphyletic in the phylogeny based on the POA-aligned ITS sequences.
Several taxa do not fit into one of those four clades, andcluster together into morphologically diverse groups.
Only Clade 1, or even only part of Clade 1 is bootstrapsupported, though the whole clade is present in all consen-sus trees. The other major clades are not bootstrap supported,but again, the clades are all recovered in the consensus trees.
Fourteen different hypotheses, mostly concerning the va-lidity of the sections, were thoroughly tested for the ITS dataset. Inclusion of both Chamaemyces fracidus and Coprinuscomatus (and C. sterquilinus) cannot be rejected (Tab. 2) (thecombined ITS-LSU data set gives the same results; Tab. 3);but Coprinus within Lepiota is very unlikely (in both analyses)(Tab. 2 & 3 – note that Lepiota has been taken in a restrictedsense in the analyses for Tab. 2, and in a wide sense in the ana-lyses for Tab. 3). The hypothesis that Chamaemyces fracidusbelongs to Lepiota (in restricted or in a wide sense) is not un-ambiguously supported, as the two different data sets give dif-ferent outcomes (Tab. 2 & 3).
The ITS data set does not support sect. Ovisporae, whosemembers are divided between Clade 1 and Clade 2 in the phy-logenies. Monophyly of sect. Lilaceae, of sect. Echinatae, andof Cystolepiota, Pulverolepiota, Melanophyllum and sect.Echinatae (Clade 4) together cannot be rejected (Tab. 2). Butmonophyly of Lepiota including Cystolepiota, Pulverolepiota,Melanophyllum, and sect. Echinatae, nor Lepiota excludingthose taxa can be accepted; in all those cases Coprinus coma-tus and allies, and Chamaemyces fracidus were not includedin Lepiota.
Discussion
Overview
In the analysis of the entire family, the tribus Lepioteae withoutChamaemyces, but including Coprinus comatus and allies,was shown to be monophyletic; Melanophyllum was shownto belong to this very clade (VELLINGA 2003).
These same results emerge from the present analyses of amuch more restricted data set. The resulting clade again alsocontains Coprinus comatus and C. sterquilinus.
Chamaemyces
The position of Chamaemyces fracidus is still not unambi-guous. In the analyses of the ITS, LSU or combined ITS-LSUdata sets of the family Agaricaceae as a whole (VELLINGA
2003), Chamaemyces fracidus is always basal to the family,and inclusion in Lepiota has to be rejected. In the present ana-lyses, based on a smaller sample from fewer lineages, its place-ment varies with the data; according to the ITS data a placewithin Lepiota is acceptable (Fig. 1; Tab. 2), but it is excluded
based on the results of the Kishino-Hasegawa test on the com-bined data set, where Lepiota is taken in a wider sense (includ-ing Cystolepiota, sect. Echinatae, Melanophyllum and Pul-verolepiota) (Table 3).
The morphology of Chamaemyces fracidus, and especial-ly its development (see under ‘Developmental characters’), isquite different from Lepiota itself, and a position at the baseof the family seems much more plausible than one withinLepiota.
Coprinus comatus
Despite ending up within the Lepiota clade, the position ofCoprinus comatus and C. sterquilinus is ambiguous. Inclu-sion of Coprinus comatus and C. sterquilinus within Lepiotais not supported by the ITS data, nor by the ITS-LSU data;though a position within Lepiota for this group and for Cha-maemyces cannot be rejected (Tab. 3). However, MONCALVO
et al. (2002) showed, based on LSU analyses, that these taxatogether with three Montagnea species form a distinct clade,separate from Lepiota and the other monophyletic groups.These authors examined the same seven Lepiota and Cysto-lepiota taxa as JOHNSON & VILGALYS (1998), plus two Melano-phyllum sequences.
Morphologically, the coprinoid taxa are totally differentfrom all taxa in the Lepiota clade, and a separate position,which cannot be ruled out by our analyses, does seem an ap-propriate solution.
Lineages
Contrary to expectations, essentially none of the morpholo-gically recognized genera, subgenera and sections form at firstglance monophyletic groups; Pulverolepiota and Melano-phyllum form the exception. Though when tested with the Kis-hino-Hasegawa test, monophyly of sections Lilaceae andEchinatae cannot be rejected (ITS data; Tab. 2).
Based on morphology and outcome of the present analysesthe following four clades are accepted, and briefly describedand discussed.
Clade 1
Clade 1 is characterized by trichodermal pileus covering madeup of long and short, cylindrical to narrowly clavate, elements.The spore shape varies: fusiform (e.g. in L. erminea and L.clypeolaria), penguin-shaped (in L. spheniscispora, L. magni-spora, and L. cortinarius), and broadly fusiform to ellipsoid(e.g. in L. subgracilis, L. xanthophylla). Within the Lepioteae,Clade 1 always gets relatively high bootstrap support. Thisclade harbours all species from sect. Lepiota and a part of sect.Ovisporae.
There has been disagreement about the position of L. cor-tinarius; BON (1981; 1993) placed the species in sect. Steno-sporae, but VELLINGA (1992) placed it in sect. Lepiota closeto L. ventriosospora. Both authors used spore shape as theoverruling character for their decision. A position in sect.

314 Phylogeny of Lepiota
Lepiota is supported by the present, molecular data, and bythe structure of the pileus covering.
The collection from which the GenBank sequence of L.subincarnata has been derived must have been misidentified,as the present data do not place it with our collections of thisspecies, characterized by the lack of short elements in the pi-leus covering.
Several collections, morphologically identified as L. cly-peolaria, turned out to be polymorphic with regard to theirITS sequences; these collections were not included in thepresent data set.
Clade 2
This clade harbours species with similarly varied spore shape,and with two types of pileus covering, either a trichoderm oflong elements only or a loosely arranged cutis with articulate,clamped hyphae. Species with ellipsoid and with spurred spo-res both occur in this clade, representing sect. Stenosporae (partof genus Lepiotula) and sect. Ovisporae subsect. Helveolinae.Three subclades can be recognized: 2a with ellipsoid spores anda trichoderm, 2c with spurred spores and a trichoderm, and 2bwith spurred spores and a cutis-like pileus covering.
The most infamous amanitin containing species, L. sub-incarnata (and its synonym L. josserandii), L. elaiophylla, andL. brunneoincarnata constitute Clade 2a.
Species limitations in the complex of Lepiota castaneahave been examined by MIGLIOZZI & ZECCHIN (1997), whorecognized three species, mainly on spore size. However, VEL-LINGA (2001a) found the boundaries between their taxa werenot clear-cut and still considered L. castanea a single very va-riable species. Two samples of this group were sequenced,and it appears that indeed, a small-spored taxon, tentativelycalled L. ignicolor, with relatively short elements in the pileuscovering can be separated from a long-spored taxon, viz. L.castanea, with long elements. Lepiota ignicolor is a sister ta-xon of L. subalba, with equally short pileus covering elements.A thorough examination of this group is needed, using a widesample from all over the Northern Hemisphere; the morpho-logical variability and the molecular data deserve equal at-tention. The morphologically relevant characters are generalbasidiocarp shape and colour, spore size, length and numberof septa of the pileus covering elements and the position ofthese on the pileus.
Clade 3
Clade 3 is characterized by a hymenidermal pileus covering,whereas the spore shape is variable, and so are the chemicalreactions and the number of nuclei per spore (see VELLINGA
& HUIJSER (1999) for an extensive morphological overviewof the relevant species). Hymenidermal species assigned toLepiotula, and to sect. Lilaceae belong here. Three species, L.pyrochroa, L. lilacea and L. ochraceofulva, do not fall intothis clade on molecular grounds, though they fit morphologi-cally; their inclusion cannot be rejected (Tab. 2). Surprisingly,L. rufipes and L. pyrochroa are not sister taxa, as suggested
by VELLINGA & HUIJSER (1999), nor does L. rufipes stand outfrom the other taxa in this clade, despite its distinctive mor-phology. Spores of several species in this clade have meta-chromatic, non-dextrinoid walls. Pink colouration of the sporewall in Cresyl Blue, a metachromatic reaction, is characteristicfor, but not restricted to, the genera Leucoagaricus, Leucoco-prinus, Macrolepiota and Chlorophyllum. In these genera, thespore wall is also dextrinoid.
Lepiota scaberula, L. cystophoroides, and L. luteophyllado not have a simple hymenidermal pileus covering, but onegiving rise to loose cells. On account of this architecture of thepileus covering, L. luteophylla was accommodated in Cysto-lepiota (KNUDSEN 1978). That position does not seem accept-able, in light of the present data. As in many other species ofthis clade, the spores are not dextrinoid.
Chamaemyces fracidus possesses the characteristic hy-meniform pileus covering of Clade 3, and is accommodatedthere in trees based on the combined data set. However, thedevelopment of the basidiocarp in Ch. fracidus is quite dif-ferent from that in L. cristata (REIJNDERS 1975; ATKINSON
1916 for L. cristata).
Clade 4
The fourth clade is not monophyletic in phylogenies based onthe ITS-data, though its monophyly cannot be rejected (Tab. 2).This group is made up of species assigned to sect. Echinatae,and the genera Cystolepiota, Melanophyllum and Pulvero-lepiota. In the ITS-phylogenies, Coprinus comatus and C.sterquilinus also belong to this clade. The position of thesetwo taxa is far from resolved, as ITS and LSU data give con-flicting signals. Section Echinatae is split into two subclades,a ‘large-spored’ group (L. aspera, L. hystrix, L. perplexa), anda ‘small-spored’ group (L. jacobi, L. echinacea and some un-described species from California). Amazingly, in this casespore shape and size do seem to distinguish the two subclades.Variation in ITS among the species in this clade as a whole isquite substantial.
Additional visual alignment of the POA-aligned ITS dataset renders clade 4 paraphyletic, and split up into small sub-clades (results not shown).
It is not difficult to distinguish Cystolepiota species fromspecies in Lepiota sect. Echinatae; spore characters and pileuscovering are decisive. However, misidentifications have beenmade, e.g. ENDERLE & KRIEGLSTEINER (1989) misapplied thename L. pseudoasperula Knudsen to C. moelleri Knudsen.
A close relationship of Melanophyllum and Cystolepiotais not surprising, as both taxa are morphologically and deve-lopmentally very similar, they differ only in colouration andornamentation of the spores.
It is also clear that Melanophyllum haematospermum ismore likely to be a species complex, than one phylogeneticspecies. The three collections sampled here differ in size ofthe basidiocarps, from small to medium-sized, with short orlong stipe. World-wide sampling and morphological andmolecular comparisons of the collections are necessary.
© DGfM 2003

Mycological Progress 2(4) / 2003 315
Taxa outside the four main clades
The position of some species which are morphologically dis-tinct from all the others is just as problematic on moleculargrounds. For example, L. fuscovinacea and L. nigrescentipesare not closely related to any other species in the present dataset. Lepiota nigrescentipes is closely related to L. parvannu-lata (Lasch: Fr.) Gillet (the two may be synonymous), and ischaracterized by small, non-dextrinoid, uni-nucleate spores,absence of cheilocystidia, and a cutis-like pileus covering, withclamp-connections. Lepiota fuscovinacea differs from all otherexamined species in the absence of clamp-connections, theuni-nucleate spores, and the cutis-like copious pileus and stipecovering.
The position of L. lilacea, L. ochraceofulva, and C. icte-rina in the cladograms based on ITS data (e.g. Fig. 1) is hardto explain. The first two species have a hymenidermal pileuscovering, and the latter is a unique Cystolepiota species be-cause of its yellow tinges and the dextrinoid spores.
Character combinations
Clades 1, 2 and 3 each harbour species with different sporeshapes, an indication that features such as spurred spores haveevolved independently several times. Spore shape in Clade 4also varies, though less dramatically. Since spore shape failsto characterize the clades, is there any other morphologicalcharacter that does? An answer may be found in the structureof the pileus covering, and perhaps in the developmental stagesof the basidiocarps that give rise to it.
The pileus covering of the species in Clade 1 is a tricho-derm, made up of both long cylindrical elements and short,clavate to cylindrical elements. Clade 2 is characterized byhaving a trichoderm or a cutis-like pileus covering, but bothtypes of covering are made up of long cylindrical elementsonly. Taxa in Clade 3 have a hymenidermal pileus covering.The elements of the pileus covering in Clade 4 are globoseto inflated, either loosely arranged or in agglutinated chains.
Despite the variation in spore shape within each clade,some character combinations have not evolved; for example,there are no taxa with a hymenidermal pileus covering andfusiform uni-nucleate spores.
Developmental characters
A key character in the Agaricaceae is the presence of protec-tive layers (veils or vela); their structure and developmentalpathways are both important. Ontogenesis has been morpho-logically studied in several species: L. clypeolaria (ATKINSON
1914), L. cristata (ATKINSON 1916), L. aspera (as L. acute-squamosa; GREIS 1937), L. castanea (as L. ignicolor; HUGUE-NEY 1966), C. seminuda (ATKINSON 1916), C. hetieri and Me-lanophyllum haematospermum (REIJNDERS 1963). REIJNDERS
(1975) treated the family as a whole. CLÉMENÇON (1997) in-troduced a new terminology for the carpogenesis, and re-interpreted some of the existing data. The development of L.clypeolaria differs substantially from that of L. castanea(CLÉMENÇON 1997); in L. castanea a separate hyphal cutis-
like layer covers the whole pileus, and the underlying tricho-derm is interpreted as the pileipellis by CLÉMENÇON (1997).This outer layer is absent in L. clypeolaria, and the trichodermis interpreted as of velar origin. This may supply a clue to thedifferences between Clades 1 and 2, though without furtherwork we should not assume that the development of all thespecies in those clades is the same as for the two representa-tives studied. Lepiota cristata deviates in the absence of velarmaterial on the stipe of the mature basidiocarps, though inother members of Clade 3, e.g. L. hymenoderma, there areremnants of the universal veil present at the stipe base. It alsoseems that C. seminuda follows a different developmentalpathway than the other species, so it is surprising that it is soclose to L. aspera in the molecular analyses, which place bothtaxa in Clade 4. The absence of the ‘metablematogenic’ layerin the primordial stages of C. seminuda, C. hetieri, and Mela-nophyllum haematospermum is striking, as this layer is veryprominent in the early stages of the development of L. aspera(compare GREIS (1937) and ATKINSON (1916)). Unfortunatelynothing is known about the genetic basis of these different de-velopmental pathways.
Chamaemyces fracidus differs considerably in developmentfrom Lepiota (REIJNDERS 1975). It is monovelangiocarpic,whereas all other taxa are bivelangiocarpic, and furthermore,the stipe is the first to develop (stipitocarpy); Lepiota speciesare pileostipitocarpic.
Character evolution
As the deeper branches are poorly resolved, and the variousanalyses yield slightly different results, it is not easy to sayanything with certainty about the characters of the ancestralLepiota. BON (1993), speculating on the evolutionary lineswithin the group, considered Pseudobaeospora ancestral, andregarded L. parvannulata as the most primitive species. He en-visaged lineages originating with L. parvannulata and leadingto Cystolepiota; to sect. Lepiota, and, via sect. Ovisporae, toEchinoderma, while sect. Stenosporae was a side branch.Lepiota parvannulata was not included in our sample but L.nigrescentipes, which is morphologically very close to it, doesnot seem to be a good candidate for the proto-Lepiota. Otheraspects of BON’s ‘phylogeny’ are contradicted by the presentanalyses, e.g. the close relationship of Cystolepiota and Echi-noderma. But it is premature to speculate further. Spurred and‘penguin-shaped’ spores do seem to be a derived character,having originated several times in different lineages.
Vicariance and speciation
An intriguing aspect of the present study is that it draws at-tention to several pairs of sister taxa with the same disjunctdistribution, with one species in Europe and the other in wes-tern North America. Examples are L. cystophoroides pairedwith L. scaberula, and C. pulverulenta with L. petasiformis.This phenomenon also occurs in other members of the family.It might indicate that speciation took place after the two areasbecame isolated from each other. Unfortunately, for many of
© DGfM 2003

316 Phylogeny of Lepiota
these taxa data on their occurrence in eastern North Americaand in Asia are not available. Data on sexual compatibility ofthese taxa are also needed to be more explicit on the mecha-nisms of speciation.
Secotioid forms
Recently a secotioid species was described from Sardinia, viz.Notholepiota sardoa Padovan & Contu, which resemblesthe grassland species L.oreadiformis Velen. and L. erminea(PADOVAN & CONTU 2001). Judging from the description byPADOVAN & CONTU (2001) this species belongs to LepiotaClade 1. Notholepiota areolata (Stevenson) Horak from NewZealand, which was suggested to be closely related to Lepiotaon account of the pale fusiform spores and the dextrinoid re-action of the spore wall (HORAK 1971), has turned out to beclosely related to Boletus (VELLINGA 2003). Other secotioidforms within this group of the family Agaricaceae are Mon-tagnea species in the Coprinus clade. The phenomenon of ad-apting to dry conditions by becoming secotioid has evolvedseveral times within the genus Agaricus; examples are Gyro-phragmium dunalii (Fr.) Zeller, Longula texensis (Berk. &M.A. Curtis) Zeller, and Agaricus inapertus Vellinga (syn.Endoptychum depressum Singer & A.H. Sm.).
In conclusion
It is premature to base a new classification on the phyloge-netic hypotheses presented here; bootstrap supports for theclades are low, the different data sets do not always presentthe same topologies, and the deeper branching is unresolved.However, morphological characters strongly support the cla-des, and the main clades are always present in the consensustrees. In other basidiomycete genera ITS sequences have suc-cessfully been used to recognize infrageneric taxa, e.g. in Rhi-zopogon (GRUBISHA et al. 2002).
Chamaemyces fracidus should be excluded from Lepio-ta; a place at the base of the family Agaricaceae seems ap-propriate on account of both morphological and molecularcharacters. Inclusion of Coprinus comatus and allies withinLepiota does not seem acceptable either.
Results indicate that sections Lepiota and section Steno-sporae have to include species with different spore shapes thanaccepted hitherto, and a re-evaluation of section Ovisporae isneeded. Also the recognition of separate genera for Cysto-lepiota, sect. Echinatae, Melanophyllum and Pulverolepiotahas to be reconsidered. One genus for this group seems a goodalternative as already advocated by KNUDSEN (1978).
The structure of the pileus covering, and the developmentof the basidiocarps are key characters in the evolution andclassification of this group.
The present study suggests several areas where further re-search may elucidate the phylogeny of Lepiota and its allies:i. molecular research focusing on other genes, preferablymarkers which give a good phylogenetic signal above specieslevel to elucidate the basal branching; ii. molecular and mor-
phological research on tropical taxa, and on taxa with uniquecharacter combinations, such as the neotropical species L.rubella Bres. with a cutis-like pileus covering of thick-walledelements and small ellipsoid spores, and the neotropical genusSmithiomyces, with its unique structure of the pileus covering,a genus which might be closely related to Lepiota; iii. deve-lopmental studies, especially focusing on the ontogeny of thebasidiocarps and the origin and formation of the velar layers,but also paying attention to the developmental differences be-tween agaricoid and secotioid basidiocarps.
But the foremost requirement is for descriptive taxonomicand morphological work in all areas of the world. Even in Eu-rope, where the situation is best, new Lepiota species are de-scribed almost every year. Some recent examples: FILLION
(1997) described L. glareophila from the French Alps, a spe-cies which probably belongs to Clade 1 in the present presen-tation, and VELLINGA & HUIJSER (1999) published two newspecies in sect. Lilaceae (Clade 3). In North America, ourknowledge of the Lepiota flora is still very limited, and manyundescribed species occur.
Acknowledgments
The basis for this analysis was formed by collections made bymany people. Curators of the herbaria at C, L, and MICH, andthe following individuals are acknowledged for their contri-butions: Henk A. Huijser, Rob Chrispijn, Manfred Enderle,Thomas Læssøe, Pierre Roux, Nico J. Dam, Fred Stevens,Nancy Ironside, Rod E. Tulloss, Zhu-Liang Yang, P. Bran-don Matheny, and Lothar J. Krieglsteiner. The MSA awardedthe A.H. and H.V. Smith award to visit the herbarium in AnnArbor, Michigan in 2000. The Rijksherbariumfonds Dr. E.Kits van Waveren provided funding for laboratory work in1999. The Bruns Lab (University of California at Berkeley,U.S.A.) supported laboratory work during the years 2000-2002. Comments on the manuscript by John Lennie, PieterBaas, Machiel E. Noordeloos, and Thomas W. Kuyper aregreatly appreciated.
References
ARORA D (1986) Mushrooms Demystified. A Comprehensive Guide tothe Fleshy Fungi, 2nd ed. Ten Speed Press, Berkeley.
ATKINSON GF (1914) The development of Lepiota clypeolaria. – An-nales Mycologici 12: 346-356, pl. 13-16.
ATKINSON GF (1916) The development of Lepiota cristata and L. se-minuda. – Memoirs New York Botanical Garden 6: 209-228.
BON M (1981) Clé monographique des 'Lépiotes' d'Europe. – Docu-ments Mycologiques 11(43): 1-77.
BON M (1993) Flore mycologique d'Europe 3. Les Lépiotes. DocumentsMycologiques. – Mémoire hors série 3: 1-153. Lepiotaceae Roze.
CANDUSSO M, LANZONI G (1990) Lepiota s.l. Fungi europaei 4. Gio-vanna Biella, Saronno.
CLÉMENÇON H (1997) Anatomie der Hymenomyceten. Flück-Wirth,Teufen AG.
© DGfM 2003

Mycological Progress 2(4) / 2003 317
ENDERLE M, KRIEGLSTEINER GJ (1989) Die Gattung Lepiota (Pers.) S.F.Gray emend. Pat. in der Bundesrepublik Deutschland (Mitteleu-ropa). – Zeitschrift für Mykologie 55: 43-104.
FELSENSTEIN J (1985) Confidence limits on phylogenies: an approachusing the bootstrap. – Evolution 39: 783-791.
FILLION R (1997) Lepiota glareophila espèce nouvelle. – Bulletin tri-mestriel de la Fédération Mycologique Dauphiné-Savoie 144:7-11.
FRIES EM (1821) Systema Mycologicum. 1. Gryphiswaldiae. “Sumti-bus Ernesti Mayritii”.
GARDES M, BRUNS TD (1993) ITS primers with enhanced specificityfor Basidiomycetes. Application to the identification of mycor-rhizae and rusts. – Molecular Ecology 2: 113-118.
GARDES M, BRUNS TD (1996) Community structure of ectomycorrhi-zal fungi in a Pinus muricata forest: above- and below-groundviews. – Canadian Journal of Botany 74: 1572-1583.
GREIS H (1937) Zur Entwicklungsgeschichte von Lepiota acutesqua-mosa Weinm. – Jahrbücher für Wissenschaftliche Botanik 84:449-480.
GRUBISHA LC, TRAPPE JM, MOLINA R, SPATAFORA JW (2002) Biologyof the ecto-mycorrhizal genus Rhizopogon. VI. Re-examinationof infrageneric relationships inferred from phylogenetic analy-ses of ITS sequences. – Mycologia 94: 607-619.
HARMAJA H (2002) Amylolepiota, Clavicybe and Cystodermella, newgenera of the Agaricales. – Karstenia 42: 39-48.
HORAK E (1971) Contributions to the knowledge of the Agaricales s.l.(Fungi) of New Zealand. – New Zealand Journal of Botany 9:463-493.
HORAK E (1979) Three new genera from Papua New Guinea. – Beiheftezur Sydowia 8: 202-208.
HUGUENEY R (1966) Sur la présence de deux voiles universels chezLepiota ignipes Locquin. – Comptes Rendus Hebdomadaires desSéances de l'Académie des Sciences, Paris, série D, 263: 491-493.
JOHNSON J (1999) Phylogenetic relationships within Lepiota sensu latobased on morphological and molecular data. – Mycologia 91:443-458.
JOHNSON J, VILGALYS R (1998) Phylogenetic systematics of Lepiotasensu lato based on nuclear large subunit rDNA evidence. – My-cologia 90: 971-979.
JØRGENSEN PM, RYMAN S, GAMS W, STALPERS JA (2001) (1486) Pro-posal to conserve the name Coprinus Pers. (Basidiomycota) witha conserved type. – Taxon 50: 909-910.
KERRIGAN RW (1986) The Agaricales of California. 6. Agaricaceae.Mad River Press, Eureka, CA.
KISHINO H, HASEGAWA M (1989) Evaluation of the maximum likelihoodestimate of the evolutionary tree topologies from DNA sequencedata and the branching order in Hominoidea. – Journal of Mole-cular Evolution 29: 170-179.
KNUDSEN H (1978) Notes on Cystolepiota Sing. and Lepiota S.F. Gray.– Botanisk Tidsskrift 73: 124-136.
KNUDSEN H (1980) A revision of Lepiota sect. Echinatae and Amy-loideae (Agaricaceae) in Europe. – Botanisk Tidsskrift 75: 121-155.
KRÜGER D, BINDER M, FISCHER M, KREISEL H (2001) The Lycoperda-les. A molecular approach to the systematics of some gasteroidmushrooms. – Mycologia 93: 947-957.
KÜHNER R (1936) Recherches sur le genre Lepiota. – Bulletin Trimest-riel de la Societé Mycologique de France 52: 177-238.
LEE C, GRASSO C, SHARLOW M (2002) Multiple sequence alignmentusing partial order graphs. – Bioinformatics 18: 452-464.
LINCOFF GH (1981) The Audubon Society Field Guide to North Ame-rican Mushrooms. Alfred A. Knopf, New York. 926 pp.
MIGLIOZZI V, ZECCHIN G (1997) Studio della sezione Stenosporae (Lan-ge) Kühner del genere Lepiota s.s. 2a parte. Lepiota ignicolor Bre-sadola, Lepiota castanea Quélet sensu stricto e Lepiota castaneaQuélet sensu auctores. – Micologia Italiana 26 (2): 11-22.
MILLER OK JR, VOLK TJ, BESSETTE AE (1996) A new genus, Leuco-pholiota, in the Tricholomataceae (Agaricales) to accommodatean unusual taxon. – Mycologia 88: 137-139.
MONCALVO J-M, VILGALYS R, REDHEAD SA, JOHNSON JE, JAMES TY,AIME MC, HOFSTETTER V, VERDUIN SJW, LARSSON E, BARONI
TJ, THORN RG, JACOBSSON S, CLÉMENÇON H, MILLER OK Jr(2002) One hundred and seventeen clades of Euagarics. – Mole-cular Phylogenetics and Evolution 23: 357-400.
PADOVAN F, CONTU M (2001) Notholepiota sardoa sp. nov., un nuovogasteromicete filogeneticamente correlato alle Lepiotaceae sco-perto in Sardegna. – Bollettino del Gruppo Micologico ‘G. Bre-sadola’, n.S., 44 (2): 31-36.
REDHEAD SA, VILGALYS R, MONCALVO J-M, JOHNSON J, HOPPLE JS Jr.(2001) Coprinus Pers. and the disposition of Coprinus speciessensu lato. – Taxon 50: 203-241.
REIJNDERS AFM (1963) Les Problèmes du Développement des Carpo-phores des Agaricales et de quelques Groupes Voisins. Uitge-verij Dr. W. Junk, Den Haag.
REIJNDERS AFM (1975) The development of three species of the Aga-ricaceae and the ontogenetic pattern of this family as a whole. –Persoonia 8: 307-319.
SINGER R (1986) The Agaricales in Modern Taxonomy, 4th edn. KoeltzScientific Books, Koenigstein.
SINGER R, GRINLING K (1967) Some Agaricales from the Congo. – Per-soonia 4: 355-377.
SWOFFORD DL (2002) PAUP*. Phylogenetic Analysis Using Parsimony(*and Other Methods). Version 4. Sinauer Associates, Sunder-land, Massachusetts.
THOMPSON JD, GIBSON TJ PLEWNIAK F, JEANMOUGIN F, HIGGINS DG(1997) The CLUSTAL_X windows interface: flexible strategies formultiple sequence alignment aided by quality analysis tools. –Acids Research 25: 4876-4882.
VELLINGA EC (1992) Notulae ad floram agaricina neerlandicam – XVIII.Some notes on Cystolepiota and Lepiota. – Persoonia 14: 407-415.
VELLINGA EC (1999) An American in a Belgian swimmingpool. – My-cologist 13: 50-53.
VELLINGA EC (2001a) Lepiota. In Noordeloos ME, Kuyper ThW, Vel-linga EC (eds). Flora Agaricina Neerlandica 5, pp 109-151. A.A.Balkema Publishers, Lisse, Abingdon, Exton (PA), Tokyo.
VELLINGA EC (2001b) Studies in Lepiota III. Some species from Cali-fornia, U.S.A. – Mycotaxon 80: 285-295.
VELLINGA EC (2001c) Studies in Lepiota IV. Lepiota cristata and L.castaneidisca. – Mycotaxon 80: 297-306.
VELLINGA EC (2003) Phylogeny and Taxonomy of Lepiotaceous Fungi.PhD thesis, Universiteit Leiden, the Netherlands.
VELLINGA EC, HUIJSER HA ('1998' 1999). Studies in Lepiota I. Specieswith a hymeniform pileus covering. – Belgian Journal of Botany131: 191-210.
VELLINGA EC, DE KOK RPJ, BRUNS, TD (2003). Phylogeny and taxo-nomy of Macrolepiota (Agaricaceae). – Mycologia 95: 442-456.
WASSER SP (2002) Biodiversity of Cyanoprocaryotes, Algae and Fungiof Israel. Family Agaricaceae (Fr.) Cohn (Basidiomycetes) of Is-rael mycobiota. I. Tribe Agariceae Pat. A.R.A. Gantner Verlag,Ruggell.
Accepted: 22.8.2003
© DGfM 2003

Species Classification Collection Particulars ITS LSUaccording to GenBank GenBankBON (1993) accession # accession #
Agaricus subrutilescens (Kauffman) n.a. U.S.A., California, Sonoma Co., Salt Point State AF482832Hotson & Stuntz Park, 15.12.1999, E.C. Vellinga 2418 (UC)
Chamaemyces fracidus (Fr.) Donk Chamaemyces Belgium, prov. Luxembourg, Ave et Auffe, Le AY176343 AY176344Roptai, 10.1977, Th.W. Kuyper 960 (L)
Chlorophyllum rachodes (Vittad.) Macrolepiota sect. The Netherlands, prov. Zuid-Holland, Leiden, AF482849Vellinga Laevistipedes 24.9.1997, E.C. Vellinga 2106 (L)
Chlorophyllum rachodes Macrolepiota sect. GenBank U85277Laevistipedes
Coprinus comatus (O.F. Müll.: Fr.) n.a. U.S.A., California, Alameda Co., Berkeley, AY176346Pers. campus University of California at Berkeley, near
Warren Hall, 3.12.2001, E.C. Vellinga 2766 (UC)Coprinus comatus n.a. GenBank AF041529
Coprinus sterquilinus (Fr.: Fr.) Fr. n.a. GenBank AF345821 AF041530
Cystolepiota bucknallii (Berk. & Br.) Cystolepiota sect. The Netherlands, prov. Limburg, Bemelen, AY176458Clémençon & Singer Pseudoamyloideae Bemelerberg, 9.10.1991, E.C. Vellinga 1761 (L)
Cystolepiota cystidiosa (A.H. Sm.) Bon sect. Cystolepiota GenBank U85333 U85298
Cystolepiota hetieri (Boud.) Singer Cystolepiota sect. The Netherlands, prov. Limburg, Elsloo- Geulle, AY176459Cystolepiota Bunderbos, 19.9.1998, E.C. Vellinga 2237 (L)
Cystolepiota icterina Knudsen Cystolepiota sect. Denmark, Bornholm, Kobbeådalen, 9.9.1992, AY176460Pseudoamyloideae R. Ejrnæs RE090992-1 (C)
Cystolepiota pulverulenta (Huijsman) Pulverolepiota Great Britain, Surrey, Richmond, Kew Botanical AF391036 AY176349Vellinga Garden, 8.9.1992, E.C. Vellinga 1872 (L)
Cystolepiota seminuda (Lasch) Bon Cystolepiota sect. The Netherlands, prov. Limburg, Savelsbos, AY176350 AY176351Cystolepiota 4.10.1989, H.A. Huijser s.n. (herb. Huijser)
Lepiota andegavensis Mornand Sect. Stenosporae France, Coudré, congrès de Thonon, 8.10.1994, AY176461P.D.H. Roux 2121 (herb. Roux)
Lepiota apatelia Vellinga & Huijser Sect. Lilaceae The Netherlands, prov. Limburg, Brunssum, mine AY176462Hendrik, 26.9.1990, H.A. Huijser s.n. (herb. Huijser)
Lepiota aspera (Pers.: Fr.) Quél. Echinoderma The Netherlands, prov. Limburg, Elsloo- Geulle, AY176354 Bunderbos, 19.9.1998, E.C. Vellinga 2233 (L)
Lepiota aspera Echinoderma GenBank U85293
Lepiota boudieri Rea Sect. Stenosporae The Netherlands, prov. Noord-Holland, Amsterdam- AF391025Buitenveldert, 17.9.1987, E.C. Vellinga 1180 (L)
Lepiota brunneoincarnata Chodat & Sect. Felinae The Netherlands, prov. Zeeland, Schouwen- AY176355C. Martin Duiveland, Verklikkerduinen, 7.10.1998, E.C.
Vellinga 2295 (L)Lepiota brunneoincarnata Sect. Felinae Germany, Baden-Württemberg, Ulm-Grimmelfingen, AF482896
'Wald "Hörnle" 29.8.1996, M. Enderle s.n. (L)
Lepiota castanea Quél. Sect. Stenosporae The Netherlands, prov. Gelderland, Nijmegen, AY176463Staddijk, 27.7.1997, N. Dam 97020 (herb. Dam)
Lepiota castaneidisca Murrill Sect. Cristatae U.S.A., California, San Mateo Co., San Mateo AF391057County Memorial Park, 4.12.1998, F. Stevens (coll. E.C. Vellinga 2300) (UC)
Lepiota castaneidisca Sect. Cristatae U.S.A., California, Marin Co., Samuel P. Taylor AY176464State Park, 28.11.2001, E.C. Vellinga 2758 (UC)
Lepiota castaneidisca Sect. Cristatae U.S.A., California, San Mateo Co., Skyline Blv, AY17635613.12.1999, E.C. Vellinga 2411 (UC)
Lepiota cf. clypeolaria (Bull.: Fr.) Sect. Lepiota Canada, Ontario, Manitoulin Island, Co. Mani- AY176465Kummer toulin, Robinson, 29.9.2001, N. Ironside s.n. (UC)
Lepiota cf. cristata (Bolt.: Fr.) Sect. Cristatae U.S.A., California, Mendocino Co., Navarro River AY176466 Kummer Redwood State Park, 18.11.2001, E.C. Vellinga
2677 (UC)
Lepiota cf. erminea (Fr.: Fr.) Kummer Sect. Lepiota France, dépt. l'Hérault, Bédarieux, Oct. 1997, AY176467E.C. Vellinga 2166 (L)
Lepiota cf. erminea Sect. Lepiota U.S.A., Idaho, Owyhee Co. Cow Creek, 12.6.1972, AY176357 AY176358N.S. Weber 2947 (MICH)
Tab. 1. Overview of species and collections and the GenBank accession numbers for the ITS and LSU sequences
© DGfM 2003
318 Phylogeny of Lepiota

Species Classification Collection Particulars ITS LSUaccording to GenBank GenBankBON (1993) accession # accession #
Lepiota cingulum Kelderman Sect. Stenosporae Germany, Bavaria, 20.9.1995, M. Enderle s.n. (L) AY176359 AY176360
Lepiota clypeolaria (Bull.: Fr.) Kummer Sect. Lepiota Germany, Rheinland-Pfalz, Eifel, Gerolstein, AY176361Berlinger Bach, 17.9.1990, E.C. Vellinga 1683 (L)
Lepiota clypeolaria Sect. Lepiota Belgium, prov. Luxembourg, Resteigne, Bois de AY176362Resteigne, 4.10.1998, H.A. Huijser s.n. (herb. Huijser)
Lepiota cortinarius J.E. Lange Sect. Stenosporae U.S.A., Connecticut, Middlesex Co., Meshomasic AY176468State Forest, 25.9.1999, R.E. Tulloss s.n. (UC)
Lepiota cristata (Bolt.: Fr.) Kummer Sect. Cristatae Luxembourg, Hollenfels, 28.9.1988, E.C. Vellinga AF3910271445 (L)
Lepiota cristata Sect. Cristatae GenBank U85292
Lepiota cristatoides Einhellinger Sect. Integrellae The Netherlands, prov. Limburg, Cadier en Keer, AY176363 AY176364Riesenberg, 5.9.1996, H.A. Huijser s.n. (herb. Huijser)
Lepiota cystophoroides Joss. & Sect. Lilaceae France, dépt l'Hérault, Cêdraie d'Escandorgue, AF391031 AY176365Riousset 20.10.1997, E.C. Vellinga 2142 (L)
Lepiota echinacea J.E. Lange Echinoderma The Netherlands, prov. Noord-Brabant, Best, Steen- AY176469fabriek, 21.9.1998, H.A. Huijser s.n. (herb. Huijser)
Lepiota echinella Quél. & Bernard Sect. Felinae Belgium, prov. Luxembourg, Bois de Resteigne, AY176366 AY1763674.10.1998, H.A. Huijser s.n. (herb. Huijser)
Lepiota elaiophylla Vellinga & Huijser Sect. Helveolae The Netherlands, prov. Noord-Holland, Uithoorn, AF391024de Kwakel, in greenhouse, 15.9.1986, E.J.M. Uljé (coll. E.C. Vellinga 1006) (L)
Lepiota erminea (Fr.: Fr.) Kummer Sect. Lepiota The Netherlands, prov. Zeeland, Schouwen- AY176470Duiveland, Meeuwenduinen, 6.11.1998, E.C. Vellinga 2290 (L)
Lepiota farinolens Bon & G. Riousset Sect. Felinae France, dépt. L’Hérault, Graissesac, 21.10.1997, AY176368 AY176369E.C. Vellinga 2146 (L)
Lepiota felina (Pers.) P. Karst. Sect. Felinae GenBank U85330 U85295
Lepiota forquignonii Quél. Sect. Felinae The Netherlands, prov. Limburg, Valkenburg, AY176370 AY176371Schaelsberg, 10.10.1998, E.C. Vellinga 2284 (L)
Lepiota fuscovinacea F.H. Møller & Sect. Fuscovinaceae The Netherlands, prov. Noord-Holland, Amsterdam, AY176372 AY176373J.E. Lange Amstelpark, 23.9.1998, E.C. Vellinga 2255 (L)
Lepiota grangei (Eyre) Kühner Sect. Stenosporae Belgium, prov. Luxembourg, Bois de Resteigne, AY1764714.10.1998, H.A. Huijser s.n. (herb. Huijser)
Lepiota hymenoderma D.A. Reid Sect. Lilaceae The Netherlands, prov. Limburg, Elsloo-Geulle, AF391083 AY176376Bunderbos, 7.9.1996, E.C. Vellinga 2017 (L)
Lepiota hystrix F.H. Møller & Echinoderma France, Beaulieu-en-Argonne, Triancourt, AY176377 AY176378J.E. Lange 25.10.1998, H.A. Huijser s.n. (herb. Huijser)
Lepiota ignicolor Bres. Sect. Stenosporae The Netherlands, prov. Limburg, Savelsbos, Trich- AY176472terberg, 17.10.1999, H.A. Huijser s.n. (herb. Huijser)
Lepiota ignivolvata Bousset & Joss. Sect. Lepiota France, dépt. Aude, Saissac, exhibition, AY17647316.10.1997, E.C. Vellinga 2127 (L)
Lepiota jacobi Vellinga & Knudsen Echinoderma The Netherlands, prov. Noord-Holland, Amsterdam, AY176474Amstelpark, 23.9.1998, E.C. Vellinga 2257 (L)
Lepiota lilacea Bres. Sect. Lepiota U.S.A., Michigan, Washtenaw Co., Ann Arbor, AY176379 AY1763806.8.2000, E.C. Vellinga 2451 (UC)
Lepiota luteophylla Sundb. Sect. Lepiota U.S.A., Michigan, Washtenaw Co., Sharon AY176475Hollow, 25.8.1972, H.V. Smith 284 (MICH)
Lepiota magnispora Murrill Sect. Lepiota Germany, Baden-Württemberg, NE of Riedheim, AF3910052.9.1994, M. Enderle s.n. (L)
Lepiota magnispora Sect. Lepiota U.S.A., California, Mendocino Co., Van Damme AF391010State Park, 19.11.2000, E.C. Vellinga 2520 (UC)
Lepiota magnispora Sect. Lepiota The Netherlands, prov. Limburg, Hopel-Kerkrade, AF391021mine Laura-Julia, 19.9.1998, E.C. Vellinga 2246 (L)
Lepiota magnispora Sect. Lepiota U.S.A., Arizona, Cochise Co., Chiricahua AY176381Mountains, Onion Pass, 4.9.1993, J. States 1830 (AEF1015) (MICH)
© DGfM 2003
Mycological Progress 2(4) / 2003 319
Tab. 1. Continued

Species Classification Collection Particulars ITS LSUaccording to GenBank GenBankBON (1993) accession # accession #
Lepiota nigrescentipes G. Riousset Sect. Helveolae France, dépt. Bouche du Rhône, Maillane, AY176382 AY1763835.10.1993, G. Riousset 93051001 (herb. Riousset)
Lepiota ochraceofulva P.D. Orton Sect. Felinae The Netherlands, prov. Zuid-Holland, Rijswijk, AY176386Ter Werve, 30.9.1998, C.B. Uljé (coll. E.C. Vellinga 2273) (L)
Lepiota ochraceofulva Sect. Felinae The Netherlands, prov. Flevoland, Noordoostpolder, AY176387Urkerbos, 23.9.1998, E.C. Vellinga 2267 (L)
Lepiota perplexa Knudsen Echinoderma The Netherlands, prov. Limburg, Elsloo- Geulle, AY176388 AY176389Bunderbos, 19.9.1998, E.C. Vellinga 2228 (L)
Lepiota petasiformis Murrill Sect. Fuscovinaceae U.S.A., California, San Mateo Co., San Francisco AF391035Watershed, near Pulgas Water Temple, 8.12.2000, E.C. Vellinga 2572 (UC)
Lepiota pilodes Vellinga & Huijser Sect. Stenosporae The Netherlands, prov. Limburg, Elsloo-Geulle, AY176476Bunderbos, 19.9.1998, H.A. Huijser s.n. (herb. Huijser)
Lepiota psalion Huijser & Vellinga n.a. The Netherlands, prov. Limburg, Valkenburg, Schaels- AY176391berg, 23.8.1999, H.A. Huijser s.n. (herb. Huijser)
Lepiota psalion n.a. The Netherlands, prov. Limburg, Valkenburg, AY176390Schaelsberg, 15.9.1999, H.A. Huijser s.n. (herb. Huijser)
Lepiota pseudolilacea Huijsman Sect. Felinae The Netherlands, prov. Noord-Holland, AY176392Amsterdamse Waterleidingduinen, 4.10.1998,E.C. Vellinga 2278 (L)
Lepiota pseudolilacea Sect. Felinae The Netherlands, prov. Limburg, Brunssum, mine AY176393Hendrik, 7.10.1998, H.A. Huijser s.n. (herb. Huijser)
Lepiota pyrochroa Malençon Sect. Integrellae The Netherlands, prov. Limburg, Elsloo- Geulle, AY176477Bunderbos, 7.9.1996, E.C. Vellinga 2006 (L)
Lepiota rufipes Morgan Sect. Integrellae The Netherlands, prov. Limburg, Bemelen, AF391066 AF4828979.10.1991, H.A. Huijser s.n. (herb. Huijser)
Lepiota scaberula Vellinga Sect. Lilaceae U.S.A., California, San Mateo Co., San Francisco AF391030Watershed, near Pulgas Water Temple, 8.12.2000, E.C. Vellinga 2595 (UC)
Lepiota scaberula Sect. Lilaceae U.S.A., California, San Mateo Co., Wunderlich AY176396County Park, 26.12.1998, E.C. Vellinga 2307 (UC)
Lepiota spec. Sect. Helveolae U.S.A., Michigan, Washtenaw Co., Ann Arbor, AY1764789.8.2000, E.C. Vellinga 2461 (UC)
Lepiota spec. Sect. Lepiota China, Sichuan Prov., Daocheng co., on the way AY176486from Haizishan to Sangdui, 1.7.1998, Z.-L. Yang 1957 (HKAS 32150; L)
Lepiota spec. n.a. GenBank PA409 AF079752 AF079670
Lepiota spec. n.a. GenBank (as L. subincarnata) U85329 U85294
Lepiota spec. n.a. U.S.A., Hawaii, Kawai, Koke’e State Park, Waimea AY176402 AY176403Lamgon, 28.12.1999, P.B. Matheny 1827 (WTU)
Lepiota spec. n.a. U.S.A., Missouri, Taney Co., Coy Bald at Hercules AY176400 AY176401Glade, 4.7.1999, P.B. Matheny 1486 (WTU)
Lepiota spec. Sect. Stenosporae U.S.A., California, San Mateo Co., San Francisco AY176479Watershed, 8.12.2000, E.C. Vellinga 2601 (UC)
Lepiota spec. Sect. Stenosporae U.S.A., California, San Mateo Co., San Francisco AY176480Watershed, 8.12.2000, E.C. Vellinga 2610 (UC)
Lepiota spec. Sect. Stenosporae U.S.A., California, San Mateo Co., San Francisco AY176484Watershed, 8.12.2000, E.C. Vellinga 2574 (UC)
Lepiota spec. Sect. Stenosporae U.S.A., California, San Mateo Co., San Francisco AY176481Watershed, 8.12.2000, E.C. Vellinga 2603 (UC)
Lepiota spec. n.a. U.S.A., California, San Mateo Co., San Francisco AY176485Watershed, 8.12.2000, E.C. Vellinga 2590 (UC)
Lepiota spec. Echinoderma U.S.A., California, Alameda Co., Berkeley, AY176488Keeler Ave, 19.12.2001, E.C. Vellinga 2806 (UC)
© DGfM 2003
320 Phylogeny of Lepiota
Tab. 1. Continued

Species Classification Collection Particulars ITS LSUaccording to GenBank GenBankBON (1993) accession # accession #
Lepiota spec. Echinoderma U.S.A., California, San Mateo Co., San Francisco AY176482Watershed, 8.12.2000, E.C. Vellinga 2608 (UC)
Lepiota spec. Echinoderma U.S.A., California, San Mateo Co., San Francisco AY176483Watershed, 8.12.2000, E.C. Vellinga 2609 (UC)
Lepiota spec. Echinoderma China, Sichuan Prov., Xiangcheng Co., in the AY176487mountains near Shagong, 3100 m asl, 1.7. 1998, Z.L. Yang 2280 (HKAS 32361; L)
Lepiota spheniscispora Vellinga Sect. Lepiota U.S.A., California, San Mateo Co., San Francisco AF391000Watershed, near Pulgas Water Temple, 4.12.1998,E.C. Vellinga 2298 (UC)
Lepiota spheniscispora Sect. Lepiota U.S.A., California, Marin Co., Mt. Tamalpais, AY17640415.2.2000, E.C. Vellinga 2438 (UC)
Lepiota subalba P.D. Orton Sect. Stenosporae The Netherlands, prov. Limburg, Elsloo- Geulle, AY176489Bunderbos, 19.9.1998, E.C. Vellinga 2242 (L)
Lepiota subgracilis Kühner Sect. Lepiota The Netherlands, prov. Limburg, Valkenburg, AY176490Schaelsberg, 8.10.1991, P.H. Kelderman(coll. E.C. Vellinga 1783) (L)
Lepiota subincarnata J.E. Lange Sect. Helveolae The Netherlands, prov. Limburg, Elsloo- Geulle, AY176491Bunderbos, 19.9.1998, E.C. Vellinga 2234 (L)
Lepiota thiersii Sundb. n.a. U.S.A., California, San Mateo Co., San Francisco AY176492Watershed, 8.12.2000, E.C. Vellinga 2602 (UC)
Lepiota xanthophylla P.D. Orton Sect. Felinae The Netherlands, prov. Limburg, Elsloo- Geulle, AY176405 AY176406Bunderbos, 19.9.1998, E.C. Vellinga 2240 (L)
Leucoagaricus leucothites (Vittad.) Leucoagaricus The Netherlands, prov. Noord-Holland, Amster- AF482865Wasser sect. Annulati damse Waterleidingduinen, Van Lennepkanaal,
15.10.1996, E.C. Vellinga 2050 (L)Leucoagaricus leucothites Leucoagaricus GenBank U85280
sect. Annulati
Limacella glioderma (Fr.: Fr.) Maire n.a. The Netherlands, prov. Limburg, Elsloo- Geulle, AY176451 AY176452Bunderbos, 19.9.1998, E.C. Vellinga 2241 (L)
Limacella glioderma n.a. U.S.A., Michigan, Washtenaw Co., Ann Arbor, AY176453 AY1764547.8.2000, E.C. Vellinga 2456 (UC)
Macrolepiota procera (Vittad.) Singer Macrolepiota sect. The Netherlands, prov. Zeeland, Schouwen- AF482848Macrolepiota Duiveland, Haamstede, 7.11.1998, E.C.
Vellinga 2293 (L)Macrolepiota procera Macrolepiota sect. The Netherlands, prov. Groningen, Ter Apel, AF482880
Macrolepiota Ter Haar, 18.10.1990, R.P.J. de Kok s.n. (L)
Melanophyllum eyrei (Massee) Singer Melanophyllum Sweden, Skåne, par. Vittskövle, Segesholm, AY176493Herremöllan, 27.9.2001, T. Læssøe TL6692 (C)
Melanophyllum haematospermum Melanophyllum U.S.A., California, Mendocino Co., Navarro AF391039 AY176456(Bull.: Fr.) Kreisel River Redwoods State Park, 19.11.2000, E.C.
Vellinga 2517 (UC) Melanophyllum haematospermum Melanophyllum The Netherlands, prov. Limburg, Hopel-Kerkrade, AF391038 AY176455
mine waste heap Laura-Julia, 19.9.1998, E.C. Vellinga 2249 (L)
Melanophyllum haematospermum Melanophyllum The Netherlands, prov. Noord-Holland, Bloem- AY176494endaal, Caprera, 27.9.1997, E.C. Vellinga 2111 (L)
Pseudobaeospora pyrifera Bas & n.a. Germany, Bavaria, Kitzinger, Klosterforst, 10.9.1995, AF391034 AY176457L.G. Krieglst. L. Krieglsteiner s.n., Isotype (REG)
© DGfM 2003
Mycological Progress 2(4) / 2003 321
Tab. 1. Continued

Constraint Tree length -lnL Diff –ln L P
None 2768 12375.63188 (best)
Lepiota including Cystolepiota, Pulverolepiota, 2795 12451.19875 75.56687 0.002*Melanophyllum and sect. Echinatae monophyletic
Lepiota excluding Cystolepiota, Pulverolepiota, 2790 12447.83974 72.20785 0.014*Melanophyllum and sect. Echinatae monophyletic
Cystolepiota s. str. monophyletic 2785 12401.26433 25.63245 0.404
Cystolepiota plus Pulverolepiota monophyletic 2785 12406.70304 31.07116 0.100
Cystolepiota plus Pulverolepiota and Melanophyllum 2780 12408.01734 32.38546 0.190monophyletic
Sect. Echinatae monophyletic 2771 12384.78846 9.15658 0.707
Sect. Ovisporae monophyletic 2802 12480.13909 104.50721 0.000*
Melanophyllum monophyletic 2769 12378.59818 2.96630 0.909
Sect. Lilaceae monophyletic 2775 12394.77661 19.14473 0.216
Chamaemyces fracidus within Lepiota 2776 12390.31366 14.68178 0.498
Coprinus comatus and allies included in Lepiota 2791 12432.31070 56.67882 0.046*
Coprinus comatus et al. and Chamaemyces fracidus 2785 12407.20977 31.57788 0.282within Lepiota
Sect. Echinatae, Cystolepiota, Pulverolepiota plus 2775 12400.29636 24.66448 0.397Melanophyllum monophyletic
Sect. Echinatae, Cystolepiota, Pulverolepiota plus Melano- 2780 12394.77661 19.14473 0.216phyllum monophyletic, and sect. Lilaceae monophyletic
Tab. 2. Results from two-tailed test Kishino-Hasegawa tests for the ITS data set, using RELL bootstrap; the number of boot-strap replicates = 1000 P is the probability of getting a more extreme T-value under the null hypothesis of no difference between the two trees (two-tailed test).* P < 0.05
Tab. 3. Results from the two-tailed Kishino-Hasegawa test using RELL bootstrap of the ITS-LSU data set. The number of boot-strap replicates = 1000 P is the probability of getting a more extreme T-value under the null hypothesis of no difference between the two trees (two-tailed test).* P < 0.05
N.B. Lepiota has been taken in a wide sense, including Cystolepiota, sect. Echinatae, Melanophyllum and Pulverolepiota
Constraint Tree length -lnL Diff –ln L P
none 1987 8842.94713 (best)
Lepiota including Cystolepiota, sect. Echinatae, 1992 8877.36421 34.41708 0.152Melanophyllum and Pulverolepiota monophyletic
Sect. Echinatae monophyletic 1988 8843.51678 0.56965 0.940
Chamaemyces fracidus within Lepiota 2006 8904.64689 61.69976 0.005*
Chamaemyces fracidus and Coprinus comatus and 2006 8869.26898 26.32185 0.149allies within Lepiota
Coprinus comatus and allies within Lepiota, 2012 8912.68769 69.74056 0.000*Chamaemyces not
© DGfM 2003
322 Phylogeny of Lepiota





























