Embed Size (px)
Citation preview

This article was downloaded by:[Canadian Research Knowledge Network]On: 15 February 2008Access Details: [subscription number 789349994]Publisher: Informa HealthcareInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Pharmaceutical BiologyPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t713721640
Secondary Assays for Testing the Mode of Action ofNatural Products with Bioactivity Against FungiRobert Phillip Smith a; Isabel Cruz a; Ashkan Golshani a; Claire Chesnais a; MyronL. Smith aa Department of Biology, Carleton University, Ottawa, Ontario, Canada
Online Publication Date: 01 January 2008To cite this Article: Smith, Robert Phillip, Cruz, Isabel, Golshani, Ashkan,Chesnais, Claire and Smith, Myron L. (2008) 'Secondary Assays for Testing theMode of Action of Natural Products with Bioactivity Against Fungi ', PharmaceuticalBiology, 46:1, 16 - 25To link to this article: DOI: 10.1080/13880200701729695URL: http://dx.doi.org/10.1080/13880200701729695
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article maybe used for research, teaching and private study purposes. Any substantial or systematic reproduction,re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expresslyforbidden.
The publisher does not give any warranty express or implied or make any representation that the contents will becomplete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should beindependently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings,demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with orarising out of the use of this material.

Dow
nloa
ded
By:
[Can
adia
n R
esea
rch
Kno
wle
dge
Net
wor
k] A
t: 16
:58
15 F
ebru
ary
2008
Pharmaceutical Biology2008, Vol. 46, Nos. 1–2, pp. 16–25
Secondary Assays for Testing the Mode of Action of Natural
Products with Bioactivity Against Fungi∗
Robert Phillip Smith,1 Isabel Cruz,1 Ashkan Golshani,1 Claire Chesnais,1 and Myron L. Smith1
Department of Biology, Carleton University, Ottawa, Ontario, Canada
Abstract
Identification of novel antifungals is needed, and plant-derived compounds offer important leads in this respect.This article initiates a compendium of secondary assaysfor testing hypotheses on mode of action of plant-derivedcompounds. Three assays were developed to evaluate com-pounds that are hypothesized to effect cell wall function,transcription, and oxidative stress. A hypothesized pertur-bation on cell wall integrity by chimaphilin and Echinacearoot extracts was supported using sonication and heat shockassays. Heat shock treatments reduced the number of viablecells recovered when yeast was grown in the presence ofsubinhibitory concentrations of chimaphilin. A reductionin cell numbers was also observed with both sonication-based and heat shock–based assays when yeast was grownin the presence of Echinacea purpurea (L.) Moench (Aster-aceae) extracts. The hypothesis that chimaphilin impedesmRNA transcription processes was also supported by a β-galactosidase reporter gene assay. The antioxidant activityof nordihydroguaiaretic acid (NDGA) and ascorbic acidwere assessed using a yeast strain that does not grow un-der hyperoxic conditions. At the concentrations assayed,NDGA did not remediate this sensitivity to hyperoxia,whereas ascorbic acid did. These and additional assays arefurther reviewed with an aim to collate and standardizesecondary assays and streamline antifungal mode of actiondeterminations.
Keywords: Antifungal activity, antioxidant activity, cellwall integrity, gene expression, plant-derived mycotics, sec-ondary assays, traditional medicine.
∗Dedicated to Professor John Tror Arnason of the University of Ottawa, Department of Biology, on the occasion of his sixtieth birthday.
Accepted: September 24, 2007.
Address correspondence to: Robert Phillip Smith, Department of Biology, Carleton University, 1125 Colonel By Drive, Ottawa, Ontario,Canada, K1S 5B7. Tel.: 613 520–2600 ext.1848; Fax: 613 520–3439; E-mail: [email protected]
Introduction
Fungi have beneficial roles in human affairs such as in pro-duction of food, as symbionts, and in nutrient cycling. Fungialso have significant harmful effects on human health, agri-culture, and in food and materials spoilage. As examples,the rice blast fungus, Magnaporthe grisea (Hebert) Barr(Magnaporthaceae), destroys millions of tons of rice eachyear (Valent, 2004), and other fungi cause mycoses that re-sult in livestock loss and increased health care costs. Controlstrategies that include the use of antifungals are thereforerequired to limit which, when, and where fungi grow in ourenvironment. Currently, six main classes of antifungals ex-ist: echinocandins, griseofulvins, polyene macrolides, flu-orinated pyrimidines, allylamines, and azole derivatives(Francois et al., 2005). Widespread use of selected antifun-gals has led to an increase in the incidence of drug-resistantfungal strains. This, along with a limited effectiveness ofantifungals against some pathogenic fungi, has emphasizeda need to develop new antifungals with different activityspectra. The development of new antifungals is compli-cated because, as eukaryotes, fungi are evolutionarily re-lated to plants and animals (Anderson, 2005). Therefore,suitable antifungals must have a high index of specificity sothat they affect fungal-specific processes and do not harmnontarget species. In this respect, it is desirable to obtainantifungals that interfere with diverse, essential, fungal-specific pathways and/or that target multiple pathways sothat resistance is unlikely to readily arise. Consequently, theinitial process of antifungal discovery involves recognitionof possible leads and characterization of the compound,including specificity and mode of action determinations.
DOI: 10.1080/13880200701729695 C© 2008 Informa Healthcare USA, Inc.

Dow
nloa
ded
By:
[Can
adia
n R
esea
rch
Kno
wle
dge
Net
wor
k] A
t: 16
:58
15 F
ebru
ary
2008
Secondary assays for antifungals 17
Plant-derived compounds offer promising leads in thediscovery of new and effective antifungals. Several plant-derived antifungals have been described (review in Cowan,1999). Many of these antifungals are secondary metabolitesthat are likely produced by the plant as a defense against fun-gal pathogens. Clearly, there has been a strong evolutionarypressure on plants to develop chemical defense strategiesthat are not readily overcome by pathogen resistance mech-anisms. Traditional healers have long recognized antisepticproperties of specific plant species, and this knowledge baseserves as a useful starting point in the discovery process.For example, in a study by Arnason’s group (Jones et al.,2000), it was found that plants used most often for treat-ing infection-like symptoms by First Nations Peoples ofeastern Canada had higher overall antifungal activities thanmedicinal plants used for other purposes.
Once identified, isolation and characterization of antifun-gal compound(s) from a plant is effectively done throughbioassay-guided fractionation, a process by which a crudeplant extract is fractionated based on chemical character-istics (solubility, polarity, etc.) and each fraction is subse-quently assayed for antifungal activity (Ficker et al., 2003).Activity can be measured using methods such as disk diffu-sion and broth dilution assays. In addition, high-throughputmethods involving gene deletion arrays (GDAs) and DNAmicroarrays are being developed to elucidate mode of actionof antifungals. GDA analysis, in particular, holds promisein antifungal mode of action determinations. GDA basedon the yeast Saccharomyces cerevisiae Meyen ex. E. C.Hansen (Saccharomycetaceae) consists of inoculating eachof ∼4700 viable deletion mutants onto medium contain-ing quasi-inhibitory concentrations of the test compoundfollowed by growth analyses of each mutant after a suit-able incubation period. A synthetic sick phenotype resultsfrom the combination of a specific gene deletion and achemical interaction (Tong et al., 2004). Recognition ofthis synthetic sick phenotype can be automated by digitalanalysis of colony size (Memarian et al., 2007). Inferringmode of action from GDA data sets requires analysis of thefunction of knockout genes in supersensitive mutants fol-lowed by identification of candidate parallel pathways thatmay be disrupted by the chemical. Mode of action usingthis approach was demonstrated with rapamycin, a natu-rally occurring drug produced by Streptomyces hygroscop-icus (Jensen) Waksman and Henrici (Streptomycetaceae)(Xie et al., 2005) and is ongoing with several ethnobotan-ical compounds including those found in Echinacea, andwith chimaphilin and berberine (Mirrashed, 2007; Smith,Arnason, Golshani et al., personal communication). Re-cent developments to automate this data analysis step willprovide rapid inference on mode of action from GDA ex-periments (Dumontier, 2006). Aside from the complexityof identifying antifungal mode of action from GDA datasets, the incidence of false positives associated with high-throughput assays must be acknowledged. It is thereforedesirable to test hypotheses on mode of action obtained inprimary assays using secondary assays.
Secondary assays to verify antifungal mode of actionare not standardized, and the methods are dispersed in theliterature. The goal of this study is to initiate a compendiumof secondary assays that evaluate antifungal mode of action.Assays are developed that set out to test the in vivo effectsof phytochemicals on fungal mRNA transcription, cell wallintegrity, and antioxidant activities, and additional assaysare reviewed.
Materials and Methods
General methods
Echinacea purpurea (L.) Moench (Asteraceae) roots wereextracted and fractionated as described in Hudson et al.(2005). Chimaphilin was purified from dried Chimaphilaumbellata (L.) W. Bart (Pyrolaceae) as described by Galvanet al. (Galvan et al., 2007). Nordihydroguaiaretic acid(NDGA) and ascorbic acid were obtained from Calbiochem(Darmstadt, Germany). Four S. cerevisiae strains wereused in this study: S288C (MATα SUC2 gal2 mal melflo1 flo8–1 hap1; Mortimer & Johnston, 1986), DAN1�
(MATαA his3�1/his3�1 leu2�0 /leu2�0 lys2�0/LYS2MET15/met15�0 ura3�0 /ura3�0 YJR150C::G418R; Gi-aever et al., 2002), SOD1� (MATαA his3�1/his3�1leu2�0 /leu2�0 lys2�0/LYS2 MET15/met15�0 ura3�0/ura3�0 YJR104C::G418R; Giaever et al., 2002),and YBR115C� (MATαA his3�1/his3�1 leu2�0/leu2�0 lys2�0/LYS2 MET15/met15�0 ura3�0 /ura3�0YBR115C::G418R; Giaever et al., 2002). Yeast strains werecultured by standard methods (Ausubel et al., 1997) witheither YPD (1% Bacto yeast extract, 2% Bacto peptone, 2%glucose; BD, Sparks, MD, USA) or Sabouraud’s medium(EM Science, Gibbstown, NJ, USA), with or without 2%Agar (BD), or with SC-ura medium (0.67% nitrogen base,2% glucose, 20 mg/mL amino acids; Sigma-Aldrich, St.Louis, MO, USA).
Transcriptional activity assay
For this assay, we used strain YBR115C� expressing theconstruct pUKC815 that contained the β-galactosidasereporter gene. Mid-log phase cells were grown in SC-ura with subinhibitory amounts of chimaphilin, or withthe carrier control (EtOH), for 4 h at 30◦C. Yeast cellswere harvested by centrifugation, resuspended in Z-buffer(60 mM Na2HPO4, 40 mM NaH2PO4, 10 mM KCl, 1 mMMgSO4·7H2O, 0.27% β-mercaptoethanol, pH 7.0) and kepton ice. Initial cell density was measured at OD600, and β-galactosidase activity was measured as described by Miller(1972). Briefly, 20 µL of 0.1% SDS and 50 µL of chlo-roform were added to each sample, equilibrated in a 30◦Cwater bath, and mixed with 200 µL of prewarmed ONPG(o-nitrophenyl-β-D-galactopyranoside, 4 mg/mL in 0.1 Mpotassium phosphate; Sigma-Aldrich). The reaction wasstopped with 500 µL of 1 M Na2CO3. The cell debriswas removed by centrifugation and units of β-galactosidase
Dow
nloa
ded
By:
[Can
adia
n R
esea
rch
Kno
wle
dge
Net
wor
k] A
t: 16
:58
15 F
ebru
ary
2008
18 R.P. Smith et al.
activity was quantified spectrophotometrically using thefollowing equation: U = 1000[OD420 – (1.75 · OD550)/(10(t · V · OD600)], where V is volume (mL) of cell suspensionused and t is time in minutes.
Cell wall integrity assays
Yeast strain S228C was grown overnight in YPD to mid-log phase and adjusted to approximately 104 cells/mL.The culture (2 mL) was transferred to a disposable cul-ture tube and incubated overnight with gentle shaking at30◦C with either chimaphilin (3.0 µg/mL) or Echinaceaextract (1.42 mg/mL of 70% EtOH extract). Controls con-sisted of cultures to which the equivalent volume of carriersolvent (EtOH) was added without compound. After incu-bation, the cell density was adjusted to 107cells/mL. Cellwall disruption was done either by heat shock treatments ina 65◦C water bath for 0, 30, 40, and 50 s or by sonication(Vibra-Cell, Sonics and Materials, Danbury, CT, USA) ata 3-µm amplitude for durations of 0, 135, and 140 s. Af-ter heat shock or sonication treatments, cell viability wasestimated by drop-plate assays as described in Chen et al.(2003) using a 10-fold dilution series.
Antioxidant activity assay
The knockout mutants DAN1� [referred to as wild-type(wt) hereafter because it is not sensitive to hyperoxia] and
SOD1� were grown overnight in YPD to mid-log phaseand 10x serially diluted before drop-plate assays were doneon YPD medium containing NDGA (0, 3.13, 6.25, 12.5, 25,50, and 100 µM), ascorbic acid (0, 2.5, 5, 10, 20, 40, and80 mM), or equivalent volumes of carrier solvent (EtOHand water, respectively). Replicate plates were incubatedseparately under normoxic (ca. 21% O2) and hyperoxic (ca.100% O2) conditions at 30◦C for 48 h at which time colonycounts were done.
Results and Discussion
Reporter gene expression assay to test effects on fungaltranscription
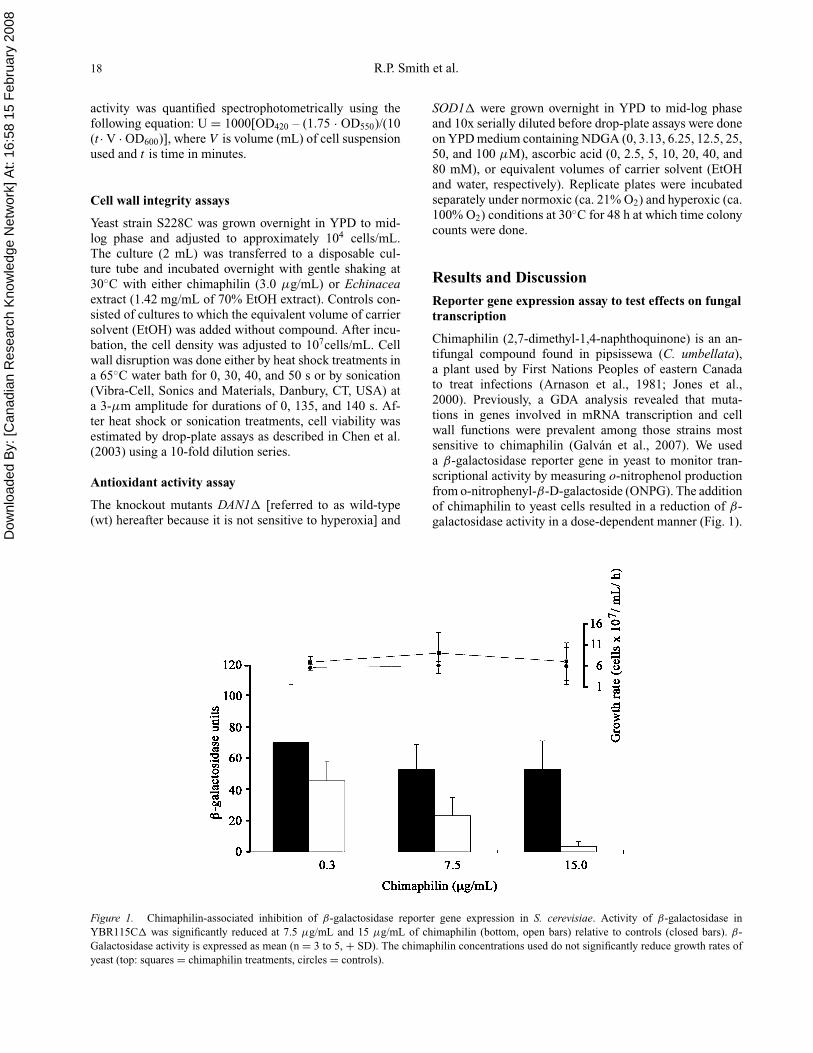
Chimaphilin (2,7-dimethyl-1,4-naphthoquinone) is an an-tifungal compound found in pipsissewa (C. umbellata),a plant used by First Nations Peoples of eastern Canadato treat infections (Arnason et al., 1981; Jones et al.,2000). Previously, a GDA analysis revealed that muta-tions in genes involved in mRNA transcription and cellwall functions were prevalent among those strains mostsensitive to chimaphilin (Galvan et al., 2007). We useda β-galactosidase reporter gene in yeast to monitor tran-scriptional activity by measuring o-nitrophenol productionfrom o-nitrophenyl-β-D-galactoside (ONPG). The additionof chimaphilin to yeast cells resulted in a reduction of β-galactosidase activity in a dose-dependent manner (Fig. 1).
Figure 1. Chimaphilin-associated inhibition of β-galactosidase reporter gene expression in S. cerevisiae. Activity of β-galactosidase inYBR115C� was significantly reduced at 7.5 µg/mL and 15 µg/mL of chimaphilin (bottom, open bars) relative to controls (closed bars). β-Galactosidase activity is expressed as mean (n = 3 to 5, + SD). The chimaphilin concentrations used do not significantly reduce growth rates ofyeast (top: squares = chimaphilin treatments, circles = controls).

Dow
nloa
ded
By:
[Can
adia
n R
esea
rch
Kno
wle
dge
Net
wor
k] A
t: 16
:58
15 F
ebru
ary
2008
Secondary assays for antifungals 19
Addition of 0.3 µg/mL of chimaphilin had a small effect inreducing o-nitrophenol production by yeast cells (p = 0.40),whereas at concentrations of 7.5 and 15 µg/mL, a signifi-cant reduction was observed, compared with the carrier sol-vent controls (p = 0.01 and p =<0.001, respectively; pairedt-test). To verify that chimaphilin concentrations used weresubinhibitory to growth, cell density was measured beforeand after assay and used to calculate growth rates (Fig. 1).There was no significant difference in growth rates of thecells (p = 0.43) at the chimaphilin and EtOH (control) con-centrations used. Therefore, chimaphilin appears to inhibitβ-galactosidase expression, consistent with the hypothe-sis that chimaphilin interferes with mRNA transcriptionprocesses. However, from these experiments, we cannotrule out the possibility that chimaphilin interferes with pro-tein synthesis. Additional assays that set out to monitorwhether a compound inhibits protein synthesis, as describedbelow, could offer additional insight into chimaphilin’smode of action. More directly, developing an RT-PCR as-say that will determine transcription rates in cells exposedto chimaphilin is worthwhile given the results presentedhere. Such RT-PCR methods are highly quantitative andfast but otherwise similar to those that used Northern anal-ysis to test the mode of action of azoxybacilin (Aoki et al.,1996).
Cell wall integrity assays
The cell wall is an obvious fungal-specific target for anti-fungal development. Fungal cell walls are composed pri-marily of β-glucans and chitin and are integral to survivalof most free-living fungi (Debono & Gordee, 1994). Tradi-tionally, incorporation of calcofluor white into the growth
medium has been used to assay for fungal cell wall mu-tants (Ram et al., 1994). Calcofluor white binds to cellwalls and appears to inhibit cell wall biogenesis or otherfunctions. Mutants that are viable but defective in somecell wall factors have increased sensitivity to and growpoorly in the presence of calcofluor white (Ram et al.,1994). This assay may not be suitable for evaluating an-tifungal effects on cell walls, however, as both calcofluorwhite and the antifungal compound must be added to thegrowth medium and this could result in synergistic or con-founding interactions. It is therefore desirable to assay forcell wall integrity using physical stress. To test hypothe-ses based on previous GDA experiments that chimaphilinand Echinacea extracts interfere with cell wall–related pro-cesses, we used heat shock–based and sonication-basedassays.
To examine cell wall integrity in yeast challenged withchimaphilin, S288C cells were grown overnight in the pres-ence of subinhibitory (3.0 µg/mL) levels of chimaphilin or,as controls, in an equivalent volume of the carrier solvent(95% EtOH). Cultures were then heat shocked at 65◦C andcell viability was estimated by drop-plate assays on YPDplates. Figure 2 shows how the combination of chimaphilinwith 30 or 40 s of exposure to 65◦C, reduced the numberof viable cells by 10-fold compared with the EtOH control.With 50 s of 65◦C treatment, viable cells no longer occurredin either the chimaphilin- or EtOH-treated cultures. The in-creased sensitivity to the shorter heat shock durations usedsupports the hypothesis that chimaphilin interferes withfungal cell wall functions. A rapid thermal expansion ofcells likely results in increased frequencies of death whencell wall integrity is compromised through chimaphilin ex-posure. From these observations, it cannot be ruled out thatproteins involved in heat shock recovery may be targets of
Figure 2. Drop-plate assays of S288C treated with 3.0 µg/mL of chimaphilin followed by heat shock at 65◦C. This assay suggests a loss of cellwall integrity occurs when yeast cells are exposed to chimaphilin (Ch) compared with controls (CC).

Dow
nloa
ded
By:
[Can
adia
n R
esea
rch
Kno
wle
dge
Net
wor
k] A
t: 16
:58
15 F
ebru
ary
2008
20 R.P. Smith et al.
Figure 3. The effect of sonication on S288C cells treated with 1.42 mg/mL of 70% EtOH extracts of E. purpurea root. Increased sensitivityto 2.15 and 2.30 min of sonication in Echinacea-treated cells (open bars) compared with controls (closed bars) is evident by determining colonyforming units (CFU) in drop-out plates. Values are means from two independent experiments.
chimaphilin. This is unlikely, however, as GDA analysis didnot reveal interactions between chimaphilin and heat shockproteins.
E. purpurea root extracts are also hypothesized to inhibitfungal cell wall functions based on previous GDA experi-ments (Mirrashed, 2007). We tested this hypothesis using asonication assay on yeast S228C cells grown overnight inconcentrations of E. purpurea extract that caused a 20% re-duction in growth rate. Sonication was previously describedas an efficient assay to screen for cell wall mutants (Ruizet al., 1999). Treated cells sonicated for 2.15 and 2.30 minshowed a marked reduction in cell viability based on drop-plate assays compared with cells grown in the presenceof equivalent volumes of the 70% EtOH carrier solvent(Fig. 3). The results of this sonication assay and heat shockassays (data not shown) were congruent, and both lend sup-port to the hypothesis that E. purpurea extracts interferewith cell wall functions.
Further studies are required to reveal the cell wall tar-get(s) of chimaphilin and Echinacea extracts. For this pur-pose, enzymes involved in cell wall biogenesis can be as-sayed in vitro using previously described methods. Suchapproaches may require considerable trial and error, butmay be effective in high-throughput screens. For exam-ple, the targets of the triterpene glycoside FR227244 wereinvestigated using an in vitro enzymatic assay containingglucan or chitin synthase that monitored the incorpora-tion of radiolabeled UDP-glucose into cell wall subunits(Kobayashi et al., 2004). Monitoring cell permeability withfluoresecent dyes such as Sytox green may provide fur-ther indications of cell wall defects caused by antifungals(Munoz et al., 2006). A judicious approach involves the useof multiple different assays to test hypotheses on mode ofaction as cellular processes are complex and highly inter-related. Cell wall defects, for example, could result from
specific transcriptional or translational alterations or fromperturbations to polymerization, deposition, or stabiliza-tion processes involving cell wall components. It is alsoimportant to keep in mind that plant-based antifungals mayinhibit multiple unrelated cellular processes. This would bean appropriate strategy to avoid pathogen resistance mech-anisms.
In vivo antioxidant assays
Wound healing is a complex process that can be enhancedby application of antibiotics and other biologically activecompounds such as antioxidants. In addition, several an-tioxidants, such as NDGA, have been shown to have a syn-ergistic effect when combined with an antifungal (Beggs etal., 1978). NDGA is the primary antioxidant found in thecreosote bush [Larrea tridentate (Sesse & Moc. ex DC.)Coville (Zygophyllaceae)], which has been used in tradi-tional medicine against at least 50 ailments including kid-ney stones, diabetes, inflammation, and arthritis (Arteagaet al., 2005). Previous studies also confirmed NDGA asan antifungal against pathogenic fungi (Beggs et al., 1978;Martinez-Tellez et al., 2005) and that it has antioxidantactivity in vitro and in vivo (Harper et al., 1999). We exam-ined antioxidant activities of subinhibitory levels of NDGAusing an oxidative stress assay with SOD1�, a superox-ide dismutase knockout strain. The SOD1� mutant doesnot normally grow under hyperoxic conditions as evidentfrom drop-plate assays in which cells were incubated for2 days under 100% oxygen conditions (Fig. 4A). Growthof both wild-type and SOD1� strains occurs under nor-moxia (Fig. 4B). As a positive antioxidant control, ascorbicacid was included in the medium and clearly remediatedhyperoxic sensitivity of SOD1� cells (Fig. 4A; Krzepilko

Dow
nloa
ded
By:
[Can
adia
n R
esea
rch
Kno
wle
dge
Net
wor
k] A
t: 16
:58
15 F
ebru
ary
2008
Secondary assays for antifungals 21
Figure 4. The effects of ascorbic acid and NDGA on wild-type (WT) and SOD1�-defective strain growth under normoxia and hyperoxia.Drop-plates of WT and SOD1� strains grown (A) under hyperoxia where growth of SOD1� is remediated by ascorbic acid and (B) undernormoxia where both strains grow regardless of additives used. (C) Plot showing how NDGA does not remediate hyperoxia sensitivity of SOD1�
whereas ascorbic acid does. Relative concentrations are given at bottom, where “x” is 6.25 µL carrier solvent (water and EtOH for ascorbic acidand NDGA), with 2.5 mM ascorbic acid or 3.125 µM NDGA.

Dow
nloa
ded
By:
[Can
adia
n R
esea
rch
Kno
wle
dge
Net
wor
k] A
t: 16
:58
15 F
ebru
ary
2008
22 R.P. Smith et al.
Table 1. Initial compendium of secondary mode of action assays to test antifungal activity in natural products.
Function assayed Test type Reference Method
Cell wall (CW)CW integrity In vivo This study Sonication of extract-treated cells followed by CFU determination.CW integrity In vivo This study Heat shock–treated cells followed by CFU determination.CW integrity In vivo Ruiz et al., 1999 Sonication followed by cell damage assessment by flow cytometry.CW synthesis In vivo Straede et al., 2007 Cell wall biosynthesis transcription factor (Rlm1p) fused to β-gal to
monitor β-gal activity during cell wall stress.CW synthesis In vivo Evans et al., 2002 Pico Green fluorescence measured after cells were subjected to
osmotic shock.Chitin and glucansynthase inhibition
In vitro Kobayashi et al., 2004 Enzyme assay measures incorporation of 14C-UDP-glucose into thecell wall.
1,3-β-D-Glucan synthaseinhibition
In vitro Kurtz et al., 1994 Measured rate of 3H-UDP-glucose incorporation into TCA precipitategenerated by fungal cell membrane extracts.
1,3 and 1,6-β-D-Glucansynthesis inhibition
In vivo Feldmesser et al., 2000 1,3 or 1,6-β-D-Glucan antisera used with immunoelectron microscopy.
Chitin degradation In vitro Mavromatis et al., 2003 Antifungals incubated with chitin and reaction products observedusing HPLC.
Chitin synthesis In vivo Larcher et al., 2004 Treated cells examined microscopically for chitin deposition usingFITC-conjugated wheat germ agglutinin.
CW permeation In vivo Munoz et al., 2006 Observe uptake of Sytox green after incubation with antifungal.Cell membrane (CM)
CM permeation In vivo Endo et al., 1997 Examines release of glutamine and arginine.CM permeation In vivo Terras et al., 1993 Quantified K+ efflux from cells using atomic absorption spectrometry.Sterol biosynthesis In vivo Sanati et al., 1997 Quantified and identified sterols extracted from cells challenged with
antifungal.Sterol biosynthesis In vivo Ryder et al., 1986 Examined the incorporation of U-14C-acetate and
L-methyl-14C-methionine into ergosterol.Sterol biosynthesis In vitro Ryder et al., 1986 Examine production of ergosterol from 2−14C-mevalonate.Sterol biosynthesis In vivo Buurman et al., 2004 ERG10 fused to β-galactosidase reporter to measure β-gal activity in
response to ergosterol synthesis inhibitors.Transcription
mRNA transcription In vivo This study β-Galactosidase reporter to examine global transcription.RNA polymeraseinhibitor
In vitro Jimenez et al., 1973 Measured incorpotation of 14C-UTP into RNA using extractedRNA-polymerases.
Gene expression In vivo Aoki et al., 1996 Examined expression of putative target genes using Northern blot.Translation
Protein synthesiselongation
In vitro Dominguez et al., 1998 Monitored incorporation of 14C phenylalanine into TCA precipitate.
Aminoacyl-tRNAinhibition
In vitro Ziegelbauer et al., 1998 Monitered the incorporation of 14C-iosleucine and 14C-lysine intoTCA precipitate using aminocyl-tRNA.
Ribosome signaling In vivo Foiani et al., 1991 Examine alteration in ratios of ribosomal subunits.Translation fidelity In vivo Carr-Schmid et al., 1999 Monitor increase in β-gal activity from construct with premature stop
codon.Translation In vivo Gaynor et al., 1994 Measure incorporation of 35S into proteins.
OtherGlobal macromolecularsynthesis
In vivo Onishi et al., 2000 Pulse-labeled cells with various radiolabelled precursors.
Sulfur fixation In vitro Aoki et al. 1996 Monitored incorporation of 35SO4 into acid-soluble fractions.Topoisomerase inhibition In vivo Spaltmann et al., 1999
Khan et al., 2002Used topoisomerase I and II knockouts to show remediation of
sensitivity antifungals.Proton pump inhibition In vitro Lunde and Kubo, 2000 Measured ATP-hydrolysis of extracted protein pumps.Pyrimidine salvaging In vitro Diasio et al., 1978 Measured incorporation of 3H-dUMP by thymidylate synthetase.Respiration In vivo Uno et al., 1982 Measured O2consumption during growth.Interaction with thiols In vitro Miron et al., 2000 Monitored glutathione and 2-nitro-5-thiobenzoate using DNTB.Calcineurin signaling In vivo Parsons et al., 2006 Calcineurin-dependent response elements fused to β-galactosidase
reporter.DNA synthesis In vitro Smith et al., 2004 PCR-based assay for bioadduct formation.

Dow
nloa
ded
By:
[Can
adia
n R
esea
rch
Kno
wle
dge
Net
wor
k] A
t: 16
:58
15 F
ebru
ary
2008
Secondary assays for antifungals 23
et al., 2004). As seen in Figure 4C, addition of ascorbicacid restores growth of SOD1� in a dose-dependent man-ner and completely restored hyperoxic growth of SOD1�
to near wild-type levels when 80 mM ascorbic acid wasadded to the medium. In contrast, the addition of NDGA tothe medium did not decrease sensitivity of SOD1� to hy-peroxia, suggesting that NDGA is not an effective antioxi-dant in this in vitro assay. The apparent lack of antioxidantactivity by NDGA in our assay could relate to bioavailabil-ity, as NDGA is not water-soluble. Interestingly, NDGAdid not appear to inhibit yeast growth, even at the high-est concentrations we assayed. Higher concentrations ofNDGA, up to levels that clearly inhibit growth of yeast,should therefore be tested for antioxidant activity. Thereare several additional yeast knockouts involving distinctpathways from those of SOD1� that are also unable togrow under hyperoxic conditions (Golshani et al., personalcommunication). It is possible, therefore, that NDGA mayspecifically interact with other pathways as an antioxidant,and further in vivo studies with these other mutants arerecommended.
Inhibition of protein synthesis and other assays
The process of translation or protein synthesis is one ofthe most essential and complex cellular pathways, and be-cause of its absolute requirement for the survival of the cell,many naturally occurring antifungals, such as hygromycinB, target this process for their activities. Consequently, it isuseful to establish a systematic approach to analyze the ef-fect of a newly identified antifungal on this process. Trans-lation may be divided into three subcategories: ribosomebiogenesis, translation fidelity, and translation efficiency.Ribosome biogenesis refers to all the pathways leading tothe formation of mature ribosomes. One way to examinethe effect of a target compound on ribosome biogenesisis through ribosome profile analysis (Foiani et al., 1991).Ribosome profiling refers to the determination of the ra-tio of cellular ribosomal subunits to each other. A typicalprofile in the presence of a target compound may indicatealterations for the ratios of free 40S to 60S subunits, 40Sto 80S subunits, or 40S to polysomes, as well as produc-tion of premature ribosomal subunits. Translation fidelitycan be most readily studied by measuring the in vivo rateof premature stop codon read-through. This can be deter-mined using specialized β-gal expression plasmids such aspUKC817, pUKC818, and pUKC819 (Carr-Schmid et al.,1999), each containing a different premature stop codon inthe coding regions of their β-gal genes. The presence of abioactive compound that targets translation fidelity can in-crease the production of β-gal from such reporter cassettes.It is noteworthy that an increase in β-gal production in thesesystems may also be explained by other means such as anincrease in the rate of transcription (as in the transcriptionassay described above). Therefore, proper controls, such aswild-type β-gal reporter plasmids, and accompanying ex-
periments, such as determination of mRNA content, shouldalso be considered. Other bioactive compounds may af-fect the rate of translation. This can be analyzed by mea-suring the rate of incorporation of radioactively labeledamino acids into newly synthesized proteins. The presenceof 35S-methionine in the medium (Gaynor et al., 1994) isthe simplest and most common approach for this type ofanalysis.
Table 1 provides a list of secondary assays used in thispaper and from literature sources. The list includes assaysfor processes involving cell wall, cell membrane, transcrip-tion, translation, and other functions. As the study of plant-based antifungals develops, it is recommended that thiscompendium develop into a set of standard methods thatcan be used to further test mode of action.
Conclusions
The compendium initiated here is intended as a tool toguide investigations into mode of action in novel or unchar-acterized antifungals. Three assays were used to examinethe effects of antifungals on the cell wall and mRNA tran-scription in fungi. In addition, an in vivo assay was usedto examine antioxidant activity of the compound NDGA.These secondary assays are based on the yeast system butcan readily be adapted to test similar mode of action pro-cesses in fungal species that are more relevant to plant andanimal pathology.
Acknowledgments
This work was supported by OGS and NSERC CGS toR.P.S. and by NSERC Discovery Grants to M.L.S and A.G.
References
Anderson JB (2005): Evolution of antifungal drug resistance:Mechanisms and pathogen fitness. Nature Rev 3: 547–556.
Aoki Y, Yamamoto A, Hosseini-Mazinani SM, Koshikawa N,Sugimoto K, Arisawa M (1996): Antifungal azoxybacilinexhibits activity by inhibiting gene expression of sulfite re-ductase. Antimicrob Agents Chemother 40: 127–132.
Arnason T, Hebda RJ, Johns T (1981): Use of plants for food andmedicine by native peoples of eastern Canada. Can J Bot 59:2189–2325.
Arteaga S, Andrade-Cetto A, Cardenas R (2005): Larrea triden-tata (Creosote bush), an abundant plant of Mexican and US-American deserts and its metabolite nordihydroguaiareticacid. J Ethnopharmacol 98: 231–239.
Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG,Smith JA, Struhl K (1997): Current Protocols in MolecularBiology. New York, John Wiley & Sons, pp. 13.1–13.3.
Beggs WH, Andrews FA, Sarosi GA (1978): Synergistic action ofamphotericn B and antioxidants against certain opportunisticyeast pathogens. Antimicrob Agents Chemother 13: 266–270.

Dow
nloa
ded
By:
[Can
adia
n R
esea
rch
Kno
wle
dge
Net
wor
k] A
t: 16
:58
15 F
ebru
ary
2008
24 R.P. Smith et al.
Buurman ET, Blodgett AE, Hull KG, Carcanague D (2004):Pyridines and pymidines mediating activity against andefflux-negative strain of Candida albicans through puta-tive inhibition of lanosterol demethylase. Antimicrob AgentsChemother 48: 313–318.
Carr-Schmid A, Durko N, Cavallius J, Merrick WC, Kinzy TG(1999): Mutations in a GTP-binding motif of eukaryoticelongation factor 1A reduce both translational fidelity andthe requirement for nucleotide exchange. J Biol Chem 274:30297–30302.
Chen C, Nace GW, Irwin PL (2003): A 6 × 6 drop plate methodfor simultaneous colony counting and MPN enumerationof Campylobacter jejuni, Listeria monocytogenes, and Es-cherichia coli. J Microbiol Methods 55: 475–479.
Cowan MM (1999): Plant products as antimicrobial agents. ClinMicrobiol Rev 12: 564–582.
Debono M, Gordee RS (1994): Antibiotics that inhibit fungal cellwall development. Annu Rev Microbiol 48: 471–497.
Diasio RB, Bennett JE, Myers CE (1978): Mode of action of5-fluorocytosine. Biochem Pharmacol 27: 703–707.
Dominguez JM, Kelly VA, Kinsman OS, Marriott MS, De lasHeras FG, Martin JJ (1998): Sordarins: A new class of an-tifungals with selective inhibition of the protein synthesiselongation cycle in yeast. Antimicrob Agents Chemother 42:2274–2278.
Dumontier M (2006): YeastFeatures: Identifying significantfeatures shared among yeast proteins for functional ge-nomics. http://software.dumontierlab.com/yeastfeatures. Ac-cessed December 17th, 2007.
Endo M, Takesako K, Kato I, Yamaguhi H (1997): Fungicidalaction of aureobasidin A, a cyclic depsipeptide antifungalantibiotic, against Saccharomyces cerevisiae. AntimicrobiolAgents Chemother 41: 672–676.
Evans JM, Zaworski PG, Parker CN (2002): A high throughputscreen for inhibitors of fungal cell wall synthesis. J BiomolScreening 7: 359–366.
Feldmesser M, Kress Y, Mednick A, Casadevall A (2000): Theeffect of the echinocandin analogue caspofungin on cell wallglucan synthesis by Cryptococcus neoformans. J Infect Dis182: 1791–1795.
Ficker C, Smith ML, Akpagana K, Gbeassor M, Zhang J, DurstTR, Assabgui R, Arnason JT (2003): Bioassay-guided isola-tion and identification of antifungal compounds from ginger.Phytother Res 17: 897–902.
Foiani M, Cigan AM, Paddon CJ, Harashima S, Hinnebusch AG(1991): GCD2, a translational repressor of the GCN4 gene,has a general function in the initiation of protein synthe-sis in Saccharomyces cerevisiae. Mol Cell Biol 11: 3203–3216.
Francois IEJA, Aers AM, Cammue BPA, Thevissen K (2005):Currently used antimycotics: Spectrum, mode of action andresistance occurrence. Curr Drug Targets 6: 895–907.
Galvan IJ, Mir-Rashed N, Jessulat M, Atanya M, Golshani A,Durst T, Petit P, Treyvaud-Amiguet V, Boekhout T, Sum-merbell R, Cruz I, Arnason JT, Smith ML (2007): Anti-fungal and antioxidant activities of the phytomedicine Pip-
sissewa, Chimaphila umbellata. Phytochemistry. Availableonline ahead of print. (doi: 10.1016).
Gaynor EC, te Heesen S, Graham TR, Aebi M, Emr SD (1994):Signal-mediated retrieval of a membrane protein from theGolgi to the ER in yeast. J Cell Biol 127: 653–665.
Giaever G, Chu AM., Ni L, et al. (2002): Functional profiling ofthe Saccharomyces cerevisiae genome. Nature 418: 387–391.
Harper A, Kerr DJ, Gescher A, Chipman JK (1999): Antioxidanteffects of isoflavonoids and lignans, and protection againstDNA oxidation. Free Radic Res 31: 149–160.
Hudson J, Vimalanathan S, Kang L, Treyvaud Amiguet V, LiveseyJ, Arnason JT (2005): Characterization of antiviral activitiesin Echinacea root preparations. Pharm Biol 43: 790–796.
Jimenez A, Tipper DJ, Davie J (1973): Mode of action of thiolutin,an inhibitor of macromolecular synthesis in Saccharomycescerevisiae. Antimicrob Agents Chemother 3: 729–738.
Jones NP, Arnason JT, Abou-Zaid M, Akpagana K, Sanchez-Vindas P, Smith ML (2000): Antifungal activity of extractsfrom medicinal plants used by First Nations Peoples of east-ern Canada. J Ethnopharmacol 73: 191–198.
Khan SI, Nimrod AC, Mehrpooya M, Nitiss JL, Walker LA, ClarkAM (2002): Antifungal activity of eupolauridine and its ac-tion on DNA topoisomerases. Antimicrob Agents Chemother46: 1785–1792.
Kobayashi M, Sato I, Abe F, Nitta K, Hashimoto M, Fujie A,Hino M, Hori Y (2004): FR227244, a novel antifungalantibiotic from Myrothecium cinctum No. 002 II. Biolog-ical properties and mode of action. J Antibiot 57: 788–796.
Krzepilko A, Swiecilo A, Wawryn J, Zadrag R, Koziol S,Bartosz G, Bilinski T (2004): Ascobate restores lifespan ofsuperoxide-dismutase deficient yeast. Free Radic Biol 38:1019–1024.
Kurtz MB, Douglas C, Marrinan J, Nollstadt K, Onishi J, DreikornS, Milligan J, Mandala S, Thompson J, Balkovec JM,Bouffard FA, Dropinski JF, Hammond ML, Zambias RA,Abruzzo G, Bartizal K, McManus B, Garcia ML (1994):Increased antifungal activity of L-733,560, a water-soluble,semisynthetic pneumocandin, is due to enhanced inhibitionof cell wall synthesis. Antimicrob Agents Chemother 38:2750–2757.
Larcher G, Morel C, Tronchin G, Landreau G, Seraphin D,Richomme S, Boucharra J (2004): Investigation of theantifungal activity of caledonixanthone E and other xan-thones against Aspergillus fumigatus. Planta Med 70: 569–571.
Lunde CS, Kubo I (2000): Effect of polygodial on the mito-chondrial ATPase of Saccharomyces cerevisiae. AntimicrobAgents Chemother 44: 1943–1953.
Martınez-Tellez MA, Vargas-Arispuro I, Reyes-Baez R, Rivera-Castaneda G, Rivero-Espejel I (2005): Antifungal lignansfrom the creosotebush (Larrea tridentata). Industrial Cropsand Products 22: 101–107.
Mavromatis K, Lorito M, Woo SL, Bouriotis V (2003): Mode ofaction and antifungal properties of two cold-adapted chiti-nases. Extemeophiles 7: 385–390.

Dow
nloa
ded
By:
[Can
adia
n R
esea
rch
Kno
wle
dge
Net
wor
k] A
t: 16
:58
15 F
ebru
ary
2008
Secondary assays for antifungals 25
Memarian N, Jessulat M, Alirezaie J, Mir-Rashed N, Xu J, ZareireM, Smith M, Golshani A (2007): Colony size measurementof the yeast gene deletion strains for functional genomics.BMC Bioinformatics 8: 117–128.
Miller J (1972): Experiments in Molecular Genetics. Cold SpringHarbor, NY, Cold Spring Harbor Laboratory, pp. 352–355.
Mirrashed NH (2007): Using Saccharomyces cerevisiae geneticarray technologies to understand mode of action of ethnob-otanical mycotics. Ph.D. Dissertation, Carleton University,Ontario, p. 72–93.
Miron T, Rabinkov A, Mirelman D, Wilchek M, Weiner L (2000):The mode of action of allicin: Its ready permeability throughphospholipid membranes may contribute to its biological ac-tivity. Biochim Biophys Acta 1463: 20–30.
Mortimer RK, Johnston JR (1986): Genealogy of principal strainsof the yeast genetic stock center. Genetics 113: 35–43.
Munoz A, Lopez-Garcia B, Marcos JF (2006): Studies on the modeof action of the antifungal hexapeptide PAF26. AntimicrobAgents Chemother 50: 3847–3855.
Onishi J, Meinz M, Thompson J, et al. (2000): Discovery of novelantifungal (1,3)-β-D-glucan synthase inhibitors. AntimicrobAgents Chemother 44: 368–377.
Parsons AB, Lopez A, Givoni IE, et al. (2006): Exploring themode-of-action of bioactive compounds by chemical-geneticprofiling in yeast. Cell 126: 611–625.
Ram AF, Wolters A, Ten Toopen R, Klis FM (1994): A new ap-proach for isolating cell wall mutants in Saccharomyces cere-visiae by screening for hypersensitivity to calcofluor white.Yeast 10: 1019–1030.
Ruiz C, Cid VJ, Lussier M, Molina M, Nombela C (1999): A largescale sonication assay for cell wall mutant analysis in yeast.Yeast 15: 1001–1008.
Ryder NS, Frank I, Dupont M (1986): Ergosterol biosynthesisinhibition by the thiocarbamate antifungal agents tolnaftateand tolciclate. Antimicrob Agents Chemother 29: 858–860.
Sanati H, Belanger P, Fratti R, Ghannoum M (1997): A new tria-zole, voriconazole (UK-109,496), blocks sterol biosynthesisin Candida albicans and Candida krusei. Antimicrob AgentsChemother 41: 2492–2496.
Smith ML, Gregory P, Bafi-Yeboa NFA, Arnason JT (2004): In-hibition of DNA polymerization and antifungal specificityof furanocoumarins present in traditional medicines. Pho-tochem Photobiol 79: 506–509.
Spaltmann F, Lohrmann E, Ziegelbauer K (1999): A target-specificcellular assay for screening of topoisomerase I inhibitors. JBiomol Screening 4: 93–100.
Straede A, Corran A, Bundy J, Heinisch JJ (2007): The effect oftea tree oil and antifungal agents on a reporter for yeast cellintegrity signaling. Yeast 24: 321–334.
Terras FRG, Schoofs HME, Thevissen K, Osborn RW,Vanerleyden J, Cammue BPA, Broekaert WF (1993): Syn-ergistic enhancement of the antifungal activity of wheatand barley thionins by radish and oilseed rape 2S albuminsand by barley trypsin inhibitors. Plant Physiol 103: 1311–1319.
Tong AHY, Lesage G, Bader GD, et al. (2004): Global mapping ofthe yeast genetic interaction network. Science 303: 808–813.
Uno J, Shigematsu ML, Arai T (1982): Primary site of actionof ketoconazole on Candida albicans. Antimicrob AgentsChemother 21: 912–918.
Valent B (2004): Underground life for rice foe. Nature 431: 516–517.
Xie MW, Jin F, Hwang H, Hwang S, Anand V, Duncan MC,Huang J (2005): Insights into TOR function and rapamycinresponse: Chemical genomic profiling by using high-densitycell array method. Proc Natl Acad Sci USA 192: 7215–7220.
Ziegelbauer K, Babczinski P, Schonfeld W (1998): Molecularmode of action of the antifungal β-amino acid BAY 10–8888.Antimicrob Agents Chemother 42: 2197–2205.





























