Embed Size (px)
Citation preview

Ecology and Evolution 201991985ndash2003 emsp|emsp1985wwwecolevolorg
Received25September2018emsp |emsp Revised5November2018emsp |emsp Accepted7December2018DOI101002ece34890
O R I G I N A L R E S E A R C H
Partitioning global change Assessing the relative importance of changes in climate and land cover for changes in avian distribution
Matthew J Clement12 emsp|emspJames D Nichols1 emsp|emspJaime A Collazo3emsp|emsp Adam J Terando4 emsp|emspJames E Hines1 emsp|emspSteven G Williams5
ThisisanopenaccessarticleunderthetermsoftheCreativeCommonsAttributionLicensewhichpermitsusedistributionandreproductioninanymediumprovidedtheoriginalworkisproperlycitedPublished2019ThisarticleisaUSGovernmentworkandisinthepublicdomainintheUSAEcology and Evolution
1UnitedStatesGeologicalSurveyPatuxentWildlifeResearchCenterLaurelMaryland2ArizonaGameandFishDepartmentResearchBranchPhoenixArizona3UnitedStatesGeologicalSurveyNorthCarolinaCooperativeFishandWildlifeResearchUnitDepartmentofAppliedEcologyNorthCarolinaStateUniversityRaleighNorthCarolina4SoutheastClimateAdaptationScienceCenterUSGeologicalSurveyRaleighNorthCarolina5NorthCarolinaCooperativeFishandWildlifeResearchUnitDepartmentofAppliedEcologyNorthCarolinaStateUniversityRaleighNorthCarolina
CorrespondenceMatthewJClementArizonaGameandFishDepartmentResearchBranchPhoenixAZEmailmclementgmailcom
Funding informationAssociationofFishandWildlifeAgenciesGrantAwardNumberNCM‐1‐HMUSGeologicalSurveyNorthCarolinaCooperativeFishandWildlifeResearchUnitResearchWorkOrder119
AbstractUnderstanding the relative impact of climate change and land cover change onchangesinaviandistributionhasimplicationsforthefuturecourseofaviandistribu‐tionsandappropriatemanagementstrategiesDuetothedynamicnatureofclimatechangeourgoalwastoinvestigatetheprocessesthatshapespeciesdistributionsratherthanthecurrentdistributionalpatternsTothisendweanalyzedchangesinthe distribution of EasternWood Pewees (Contopus virens) and Red‐eyed Vireos(Vireo olivaceus) from1997 to2012usingBreedingBirdSurveydataanddynamiccorrelated‐detectionoccupancymodelsWeestimatedthelocalcolonizationandex‐tinction ratesof thesespecies in relation tochanges inclimate (hoursofextremetemperature)andchangesinlandcover(amountofnestinghabitat)WefitsixnestedmodelstopartitionthedevianceexplainedbyspatialandtemporalcomponentsoflandcoverandclimateWeisolatedthetemporalcomponentsofenvironmentalvari‐ablesbecausethisistheessenceofglobalchangeForbothspeciesmodelfitwassignificantlyimprovedwhenwemodeledvitalratesasafunctionofspatialvariationinclimateandlandcoverModelfitimprovedonlymarginallywhenweaddedtempo‐ralvariation inclimateand landcover to themodelTemporalvariation inclimateexplainedmoredeviancethantemporalvariationinlandcoveralthoughbothcom‐binedonlyexplained20(EasternWoodPewee)and6(Red‐eyedVireo)oftempo‐ralvariationinvitalratesOurresultsshowingasignificantcorrelationbetweeninitialoccupancyandenvironmentalcovariatesareconsistentwithbiologicalexpectationandpreviousstudiesTheweakcorrelationbetweenvitalratesandtemporalchangesincovariatesindicatedthatwehaveyettoidentifythemostrelevantcomponentsofglobalchangeinfluencingthedistributionsofthesespeciesandmoreimportantlythatspatiallysignificantcovariatesarenotnecessarilydrivingtemporalshiftsinaviandistributions
K E Y W O R D S
dynamicoccupancymodelsEasternWoodPeweelocalcolonizationlocalextinctionNorthAmericanBreedingBirdSurveyRed‐eyedVireo
1986emsp |emsp emspensp CLEMENT ET aL
1emsp |emspINTRODUC TION
In recent decades the distributions of many bird species havechanged inBritain (Fuller et al 1995ThomasampLennon1999)continental Europe (Boumlhning‐Gaese amp Bauer 1996 BrommerLehikoinen amp Valkama 2012) South Africa (Hockey SiramiRidleyMidgleyampBabiker2011)andNorthAmerica(SauerLinkFallon Pardieck amp Ziolkowski 2013) These shifts are of bothecological and conservation interest and they have stimulateda great deal of researchWhile various factorsmay be contrib‐uting to distributional shifts including invasive species (Crowl CristParmenterBelovskyampLugo2008)pollution(Trathanetal2015) exploitation (Laursen amp Frikke 2008) and other causesclimatechangeandlandcoverchangesarelikelytobetwoofthestrongest drivers for many species (Barbet‐Massin Thuiller ampJiguet2012LemoineBauerPeintingerampBoumlhning‐Gaese2007Travis2003)Understandingtherelativeimpactofthesedriversof distributional change has implications for ecological under‐standingpredictionsoffuturechangesappropriatemanagementstrategiesandtheallocationofconservationresourcesHoweverspeciesdistributionmodelsfocusedontheeffectsofclimatearefarmorecommonthananalysesthatcomparethecontributionsofclimateandlandcover(Siramietal2017)
Ecologists have long debated the factors that affect speciesranges(Grinnell1917MacArthur1972)becauseoftherelevanceto ecology biogeography and community ecology In recent de‐cades global changehasgivenmoreurgency to the topic (Urban2015) and the expectation that both the global environment andspecies distributionswill continue to changehas stimulatedmuchresearchintopredictingthefuturecourseofchange(Huntleyetal2006)Questionsabout the relative impactof climatechangeandland cover change are important because of the implications forthesepredictions(Siramietal2017)Forexampleclimatechangeis expected tooccur alonga latitudinal gradientwhile changes inlandcovermayexhibitlessdirectionalityiffactorssuchasurbaniza‐tionorconversiontoorfromagriculturalusearesignificantdriversof landcoverchange (Lemoineetal2007Travis2003)Asare‐sultspeciesdistributionsthatrespondprimarilytoclimatearemorelikely to also exhibit directionality (Brommer et al 2012Hockeyetal2011ParmesanampYohe2003)Whetherspeciesdistributionsrespondtolandcoverorclimatechangehasimplicationsforappro‐priateresponsessuchasthedesignandlocationofreserves(ArauacutejoCabezaThuillerHannahampWilliams2004)habitatrestorationandassistedmigration (Hoegh‐Guldbergetal2008)Accordinglypo‐liticallyandeconomicallysignificantdecisionsmaybeinfluencedbytheseprojectionsTherefore there isaneed for research into therelativeeffectsofclimatechangeandlandcoverchangeonchangesinthedistributionofspecies
Whenthereisinterestinprojectingchangesinspeciesdistri‐butions a crucial insight is that environmental factors that cor‐relatewiththecurrentspeciesdistributionmaynotcorrelatewithfuture changes in distribution (Barbet‐Massin et al 2012) Onereason is that projections developed from current distribution
patterns assume that species are in equilibriumwith their envi‐ronment(ElithKearneyampPhillips2010)whichmaynotbetrue(ZhuWoodallampClark2012)A second reason is thatdispersallimitations may prevent species from occupying their preferredhabitat in the future (Devictor JulliardCouvetampJiguet2008)Therefore investigating theprocesses that shape species distri‐butionsratherthancurrentspeciesdistributionpatternsislikelytogenerategreaterecologicalunderstandingandbetterprojecteddistributions (YackulicNicholsReidampDer2015)ForexampletherecentdistributionofbreedingLouisianaWaterthrush(Parkesia motacilla) inNorthAmerica is best describedbymean tempera‐turebutchangesinthatdistributioncorrelatewithmeanprecip‐itation aswell asmean temperature indicating that theprocesscannotbefullydescribedbythepatternobservedatonepointintime(ClementHinesNicholsPardieckampZiolkowski2016)WefurthernotethatdataonenvironmentalvariablestypicallyincludebothspatialandtemporalcomponentsForexampletemperaturemayvaryalonganorthndashsouthgradient aswell as through timeWhentheprimaryresearchinterestisintemporalchangesindis‐tributions itmakessensetoisolatethetemporalcomponentsofenvironmentalvariables(whicharetheessenceofglobalchange)wheninvestigatingspeciesresponsetoglobalchange
HereweassesstherelativeimportanceofchangesinclimateandlandcovertochangesindistributionsfortwobirdspeciestheRed‐eyedVireo(Vireo olivaceus)andtheEasternWoodPewee(Contopus virens)Weselectedjusttwospecieswithsimilarbiologybutdiffer‐ingpopulationtrendssothatwecoulddevelopspecies‐specifichy‐pothesesaboutdistributionalshiftsWepartitionedthespatialandtemporal componentsofourenvironmentalpredictors to improveour inferencesabout theprocesses affecting speciesdistributionsandaboutthelikelyconsequencesoffutureglobalchangeWeuseddynamic occupancymodeling to explicitly estimate the vital ratesthatgovernchanges indistributionssoastoavoidtheequilibriumassumptionimplicitinstaticspeciesdistributionmodelsFinallyweusedananalysisofdevianceapproachtoassesstherelativeimpor‐tanceofclimateand landcovertothechanges indistributionsforthesetwospecies
2emsp |emspMATERIAL S AND METHODS
21emsp|emspHypotheses
Ourgoalwastoassesstherelativeimportanceofclimateandlandcovertochanges inthedistributionsoftwobirdspeciestheRed‐eyedVireoandtheEasternWoodPeweeWeselectedthesebirdsbecausetheyaremigratoryinsectivorousandinhabitsimilarplantcommunities(Hamel1992)butexhibitdivergenttrendsinrelativeabundancereportedbytheBBS(Saueretal2015)TheRed‐eyedVireobreedsintheeasternandnorthernUnitedStatesandmuchofsouthernCanadaandwintersinSouthAmericaItisacommonandwidespreadspeciesthatusesawidevarietyofforesthabitatstonestandgleaninsectsfromfoliageCountsofRed‐eyedVireoshavebeenincreasingby08annuallysince1966andby10annuallysince
emspensp emsp | emsp1987CLEMENT ET aL
2003withsomelocalizeddeclinesinFloridaTexasandthePacificNorthwest(Saueretal2015)TheEasternWoodPeweebreedsintheeasternUnitedStatesandpartsofsoutheasternCanadaIncon‐trasttoRed‐eyedVireoscountsoftheEasternWoodPeweehavebeendecliningby15since1966andby12since2003(Saueretal2015)DespitethegenerallynegativetrendtherehavebeenlocalizedincreasesincountsintheMidwest
Toestimatetherelativeimportanceofclimateandlandcovertochangesinbirddistributionswegeneratedtestablehypothesesex‐pressingrelationshipsbetweenbirdsandenvironmentalcovariatesAcommonapproachistogenerateanextensivesetofhypothesesfromasuiteofenvironmentalmetricsWecanthenfitthespecifiedmodelsandobservewhichmetricsyieldsignificantresultsHowevertestingmanystatisticalhypotheseshasapropensitytoidentifyspu‐riousrelationshipsandsuchanalysesshouldbeconsideredexplor‐atory(AndersonBurnhamGouldampCherry2001Ioannidis2005)Inthisstudywedevelopedtwoapriorihypothesesfromecologicalprinciples
1 Changes in the total annual periods of extreme temperaturedrive changes in bird distributionsWe focus on extreme tem‐perature because we expect these are the periods of greateststress due to cold or heat stress increased metabolic costsreduced resource availability or other mechanisms (Dawson ampWhittow 2000 Root 1988) We defined ldquoextremerdquo relativetothethermoneutralzone(TNZ)whichistherangeofambienttemperaturesunderwhichendothermscanmaintain theirbodytemperature without deviating from their basal metabolic rate(Calder amp King 1974) The TNZ varies among species but hasbeenestimatedas18ndash38degC fora ldquogenericrdquobird (CalderampKing1974) Several small temperate‐zone passerines (similar to ourstudy species) including the Northern Cardinal Verdin HouseSparrow and Red‐breasted Nuthatch have been documentedto have TNZs similar to that of this ldquogenericrdquo bird (Dawson1958 Goldstein 1974 Hinds amp Calder 1973 Khaliq HofPrinzinger Boumlhning‐Gaese amp Pfenninger 2014) Therefore weexpected temperatures below 18degC and above 38degC to beassociated with changes in bird distributions
2 Changes in the amount of appropriate habitat locally availableduringthebreedingseasondrivechangesinbirddistributionsbe‐cause inappropriatehabitatwillnotprovidesufficient foodandshelterforbreedingbirds(FriggensampFinch2015IgleciaCollazoampMcKerrow2012) For theEasternWoodPeweewedefinedappropriatehabitats asdeciduous forest evergreen forest andmixedforest(Hamel1992)FortheRed‐eyedVireowedefinedappropriate habitats as the same forest types and shrubndashscrub(Hamel1992)
Ofcoursethesehypothesesarenotexhaustivebutweselectedthem as logical general mechanisms that might underlie avian re‐sponsestoclimateandlandcoverchangeWeexpectedthepaucityand specificity of our hypotheseswould reduceour chancesof ob‐taining statistically significant resultsbychanceForexample anull
hypothesisthatbirddistributionswillnotexpandataspecifictempera‐tureishardertorejectthanthenullhypothesisthatbirddistributionswillnotexpandatanytemperatureWeviewamorespecifichypothe‐sisasamorepowerfuldiscriminatorytool(Platt1964Popper1959)moreinkeepingwiththehypothetico‐deductivemethod(Chamberlin1890Romesburg1981) andwith aType1error rate closer to thenominallevel(Andersonetal2001Ioannidis2005)
Westatedourhypothesizedeffectsintermsofchanges inbirddistribution(iebirdswillincreasewherehabitatisavailable)ratherthan static patterns of bird distribution (ie birds will be locatedwhere habitat is available)We prefer this formulation because itfocusesontheecologicalprocessesthatunderliespeciesdistribu‐tionsandtheirdynamics Inmetapopulationtheorythevitalratesdescribingtheseprocessesareoftenthelocalcolonizationandex‐tinction rates Models that account for ecological vital rates (iedynamicmodels)areusefulbecausetheyarerelativelymechanisticthereby strengthening the generality of findings and contributingmoretoecologicalunderstandingIncontrastinordertobeusefulforforecastingandprojectionrelationshipsbetweenenvironmentalcovariates andoccurrence require an assumption that the speciesinquestion is inequilibriumwith theenvironmental factorsunderstudy (Elith et al 2010) Themore phenomenological orientationofmodelsthatfocusonoccurrencecanbelimitingwhenprojectingmodelresultsintonovelsituationssuchasfutureclimateandlandcoverconditions(Cuddingtonetal2013Gustafson2013)Incon‐trast dynamicmodels avoid the equilibrium assumption enablingmore robust predictions of occurrence under future conditions(Clementetal2016Yackulicetal2015)
Bothclimateand landcovervary spatiallyand temporally andeachprocess could impactbirddistributions It is possible for thelevelofcorrelationwithbirdvital rates todifferacross those twodimensions For example consider an environmental variable thatiscompletelystaticthroughagiventimeperiodandaspeciesdistri‐butionthatshiftsduringthesametimeperiodInthiscaseitisnotpossibleforchangeinthestaticvariabletohavecausedthedistri‐butionalshiftbecausenosuchchangeoccurredHoweveritisstillpossibleforcolonizationandextinctiontocorrelatewiththeenvi‐ronmentalvariablebecauseofaspatialcorrelationbetweenthemHere our goalwas to incorporate temporal variation in covariatevalues intoouranalysisbecausemuchresearch intoglobalchangeismotivatedbyinterestintemporalchangesinspeciesdistributionsTherefore our hypotheses and models are structured to isolateandevaluatetheabilityofenvironmentalcovariatestoaccountforspatial and temporal variation in local colonization and extinctionprobabilities
22emsp|emspData
We obtained data on the detection and nondetection of breed‐ing Red‐eyed Vireos and Eastern Wood Pewees from the NorthAmerican Breeding Bird Survey (Pardieck Ziolkowski amp Hudson2015 wwwpwrcusgsgovBBSRawData) The Breeding BirdSurvey (BBS) is a joint project of the US Geological Survey and
1988emsp |emsp emspensp CLEMENT ET aL
theCanadianWildlifeServicedesignedtomonitor thestatusandtrendsofbirdsbreedinginNorthAmerica(Saueretal2013)Sinceitsinceptionin1966theBBShasexpandedtoincludeover5000survey routes across theUnited States and southernCanada ap‐proximately3000ofwhichare surveyedeachyearEach route issurveyedonceperyearbyaskilledvolunteerThedateofthesurveyistimedtooccurduringpeakterritorialbehaviortypicallylateMaytoearlyJulydependingon latitudeSurveyorsfollowaprescribedroutealongsecondaryroadsstoppingatapproximately800‐min‐tervalsandperforming3‐minroadsidepointcountsgenerating50countsper394kmrouteWeselectedBBSdatabecausetheirgreatgeographicand temporalextent is suitable for investigatingdistri‐butionalchanges
We selected a subsetof the availableBBSdata for use inouranalysis Our modeling approach described below makes use ofstop‐specificsurveyresultsbutthesedetailedresultsarecurrentlyavailableindigitalformatsonlysince1997Thereforeweuseddatafrom1997to2012BecauseourstudyspeciesarefoundinforestedregionsofeasternNorthAmericawedelineatedthestudyareabyaggregatingthreeOmernikLevelIclassesrepresentingeasternfor‐estecoregionsNorthernForestsEasternTemperateForestsandTropicalWetForests (CommissionforEnvironmentalCooperation1997Omernik19952004OmernikampGriffith2014)ThisstudyareaincludestheUnitedStateseastoftheGreatPlains(29millionkm2) This region encompassed the entire breeding range of theEasternWoodPeweeexceptsomeareasinCanadanotcoveredbyourclimateandhabitatcovariatesWeselectedBBSroutesentirelywithinthisstudyareaforanalysisWeexcludedroutesthatwerenotclassifiedasldquoactiverdquobytheBBSandroutesthatdifferedfromtherecommendedlength(394km)bymorethan25Weonlyanalyzedsurveys conducted under acceptable conditions (ie acceptableweatherdatetimeofdaystopsreportedbyBBSasldquoruntype=1rdquo)OntherareoccasionsthataroutehadmultipleacceptablesurveysperyearweuseddatafromthefirstacceptablesurveyThisyielded1371routesforouranalysisFinallyweconvertedcountsofbirdsateachstopalongeachBBSroutetodetectionnondetectiondatasuitableforpresencendashabsencemodelingThisconversionsacrificeddata richness but allowed us to account for imperfect detectionof species Using the full count data typically requires additionalassumptions (Illaacuten et al 2014) or incorporation of additional datasources(HootenWikleDorazioampRoyle2007)
ClimatedatawereobtainedfromthePRISMproject(Dalyetal2008)ThePRISMoutputuseslocallyweightedregressionmodelstointerpolatelong‐termweatherstationobservationstoa4kmresolu‐tiongridDailyinterpolatedclimatedataareavailablefrom1981tothepresentFromthesedatawecalculatedtwotemperature‐basedindices based on our first hypothesis described above The firstindexwascalculatedasthetotalannualhoursabove38degCwhilethesecondindexwascalculatedasthetotalannualhoursbelow18degC
Becausethetemperature‐basedindiceswerebasedonhoursofexposureitwasnecessarytointerpolatethedailyPRISMdatatoes‐timatehourlyvaluesWeconsideredasimplelinearinterpolationtoimputevaluesbetweendailymaximumandminimumtemperatures
However for extreme temperatures thismethodwould likely re‐sult in estimated exposure durations that are biased low InsteadweadoptedthemethoddescribedbyCesaraccioSpanoDuceandSnyder(2001seeFigure3)andEccel (2010)usingtheRsoftwarepackageldquoInterpolTrdquo (EccelampCordano2013) Inthismethodcan‐didatefunctionsarefit toobservedhourlytemperaturedatafromnearby high‐quality ASOS (Automated Station Observing System)weather stations in the study regionusing a combinationof sinu‐soidalandquadratic functionsForeach4kmPRISMgridcell theresultingcalibrationfunctionfromthenearestASOSstationisthenappliedtoeachdaysmaximumandminimumtemperaturetopredicttheinterveninghourlyvalues
Usingtheinterpolatedhourlytemperatureestimateswecalcu‐latedthetotalannualhourswithinthezonesofheatstressorcoldstress for each year from 1981 to 2012 Annual valueswere cal‐culatedover theperiodJune1ndashMay31 to represent thebreedingcycleOncetheannualresidencetimeswereobtainedweapplieda15‐yearmovingaveragewindowtothedataresultingin16mov‐ingaverageperiodsstartingwiththe198182ndash199596periodandendingwiththe199697ndash201112periodApplyingamovingaver‐ageactsasahigh‐passfilterthatattenuatesinterannualvariabilitywhileamplifyinganylong‐termclimatechangesignalsOtherperiodlengths could have been chosen however we hypothesized that15years represented a reasonable tradeoff between (a) reducingthemovingaveragelengthsoasnottoexceedthelengthofthedailyPRISMdataserieswhichbeginsin1981(b)increasingthemovingaveragelengthsothatclimaticchanges(asopposedtobackgroundclimaticvariation)canbeidentifiedand(c)usingaperiodthatisbio‐logicallymeaningfulinthatwecanreasonablyexpectanylong‐termclimaticchangesinthemovingaverageswouldbeoutsidethehis‐torical rangeofvariabilityand thusshouldaffectcolonizationandextinctionratesofsensitivespecies
We obtained land cover data from the National Land CoverDatabase (NLCD) for 2001 (Homer et al 2007) 2006 (Fry et al2011)and2011(Homeretal2015)WereclassifiedthegivenlandcovertypesintohabitatornonhabitatforeachbirdspeciesbasedonoursecondhypothesisdescribedaboveWecalculatedtheper‐centof landareathatwasasuitablehabitatwithin400mofeachrouteandusedthisasacovariate inouranalysisWeused400mbecausethisishalfthedistancebetweenstopsonBBSroutesForyearswithoutlandcoverdataweusedthelandcovervaluesofthesubsequent NLCD survey Specifically we used 2001 values for1997ndash20012006valuesfor2002ndash2006and2011valuesfor2007ndash2012Weusedaz‐transformationtocenterandscaleallcovariatespriortoanalysis
23emsp|emspAnalysis
OurgoalwastoassesstherelativeimportanceofclimateandlandcovertochangesinthedistributionofRed‐eyedVireosandEasternWoodPeweesWhileourbirddataconsistedofdetectionnonde‐tection data from BBS surveys our parameters of interest weretheprobabilitiesoflocalcolonizationandlocalextinctionasthese
emspensp emsp | emsp1989CLEMENT ET aL
processes drive changes in distribution Our predictor variableswereourmeasuresof climate and land cover suitability for eachspeciesdescribedaboveWeuseddynamic correlated‐detectionoccupancymodels with detection heterogeneity to estimate therelationshipbetweenourpredictorsandbirddistributiondynam‐ics (Clement et al 2016HinesNichols amp Collazo 2014Hinesetal2010)Occupancymodelsareaclassofmodelsthatestimatetheprobabilityofpresencewhileaccountingforimperfectdetec‐tionofspeciesbyanalyzingreplicated(usuallytemporally)surveys(MacKenzieetal20022018)Dynamicoccupancymodelsmodelinterannual changes in occupancy by positing aMarkov processinwhich the probability of occurrence at time t is a function ofthe probability of occurrence at time tminus1 (MacKenzie NicholsHinesKnutsonampFranklin2003)IncontrasttostaticmodelsthisMarkovprocess recognizes that thedistributionof a species is afunctionofpreviousenvironmentalconditionsanddispersalcon‐straintsaswellascurrentenvironmentalconditions(Dormannetal2012KeacuteryGuillera‐ArroitaampLahoz‐Monfort2013)Dynamiccorrelated‐detectionoccupancymodels areamodelextension inwhichspatiallycorrelatedreplicatedsurveysareusedtoestimatedetectionprobabilities (Hinesetal20142010)Weselectedanoccupancymodelingapproachbecause failure toaccount for im‐perfect detection can bias estimates of colonization and extinc‐tionrates(Ruiz‐GutieacuterrezampZipkin2011)Weselectedadynamicapproachbecausedynamicmodelsoffermoreecological realismmoreaccurateprojectionsandgreatergeneralizabilitythanstaticmodels (Clementetal2016Yackulicetal2015)Weusedcor‐related‐detection models because the BBS generates spatiallyreplicatedsurveysandfailuretoaccountforspatialcorrelationofreplicatescanbiasoccupancyestimates(Hinesetal20142010)Finallywe used a finitemixturemodel to account for detectionheterogeneity because of the variation in habitat and focal spe‐ciesabundanceacrossthestudyarea (Clementetal2016)Afi‐nitemixturemodel approximates the heterogeneity of detectionprobabilitiesbypositingthatthepopulationconsistsofamixtureofrouteswhichhaveeitherarelativelyhighdetectionprobabilityorarelativelylowdetectionprobability
AsdetailedaboveBBSdata consistofnumerous routes eachcomposedofbirdobservationsat50 stopsA speciesmaybeab‐sentfromindividualstopswhenitispresentonaroutebutnotviceversaDetectionof a species is taken asproofof presence but anondetectionmayresultfromatrueabsenceorafalseabsenceForclarityweusethetermldquooccupiedrdquotoindicateaspeciesispresentonarouteandthetermldquoavailablerdquotoindicateaspeciesislocallypres‐entataspecificstopatthetimeofthesurvey(NicholsThomasampConn2009)Bythisterminologythedynamiccorrelated‐detectionoccupancymodelincludesthefollowingparameters
ψ=Pr(routeoccupiedduringthefirstseasonofsurveys)θ = Pr (species available at stop | route occupied and species unavailable
at previous stop)θprime= Pr (species available at stop | route occupied and species available
at previous stop)
p1 = Pr (detection at a stop for low‐detection routes | route occupied and species available at stop)
p2 = Pr (detection at a stop for high‐detection routes | route occupied and species available at stop)
π = Pr (probability that a route is a low‐detection route)εt = Pr (route is not occupied in season t + 1 | occupied in season t) andγt = Pr (route is occupied in season t + 1 | not occupied in season t)
Hinesetal(20142010)developedamodellikelihoodthatallowstheseparameterstobemodeledasfunctionsofroute‐andyear‐spe‐cificcovariateswhiledetectionparameterscanalsobeinfluencedbystop‐specificcovariatesParameterscanthenbeestimatedusingmax‐imum‐likelihoodestimationinprogramPRESENCE(Hines2006)
WedevelopedsixspecificmodelsofγandεforeachbirdspeciesWepartitionedspatialandtemporalvariationincovariatesbyparti‐tioningcovariatesintotheaveragevalueovertimeandtheannualdeviationfromthataverageInthiswaytheaveragedcovariatexiincludedspatialvariationonlywhilethedeviationcomponentΔxiyincludedtemporalvariationonlyUsingthesecovariatesourmodelsincluded(a)anullmodelinwhichγandεareconstantthroughtimeandspaceand(b)aglobal(mostgeneral)modelinwhichγandεvarythroughspaceaccordingtomeanclimateandlandcoversuitabilityandthroughtimeusingadummycovariate foreachyearWealsoconsideredintermediatemodelstoassesstheexplanatorypowerofourcovariates(c)γandεvaryspatiallywithmeanclimateandlandcover suitability but notwith time (d) γ and ε vary spatiallywithmeanclimateandlandcoversuitabilityandtemporallywithannualchangesinclimateonly(e)γandεvaryspatiallywithmeanclimateand land cover suitability and temporally with annual changes inlandcoveronlyand(f)γandεvaryspatiallywithmeanclimateandlandcoversuitabilityandtemporallywithannualchangesinclimateandlandcover
Prior to comparing these models we worked to achieve ade‐quatefitfortheothermodelparametersBecauseofthenumberofmodelparametersweconsideredthemsequentiallyInitiallywefittheglobalmodelforγandεandwemodeledθθprimeandψasfunctionsofclimateandlandcovercovariatesInthisinitialmodelweusedafinitemixturemodelonptoaccountforheterogeneitypresumablycausedbydifferencesamongroutesinhabitatobserversbirdabun‐danceandotherfeaturesthatwerenotexplicitlymodeled(Clementetal2016)Wealsomodeledpasa functionofclimateand landcover covariates and year Because bird activity often varieswithtimeofdaywealsomodeleddetectionasaquadratic functionofstopnumberwhichweconsidertobeaproxyfortimeofdayFromthis highly parameterizedmodelwe fit a reducedmodel by elim‐inating the covariatewith the smallest estimate‐to‐standard errorratioWecontinuedtoeliminatecovariatesinthiswayuntilAICin‐creasedandthenselectedthemodelwiththelowestAICWeusedthisparameterreductionprotocolwithψθθprimeandpinturnmain‐tainingthemodelstructureonγandεAfteridentifyingparsimoni‐ousmodelsforψθθprimeandpwefitthesetofsixmodelsforγandεdescribedaboveThesequentialapproachmaynotyieldidenticalresultsasasingleanalysisbutitisapragmaticapproachfordealing
1990emsp |emsp emspensp CLEMENT ET aL
with complexmodels (MacKenzie et al 2018) By beginningwithhighlyparameterizedmodelsandprogressingtowardreducedmod‐elswereducedtheriskofconfoundingtheoccupancyanddetectionprocesses (MacKenzieet al2018)Wealsochecked forpotentialmulti‐collinearityissuesbycalculatingPearsonscorrelationcoeffi‐cientforthedifferentcovariatesusedinthemodelsWetreatedanR2lt05asindicativeofalackofmulti‐collinearity
WealsoevaluatedthegoodnessoffitoftheglobalmodelforeachspeciesusingthenaiumlvecolonizationandextinctionratestocalculatetheteststatisticofaHosmerndashLemeshowtest(HosmerampLemeshow2000) Inthiscontext thenaiumlvecolonizationrate is thenumberofsitesthatchangedfromnondetectionsitesinyeart todetectionsitesinyear t+1while thenaiumlveextinction rate is thenumberof sitesthatchangedfromdetectionsitesinyeart tonondetectionsitesinyeart+1Wedividedtheroutesintodecilesbasedontheexpectedcolonizationandextinctionratesandcomparedthepredictedratesto theobservedrateswithchi‐squaretests (α=005) Ifnecessarywecombineddecilestoensurethatthepredictednumberofdetec‐tionsinadecilewasgt4AlthoughtheHosmerndashLemeshowteststatis‐ticisoftencalculatedfrompredictedpresencevitalratesseemedtobemoreappropriatemeasuresinthiscasebecausedynamicmodelsestimatechangesinpresenceratherthanpresence(Clementetal2016)Wenotethatthisuseofnaiumlveratesfocusesonproductsofmodelparameters(ieγεp ψ ϑ)andthereforewedonottestthefitofthefullyparameterizedmodelWefocusedonnaiumlveratesbe‐causethetruevital rates (ieγandε)cannotbedirectlyobservedwhendetectionisimperfectwhilethenaiumlveratescanbeobserved
We used an analysis of deviance approach (ANODEV egSkalski 1996) to evaluate the relative importance of climate andland cover to changes in bird distributions In this procedure wecompared thedeviance explainedbyour parameter of interest tothedevianceexplainedbythemostfullyparameterizedmodelinthemodelset
Dev(Mnull)isthedevianceofModel3whichaccountsforspatialbutnottemporalvariationincolonizationandextinctionprobabili‐tiesDev(Mfull)isthedevianceofModel2theglobalmodelwithbothspatialandtemporalvariationinγandεWethenevaluatedthedevi‐anceexplainedbyclimateandorlandcoverbyusingmodels45and6 forDev(Mcov)HighervaluesofR
2
Dev indicategreaterexplanatory
powerwith0indicatingnopowerand1indicatingpowerequaltothatofthefullyparameterizedmodelalthoughR2
Devcouldpotentially
exceed1becauseourcovariatemodelsarenotstrictlynestedwithinModel2ThereforewecalculatedandreportR2
Dev forbothclimate
and landcoverThis isnot theonlypossibleapproach toassessingrelativeimportanceofexplanatoryvariablesbutitissoundandhasbeenusedandrecommendedbyothers(Grosboisetal2008)
The general ANODEV approach to partitioning variation ledustofocusonhypothesistestingasameansofassessingtheneedforextramodelparameters (sourcesofvariation)Thesmallsetof
specificapriorihypothesesalsosets thisworkapart fromexplor‐atorystudiesinvestigatingtheadequacyofmanydifferentmodelsForthesereasonsweusedlikelihoodratiotestsasameansofcom‐paringdifferentmodelsalthoughwenotethatuseofmodelselec‐tioncriteria(egAICc)yieldedsimilarinferences
3emsp |emspRESULTS
Ourcovariatesindicatedasmalllossofhabitatandincreasedtem‐peraturesduringthestudyperioddespitetherelativelyshorttimescaleForEasternWoodPeweehabitatchangewasgt1on38ofrouteswiththemeanshareofsuitablehabitatnearroutesde‐cliningfrom416to409ForRed‐eyedVireohabitatchangewasgt1on23of routeswithmeansuitablehabitatdecliningfrom480to476Routetemperatureswererarelyabovetheheatstress thresholdbut theaveragetimeabovethis thresholdper route increased from 05hr per year from 1982 to 1997 to17hrperyearfrom1997to2012Wealsoobservedadeclineinperiodsofcoldstressfrom5450to5291hrperrouteperyearThePearsons correlation coefficient did not indicate anymulti‐collinearityproblemsamongourcovariates(R2lt02inallcases)
31emsp|emspEastern Wood Pewee
ForEasternWoodPeweethebest‐supportedmodelincludedbothclimateandhabitatmeasuresas covariatesofψθ andθprimeWeex‐cludedclimateandhabitatcovariatesfromthedetectionprobabilitymodelbecausesomeofthosemodelsdidnotconvergeThereforethedetectionmodelforeachspeciesincludedyearandstopnum‐ber as covariates As a result the global model included 85 pa‐rameters (Table1)TheHosmerndashLemeshowtest indicatedthattheglobalmodel fit thedataadequatelywithnoevidenceofdiscrep‐ancies between estimated and observed naiumlve colonization rates(χ2=441 df =8 p=082) and naiumlve extinction rates (χ2=1117df =8p=019)
EasternWoodPeweewerewidespreadinthestudyareawithanaverageprobabilityofoccupancyonBBSroutesin1997of090intheglobalmodel(Figure1)Occupancydeclinedslightlyto088by2012mirroringthedeclineincountsreportedbytheBBSInitialoccupancywaspositivelyrelatedtoavailablehabitatandhoursofcoldstress(Table1)Probabilityofdetectionatthefirstindividualstopduring1997waslowat013whileprobabilityofdetectionat the route level was much higher at 095 because birds weretypically available atmany stopswithina routeModel3whichincludedspatialvariationinclimateandhabitatinthecolonizationandextinctionmodelssignificantlyimprovedmodelfitrelativetoModel1(likelihoodratioχ2=4419df =6plt0001)Underthismodelestimatedcolonizationratesin1998rangedamongroutesfrom 008 to 028 (with amean of 018) while extinction ratesvariedfrom001to004(withameanof002Figure2)Model2differedfromModel3inthat itallowedcolonizationandextinc‐tionratestovaryeachyearModel2improvedmodelfitrelative
R2
Dev=Dev(Mnull)minusDev(Mcov)
Dev(Mnull)minusDev(Mfull)
emspensp emsp | emsp1991CLEMENT ET aL
TA B L E 1 emspEstimatedparametersforcorrelated‐detectiondynamicoccupancyModel2relatinghabitatandclimatecovariatestooccupancydynamicsofbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012Parameterestimatesindicatechangeinlogoddsofparametersinresponsetoz‐transformedcovariatesParametersψϑϑprimeγεp1p2andπdefinedintext
Parameters
Eastern Wood Pewee Red‐eyed Vireo
Estimate SE Estimate SE
Ψintercept 2367 0177 4136 0404
Ψhabitat 0556 0163 2315 0300
Ψheatstress minus0011 0198 minus0337 0281
Ψcoldstress 0598 0166 1020 0258
ϑintercept minus1821 0026 minus1385 0010
ϑhabitat minus0055 0026 0516 0013
ϑheatstress minus0094 0030 0017 0008
ϑcoldstress 0172 0040 0258 0013
ϑprimeintercept 2292 0045 0426 0020
ϑprimehabitat 0316 0036 1210 0018
ϑprimeheatstress minus0006 0038 0054 0013
ϑprimecoldstress minus0622 0064 0876 0019
γintercept minus0884 0353 0010 0424
γ 1999 minus0601 0585 minus0150 0532
γ 2000 minus1083 0633 0516 0464
γ 2001 minus0300 0484 0228 0483
γ 2002 minus1113 0577 0262 0511
γ 2003 minus1055 0564 minus0311 0551
γ2004 minus0656 0525 minus0179 0534
γ 2005 minus0677 0533 0468 0485
γ2006 minus1014 0620 minus0217 0579
γ2007 minus0162 0453 minus0093 0548
γ 2008 minus23545 70483373 minus0478 0607
γ 2009 minus0428 0455 minus0253 0566
γ 2010 minus0292 0478 minus0159 0517
γ 2011 minus1136 0607 0012 0490
γ 2012 minus0498 0474 0141 0497
γaveragehabitat 0095 0091 0801 0112
γaverageheatstress minus0024 0067 0005 0062
γaveragecoldstress 0227 0071 0358 0120
εintercept minus3957 0386 minus3767 0242
ε 1999 0365 0482 minus0189 0337
ε 2000 0049 0512 minus0921 0399
ε 2001 minus0266 0548 minus0532 0362
ε 2002 minus0576 0660 minus1156 0398
ε 2003 minus0298 0595 minus0692 0355
ε2004 0113 0509 minus0621 0343
ε 2005 0173 0482 minus1096 0429
ε2006 minus0145 0561 minus1173 0463
ε2007 0451 0458 minus0894 0366
ε 2008 0283 0480 minus0976 0424
ε 2009 0218 0496 minus0898 0381
(Continues)
1992emsp |emsp emspensp CLEMENT ET aL
Parameters
Eastern Wood Pewee Red‐eyed Vireo
Estimate SE Estimate SE
ε 2010 minus0397 0605 minus0348 0304
ε 2011 0643 0455 minus0999 0406
ε 2012 0153 0644 minus0803 0410
εaveragehabitat 0238 0090 minus1670 0122
εaverageheatstress minus0071 0092 minus0075 0074
εaveragecoldstress minus0307 0141 minus0904 0104
p1intercept minus2172 0043 1352 0038
p1 1998 minus0118 0048 minus0127 0058
p1 1999 minus0112 0049 minus0043 0058
p1 2000 minus0166 0050 minus0016 0059
p1 2001 minus0156 0049 0031 0060
p1 2002 minus0179 0049 0076 0061
p1 2003 minus0216 0051 0161 0064
p12004 minus0242 0051 0066 0060
p1 2005 minus0124 0050 0149 0060
p12006 minus0182 0051 0124 0059
p12007 minus0149 0050 0184 0061
p1 2008 minus0202 0052 0192 0063
p1 2009 minus0221 0052 0135 0061
p1 2010 minus0195 0051 0062 0059
p1 2011 minus0122 0051 0203 0062
p1 2012 minus0227 0055 0194 0062
p1stopnumber 0033 0009 minus0055 0011
p1stopnumbersquared minus0069 0010 minus0247 0013
p2intercept minus0921 0018 minus0557 0036
p2 1998 minus0138 0033 minus0106 0050
p2 1999 minus0137 0032 0000 0052
p2 2000 minus0164 0032 minus0027 0051
p2 2001 minus0077 0032 minus0053 0053
p2 2002 minus0101 0032 minus0019 0052
p2 2003 minus0199 0033 0086 0053
p22004 minus0186 0032 0124 0057
p2 2005 minus0146 0032 0076 0051
p22006 minus0155 0032 0128 0054
p22007 minus0153 0032 0120 0054
p2 2008 minus0168 0032 0072 0056
p2 2009 minus0077 0033 0101 0055
p2 2010 minus0114 0033 0085 0060
p2 2011 minus0079 0032 0124 0054
p2 2012 minus0186 0033 0036 0061
p2stopnumber 0045 0007 0165 0010
p2stopnumbersquared minus0105 0008 minus0183 0010
π 0533 0064 minus0505 0065
TA B L E 1 emsp (Continued)
emspensp emsp | emsp1993CLEMENT ET aL
toModel3althoughnotsignificantlyduetothenumberofaddi‐tional parameters (χ2=2679df =28p=053)UnderModel 2estimatedcolonizationratesvariedfrom000in2008to030in1998whileextinctionratesvariedfrom001 in2001ndash2003and2010to004in2011WeusedModels45and6toestimatethepercentof improvedmodel fit (measuredby thechange indevi‐ance)generatedbyModel2thatcouldbeaccountedforbytempo‐ralvariationinclimateandhabitatcovariates(Table2)Temporalvariationinhabitataccountedfor04ofthechangeindeviancewhile temporal variation in climate accounted for 196Model6whichaccountedfortemporalvariationinbothhabitatandcli‐mateindicatedthatin1998colonizationinthesouthwestportionofthestudyareawasabovethelocalaveragewhileextinctioninthesouthwestwasbelowthelocalaverageindicatingarelativelystrongoccupancy trend in thesouthwest in thatyear (Figure3)By2012thispatternhadreversedyieldingarelativelyweakoc‐cupancytrendinthesouthwest(Figure3)NoneofthesemodelsrepresentedasignificantimprovementoverModel3(pgt026)Insomecasesthesignoftheestimatedeffectoftemporalvariationinclimateandhabitatwasoppositeoftheexpectedeffectbutinnoinstancesweretheseestimatessignificant(Table3)
32emsp|emspRed‐eyed Vireo
The best‐supportedmodel for Red‐eyedVireo included the samecovariatesasforEasternWoodPeweeyieldingaglobalmodelwith85parameters (Table1)TheHosmerndashLemeshowtest indicatedfitwasadequate for theRed‐eyedVireoglobalmodel forbothnaiumlvecolonization rates (χ2=540df =4p=025) and naiumlve extinctionrates(χ2=100df =4p=091)
Red‐eyedVireowere similarlywidespread in the study areawith an average probability of occupancy of 091 in the global
model (Figure1)Occupancydecreasedslightlyto090by2012incontrasttothe increase incountsreportedbytheBBS Initialoccupancywas positively related to available habitat and hoursofcoldstress (Table1)Probabilityofdetectionat the first stopwasrelativelyhighat038whileprobabilityofdetectionattheroute levelnearlyperfectat098Model3which includedspa‐tialvariationinclimateandhabitattothecolonizationandextinc‐tionmodelsimprovedmodelfitdramatically(χ2=41884df =6plt0001)relativetothenullmodelInkeepingwiththegreaterchangeindevianceforRed‐eyedVireoModel3estimatedmorespatialvariationinvitalratescomparedtoEasternWoodPeweeIn 1998 estimated colonization rates varied across BBS routesfrom 010 to 088 (mean of 049)while extinction rates rangedfrom000to048(meanof004Figure4)Model2whichallowedcolonization and extinction rates to vary each year improvedmodelfitrelativetoModel3butnotsignificantlyduetothenum‐berofadditionalparameters(χ2=3427df =28p=019)UnderModel2estimatedcolonizationratesvariedfrom039in2008to060 in2000while estimatedextinction rates varied from003in2006to007in1998WeusedModels45and6toestimatethepercentofthedeviancereductioninModel2thatcouldbeac‐countedforbytemporalvariationinclimateandhabitatcovariates(Table2)Wefoundthattemporalvariation inhabitataccountedfor01of the change indeviancewhile climate accounted for56Model 6 which accounted for temporal variation in bothhabitatandclimateindicatedthatin1998bothcolonizationandextinctionwereaboveaverageindicatinghigherturnoverearlyinthestudyperiod(Figure5)By2012bothcolonizationandextinc‐tion rateshaddeclined indicatinggreater stability inoccupancy(Figure 5) None of these models represented a significant im‐provementoverModel3(pgt074)Someoftheestimatedeffects
F I G U R E 1 emspOccupancyprobabilityfor(a)EasternWoodPeweeand(b)Red‐eyedVireointheeasternUnitedStates1997underModel2Occupancyvariesbytheamountofsuitablehabitathoursofheatstressandhoursofcoldstressin1997
1994emsp |emsp emspensp CLEMENT ET aL
oftemporalvariationinclimateandhabitatdifferedfromexpecta‐tionsbutnotsignificantly(Table3)
4emsp |emspDISCUSSION
Wefound that forbothbirdspecies initialoccupancycoloniza‐tionratesandextinctionratescorrelatedwithspatialvariationinclimateandhabitatcovariatesHowevercolonizationandextinc‐tionrateswerenotcorrelatedwithtemporalvariation inclimateand habitat covariates Our results correlating initial occupancywith environmental covariates are broadly speaking consistentwithbiologicalexpectationandaraftofpreviousstudiesthathaveindicatedasignificantrelationshipbetweenspeciesdistributionsandhabitat(RobinsonWilsonampCrick2001ThogmartinSauerampKnutson 2007) climate (Barbet‐Massinamp Jetz 2014BellardBertelsmeier Leadley Thuiller amp Courchamp 2012) or both(Seoane Bustamante amp Diacuteaz‐Delgado 2004 Sohl 2014)Howeverwesuggestthatestimatedrelationshipsbetweenspatialvariationinenvironmentalcovariatesandcolonizationandextinc‐tion rates are of greater ecological interest than relatively phe‐nomenological species distribution models or climate envelopemodels(Clementetal2016)Theestimatedvitalratesofcoloni‐zationandextinctionbringusclosertounderstandingthedynamicprocessunderlying thedistributionof these speciesGivencon‐stantvital rateswecanalsoproject theequilibrium levelofoc‐cupancyatsites ( i
i+i
atsite i)Wecancomparetheseprojected
equilibria tocurrentoccupancytoproject trends for thespeciesunder current environmental conditions We can also use
projectedchangesinkeyenvironmentalvariablestodeveloppre‐dicted rangechanges in the futureWeargue that theseprojec‐tions are more robust than those from static climate envelopemodels which assume that species are currently in equilibrium(Yackulicetal2015)Incontraststaticmodelscanoverestimaterange changes when the equilibrium does not hold (Morin ampThuiller2009)
Given the importanceofglobal changeweweremotivated toinvestigate factors affecting temporal aswell as spatial variationin vital rates In this case spatial variation must be incorporatedintosuchanalysesinordertoaccommodatethedifferentdynamicsexpectedindifferentportionsofaspeciesrangeWeviewthere‐lationshipbetweentemporalvariationincovariatevaluesandvitalrates as the most relevant to understanding the effect of globalchangeontherecentandfuturedistributionsofourstudyspeciesFurthermore a correlation between vital rates and spatially vary‐ingcovariatesmaynot indicateasimilarcorrelationwithtemporalchangesinthosesamecovariatesForexampleaspecieswithstrongdispersal constraintsmight have a strong spatial relationshipwithacovariatebutaweaktemporalrelationshipduetoits immobility(Devictor et al 2008) Similarly an estimated spatial relationshipmaybepurelyphenomenologicalduetothenatureofspatialvari‐ablesWhen spatial variables followgradients it is relatively easytoobtainstatisticallysignificantcorrelationsabsentanycausalre‐lationship(Grosboisetal2008)ForexampleifcolonizationratesandtemperaturesbothfollownorthndashsouthgradientstheymaybespatiallycorrelatedevenifcolonizationisgovernedbysomeotherunidentifiedfactorInthiscasewewouldexpecttheweakcorrela‐tion between temporal changes in temperature and colonization
F I G U R E 2 emspProbabilityof(a)colonizationand(b)extinctionforEasternWoodPeweeintheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1995CLEMENT ET aL
ratestobemoreindicativeofthetruerelationshipthanthestrongcorrelationwithspatialvariationintemperature
BBS surveys indicate different abundance trajectories for ourstudyspecieswithcounts increasingforRed‐eyedVireosandde‐creasing for EasternWood Pewees (Sauer et al 2015) Becauseabundanceandoccupancytendtobelinked(Gastonetal2000)weexpectedsomedivergenceinoccupancydynamicsWefoundthatestimatedoccupancywashighandstableforbothspeciesalthoughtherewasgreaterturnoverforRed‐eyedVireosForexampleunderModel3 (spatialvariationonly incovariates)bothaveragecoloni‐zationrates(049)andaverageextinctionrates(004)wereatleasttwiceashighforRed‐eyedVireoscomparedtoEasternWoodPewee(018and002respectively)Red‐eyedVireosexhibitedgreaterspa‐tialvariationincolonizationandextinctionrates(Figure4)althoughour selected covariates explained little temporal variation for thisspeciesEasternWoodPeweecolonizationandextinctionratesex‐hibitedmoreofalatitudinalgradientreflectingaspatialcorrelationwith extreme temperature (Figure 2)Our results also hinted at agreatercorrelationbetweentemporalchanges in temperatureandoccupancyvitalratesforEasternWoodPeweewhichcouldplayaroleinthedeclineofthisspeciesalthoughtheresultswerenotsig‐nificant(Table3)Alternatively ithasbeensuggestedthatEasternWoodPeweemaybedecliningduetoa lossofforestedwinteringhabitat(RobbinsSauerGreenbergampDroege1989)orduetodeer‐mediatedchanges in foreststructureonbreedinggrounds (deCal‐esta 1994) illustrating the complexity of ascertaining cause andeffectusingobservationalstudiesinthesenaturalsystems
Althoughrelativelyrare(Siramietal2017)otherstudieshaveinvestigated both climate and land cover effects on bird distribu‐tions Inagreementwithour finding that temporalvariation incli‐mate explainedmuchmore of themodel deviance than temporalvariation in habitat climate‐only species distribution models hadbetter cross‐validation support than habitat‐only models for 409Europeanbirdspecies(Barbet‐Massinetal2012)SimilarlymodelsestimatingthenumberofnewlyoccupiedareasbyHoodedWarblers(Wilsonia citrina) inOntariousingclimatedatahadbetter informa‐tioncriterionsupportthanmodelsusingforestdata(MellesFortinLindsay amp Badzinski 2011) In contrast land‐use models outper‐formedclimatemodels for18 farmlandbird species in theUnitedKingdom(EglingtonampPearce‐Higgins2012)Similarlyvegetation‐based species distributionmodels had superior performance thanclimate‐based models for 79 Spanish bird species (Seoane et al2004)andforthreebirdspeciesinNewMexico(FriggensampFinch2015) Two studies focused on colonization and extinction ratesfoundmixed results ForGardenWarblers (Sylvia borin) inBritainvegetationexplainedmorevariationincolonizationwhiletempera‐turesexplainedmorevariationinlocalextinction(MustinAmarampRedpath2014)For122birdspeciesinOntariothebest‐supportedcovariates (land cover or climate change) varied among species(YalcinampLeroux2018)
Thedivergentresultsamongstudiesregardingtheimportanceofclimateandlandcovermaypartlybeduetothebiologyofindividualspecies (Yalcinamp Leroux 2018) For example farmland birdsmayTA
BLE
2emspR2 DevfordifferentmodelsofcolonizationandextinctionprobabilitiesforbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012
Mod
el
Cova
riate
s on
colo
niza
tion
and
extin
ctio
nEa
ster
n W
ood
Pew
eeRe
d‐ey
ed V
ireo
Inte
rcep
tAv
erag
e H
abita
tAv
erag
e Cl
imat
eH
abita
t Dev
iatio
nCl
imat
e D
evia
tion
Year
Eff
ect
Δ minus
2LL
R2 Dev
Δ minus
2LL
R2 Dev
2X
XX
X0
0010
00
0010
0
6X
XX
XX
minus2138
202
minus3230
57
4X
XX
Xminus2154
196
minus3234
56
5X
XX
Xminus2667
04
minus3422
01
3X
XX
minus2679
0minus3427
0
1X
minus7098
NA
minus45311
NA
Not
eNAnotapplicable
1996emsp |emsp emspensp CLEMENT ET aL
beparticularlyaffectedbychangesinagriculturalintensitybecauseoftheirextensiveuseoffarms(EglingtonampPearce‐Higgins2012)Alternativelyithasbeenarguedthatspatialscalemayplayaroleintherelativeimportanceofglobalchangefactorswithclimatemoreimportantatlargescales(PearsonampDawson2003)Wenotethattwostudiesthatfoundalargerroleforclimatebothreliedonlower‐resolution bird surveys (05deg atlas in Barbet‐Massin et al 2012
10km2atlasinMellesetal2011)IntheBBSdatathatweanalyzedsurveyswereconductedevery800mbutwecalculatedoccupancyacross each 394km transect In addition the cited studies useddifferentdata sources anddiverse analysismethods complicatinginterpretationofthedifferingresults
While we estimated that climate explained more model de‐viance than habitat these estimated relationships were not
F I G U R E 3 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionrates((t)minus (t)minus )forEasternWoodPeweeintheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
emspensp emsp | emsp1997CLEMENT ET aL
TA B L E 3 emspEstimatedparametersforcorrelated‐detectiondynamicoccupancyModel6relatinghabitatandclimatecovariatestooccupancydynamicsofbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012Parameterestimatesindicatechangeinlogoddsofparametersinresponsetoz‐transformedcovariatesParametersψϑϑprimeγε p1p2andπdefinedintextForkeyparameters(boldtext)expectedsignsincluded
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
Ψintercept 2367 0177 4087 0381
Ψhabitat 0556 0163 2323 0288
Ψheatstress minus0011 0198 minus0322 0275
Ψcoldstress 0598 0166 1054 0251
ϑintercept minus1821 0026 minus1385 0010
ϑhabitat minus0055 0026 0516 0013
ϑheatstress minus0094 0030 0017 0008
ϑcoldstress 0172 0040 0256 0013
ϑprimeintercept 2292 0045 0426 0020
ϑprimehabitat 0316 0036 1210 0018
ϑprimeheatstress minus0006 0038 0054 0013
ϑprimecoldstress minus0622 0064 0875 0019
γintercept minus0884 0353 0023 0162
γhabitatdeviation + minus0292 0478 minus0240 1768
γheatstressdeviation minus minus1136 0607 0064 0194
γcoldstressdeviation minus minus0498 0474 2398 2253
γaveragehabitat 0095 0091 0788 0111
γaverageheatstress minus0024 0067 0004 0062
γaveragecoldstress 0227 0071 0333 0119
εintercept minus3957 0386 minus4471 0141
εhabitatdeviation minus minus0397 0605 0121 1492
εheatstressdeviation + 0643 0455 minus0034 0182
εcoldstressdeviation + 0153 0644 2042 2072
εaveragehabitat 0238 0090 minus1664 0123
εaverageheatstress minus0071 0092 minus0080 0074
εaveragecoldstress minus0307 0141 minus0939 0104
p1intercept minus2172 0043 1351 0039
p1 1998 minus0118 0048 minus0128 0058
p1 1999 minus0112 0049 minus0043 0058
p1 2000 minus0166 0050 minus0017 0059
p1 2001 minus0156 0049 0031 0061
p1 2002 minus0179 0049 0076 0062
p1 2003 minus0216 0051 0160 0064
p12004 minus0242 0051 0066 0061
p1 2005 minus0124 0050 0149 0061
p12006 minus0182 0051 0124 0060
p12007 minus0149 0050 0183 0062
p1 2008 minus0202 0052 0191 0064
p1 2009 minus0221 0052 0134 0062
p1 2010 minus0195 0051 0062 0060
p1 2011 minus0122 0051 0202 0063
(Continues)
1998emsp |emsp emspensp CLEMENT ET aL
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
p1 2012 minus0227 0055 0193 0063
p1stopnumber 0033 0009 minus0055 0011
p1stopnumbersquared minus0069 0010 minus0247 0013
p2intercept minus0921 0018 minus0555 0037
p2 1998 minus0138 0033 minus0121 0050
p2 1999 minus0137 0032 minus0025 0052
p2 2000 minus0164 0032 minus0027 0051
p2 2001 minus0077 0032 minus0059 0053
p2 2002 minus0101 0032 minus0012 0052
p2 2003 minus0199 0033 0085 0053
p22004 minus0186 0032 0113 0059
p2 2005 minus0146 0032 0082 0052
p22006 minus0155 0032 0135 0054
p22007 minus0153 0032 0124 0055
p2 2008 minus0168 0032 0073 0057
p2 2009 minus0077 0033 0100 0056
p2 2010 minus0114 0033 0071 0062
p2 2011 minus0079 0032 0120 0055
p2 2012 minus0186 0033 0030 0063
p2stopnumber 0045 0007 0165 0010
p2stopnumbersquared minus0105 0008 minus0182 0011
π 0533 0064 minus0505 0065
TA B L E 3 emsp (Continued)
F I G U R E 4 emspProbabilityof(a)colonizationand(b)extinctionforRed‐eyedVireointheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1999CLEMENT ET aL
significantanddidnotexplainalargefractionofvariationThereareseveralpotentialexplanationsforwhywedidnotfindasub‐stantialresponsetochangesinhabitatandclimateFirstourdatawerelackingindynamiceventstoinformourmodelsThecovari‐ateschangedrelativelylittleoverthetimeperiodmakingitdiffi‐culttodetectaresponseThelackofchangewaspartlybecauseoftherelativelyshort(16‐year)timeperiodstudiedAlsoweusedamovingaverageforclimatedatawhichtendstoreducedifferences
overshorttimeperiods Inadditionthestudyregionhasexperi‐encedlesswarmingoverthepastcenturythanotherpartsoftheplanet (Walshetal2014)Thebirddistributionsalso lackeddy‐namismBecauseextinctionrateswerelowtherewerefewlocalextinctioneventstoinformthemodelAlthoughcolonizationrateswerehighthefactthatinitialoccupancywashighandextinctionrateswerelowmeanttherewerefewopportunitiesforcoloniza‐tionevents
F I G U R E 5 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionratesforRed‐eyedVireointheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
2000emsp |emsp emspensp CLEMENT ET aL
Second measurement error may have obscured responses toglobalchangeThePRISMclimatedataareestimatedbyinterpola‐tionratherthandirectmeasurementacrossthestudyarea(Dalyetal2008)AssuchtheremaybeartifactsoranomaliesthatimpededourestimationandincreaseduncertaintySimilarlyNLCDdataareremotelysensedandmay includesomemeasurementerrorwhichcan increase uncertainty in our estimates (Homer et al 2015)Additionallylandcoverwasonlymeasuredinthreeyearsrequiringus to imputevalues in13yearsAsa result all landcover changethatoccurredbetween2002and2006wasrecordedin2002whilechangesbetween2007and2011wererecorded in2007Thisap‐proximation isnot idealbutwasnecessarygiventhe5‐yearreso‐lutionofthedataThislowerresolutionofthelandcoverdatamayhavereducedthesensitivityofthemodel
Third wemay havemisspecified ourmodelWe tried to de‐velopbiologicallymeaningfulcovariatesbutwemightnotunder‐standthesystemaswellaswehopedItispossiblethatbirdvitalrates respond to climateon adifferent time scale (ie not a15‐yearmoving average) or to climateduringdifferent timesof theyearortodifferenttemperaturethresholdsortodifferentclimatecomponents (egprecipitation Illaacutenetal2014) It isalsopossi‐ble that thebirds respond to finerhabitat features than the landcovertypesweusedoratadifferentspatialscale(ienot400mPearsonampDawson2003)Alternatively theremaybe importantregionaldifferencesor interactionsamongcovariatesthatwedidnotcaptureOritispossiblethatchangesinclimateandhabitatintheirbreedingareassimplyhavelittleeffectonvitalratesforthesespecies relative to other factors such as competitionwith otherbirdspeciesorglobalchangeoccurringontheirwinteringgroundsinSouthAmerica(Dormann2007)Ofcoursethesemisspecifica‐tionissuesareinherenttoallobservationalstudiesandnotuniquetothisstudy
Itwould be possible for us to explore some of themisspec‐ification issues just mentioned For example we could rerunthe samemodelsusingnewcovariates representing a varietyoftemperature thresholds and habitat buffer sizes to search for astrongercorrelationExperiencesuggeststhatpersistencewouldeventuallyberewardedwithap‐valuelt005ora lowmodelse‐lection valueHowever one reason to avoid such exploration isthat it tends to lead toType Ierrorsoverfitmodels andbiasedparameterestimates(FiebergampJohnson2015)Furthermorewefind thecontrastbetweensignificant correlations for spatial co‐variatesandnonsignificantcorrelationsfortemporalcovariatesil‐luminatingIntypicalclimateenvelopestudieswewouldestimatethe relationshipbetweencovariatesand thecurrentdistributionofaspeciesmuchlikeourestimateofinitialoccupancy(Dormann2007)BecauseweobtainedsignificantparameterestimateswemightconcludethatwehaveidentifiedcovariatesthatdeterminetherangeofthisspeciesThenwemightusetheseresultstoproj‐ectthefuturerangeofthespeciesundervariousclimatescenariosbased on the (questionable) assumption that the covariatendashspe‐cies relationship will not change as climate and habitat change(Cuddington et al 2013 Elith et al 2010Gustafson 2013) In
this case the insignificant correlationwith the temporal covari‐atessuggeststhatouroccupancycovariatesmerelydescribetherangeofthesebirdswithoutdeterminingtherangeFurthermoreour results suggest that despite significant results in the staticportionof themodelwehaveyet to identify themost relevantcomponents of climate change shaping thedistributionof thesebirds although itmay be difficult to identify these componentsgiven the lackofobservedchangeduringa relatively short timeperiodFinallyourresultssuggestthatprojectionsbasedonthestaticportionofourmodelareunlikelytobereliabledespitetheirstatisticalsignificance
Ifourinterpretationiscorrectitsuggeststhatsomeprojectionsof futurerangeshiftsbasedonstaticmodelsrelyonunsupportedassumptionsandarenot reliableWehavedemonstratedadiffer‐entapproachthatdoesnotrequirethestrongassumptionsofstaticmodelingapproachesOurapproachwouldbemoreapttoidentifyfactorsthatguidespeciesrsquoresponsetoglobalchangeifboththespe‐ciesandthecovariatesexhibitedmoredynamismandifweanalyzedalongertimeperiodthatencompassedgreaterchange
ACKNOWLEDG MENTS
WethankKeithPardieckDaveZiolkowskiJrandthethousandsofhighlyskilledvolunteerswhohavecoordinatedandperformedtheNorthAmericanBreedingBirdSurvey for half a centuryWealsothanktheClimateResearchUnitattheUniversityofEastAngliaformakingvaluableclimatedataeasilyaccessibleMJCwasfinanciallysupported by the BBS and the USGS Northeast Climate ScienceCenterThecontentsofthisdocumentaretheresponsibilityoftheauthorsanddonotnecessarilyrepresenttheviewsoftheUSGSAnyuseoftradeproductorfirmnamesisfordescriptivepurposesonlyanddoesnotimplyendorsementbytheUSGovernment
DEDIC ATION
We dedicate this paper to the late Chandler S Robbins ldquoFatherrdquooftheNorthAmericanBreedingBirdSurvey (BBS)DatafromtheBBSformthebasisforthisspecificpaperandforhundredsofothersfocusedon topics ranging from local trends tocontinentalmacro‐ecologyChanwasauniquecombinationofbird‐loverandscientistwhosevisionandinquisitiveenthusiasmwereaninspirationtoallofusfortunateenoughtohaveknownhim
CONFLIC T OF INTERE S T
Nonedeclared
AUTHORSrsquo CONTRIBUTION
JACandJDNconceivedthestudyAJTandSWanalyzedhabitatandclimatedataMJCJEHandJDNperformedtheoccupancyanaly‐sisMJCledthewritingofthemanuscriptAllauthorsgavefinalap‐provalforpublication
emspensp emsp | emsp2001CLEMENT ET aL
DATA ACCE SSIBILIT Y
Raw data are available from the following sources Breeding BirdSurvey httpswwwpwrcusgsgovbbsrawdata PRISM climatedata httpwwwprismoregonstateedu NLCD land cover datahttpswwwmrlcgov Processed data will be archived with theDryadDigitalRepository
ORCID
Matthew J Clement httpsorcidorg0000‐0003‐4231‐7949
James D Nichols httpsorcidorg0000‐0002‐7631‐2890
Adam J Terando httpsorcidorg0000‐0002‐9280‐043X
James E Hines httpsorcidorg0000‐0001‐5478‐7230
Steven G Williams httpsorcidorg0000‐0003‐3760‐6818
R E FE R E N C E S
Anderson D R Burnham K P Gould W R amp Cherry S (2001)Concerns about finding effects that are actually spuriousWildlife Society Bulletin29311ndash316
Arauacutejo M B Cabeza M Thuiller W Hannah L amp WilliamsP H (2004) Would climate change drive species out of re‐serves An assessment of existing reserve‐selection meth‐ods Global Change Biology 10 1618ndash1626 httpsdoiorg101111j1365‐2486200400828x
Barbet‐MassinMampJetzW(2014)A40‐yearcontinent‐widemulti‐speciesassessmentofrelevantclimatepredictorsforspeciesdistri‐butionmodellingDiversity and Distributions201285ndash1295httpsdoiorg101111ddi12229
Barbet‐Massin M Thuiller W amp Jiguet F (2012) The fate ofEuropean breeding birds under climate land‐use and disper‐sal scenarios Global Change Biology 18 881ndash890 httpsdoiorg101111j1365‐2486201102552x
Bellard C Bertelsmeier C Leadley P Thuiller W ampCourchamp F (2012) Impacts of climate change on the fu‐ture of biodiversity Ecology Letters 15 365ndash377 httpsdoiorg101111j1461‐0248201101736x
Boumlhning‐Gaese K amp Bauer H G (1996) Changes in species abun‐dance distribution and diversity in a central European birdcommunity Conservation Biology 10 175ndash187 httpsdoiorg101046j1523‐1739199610010175x
BrommerJELehikoinenAampValkamaJ(2012)Thebreedingrangesof central European and Arctic bird speciesmove poleward PLoS ONE7e43648httpsdoiorg101371journalpone0043648
CalderWAampKingJR(1974)ThermalandcaloricrelationsofbirdsInDSFarnerampJRKing(Eds)Avian biology(Vol4pp259ndash413)NewYorkNYAcademicPress
CesaraccioCSpanoDDucePampSnyderRL(2001)Animprovedmodel for determining degree‐day values from daily temperaturedataInternational Journal of Biometeorology45161ndash169httpsdoiorg101007s004840100104
ChamberlinTC (1890)ThemethodofmultipleworkinghypothesesScience1592ndash96
ClementMJHinesJENicholsJDPardieckKLampZiolkowskiDJJr (2016)Estimating indicesofrangeshifts inbirdsusingdy‐namicmodelswhendetectionisimperfectGlobal Change Biology223273ndash3285httpsdoiorg101111gcb13283
Commission for Environmental Cooperation (1997) Ecological regionsofNorthAmericaTowardacommonperspectiveCommission for
EnvironmentalCooperationMontrealRetrievedfromftpftpepagovwedecoregionscec_naCEC_NAecopdf
Crowl TA Crist TO Parmenter R R BelovskyGamp LugoA E(2008)Thespreadofinvasivespeciesandinfectiousdiseaseasdriv‐ersofecosystemchangeFrontiers in Ecology and the Environment6238ndash246httpsdoiorg101890070151
Cuddington K FortinM J Gerber L RHastings A Liebhold AOConnorMampRayC(2013)Process‐basedmodelsarerequiredtomanageecologicalsystemsinachangingworldEcosphere41ndash12httpsdoiorg101890ES12‐001781
DalyCHalbleibMSmithJIGibsonWPDoggettMKTaylorGHhellipPasterisPP(2008)Physiographicallysensitivemappingofcli‐matologicaltemperatureandprecipitationacrosstheconterminousUnited States International Journal of Climatology 28 2031ndash2064httpsdoiorg101002joc1688
DawsonW (1958) Relation of oxygen consumption and evaporativewater losstotemperatureintheCardinalPhysiological Zoology3137ndash48httpsdoiorg101086physzool31130155377
DawsonWRampWhittowGC (2000)Regulationofbodytempera‐ture InG CWhittow (Ed)Sturkiersquos avian physiology (5th ed pp343ndash390)SanDiegoCAAcademicPress
DeCalestaDS(1994)Effectofwhite‐taileddeeronsongbirdswithinmanagedforestsinPennsylvaniaJournal of Wildlife Management58711ndash718httpsdoiorg1023073809685
DevictorVJulliardRCouvetDampJiguetF(2008)Birdsaretrack‐ing climatewarming but not fast enoughProceedings of the Royal Society of London B Biological Sciences2752743ndash2748httpsdoiorg101098rspb20080878
Dormann C F (2007) Promising the future Global change projec‐tionsofspeciesdistributionsBasic and Applied Ecology8387ndash397httpsdoiorg101016jbaae200611001
Dormann C F Schymanski S J Cabral J Chuine I Graham CHartigFhellipSingerA(2012)Correlationandprocessinspeciesdis‐tributionmodelsBridgingadichotomyJournal of Biogeography392119ndash2131httpsdoiorg101111j1365‐2699201102659x
EccelE(2010)WhatwecanasktohourlytemperaturerecordingPartIIHourlyinterpolationoftemperaturesforclimatologyandmodel‐lingItalian Journal of Agrometeorology1545ndash50
EccelEampCordanoE(2013)InterpolTHourlyinterpolationofmulti‐pletemperaturedailyseriesRpackageversion211RetrievedfromhttpsCRANR‐projectorgpackage=InterpolT
EglingtonSMampPearce‐HigginsJW(2012)Disentanglingtherela‐tiveimportanceofchangesinclimateandland‐useintensityindriv‐ingrecentbirdpopulationtrendsPLoS ONE7e30407httpsdoiorg101371journalpone0030407
Elith JKearneyMampPhillipsS (2010)Theartofmodelling range‐shifting species Methods in Ecology and Evolution 1 330ndash342httpsdoiorg101111j2041‐210X201000036x
FiebergJampJohnsonDH(2015)MMIMultimodelinferenceormod‐elswithmanagement implications Journal of Wildlife Management79708ndash718
FriggensMMampFinchDM (2015) Implicationsof climatechangeforbirdconservationinthesouthwesternUSunderthreealternativefuturesPLoS ONE10e0144089
FryJXianGZJinSDewitzJHomerCGYangLhellipWickhamJD (2011)Completionof the2006national landcoverdatabasefor theconterminousUnitedStatesPhotogrammetric Engineering amp Remote Sensing77858ndash864
Fuller R J Gregory R D Gibbons D W Marchant J HWilson J D Baillie S R amp Carter N (1995) Population de‐clines and range contractions among lowland farmland birdsin Britain Conservation Biology 9 1425ndash1441 httpsdoiorg101046j1523‐1739199509061425x
Gaston K J Blackburn T M Greenwood J J Gregory R DQuinn R M amp Lawton J H (2000) Abundancendashoccupancy
2002emsp |emsp emspensp CLEMENT ET aL
relationships Journal of Applied Ecology 37 39ndash59 httpsdoiorg101046j1365‐2664200000485x
GoldsteinRB(1974)RelationofmetabolismtoambienttemperatureintheverdinCondor76116ndash119httpsdoiorg1023071365995
GrinnellJ(1917)FieldtestsoftheoriesconcerningdistributionalcontrolAmerican Naturalist51115ndash128httpsdoiorg101086279591
GrosboisVGimenezOGaillardJMPradelRBarbraudCClobertJhellipWeimerskirchH(2008)Assessingtheimpactofclimatevari‐ation on survival in vertebrate populationsBiological Reviews 83357ndash399httpsdoiorg101111j1469‐185X200800047x
GustafsonEJ(2013)WhenrelationshipsestimatedinthepastcannotbeusedtopredictthefutureUsingmechanisticmodelstopredictlandscapeecologicaldynamicsinachangingworldLandscape Ecology281429ndash1437httpsdoiorg101007s10980‐013‐9927‐4
Hamel P B (1992) LandManagers guide to the birds of the SouthGeneralTechnicalReportSE‐22USDAForestServiceSoutheasternForestExperimentStationandTheNatureConservancyChapelHillNC
Hinds D S amp Calder W A (1973) Temperature regulation of thePyrrhuloxiaandtheArizonaCardinalPhysiological Zoology4655ndash71httpsdoiorg101086physzool46130152517
HinesJE(2006)PRESENCE Software to estimate patch occupancy and related parameters Laurel MD US Geological survey PatuxentWildlife Research Center Retrieved from httpwwwmbr‐pwrcusgsgovsoftwarepresencehtml
HinesJENicholsJDampCollazoJA(2014)Multiseasonoccupancymodels for correlated replicate surveys Methods in Ecology and Evolution5583ndash591httpsdoiorg1011112041‐210X12186
HinesJENicholsJDRoyleJAMacKenzieDIGopalaswamyAMKumarNampKaranthKU(2010)TigersontrailsOccupancymodeling for cluster sampling Ecological Applications 20 1456ndash1466httpsdoiorg10189009‐03211
HockeyPA SiramiCRidleyARMidgleyGFampBabikerHA(2011) Interrogating recent range changes in South African birdsConfounding signals from land use and climate change present achallenge for attribution Diversity and Distributions 17 254ndash261httpsdoiorg101111j1472‐4642201000741x
Hoegh‐Guldberg O Hughes L McIntyre S Lindenmayer D BParmesanCPossinghamHPampThomasCD (2008)AssistedcolonizationandrapidclimatechangeScience321345ndash346
Homer C Dewitz J Fry J Coan M Hossain N Larson C hellipWickham J (2007) Completion of the 2001 national land coverdatabase for the conterminous United States Photogrammetric Engineering amp Remote Sensing73337
HomerCDewitzJYangLJinSDanielsonPXianGhellipMegownK(2015)Completionofthe2011NationalLandCoverDatabasefortheconterminousUnitedStatesndashrepresentingadecadeoflandcoverchange informationPhotogrammetric Engineering amp Remote Sensing81345ndash354
HootenMBWikleCKDorazioRMampRoyleJA(2007)HierarchicalspatiotemporalmatrixmodelsforcharacterizinginvasionsBiometrics63558ndash567httpsdoiorg101111j1541‐0420200600725x
HosmerDWJrampLemeshowS(2000)Applied logistic regression(2nded)NewYorkNYJohnWileyampSons
Huntley B Collingham Y CGreen R EHiltonGM RahbekCamp Willis S G (2006) Potential impacts of climatic change upongeographical distributions of birds Ibis 148 8ndash28 httpsdoiorg101111j1474‐919X200600523x
IgleciaMNCollazoJAampMcKerrowAJ(2012)Useofoccupancymodels to evaluate expert knowledge‐based species‐habitat rela‐tionshipsAvian Conservation and Ecology7(2)13pRetrievedfromhttpwwwace‐ecoorgvol7iss2art5
IllaacutenJGThomasCDJonesJAWongWKShirleySMampBettsMG(2014)Precipitationandwintertemperaturepredictlong‐termrange‐scale abundancechanges inWesternNorthAmericanbirds
Global Change Biology 20 3351ndash3364 httpsdoiorg101111gcb12642
IoannidisJP (2005)WhymostpublishedresearchfindingsarefalsePLoS Medicine2e124
KeacuteryMGuillera‐ArroitaGampLahoz‐Monfort J J (2013)Analysingand mapping species range dynamics using occupancy modelsJournal of Biogeography 40 1463ndash1474 httpsdoiorg101111jbi12087
Khaliq I Hof C Prinzinger R Boumlhning‐Gaese K amp PfenningerM(2014) Global variation in thermal tolerances and vulnerability ofendotherms to climate change Proceedings of the Royal Society B Biological Sciences 281(1789) 20141097 httpsdoiorg101098rspb20141097
Laursen K amp Frikke J (2008) Hunting from motorboats displacesWadden Sea eiders Somateria mollissima from their favouredfeeding distribution Wildlife Biology 14 423ndash433 httpsdoiorg1029810909‐6396‐144423
LemoineNBauerHGPeintingerMampBoumlhning‐GaeseK (2007)Effectsof climate and land‐use changeon species abundance in acentralEuropeanbirdcommunityConservation Biology21495ndash503httpsdoiorg101111j1523‐1739200600633x
MacArthurRH(1972)Geographical ecology Patterns in the distribution of speciesNewYorkNYHarperampRow
MacKenzieDINicholsJDHinesJEKnutsonMGampFranklinAB(2003)Estimatingsiteoccupancycolonizationandlocalextinc‐tionwhenaspeciesisdetectedimperfectlyEcology842200ndash2207httpsdoiorg10189002‐3090
MacKenzieDINicholsJDLachmanGBDroegeSRoyleJAampLangtimmCA(2002)Estimatingsiteoccupancyrateswhendetec‐tionprobabilitiesarelessthanoneEcology832248ndash2255httpsdoiorg1018900012‐9658(2002)083[2248ESORWD]20CO2
MacKenzieDINicholsJDRoyleJAPollockKHBaileyLampHinesJE(2018)Occupancy estimation and modeling Inferring pat‐terns and dynamics of species occurrence (2nd ed) BurlingtonMAAcademicPress
MellesSJFortinMJLindsayKampBadzinskiD(2011)ExpandingnorthwardInfluenceofclimatechangeforestconnectivityandpop‐ulationprocessesonathreatenedspeciesrangeshiftGlobal Change Biology1717ndash31httpsdoiorg101111j1365‐2486201002214x
Morin X amp Thuiller W (2009) Comparing niche‐and process‐based models to reduce prediction uncertainty in species rangeshifts under climate change Ecology 90 1301ndash1313 httpsdoiorg10189008‐01341
MustinKAmarAampRedpathSM(2014)Colonizationandextinctiondynamicsofadecliningmigratorybirdareinfluencedbyclimateandhabitat degradation Ibis 156 788ndash798 httpsdoiorg101111ibi12173
NicholsJDThomasLampConnPB(2009)InferencesaboutlandbirdabundancefromcountdataRecentadvancesandfuturedirectionsInDLThomsonEJCoochampMJConroy(Eds)Modeling demo‐graphic processes in marked populations(pp201ndash235)NewYorkNYSpringer
OmernikJM(1995)EcoregionsAspatialframeworkforenvironmen‐talmanagement InWSDavisampTSimon(Eds)Biological assess‐ment and criteria Tools for water resource planning and decision making (pp49ndash62)BocaRatonFLLewisPublishers
OmernikJM(2004)Perspectivesonthenatureanddefinitionofeco‐logicalregionsEnvironmental Management34(Supplement1)S27ndashS38httpsdoiorg101007s00267‐003‐5197‐2
Omernik J M amp Griffith G E (2014) Ecoregions of the contermi‐nousUnited States Evolution of a hierarchical spatial frameworkEnvironmental Management541249ndash1266httpsdoiorg101007s00267‐014‐0364‐1
Pardieck K L ZiolkowskiD J JrampHudsonM‐A R (2015)NorthAmericanBreedingBirdSurveyDataset1966ndash2014version20140
emspensp emsp | emsp2003CLEMENT ET aL
USGeologicalSurveyPatuxentWildlifeResearchCenterRetrievedfromhttpswwwpwrcusgsgovbbsrawdata
ParmesanCampYoheG (2003)Agloballycoherentfingerprintofcli‐mate change impacts across natural systemsNature 421 37ndash42httpsdoiorg101038nature01286
PearsonRGampDawsonTP(2003)PredictingtheimpactsofclimatechangeonthedistributionofspeciesArebioclimateenvelopemod‐elsusefulGlobal Ecology and Biogeography12361ndash371
PlattJR(1964)StronginferenceScience146347ndash353PopperK(1959)The logic of scientific discoveryLondonUKHutchinsonRobbins C S Sauer J R Greenberg R S amp Droege S (1989)
Population declines in North American birds that migrate to theNeotropics Proceedings of the National Academy of Sciences 867658ndash7662httpsdoiorg101073pnas86197658
Robinson R A Wilson J D amp Crick H Q (2001) The impor‐tance of arable habitat for farmland birds in grassland land‐scapes Journal of Applied Ecology 38 1059ndash1069 httpsdoiorg101046j1365‐2664200100654x
Romesburg H C (1981) Wildlife science Gaining reliable knowl‐edge Journal of Wildlife Management 45 293ndash313 httpsdoiorg1023073807913
Root T (1988) Energy constraints on avian distributions and abun‐dancesEcology69330ndash339httpsdoiorg1023071940431
Ruiz‐GutieacuterrezVampZipkinEF (2011)Detectionbiasesyieldmis‐leadingpatternsof speciespersistence and colonization in frag‐mented landscapesEcosphere2 1ndash14 httpsdoiorg101890ES10‐002071
SauerJRHinesJEFallonJEPardieckKLZiolkowskiDJJrampLinkWA(2015)The North American breeding bird survey results and analysis 1966ndash2013 Version 01302015LaurelMDUSGSPatuxentWildlifeResearchCenterhttpswwwmbr‐pwrcusgsgovbbsbbshtml
Sauer J R LinkWA Fallon J E Pardieck K L amp ZiolkowskiDJJr(2013)TheNorthAmericanbreedingbirdsurvey1966ndash2011SummaryanalysisandspeciesaccountsNorth American Fauna791ndash32httpsdoiorg103996nafa790001
SeoaneJBustamanteJampDiacuteaz‐DelgadoR(2004)Competingrolesforlandscapevegetationtopographyandclimateinpredictivemod‐elsofbirddistributionEcological Modelling171209ndash222httpsdoiorg101016jecolmodel200308006
Sirami C Caplat P Popy S Clamens A Arlettaz R Jiguet F hellipMartin J‐L (2017) Impacts of global change on species distri‐butions Obstacles and solutions to integrate climate and landuse Global Ecology and Biogeography 26 385ndash394 httpsdoiorg101111geb12555
SkalskiJR(1996)Regressionofabundanceestimatesfrommarkrecap‐ture surveys against environmental covariatesCanadian Journal of
Fisheries and Aquatic Sciences53196ndash204httpsdoiorg101139f95‐169
SohlTL(2014)Therelativeimpactsofclimateandland‐usechangeonconterminousUnitedStatesbird species from2001 to2075PLoS ONE9e112251httpsdoiorg101371journalpone0112251
ThogmartinWESauerJRampKnutsonMG(2007)ModelingandmappingabundanceofAmericanwoodcockacrosstheMidwesternandNortheasternUnitedStatesJournal of Wildlife Management71376ndash382httpsdoiorg1021932005‐680
ThomasCDampLennonJJ (1999)Birdsextendtheirrangesnorth‐wardsNature399213httpsdoiorg10103820335
Trathan P N Garciacutea‐Borboroglu P Boersma D Bost C ACrawford R J CrossinG ThellipWienecke B (2015) Pollutionhabitatlossfishingandclimatechangeascriticalthreatstopen‐guins Conservation Biology 29 31ndash41 httpsdoiorg101111cobi12349
TravisJMJ(2003)Climatechangeandhabitatdestructionadeadlyanthropogenic cocktail Proceedings of the Royal Society of London Series B Biological Sciences 270(1514) 467ndash473 httpsdoiorg101098rspb20022246
UrbanMC (2015)Acceleratingextinction risk fromclimatechangeScience348571ndash573httpsdoiorg101126scienceaaa4984
WalshJWuebblesDHayhoeKKossinJKunkelKStephensGhellipSomervilleR(2014)OurchangingclimateInJMMelilloTCRichmondampGWYohe(Eds)Climate change impacts in the United States The Third National climate assessment(pp19ndash67)WashingtonDCUSGovernmentPrintingOffice
YackulicCBNichols JDReid JampDerR (2015)Topredict thenichemodelcolonizationandextinctionEcology9616ndash23httpsdoiorg10189014‐13611
Yalcin S amp Leroux S J (2018) An empirical test of the relative andcombinedeffectsofland‐coverandclimatechangeonlocalcoloni‐zationandextinctionGlobal Change Biology243849ndash3861httpsdoiorg101111gcb14169
ZhuKWoodallCWampClarkJS(2012)FailuretomigrateLackoftreerangeexpansioninresponsetoclimatechangeGlobal Change Biology181042ndash1052httpsdoiorg101111j1365‐2486201102571x
How to cite this articleClementMJNicholsJDCollazoJATerandoAJHinesJEWilliamsSGPartitioningglobalchangeAssessingtherelativeimportanceofchangesinclimateandlandcoverforchangesinaviandistributionEcol Evol 201991985ndash2003 httpsdoiorg101002ece34890

1986emsp |emsp emspensp CLEMENT ET aL
1emsp |emspINTRODUC TION
In recent decades the distributions of many bird species havechanged inBritain (Fuller et al 1995ThomasampLennon1999)continental Europe (Boumlhning‐Gaese amp Bauer 1996 BrommerLehikoinen amp Valkama 2012) South Africa (Hockey SiramiRidleyMidgleyampBabiker2011)andNorthAmerica(SauerLinkFallon Pardieck amp Ziolkowski 2013) These shifts are of bothecological and conservation interest and they have stimulateda great deal of researchWhile various factorsmay be contrib‐uting to distributional shifts including invasive species (Crowl CristParmenterBelovskyampLugo2008)pollution(Trathanetal2015) exploitation (Laursen amp Frikke 2008) and other causesclimatechangeandlandcoverchangesarelikelytobetwoofthestrongest drivers for many species (Barbet‐Massin Thuiller ampJiguet2012LemoineBauerPeintingerampBoumlhning‐Gaese2007Travis2003)Understandingtherelativeimpactofthesedriversof distributional change has implications for ecological under‐standingpredictionsoffuturechangesappropriatemanagementstrategiesandtheallocationofconservationresourcesHoweverspeciesdistributionmodelsfocusedontheeffectsofclimatearefarmorecommonthananalysesthatcomparethecontributionsofclimateandlandcover(Siramietal2017)
Ecologists have long debated the factors that affect speciesranges(Grinnell1917MacArthur1972)becauseoftherelevanceto ecology biogeography and community ecology In recent de‐cades global changehasgivenmoreurgency to the topic (Urban2015) and the expectation that both the global environment andspecies distributionswill continue to changehas stimulatedmuchresearchintopredictingthefuturecourseofchange(Huntleyetal2006)Questionsabout the relative impactof climatechangeandland cover change are important because of the implications forthesepredictions(Siramietal2017)Forexampleclimatechangeis expected tooccur alonga latitudinal gradientwhile changes inlandcovermayexhibitlessdirectionalityiffactorssuchasurbaniza‐tionorconversiontoorfromagriculturalusearesignificantdriversof landcoverchange (Lemoineetal2007Travis2003)Asare‐sultspeciesdistributionsthatrespondprimarilytoclimatearemorelikely to also exhibit directionality (Brommer et al 2012Hockeyetal2011ParmesanampYohe2003)Whetherspeciesdistributionsrespondtolandcoverorclimatechangehasimplicationsforappro‐priateresponsessuchasthedesignandlocationofreserves(ArauacutejoCabezaThuillerHannahampWilliams2004)habitatrestorationandassistedmigration (Hoegh‐Guldbergetal2008)Accordinglypo‐liticallyandeconomicallysignificantdecisionsmaybeinfluencedbytheseprojectionsTherefore there isaneed for research into therelativeeffectsofclimatechangeandlandcoverchangeonchangesinthedistributionofspecies
Whenthereisinterestinprojectingchangesinspeciesdistri‐butions a crucial insight is that environmental factors that cor‐relatewiththecurrentspeciesdistributionmaynotcorrelatewithfuture changes in distribution (Barbet‐Massin et al 2012) Onereason is that projections developed from current distribution
patterns assume that species are in equilibriumwith their envi‐ronment(ElithKearneyampPhillips2010)whichmaynotbetrue(ZhuWoodallampClark2012)A second reason is thatdispersallimitations may prevent species from occupying their preferredhabitat in the future (Devictor JulliardCouvetampJiguet2008)Therefore investigating theprocesses that shape species distri‐butionsratherthancurrentspeciesdistributionpatternsislikelytogenerategreaterecologicalunderstandingandbetterprojecteddistributions (YackulicNicholsReidampDer2015)ForexampletherecentdistributionofbreedingLouisianaWaterthrush(Parkesia motacilla) inNorthAmerica is best describedbymean tempera‐turebutchangesinthatdistributioncorrelatewithmeanprecip‐itation aswell asmean temperature indicating that theprocesscannotbefullydescribedbythepatternobservedatonepointintime(ClementHinesNicholsPardieckampZiolkowski2016)WefurthernotethatdataonenvironmentalvariablestypicallyincludebothspatialandtemporalcomponentsForexampletemperaturemayvaryalonganorthndashsouthgradient aswell as through timeWhentheprimaryresearchinterestisintemporalchangesindis‐tributions itmakessensetoisolatethetemporalcomponentsofenvironmentalvariables(whicharetheessenceofglobalchange)wheninvestigatingspeciesresponsetoglobalchange
HereweassesstherelativeimportanceofchangesinclimateandlandcovertochangesindistributionsfortwobirdspeciestheRed‐eyedVireo(Vireo olivaceus)andtheEasternWoodPewee(Contopus virens)Weselectedjusttwospecieswithsimilarbiologybutdiffer‐ingpopulationtrendssothatwecoulddevelopspecies‐specifichy‐pothesesaboutdistributionalshiftsWepartitionedthespatialandtemporal componentsofourenvironmentalpredictors to improveour inferencesabout theprocesses affecting speciesdistributionsandaboutthelikelyconsequencesoffutureglobalchangeWeuseddynamic occupancymodeling to explicitly estimate the vital ratesthatgovernchanges indistributionssoastoavoidtheequilibriumassumptionimplicitinstaticspeciesdistributionmodelsFinallyweusedananalysisofdevianceapproachtoassesstherelativeimpor‐tanceofclimateand landcovertothechanges indistributionsforthesetwospecies
2emsp |emspMATERIAL S AND METHODS
21emsp|emspHypotheses
Ourgoalwastoassesstherelativeimportanceofclimateandlandcovertochanges inthedistributionsoftwobirdspeciestheRed‐eyedVireoandtheEasternWoodPeweeWeselectedthesebirdsbecausetheyaremigratoryinsectivorousandinhabitsimilarplantcommunities(Hamel1992)butexhibitdivergenttrendsinrelativeabundancereportedbytheBBS(Saueretal2015)TheRed‐eyedVireobreedsintheeasternandnorthernUnitedStatesandmuchofsouthernCanadaandwintersinSouthAmericaItisacommonandwidespreadspeciesthatusesawidevarietyofforesthabitatstonestandgleaninsectsfromfoliageCountsofRed‐eyedVireoshavebeenincreasingby08annuallysince1966andby10annuallysince
emspensp emsp | emsp1987CLEMENT ET aL
2003withsomelocalizeddeclinesinFloridaTexasandthePacificNorthwest(Saueretal2015)TheEasternWoodPeweebreedsintheeasternUnitedStatesandpartsofsoutheasternCanadaIncon‐trasttoRed‐eyedVireoscountsoftheEasternWoodPeweehavebeendecliningby15since1966andby12since2003(Saueretal2015)DespitethegenerallynegativetrendtherehavebeenlocalizedincreasesincountsintheMidwest
Toestimatetherelativeimportanceofclimateandlandcovertochangesinbirddistributionswegeneratedtestablehypothesesex‐pressingrelationshipsbetweenbirdsandenvironmentalcovariatesAcommonapproachistogenerateanextensivesetofhypothesesfromasuiteofenvironmentalmetricsWecanthenfitthespecifiedmodelsandobservewhichmetricsyieldsignificantresultsHowevertestingmanystatisticalhypotheseshasapropensitytoidentifyspu‐riousrelationshipsandsuchanalysesshouldbeconsideredexplor‐atory(AndersonBurnhamGouldampCherry2001Ioannidis2005)Inthisstudywedevelopedtwoapriorihypothesesfromecologicalprinciples
1 Changes in the total annual periods of extreme temperaturedrive changes in bird distributionsWe focus on extreme tem‐perature because we expect these are the periods of greateststress due to cold or heat stress increased metabolic costsreduced resource availability or other mechanisms (Dawson ampWhittow 2000 Root 1988) We defined ldquoextremerdquo relativetothethermoneutralzone(TNZ)whichistherangeofambienttemperaturesunderwhichendothermscanmaintain theirbodytemperature without deviating from their basal metabolic rate(Calder amp King 1974) The TNZ varies among species but hasbeenestimatedas18ndash38degC fora ldquogenericrdquobird (CalderampKing1974) Several small temperate‐zone passerines (similar to ourstudy species) including the Northern Cardinal Verdin HouseSparrow and Red‐breasted Nuthatch have been documentedto have TNZs similar to that of this ldquogenericrdquo bird (Dawson1958 Goldstein 1974 Hinds amp Calder 1973 Khaliq HofPrinzinger Boumlhning‐Gaese amp Pfenninger 2014) Therefore weexpected temperatures below 18degC and above 38degC to beassociated with changes in bird distributions
2 Changes in the amount of appropriate habitat locally availableduringthebreedingseasondrivechangesinbirddistributionsbe‐cause inappropriatehabitatwillnotprovidesufficient foodandshelterforbreedingbirds(FriggensampFinch2015IgleciaCollazoampMcKerrow2012) For theEasternWoodPeweewedefinedappropriatehabitats asdeciduous forest evergreen forest andmixedforest(Hamel1992)FortheRed‐eyedVireowedefinedappropriate habitats as the same forest types and shrubndashscrub(Hamel1992)
Ofcoursethesehypothesesarenotexhaustivebutweselectedthem as logical general mechanisms that might underlie avian re‐sponsestoclimateandlandcoverchangeWeexpectedthepaucityand specificity of our hypotheseswould reduceour chancesof ob‐taining statistically significant resultsbychanceForexample anull
hypothesisthatbirddistributionswillnotexpandataspecifictempera‐tureishardertorejectthanthenullhypothesisthatbirddistributionswillnotexpandatanytemperatureWeviewamorespecifichypothe‐sisasamorepowerfuldiscriminatorytool(Platt1964Popper1959)moreinkeepingwiththehypothetico‐deductivemethod(Chamberlin1890Romesburg1981) andwith aType1error rate closer to thenominallevel(Andersonetal2001Ioannidis2005)
Westatedourhypothesizedeffectsintermsofchanges inbirddistribution(iebirdswillincreasewherehabitatisavailable)ratherthan static patterns of bird distribution (ie birds will be locatedwhere habitat is available)We prefer this formulation because itfocusesontheecologicalprocessesthatunderliespeciesdistribu‐tionsandtheirdynamics Inmetapopulationtheorythevitalratesdescribingtheseprocessesareoftenthelocalcolonizationandex‐tinction rates Models that account for ecological vital rates (iedynamicmodels)areusefulbecausetheyarerelativelymechanisticthereby strengthening the generality of findings and contributingmoretoecologicalunderstandingIncontrastinordertobeusefulforforecastingandprojectionrelationshipsbetweenenvironmentalcovariates andoccurrence require an assumption that the speciesinquestion is inequilibriumwith theenvironmental factorsunderstudy (Elith et al 2010) Themore phenomenological orientationofmodelsthatfocusonoccurrencecanbelimitingwhenprojectingmodelresultsintonovelsituationssuchasfutureclimateandlandcoverconditions(Cuddingtonetal2013Gustafson2013)Incon‐trast dynamicmodels avoid the equilibrium assumption enablingmore robust predictions of occurrence under future conditions(Clementetal2016Yackulicetal2015)
Bothclimateand landcovervary spatiallyand temporally andeachprocess could impactbirddistributions It is possible for thelevelofcorrelationwithbirdvital rates todifferacross those twodimensions For example consider an environmental variable thatiscompletelystaticthroughagiventimeperiodandaspeciesdistri‐butionthatshiftsduringthesametimeperiodInthiscaseitisnotpossibleforchangeinthestaticvariabletohavecausedthedistri‐butionalshiftbecausenosuchchangeoccurredHoweveritisstillpossibleforcolonizationandextinctiontocorrelatewiththeenvi‐ronmentalvariablebecauseofaspatialcorrelationbetweenthemHere our goalwas to incorporate temporal variation in covariatevalues intoouranalysisbecausemuchresearch intoglobalchangeismotivatedbyinterestintemporalchangesinspeciesdistributionsTherefore our hypotheses and models are structured to isolateandevaluatetheabilityofenvironmentalcovariatestoaccountforspatial and temporal variation in local colonization and extinctionprobabilities
22emsp|emspData
We obtained data on the detection and nondetection of breed‐ing Red‐eyed Vireos and Eastern Wood Pewees from the NorthAmerican Breeding Bird Survey (Pardieck Ziolkowski amp Hudson2015 wwwpwrcusgsgovBBSRawData) The Breeding BirdSurvey (BBS) is a joint project of the US Geological Survey and
1988emsp |emsp emspensp CLEMENT ET aL
theCanadianWildlifeServicedesignedtomonitor thestatusandtrendsofbirdsbreedinginNorthAmerica(Saueretal2013)Sinceitsinceptionin1966theBBShasexpandedtoincludeover5000survey routes across theUnited States and southernCanada ap‐proximately3000ofwhichare surveyedeachyearEach route issurveyedonceperyearbyaskilledvolunteerThedateofthesurveyistimedtooccurduringpeakterritorialbehaviortypicallylateMaytoearlyJulydependingon latitudeSurveyorsfollowaprescribedroutealongsecondaryroadsstoppingatapproximately800‐min‐tervalsandperforming3‐minroadsidepointcountsgenerating50countsper394kmrouteWeselectedBBSdatabecausetheirgreatgeographicand temporalextent is suitable for investigatingdistri‐butionalchanges
We selected a subsetof the availableBBSdata for use inouranalysis Our modeling approach described below makes use ofstop‐specificsurveyresultsbutthesedetailedresultsarecurrentlyavailableindigitalformatsonlysince1997Thereforeweuseddatafrom1997to2012BecauseourstudyspeciesarefoundinforestedregionsofeasternNorthAmericawedelineatedthestudyareabyaggregatingthreeOmernikLevelIclassesrepresentingeasternfor‐estecoregionsNorthernForestsEasternTemperateForestsandTropicalWetForests (CommissionforEnvironmentalCooperation1997Omernik19952004OmernikampGriffith2014)ThisstudyareaincludestheUnitedStateseastoftheGreatPlains(29millionkm2) This region encompassed the entire breeding range of theEasternWoodPeweeexceptsomeareasinCanadanotcoveredbyourclimateandhabitatcovariatesWeselectedBBSroutesentirelywithinthisstudyareaforanalysisWeexcludedroutesthatwerenotclassifiedasldquoactiverdquobytheBBSandroutesthatdifferedfromtherecommendedlength(394km)bymorethan25Weonlyanalyzedsurveys conducted under acceptable conditions (ie acceptableweatherdatetimeofdaystopsreportedbyBBSasldquoruntype=1rdquo)OntherareoccasionsthataroutehadmultipleacceptablesurveysperyearweuseddatafromthefirstacceptablesurveyThisyielded1371routesforouranalysisFinallyweconvertedcountsofbirdsateachstopalongeachBBSroutetodetectionnondetectiondatasuitableforpresencendashabsencemodelingThisconversionsacrificeddata richness but allowed us to account for imperfect detectionof species Using the full count data typically requires additionalassumptions (Illaacuten et al 2014) or incorporation of additional datasources(HootenWikleDorazioampRoyle2007)
ClimatedatawereobtainedfromthePRISMproject(Dalyetal2008)ThePRISMoutputuseslocallyweightedregressionmodelstointerpolatelong‐termweatherstationobservationstoa4kmresolu‐tiongridDailyinterpolatedclimatedataareavailablefrom1981tothepresentFromthesedatawecalculatedtwotemperature‐basedindices based on our first hypothesis described above The firstindexwascalculatedasthetotalannualhoursabove38degCwhilethesecondindexwascalculatedasthetotalannualhoursbelow18degC
Becausethetemperature‐basedindiceswerebasedonhoursofexposureitwasnecessarytointerpolatethedailyPRISMdatatoes‐timatehourlyvaluesWeconsideredasimplelinearinterpolationtoimputevaluesbetweendailymaximumandminimumtemperatures
However for extreme temperatures thismethodwould likely re‐sult in estimated exposure durations that are biased low InsteadweadoptedthemethoddescribedbyCesaraccioSpanoDuceandSnyder(2001seeFigure3)andEccel (2010)usingtheRsoftwarepackageldquoInterpolTrdquo (EccelampCordano2013) Inthismethodcan‐didatefunctionsarefit toobservedhourlytemperaturedatafromnearby high‐quality ASOS (Automated Station Observing System)weather stations in the study regionusing a combinationof sinu‐soidalandquadratic functionsForeach4kmPRISMgridcell theresultingcalibrationfunctionfromthenearestASOSstationisthenappliedtoeachdaysmaximumandminimumtemperaturetopredicttheinterveninghourlyvalues
Usingtheinterpolatedhourlytemperatureestimateswecalcu‐latedthetotalannualhourswithinthezonesofheatstressorcoldstress for each year from 1981 to 2012 Annual valueswere cal‐culatedover theperiodJune1ndashMay31 to represent thebreedingcycleOncetheannualresidencetimeswereobtainedweapplieda15‐yearmovingaveragewindowtothedataresultingin16mov‐ingaverageperiodsstartingwiththe198182ndash199596periodandendingwiththe199697ndash201112periodApplyingamovingaver‐ageactsasahigh‐passfilterthatattenuatesinterannualvariabilitywhileamplifyinganylong‐termclimatechangesignalsOtherperiodlengths could have been chosen however we hypothesized that15years represented a reasonable tradeoff between (a) reducingthemovingaveragelengthsoasnottoexceedthelengthofthedailyPRISMdataserieswhichbeginsin1981(b)increasingthemovingaveragelengthsothatclimaticchanges(asopposedtobackgroundclimaticvariation)canbeidentifiedand(c)usingaperiodthatisbio‐logicallymeaningfulinthatwecanreasonablyexpectanylong‐termclimaticchangesinthemovingaverageswouldbeoutsidethehis‐torical rangeofvariabilityand thusshouldaffectcolonizationandextinctionratesofsensitivespecies
We obtained land cover data from the National Land CoverDatabase (NLCD) for 2001 (Homer et al 2007) 2006 (Fry et al2011)and2011(Homeretal2015)WereclassifiedthegivenlandcovertypesintohabitatornonhabitatforeachbirdspeciesbasedonoursecondhypothesisdescribedaboveWecalculatedtheper‐centof landareathatwasasuitablehabitatwithin400mofeachrouteandusedthisasacovariate inouranalysisWeused400mbecausethisishalfthedistancebetweenstopsonBBSroutesForyearswithoutlandcoverdataweusedthelandcovervaluesofthesubsequent NLCD survey Specifically we used 2001 values for1997ndash20012006valuesfor2002ndash2006and2011valuesfor2007ndash2012Weusedaz‐transformationtocenterandscaleallcovariatespriortoanalysis
23emsp|emspAnalysis
OurgoalwastoassesstherelativeimportanceofclimateandlandcovertochangesinthedistributionofRed‐eyedVireosandEasternWoodPeweesWhileourbirddataconsistedofdetectionnonde‐tection data from BBS surveys our parameters of interest weretheprobabilitiesoflocalcolonizationandlocalextinctionasthese
emspensp emsp | emsp1989CLEMENT ET aL
processes drive changes in distribution Our predictor variableswereourmeasuresof climate and land cover suitability for eachspeciesdescribedaboveWeuseddynamic correlated‐detectionoccupancymodels with detection heterogeneity to estimate therelationshipbetweenourpredictorsandbirddistributiondynam‐ics (Clement et al 2016HinesNichols amp Collazo 2014Hinesetal2010)Occupancymodelsareaclassofmodelsthatestimatetheprobabilityofpresencewhileaccountingforimperfectdetec‐tionofspeciesbyanalyzingreplicated(usuallytemporally)surveys(MacKenzieetal20022018)Dynamicoccupancymodelsmodelinterannual changes in occupancy by positing aMarkov processinwhich the probability of occurrence at time t is a function ofthe probability of occurrence at time tminus1 (MacKenzie NicholsHinesKnutsonampFranklin2003)IncontrasttostaticmodelsthisMarkovprocess recognizes that thedistributionof a species is afunctionofpreviousenvironmentalconditionsanddispersalcon‐straintsaswellascurrentenvironmentalconditions(Dormannetal2012KeacuteryGuillera‐ArroitaampLahoz‐Monfort2013)Dynamiccorrelated‐detectionoccupancymodels areamodelextension inwhichspatiallycorrelatedreplicatedsurveysareusedtoestimatedetectionprobabilities (Hinesetal20142010)Weselectedanoccupancymodelingapproachbecause failure toaccount for im‐perfect detection can bias estimates of colonization and extinc‐tionrates(Ruiz‐GutieacuterrezampZipkin2011)Weselectedadynamicapproachbecausedynamicmodelsoffermoreecological realismmoreaccurateprojectionsandgreatergeneralizabilitythanstaticmodels (Clementetal2016Yackulicetal2015)Weusedcor‐related‐detection models because the BBS generates spatiallyreplicatedsurveysandfailuretoaccountforspatialcorrelationofreplicatescanbiasoccupancyestimates(Hinesetal20142010)Finallywe used a finitemixturemodel to account for detectionheterogeneity because of the variation in habitat and focal spe‐ciesabundanceacrossthestudyarea (Clementetal2016)Afi‐nitemixturemodel approximates the heterogeneity of detectionprobabilitiesbypositingthatthepopulationconsistsofamixtureofrouteswhichhaveeitherarelativelyhighdetectionprobabilityorarelativelylowdetectionprobability
AsdetailedaboveBBSdata consistofnumerous routes eachcomposedofbirdobservationsat50 stopsA speciesmaybeab‐sentfromindividualstopswhenitispresentonaroutebutnotviceversaDetectionof a species is taken asproofof presence but anondetectionmayresultfromatrueabsenceorafalseabsenceForclarityweusethetermldquooccupiedrdquotoindicateaspeciesispresentonarouteandthetermldquoavailablerdquotoindicateaspeciesislocallypres‐entataspecificstopatthetimeofthesurvey(NicholsThomasampConn2009)Bythisterminologythedynamiccorrelated‐detectionoccupancymodelincludesthefollowingparameters
ψ=Pr(routeoccupiedduringthefirstseasonofsurveys)θ = Pr (species available at stop | route occupied and species unavailable
at previous stop)θprime= Pr (species available at stop | route occupied and species available
at previous stop)
p1 = Pr (detection at a stop for low‐detection routes | route occupied and species available at stop)
p2 = Pr (detection at a stop for high‐detection routes | route occupied and species available at stop)
π = Pr (probability that a route is a low‐detection route)εt = Pr (route is not occupied in season t + 1 | occupied in season t) andγt = Pr (route is occupied in season t + 1 | not occupied in season t)
Hinesetal(20142010)developedamodellikelihoodthatallowstheseparameterstobemodeledasfunctionsofroute‐andyear‐spe‐cificcovariateswhiledetectionparameterscanalsobeinfluencedbystop‐specificcovariatesParameterscanthenbeestimatedusingmax‐imum‐likelihoodestimationinprogramPRESENCE(Hines2006)
WedevelopedsixspecificmodelsofγandεforeachbirdspeciesWepartitionedspatialandtemporalvariationincovariatesbyparti‐tioningcovariatesintotheaveragevalueovertimeandtheannualdeviationfromthataverageInthiswaytheaveragedcovariatexiincludedspatialvariationonlywhilethedeviationcomponentΔxiyincludedtemporalvariationonlyUsingthesecovariatesourmodelsincluded(a)anullmodelinwhichγandεareconstantthroughtimeandspaceand(b)aglobal(mostgeneral)modelinwhichγandεvarythroughspaceaccordingtomeanclimateandlandcoversuitabilityandthroughtimeusingadummycovariate foreachyearWealsoconsideredintermediatemodelstoassesstheexplanatorypowerofourcovariates(c)γandεvaryspatiallywithmeanclimateandlandcover suitability but notwith time (d) γ and ε vary spatiallywithmeanclimateandlandcoversuitabilityandtemporallywithannualchangesinclimateonly(e)γandεvaryspatiallywithmeanclimateand land cover suitability and temporally with annual changes inlandcoveronlyand(f)γandεvaryspatiallywithmeanclimateandlandcoversuitabilityandtemporallywithannualchangesinclimateandlandcover
Prior to comparing these models we worked to achieve ade‐quatefitfortheothermodelparametersBecauseofthenumberofmodelparametersweconsideredthemsequentiallyInitiallywefittheglobalmodelforγandεandwemodeledθθprimeandψasfunctionsofclimateandlandcovercovariatesInthisinitialmodelweusedafinitemixturemodelonptoaccountforheterogeneitypresumablycausedbydifferencesamongroutesinhabitatobserversbirdabun‐danceandotherfeaturesthatwerenotexplicitlymodeled(Clementetal2016)Wealsomodeledpasa functionofclimateand landcover covariates and year Because bird activity often varieswithtimeofdaywealsomodeleddetectionasaquadratic functionofstopnumberwhichweconsidertobeaproxyfortimeofdayFromthis highly parameterizedmodelwe fit a reducedmodel by elim‐inating the covariatewith the smallest estimate‐to‐standard errorratioWecontinuedtoeliminatecovariatesinthiswayuntilAICin‐creasedandthenselectedthemodelwiththelowestAICWeusedthisparameterreductionprotocolwithψθθprimeandpinturnmain‐tainingthemodelstructureonγandεAfteridentifyingparsimoni‐ousmodelsforψθθprimeandpwefitthesetofsixmodelsforγandεdescribedaboveThesequentialapproachmaynotyieldidenticalresultsasasingleanalysisbutitisapragmaticapproachfordealing
1990emsp |emsp emspensp CLEMENT ET aL
with complexmodels (MacKenzie et al 2018) By beginningwithhighlyparameterizedmodelsandprogressingtowardreducedmod‐elswereducedtheriskofconfoundingtheoccupancyanddetectionprocesses (MacKenzieet al2018)Wealsochecked forpotentialmulti‐collinearityissuesbycalculatingPearsonscorrelationcoeffi‐cientforthedifferentcovariatesusedinthemodelsWetreatedanR2lt05asindicativeofalackofmulti‐collinearity
WealsoevaluatedthegoodnessoffitoftheglobalmodelforeachspeciesusingthenaiumlvecolonizationandextinctionratestocalculatetheteststatisticofaHosmerndashLemeshowtest(HosmerampLemeshow2000) Inthiscontext thenaiumlvecolonizationrate is thenumberofsitesthatchangedfromnondetectionsitesinyeart todetectionsitesinyear t+1while thenaiumlveextinction rate is thenumberof sitesthatchangedfromdetectionsitesinyeart tonondetectionsitesinyeart+1Wedividedtheroutesintodecilesbasedontheexpectedcolonizationandextinctionratesandcomparedthepredictedratesto theobservedrateswithchi‐squaretests (α=005) Ifnecessarywecombineddecilestoensurethatthepredictednumberofdetec‐tionsinadecilewasgt4AlthoughtheHosmerndashLemeshowteststatis‐ticisoftencalculatedfrompredictedpresencevitalratesseemedtobemoreappropriatemeasuresinthiscasebecausedynamicmodelsestimatechangesinpresenceratherthanpresence(Clementetal2016)Wenotethatthisuseofnaiumlveratesfocusesonproductsofmodelparameters(ieγεp ψ ϑ)andthereforewedonottestthefitofthefullyparameterizedmodelWefocusedonnaiumlveratesbe‐causethetruevital rates (ieγandε)cannotbedirectlyobservedwhendetectionisimperfectwhilethenaiumlveratescanbeobserved
We used an analysis of deviance approach (ANODEV egSkalski 1996) to evaluate the relative importance of climate andland cover to changes in bird distributions In this procedure wecompared thedeviance explainedbyour parameter of interest tothedevianceexplainedbythemostfullyparameterizedmodelinthemodelset
Dev(Mnull)isthedevianceofModel3whichaccountsforspatialbutnottemporalvariationincolonizationandextinctionprobabili‐tiesDev(Mfull)isthedevianceofModel2theglobalmodelwithbothspatialandtemporalvariationinγandεWethenevaluatedthedevi‐anceexplainedbyclimateandorlandcoverbyusingmodels45and6 forDev(Mcov)HighervaluesofR
2
Dev indicategreaterexplanatory
powerwith0indicatingnopowerand1indicatingpowerequaltothatofthefullyparameterizedmodelalthoughR2
Devcouldpotentially
exceed1becauseourcovariatemodelsarenotstrictlynestedwithinModel2ThereforewecalculatedandreportR2
Dev forbothclimate
and landcoverThis isnot theonlypossibleapproach toassessingrelativeimportanceofexplanatoryvariablesbutitissoundandhasbeenusedandrecommendedbyothers(Grosboisetal2008)
The general ANODEV approach to partitioning variation ledustofocusonhypothesistestingasameansofassessingtheneedforextramodelparameters (sourcesofvariation)Thesmallsetof
specificapriorihypothesesalsosets thisworkapart fromexplor‐atorystudiesinvestigatingtheadequacyofmanydifferentmodelsForthesereasonsweusedlikelihoodratiotestsasameansofcom‐paringdifferentmodelsalthoughwenotethatuseofmodelselec‐tioncriteria(egAICc)yieldedsimilarinferences
3emsp |emspRESULTS
Ourcovariatesindicatedasmalllossofhabitatandincreasedtem‐peraturesduringthestudyperioddespitetherelativelyshorttimescaleForEasternWoodPeweehabitatchangewasgt1on38ofrouteswiththemeanshareofsuitablehabitatnearroutesde‐cliningfrom416to409ForRed‐eyedVireohabitatchangewasgt1on23of routeswithmeansuitablehabitatdecliningfrom480to476Routetemperatureswererarelyabovetheheatstress thresholdbut theaveragetimeabovethis thresholdper route increased from 05hr per year from 1982 to 1997 to17hrperyearfrom1997to2012Wealsoobservedadeclineinperiodsofcoldstressfrom5450to5291hrperrouteperyearThePearsons correlation coefficient did not indicate anymulti‐collinearityproblemsamongourcovariates(R2lt02inallcases)
31emsp|emspEastern Wood Pewee
ForEasternWoodPeweethebest‐supportedmodelincludedbothclimateandhabitatmeasuresas covariatesofψθ andθprimeWeex‐cludedclimateandhabitatcovariatesfromthedetectionprobabilitymodelbecausesomeofthosemodelsdidnotconvergeThereforethedetectionmodelforeachspeciesincludedyearandstopnum‐ber as covariates As a result the global model included 85 pa‐rameters (Table1)TheHosmerndashLemeshowtest indicatedthattheglobalmodel fit thedataadequatelywithnoevidenceofdiscrep‐ancies between estimated and observed naiumlve colonization rates(χ2=441 df =8 p=082) and naiumlve extinction rates (χ2=1117df =8p=019)
EasternWoodPeweewerewidespreadinthestudyareawithanaverageprobabilityofoccupancyonBBSroutesin1997of090intheglobalmodel(Figure1)Occupancydeclinedslightlyto088by2012mirroringthedeclineincountsreportedbytheBBSInitialoccupancywaspositivelyrelatedtoavailablehabitatandhoursofcoldstress(Table1)Probabilityofdetectionatthefirstindividualstopduring1997waslowat013whileprobabilityofdetectionat the route level was much higher at 095 because birds weretypically available atmany stopswithina routeModel3whichincludedspatialvariationinclimateandhabitatinthecolonizationandextinctionmodelssignificantlyimprovedmodelfitrelativetoModel1(likelihoodratioχ2=4419df =6plt0001)Underthismodelestimatedcolonizationratesin1998rangedamongroutesfrom 008 to 028 (with amean of 018) while extinction ratesvariedfrom001to004(withameanof002Figure2)Model2differedfromModel3inthat itallowedcolonizationandextinc‐tionratestovaryeachyearModel2improvedmodelfitrelative
R2
Dev=Dev(Mnull)minusDev(Mcov)
Dev(Mnull)minusDev(Mfull)
emspensp emsp | emsp1991CLEMENT ET aL
TA B L E 1 emspEstimatedparametersforcorrelated‐detectiondynamicoccupancyModel2relatinghabitatandclimatecovariatestooccupancydynamicsofbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012Parameterestimatesindicatechangeinlogoddsofparametersinresponsetoz‐transformedcovariatesParametersψϑϑprimeγεp1p2andπdefinedintext
Parameters
Eastern Wood Pewee Red‐eyed Vireo
Estimate SE Estimate SE
Ψintercept 2367 0177 4136 0404
Ψhabitat 0556 0163 2315 0300
Ψheatstress minus0011 0198 minus0337 0281
Ψcoldstress 0598 0166 1020 0258
ϑintercept minus1821 0026 minus1385 0010
ϑhabitat minus0055 0026 0516 0013
ϑheatstress minus0094 0030 0017 0008
ϑcoldstress 0172 0040 0258 0013
ϑprimeintercept 2292 0045 0426 0020
ϑprimehabitat 0316 0036 1210 0018
ϑprimeheatstress minus0006 0038 0054 0013
ϑprimecoldstress minus0622 0064 0876 0019
γintercept minus0884 0353 0010 0424
γ 1999 minus0601 0585 minus0150 0532
γ 2000 minus1083 0633 0516 0464
γ 2001 minus0300 0484 0228 0483
γ 2002 minus1113 0577 0262 0511
γ 2003 minus1055 0564 minus0311 0551
γ2004 minus0656 0525 minus0179 0534
γ 2005 minus0677 0533 0468 0485
γ2006 minus1014 0620 minus0217 0579
γ2007 minus0162 0453 minus0093 0548
γ 2008 minus23545 70483373 minus0478 0607
γ 2009 minus0428 0455 minus0253 0566
γ 2010 minus0292 0478 minus0159 0517
γ 2011 minus1136 0607 0012 0490
γ 2012 minus0498 0474 0141 0497
γaveragehabitat 0095 0091 0801 0112
γaverageheatstress minus0024 0067 0005 0062
γaveragecoldstress 0227 0071 0358 0120
εintercept minus3957 0386 minus3767 0242
ε 1999 0365 0482 minus0189 0337
ε 2000 0049 0512 minus0921 0399
ε 2001 minus0266 0548 minus0532 0362
ε 2002 minus0576 0660 minus1156 0398
ε 2003 minus0298 0595 minus0692 0355
ε2004 0113 0509 minus0621 0343
ε 2005 0173 0482 minus1096 0429
ε2006 minus0145 0561 minus1173 0463
ε2007 0451 0458 minus0894 0366
ε 2008 0283 0480 minus0976 0424
ε 2009 0218 0496 minus0898 0381
(Continues)
1992emsp |emsp emspensp CLEMENT ET aL
Parameters
Eastern Wood Pewee Red‐eyed Vireo
Estimate SE Estimate SE
ε 2010 minus0397 0605 minus0348 0304
ε 2011 0643 0455 minus0999 0406
ε 2012 0153 0644 minus0803 0410
εaveragehabitat 0238 0090 minus1670 0122
εaverageheatstress minus0071 0092 minus0075 0074
εaveragecoldstress minus0307 0141 minus0904 0104
p1intercept minus2172 0043 1352 0038
p1 1998 minus0118 0048 minus0127 0058
p1 1999 minus0112 0049 minus0043 0058
p1 2000 minus0166 0050 minus0016 0059
p1 2001 minus0156 0049 0031 0060
p1 2002 minus0179 0049 0076 0061
p1 2003 minus0216 0051 0161 0064
p12004 minus0242 0051 0066 0060
p1 2005 minus0124 0050 0149 0060
p12006 minus0182 0051 0124 0059
p12007 minus0149 0050 0184 0061
p1 2008 minus0202 0052 0192 0063
p1 2009 minus0221 0052 0135 0061
p1 2010 minus0195 0051 0062 0059
p1 2011 minus0122 0051 0203 0062
p1 2012 minus0227 0055 0194 0062
p1stopnumber 0033 0009 minus0055 0011
p1stopnumbersquared minus0069 0010 minus0247 0013
p2intercept minus0921 0018 minus0557 0036
p2 1998 minus0138 0033 minus0106 0050
p2 1999 minus0137 0032 0000 0052
p2 2000 minus0164 0032 minus0027 0051
p2 2001 minus0077 0032 minus0053 0053
p2 2002 minus0101 0032 minus0019 0052
p2 2003 minus0199 0033 0086 0053
p22004 minus0186 0032 0124 0057
p2 2005 minus0146 0032 0076 0051
p22006 minus0155 0032 0128 0054
p22007 minus0153 0032 0120 0054
p2 2008 minus0168 0032 0072 0056
p2 2009 minus0077 0033 0101 0055
p2 2010 minus0114 0033 0085 0060
p2 2011 minus0079 0032 0124 0054
p2 2012 minus0186 0033 0036 0061
p2stopnumber 0045 0007 0165 0010
p2stopnumbersquared minus0105 0008 minus0183 0010
π 0533 0064 minus0505 0065
TA B L E 1 emsp (Continued)
emspensp emsp | emsp1993CLEMENT ET aL
toModel3althoughnotsignificantlyduetothenumberofaddi‐tional parameters (χ2=2679df =28p=053)UnderModel 2estimatedcolonizationratesvariedfrom000in2008to030in1998whileextinctionratesvariedfrom001 in2001ndash2003and2010to004in2011WeusedModels45and6toestimatethepercentof improvedmodel fit (measuredby thechange indevi‐ance)generatedbyModel2thatcouldbeaccountedforbytempo‐ralvariationinclimateandhabitatcovariates(Table2)Temporalvariationinhabitataccountedfor04ofthechangeindeviancewhile temporal variation in climate accounted for 196Model6whichaccountedfortemporalvariationinbothhabitatandcli‐mateindicatedthatin1998colonizationinthesouthwestportionofthestudyareawasabovethelocalaveragewhileextinctioninthesouthwestwasbelowthelocalaverageindicatingarelativelystrongoccupancy trend in thesouthwest in thatyear (Figure3)By2012thispatternhadreversedyieldingarelativelyweakoc‐cupancytrendinthesouthwest(Figure3)NoneofthesemodelsrepresentedasignificantimprovementoverModel3(pgt026)Insomecasesthesignoftheestimatedeffectoftemporalvariationinclimateandhabitatwasoppositeoftheexpectedeffectbutinnoinstancesweretheseestimatessignificant(Table3)
32emsp|emspRed‐eyed Vireo
The best‐supportedmodel for Red‐eyedVireo included the samecovariatesasforEasternWoodPeweeyieldingaglobalmodelwith85parameters (Table1)TheHosmerndashLemeshowtest indicatedfitwasadequate for theRed‐eyedVireoglobalmodel forbothnaiumlvecolonization rates (χ2=540df =4p=025) and naiumlve extinctionrates(χ2=100df =4p=091)
Red‐eyedVireowere similarlywidespread in the study areawith an average probability of occupancy of 091 in the global
model (Figure1)Occupancydecreasedslightlyto090by2012incontrasttothe increase incountsreportedbytheBBS Initialoccupancywas positively related to available habitat and hoursofcoldstress (Table1)Probabilityofdetectionat the first stopwasrelativelyhighat038whileprobabilityofdetectionattheroute levelnearlyperfectat098Model3which includedspa‐tialvariationinclimateandhabitattothecolonizationandextinc‐tionmodelsimprovedmodelfitdramatically(χ2=41884df =6plt0001)relativetothenullmodelInkeepingwiththegreaterchangeindevianceforRed‐eyedVireoModel3estimatedmorespatialvariationinvitalratescomparedtoEasternWoodPeweeIn 1998 estimated colonization rates varied across BBS routesfrom 010 to 088 (mean of 049)while extinction rates rangedfrom000to048(meanof004Figure4)Model2whichallowedcolonization and extinction rates to vary each year improvedmodelfitrelativetoModel3butnotsignificantlyduetothenum‐berofadditionalparameters(χ2=3427df =28p=019)UnderModel2estimatedcolonizationratesvariedfrom039in2008to060 in2000while estimatedextinction rates varied from003in2006to007in1998WeusedModels45and6toestimatethepercentofthedeviancereductioninModel2thatcouldbeac‐countedforbytemporalvariationinclimateandhabitatcovariates(Table2)Wefoundthattemporalvariation inhabitataccountedfor01of the change indeviancewhile climate accounted for56Model 6 which accounted for temporal variation in bothhabitatandclimateindicatedthatin1998bothcolonizationandextinctionwereaboveaverageindicatinghigherturnoverearlyinthestudyperiod(Figure5)By2012bothcolonizationandextinc‐tion rateshaddeclined indicatinggreater stability inoccupancy(Figure 5) None of these models represented a significant im‐provementoverModel3(pgt074)Someoftheestimatedeffects
F I G U R E 1 emspOccupancyprobabilityfor(a)EasternWoodPeweeand(b)Red‐eyedVireointheeasternUnitedStates1997underModel2Occupancyvariesbytheamountofsuitablehabitathoursofheatstressandhoursofcoldstressin1997
1994emsp |emsp emspensp CLEMENT ET aL
oftemporalvariationinclimateandhabitatdifferedfromexpecta‐tionsbutnotsignificantly(Table3)
4emsp |emspDISCUSSION
Wefound that forbothbirdspecies initialoccupancycoloniza‐tionratesandextinctionratescorrelatedwithspatialvariationinclimateandhabitatcovariatesHowevercolonizationandextinc‐tionrateswerenotcorrelatedwithtemporalvariation inclimateand habitat covariates Our results correlating initial occupancywith environmental covariates are broadly speaking consistentwithbiologicalexpectationandaraftofpreviousstudiesthathaveindicatedasignificantrelationshipbetweenspeciesdistributionsandhabitat(RobinsonWilsonampCrick2001ThogmartinSauerampKnutson 2007) climate (Barbet‐Massinamp Jetz 2014BellardBertelsmeier Leadley Thuiller amp Courchamp 2012) or both(Seoane Bustamante amp Diacuteaz‐Delgado 2004 Sohl 2014)Howeverwesuggestthatestimatedrelationshipsbetweenspatialvariationinenvironmentalcovariatesandcolonizationandextinc‐tion rates are of greater ecological interest than relatively phe‐nomenological species distribution models or climate envelopemodels(Clementetal2016)Theestimatedvitalratesofcoloni‐zationandextinctionbringusclosertounderstandingthedynamicprocessunderlying thedistributionof these speciesGivencon‐stantvital rateswecanalsoproject theequilibrium levelofoc‐cupancyatsites ( i
i+i
atsite i)Wecancomparetheseprojected
equilibria tocurrentoccupancytoproject trends for thespeciesunder current environmental conditions We can also use
projectedchangesinkeyenvironmentalvariablestodeveloppre‐dicted rangechanges in the futureWeargue that theseprojec‐tions are more robust than those from static climate envelopemodels which assume that species are currently in equilibrium(Yackulicetal2015)Incontraststaticmodelscanoverestimaterange changes when the equilibrium does not hold (Morin ampThuiller2009)
Given the importanceofglobal changeweweremotivated toinvestigate factors affecting temporal aswell as spatial variationin vital rates In this case spatial variation must be incorporatedintosuchanalysesinordertoaccommodatethedifferentdynamicsexpectedindifferentportionsofaspeciesrangeWeviewthere‐lationshipbetweentemporalvariationincovariatevaluesandvitalrates as the most relevant to understanding the effect of globalchangeontherecentandfuturedistributionsofourstudyspeciesFurthermore a correlation between vital rates and spatially vary‐ingcovariatesmaynot indicateasimilarcorrelationwithtemporalchangesinthosesamecovariatesForexampleaspecieswithstrongdispersal constraintsmight have a strong spatial relationshipwithacovariatebutaweaktemporalrelationshipduetoits immobility(Devictor et al 2008) Similarly an estimated spatial relationshipmaybepurelyphenomenologicalduetothenatureofspatialvari‐ablesWhen spatial variables followgradients it is relatively easytoobtainstatisticallysignificantcorrelationsabsentanycausalre‐lationship(Grosboisetal2008)ForexampleifcolonizationratesandtemperaturesbothfollownorthndashsouthgradientstheymaybespatiallycorrelatedevenifcolonizationisgovernedbysomeotherunidentifiedfactorInthiscasewewouldexpecttheweakcorrela‐tion between temporal changes in temperature and colonization
F I G U R E 2 emspProbabilityof(a)colonizationand(b)extinctionforEasternWoodPeweeintheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1995CLEMENT ET aL
ratestobemoreindicativeofthetruerelationshipthanthestrongcorrelationwithspatialvariationintemperature
BBS surveys indicate different abundance trajectories for ourstudyspecieswithcounts increasingforRed‐eyedVireosandde‐creasing for EasternWood Pewees (Sauer et al 2015) Becauseabundanceandoccupancytendtobelinked(Gastonetal2000)weexpectedsomedivergenceinoccupancydynamicsWefoundthatestimatedoccupancywashighandstableforbothspeciesalthoughtherewasgreaterturnoverforRed‐eyedVireosForexampleunderModel3 (spatialvariationonly incovariates)bothaveragecoloni‐zationrates(049)andaverageextinctionrates(004)wereatleasttwiceashighforRed‐eyedVireoscomparedtoEasternWoodPewee(018and002respectively)Red‐eyedVireosexhibitedgreaterspa‐tialvariationincolonizationandextinctionrates(Figure4)althoughour selected covariates explained little temporal variation for thisspeciesEasternWoodPeweecolonizationandextinctionratesex‐hibitedmoreofalatitudinalgradientreflectingaspatialcorrelationwith extreme temperature (Figure 2)Our results also hinted at agreatercorrelationbetweentemporalchanges in temperatureandoccupancyvitalratesforEasternWoodPeweewhichcouldplayaroleinthedeclineofthisspeciesalthoughtheresultswerenotsig‐nificant(Table3)Alternatively ithasbeensuggestedthatEasternWoodPeweemaybedecliningduetoa lossofforestedwinteringhabitat(RobbinsSauerGreenbergampDroege1989)orduetodeer‐mediatedchanges in foreststructureonbreedinggrounds (deCal‐esta 1994) illustrating the complexity of ascertaining cause andeffectusingobservationalstudiesinthesenaturalsystems
Althoughrelativelyrare(Siramietal2017)otherstudieshaveinvestigated both climate and land cover effects on bird distribu‐tions Inagreementwithour finding that temporalvariation incli‐mate explainedmuchmore of themodel deviance than temporalvariation in habitat climate‐only species distribution models hadbetter cross‐validation support than habitat‐only models for 409Europeanbirdspecies(Barbet‐Massinetal2012)SimilarlymodelsestimatingthenumberofnewlyoccupiedareasbyHoodedWarblers(Wilsonia citrina) inOntariousingclimatedatahadbetter informa‐tioncriterionsupportthanmodelsusingforestdata(MellesFortinLindsay amp Badzinski 2011) In contrast land‐use models outper‐formedclimatemodels for18 farmlandbird species in theUnitedKingdom(EglingtonampPearce‐Higgins2012)Similarlyvegetation‐based species distributionmodels had superior performance thanclimate‐based models for 79 Spanish bird species (Seoane et al2004)andforthreebirdspeciesinNewMexico(FriggensampFinch2015) Two studies focused on colonization and extinction ratesfoundmixed results ForGardenWarblers (Sylvia borin) inBritainvegetationexplainedmorevariationincolonizationwhiletempera‐turesexplainedmorevariationinlocalextinction(MustinAmarampRedpath2014)For122birdspeciesinOntariothebest‐supportedcovariates (land cover or climate change) varied among species(YalcinampLeroux2018)
Thedivergentresultsamongstudiesregardingtheimportanceofclimateandlandcovermaypartlybeduetothebiologyofindividualspecies (Yalcinamp Leroux 2018) For example farmland birdsmayTA
BLE
2emspR2 DevfordifferentmodelsofcolonizationandextinctionprobabilitiesforbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012
Mod
el
Cova
riate
s on
colo
niza
tion
and
extin
ctio
nEa
ster
n W
ood
Pew
eeRe
d‐ey
ed V
ireo
Inte
rcep
tAv
erag
e H
abita
tAv
erag
e Cl
imat
eH
abita
t Dev
iatio
nCl
imat
e D
evia
tion
Year
Eff
ect
Δ minus
2LL
R2 Dev
Δ minus
2LL
R2 Dev
2X
XX
X0
0010
00
0010
0
6X
XX
XX
minus2138
202
minus3230
57
4X
XX
Xminus2154
196
minus3234
56
5X
XX
Xminus2667
04
minus3422
01
3X
XX
minus2679
0minus3427
0
1X
minus7098
NA
minus45311
NA
Not
eNAnotapplicable
1996emsp |emsp emspensp CLEMENT ET aL
beparticularlyaffectedbychangesinagriculturalintensitybecauseoftheirextensiveuseoffarms(EglingtonampPearce‐Higgins2012)Alternativelyithasbeenarguedthatspatialscalemayplayaroleintherelativeimportanceofglobalchangefactorswithclimatemoreimportantatlargescales(PearsonampDawson2003)Wenotethattwostudiesthatfoundalargerroleforclimatebothreliedonlower‐resolution bird surveys (05deg atlas in Barbet‐Massin et al 2012
10km2atlasinMellesetal2011)IntheBBSdatathatweanalyzedsurveyswereconductedevery800mbutwecalculatedoccupancyacross each 394km transect In addition the cited studies useddifferentdata sources anddiverse analysismethods complicatinginterpretationofthedifferingresults
While we estimated that climate explained more model de‐viance than habitat these estimated relationships were not
F I G U R E 3 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionrates((t)minus (t)minus )forEasternWoodPeweeintheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
emspensp emsp | emsp1997CLEMENT ET aL
TA B L E 3 emspEstimatedparametersforcorrelated‐detectiondynamicoccupancyModel6relatinghabitatandclimatecovariatestooccupancydynamicsofbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012Parameterestimatesindicatechangeinlogoddsofparametersinresponsetoz‐transformedcovariatesParametersψϑϑprimeγε p1p2andπdefinedintextForkeyparameters(boldtext)expectedsignsincluded
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
Ψintercept 2367 0177 4087 0381
Ψhabitat 0556 0163 2323 0288
Ψheatstress minus0011 0198 minus0322 0275
Ψcoldstress 0598 0166 1054 0251
ϑintercept minus1821 0026 minus1385 0010
ϑhabitat minus0055 0026 0516 0013
ϑheatstress minus0094 0030 0017 0008
ϑcoldstress 0172 0040 0256 0013
ϑprimeintercept 2292 0045 0426 0020
ϑprimehabitat 0316 0036 1210 0018
ϑprimeheatstress minus0006 0038 0054 0013
ϑprimecoldstress minus0622 0064 0875 0019
γintercept minus0884 0353 0023 0162
γhabitatdeviation + minus0292 0478 minus0240 1768
γheatstressdeviation minus minus1136 0607 0064 0194
γcoldstressdeviation minus minus0498 0474 2398 2253
γaveragehabitat 0095 0091 0788 0111
γaverageheatstress minus0024 0067 0004 0062
γaveragecoldstress 0227 0071 0333 0119
εintercept minus3957 0386 minus4471 0141
εhabitatdeviation minus minus0397 0605 0121 1492
εheatstressdeviation + 0643 0455 minus0034 0182
εcoldstressdeviation + 0153 0644 2042 2072
εaveragehabitat 0238 0090 minus1664 0123
εaverageheatstress minus0071 0092 minus0080 0074
εaveragecoldstress minus0307 0141 minus0939 0104
p1intercept minus2172 0043 1351 0039
p1 1998 minus0118 0048 minus0128 0058
p1 1999 minus0112 0049 minus0043 0058
p1 2000 minus0166 0050 minus0017 0059
p1 2001 minus0156 0049 0031 0061
p1 2002 minus0179 0049 0076 0062
p1 2003 minus0216 0051 0160 0064
p12004 minus0242 0051 0066 0061
p1 2005 minus0124 0050 0149 0061
p12006 minus0182 0051 0124 0060
p12007 minus0149 0050 0183 0062
p1 2008 minus0202 0052 0191 0064
p1 2009 minus0221 0052 0134 0062
p1 2010 minus0195 0051 0062 0060
p1 2011 minus0122 0051 0202 0063
(Continues)
1998emsp |emsp emspensp CLEMENT ET aL
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
p1 2012 minus0227 0055 0193 0063
p1stopnumber 0033 0009 minus0055 0011
p1stopnumbersquared minus0069 0010 minus0247 0013
p2intercept minus0921 0018 minus0555 0037
p2 1998 minus0138 0033 minus0121 0050
p2 1999 minus0137 0032 minus0025 0052
p2 2000 minus0164 0032 minus0027 0051
p2 2001 minus0077 0032 minus0059 0053
p2 2002 minus0101 0032 minus0012 0052
p2 2003 minus0199 0033 0085 0053
p22004 minus0186 0032 0113 0059
p2 2005 minus0146 0032 0082 0052
p22006 minus0155 0032 0135 0054
p22007 minus0153 0032 0124 0055
p2 2008 minus0168 0032 0073 0057
p2 2009 minus0077 0033 0100 0056
p2 2010 minus0114 0033 0071 0062
p2 2011 minus0079 0032 0120 0055
p2 2012 minus0186 0033 0030 0063
p2stopnumber 0045 0007 0165 0010
p2stopnumbersquared minus0105 0008 minus0182 0011
π 0533 0064 minus0505 0065
TA B L E 3 emsp (Continued)
F I G U R E 4 emspProbabilityof(a)colonizationand(b)extinctionforRed‐eyedVireointheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1999CLEMENT ET aL
significantanddidnotexplainalargefractionofvariationThereareseveralpotentialexplanationsforwhywedidnotfindasub‐stantialresponsetochangesinhabitatandclimateFirstourdatawerelackingindynamiceventstoinformourmodelsThecovari‐ateschangedrelativelylittleoverthetimeperiodmakingitdiffi‐culttodetectaresponseThelackofchangewaspartlybecauseoftherelativelyshort(16‐year)timeperiodstudiedAlsoweusedamovingaverageforclimatedatawhichtendstoreducedifferences
overshorttimeperiods Inadditionthestudyregionhasexperi‐encedlesswarmingoverthepastcenturythanotherpartsoftheplanet (Walshetal2014)Thebirddistributionsalso lackeddy‐namismBecauseextinctionrateswerelowtherewerefewlocalextinctioneventstoinformthemodelAlthoughcolonizationrateswerehighthefactthatinitialoccupancywashighandextinctionrateswerelowmeanttherewerefewopportunitiesforcoloniza‐tionevents
F I G U R E 5 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionratesforRed‐eyedVireointheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
2000emsp |emsp emspensp CLEMENT ET aL
Second measurement error may have obscured responses toglobalchangeThePRISMclimatedataareestimatedbyinterpola‐tionratherthandirectmeasurementacrossthestudyarea(Dalyetal2008)AssuchtheremaybeartifactsoranomaliesthatimpededourestimationandincreaseduncertaintySimilarlyNLCDdataareremotelysensedandmay includesomemeasurementerrorwhichcan increase uncertainty in our estimates (Homer et al 2015)Additionallylandcoverwasonlymeasuredinthreeyearsrequiringus to imputevalues in13yearsAsa result all landcover changethatoccurredbetween2002and2006wasrecordedin2002whilechangesbetween2007and2011wererecorded in2007Thisap‐proximation isnot idealbutwasnecessarygiventhe5‐yearreso‐lutionofthedataThislowerresolutionofthelandcoverdatamayhavereducedthesensitivityofthemodel
Third wemay havemisspecified ourmodelWe tried to de‐velopbiologicallymeaningfulcovariatesbutwemightnotunder‐standthesystemaswellaswehopedItispossiblethatbirdvitalrates respond to climateon adifferent time scale (ie not a15‐yearmoving average) or to climateduringdifferent timesof theyearortodifferenttemperaturethresholdsortodifferentclimatecomponents (egprecipitation Illaacutenetal2014) It isalsopossi‐ble that thebirds respond to finerhabitat features than the landcovertypesweusedoratadifferentspatialscale(ienot400mPearsonampDawson2003)Alternatively theremaybe importantregionaldifferencesor interactionsamongcovariatesthatwedidnotcaptureOritispossiblethatchangesinclimateandhabitatintheirbreedingareassimplyhavelittleeffectonvitalratesforthesespecies relative to other factors such as competitionwith otherbirdspeciesorglobalchangeoccurringontheirwinteringgroundsinSouthAmerica(Dormann2007)Ofcoursethesemisspecifica‐tionissuesareinherenttoallobservationalstudiesandnotuniquetothisstudy
Itwould be possible for us to explore some of themisspec‐ification issues just mentioned For example we could rerunthe samemodelsusingnewcovariates representing a varietyoftemperature thresholds and habitat buffer sizes to search for astrongercorrelationExperiencesuggeststhatpersistencewouldeventuallyberewardedwithap‐valuelt005ora lowmodelse‐lection valueHowever one reason to avoid such exploration isthat it tends to lead toType Ierrorsoverfitmodels andbiasedparameterestimates(FiebergampJohnson2015)Furthermorewefind thecontrastbetweensignificant correlations for spatial co‐variatesandnonsignificantcorrelationsfortemporalcovariatesil‐luminatingIntypicalclimateenvelopestudieswewouldestimatethe relationshipbetweencovariatesand thecurrentdistributionofaspeciesmuchlikeourestimateofinitialoccupancy(Dormann2007)BecauseweobtainedsignificantparameterestimateswemightconcludethatwehaveidentifiedcovariatesthatdeterminetherangeofthisspeciesThenwemightusetheseresultstoproj‐ectthefuturerangeofthespeciesundervariousclimatescenariosbased on the (questionable) assumption that the covariatendashspe‐cies relationship will not change as climate and habitat change(Cuddington et al 2013 Elith et al 2010Gustafson 2013) In
this case the insignificant correlationwith the temporal covari‐atessuggeststhatouroccupancycovariatesmerelydescribetherangeofthesebirdswithoutdeterminingtherangeFurthermoreour results suggest that despite significant results in the staticportionof themodelwehaveyet to identify themost relevantcomponents of climate change shaping thedistributionof thesebirds although itmay be difficult to identify these componentsgiven the lackofobservedchangeduringa relatively short timeperiodFinallyourresultssuggestthatprojectionsbasedonthestaticportionofourmodelareunlikelytobereliabledespitetheirstatisticalsignificance
Ifourinterpretationiscorrectitsuggeststhatsomeprojectionsof futurerangeshiftsbasedonstaticmodelsrelyonunsupportedassumptionsandarenot reliableWehavedemonstratedadiffer‐entapproachthatdoesnotrequirethestrongassumptionsofstaticmodelingapproachesOurapproachwouldbemoreapttoidentifyfactorsthatguidespeciesrsquoresponsetoglobalchangeifboththespe‐ciesandthecovariatesexhibitedmoredynamismandifweanalyzedalongertimeperiodthatencompassedgreaterchange
ACKNOWLEDG MENTS
WethankKeithPardieckDaveZiolkowskiJrandthethousandsofhighlyskilledvolunteerswhohavecoordinatedandperformedtheNorthAmericanBreedingBirdSurvey for half a centuryWealsothanktheClimateResearchUnitattheUniversityofEastAngliaformakingvaluableclimatedataeasilyaccessibleMJCwasfinanciallysupported by the BBS and the USGS Northeast Climate ScienceCenterThecontentsofthisdocumentaretheresponsibilityoftheauthorsanddonotnecessarilyrepresenttheviewsoftheUSGSAnyuseoftradeproductorfirmnamesisfordescriptivepurposesonlyanddoesnotimplyendorsementbytheUSGovernment
DEDIC ATION
We dedicate this paper to the late Chandler S Robbins ldquoFatherrdquooftheNorthAmericanBreedingBirdSurvey (BBS)DatafromtheBBSformthebasisforthisspecificpaperandforhundredsofothersfocusedon topics ranging from local trends tocontinentalmacro‐ecologyChanwasauniquecombinationofbird‐loverandscientistwhosevisionandinquisitiveenthusiasmwereaninspirationtoallofusfortunateenoughtohaveknownhim
CONFLIC T OF INTERE S T
Nonedeclared
AUTHORSrsquo CONTRIBUTION
JACandJDNconceivedthestudyAJTandSWanalyzedhabitatandclimatedataMJCJEHandJDNperformedtheoccupancyanaly‐sisMJCledthewritingofthemanuscriptAllauthorsgavefinalap‐provalforpublication
emspensp emsp | emsp2001CLEMENT ET aL
DATA ACCE SSIBILIT Y
Raw data are available from the following sources Breeding BirdSurvey httpswwwpwrcusgsgovbbsrawdata PRISM climatedata httpwwwprismoregonstateedu NLCD land cover datahttpswwwmrlcgov Processed data will be archived with theDryadDigitalRepository
ORCID
Matthew J Clement httpsorcidorg0000‐0003‐4231‐7949
James D Nichols httpsorcidorg0000‐0002‐7631‐2890
Adam J Terando httpsorcidorg0000‐0002‐9280‐043X
James E Hines httpsorcidorg0000‐0001‐5478‐7230
Steven G Williams httpsorcidorg0000‐0003‐3760‐6818
R E FE R E N C E S
Anderson D R Burnham K P Gould W R amp Cherry S (2001)Concerns about finding effects that are actually spuriousWildlife Society Bulletin29311ndash316
Arauacutejo M B Cabeza M Thuiller W Hannah L amp WilliamsP H (2004) Would climate change drive species out of re‐serves An assessment of existing reserve‐selection meth‐ods Global Change Biology 10 1618ndash1626 httpsdoiorg101111j1365‐2486200400828x
Barbet‐MassinMampJetzW(2014)A40‐yearcontinent‐widemulti‐speciesassessmentofrelevantclimatepredictorsforspeciesdistri‐butionmodellingDiversity and Distributions201285ndash1295httpsdoiorg101111ddi12229
Barbet‐Massin M Thuiller W amp Jiguet F (2012) The fate ofEuropean breeding birds under climate land‐use and disper‐sal scenarios Global Change Biology 18 881ndash890 httpsdoiorg101111j1365‐2486201102552x
Bellard C Bertelsmeier C Leadley P Thuiller W ampCourchamp F (2012) Impacts of climate change on the fu‐ture of biodiversity Ecology Letters 15 365ndash377 httpsdoiorg101111j1461‐0248201101736x
Boumlhning‐Gaese K amp Bauer H G (1996) Changes in species abun‐dance distribution and diversity in a central European birdcommunity Conservation Biology 10 175ndash187 httpsdoiorg101046j1523‐1739199610010175x
BrommerJELehikoinenAampValkamaJ(2012)Thebreedingrangesof central European and Arctic bird speciesmove poleward PLoS ONE7e43648httpsdoiorg101371journalpone0043648
CalderWAampKingJR(1974)ThermalandcaloricrelationsofbirdsInDSFarnerampJRKing(Eds)Avian biology(Vol4pp259ndash413)NewYorkNYAcademicPress
CesaraccioCSpanoDDucePampSnyderRL(2001)Animprovedmodel for determining degree‐day values from daily temperaturedataInternational Journal of Biometeorology45161ndash169httpsdoiorg101007s004840100104
ChamberlinTC (1890)ThemethodofmultipleworkinghypothesesScience1592ndash96
ClementMJHinesJENicholsJDPardieckKLampZiolkowskiDJJr (2016)Estimating indicesofrangeshifts inbirdsusingdy‐namicmodelswhendetectionisimperfectGlobal Change Biology223273ndash3285httpsdoiorg101111gcb13283
Commission for Environmental Cooperation (1997) Ecological regionsofNorthAmericaTowardacommonperspectiveCommission for
EnvironmentalCooperationMontrealRetrievedfromftpftpepagovwedecoregionscec_naCEC_NAecopdf
Crowl TA Crist TO Parmenter R R BelovskyGamp LugoA E(2008)Thespreadofinvasivespeciesandinfectiousdiseaseasdriv‐ersofecosystemchangeFrontiers in Ecology and the Environment6238ndash246httpsdoiorg101890070151
Cuddington K FortinM J Gerber L RHastings A Liebhold AOConnorMampRayC(2013)Process‐basedmodelsarerequiredtomanageecologicalsystemsinachangingworldEcosphere41ndash12httpsdoiorg101890ES12‐001781
DalyCHalbleibMSmithJIGibsonWPDoggettMKTaylorGHhellipPasterisPP(2008)Physiographicallysensitivemappingofcli‐matologicaltemperatureandprecipitationacrosstheconterminousUnited States International Journal of Climatology 28 2031ndash2064httpsdoiorg101002joc1688
DawsonW (1958) Relation of oxygen consumption and evaporativewater losstotemperatureintheCardinalPhysiological Zoology3137ndash48httpsdoiorg101086physzool31130155377
DawsonWRampWhittowGC (2000)Regulationofbodytempera‐ture InG CWhittow (Ed)Sturkiersquos avian physiology (5th ed pp343ndash390)SanDiegoCAAcademicPress
DeCalestaDS(1994)Effectofwhite‐taileddeeronsongbirdswithinmanagedforestsinPennsylvaniaJournal of Wildlife Management58711ndash718httpsdoiorg1023073809685
DevictorVJulliardRCouvetDampJiguetF(2008)Birdsaretrack‐ing climatewarming but not fast enoughProceedings of the Royal Society of London B Biological Sciences2752743ndash2748httpsdoiorg101098rspb20080878
Dormann C F (2007) Promising the future Global change projec‐tionsofspeciesdistributionsBasic and Applied Ecology8387ndash397httpsdoiorg101016jbaae200611001
Dormann C F Schymanski S J Cabral J Chuine I Graham CHartigFhellipSingerA(2012)Correlationandprocessinspeciesdis‐tributionmodelsBridgingadichotomyJournal of Biogeography392119ndash2131httpsdoiorg101111j1365‐2699201102659x
EccelE(2010)WhatwecanasktohourlytemperaturerecordingPartIIHourlyinterpolationoftemperaturesforclimatologyandmodel‐lingItalian Journal of Agrometeorology1545ndash50
EccelEampCordanoE(2013)InterpolTHourlyinterpolationofmulti‐pletemperaturedailyseriesRpackageversion211RetrievedfromhttpsCRANR‐projectorgpackage=InterpolT
EglingtonSMampPearce‐HigginsJW(2012)Disentanglingtherela‐tiveimportanceofchangesinclimateandland‐useintensityindriv‐ingrecentbirdpopulationtrendsPLoS ONE7e30407httpsdoiorg101371journalpone0030407
Elith JKearneyMampPhillipsS (2010)Theartofmodelling range‐shifting species Methods in Ecology and Evolution 1 330ndash342httpsdoiorg101111j2041‐210X201000036x
FiebergJampJohnsonDH(2015)MMIMultimodelinferenceormod‐elswithmanagement implications Journal of Wildlife Management79708ndash718
FriggensMMampFinchDM (2015) Implicationsof climatechangeforbirdconservationinthesouthwesternUSunderthreealternativefuturesPLoS ONE10e0144089
FryJXianGZJinSDewitzJHomerCGYangLhellipWickhamJD (2011)Completionof the2006national landcoverdatabasefor theconterminousUnitedStatesPhotogrammetric Engineering amp Remote Sensing77858ndash864
Fuller R J Gregory R D Gibbons D W Marchant J HWilson J D Baillie S R amp Carter N (1995) Population de‐clines and range contractions among lowland farmland birdsin Britain Conservation Biology 9 1425ndash1441 httpsdoiorg101046j1523‐1739199509061425x
Gaston K J Blackburn T M Greenwood J J Gregory R DQuinn R M amp Lawton J H (2000) Abundancendashoccupancy
2002emsp |emsp emspensp CLEMENT ET aL
relationships Journal of Applied Ecology 37 39ndash59 httpsdoiorg101046j1365‐2664200000485x
GoldsteinRB(1974)RelationofmetabolismtoambienttemperatureintheverdinCondor76116ndash119httpsdoiorg1023071365995
GrinnellJ(1917)FieldtestsoftheoriesconcerningdistributionalcontrolAmerican Naturalist51115ndash128httpsdoiorg101086279591
GrosboisVGimenezOGaillardJMPradelRBarbraudCClobertJhellipWeimerskirchH(2008)Assessingtheimpactofclimatevari‐ation on survival in vertebrate populationsBiological Reviews 83357ndash399httpsdoiorg101111j1469‐185X200800047x
GustafsonEJ(2013)WhenrelationshipsestimatedinthepastcannotbeusedtopredictthefutureUsingmechanisticmodelstopredictlandscapeecologicaldynamicsinachangingworldLandscape Ecology281429ndash1437httpsdoiorg101007s10980‐013‐9927‐4
Hamel P B (1992) LandManagers guide to the birds of the SouthGeneralTechnicalReportSE‐22USDAForestServiceSoutheasternForestExperimentStationandTheNatureConservancyChapelHillNC
Hinds D S amp Calder W A (1973) Temperature regulation of thePyrrhuloxiaandtheArizonaCardinalPhysiological Zoology4655ndash71httpsdoiorg101086physzool46130152517
HinesJE(2006)PRESENCE Software to estimate patch occupancy and related parameters Laurel MD US Geological survey PatuxentWildlife Research Center Retrieved from httpwwwmbr‐pwrcusgsgovsoftwarepresencehtml
HinesJENicholsJDampCollazoJA(2014)Multiseasonoccupancymodels for correlated replicate surveys Methods in Ecology and Evolution5583ndash591httpsdoiorg1011112041‐210X12186
HinesJENicholsJDRoyleJAMacKenzieDIGopalaswamyAMKumarNampKaranthKU(2010)TigersontrailsOccupancymodeling for cluster sampling Ecological Applications 20 1456ndash1466httpsdoiorg10189009‐03211
HockeyPA SiramiCRidleyARMidgleyGFampBabikerHA(2011) Interrogating recent range changes in South African birdsConfounding signals from land use and climate change present achallenge for attribution Diversity and Distributions 17 254ndash261httpsdoiorg101111j1472‐4642201000741x
Hoegh‐Guldberg O Hughes L McIntyre S Lindenmayer D BParmesanCPossinghamHPampThomasCD (2008)AssistedcolonizationandrapidclimatechangeScience321345ndash346
Homer C Dewitz J Fry J Coan M Hossain N Larson C hellipWickham J (2007) Completion of the 2001 national land coverdatabase for the conterminous United States Photogrammetric Engineering amp Remote Sensing73337
HomerCDewitzJYangLJinSDanielsonPXianGhellipMegownK(2015)Completionofthe2011NationalLandCoverDatabasefortheconterminousUnitedStatesndashrepresentingadecadeoflandcoverchange informationPhotogrammetric Engineering amp Remote Sensing81345ndash354
HootenMBWikleCKDorazioRMampRoyleJA(2007)HierarchicalspatiotemporalmatrixmodelsforcharacterizinginvasionsBiometrics63558ndash567httpsdoiorg101111j1541‐0420200600725x
HosmerDWJrampLemeshowS(2000)Applied logistic regression(2nded)NewYorkNYJohnWileyampSons
Huntley B Collingham Y CGreen R EHiltonGM RahbekCamp Willis S G (2006) Potential impacts of climatic change upongeographical distributions of birds Ibis 148 8ndash28 httpsdoiorg101111j1474‐919X200600523x
IgleciaMNCollazoJAampMcKerrowAJ(2012)Useofoccupancymodels to evaluate expert knowledge‐based species‐habitat rela‐tionshipsAvian Conservation and Ecology7(2)13pRetrievedfromhttpwwwace‐ecoorgvol7iss2art5
IllaacutenJGThomasCDJonesJAWongWKShirleySMampBettsMG(2014)Precipitationandwintertemperaturepredictlong‐termrange‐scale abundancechanges inWesternNorthAmericanbirds
Global Change Biology 20 3351ndash3364 httpsdoiorg101111gcb12642
IoannidisJP (2005)WhymostpublishedresearchfindingsarefalsePLoS Medicine2e124
KeacuteryMGuillera‐ArroitaGampLahoz‐Monfort J J (2013)Analysingand mapping species range dynamics using occupancy modelsJournal of Biogeography 40 1463ndash1474 httpsdoiorg101111jbi12087
Khaliq I Hof C Prinzinger R Boumlhning‐Gaese K amp PfenningerM(2014) Global variation in thermal tolerances and vulnerability ofendotherms to climate change Proceedings of the Royal Society B Biological Sciences 281(1789) 20141097 httpsdoiorg101098rspb20141097
Laursen K amp Frikke J (2008) Hunting from motorboats displacesWadden Sea eiders Somateria mollissima from their favouredfeeding distribution Wildlife Biology 14 423ndash433 httpsdoiorg1029810909‐6396‐144423
LemoineNBauerHGPeintingerMampBoumlhning‐GaeseK (2007)Effectsof climate and land‐use changeon species abundance in acentralEuropeanbirdcommunityConservation Biology21495ndash503httpsdoiorg101111j1523‐1739200600633x
MacArthurRH(1972)Geographical ecology Patterns in the distribution of speciesNewYorkNYHarperampRow
MacKenzieDINicholsJDHinesJEKnutsonMGampFranklinAB(2003)Estimatingsiteoccupancycolonizationandlocalextinc‐tionwhenaspeciesisdetectedimperfectlyEcology842200ndash2207httpsdoiorg10189002‐3090
MacKenzieDINicholsJDLachmanGBDroegeSRoyleJAampLangtimmCA(2002)Estimatingsiteoccupancyrateswhendetec‐tionprobabilitiesarelessthanoneEcology832248ndash2255httpsdoiorg1018900012‐9658(2002)083[2248ESORWD]20CO2
MacKenzieDINicholsJDRoyleJAPollockKHBaileyLampHinesJE(2018)Occupancy estimation and modeling Inferring pat‐terns and dynamics of species occurrence (2nd ed) BurlingtonMAAcademicPress
MellesSJFortinMJLindsayKampBadzinskiD(2011)ExpandingnorthwardInfluenceofclimatechangeforestconnectivityandpop‐ulationprocessesonathreatenedspeciesrangeshiftGlobal Change Biology1717ndash31httpsdoiorg101111j1365‐2486201002214x
Morin X amp Thuiller W (2009) Comparing niche‐and process‐based models to reduce prediction uncertainty in species rangeshifts under climate change Ecology 90 1301ndash1313 httpsdoiorg10189008‐01341
MustinKAmarAampRedpathSM(2014)Colonizationandextinctiondynamicsofadecliningmigratorybirdareinfluencedbyclimateandhabitat degradation Ibis 156 788ndash798 httpsdoiorg101111ibi12173
NicholsJDThomasLampConnPB(2009)InferencesaboutlandbirdabundancefromcountdataRecentadvancesandfuturedirectionsInDLThomsonEJCoochampMJConroy(Eds)Modeling demo‐graphic processes in marked populations(pp201ndash235)NewYorkNYSpringer
OmernikJM(1995)EcoregionsAspatialframeworkforenvironmen‐talmanagement InWSDavisampTSimon(Eds)Biological assess‐ment and criteria Tools for water resource planning and decision making (pp49ndash62)BocaRatonFLLewisPublishers
OmernikJM(2004)Perspectivesonthenatureanddefinitionofeco‐logicalregionsEnvironmental Management34(Supplement1)S27ndashS38httpsdoiorg101007s00267‐003‐5197‐2
Omernik J M amp Griffith G E (2014) Ecoregions of the contermi‐nousUnited States Evolution of a hierarchical spatial frameworkEnvironmental Management541249ndash1266httpsdoiorg101007s00267‐014‐0364‐1
Pardieck K L ZiolkowskiD J JrampHudsonM‐A R (2015)NorthAmericanBreedingBirdSurveyDataset1966ndash2014version20140
emspensp emsp | emsp2003CLEMENT ET aL
USGeologicalSurveyPatuxentWildlifeResearchCenterRetrievedfromhttpswwwpwrcusgsgovbbsrawdata
ParmesanCampYoheG (2003)Agloballycoherentfingerprintofcli‐mate change impacts across natural systemsNature 421 37ndash42httpsdoiorg101038nature01286
PearsonRGampDawsonTP(2003)PredictingtheimpactsofclimatechangeonthedistributionofspeciesArebioclimateenvelopemod‐elsusefulGlobal Ecology and Biogeography12361ndash371
PlattJR(1964)StronginferenceScience146347ndash353PopperK(1959)The logic of scientific discoveryLondonUKHutchinsonRobbins C S Sauer J R Greenberg R S amp Droege S (1989)
Population declines in North American birds that migrate to theNeotropics Proceedings of the National Academy of Sciences 867658ndash7662httpsdoiorg101073pnas86197658
Robinson R A Wilson J D amp Crick H Q (2001) The impor‐tance of arable habitat for farmland birds in grassland land‐scapes Journal of Applied Ecology 38 1059ndash1069 httpsdoiorg101046j1365‐2664200100654x
Romesburg H C (1981) Wildlife science Gaining reliable knowl‐edge Journal of Wildlife Management 45 293ndash313 httpsdoiorg1023073807913
Root T (1988) Energy constraints on avian distributions and abun‐dancesEcology69330ndash339httpsdoiorg1023071940431
Ruiz‐GutieacuterrezVampZipkinEF (2011)Detectionbiasesyieldmis‐leadingpatternsof speciespersistence and colonization in frag‐mented landscapesEcosphere2 1ndash14 httpsdoiorg101890ES10‐002071
SauerJRHinesJEFallonJEPardieckKLZiolkowskiDJJrampLinkWA(2015)The North American breeding bird survey results and analysis 1966ndash2013 Version 01302015LaurelMDUSGSPatuxentWildlifeResearchCenterhttpswwwmbr‐pwrcusgsgovbbsbbshtml
Sauer J R LinkWA Fallon J E Pardieck K L amp ZiolkowskiDJJr(2013)TheNorthAmericanbreedingbirdsurvey1966ndash2011SummaryanalysisandspeciesaccountsNorth American Fauna791ndash32httpsdoiorg103996nafa790001
SeoaneJBustamanteJampDiacuteaz‐DelgadoR(2004)Competingrolesforlandscapevegetationtopographyandclimateinpredictivemod‐elsofbirddistributionEcological Modelling171209ndash222httpsdoiorg101016jecolmodel200308006
Sirami C Caplat P Popy S Clamens A Arlettaz R Jiguet F hellipMartin J‐L (2017) Impacts of global change on species distri‐butions Obstacles and solutions to integrate climate and landuse Global Ecology and Biogeography 26 385ndash394 httpsdoiorg101111geb12555
SkalskiJR(1996)Regressionofabundanceestimatesfrommarkrecap‐ture surveys against environmental covariatesCanadian Journal of
Fisheries and Aquatic Sciences53196ndash204httpsdoiorg101139f95‐169
SohlTL(2014)Therelativeimpactsofclimateandland‐usechangeonconterminousUnitedStatesbird species from2001 to2075PLoS ONE9e112251httpsdoiorg101371journalpone0112251
ThogmartinWESauerJRampKnutsonMG(2007)ModelingandmappingabundanceofAmericanwoodcockacrosstheMidwesternandNortheasternUnitedStatesJournal of Wildlife Management71376ndash382httpsdoiorg1021932005‐680
ThomasCDampLennonJJ (1999)Birdsextendtheirrangesnorth‐wardsNature399213httpsdoiorg10103820335
Trathan P N Garciacutea‐Borboroglu P Boersma D Bost C ACrawford R J CrossinG ThellipWienecke B (2015) Pollutionhabitatlossfishingandclimatechangeascriticalthreatstopen‐guins Conservation Biology 29 31ndash41 httpsdoiorg101111cobi12349
TravisJMJ(2003)Climatechangeandhabitatdestructionadeadlyanthropogenic cocktail Proceedings of the Royal Society of London Series B Biological Sciences 270(1514) 467ndash473 httpsdoiorg101098rspb20022246
UrbanMC (2015)Acceleratingextinction risk fromclimatechangeScience348571ndash573httpsdoiorg101126scienceaaa4984
WalshJWuebblesDHayhoeKKossinJKunkelKStephensGhellipSomervilleR(2014)OurchangingclimateInJMMelilloTCRichmondampGWYohe(Eds)Climate change impacts in the United States The Third National climate assessment(pp19ndash67)WashingtonDCUSGovernmentPrintingOffice
YackulicCBNichols JDReid JampDerR (2015)Topredict thenichemodelcolonizationandextinctionEcology9616ndash23httpsdoiorg10189014‐13611
Yalcin S amp Leroux S J (2018) An empirical test of the relative andcombinedeffectsofland‐coverandclimatechangeonlocalcoloni‐zationandextinctionGlobal Change Biology243849ndash3861httpsdoiorg101111gcb14169
ZhuKWoodallCWampClarkJS(2012)FailuretomigrateLackoftreerangeexpansioninresponsetoclimatechangeGlobal Change Biology181042ndash1052httpsdoiorg101111j1365‐2486201102571x
How to cite this articleClementMJNicholsJDCollazoJATerandoAJHinesJEWilliamsSGPartitioningglobalchangeAssessingtherelativeimportanceofchangesinclimateandlandcoverforchangesinaviandistributionEcol Evol 201991985ndash2003 httpsdoiorg101002ece34890

emspensp emsp | emsp1987CLEMENT ET aL
2003withsomelocalizeddeclinesinFloridaTexasandthePacificNorthwest(Saueretal2015)TheEasternWoodPeweebreedsintheeasternUnitedStatesandpartsofsoutheasternCanadaIncon‐trasttoRed‐eyedVireoscountsoftheEasternWoodPeweehavebeendecliningby15since1966andby12since2003(Saueretal2015)DespitethegenerallynegativetrendtherehavebeenlocalizedincreasesincountsintheMidwest
Toestimatetherelativeimportanceofclimateandlandcovertochangesinbirddistributionswegeneratedtestablehypothesesex‐pressingrelationshipsbetweenbirdsandenvironmentalcovariatesAcommonapproachistogenerateanextensivesetofhypothesesfromasuiteofenvironmentalmetricsWecanthenfitthespecifiedmodelsandobservewhichmetricsyieldsignificantresultsHowevertestingmanystatisticalhypotheseshasapropensitytoidentifyspu‐riousrelationshipsandsuchanalysesshouldbeconsideredexplor‐atory(AndersonBurnhamGouldampCherry2001Ioannidis2005)Inthisstudywedevelopedtwoapriorihypothesesfromecologicalprinciples
1 Changes in the total annual periods of extreme temperaturedrive changes in bird distributionsWe focus on extreme tem‐perature because we expect these are the periods of greateststress due to cold or heat stress increased metabolic costsreduced resource availability or other mechanisms (Dawson ampWhittow 2000 Root 1988) We defined ldquoextremerdquo relativetothethermoneutralzone(TNZ)whichistherangeofambienttemperaturesunderwhichendothermscanmaintain theirbodytemperature without deviating from their basal metabolic rate(Calder amp King 1974) The TNZ varies among species but hasbeenestimatedas18ndash38degC fora ldquogenericrdquobird (CalderampKing1974) Several small temperate‐zone passerines (similar to ourstudy species) including the Northern Cardinal Verdin HouseSparrow and Red‐breasted Nuthatch have been documentedto have TNZs similar to that of this ldquogenericrdquo bird (Dawson1958 Goldstein 1974 Hinds amp Calder 1973 Khaliq HofPrinzinger Boumlhning‐Gaese amp Pfenninger 2014) Therefore weexpected temperatures below 18degC and above 38degC to beassociated with changes in bird distributions
2 Changes in the amount of appropriate habitat locally availableduringthebreedingseasondrivechangesinbirddistributionsbe‐cause inappropriatehabitatwillnotprovidesufficient foodandshelterforbreedingbirds(FriggensampFinch2015IgleciaCollazoampMcKerrow2012) For theEasternWoodPeweewedefinedappropriatehabitats asdeciduous forest evergreen forest andmixedforest(Hamel1992)FortheRed‐eyedVireowedefinedappropriate habitats as the same forest types and shrubndashscrub(Hamel1992)
Ofcoursethesehypothesesarenotexhaustivebutweselectedthem as logical general mechanisms that might underlie avian re‐sponsestoclimateandlandcoverchangeWeexpectedthepaucityand specificity of our hypotheseswould reduceour chancesof ob‐taining statistically significant resultsbychanceForexample anull
hypothesisthatbirddistributionswillnotexpandataspecifictempera‐tureishardertorejectthanthenullhypothesisthatbirddistributionswillnotexpandatanytemperatureWeviewamorespecifichypothe‐sisasamorepowerfuldiscriminatorytool(Platt1964Popper1959)moreinkeepingwiththehypothetico‐deductivemethod(Chamberlin1890Romesburg1981) andwith aType1error rate closer to thenominallevel(Andersonetal2001Ioannidis2005)
Westatedourhypothesizedeffectsintermsofchanges inbirddistribution(iebirdswillincreasewherehabitatisavailable)ratherthan static patterns of bird distribution (ie birds will be locatedwhere habitat is available)We prefer this formulation because itfocusesontheecologicalprocessesthatunderliespeciesdistribu‐tionsandtheirdynamics Inmetapopulationtheorythevitalratesdescribingtheseprocessesareoftenthelocalcolonizationandex‐tinction rates Models that account for ecological vital rates (iedynamicmodels)areusefulbecausetheyarerelativelymechanisticthereby strengthening the generality of findings and contributingmoretoecologicalunderstandingIncontrastinordertobeusefulforforecastingandprojectionrelationshipsbetweenenvironmentalcovariates andoccurrence require an assumption that the speciesinquestion is inequilibriumwith theenvironmental factorsunderstudy (Elith et al 2010) Themore phenomenological orientationofmodelsthatfocusonoccurrencecanbelimitingwhenprojectingmodelresultsintonovelsituationssuchasfutureclimateandlandcoverconditions(Cuddingtonetal2013Gustafson2013)Incon‐trast dynamicmodels avoid the equilibrium assumption enablingmore robust predictions of occurrence under future conditions(Clementetal2016Yackulicetal2015)
Bothclimateand landcovervary spatiallyand temporally andeachprocess could impactbirddistributions It is possible for thelevelofcorrelationwithbirdvital rates todifferacross those twodimensions For example consider an environmental variable thatiscompletelystaticthroughagiventimeperiodandaspeciesdistri‐butionthatshiftsduringthesametimeperiodInthiscaseitisnotpossibleforchangeinthestaticvariabletohavecausedthedistri‐butionalshiftbecausenosuchchangeoccurredHoweveritisstillpossibleforcolonizationandextinctiontocorrelatewiththeenvi‐ronmentalvariablebecauseofaspatialcorrelationbetweenthemHere our goalwas to incorporate temporal variation in covariatevalues intoouranalysisbecausemuchresearch intoglobalchangeismotivatedbyinterestintemporalchangesinspeciesdistributionsTherefore our hypotheses and models are structured to isolateandevaluatetheabilityofenvironmentalcovariatestoaccountforspatial and temporal variation in local colonization and extinctionprobabilities
22emsp|emspData
We obtained data on the detection and nondetection of breed‐ing Red‐eyed Vireos and Eastern Wood Pewees from the NorthAmerican Breeding Bird Survey (Pardieck Ziolkowski amp Hudson2015 wwwpwrcusgsgovBBSRawData) The Breeding BirdSurvey (BBS) is a joint project of the US Geological Survey and
1988emsp |emsp emspensp CLEMENT ET aL
theCanadianWildlifeServicedesignedtomonitor thestatusandtrendsofbirdsbreedinginNorthAmerica(Saueretal2013)Sinceitsinceptionin1966theBBShasexpandedtoincludeover5000survey routes across theUnited States and southernCanada ap‐proximately3000ofwhichare surveyedeachyearEach route issurveyedonceperyearbyaskilledvolunteerThedateofthesurveyistimedtooccurduringpeakterritorialbehaviortypicallylateMaytoearlyJulydependingon latitudeSurveyorsfollowaprescribedroutealongsecondaryroadsstoppingatapproximately800‐min‐tervalsandperforming3‐minroadsidepointcountsgenerating50countsper394kmrouteWeselectedBBSdatabecausetheirgreatgeographicand temporalextent is suitable for investigatingdistri‐butionalchanges
We selected a subsetof the availableBBSdata for use inouranalysis Our modeling approach described below makes use ofstop‐specificsurveyresultsbutthesedetailedresultsarecurrentlyavailableindigitalformatsonlysince1997Thereforeweuseddatafrom1997to2012BecauseourstudyspeciesarefoundinforestedregionsofeasternNorthAmericawedelineatedthestudyareabyaggregatingthreeOmernikLevelIclassesrepresentingeasternfor‐estecoregionsNorthernForestsEasternTemperateForestsandTropicalWetForests (CommissionforEnvironmentalCooperation1997Omernik19952004OmernikampGriffith2014)ThisstudyareaincludestheUnitedStateseastoftheGreatPlains(29millionkm2) This region encompassed the entire breeding range of theEasternWoodPeweeexceptsomeareasinCanadanotcoveredbyourclimateandhabitatcovariatesWeselectedBBSroutesentirelywithinthisstudyareaforanalysisWeexcludedroutesthatwerenotclassifiedasldquoactiverdquobytheBBSandroutesthatdifferedfromtherecommendedlength(394km)bymorethan25Weonlyanalyzedsurveys conducted under acceptable conditions (ie acceptableweatherdatetimeofdaystopsreportedbyBBSasldquoruntype=1rdquo)OntherareoccasionsthataroutehadmultipleacceptablesurveysperyearweuseddatafromthefirstacceptablesurveyThisyielded1371routesforouranalysisFinallyweconvertedcountsofbirdsateachstopalongeachBBSroutetodetectionnondetectiondatasuitableforpresencendashabsencemodelingThisconversionsacrificeddata richness but allowed us to account for imperfect detectionof species Using the full count data typically requires additionalassumptions (Illaacuten et al 2014) or incorporation of additional datasources(HootenWikleDorazioampRoyle2007)
ClimatedatawereobtainedfromthePRISMproject(Dalyetal2008)ThePRISMoutputuseslocallyweightedregressionmodelstointerpolatelong‐termweatherstationobservationstoa4kmresolu‐tiongridDailyinterpolatedclimatedataareavailablefrom1981tothepresentFromthesedatawecalculatedtwotemperature‐basedindices based on our first hypothesis described above The firstindexwascalculatedasthetotalannualhoursabove38degCwhilethesecondindexwascalculatedasthetotalannualhoursbelow18degC
Becausethetemperature‐basedindiceswerebasedonhoursofexposureitwasnecessarytointerpolatethedailyPRISMdatatoes‐timatehourlyvaluesWeconsideredasimplelinearinterpolationtoimputevaluesbetweendailymaximumandminimumtemperatures
However for extreme temperatures thismethodwould likely re‐sult in estimated exposure durations that are biased low InsteadweadoptedthemethoddescribedbyCesaraccioSpanoDuceandSnyder(2001seeFigure3)andEccel (2010)usingtheRsoftwarepackageldquoInterpolTrdquo (EccelampCordano2013) Inthismethodcan‐didatefunctionsarefit toobservedhourlytemperaturedatafromnearby high‐quality ASOS (Automated Station Observing System)weather stations in the study regionusing a combinationof sinu‐soidalandquadratic functionsForeach4kmPRISMgridcell theresultingcalibrationfunctionfromthenearestASOSstationisthenappliedtoeachdaysmaximumandminimumtemperaturetopredicttheinterveninghourlyvalues
Usingtheinterpolatedhourlytemperatureestimateswecalcu‐latedthetotalannualhourswithinthezonesofheatstressorcoldstress for each year from 1981 to 2012 Annual valueswere cal‐culatedover theperiodJune1ndashMay31 to represent thebreedingcycleOncetheannualresidencetimeswereobtainedweapplieda15‐yearmovingaveragewindowtothedataresultingin16mov‐ingaverageperiodsstartingwiththe198182ndash199596periodandendingwiththe199697ndash201112periodApplyingamovingaver‐ageactsasahigh‐passfilterthatattenuatesinterannualvariabilitywhileamplifyinganylong‐termclimatechangesignalsOtherperiodlengths could have been chosen however we hypothesized that15years represented a reasonable tradeoff between (a) reducingthemovingaveragelengthsoasnottoexceedthelengthofthedailyPRISMdataserieswhichbeginsin1981(b)increasingthemovingaveragelengthsothatclimaticchanges(asopposedtobackgroundclimaticvariation)canbeidentifiedand(c)usingaperiodthatisbio‐logicallymeaningfulinthatwecanreasonablyexpectanylong‐termclimaticchangesinthemovingaverageswouldbeoutsidethehis‐torical rangeofvariabilityand thusshouldaffectcolonizationandextinctionratesofsensitivespecies
We obtained land cover data from the National Land CoverDatabase (NLCD) for 2001 (Homer et al 2007) 2006 (Fry et al2011)and2011(Homeretal2015)WereclassifiedthegivenlandcovertypesintohabitatornonhabitatforeachbirdspeciesbasedonoursecondhypothesisdescribedaboveWecalculatedtheper‐centof landareathatwasasuitablehabitatwithin400mofeachrouteandusedthisasacovariate inouranalysisWeused400mbecausethisishalfthedistancebetweenstopsonBBSroutesForyearswithoutlandcoverdataweusedthelandcovervaluesofthesubsequent NLCD survey Specifically we used 2001 values for1997ndash20012006valuesfor2002ndash2006and2011valuesfor2007ndash2012Weusedaz‐transformationtocenterandscaleallcovariatespriortoanalysis
23emsp|emspAnalysis
OurgoalwastoassesstherelativeimportanceofclimateandlandcovertochangesinthedistributionofRed‐eyedVireosandEasternWoodPeweesWhileourbirddataconsistedofdetectionnonde‐tection data from BBS surveys our parameters of interest weretheprobabilitiesoflocalcolonizationandlocalextinctionasthese
emspensp emsp | emsp1989CLEMENT ET aL
processes drive changes in distribution Our predictor variableswereourmeasuresof climate and land cover suitability for eachspeciesdescribedaboveWeuseddynamic correlated‐detectionoccupancymodels with detection heterogeneity to estimate therelationshipbetweenourpredictorsandbirddistributiondynam‐ics (Clement et al 2016HinesNichols amp Collazo 2014Hinesetal2010)Occupancymodelsareaclassofmodelsthatestimatetheprobabilityofpresencewhileaccountingforimperfectdetec‐tionofspeciesbyanalyzingreplicated(usuallytemporally)surveys(MacKenzieetal20022018)Dynamicoccupancymodelsmodelinterannual changes in occupancy by positing aMarkov processinwhich the probability of occurrence at time t is a function ofthe probability of occurrence at time tminus1 (MacKenzie NicholsHinesKnutsonampFranklin2003)IncontrasttostaticmodelsthisMarkovprocess recognizes that thedistributionof a species is afunctionofpreviousenvironmentalconditionsanddispersalcon‐straintsaswellascurrentenvironmentalconditions(Dormannetal2012KeacuteryGuillera‐ArroitaampLahoz‐Monfort2013)Dynamiccorrelated‐detectionoccupancymodels areamodelextension inwhichspatiallycorrelatedreplicatedsurveysareusedtoestimatedetectionprobabilities (Hinesetal20142010)Weselectedanoccupancymodelingapproachbecause failure toaccount for im‐perfect detection can bias estimates of colonization and extinc‐tionrates(Ruiz‐GutieacuterrezampZipkin2011)Weselectedadynamicapproachbecausedynamicmodelsoffermoreecological realismmoreaccurateprojectionsandgreatergeneralizabilitythanstaticmodels (Clementetal2016Yackulicetal2015)Weusedcor‐related‐detection models because the BBS generates spatiallyreplicatedsurveysandfailuretoaccountforspatialcorrelationofreplicatescanbiasoccupancyestimates(Hinesetal20142010)Finallywe used a finitemixturemodel to account for detectionheterogeneity because of the variation in habitat and focal spe‐ciesabundanceacrossthestudyarea (Clementetal2016)Afi‐nitemixturemodel approximates the heterogeneity of detectionprobabilitiesbypositingthatthepopulationconsistsofamixtureofrouteswhichhaveeitherarelativelyhighdetectionprobabilityorarelativelylowdetectionprobability
AsdetailedaboveBBSdata consistofnumerous routes eachcomposedofbirdobservationsat50 stopsA speciesmaybeab‐sentfromindividualstopswhenitispresentonaroutebutnotviceversaDetectionof a species is taken asproofof presence but anondetectionmayresultfromatrueabsenceorafalseabsenceForclarityweusethetermldquooccupiedrdquotoindicateaspeciesispresentonarouteandthetermldquoavailablerdquotoindicateaspeciesislocallypres‐entataspecificstopatthetimeofthesurvey(NicholsThomasampConn2009)Bythisterminologythedynamiccorrelated‐detectionoccupancymodelincludesthefollowingparameters
ψ=Pr(routeoccupiedduringthefirstseasonofsurveys)θ = Pr (species available at stop | route occupied and species unavailable
at previous stop)θprime= Pr (species available at stop | route occupied and species available
at previous stop)
p1 = Pr (detection at a stop for low‐detection routes | route occupied and species available at stop)
p2 = Pr (detection at a stop for high‐detection routes | route occupied and species available at stop)
π = Pr (probability that a route is a low‐detection route)εt = Pr (route is not occupied in season t + 1 | occupied in season t) andγt = Pr (route is occupied in season t + 1 | not occupied in season t)
Hinesetal(20142010)developedamodellikelihoodthatallowstheseparameterstobemodeledasfunctionsofroute‐andyear‐spe‐cificcovariateswhiledetectionparameterscanalsobeinfluencedbystop‐specificcovariatesParameterscanthenbeestimatedusingmax‐imum‐likelihoodestimationinprogramPRESENCE(Hines2006)
WedevelopedsixspecificmodelsofγandεforeachbirdspeciesWepartitionedspatialandtemporalvariationincovariatesbyparti‐tioningcovariatesintotheaveragevalueovertimeandtheannualdeviationfromthataverageInthiswaytheaveragedcovariatexiincludedspatialvariationonlywhilethedeviationcomponentΔxiyincludedtemporalvariationonlyUsingthesecovariatesourmodelsincluded(a)anullmodelinwhichγandεareconstantthroughtimeandspaceand(b)aglobal(mostgeneral)modelinwhichγandεvarythroughspaceaccordingtomeanclimateandlandcoversuitabilityandthroughtimeusingadummycovariate foreachyearWealsoconsideredintermediatemodelstoassesstheexplanatorypowerofourcovariates(c)γandεvaryspatiallywithmeanclimateandlandcover suitability but notwith time (d) γ and ε vary spatiallywithmeanclimateandlandcoversuitabilityandtemporallywithannualchangesinclimateonly(e)γandεvaryspatiallywithmeanclimateand land cover suitability and temporally with annual changes inlandcoveronlyand(f)γandεvaryspatiallywithmeanclimateandlandcoversuitabilityandtemporallywithannualchangesinclimateandlandcover
Prior to comparing these models we worked to achieve ade‐quatefitfortheothermodelparametersBecauseofthenumberofmodelparametersweconsideredthemsequentiallyInitiallywefittheglobalmodelforγandεandwemodeledθθprimeandψasfunctionsofclimateandlandcovercovariatesInthisinitialmodelweusedafinitemixturemodelonptoaccountforheterogeneitypresumablycausedbydifferencesamongroutesinhabitatobserversbirdabun‐danceandotherfeaturesthatwerenotexplicitlymodeled(Clementetal2016)Wealsomodeledpasa functionofclimateand landcover covariates and year Because bird activity often varieswithtimeofdaywealsomodeleddetectionasaquadratic functionofstopnumberwhichweconsidertobeaproxyfortimeofdayFromthis highly parameterizedmodelwe fit a reducedmodel by elim‐inating the covariatewith the smallest estimate‐to‐standard errorratioWecontinuedtoeliminatecovariatesinthiswayuntilAICin‐creasedandthenselectedthemodelwiththelowestAICWeusedthisparameterreductionprotocolwithψθθprimeandpinturnmain‐tainingthemodelstructureonγandεAfteridentifyingparsimoni‐ousmodelsforψθθprimeandpwefitthesetofsixmodelsforγandεdescribedaboveThesequentialapproachmaynotyieldidenticalresultsasasingleanalysisbutitisapragmaticapproachfordealing
1990emsp |emsp emspensp CLEMENT ET aL
with complexmodels (MacKenzie et al 2018) By beginningwithhighlyparameterizedmodelsandprogressingtowardreducedmod‐elswereducedtheriskofconfoundingtheoccupancyanddetectionprocesses (MacKenzieet al2018)Wealsochecked forpotentialmulti‐collinearityissuesbycalculatingPearsonscorrelationcoeffi‐cientforthedifferentcovariatesusedinthemodelsWetreatedanR2lt05asindicativeofalackofmulti‐collinearity
WealsoevaluatedthegoodnessoffitoftheglobalmodelforeachspeciesusingthenaiumlvecolonizationandextinctionratestocalculatetheteststatisticofaHosmerndashLemeshowtest(HosmerampLemeshow2000) Inthiscontext thenaiumlvecolonizationrate is thenumberofsitesthatchangedfromnondetectionsitesinyeart todetectionsitesinyear t+1while thenaiumlveextinction rate is thenumberof sitesthatchangedfromdetectionsitesinyeart tonondetectionsitesinyeart+1Wedividedtheroutesintodecilesbasedontheexpectedcolonizationandextinctionratesandcomparedthepredictedratesto theobservedrateswithchi‐squaretests (α=005) Ifnecessarywecombineddecilestoensurethatthepredictednumberofdetec‐tionsinadecilewasgt4AlthoughtheHosmerndashLemeshowteststatis‐ticisoftencalculatedfrompredictedpresencevitalratesseemedtobemoreappropriatemeasuresinthiscasebecausedynamicmodelsestimatechangesinpresenceratherthanpresence(Clementetal2016)Wenotethatthisuseofnaiumlveratesfocusesonproductsofmodelparameters(ieγεp ψ ϑ)andthereforewedonottestthefitofthefullyparameterizedmodelWefocusedonnaiumlveratesbe‐causethetruevital rates (ieγandε)cannotbedirectlyobservedwhendetectionisimperfectwhilethenaiumlveratescanbeobserved
We used an analysis of deviance approach (ANODEV egSkalski 1996) to evaluate the relative importance of climate andland cover to changes in bird distributions In this procedure wecompared thedeviance explainedbyour parameter of interest tothedevianceexplainedbythemostfullyparameterizedmodelinthemodelset
Dev(Mnull)isthedevianceofModel3whichaccountsforspatialbutnottemporalvariationincolonizationandextinctionprobabili‐tiesDev(Mfull)isthedevianceofModel2theglobalmodelwithbothspatialandtemporalvariationinγandεWethenevaluatedthedevi‐anceexplainedbyclimateandorlandcoverbyusingmodels45and6 forDev(Mcov)HighervaluesofR
2
Dev indicategreaterexplanatory
powerwith0indicatingnopowerand1indicatingpowerequaltothatofthefullyparameterizedmodelalthoughR2
Devcouldpotentially
exceed1becauseourcovariatemodelsarenotstrictlynestedwithinModel2ThereforewecalculatedandreportR2
Dev forbothclimate
and landcoverThis isnot theonlypossibleapproach toassessingrelativeimportanceofexplanatoryvariablesbutitissoundandhasbeenusedandrecommendedbyothers(Grosboisetal2008)
The general ANODEV approach to partitioning variation ledustofocusonhypothesistestingasameansofassessingtheneedforextramodelparameters (sourcesofvariation)Thesmallsetof
specificapriorihypothesesalsosets thisworkapart fromexplor‐atorystudiesinvestigatingtheadequacyofmanydifferentmodelsForthesereasonsweusedlikelihoodratiotestsasameansofcom‐paringdifferentmodelsalthoughwenotethatuseofmodelselec‐tioncriteria(egAICc)yieldedsimilarinferences
3emsp |emspRESULTS
Ourcovariatesindicatedasmalllossofhabitatandincreasedtem‐peraturesduringthestudyperioddespitetherelativelyshorttimescaleForEasternWoodPeweehabitatchangewasgt1on38ofrouteswiththemeanshareofsuitablehabitatnearroutesde‐cliningfrom416to409ForRed‐eyedVireohabitatchangewasgt1on23of routeswithmeansuitablehabitatdecliningfrom480to476Routetemperatureswererarelyabovetheheatstress thresholdbut theaveragetimeabovethis thresholdper route increased from 05hr per year from 1982 to 1997 to17hrperyearfrom1997to2012Wealsoobservedadeclineinperiodsofcoldstressfrom5450to5291hrperrouteperyearThePearsons correlation coefficient did not indicate anymulti‐collinearityproblemsamongourcovariates(R2lt02inallcases)
31emsp|emspEastern Wood Pewee
ForEasternWoodPeweethebest‐supportedmodelincludedbothclimateandhabitatmeasuresas covariatesofψθ andθprimeWeex‐cludedclimateandhabitatcovariatesfromthedetectionprobabilitymodelbecausesomeofthosemodelsdidnotconvergeThereforethedetectionmodelforeachspeciesincludedyearandstopnum‐ber as covariates As a result the global model included 85 pa‐rameters (Table1)TheHosmerndashLemeshowtest indicatedthattheglobalmodel fit thedataadequatelywithnoevidenceofdiscrep‐ancies between estimated and observed naiumlve colonization rates(χ2=441 df =8 p=082) and naiumlve extinction rates (χ2=1117df =8p=019)
EasternWoodPeweewerewidespreadinthestudyareawithanaverageprobabilityofoccupancyonBBSroutesin1997of090intheglobalmodel(Figure1)Occupancydeclinedslightlyto088by2012mirroringthedeclineincountsreportedbytheBBSInitialoccupancywaspositivelyrelatedtoavailablehabitatandhoursofcoldstress(Table1)Probabilityofdetectionatthefirstindividualstopduring1997waslowat013whileprobabilityofdetectionat the route level was much higher at 095 because birds weretypically available atmany stopswithina routeModel3whichincludedspatialvariationinclimateandhabitatinthecolonizationandextinctionmodelssignificantlyimprovedmodelfitrelativetoModel1(likelihoodratioχ2=4419df =6plt0001)Underthismodelestimatedcolonizationratesin1998rangedamongroutesfrom 008 to 028 (with amean of 018) while extinction ratesvariedfrom001to004(withameanof002Figure2)Model2differedfromModel3inthat itallowedcolonizationandextinc‐tionratestovaryeachyearModel2improvedmodelfitrelative
R2
Dev=Dev(Mnull)minusDev(Mcov)
Dev(Mnull)minusDev(Mfull)
emspensp emsp | emsp1991CLEMENT ET aL
TA B L E 1 emspEstimatedparametersforcorrelated‐detectiondynamicoccupancyModel2relatinghabitatandclimatecovariatestooccupancydynamicsofbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012Parameterestimatesindicatechangeinlogoddsofparametersinresponsetoz‐transformedcovariatesParametersψϑϑprimeγεp1p2andπdefinedintext
Parameters
Eastern Wood Pewee Red‐eyed Vireo
Estimate SE Estimate SE
Ψintercept 2367 0177 4136 0404
Ψhabitat 0556 0163 2315 0300
Ψheatstress minus0011 0198 minus0337 0281
Ψcoldstress 0598 0166 1020 0258
ϑintercept minus1821 0026 minus1385 0010
ϑhabitat minus0055 0026 0516 0013
ϑheatstress minus0094 0030 0017 0008
ϑcoldstress 0172 0040 0258 0013
ϑprimeintercept 2292 0045 0426 0020
ϑprimehabitat 0316 0036 1210 0018
ϑprimeheatstress minus0006 0038 0054 0013
ϑprimecoldstress minus0622 0064 0876 0019
γintercept minus0884 0353 0010 0424
γ 1999 minus0601 0585 minus0150 0532
γ 2000 minus1083 0633 0516 0464
γ 2001 minus0300 0484 0228 0483
γ 2002 minus1113 0577 0262 0511
γ 2003 minus1055 0564 minus0311 0551
γ2004 minus0656 0525 minus0179 0534
γ 2005 minus0677 0533 0468 0485
γ2006 minus1014 0620 minus0217 0579
γ2007 minus0162 0453 minus0093 0548
γ 2008 minus23545 70483373 minus0478 0607
γ 2009 minus0428 0455 minus0253 0566
γ 2010 minus0292 0478 minus0159 0517
γ 2011 minus1136 0607 0012 0490
γ 2012 minus0498 0474 0141 0497
γaveragehabitat 0095 0091 0801 0112
γaverageheatstress minus0024 0067 0005 0062
γaveragecoldstress 0227 0071 0358 0120
εintercept minus3957 0386 minus3767 0242
ε 1999 0365 0482 minus0189 0337
ε 2000 0049 0512 minus0921 0399
ε 2001 minus0266 0548 minus0532 0362
ε 2002 minus0576 0660 minus1156 0398
ε 2003 minus0298 0595 minus0692 0355
ε2004 0113 0509 minus0621 0343
ε 2005 0173 0482 minus1096 0429
ε2006 minus0145 0561 minus1173 0463
ε2007 0451 0458 minus0894 0366
ε 2008 0283 0480 minus0976 0424
ε 2009 0218 0496 minus0898 0381
(Continues)
1992emsp |emsp emspensp CLEMENT ET aL
Parameters
Eastern Wood Pewee Red‐eyed Vireo
Estimate SE Estimate SE
ε 2010 minus0397 0605 minus0348 0304
ε 2011 0643 0455 minus0999 0406
ε 2012 0153 0644 minus0803 0410
εaveragehabitat 0238 0090 minus1670 0122
εaverageheatstress minus0071 0092 minus0075 0074
εaveragecoldstress minus0307 0141 minus0904 0104
p1intercept minus2172 0043 1352 0038
p1 1998 minus0118 0048 minus0127 0058
p1 1999 minus0112 0049 minus0043 0058
p1 2000 minus0166 0050 minus0016 0059
p1 2001 minus0156 0049 0031 0060
p1 2002 minus0179 0049 0076 0061
p1 2003 minus0216 0051 0161 0064
p12004 minus0242 0051 0066 0060
p1 2005 minus0124 0050 0149 0060
p12006 minus0182 0051 0124 0059
p12007 minus0149 0050 0184 0061
p1 2008 minus0202 0052 0192 0063
p1 2009 minus0221 0052 0135 0061
p1 2010 minus0195 0051 0062 0059
p1 2011 minus0122 0051 0203 0062
p1 2012 minus0227 0055 0194 0062
p1stopnumber 0033 0009 minus0055 0011
p1stopnumbersquared minus0069 0010 minus0247 0013
p2intercept minus0921 0018 minus0557 0036
p2 1998 minus0138 0033 minus0106 0050
p2 1999 minus0137 0032 0000 0052
p2 2000 minus0164 0032 minus0027 0051
p2 2001 minus0077 0032 minus0053 0053
p2 2002 minus0101 0032 minus0019 0052
p2 2003 minus0199 0033 0086 0053
p22004 minus0186 0032 0124 0057
p2 2005 minus0146 0032 0076 0051
p22006 minus0155 0032 0128 0054
p22007 minus0153 0032 0120 0054
p2 2008 minus0168 0032 0072 0056
p2 2009 minus0077 0033 0101 0055
p2 2010 minus0114 0033 0085 0060
p2 2011 minus0079 0032 0124 0054
p2 2012 minus0186 0033 0036 0061
p2stopnumber 0045 0007 0165 0010
p2stopnumbersquared minus0105 0008 minus0183 0010
π 0533 0064 minus0505 0065
TA B L E 1 emsp (Continued)
emspensp emsp | emsp1993CLEMENT ET aL
toModel3althoughnotsignificantlyduetothenumberofaddi‐tional parameters (χ2=2679df =28p=053)UnderModel 2estimatedcolonizationratesvariedfrom000in2008to030in1998whileextinctionratesvariedfrom001 in2001ndash2003and2010to004in2011WeusedModels45and6toestimatethepercentof improvedmodel fit (measuredby thechange indevi‐ance)generatedbyModel2thatcouldbeaccountedforbytempo‐ralvariationinclimateandhabitatcovariates(Table2)Temporalvariationinhabitataccountedfor04ofthechangeindeviancewhile temporal variation in climate accounted for 196Model6whichaccountedfortemporalvariationinbothhabitatandcli‐mateindicatedthatin1998colonizationinthesouthwestportionofthestudyareawasabovethelocalaveragewhileextinctioninthesouthwestwasbelowthelocalaverageindicatingarelativelystrongoccupancy trend in thesouthwest in thatyear (Figure3)By2012thispatternhadreversedyieldingarelativelyweakoc‐cupancytrendinthesouthwest(Figure3)NoneofthesemodelsrepresentedasignificantimprovementoverModel3(pgt026)Insomecasesthesignoftheestimatedeffectoftemporalvariationinclimateandhabitatwasoppositeoftheexpectedeffectbutinnoinstancesweretheseestimatessignificant(Table3)
32emsp|emspRed‐eyed Vireo
The best‐supportedmodel for Red‐eyedVireo included the samecovariatesasforEasternWoodPeweeyieldingaglobalmodelwith85parameters (Table1)TheHosmerndashLemeshowtest indicatedfitwasadequate for theRed‐eyedVireoglobalmodel forbothnaiumlvecolonization rates (χ2=540df =4p=025) and naiumlve extinctionrates(χ2=100df =4p=091)
Red‐eyedVireowere similarlywidespread in the study areawith an average probability of occupancy of 091 in the global
model (Figure1)Occupancydecreasedslightlyto090by2012incontrasttothe increase incountsreportedbytheBBS Initialoccupancywas positively related to available habitat and hoursofcoldstress (Table1)Probabilityofdetectionat the first stopwasrelativelyhighat038whileprobabilityofdetectionattheroute levelnearlyperfectat098Model3which includedspa‐tialvariationinclimateandhabitattothecolonizationandextinc‐tionmodelsimprovedmodelfitdramatically(χ2=41884df =6plt0001)relativetothenullmodelInkeepingwiththegreaterchangeindevianceforRed‐eyedVireoModel3estimatedmorespatialvariationinvitalratescomparedtoEasternWoodPeweeIn 1998 estimated colonization rates varied across BBS routesfrom 010 to 088 (mean of 049)while extinction rates rangedfrom000to048(meanof004Figure4)Model2whichallowedcolonization and extinction rates to vary each year improvedmodelfitrelativetoModel3butnotsignificantlyduetothenum‐berofadditionalparameters(χ2=3427df =28p=019)UnderModel2estimatedcolonizationratesvariedfrom039in2008to060 in2000while estimatedextinction rates varied from003in2006to007in1998WeusedModels45and6toestimatethepercentofthedeviancereductioninModel2thatcouldbeac‐countedforbytemporalvariationinclimateandhabitatcovariates(Table2)Wefoundthattemporalvariation inhabitataccountedfor01of the change indeviancewhile climate accounted for56Model 6 which accounted for temporal variation in bothhabitatandclimateindicatedthatin1998bothcolonizationandextinctionwereaboveaverageindicatinghigherturnoverearlyinthestudyperiod(Figure5)By2012bothcolonizationandextinc‐tion rateshaddeclined indicatinggreater stability inoccupancy(Figure 5) None of these models represented a significant im‐provementoverModel3(pgt074)Someoftheestimatedeffects
F I G U R E 1 emspOccupancyprobabilityfor(a)EasternWoodPeweeand(b)Red‐eyedVireointheeasternUnitedStates1997underModel2Occupancyvariesbytheamountofsuitablehabitathoursofheatstressandhoursofcoldstressin1997
1994emsp |emsp emspensp CLEMENT ET aL
oftemporalvariationinclimateandhabitatdifferedfromexpecta‐tionsbutnotsignificantly(Table3)
4emsp |emspDISCUSSION
Wefound that forbothbirdspecies initialoccupancycoloniza‐tionratesandextinctionratescorrelatedwithspatialvariationinclimateandhabitatcovariatesHowevercolonizationandextinc‐tionrateswerenotcorrelatedwithtemporalvariation inclimateand habitat covariates Our results correlating initial occupancywith environmental covariates are broadly speaking consistentwithbiologicalexpectationandaraftofpreviousstudiesthathaveindicatedasignificantrelationshipbetweenspeciesdistributionsandhabitat(RobinsonWilsonampCrick2001ThogmartinSauerampKnutson 2007) climate (Barbet‐Massinamp Jetz 2014BellardBertelsmeier Leadley Thuiller amp Courchamp 2012) or both(Seoane Bustamante amp Diacuteaz‐Delgado 2004 Sohl 2014)Howeverwesuggestthatestimatedrelationshipsbetweenspatialvariationinenvironmentalcovariatesandcolonizationandextinc‐tion rates are of greater ecological interest than relatively phe‐nomenological species distribution models or climate envelopemodels(Clementetal2016)Theestimatedvitalratesofcoloni‐zationandextinctionbringusclosertounderstandingthedynamicprocessunderlying thedistributionof these speciesGivencon‐stantvital rateswecanalsoproject theequilibrium levelofoc‐cupancyatsites ( i
i+i
atsite i)Wecancomparetheseprojected
equilibria tocurrentoccupancytoproject trends for thespeciesunder current environmental conditions We can also use
projectedchangesinkeyenvironmentalvariablestodeveloppre‐dicted rangechanges in the futureWeargue that theseprojec‐tions are more robust than those from static climate envelopemodels which assume that species are currently in equilibrium(Yackulicetal2015)Incontraststaticmodelscanoverestimaterange changes when the equilibrium does not hold (Morin ampThuiller2009)
Given the importanceofglobal changeweweremotivated toinvestigate factors affecting temporal aswell as spatial variationin vital rates In this case spatial variation must be incorporatedintosuchanalysesinordertoaccommodatethedifferentdynamicsexpectedindifferentportionsofaspeciesrangeWeviewthere‐lationshipbetweentemporalvariationincovariatevaluesandvitalrates as the most relevant to understanding the effect of globalchangeontherecentandfuturedistributionsofourstudyspeciesFurthermore a correlation between vital rates and spatially vary‐ingcovariatesmaynot indicateasimilarcorrelationwithtemporalchangesinthosesamecovariatesForexampleaspecieswithstrongdispersal constraintsmight have a strong spatial relationshipwithacovariatebutaweaktemporalrelationshipduetoits immobility(Devictor et al 2008) Similarly an estimated spatial relationshipmaybepurelyphenomenologicalduetothenatureofspatialvari‐ablesWhen spatial variables followgradients it is relatively easytoobtainstatisticallysignificantcorrelationsabsentanycausalre‐lationship(Grosboisetal2008)ForexampleifcolonizationratesandtemperaturesbothfollownorthndashsouthgradientstheymaybespatiallycorrelatedevenifcolonizationisgovernedbysomeotherunidentifiedfactorInthiscasewewouldexpecttheweakcorrela‐tion between temporal changes in temperature and colonization
F I G U R E 2 emspProbabilityof(a)colonizationand(b)extinctionforEasternWoodPeweeintheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1995CLEMENT ET aL
ratestobemoreindicativeofthetruerelationshipthanthestrongcorrelationwithspatialvariationintemperature
BBS surveys indicate different abundance trajectories for ourstudyspecieswithcounts increasingforRed‐eyedVireosandde‐creasing for EasternWood Pewees (Sauer et al 2015) Becauseabundanceandoccupancytendtobelinked(Gastonetal2000)weexpectedsomedivergenceinoccupancydynamicsWefoundthatestimatedoccupancywashighandstableforbothspeciesalthoughtherewasgreaterturnoverforRed‐eyedVireosForexampleunderModel3 (spatialvariationonly incovariates)bothaveragecoloni‐zationrates(049)andaverageextinctionrates(004)wereatleasttwiceashighforRed‐eyedVireoscomparedtoEasternWoodPewee(018and002respectively)Red‐eyedVireosexhibitedgreaterspa‐tialvariationincolonizationandextinctionrates(Figure4)althoughour selected covariates explained little temporal variation for thisspeciesEasternWoodPeweecolonizationandextinctionratesex‐hibitedmoreofalatitudinalgradientreflectingaspatialcorrelationwith extreme temperature (Figure 2)Our results also hinted at agreatercorrelationbetweentemporalchanges in temperatureandoccupancyvitalratesforEasternWoodPeweewhichcouldplayaroleinthedeclineofthisspeciesalthoughtheresultswerenotsig‐nificant(Table3)Alternatively ithasbeensuggestedthatEasternWoodPeweemaybedecliningduetoa lossofforestedwinteringhabitat(RobbinsSauerGreenbergampDroege1989)orduetodeer‐mediatedchanges in foreststructureonbreedinggrounds (deCal‐esta 1994) illustrating the complexity of ascertaining cause andeffectusingobservationalstudiesinthesenaturalsystems
Althoughrelativelyrare(Siramietal2017)otherstudieshaveinvestigated both climate and land cover effects on bird distribu‐tions Inagreementwithour finding that temporalvariation incli‐mate explainedmuchmore of themodel deviance than temporalvariation in habitat climate‐only species distribution models hadbetter cross‐validation support than habitat‐only models for 409Europeanbirdspecies(Barbet‐Massinetal2012)SimilarlymodelsestimatingthenumberofnewlyoccupiedareasbyHoodedWarblers(Wilsonia citrina) inOntariousingclimatedatahadbetter informa‐tioncriterionsupportthanmodelsusingforestdata(MellesFortinLindsay amp Badzinski 2011) In contrast land‐use models outper‐formedclimatemodels for18 farmlandbird species in theUnitedKingdom(EglingtonampPearce‐Higgins2012)Similarlyvegetation‐based species distributionmodels had superior performance thanclimate‐based models for 79 Spanish bird species (Seoane et al2004)andforthreebirdspeciesinNewMexico(FriggensampFinch2015) Two studies focused on colonization and extinction ratesfoundmixed results ForGardenWarblers (Sylvia borin) inBritainvegetationexplainedmorevariationincolonizationwhiletempera‐turesexplainedmorevariationinlocalextinction(MustinAmarampRedpath2014)For122birdspeciesinOntariothebest‐supportedcovariates (land cover or climate change) varied among species(YalcinampLeroux2018)
Thedivergentresultsamongstudiesregardingtheimportanceofclimateandlandcovermaypartlybeduetothebiologyofindividualspecies (Yalcinamp Leroux 2018) For example farmland birdsmayTA
BLE
2emspR2 DevfordifferentmodelsofcolonizationandextinctionprobabilitiesforbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012
Mod
el
Cova
riate
s on
colo
niza
tion
and
extin
ctio
nEa
ster
n W
ood
Pew
eeRe
d‐ey
ed V
ireo
Inte
rcep
tAv
erag
e H
abita
tAv
erag
e Cl
imat
eH
abita
t Dev
iatio
nCl
imat
e D
evia
tion
Year
Eff
ect
Δ minus
2LL
R2 Dev
Δ minus
2LL
R2 Dev
2X
XX
X0
0010
00
0010
0
6X
XX
XX
minus2138
202
minus3230
57
4X
XX
Xminus2154
196
minus3234
56
5X
XX
Xminus2667
04
minus3422
01
3X
XX
minus2679
0minus3427
0
1X
minus7098
NA
minus45311
NA
Not
eNAnotapplicable
1996emsp |emsp emspensp CLEMENT ET aL
beparticularlyaffectedbychangesinagriculturalintensitybecauseoftheirextensiveuseoffarms(EglingtonampPearce‐Higgins2012)Alternativelyithasbeenarguedthatspatialscalemayplayaroleintherelativeimportanceofglobalchangefactorswithclimatemoreimportantatlargescales(PearsonampDawson2003)Wenotethattwostudiesthatfoundalargerroleforclimatebothreliedonlower‐resolution bird surveys (05deg atlas in Barbet‐Massin et al 2012
10km2atlasinMellesetal2011)IntheBBSdatathatweanalyzedsurveyswereconductedevery800mbutwecalculatedoccupancyacross each 394km transect In addition the cited studies useddifferentdata sources anddiverse analysismethods complicatinginterpretationofthedifferingresults
While we estimated that climate explained more model de‐viance than habitat these estimated relationships were not
F I G U R E 3 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionrates((t)minus (t)minus )forEasternWoodPeweeintheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
emspensp emsp | emsp1997CLEMENT ET aL
TA B L E 3 emspEstimatedparametersforcorrelated‐detectiondynamicoccupancyModel6relatinghabitatandclimatecovariatestooccupancydynamicsofbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012Parameterestimatesindicatechangeinlogoddsofparametersinresponsetoz‐transformedcovariatesParametersψϑϑprimeγε p1p2andπdefinedintextForkeyparameters(boldtext)expectedsignsincluded
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
Ψintercept 2367 0177 4087 0381
Ψhabitat 0556 0163 2323 0288
Ψheatstress minus0011 0198 minus0322 0275
Ψcoldstress 0598 0166 1054 0251
ϑintercept minus1821 0026 minus1385 0010
ϑhabitat minus0055 0026 0516 0013
ϑheatstress minus0094 0030 0017 0008
ϑcoldstress 0172 0040 0256 0013
ϑprimeintercept 2292 0045 0426 0020
ϑprimehabitat 0316 0036 1210 0018
ϑprimeheatstress minus0006 0038 0054 0013
ϑprimecoldstress minus0622 0064 0875 0019
γintercept minus0884 0353 0023 0162
γhabitatdeviation + minus0292 0478 minus0240 1768
γheatstressdeviation minus minus1136 0607 0064 0194
γcoldstressdeviation minus minus0498 0474 2398 2253
γaveragehabitat 0095 0091 0788 0111
γaverageheatstress minus0024 0067 0004 0062
γaveragecoldstress 0227 0071 0333 0119
εintercept minus3957 0386 minus4471 0141
εhabitatdeviation minus minus0397 0605 0121 1492
εheatstressdeviation + 0643 0455 minus0034 0182
εcoldstressdeviation + 0153 0644 2042 2072
εaveragehabitat 0238 0090 minus1664 0123
εaverageheatstress minus0071 0092 minus0080 0074
εaveragecoldstress minus0307 0141 minus0939 0104
p1intercept minus2172 0043 1351 0039
p1 1998 minus0118 0048 minus0128 0058
p1 1999 minus0112 0049 minus0043 0058
p1 2000 minus0166 0050 minus0017 0059
p1 2001 minus0156 0049 0031 0061
p1 2002 minus0179 0049 0076 0062
p1 2003 minus0216 0051 0160 0064
p12004 minus0242 0051 0066 0061
p1 2005 minus0124 0050 0149 0061
p12006 minus0182 0051 0124 0060
p12007 minus0149 0050 0183 0062
p1 2008 minus0202 0052 0191 0064
p1 2009 minus0221 0052 0134 0062
p1 2010 minus0195 0051 0062 0060
p1 2011 minus0122 0051 0202 0063
(Continues)
1998emsp |emsp emspensp CLEMENT ET aL
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
p1 2012 minus0227 0055 0193 0063
p1stopnumber 0033 0009 minus0055 0011
p1stopnumbersquared minus0069 0010 minus0247 0013
p2intercept minus0921 0018 minus0555 0037
p2 1998 minus0138 0033 minus0121 0050
p2 1999 minus0137 0032 minus0025 0052
p2 2000 minus0164 0032 minus0027 0051
p2 2001 minus0077 0032 minus0059 0053
p2 2002 minus0101 0032 minus0012 0052
p2 2003 minus0199 0033 0085 0053
p22004 minus0186 0032 0113 0059
p2 2005 minus0146 0032 0082 0052
p22006 minus0155 0032 0135 0054
p22007 minus0153 0032 0124 0055
p2 2008 minus0168 0032 0073 0057
p2 2009 minus0077 0033 0100 0056
p2 2010 minus0114 0033 0071 0062
p2 2011 minus0079 0032 0120 0055
p2 2012 minus0186 0033 0030 0063
p2stopnumber 0045 0007 0165 0010
p2stopnumbersquared minus0105 0008 minus0182 0011
π 0533 0064 minus0505 0065
TA B L E 3 emsp (Continued)
F I G U R E 4 emspProbabilityof(a)colonizationand(b)extinctionforRed‐eyedVireointheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1999CLEMENT ET aL
significantanddidnotexplainalargefractionofvariationThereareseveralpotentialexplanationsforwhywedidnotfindasub‐stantialresponsetochangesinhabitatandclimateFirstourdatawerelackingindynamiceventstoinformourmodelsThecovari‐ateschangedrelativelylittleoverthetimeperiodmakingitdiffi‐culttodetectaresponseThelackofchangewaspartlybecauseoftherelativelyshort(16‐year)timeperiodstudiedAlsoweusedamovingaverageforclimatedatawhichtendstoreducedifferences
overshorttimeperiods Inadditionthestudyregionhasexperi‐encedlesswarmingoverthepastcenturythanotherpartsoftheplanet (Walshetal2014)Thebirddistributionsalso lackeddy‐namismBecauseextinctionrateswerelowtherewerefewlocalextinctioneventstoinformthemodelAlthoughcolonizationrateswerehighthefactthatinitialoccupancywashighandextinctionrateswerelowmeanttherewerefewopportunitiesforcoloniza‐tionevents
F I G U R E 5 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionratesforRed‐eyedVireointheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
2000emsp |emsp emspensp CLEMENT ET aL
Second measurement error may have obscured responses toglobalchangeThePRISMclimatedataareestimatedbyinterpola‐tionratherthandirectmeasurementacrossthestudyarea(Dalyetal2008)AssuchtheremaybeartifactsoranomaliesthatimpededourestimationandincreaseduncertaintySimilarlyNLCDdataareremotelysensedandmay includesomemeasurementerrorwhichcan increase uncertainty in our estimates (Homer et al 2015)Additionallylandcoverwasonlymeasuredinthreeyearsrequiringus to imputevalues in13yearsAsa result all landcover changethatoccurredbetween2002and2006wasrecordedin2002whilechangesbetween2007and2011wererecorded in2007Thisap‐proximation isnot idealbutwasnecessarygiventhe5‐yearreso‐lutionofthedataThislowerresolutionofthelandcoverdatamayhavereducedthesensitivityofthemodel
Third wemay havemisspecified ourmodelWe tried to de‐velopbiologicallymeaningfulcovariatesbutwemightnotunder‐standthesystemaswellaswehopedItispossiblethatbirdvitalrates respond to climateon adifferent time scale (ie not a15‐yearmoving average) or to climateduringdifferent timesof theyearortodifferenttemperaturethresholdsortodifferentclimatecomponents (egprecipitation Illaacutenetal2014) It isalsopossi‐ble that thebirds respond to finerhabitat features than the landcovertypesweusedoratadifferentspatialscale(ienot400mPearsonampDawson2003)Alternatively theremaybe importantregionaldifferencesor interactionsamongcovariatesthatwedidnotcaptureOritispossiblethatchangesinclimateandhabitatintheirbreedingareassimplyhavelittleeffectonvitalratesforthesespecies relative to other factors such as competitionwith otherbirdspeciesorglobalchangeoccurringontheirwinteringgroundsinSouthAmerica(Dormann2007)Ofcoursethesemisspecifica‐tionissuesareinherenttoallobservationalstudiesandnotuniquetothisstudy
Itwould be possible for us to explore some of themisspec‐ification issues just mentioned For example we could rerunthe samemodelsusingnewcovariates representing a varietyoftemperature thresholds and habitat buffer sizes to search for astrongercorrelationExperiencesuggeststhatpersistencewouldeventuallyberewardedwithap‐valuelt005ora lowmodelse‐lection valueHowever one reason to avoid such exploration isthat it tends to lead toType Ierrorsoverfitmodels andbiasedparameterestimates(FiebergampJohnson2015)Furthermorewefind thecontrastbetweensignificant correlations for spatial co‐variatesandnonsignificantcorrelationsfortemporalcovariatesil‐luminatingIntypicalclimateenvelopestudieswewouldestimatethe relationshipbetweencovariatesand thecurrentdistributionofaspeciesmuchlikeourestimateofinitialoccupancy(Dormann2007)BecauseweobtainedsignificantparameterestimateswemightconcludethatwehaveidentifiedcovariatesthatdeterminetherangeofthisspeciesThenwemightusetheseresultstoproj‐ectthefuturerangeofthespeciesundervariousclimatescenariosbased on the (questionable) assumption that the covariatendashspe‐cies relationship will not change as climate and habitat change(Cuddington et al 2013 Elith et al 2010Gustafson 2013) In
this case the insignificant correlationwith the temporal covari‐atessuggeststhatouroccupancycovariatesmerelydescribetherangeofthesebirdswithoutdeterminingtherangeFurthermoreour results suggest that despite significant results in the staticportionof themodelwehaveyet to identify themost relevantcomponents of climate change shaping thedistributionof thesebirds although itmay be difficult to identify these componentsgiven the lackofobservedchangeduringa relatively short timeperiodFinallyourresultssuggestthatprojectionsbasedonthestaticportionofourmodelareunlikelytobereliabledespitetheirstatisticalsignificance
Ifourinterpretationiscorrectitsuggeststhatsomeprojectionsof futurerangeshiftsbasedonstaticmodelsrelyonunsupportedassumptionsandarenot reliableWehavedemonstratedadiffer‐entapproachthatdoesnotrequirethestrongassumptionsofstaticmodelingapproachesOurapproachwouldbemoreapttoidentifyfactorsthatguidespeciesrsquoresponsetoglobalchangeifboththespe‐ciesandthecovariatesexhibitedmoredynamismandifweanalyzedalongertimeperiodthatencompassedgreaterchange
ACKNOWLEDG MENTS
WethankKeithPardieckDaveZiolkowskiJrandthethousandsofhighlyskilledvolunteerswhohavecoordinatedandperformedtheNorthAmericanBreedingBirdSurvey for half a centuryWealsothanktheClimateResearchUnitattheUniversityofEastAngliaformakingvaluableclimatedataeasilyaccessibleMJCwasfinanciallysupported by the BBS and the USGS Northeast Climate ScienceCenterThecontentsofthisdocumentaretheresponsibilityoftheauthorsanddonotnecessarilyrepresenttheviewsoftheUSGSAnyuseoftradeproductorfirmnamesisfordescriptivepurposesonlyanddoesnotimplyendorsementbytheUSGovernment
DEDIC ATION
We dedicate this paper to the late Chandler S Robbins ldquoFatherrdquooftheNorthAmericanBreedingBirdSurvey (BBS)DatafromtheBBSformthebasisforthisspecificpaperandforhundredsofothersfocusedon topics ranging from local trends tocontinentalmacro‐ecologyChanwasauniquecombinationofbird‐loverandscientistwhosevisionandinquisitiveenthusiasmwereaninspirationtoallofusfortunateenoughtohaveknownhim
CONFLIC T OF INTERE S T
Nonedeclared
AUTHORSrsquo CONTRIBUTION
JACandJDNconceivedthestudyAJTandSWanalyzedhabitatandclimatedataMJCJEHandJDNperformedtheoccupancyanaly‐sisMJCledthewritingofthemanuscriptAllauthorsgavefinalap‐provalforpublication
emspensp emsp | emsp2001CLEMENT ET aL
DATA ACCE SSIBILIT Y
Raw data are available from the following sources Breeding BirdSurvey httpswwwpwrcusgsgovbbsrawdata PRISM climatedata httpwwwprismoregonstateedu NLCD land cover datahttpswwwmrlcgov Processed data will be archived with theDryadDigitalRepository
ORCID
Matthew J Clement httpsorcidorg0000‐0003‐4231‐7949
James D Nichols httpsorcidorg0000‐0002‐7631‐2890
Adam J Terando httpsorcidorg0000‐0002‐9280‐043X
James E Hines httpsorcidorg0000‐0001‐5478‐7230
Steven G Williams httpsorcidorg0000‐0003‐3760‐6818
R E FE R E N C E S
Anderson D R Burnham K P Gould W R amp Cherry S (2001)Concerns about finding effects that are actually spuriousWildlife Society Bulletin29311ndash316
Arauacutejo M B Cabeza M Thuiller W Hannah L amp WilliamsP H (2004) Would climate change drive species out of re‐serves An assessment of existing reserve‐selection meth‐ods Global Change Biology 10 1618ndash1626 httpsdoiorg101111j1365‐2486200400828x
Barbet‐MassinMampJetzW(2014)A40‐yearcontinent‐widemulti‐speciesassessmentofrelevantclimatepredictorsforspeciesdistri‐butionmodellingDiversity and Distributions201285ndash1295httpsdoiorg101111ddi12229
Barbet‐Massin M Thuiller W amp Jiguet F (2012) The fate ofEuropean breeding birds under climate land‐use and disper‐sal scenarios Global Change Biology 18 881ndash890 httpsdoiorg101111j1365‐2486201102552x
Bellard C Bertelsmeier C Leadley P Thuiller W ampCourchamp F (2012) Impacts of climate change on the fu‐ture of biodiversity Ecology Letters 15 365ndash377 httpsdoiorg101111j1461‐0248201101736x
Boumlhning‐Gaese K amp Bauer H G (1996) Changes in species abun‐dance distribution and diversity in a central European birdcommunity Conservation Biology 10 175ndash187 httpsdoiorg101046j1523‐1739199610010175x
BrommerJELehikoinenAampValkamaJ(2012)Thebreedingrangesof central European and Arctic bird speciesmove poleward PLoS ONE7e43648httpsdoiorg101371journalpone0043648
CalderWAampKingJR(1974)ThermalandcaloricrelationsofbirdsInDSFarnerampJRKing(Eds)Avian biology(Vol4pp259ndash413)NewYorkNYAcademicPress
CesaraccioCSpanoDDucePampSnyderRL(2001)Animprovedmodel for determining degree‐day values from daily temperaturedataInternational Journal of Biometeorology45161ndash169httpsdoiorg101007s004840100104
ChamberlinTC (1890)ThemethodofmultipleworkinghypothesesScience1592ndash96
ClementMJHinesJENicholsJDPardieckKLampZiolkowskiDJJr (2016)Estimating indicesofrangeshifts inbirdsusingdy‐namicmodelswhendetectionisimperfectGlobal Change Biology223273ndash3285httpsdoiorg101111gcb13283
Commission for Environmental Cooperation (1997) Ecological regionsofNorthAmericaTowardacommonperspectiveCommission for
EnvironmentalCooperationMontrealRetrievedfromftpftpepagovwedecoregionscec_naCEC_NAecopdf
Crowl TA Crist TO Parmenter R R BelovskyGamp LugoA E(2008)Thespreadofinvasivespeciesandinfectiousdiseaseasdriv‐ersofecosystemchangeFrontiers in Ecology and the Environment6238ndash246httpsdoiorg101890070151
Cuddington K FortinM J Gerber L RHastings A Liebhold AOConnorMampRayC(2013)Process‐basedmodelsarerequiredtomanageecologicalsystemsinachangingworldEcosphere41ndash12httpsdoiorg101890ES12‐001781
DalyCHalbleibMSmithJIGibsonWPDoggettMKTaylorGHhellipPasterisPP(2008)Physiographicallysensitivemappingofcli‐matologicaltemperatureandprecipitationacrosstheconterminousUnited States International Journal of Climatology 28 2031ndash2064httpsdoiorg101002joc1688
DawsonW (1958) Relation of oxygen consumption and evaporativewater losstotemperatureintheCardinalPhysiological Zoology3137ndash48httpsdoiorg101086physzool31130155377
DawsonWRampWhittowGC (2000)Regulationofbodytempera‐ture InG CWhittow (Ed)Sturkiersquos avian physiology (5th ed pp343ndash390)SanDiegoCAAcademicPress
DeCalestaDS(1994)Effectofwhite‐taileddeeronsongbirdswithinmanagedforestsinPennsylvaniaJournal of Wildlife Management58711ndash718httpsdoiorg1023073809685
DevictorVJulliardRCouvetDampJiguetF(2008)Birdsaretrack‐ing climatewarming but not fast enoughProceedings of the Royal Society of London B Biological Sciences2752743ndash2748httpsdoiorg101098rspb20080878
Dormann C F (2007) Promising the future Global change projec‐tionsofspeciesdistributionsBasic and Applied Ecology8387ndash397httpsdoiorg101016jbaae200611001
Dormann C F Schymanski S J Cabral J Chuine I Graham CHartigFhellipSingerA(2012)Correlationandprocessinspeciesdis‐tributionmodelsBridgingadichotomyJournal of Biogeography392119ndash2131httpsdoiorg101111j1365‐2699201102659x
EccelE(2010)WhatwecanasktohourlytemperaturerecordingPartIIHourlyinterpolationoftemperaturesforclimatologyandmodel‐lingItalian Journal of Agrometeorology1545ndash50
EccelEampCordanoE(2013)InterpolTHourlyinterpolationofmulti‐pletemperaturedailyseriesRpackageversion211RetrievedfromhttpsCRANR‐projectorgpackage=InterpolT
EglingtonSMampPearce‐HigginsJW(2012)Disentanglingtherela‐tiveimportanceofchangesinclimateandland‐useintensityindriv‐ingrecentbirdpopulationtrendsPLoS ONE7e30407httpsdoiorg101371journalpone0030407
Elith JKearneyMampPhillipsS (2010)Theartofmodelling range‐shifting species Methods in Ecology and Evolution 1 330ndash342httpsdoiorg101111j2041‐210X201000036x
FiebergJampJohnsonDH(2015)MMIMultimodelinferenceormod‐elswithmanagement implications Journal of Wildlife Management79708ndash718
FriggensMMampFinchDM (2015) Implicationsof climatechangeforbirdconservationinthesouthwesternUSunderthreealternativefuturesPLoS ONE10e0144089
FryJXianGZJinSDewitzJHomerCGYangLhellipWickhamJD (2011)Completionof the2006national landcoverdatabasefor theconterminousUnitedStatesPhotogrammetric Engineering amp Remote Sensing77858ndash864
Fuller R J Gregory R D Gibbons D W Marchant J HWilson J D Baillie S R amp Carter N (1995) Population de‐clines and range contractions among lowland farmland birdsin Britain Conservation Biology 9 1425ndash1441 httpsdoiorg101046j1523‐1739199509061425x
Gaston K J Blackburn T M Greenwood J J Gregory R DQuinn R M amp Lawton J H (2000) Abundancendashoccupancy
2002emsp |emsp emspensp CLEMENT ET aL
relationships Journal of Applied Ecology 37 39ndash59 httpsdoiorg101046j1365‐2664200000485x
GoldsteinRB(1974)RelationofmetabolismtoambienttemperatureintheverdinCondor76116ndash119httpsdoiorg1023071365995
GrinnellJ(1917)FieldtestsoftheoriesconcerningdistributionalcontrolAmerican Naturalist51115ndash128httpsdoiorg101086279591
GrosboisVGimenezOGaillardJMPradelRBarbraudCClobertJhellipWeimerskirchH(2008)Assessingtheimpactofclimatevari‐ation on survival in vertebrate populationsBiological Reviews 83357ndash399httpsdoiorg101111j1469‐185X200800047x
GustafsonEJ(2013)WhenrelationshipsestimatedinthepastcannotbeusedtopredictthefutureUsingmechanisticmodelstopredictlandscapeecologicaldynamicsinachangingworldLandscape Ecology281429ndash1437httpsdoiorg101007s10980‐013‐9927‐4
Hamel P B (1992) LandManagers guide to the birds of the SouthGeneralTechnicalReportSE‐22USDAForestServiceSoutheasternForestExperimentStationandTheNatureConservancyChapelHillNC
Hinds D S amp Calder W A (1973) Temperature regulation of thePyrrhuloxiaandtheArizonaCardinalPhysiological Zoology4655ndash71httpsdoiorg101086physzool46130152517
HinesJE(2006)PRESENCE Software to estimate patch occupancy and related parameters Laurel MD US Geological survey PatuxentWildlife Research Center Retrieved from httpwwwmbr‐pwrcusgsgovsoftwarepresencehtml
HinesJENicholsJDampCollazoJA(2014)Multiseasonoccupancymodels for correlated replicate surveys Methods in Ecology and Evolution5583ndash591httpsdoiorg1011112041‐210X12186
HinesJENicholsJDRoyleJAMacKenzieDIGopalaswamyAMKumarNampKaranthKU(2010)TigersontrailsOccupancymodeling for cluster sampling Ecological Applications 20 1456ndash1466httpsdoiorg10189009‐03211
HockeyPA SiramiCRidleyARMidgleyGFampBabikerHA(2011) Interrogating recent range changes in South African birdsConfounding signals from land use and climate change present achallenge for attribution Diversity and Distributions 17 254ndash261httpsdoiorg101111j1472‐4642201000741x
Hoegh‐Guldberg O Hughes L McIntyre S Lindenmayer D BParmesanCPossinghamHPampThomasCD (2008)AssistedcolonizationandrapidclimatechangeScience321345ndash346
Homer C Dewitz J Fry J Coan M Hossain N Larson C hellipWickham J (2007) Completion of the 2001 national land coverdatabase for the conterminous United States Photogrammetric Engineering amp Remote Sensing73337
HomerCDewitzJYangLJinSDanielsonPXianGhellipMegownK(2015)Completionofthe2011NationalLandCoverDatabasefortheconterminousUnitedStatesndashrepresentingadecadeoflandcoverchange informationPhotogrammetric Engineering amp Remote Sensing81345ndash354
HootenMBWikleCKDorazioRMampRoyleJA(2007)HierarchicalspatiotemporalmatrixmodelsforcharacterizinginvasionsBiometrics63558ndash567httpsdoiorg101111j1541‐0420200600725x
HosmerDWJrampLemeshowS(2000)Applied logistic regression(2nded)NewYorkNYJohnWileyampSons
Huntley B Collingham Y CGreen R EHiltonGM RahbekCamp Willis S G (2006) Potential impacts of climatic change upongeographical distributions of birds Ibis 148 8ndash28 httpsdoiorg101111j1474‐919X200600523x
IgleciaMNCollazoJAampMcKerrowAJ(2012)Useofoccupancymodels to evaluate expert knowledge‐based species‐habitat rela‐tionshipsAvian Conservation and Ecology7(2)13pRetrievedfromhttpwwwace‐ecoorgvol7iss2art5
IllaacutenJGThomasCDJonesJAWongWKShirleySMampBettsMG(2014)Precipitationandwintertemperaturepredictlong‐termrange‐scale abundancechanges inWesternNorthAmericanbirds
Global Change Biology 20 3351ndash3364 httpsdoiorg101111gcb12642
IoannidisJP (2005)WhymostpublishedresearchfindingsarefalsePLoS Medicine2e124
KeacuteryMGuillera‐ArroitaGampLahoz‐Monfort J J (2013)Analysingand mapping species range dynamics using occupancy modelsJournal of Biogeography 40 1463ndash1474 httpsdoiorg101111jbi12087
Khaliq I Hof C Prinzinger R Boumlhning‐Gaese K amp PfenningerM(2014) Global variation in thermal tolerances and vulnerability ofendotherms to climate change Proceedings of the Royal Society B Biological Sciences 281(1789) 20141097 httpsdoiorg101098rspb20141097
Laursen K amp Frikke J (2008) Hunting from motorboats displacesWadden Sea eiders Somateria mollissima from their favouredfeeding distribution Wildlife Biology 14 423ndash433 httpsdoiorg1029810909‐6396‐144423
LemoineNBauerHGPeintingerMampBoumlhning‐GaeseK (2007)Effectsof climate and land‐use changeon species abundance in acentralEuropeanbirdcommunityConservation Biology21495ndash503httpsdoiorg101111j1523‐1739200600633x
MacArthurRH(1972)Geographical ecology Patterns in the distribution of speciesNewYorkNYHarperampRow
MacKenzieDINicholsJDHinesJEKnutsonMGampFranklinAB(2003)Estimatingsiteoccupancycolonizationandlocalextinc‐tionwhenaspeciesisdetectedimperfectlyEcology842200ndash2207httpsdoiorg10189002‐3090
MacKenzieDINicholsJDLachmanGBDroegeSRoyleJAampLangtimmCA(2002)Estimatingsiteoccupancyrateswhendetec‐tionprobabilitiesarelessthanoneEcology832248ndash2255httpsdoiorg1018900012‐9658(2002)083[2248ESORWD]20CO2
MacKenzieDINicholsJDRoyleJAPollockKHBaileyLampHinesJE(2018)Occupancy estimation and modeling Inferring pat‐terns and dynamics of species occurrence (2nd ed) BurlingtonMAAcademicPress
MellesSJFortinMJLindsayKampBadzinskiD(2011)ExpandingnorthwardInfluenceofclimatechangeforestconnectivityandpop‐ulationprocessesonathreatenedspeciesrangeshiftGlobal Change Biology1717ndash31httpsdoiorg101111j1365‐2486201002214x
Morin X amp Thuiller W (2009) Comparing niche‐and process‐based models to reduce prediction uncertainty in species rangeshifts under climate change Ecology 90 1301ndash1313 httpsdoiorg10189008‐01341
MustinKAmarAampRedpathSM(2014)Colonizationandextinctiondynamicsofadecliningmigratorybirdareinfluencedbyclimateandhabitat degradation Ibis 156 788ndash798 httpsdoiorg101111ibi12173
NicholsJDThomasLampConnPB(2009)InferencesaboutlandbirdabundancefromcountdataRecentadvancesandfuturedirectionsInDLThomsonEJCoochampMJConroy(Eds)Modeling demo‐graphic processes in marked populations(pp201ndash235)NewYorkNYSpringer
OmernikJM(1995)EcoregionsAspatialframeworkforenvironmen‐talmanagement InWSDavisampTSimon(Eds)Biological assess‐ment and criteria Tools for water resource planning and decision making (pp49ndash62)BocaRatonFLLewisPublishers
OmernikJM(2004)Perspectivesonthenatureanddefinitionofeco‐logicalregionsEnvironmental Management34(Supplement1)S27ndashS38httpsdoiorg101007s00267‐003‐5197‐2
Omernik J M amp Griffith G E (2014) Ecoregions of the contermi‐nousUnited States Evolution of a hierarchical spatial frameworkEnvironmental Management541249ndash1266httpsdoiorg101007s00267‐014‐0364‐1
Pardieck K L ZiolkowskiD J JrampHudsonM‐A R (2015)NorthAmericanBreedingBirdSurveyDataset1966ndash2014version20140
emspensp emsp | emsp2003CLEMENT ET aL
USGeologicalSurveyPatuxentWildlifeResearchCenterRetrievedfromhttpswwwpwrcusgsgovbbsrawdata
ParmesanCampYoheG (2003)Agloballycoherentfingerprintofcli‐mate change impacts across natural systemsNature 421 37ndash42httpsdoiorg101038nature01286
PearsonRGampDawsonTP(2003)PredictingtheimpactsofclimatechangeonthedistributionofspeciesArebioclimateenvelopemod‐elsusefulGlobal Ecology and Biogeography12361ndash371
PlattJR(1964)StronginferenceScience146347ndash353PopperK(1959)The logic of scientific discoveryLondonUKHutchinsonRobbins C S Sauer J R Greenberg R S amp Droege S (1989)
Population declines in North American birds that migrate to theNeotropics Proceedings of the National Academy of Sciences 867658ndash7662httpsdoiorg101073pnas86197658
Robinson R A Wilson J D amp Crick H Q (2001) The impor‐tance of arable habitat for farmland birds in grassland land‐scapes Journal of Applied Ecology 38 1059ndash1069 httpsdoiorg101046j1365‐2664200100654x
Romesburg H C (1981) Wildlife science Gaining reliable knowl‐edge Journal of Wildlife Management 45 293ndash313 httpsdoiorg1023073807913
Root T (1988) Energy constraints on avian distributions and abun‐dancesEcology69330ndash339httpsdoiorg1023071940431
Ruiz‐GutieacuterrezVampZipkinEF (2011)Detectionbiasesyieldmis‐leadingpatternsof speciespersistence and colonization in frag‐mented landscapesEcosphere2 1ndash14 httpsdoiorg101890ES10‐002071
SauerJRHinesJEFallonJEPardieckKLZiolkowskiDJJrampLinkWA(2015)The North American breeding bird survey results and analysis 1966ndash2013 Version 01302015LaurelMDUSGSPatuxentWildlifeResearchCenterhttpswwwmbr‐pwrcusgsgovbbsbbshtml
Sauer J R LinkWA Fallon J E Pardieck K L amp ZiolkowskiDJJr(2013)TheNorthAmericanbreedingbirdsurvey1966ndash2011SummaryanalysisandspeciesaccountsNorth American Fauna791ndash32httpsdoiorg103996nafa790001
SeoaneJBustamanteJampDiacuteaz‐DelgadoR(2004)Competingrolesforlandscapevegetationtopographyandclimateinpredictivemod‐elsofbirddistributionEcological Modelling171209ndash222httpsdoiorg101016jecolmodel200308006
Sirami C Caplat P Popy S Clamens A Arlettaz R Jiguet F hellipMartin J‐L (2017) Impacts of global change on species distri‐butions Obstacles and solutions to integrate climate and landuse Global Ecology and Biogeography 26 385ndash394 httpsdoiorg101111geb12555
SkalskiJR(1996)Regressionofabundanceestimatesfrommarkrecap‐ture surveys against environmental covariatesCanadian Journal of
Fisheries and Aquatic Sciences53196ndash204httpsdoiorg101139f95‐169
SohlTL(2014)Therelativeimpactsofclimateandland‐usechangeonconterminousUnitedStatesbird species from2001 to2075PLoS ONE9e112251httpsdoiorg101371journalpone0112251
ThogmartinWESauerJRampKnutsonMG(2007)ModelingandmappingabundanceofAmericanwoodcockacrosstheMidwesternandNortheasternUnitedStatesJournal of Wildlife Management71376ndash382httpsdoiorg1021932005‐680
ThomasCDampLennonJJ (1999)Birdsextendtheirrangesnorth‐wardsNature399213httpsdoiorg10103820335
Trathan P N Garciacutea‐Borboroglu P Boersma D Bost C ACrawford R J CrossinG ThellipWienecke B (2015) Pollutionhabitatlossfishingandclimatechangeascriticalthreatstopen‐guins Conservation Biology 29 31ndash41 httpsdoiorg101111cobi12349
TravisJMJ(2003)Climatechangeandhabitatdestructionadeadlyanthropogenic cocktail Proceedings of the Royal Society of London Series B Biological Sciences 270(1514) 467ndash473 httpsdoiorg101098rspb20022246
UrbanMC (2015)Acceleratingextinction risk fromclimatechangeScience348571ndash573httpsdoiorg101126scienceaaa4984
WalshJWuebblesDHayhoeKKossinJKunkelKStephensGhellipSomervilleR(2014)OurchangingclimateInJMMelilloTCRichmondampGWYohe(Eds)Climate change impacts in the United States The Third National climate assessment(pp19ndash67)WashingtonDCUSGovernmentPrintingOffice
YackulicCBNichols JDReid JampDerR (2015)Topredict thenichemodelcolonizationandextinctionEcology9616ndash23httpsdoiorg10189014‐13611
Yalcin S amp Leroux S J (2018) An empirical test of the relative andcombinedeffectsofland‐coverandclimatechangeonlocalcoloni‐zationandextinctionGlobal Change Biology243849ndash3861httpsdoiorg101111gcb14169
ZhuKWoodallCWampClarkJS(2012)FailuretomigrateLackoftreerangeexpansioninresponsetoclimatechangeGlobal Change Biology181042ndash1052httpsdoiorg101111j1365‐2486201102571x
How to cite this articleClementMJNicholsJDCollazoJATerandoAJHinesJEWilliamsSGPartitioningglobalchangeAssessingtherelativeimportanceofchangesinclimateandlandcoverforchangesinaviandistributionEcol Evol 201991985ndash2003 httpsdoiorg101002ece34890

1988emsp |emsp emspensp CLEMENT ET aL
theCanadianWildlifeServicedesignedtomonitor thestatusandtrendsofbirdsbreedinginNorthAmerica(Saueretal2013)Sinceitsinceptionin1966theBBShasexpandedtoincludeover5000survey routes across theUnited States and southernCanada ap‐proximately3000ofwhichare surveyedeachyearEach route issurveyedonceperyearbyaskilledvolunteerThedateofthesurveyistimedtooccurduringpeakterritorialbehaviortypicallylateMaytoearlyJulydependingon latitudeSurveyorsfollowaprescribedroutealongsecondaryroadsstoppingatapproximately800‐min‐tervalsandperforming3‐minroadsidepointcountsgenerating50countsper394kmrouteWeselectedBBSdatabecausetheirgreatgeographicand temporalextent is suitable for investigatingdistri‐butionalchanges
We selected a subsetof the availableBBSdata for use inouranalysis Our modeling approach described below makes use ofstop‐specificsurveyresultsbutthesedetailedresultsarecurrentlyavailableindigitalformatsonlysince1997Thereforeweuseddatafrom1997to2012BecauseourstudyspeciesarefoundinforestedregionsofeasternNorthAmericawedelineatedthestudyareabyaggregatingthreeOmernikLevelIclassesrepresentingeasternfor‐estecoregionsNorthernForestsEasternTemperateForestsandTropicalWetForests (CommissionforEnvironmentalCooperation1997Omernik19952004OmernikampGriffith2014)ThisstudyareaincludestheUnitedStateseastoftheGreatPlains(29millionkm2) This region encompassed the entire breeding range of theEasternWoodPeweeexceptsomeareasinCanadanotcoveredbyourclimateandhabitatcovariatesWeselectedBBSroutesentirelywithinthisstudyareaforanalysisWeexcludedroutesthatwerenotclassifiedasldquoactiverdquobytheBBSandroutesthatdifferedfromtherecommendedlength(394km)bymorethan25Weonlyanalyzedsurveys conducted under acceptable conditions (ie acceptableweatherdatetimeofdaystopsreportedbyBBSasldquoruntype=1rdquo)OntherareoccasionsthataroutehadmultipleacceptablesurveysperyearweuseddatafromthefirstacceptablesurveyThisyielded1371routesforouranalysisFinallyweconvertedcountsofbirdsateachstopalongeachBBSroutetodetectionnondetectiondatasuitableforpresencendashabsencemodelingThisconversionsacrificeddata richness but allowed us to account for imperfect detectionof species Using the full count data typically requires additionalassumptions (Illaacuten et al 2014) or incorporation of additional datasources(HootenWikleDorazioampRoyle2007)
ClimatedatawereobtainedfromthePRISMproject(Dalyetal2008)ThePRISMoutputuseslocallyweightedregressionmodelstointerpolatelong‐termweatherstationobservationstoa4kmresolu‐tiongridDailyinterpolatedclimatedataareavailablefrom1981tothepresentFromthesedatawecalculatedtwotemperature‐basedindices based on our first hypothesis described above The firstindexwascalculatedasthetotalannualhoursabove38degCwhilethesecondindexwascalculatedasthetotalannualhoursbelow18degC
Becausethetemperature‐basedindiceswerebasedonhoursofexposureitwasnecessarytointerpolatethedailyPRISMdatatoes‐timatehourlyvaluesWeconsideredasimplelinearinterpolationtoimputevaluesbetweendailymaximumandminimumtemperatures
However for extreme temperatures thismethodwould likely re‐sult in estimated exposure durations that are biased low InsteadweadoptedthemethoddescribedbyCesaraccioSpanoDuceandSnyder(2001seeFigure3)andEccel (2010)usingtheRsoftwarepackageldquoInterpolTrdquo (EccelampCordano2013) Inthismethodcan‐didatefunctionsarefit toobservedhourlytemperaturedatafromnearby high‐quality ASOS (Automated Station Observing System)weather stations in the study regionusing a combinationof sinu‐soidalandquadratic functionsForeach4kmPRISMgridcell theresultingcalibrationfunctionfromthenearestASOSstationisthenappliedtoeachdaysmaximumandminimumtemperaturetopredicttheinterveninghourlyvalues
Usingtheinterpolatedhourlytemperatureestimateswecalcu‐latedthetotalannualhourswithinthezonesofheatstressorcoldstress for each year from 1981 to 2012 Annual valueswere cal‐culatedover theperiodJune1ndashMay31 to represent thebreedingcycleOncetheannualresidencetimeswereobtainedweapplieda15‐yearmovingaveragewindowtothedataresultingin16mov‐ingaverageperiodsstartingwiththe198182ndash199596periodandendingwiththe199697ndash201112periodApplyingamovingaver‐ageactsasahigh‐passfilterthatattenuatesinterannualvariabilitywhileamplifyinganylong‐termclimatechangesignalsOtherperiodlengths could have been chosen however we hypothesized that15years represented a reasonable tradeoff between (a) reducingthemovingaveragelengthsoasnottoexceedthelengthofthedailyPRISMdataserieswhichbeginsin1981(b)increasingthemovingaveragelengthsothatclimaticchanges(asopposedtobackgroundclimaticvariation)canbeidentifiedand(c)usingaperiodthatisbio‐logicallymeaningfulinthatwecanreasonablyexpectanylong‐termclimaticchangesinthemovingaverageswouldbeoutsidethehis‐torical rangeofvariabilityand thusshouldaffectcolonizationandextinctionratesofsensitivespecies
We obtained land cover data from the National Land CoverDatabase (NLCD) for 2001 (Homer et al 2007) 2006 (Fry et al2011)and2011(Homeretal2015)WereclassifiedthegivenlandcovertypesintohabitatornonhabitatforeachbirdspeciesbasedonoursecondhypothesisdescribedaboveWecalculatedtheper‐centof landareathatwasasuitablehabitatwithin400mofeachrouteandusedthisasacovariate inouranalysisWeused400mbecausethisishalfthedistancebetweenstopsonBBSroutesForyearswithoutlandcoverdataweusedthelandcovervaluesofthesubsequent NLCD survey Specifically we used 2001 values for1997ndash20012006valuesfor2002ndash2006and2011valuesfor2007ndash2012Weusedaz‐transformationtocenterandscaleallcovariatespriortoanalysis
23emsp|emspAnalysis
OurgoalwastoassesstherelativeimportanceofclimateandlandcovertochangesinthedistributionofRed‐eyedVireosandEasternWoodPeweesWhileourbirddataconsistedofdetectionnonde‐tection data from BBS surveys our parameters of interest weretheprobabilitiesoflocalcolonizationandlocalextinctionasthese
emspensp emsp | emsp1989CLEMENT ET aL
processes drive changes in distribution Our predictor variableswereourmeasuresof climate and land cover suitability for eachspeciesdescribedaboveWeuseddynamic correlated‐detectionoccupancymodels with detection heterogeneity to estimate therelationshipbetweenourpredictorsandbirddistributiondynam‐ics (Clement et al 2016HinesNichols amp Collazo 2014Hinesetal2010)Occupancymodelsareaclassofmodelsthatestimatetheprobabilityofpresencewhileaccountingforimperfectdetec‐tionofspeciesbyanalyzingreplicated(usuallytemporally)surveys(MacKenzieetal20022018)Dynamicoccupancymodelsmodelinterannual changes in occupancy by positing aMarkov processinwhich the probability of occurrence at time t is a function ofthe probability of occurrence at time tminus1 (MacKenzie NicholsHinesKnutsonampFranklin2003)IncontrasttostaticmodelsthisMarkovprocess recognizes that thedistributionof a species is afunctionofpreviousenvironmentalconditionsanddispersalcon‐straintsaswellascurrentenvironmentalconditions(Dormannetal2012KeacuteryGuillera‐ArroitaampLahoz‐Monfort2013)Dynamiccorrelated‐detectionoccupancymodels areamodelextension inwhichspatiallycorrelatedreplicatedsurveysareusedtoestimatedetectionprobabilities (Hinesetal20142010)Weselectedanoccupancymodelingapproachbecause failure toaccount for im‐perfect detection can bias estimates of colonization and extinc‐tionrates(Ruiz‐GutieacuterrezampZipkin2011)Weselectedadynamicapproachbecausedynamicmodelsoffermoreecological realismmoreaccurateprojectionsandgreatergeneralizabilitythanstaticmodels (Clementetal2016Yackulicetal2015)Weusedcor‐related‐detection models because the BBS generates spatiallyreplicatedsurveysandfailuretoaccountforspatialcorrelationofreplicatescanbiasoccupancyestimates(Hinesetal20142010)Finallywe used a finitemixturemodel to account for detectionheterogeneity because of the variation in habitat and focal spe‐ciesabundanceacrossthestudyarea (Clementetal2016)Afi‐nitemixturemodel approximates the heterogeneity of detectionprobabilitiesbypositingthatthepopulationconsistsofamixtureofrouteswhichhaveeitherarelativelyhighdetectionprobabilityorarelativelylowdetectionprobability
AsdetailedaboveBBSdata consistofnumerous routes eachcomposedofbirdobservationsat50 stopsA speciesmaybeab‐sentfromindividualstopswhenitispresentonaroutebutnotviceversaDetectionof a species is taken asproofof presence but anondetectionmayresultfromatrueabsenceorafalseabsenceForclarityweusethetermldquooccupiedrdquotoindicateaspeciesispresentonarouteandthetermldquoavailablerdquotoindicateaspeciesislocallypres‐entataspecificstopatthetimeofthesurvey(NicholsThomasampConn2009)Bythisterminologythedynamiccorrelated‐detectionoccupancymodelincludesthefollowingparameters
ψ=Pr(routeoccupiedduringthefirstseasonofsurveys)θ = Pr (species available at stop | route occupied and species unavailable
at previous stop)θprime= Pr (species available at stop | route occupied and species available
at previous stop)
p1 = Pr (detection at a stop for low‐detection routes | route occupied and species available at stop)
p2 = Pr (detection at a stop for high‐detection routes | route occupied and species available at stop)
π = Pr (probability that a route is a low‐detection route)εt = Pr (route is not occupied in season t + 1 | occupied in season t) andγt = Pr (route is occupied in season t + 1 | not occupied in season t)
Hinesetal(20142010)developedamodellikelihoodthatallowstheseparameterstobemodeledasfunctionsofroute‐andyear‐spe‐cificcovariateswhiledetectionparameterscanalsobeinfluencedbystop‐specificcovariatesParameterscanthenbeestimatedusingmax‐imum‐likelihoodestimationinprogramPRESENCE(Hines2006)
WedevelopedsixspecificmodelsofγandεforeachbirdspeciesWepartitionedspatialandtemporalvariationincovariatesbyparti‐tioningcovariatesintotheaveragevalueovertimeandtheannualdeviationfromthataverageInthiswaytheaveragedcovariatexiincludedspatialvariationonlywhilethedeviationcomponentΔxiyincludedtemporalvariationonlyUsingthesecovariatesourmodelsincluded(a)anullmodelinwhichγandεareconstantthroughtimeandspaceand(b)aglobal(mostgeneral)modelinwhichγandεvarythroughspaceaccordingtomeanclimateandlandcoversuitabilityandthroughtimeusingadummycovariate foreachyearWealsoconsideredintermediatemodelstoassesstheexplanatorypowerofourcovariates(c)γandεvaryspatiallywithmeanclimateandlandcover suitability but notwith time (d) γ and ε vary spatiallywithmeanclimateandlandcoversuitabilityandtemporallywithannualchangesinclimateonly(e)γandεvaryspatiallywithmeanclimateand land cover suitability and temporally with annual changes inlandcoveronlyand(f)γandεvaryspatiallywithmeanclimateandlandcoversuitabilityandtemporallywithannualchangesinclimateandlandcover
Prior to comparing these models we worked to achieve ade‐quatefitfortheothermodelparametersBecauseofthenumberofmodelparametersweconsideredthemsequentiallyInitiallywefittheglobalmodelforγandεandwemodeledθθprimeandψasfunctionsofclimateandlandcovercovariatesInthisinitialmodelweusedafinitemixturemodelonptoaccountforheterogeneitypresumablycausedbydifferencesamongroutesinhabitatobserversbirdabun‐danceandotherfeaturesthatwerenotexplicitlymodeled(Clementetal2016)Wealsomodeledpasa functionofclimateand landcover covariates and year Because bird activity often varieswithtimeofdaywealsomodeleddetectionasaquadratic functionofstopnumberwhichweconsidertobeaproxyfortimeofdayFromthis highly parameterizedmodelwe fit a reducedmodel by elim‐inating the covariatewith the smallest estimate‐to‐standard errorratioWecontinuedtoeliminatecovariatesinthiswayuntilAICin‐creasedandthenselectedthemodelwiththelowestAICWeusedthisparameterreductionprotocolwithψθθprimeandpinturnmain‐tainingthemodelstructureonγandεAfteridentifyingparsimoni‐ousmodelsforψθθprimeandpwefitthesetofsixmodelsforγandεdescribedaboveThesequentialapproachmaynotyieldidenticalresultsasasingleanalysisbutitisapragmaticapproachfordealing
1990emsp |emsp emspensp CLEMENT ET aL
with complexmodels (MacKenzie et al 2018) By beginningwithhighlyparameterizedmodelsandprogressingtowardreducedmod‐elswereducedtheriskofconfoundingtheoccupancyanddetectionprocesses (MacKenzieet al2018)Wealsochecked forpotentialmulti‐collinearityissuesbycalculatingPearsonscorrelationcoeffi‐cientforthedifferentcovariatesusedinthemodelsWetreatedanR2lt05asindicativeofalackofmulti‐collinearity
WealsoevaluatedthegoodnessoffitoftheglobalmodelforeachspeciesusingthenaiumlvecolonizationandextinctionratestocalculatetheteststatisticofaHosmerndashLemeshowtest(HosmerampLemeshow2000) Inthiscontext thenaiumlvecolonizationrate is thenumberofsitesthatchangedfromnondetectionsitesinyeart todetectionsitesinyear t+1while thenaiumlveextinction rate is thenumberof sitesthatchangedfromdetectionsitesinyeart tonondetectionsitesinyeart+1Wedividedtheroutesintodecilesbasedontheexpectedcolonizationandextinctionratesandcomparedthepredictedratesto theobservedrateswithchi‐squaretests (α=005) Ifnecessarywecombineddecilestoensurethatthepredictednumberofdetec‐tionsinadecilewasgt4AlthoughtheHosmerndashLemeshowteststatis‐ticisoftencalculatedfrompredictedpresencevitalratesseemedtobemoreappropriatemeasuresinthiscasebecausedynamicmodelsestimatechangesinpresenceratherthanpresence(Clementetal2016)Wenotethatthisuseofnaiumlveratesfocusesonproductsofmodelparameters(ieγεp ψ ϑ)andthereforewedonottestthefitofthefullyparameterizedmodelWefocusedonnaiumlveratesbe‐causethetruevital rates (ieγandε)cannotbedirectlyobservedwhendetectionisimperfectwhilethenaiumlveratescanbeobserved
We used an analysis of deviance approach (ANODEV egSkalski 1996) to evaluate the relative importance of climate andland cover to changes in bird distributions In this procedure wecompared thedeviance explainedbyour parameter of interest tothedevianceexplainedbythemostfullyparameterizedmodelinthemodelset
Dev(Mnull)isthedevianceofModel3whichaccountsforspatialbutnottemporalvariationincolonizationandextinctionprobabili‐tiesDev(Mfull)isthedevianceofModel2theglobalmodelwithbothspatialandtemporalvariationinγandεWethenevaluatedthedevi‐anceexplainedbyclimateandorlandcoverbyusingmodels45and6 forDev(Mcov)HighervaluesofR
2
Dev indicategreaterexplanatory
powerwith0indicatingnopowerand1indicatingpowerequaltothatofthefullyparameterizedmodelalthoughR2
Devcouldpotentially
exceed1becauseourcovariatemodelsarenotstrictlynestedwithinModel2ThereforewecalculatedandreportR2
Dev forbothclimate
and landcoverThis isnot theonlypossibleapproach toassessingrelativeimportanceofexplanatoryvariablesbutitissoundandhasbeenusedandrecommendedbyothers(Grosboisetal2008)
The general ANODEV approach to partitioning variation ledustofocusonhypothesistestingasameansofassessingtheneedforextramodelparameters (sourcesofvariation)Thesmallsetof
specificapriorihypothesesalsosets thisworkapart fromexplor‐atorystudiesinvestigatingtheadequacyofmanydifferentmodelsForthesereasonsweusedlikelihoodratiotestsasameansofcom‐paringdifferentmodelsalthoughwenotethatuseofmodelselec‐tioncriteria(egAICc)yieldedsimilarinferences
3emsp |emspRESULTS
Ourcovariatesindicatedasmalllossofhabitatandincreasedtem‐peraturesduringthestudyperioddespitetherelativelyshorttimescaleForEasternWoodPeweehabitatchangewasgt1on38ofrouteswiththemeanshareofsuitablehabitatnearroutesde‐cliningfrom416to409ForRed‐eyedVireohabitatchangewasgt1on23of routeswithmeansuitablehabitatdecliningfrom480to476Routetemperatureswererarelyabovetheheatstress thresholdbut theaveragetimeabovethis thresholdper route increased from 05hr per year from 1982 to 1997 to17hrperyearfrom1997to2012Wealsoobservedadeclineinperiodsofcoldstressfrom5450to5291hrperrouteperyearThePearsons correlation coefficient did not indicate anymulti‐collinearityproblemsamongourcovariates(R2lt02inallcases)
31emsp|emspEastern Wood Pewee
ForEasternWoodPeweethebest‐supportedmodelincludedbothclimateandhabitatmeasuresas covariatesofψθ andθprimeWeex‐cludedclimateandhabitatcovariatesfromthedetectionprobabilitymodelbecausesomeofthosemodelsdidnotconvergeThereforethedetectionmodelforeachspeciesincludedyearandstopnum‐ber as covariates As a result the global model included 85 pa‐rameters (Table1)TheHosmerndashLemeshowtest indicatedthattheglobalmodel fit thedataadequatelywithnoevidenceofdiscrep‐ancies between estimated and observed naiumlve colonization rates(χ2=441 df =8 p=082) and naiumlve extinction rates (χ2=1117df =8p=019)
EasternWoodPeweewerewidespreadinthestudyareawithanaverageprobabilityofoccupancyonBBSroutesin1997of090intheglobalmodel(Figure1)Occupancydeclinedslightlyto088by2012mirroringthedeclineincountsreportedbytheBBSInitialoccupancywaspositivelyrelatedtoavailablehabitatandhoursofcoldstress(Table1)Probabilityofdetectionatthefirstindividualstopduring1997waslowat013whileprobabilityofdetectionat the route level was much higher at 095 because birds weretypically available atmany stopswithina routeModel3whichincludedspatialvariationinclimateandhabitatinthecolonizationandextinctionmodelssignificantlyimprovedmodelfitrelativetoModel1(likelihoodratioχ2=4419df =6plt0001)Underthismodelestimatedcolonizationratesin1998rangedamongroutesfrom 008 to 028 (with amean of 018) while extinction ratesvariedfrom001to004(withameanof002Figure2)Model2differedfromModel3inthat itallowedcolonizationandextinc‐tionratestovaryeachyearModel2improvedmodelfitrelative
R2
Dev=Dev(Mnull)minusDev(Mcov)
Dev(Mnull)minusDev(Mfull)
emspensp emsp | emsp1991CLEMENT ET aL
TA B L E 1 emspEstimatedparametersforcorrelated‐detectiondynamicoccupancyModel2relatinghabitatandclimatecovariatestooccupancydynamicsofbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012Parameterestimatesindicatechangeinlogoddsofparametersinresponsetoz‐transformedcovariatesParametersψϑϑprimeγεp1p2andπdefinedintext
Parameters
Eastern Wood Pewee Red‐eyed Vireo
Estimate SE Estimate SE
Ψintercept 2367 0177 4136 0404
Ψhabitat 0556 0163 2315 0300
Ψheatstress minus0011 0198 minus0337 0281
Ψcoldstress 0598 0166 1020 0258
ϑintercept minus1821 0026 minus1385 0010
ϑhabitat minus0055 0026 0516 0013
ϑheatstress minus0094 0030 0017 0008
ϑcoldstress 0172 0040 0258 0013
ϑprimeintercept 2292 0045 0426 0020
ϑprimehabitat 0316 0036 1210 0018
ϑprimeheatstress minus0006 0038 0054 0013
ϑprimecoldstress minus0622 0064 0876 0019
γintercept minus0884 0353 0010 0424
γ 1999 minus0601 0585 minus0150 0532
γ 2000 minus1083 0633 0516 0464
γ 2001 minus0300 0484 0228 0483
γ 2002 minus1113 0577 0262 0511
γ 2003 minus1055 0564 minus0311 0551
γ2004 minus0656 0525 minus0179 0534
γ 2005 minus0677 0533 0468 0485
γ2006 minus1014 0620 minus0217 0579
γ2007 minus0162 0453 minus0093 0548
γ 2008 minus23545 70483373 minus0478 0607
γ 2009 minus0428 0455 minus0253 0566
γ 2010 minus0292 0478 minus0159 0517
γ 2011 minus1136 0607 0012 0490
γ 2012 minus0498 0474 0141 0497
γaveragehabitat 0095 0091 0801 0112
γaverageheatstress minus0024 0067 0005 0062
γaveragecoldstress 0227 0071 0358 0120
εintercept minus3957 0386 minus3767 0242
ε 1999 0365 0482 minus0189 0337
ε 2000 0049 0512 minus0921 0399
ε 2001 minus0266 0548 minus0532 0362
ε 2002 minus0576 0660 minus1156 0398
ε 2003 minus0298 0595 minus0692 0355
ε2004 0113 0509 minus0621 0343
ε 2005 0173 0482 minus1096 0429
ε2006 minus0145 0561 minus1173 0463
ε2007 0451 0458 minus0894 0366
ε 2008 0283 0480 minus0976 0424
ε 2009 0218 0496 minus0898 0381
(Continues)
1992emsp |emsp emspensp CLEMENT ET aL
Parameters
Eastern Wood Pewee Red‐eyed Vireo
Estimate SE Estimate SE
ε 2010 minus0397 0605 minus0348 0304
ε 2011 0643 0455 minus0999 0406
ε 2012 0153 0644 minus0803 0410
εaveragehabitat 0238 0090 minus1670 0122
εaverageheatstress minus0071 0092 minus0075 0074
εaveragecoldstress minus0307 0141 minus0904 0104
p1intercept minus2172 0043 1352 0038
p1 1998 minus0118 0048 minus0127 0058
p1 1999 minus0112 0049 minus0043 0058
p1 2000 minus0166 0050 minus0016 0059
p1 2001 minus0156 0049 0031 0060
p1 2002 minus0179 0049 0076 0061
p1 2003 minus0216 0051 0161 0064
p12004 minus0242 0051 0066 0060
p1 2005 minus0124 0050 0149 0060
p12006 minus0182 0051 0124 0059
p12007 minus0149 0050 0184 0061
p1 2008 minus0202 0052 0192 0063
p1 2009 minus0221 0052 0135 0061
p1 2010 minus0195 0051 0062 0059
p1 2011 minus0122 0051 0203 0062
p1 2012 minus0227 0055 0194 0062
p1stopnumber 0033 0009 minus0055 0011
p1stopnumbersquared minus0069 0010 minus0247 0013
p2intercept minus0921 0018 minus0557 0036
p2 1998 minus0138 0033 minus0106 0050
p2 1999 minus0137 0032 0000 0052
p2 2000 minus0164 0032 minus0027 0051
p2 2001 minus0077 0032 minus0053 0053
p2 2002 minus0101 0032 minus0019 0052
p2 2003 minus0199 0033 0086 0053
p22004 minus0186 0032 0124 0057
p2 2005 minus0146 0032 0076 0051
p22006 minus0155 0032 0128 0054
p22007 minus0153 0032 0120 0054
p2 2008 minus0168 0032 0072 0056
p2 2009 minus0077 0033 0101 0055
p2 2010 minus0114 0033 0085 0060
p2 2011 minus0079 0032 0124 0054
p2 2012 minus0186 0033 0036 0061
p2stopnumber 0045 0007 0165 0010
p2stopnumbersquared minus0105 0008 minus0183 0010
π 0533 0064 minus0505 0065
TA B L E 1 emsp (Continued)
emspensp emsp | emsp1993CLEMENT ET aL
toModel3althoughnotsignificantlyduetothenumberofaddi‐tional parameters (χ2=2679df =28p=053)UnderModel 2estimatedcolonizationratesvariedfrom000in2008to030in1998whileextinctionratesvariedfrom001 in2001ndash2003and2010to004in2011WeusedModels45and6toestimatethepercentof improvedmodel fit (measuredby thechange indevi‐ance)generatedbyModel2thatcouldbeaccountedforbytempo‐ralvariationinclimateandhabitatcovariates(Table2)Temporalvariationinhabitataccountedfor04ofthechangeindeviancewhile temporal variation in climate accounted for 196Model6whichaccountedfortemporalvariationinbothhabitatandcli‐mateindicatedthatin1998colonizationinthesouthwestportionofthestudyareawasabovethelocalaveragewhileextinctioninthesouthwestwasbelowthelocalaverageindicatingarelativelystrongoccupancy trend in thesouthwest in thatyear (Figure3)By2012thispatternhadreversedyieldingarelativelyweakoc‐cupancytrendinthesouthwest(Figure3)NoneofthesemodelsrepresentedasignificantimprovementoverModel3(pgt026)Insomecasesthesignoftheestimatedeffectoftemporalvariationinclimateandhabitatwasoppositeoftheexpectedeffectbutinnoinstancesweretheseestimatessignificant(Table3)
32emsp|emspRed‐eyed Vireo
The best‐supportedmodel for Red‐eyedVireo included the samecovariatesasforEasternWoodPeweeyieldingaglobalmodelwith85parameters (Table1)TheHosmerndashLemeshowtest indicatedfitwasadequate for theRed‐eyedVireoglobalmodel forbothnaiumlvecolonization rates (χ2=540df =4p=025) and naiumlve extinctionrates(χ2=100df =4p=091)
Red‐eyedVireowere similarlywidespread in the study areawith an average probability of occupancy of 091 in the global
model (Figure1)Occupancydecreasedslightlyto090by2012incontrasttothe increase incountsreportedbytheBBS Initialoccupancywas positively related to available habitat and hoursofcoldstress (Table1)Probabilityofdetectionat the first stopwasrelativelyhighat038whileprobabilityofdetectionattheroute levelnearlyperfectat098Model3which includedspa‐tialvariationinclimateandhabitattothecolonizationandextinc‐tionmodelsimprovedmodelfitdramatically(χ2=41884df =6plt0001)relativetothenullmodelInkeepingwiththegreaterchangeindevianceforRed‐eyedVireoModel3estimatedmorespatialvariationinvitalratescomparedtoEasternWoodPeweeIn 1998 estimated colonization rates varied across BBS routesfrom 010 to 088 (mean of 049)while extinction rates rangedfrom000to048(meanof004Figure4)Model2whichallowedcolonization and extinction rates to vary each year improvedmodelfitrelativetoModel3butnotsignificantlyduetothenum‐berofadditionalparameters(χ2=3427df =28p=019)UnderModel2estimatedcolonizationratesvariedfrom039in2008to060 in2000while estimatedextinction rates varied from003in2006to007in1998WeusedModels45and6toestimatethepercentofthedeviancereductioninModel2thatcouldbeac‐countedforbytemporalvariationinclimateandhabitatcovariates(Table2)Wefoundthattemporalvariation inhabitataccountedfor01of the change indeviancewhile climate accounted for56Model 6 which accounted for temporal variation in bothhabitatandclimateindicatedthatin1998bothcolonizationandextinctionwereaboveaverageindicatinghigherturnoverearlyinthestudyperiod(Figure5)By2012bothcolonizationandextinc‐tion rateshaddeclined indicatinggreater stability inoccupancy(Figure 5) None of these models represented a significant im‐provementoverModel3(pgt074)Someoftheestimatedeffects
F I G U R E 1 emspOccupancyprobabilityfor(a)EasternWoodPeweeand(b)Red‐eyedVireointheeasternUnitedStates1997underModel2Occupancyvariesbytheamountofsuitablehabitathoursofheatstressandhoursofcoldstressin1997
1994emsp |emsp emspensp CLEMENT ET aL
oftemporalvariationinclimateandhabitatdifferedfromexpecta‐tionsbutnotsignificantly(Table3)
4emsp |emspDISCUSSION
Wefound that forbothbirdspecies initialoccupancycoloniza‐tionratesandextinctionratescorrelatedwithspatialvariationinclimateandhabitatcovariatesHowevercolonizationandextinc‐tionrateswerenotcorrelatedwithtemporalvariation inclimateand habitat covariates Our results correlating initial occupancywith environmental covariates are broadly speaking consistentwithbiologicalexpectationandaraftofpreviousstudiesthathaveindicatedasignificantrelationshipbetweenspeciesdistributionsandhabitat(RobinsonWilsonampCrick2001ThogmartinSauerampKnutson 2007) climate (Barbet‐Massinamp Jetz 2014BellardBertelsmeier Leadley Thuiller amp Courchamp 2012) or both(Seoane Bustamante amp Diacuteaz‐Delgado 2004 Sohl 2014)Howeverwesuggestthatestimatedrelationshipsbetweenspatialvariationinenvironmentalcovariatesandcolonizationandextinc‐tion rates are of greater ecological interest than relatively phe‐nomenological species distribution models or climate envelopemodels(Clementetal2016)Theestimatedvitalratesofcoloni‐zationandextinctionbringusclosertounderstandingthedynamicprocessunderlying thedistributionof these speciesGivencon‐stantvital rateswecanalsoproject theequilibrium levelofoc‐cupancyatsites ( i
i+i
atsite i)Wecancomparetheseprojected
equilibria tocurrentoccupancytoproject trends for thespeciesunder current environmental conditions We can also use
projectedchangesinkeyenvironmentalvariablestodeveloppre‐dicted rangechanges in the futureWeargue that theseprojec‐tions are more robust than those from static climate envelopemodels which assume that species are currently in equilibrium(Yackulicetal2015)Incontraststaticmodelscanoverestimaterange changes when the equilibrium does not hold (Morin ampThuiller2009)
Given the importanceofglobal changeweweremotivated toinvestigate factors affecting temporal aswell as spatial variationin vital rates In this case spatial variation must be incorporatedintosuchanalysesinordertoaccommodatethedifferentdynamicsexpectedindifferentportionsofaspeciesrangeWeviewthere‐lationshipbetweentemporalvariationincovariatevaluesandvitalrates as the most relevant to understanding the effect of globalchangeontherecentandfuturedistributionsofourstudyspeciesFurthermore a correlation between vital rates and spatially vary‐ingcovariatesmaynot indicateasimilarcorrelationwithtemporalchangesinthosesamecovariatesForexampleaspecieswithstrongdispersal constraintsmight have a strong spatial relationshipwithacovariatebutaweaktemporalrelationshipduetoits immobility(Devictor et al 2008) Similarly an estimated spatial relationshipmaybepurelyphenomenologicalduetothenatureofspatialvari‐ablesWhen spatial variables followgradients it is relatively easytoobtainstatisticallysignificantcorrelationsabsentanycausalre‐lationship(Grosboisetal2008)ForexampleifcolonizationratesandtemperaturesbothfollownorthndashsouthgradientstheymaybespatiallycorrelatedevenifcolonizationisgovernedbysomeotherunidentifiedfactorInthiscasewewouldexpecttheweakcorrela‐tion between temporal changes in temperature and colonization
F I G U R E 2 emspProbabilityof(a)colonizationand(b)extinctionforEasternWoodPeweeintheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1995CLEMENT ET aL
ratestobemoreindicativeofthetruerelationshipthanthestrongcorrelationwithspatialvariationintemperature
BBS surveys indicate different abundance trajectories for ourstudyspecieswithcounts increasingforRed‐eyedVireosandde‐creasing for EasternWood Pewees (Sauer et al 2015) Becauseabundanceandoccupancytendtobelinked(Gastonetal2000)weexpectedsomedivergenceinoccupancydynamicsWefoundthatestimatedoccupancywashighandstableforbothspeciesalthoughtherewasgreaterturnoverforRed‐eyedVireosForexampleunderModel3 (spatialvariationonly incovariates)bothaveragecoloni‐zationrates(049)andaverageextinctionrates(004)wereatleasttwiceashighforRed‐eyedVireoscomparedtoEasternWoodPewee(018and002respectively)Red‐eyedVireosexhibitedgreaterspa‐tialvariationincolonizationandextinctionrates(Figure4)althoughour selected covariates explained little temporal variation for thisspeciesEasternWoodPeweecolonizationandextinctionratesex‐hibitedmoreofalatitudinalgradientreflectingaspatialcorrelationwith extreme temperature (Figure 2)Our results also hinted at agreatercorrelationbetweentemporalchanges in temperatureandoccupancyvitalratesforEasternWoodPeweewhichcouldplayaroleinthedeclineofthisspeciesalthoughtheresultswerenotsig‐nificant(Table3)Alternatively ithasbeensuggestedthatEasternWoodPeweemaybedecliningduetoa lossofforestedwinteringhabitat(RobbinsSauerGreenbergampDroege1989)orduetodeer‐mediatedchanges in foreststructureonbreedinggrounds (deCal‐esta 1994) illustrating the complexity of ascertaining cause andeffectusingobservationalstudiesinthesenaturalsystems
Althoughrelativelyrare(Siramietal2017)otherstudieshaveinvestigated both climate and land cover effects on bird distribu‐tions Inagreementwithour finding that temporalvariation incli‐mate explainedmuchmore of themodel deviance than temporalvariation in habitat climate‐only species distribution models hadbetter cross‐validation support than habitat‐only models for 409Europeanbirdspecies(Barbet‐Massinetal2012)SimilarlymodelsestimatingthenumberofnewlyoccupiedareasbyHoodedWarblers(Wilsonia citrina) inOntariousingclimatedatahadbetter informa‐tioncriterionsupportthanmodelsusingforestdata(MellesFortinLindsay amp Badzinski 2011) In contrast land‐use models outper‐formedclimatemodels for18 farmlandbird species in theUnitedKingdom(EglingtonampPearce‐Higgins2012)Similarlyvegetation‐based species distributionmodels had superior performance thanclimate‐based models for 79 Spanish bird species (Seoane et al2004)andforthreebirdspeciesinNewMexico(FriggensampFinch2015) Two studies focused on colonization and extinction ratesfoundmixed results ForGardenWarblers (Sylvia borin) inBritainvegetationexplainedmorevariationincolonizationwhiletempera‐turesexplainedmorevariationinlocalextinction(MustinAmarampRedpath2014)For122birdspeciesinOntariothebest‐supportedcovariates (land cover or climate change) varied among species(YalcinampLeroux2018)
Thedivergentresultsamongstudiesregardingtheimportanceofclimateandlandcovermaypartlybeduetothebiologyofindividualspecies (Yalcinamp Leroux 2018) For example farmland birdsmayTA
BLE
2emspR2 DevfordifferentmodelsofcolonizationandextinctionprobabilitiesforbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012
Mod
el
Cova
riate
s on
colo
niza
tion
and
extin
ctio
nEa
ster
n W
ood
Pew
eeRe
d‐ey
ed V
ireo
Inte
rcep
tAv
erag
e H
abita
tAv
erag
e Cl
imat
eH
abita
t Dev
iatio
nCl
imat
e D
evia
tion
Year
Eff
ect
Δ minus
2LL
R2 Dev
Δ minus
2LL
R2 Dev
2X
XX
X0
0010
00
0010
0
6X
XX
XX
minus2138
202
minus3230
57
4X
XX
Xminus2154
196
minus3234
56
5X
XX
Xminus2667
04
minus3422
01
3X
XX
minus2679
0minus3427
0
1X
minus7098
NA
minus45311
NA
Not
eNAnotapplicable
1996emsp |emsp emspensp CLEMENT ET aL
beparticularlyaffectedbychangesinagriculturalintensitybecauseoftheirextensiveuseoffarms(EglingtonampPearce‐Higgins2012)Alternativelyithasbeenarguedthatspatialscalemayplayaroleintherelativeimportanceofglobalchangefactorswithclimatemoreimportantatlargescales(PearsonampDawson2003)Wenotethattwostudiesthatfoundalargerroleforclimatebothreliedonlower‐resolution bird surveys (05deg atlas in Barbet‐Massin et al 2012
10km2atlasinMellesetal2011)IntheBBSdatathatweanalyzedsurveyswereconductedevery800mbutwecalculatedoccupancyacross each 394km transect In addition the cited studies useddifferentdata sources anddiverse analysismethods complicatinginterpretationofthedifferingresults
While we estimated that climate explained more model de‐viance than habitat these estimated relationships were not
F I G U R E 3 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionrates((t)minus (t)minus )forEasternWoodPeweeintheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
emspensp emsp | emsp1997CLEMENT ET aL
TA B L E 3 emspEstimatedparametersforcorrelated‐detectiondynamicoccupancyModel6relatinghabitatandclimatecovariatestooccupancydynamicsofbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012Parameterestimatesindicatechangeinlogoddsofparametersinresponsetoz‐transformedcovariatesParametersψϑϑprimeγε p1p2andπdefinedintextForkeyparameters(boldtext)expectedsignsincluded
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
Ψintercept 2367 0177 4087 0381
Ψhabitat 0556 0163 2323 0288
Ψheatstress minus0011 0198 minus0322 0275
Ψcoldstress 0598 0166 1054 0251
ϑintercept minus1821 0026 minus1385 0010
ϑhabitat minus0055 0026 0516 0013
ϑheatstress minus0094 0030 0017 0008
ϑcoldstress 0172 0040 0256 0013
ϑprimeintercept 2292 0045 0426 0020
ϑprimehabitat 0316 0036 1210 0018
ϑprimeheatstress minus0006 0038 0054 0013
ϑprimecoldstress minus0622 0064 0875 0019
γintercept minus0884 0353 0023 0162
γhabitatdeviation + minus0292 0478 minus0240 1768
γheatstressdeviation minus minus1136 0607 0064 0194
γcoldstressdeviation minus minus0498 0474 2398 2253
γaveragehabitat 0095 0091 0788 0111
γaverageheatstress minus0024 0067 0004 0062
γaveragecoldstress 0227 0071 0333 0119
εintercept minus3957 0386 minus4471 0141
εhabitatdeviation minus minus0397 0605 0121 1492
εheatstressdeviation + 0643 0455 minus0034 0182
εcoldstressdeviation + 0153 0644 2042 2072
εaveragehabitat 0238 0090 minus1664 0123
εaverageheatstress minus0071 0092 minus0080 0074
εaveragecoldstress minus0307 0141 minus0939 0104
p1intercept minus2172 0043 1351 0039
p1 1998 minus0118 0048 minus0128 0058
p1 1999 minus0112 0049 minus0043 0058
p1 2000 minus0166 0050 minus0017 0059
p1 2001 minus0156 0049 0031 0061
p1 2002 minus0179 0049 0076 0062
p1 2003 minus0216 0051 0160 0064
p12004 minus0242 0051 0066 0061
p1 2005 minus0124 0050 0149 0061
p12006 minus0182 0051 0124 0060
p12007 minus0149 0050 0183 0062
p1 2008 minus0202 0052 0191 0064
p1 2009 minus0221 0052 0134 0062
p1 2010 minus0195 0051 0062 0060
p1 2011 minus0122 0051 0202 0063
(Continues)
1998emsp |emsp emspensp CLEMENT ET aL
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
p1 2012 minus0227 0055 0193 0063
p1stopnumber 0033 0009 minus0055 0011
p1stopnumbersquared minus0069 0010 minus0247 0013
p2intercept minus0921 0018 minus0555 0037
p2 1998 minus0138 0033 minus0121 0050
p2 1999 minus0137 0032 minus0025 0052
p2 2000 minus0164 0032 minus0027 0051
p2 2001 minus0077 0032 minus0059 0053
p2 2002 minus0101 0032 minus0012 0052
p2 2003 minus0199 0033 0085 0053
p22004 minus0186 0032 0113 0059
p2 2005 minus0146 0032 0082 0052
p22006 minus0155 0032 0135 0054
p22007 minus0153 0032 0124 0055
p2 2008 minus0168 0032 0073 0057
p2 2009 minus0077 0033 0100 0056
p2 2010 minus0114 0033 0071 0062
p2 2011 minus0079 0032 0120 0055
p2 2012 minus0186 0033 0030 0063
p2stopnumber 0045 0007 0165 0010
p2stopnumbersquared minus0105 0008 minus0182 0011
π 0533 0064 minus0505 0065
TA B L E 3 emsp (Continued)
F I G U R E 4 emspProbabilityof(a)colonizationand(b)extinctionforRed‐eyedVireointheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1999CLEMENT ET aL
significantanddidnotexplainalargefractionofvariationThereareseveralpotentialexplanationsforwhywedidnotfindasub‐stantialresponsetochangesinhabitatandclimateFirstourdatawerelackingindynamiceventstoinformourmodelsThecovari‐ateschangedrelativelylittleoverthetimeperiodmakingitdiffi‐culttodetectaresponseThelackofchangewaspartlybecauseoftherelativelyshort(16‐year)timeperiodstudiedAlsoweusedamovingaverageforclimatedatawhichtendstoreducedifferences
overshorttimeperiods Inadditionthestudyregionhasexperi‐encedlesswarmingoverthepastcenturythanotherpartsoftheplanet (Walshetal2014)Thebirddistributionsalso lackeddy‐namismBecauseextinctionrateswerelowtherewerefewlocalextinctioneventstoinformthemodelAlthoughcolonizationrateswerehighthefactthatinitialoccupancywashighandextinctionrateswerelowmeanttherewerefewopportunitiesforcoloniza‐tionevents
F I G U R E 5 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionratesforRed‐eyedVireointheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
2000emsp |emsp emspensp CLEMENT ET aL
Second measurement error may have obscured responses toglobalchangeThePRISMclimatedataareestimatedbyinterpola‐tionratherthandirectmeasurementacrossthestudyarea(Dalyetal2008)AssuchtheremaybeartifactsoranomaliesthatimpededourestimationandincreaseduncertaintySimilarlyNLCDdataareremotelysensedandmay includesomemeasurementerrorwhichcan increase uncertainty in our estimates (Homer et al 2015)Additionallylandcoverwasonlymeasuredinthreeyearsrequiringus to imputevalues in13yearsAsa result all landcover changethatoccurredbetween2002and2006wasrecordedin2002whilechangesbetween2007and2011wererecorded in2007Thisap‐proximation isnot idealbutwasnecessarygiventhe5‐yearreso‐lutionofthedataThislowerresolutionofthelandcoverdatamayhavereducedthesensitivityofthemodel
Third wemay havemisspecified ourmodelWe tried to de‐velopbiologicallymeaningfulcovariatesbutwemightnotunder‐standthesystemaswellaswehopedItispossiblethatbirdvitalrates respond to climateon adifferent time scale (ie not a15‐yearmoving average) or to climateduringdifferent timesof theyearortodifferenttemperaturethresholdsortodifferentclimatecomponents (egprecipitation Illaacutenetal2014) It isalsopossi‐ble that thebirds respond to finerhabitat features than the landcovertypesweusedoratadifferentspatialscale(ienot400mPearsonampDawson2003)Alternatively theremaybe importantregionaldifferencesor interactionsamongcovariatesthatwedidnotcaptureOritispossiblethatchangesinclimateandhabitatintheirbreedingareassimplyhavelittleeffectonvitalratesforthesespecies relative to other factors such as competitionwith otherbirdspeciesorglobalchangeoccurringontheirwinteringgroundsinSouthAmerica(Dormann2007)Ofcoursethesemisspecifica‐tionissuesareinherenttoallobservationalstudiesandnotuniquetothisstudy
Itwould be possible for us to explore some of themisspec‐ification issues just mentioned For example we could rerunthe samemodelsusingnewcovariates representing a varietyoftemperature thresholds and habitat buffer sizes to search for astrongercorrelationExperiencesuggeststhatpersistencewouldeventuallyberewardedwithap‐valuelt005ora lowmodelse‐lection valueHowever one reason to avoid such exploration isthat it tends to lead toType Ierrorsoverfitmodels andbiasedparameterestimates(FiebergampJohnson2015)Furthermorewefind thecontrastbetweensignificant correlations for spatial co‐variatesandnonsignificantcorrelationsfortemporalcovariatesil‐luminatingIntypicalclimateenvelopestudieswewouldestimatethe relationshipbetweencovariatesand thecurrentdistributionofaspeciesmuchlikeourestimateofinitialoccupancy(Dormann2007)BecauseweobtainedsignificantparameterestimateswemightconcludethatwehaveidentifiedcovariatesthatdeterminetherangeofthisspeciesThenwemightusetheseresultstoproj‐ectthefuturerangeofthespeciesundervariousclimatescenariosbased on the (questionable) assumption that the covariatendashspe‐cies relationship will not change as climate and habitat change(Cuddington et al 2013 Elith et al 2010Gustafson 2013) In
this case the insignificant correlationwith the temporal covari‐atessuggeststhatouroccupancycovariatesmerelydescribetherangeofthesebirdswithoutdeterminingtherangeFurthermoreour results suggest that despite significant results in the staticportionof themodelwehaveyet to identify themost relevantcomponents of climate change shaping thedistributionof thesebirds although itmay be difficult to identify these componentsgiven the lackofobservedchangeduringa relatively short timeperiodFinallyourresultssuggestthatprojectionsbasedonthestaticportionofourmodelareunlikelytobereliabledespitetheirstatisticalsignificance
Ifourinterpretationiscorrectitsuggeststhatsomeprojectionsof futurerangeshiftsbasedonstaticmodelsrelyonunsupportedassumptionsandarenot reliableWehavedemonstratedadiffer‐entapproachthatdoesnotrequirethestrongassumptionsofstaticmodelingapproachesOurapproachwouldbemoreapttoidentifyfactorsthatguidespeciesrsquoresponsetoglobalchangeifboththespe‐ciesandthecovariatesexhibitedmoredynamismandifweanalyzedalongertimeperiodthatencompassedgreaterchange
ACKNOWLEDG MENTS
WethankKeithPardieckDaveZiolkowskiJrandthethousandsofhighlyskilledvolunteerswhohavecoordinatedandperformedtheNorthAmericanBreedingBirdSurvey for half a centuryWealsothanktheClimateResearchUnitattheUniversityofEastAngliaformakingvaluableclimatedataeasilyaccessibleMJCwasfinanciallysupported by the BBS and the USGS Northeast Climate ScienceCenterThecontentsofthisdocumentaretheresponsibilityoftheauthorsanddonotnecessarilyrepresenttheviewsoftheUSGSAnyuseoftradeproductorfirmnamesisfordescriptivepurposesonlyanddoesnotimplyendorsementbytheUSGovernment
DEDIC ATION
We dedicate this paper to the late Chandler S Robbins ldquoFatherrdquooftheNorthAmericanBreedingBirdSurvey (BBS)DatafromtheBBSformthebasisforthisspecificpaperandforhundredsofothersfocusedon topics ranging from local trends tocontinentalmacro‐ecologyChanwasauniquecombinationofbird‐loverandscientistwhosevisionandinquisitiveenthusiasmwereaninspirationtoallofusfortunateenoughtohaveknownhim
CONFLIC T OF INTERE S T
Nonedeclared
AUTHORSrsquo CONTRIBUTION
JACandJDNconceivedthestudyAJTandSWanalyzedhabitatandclimatedataMJCJEHandJDNperformedtheoccupancyanaly‐sisMJCledthewritingofthemanuscriptAllauthorsgavefinalap‐provalforpublication
emspensp emsp | emsp2001CLEMENT ET aL
DATA ACCE SSIBILIT Y
Raw data are available from the following sources Breeding BirdSurvey httpswwwpwrcusgsgovbbsrawdata PRISM climatedata httpwwwprismoregonstateedu NLCD land cover datahttpswwwmrlcgov Processed data will be archived with theDryadDigitalRepository
ORCID
Matthew J Clement httpsorcidorg0000‐0003‐4231‐7949
James D Nichols httpsorcidorg0000‐0002‐7631‐2890
Adam J Terando httpsorcidorg0000‐0002‐9280‐043X
James E Hines httpsorcidorg0000‐0001‐5478‐7230
Steven G Williams httpsorcidorg0000‐0003‐3760‐6818
R E FE R E N C E S
Anderson D R Burnham K P Gould W R amp Cherry S (2001)Concerns about finding effects that are actually spuriousWildlife Society Bulletin29311ndash316
Arauacutejo M B Cabeza M Thuiller W Hannah L amp WilliamsP H (2004) Would climate change drive species out of re‐serves An assessment of existing reserve‐selection meth‐ods Global Change Biology 10 1618ndash1626 httpsdoiorg101111j1365‐2486200400828x
Barbet‐MassinMampJetzW(2014)A40‐yearcontinent‐widemulti‐speciesassessmentofrelevantclimatepredictorsforspeciesdistri‐butionmodellingDiversity and Distributions201285ndash1295httpsdoiorg101111ddi12229
Barbet‐Massin M Thuiller W amp Jiguet F (2012) The fate ofEuropean breeding birds under climate land‐use and disper‐sal scenarios Global Change Biology 18 881ndash890 httpsdoiorg101111j1365‐2486201102552x
Bellard C Bertelsmeier C Leadley P Thuiller W ampCourchamp F (2012) Impacts of climate change on the fu‐ture of biodiversity Ecology Letters 15 365ndash377 httpsdoiorg101111j1461‐0248201101736x
Boumlhning‐Gaese K amp Bauer H G (1996) Changes in species abun‐dance distribution and diversity in a central European birdcommunity Conservation Biology 10 175ndash187 httpsdoiorg101046j1523‐1739199610010175x
BrommerJELehikoinenAampValkamaJ(2012)Thebreedingrangesof central European and Arctic bird speciesmove poleward PLoS ONE7e43648httpsdoiorg101371journalpone0043648
CalderWAampKingJR(1974)ThermalandcaloricrelationsofbirdsInDSFarnerampJRKing(Eds)Avian biology(Vol4pp259ndash413)NewYorkNYAcademicPress
CesaraccioCSpanoDDucePampSnyderRL(2001)Animprovedmodel for determining degree‐day values from daily temperaturedataInternational Journal of Biometeorology45161ndash169httpsdoiorg101007s004840100104
ChamberlinTC (1890)ThemethodofmultipleworkinghypothesesScience1592ndash96
ClementMJHinesJENicholsJDPardieckKLampZiolkowskiDJJr (2016)Estimating indicesofrangeshifts inbirdsusingdy‐namicmodelswhendetectionisimperfectGlobal Change Biology223273ndash3285httpsdoiorg101111gcb13283
Commission for Environmental Cooperation (1997) Ecological regionsofNorthAmericaTowardacommonperspectiveCommission for
EnvironmentalCooperationMontrealRetrievedfromftpftpepagovwedecoregionscec_naCEC_NAecopdf
Crowl TA Crist TO Parmenter R R BelovskyGamp LugoA E(2008)Thespreadofinvasivespeciesandinfectiousdiseaseasdriv‐ersofecosystemchangeFrontiers in Ecology and the Environment6238ndash246httpsdoiorg101890070151
Cuddington K FortinM J Gerber L RHastings A Liebhold AOConnorMampRayC(2013)Process‐basedmodelsarerequiredtomanageecologicalsystemsinachangingworldEcosphere41ndash12httpsdoiorg101890ES12‐001781
DalyCHalbleibMSmithJIGibsonWPDoggettMKTaylorGHhellipPasterisPP(2008)Physiographicallysensitivemappingofcli‐matologicaltemperatureandprecipitationacrosstheconterminousUnited States International Journal of Climatology 28 2031ndash2064httpsdoiorg101002joc1688
DawsonW (1958) Relation of oxygen consumption and evaporativewater losstotemperatureintheCardinalPhysiological Zoology3137ndash48httpsdoiorg101086physzool31130155377
DawsonWRampWhittowGC (2000)Regulationofbodytempera‐ture InG CWhittow (Ed)Sturkiersquos avian physiology (5th ed pp343ndash390)SanDiegoCAAcademicPress
DeCalestaDS(1994)Effectofwhite‐taileddeeronsongbirdswithinmanagedforestsinPennsylvaniaJournal of Wildlife Management58711ndash718httpsdoiorg1023073809685
DevictorVJulliardRCouvetDampJiguetF(2008)Birdsaretrack‐ing climatewarming but not fast enoughProceedings of the Royal Society of London B Biological Sciences2752743ndash2748httpsdoiorg101098rspb20080878
Dormann C F (2007) Promising the future Global change projec‐tionsofspeciesdistributionsBasic and Applied Ecology8387ndash397httpsdoiorg101016jbaae200611001
Dormann C F Schymanski S J Cabral J Chuine I Graham CHartigFhellipSingerA(2012)Correlationandprocessinspeciesdis‐tributionmodelsBridgingadichotomyJournal of Biogeography392119ndash2131httpsdoiorg101111j1365‐2699201102659x
EccelE(2010)WhatwecanasktohourlytemperaturerecordingPartIIHourlyinterpolationoftemperaturesforclimatologyandmodel‐lingItalian Journal of Agrometeorology1545ndash50
EccelEampCordanoE(2013)InterpolTHourlyinterpolationofmulti‐pletemperaturedailyseriesRpackageversion211RetrievedfromhttpsCRANR‐projectorgpackage=InterpolT
EglingtonSMampPearce‐HigginsJW(2012)Disentanglingtherela‐tiveimportanceofchangesinclimateandland‐useintensityindriv‐ingrecentbirdpopulationtrendsPLoS ONE7e30407httpsdoiorg101371journalpone0030407
Elith JKearneyMampPhillipsS (2010)Theartofmodelling range‐shifting species Methods in Ecology and Evolution 1 330ndash342httpsdoiorg101111j2041‐210X201000036x
FiebergJampJohnsonDH(2015)MMIMultimodelinferenceormod‐elswithmanagement implications Journal of Wildlife Management79708ndash718
FriggensMMampFinchDM (2015) Implicationsof climatechangeforbirdconservationinthesouthwesternUSunderthreealternativefuturesPLoS ONE10e0144089
FryJXianGZJinSDewitzJHomerCGYangLhellipWickhamJD (2011)Completionof the2006national landcoverdatabasefor theconterminousUnitedStatesPhotogrammetric Engineering amp Remote Sensing77858ndash864
Fuller R J Gregory R D Gibbons D W Marchant J HWilson J D Baillie S R amp Carter N (1995) Population de‐clines and range contractions among lowland farmland birdsin Britain Conservation Biology 9 1425ndash1441 httpsdoiorg101046j1523‐1739199509061425x
Gaston K J Blackburn T M Greenwood J J Gregory R DQuinn R M amp Lawton J H (2000) Abundancendashoccupancy
2002emsp |emsp emspensp CLEMENT ET aL
relationships Journal of Applied Ecology 37 39ndash59 httpsdoiorg101046j1365‐2664200000485x
GoldsteinRB(1974)RelationofmetabolismtoambienttemperatureintheverdinCondor76116ndash119httpsdoiorg1023071365995
GrinnellJ(1917)FieldtestsoftheoriesconcerningdistributionalcontrolAmerican Naturalist51115ndash128httpsdoiorg101086279591
GrosboisVGimenezOGaillardJMPradelRBarbraudCClobertJhellipWeimerskirchH(2008)Assessingtheimpactofclimatevari‐ation on survival in vertebrate populationsBiological Reviews 83357ndash399httpsdoiorg101111j1469‐185X200800047x
GustafsonEJ(2013)WhenrelationshipsestimatedinthepastcannotbeusedtopredictthefutureUsingmechanisticmodelstopredictlandscapeecologicaldynamicsinachangingworldLandscape Ecology281429ndash1437httpsdoiorg101007s10980‐013‐9927‐4
Hamel P B (1992) LandManagers guide to the birds of the SouthGeneralTechnicalReportSE‐22USDAForestServiceSoutheasternForestExperimentStationandTheNatureConservancyChapelHillNC
Hinds D S amp Calder W A (1973) Temperature regulation of thePyrrhuloxiaandtheArizonaCardinalPhysiological Zoology4655ndash71httpsdoiorg101086physzool46130152517
HinesJE(2006)PRESENCE Software to estimate patch occupancy and related parameters Laurel MD US Geological survey PatuxentWildlife Research Center Retrieved from httpwwwmbr‐pwrcusgsgovsoftwarepresencehtml
HinesJENicholsJDampCollazoJA(2014)Multiseasonoccupancymodels for correlated replicate surveys Methods in Ecology and Evolution5583ndash591httpsdoiorg1011112041‐210X12186
HinesJENicholsJDRoyleJAMacKenzieDIGopalaswamyAMKumarNampKaranthKU(2010)TigersontrailsOccupancymodeling for cluster sampling Ecological Applications 20 1456ndash1466httpsdoiorg10189009‐03211
HockeyPA SiramiCRidleyARMidgleyGFampBabikerHA(2011) Interrogating recent range changes in South African birdsConfounding signals from land use and climate change present achallenge for attribution Diversity and Distributions 17 254ndash261httpsdoiorg101111j1472‐4642201000741x
Hoegh‐Guldberg O Hughes L McIntyre S Lindenmayer D BParmesanCPossinghamHPampThomasCD (2008)AssistedcolonizationandrapidclimatechangeScience321345ndash346
Homer C Dewitz J Fry J Coan M Hossain N Larson C hellipWickham J (2007) Completion of the 2001 national land coverdatabase for the conterminous United States Photogrammetric Engineering amp Remote Sensing73337
HomerCDewitzJYangLJinSDanielsonPXianGhellipMegownK(2015)Completionofthe2011NationalLandCoverDatabasefortheconterminousUnitedStatesndashrepresentingadecadeoflandcoverchange informationPhotogrammetric Engineering amp Remote Sensing81345ndash354
HootenMBWikleCKDorazioRMampRoyleJA(2007)HierarchicalspatiotemporalmatrixmodelsforcharacterizinginvasionsBiometrics63558ndash567httpsdoiorg101111j1541‐0420200600725x
HosmerDWJrampLemeshowS(2000)Applied logistic regression(2nded)NewYorkNYJohnWileyampSons
Huntley B Collingham Y CGreen R EHiltonGM RahbekCamp Willis S G (2006) Potential impacts of climatic change upongeographical distributions of birds Ibis 148 8ndash28 httpsdoiorg101111j1474‐919X200600523x
IgleciaMNCollazoJAampMcKerrowAJ(2012)Useofoccupancymodels to evaluate expert knowledge‐based species‐habitat rela‐tionshipsAvian Conservation and Ecology7(2)13pRetrievedfromhttpwwwace‐ecoorgvol7iss2art5
IllaacutenJGThomasCDJonesJAWongWKShirleySMampBettsMG(2014)Precipitationandwintertemperaturepredictlong‐termrange‐scale abundancechanges inWesternNorthAmericanbirds
Global Change Biology 20 3351ndash3364 httpsdoiorg101111gcb12642
IoannidisJP (2005)WhymostpublishedresearchfindingsarefalsePLoS Medicine2e124
KeacuteryMGuillera‐ArroitaGampLahoz‐Monfort J J (2013)Analysingand mapping species range dynamics using occupancy modelsJournal of Biogeography 40 1463ndash1474 httpsdoiorg101111jbi12087
Khaliq I Hof C Prinzinger R Boumlhning‐Gaese K amp PfenningerM(2014) Global variation in thermal tolerances and vulnerability ofendotherms to climate change Proceedings of the Royal Society B Biological Sciences 281(1789) 20141097 httpsdoiorg101098rspb20141097
Laursen K amp Frikke J (2008) Hunting from motorboats displacesWadden Sea eiders Somateria mollissima from their favouredfeeding distribution Wildlife Biology 14 423ndash433 httpsdoiorg1029810909‐6396‐144423
LemoineNBauerHGPeintingerMampBoumlhning‐GaeseK (2007)Effectsof climate and land‐use changeon species abundance in acentralEuropeanbirdcommunityConservation Biology21495ndash503httpsdoiorg101111j1523‐1739200600633x
MacArthurRH(1972)Geographical ecology Patterns in the distribution of speciesNewYorkNYHarperampRow
MacKenzieDINicholsJDHinesJEKnutsonMGampFranklinAB(2003)Estimatingsiteoccupancycolonizationandlocalextinc‐tionwhenaspeciesisdetectedimperfectlyEcology842200ndash2207httpsdoiorg10189002‐3090
MacKenzieDINicholsJDLachmanGBDroegeSRoyleJAampLangtimmCA(2002)Estimatingsiteoccupancyrateswhendetec‐tionprobabilitiesarelessthanoneEcology832248ndash2255httpsdoiorg1018900012‐9658(2002)083[2248ESORWD]20CO2
MacKenzieDINicholsJDRoyleJAPollockKHBaileyLampHinesJE(2018)Occupancy estimation and modeling Inferring pat‐terns and dynamics of species occurrence (2nd ed) BurlingtonMAAcademicPress
MellesSJFortinMJLindsayKampBadzinskiD(2011)ExpandingnorthwardInfluenceofclimatechangeforestconnectivityandpop‐ulationprocessesonathreatenedspeciesrangeshiftGlobal Change Biology1717ndash31httpsdoiorg101111j1365‐2486201002214x
Morin X amp Thuiller W (2009) Comparing niche‐and process‐based models to reduce prediction uncertainty in species rangeshifts under climate change Ecology 90 1301ndash1313 httpsdoiorg10189008‐01341
MustinKAmarAampRedpathSM(2014)Colonizationandextinctiondynamicsofadecliningmigratorybirdareinfluencedbyclimateandhabitat degradation Ibis 156 788ndash798 httpsdoiorg101111ibi12173
NicholsJDThomasLampConnPB(2009)InferencesaboutlandbirdabundancefromcountdataRecentadvancesandfuturedirectionsInDLThomsonEJCoochampMJConroy(Eds)Modeling demo‐graphic processes in marked populations(pp201ndash235)NewYorkNYSpringer
OmernikJM(1995)EcoregionsAspatialframeworkforenvironmen‐talmanagement InWSDavisampTSimon(Eds)Biological assess‐ment and criteria Tools for water resource planning and decision making (pp49ndash62)BocaRatonFLLewisPublishers
OmernikJM(2004)Perspectivesonthenatureanddefinitionofeco‐logicalregionsEnvironmental Management34(Supplement1)S27ndashS38httpsdoiorg101007s00267‐003‐5197‐2
Omernik J M amp Griffith G E (2014) Ecoregions of the contermi‐nousUnited States Evolution of a hierarchical spatial frameworkEnvironmental Management541249ndash1266httpsdoiorg101007s00267‐014‐0364‐1
Pardieck K L ZiolkowskiD J JrampHudsonM‐A R (2015)NorthAmericanBreedingBirdSurveyDataset1966ndash2014version20140
emspensp emsp | emsp2003CLEMENT ET aL
USGeologicalSurveyPatuxentWildlifeResearchCenterRetrievedfromhttpswwwpwrcusgsgovbbsrawdata
ParmesanCampYoheG (2003)Agloballycoherentfingerprintofcli‐mate change impacts across natural systemsNature 421 37ndash42httpsdoiorg101038nature01286
PearsonRGampDawsonTP(2003)PredictingtheimpactsofclimatechangeonthedistributionofspeciesArebioclimateenvelopemod‐elsusefulGlobal Ecology and Biogeography12361ndash371
PlattJR(1964)StronginferenceScience146347ndash353PopperK(1959)The logic of scientific discoveryLondonUKHutchinsonRobbins C S Sauer J R Greenberg R S amp Droege S (1989)
Population declines in North American birds that migrate to theNeotropics Proceedings of the National Academy of Sciences 867658ndash7662httpsdoiorg101073pnas86197658
Robinson R A Wilson J D amp Crick H Q (2001) The impor‐tance of arable habitat for farmland birds in grassland land‐scapes Journal of Applied Ecology 38 1059ndash1069 httpsdoiorg101046j1365‐2664200100654x
Romesburg H C (1981) Wildlife science Gaining reliable knowl‐edge Journal of Wildlife Management 45 293ndash313 httpsdoiorg1023073807913
Root T (1988) Energy constraints on avian distributions and abun‐dancesEcology69330ndash339httpsdoiorg1023071940431
Ruiz‐GutieacuterrezVampZipkinEF (2011)Detectionbiasesyieldmis‐leadingpatternsof speciespersistence and colonization in frag‐mented landscapesEcosphere2 1ndash14 httpsdoiorg101890ES10‐002071
SauerJRHinesJEFallonJEPardieckKLZiolkowskiDJJrampLinkWA(2015)The North American breeding bird survey results and analysis 1966ndash2013 Version 01302015LaurelMDUSGSPatuxentWildlifeResearchCenterhttpswwwmbr‐pwrcusgsgovbbsbbshtml
Sauer J R LinkWA Fallon J E Pardieck K L amp ZiolkowskiDJJr(2013)TheNorthAmericanbreedingbirdsurvey1966ndash2011SummaryanalysisandspeciesaccountsNorth American Fauna791ndash32httpsdoiorg103996nafa790001
SeoaneJBustamanteJampDiacuteaz‐DelgadoR(2004)Competingrolesforlandscapevegetationtopographyandclimateinpredictivemod‐elsofbirddistributionEcological Modelling171209ndash222httpsdoiorg101016jecolmodel200308006
Sirami C Caplat P Popy S Clamens A Arlettaz R Jiguet F hellipMartin J‐L (2017) Impacts of global change on species distri‐butions Obstacles and solutions to integrate climate and landuse Global Ecology and Biogeography 26 385ndash394 httpsdoiorg101111geb12555
SkalskiJR(1996)Regressionofabundanceestimatesfrommarkrecap‐ture surveys against environmental covariatesCanadian Journal of
Fisheries and Aquatic Sciences53196ndash204httpsdoiorg101139f95‐169
SohlTL(2014)Therelativeimpactsofclimateandland‐usechangeonconterminousUnitedStatesbird species from2001 to2075PLoS ONE9e112251httpsdoiorg101371journalpone0112251
ThogmartinWESauerJRampKnutsonMG(2007)ModelingandmappingabundanceofAmericanwoodcockacrosstheMidwesternandNortheasternUnitedStatesJournal of Wildlife Management71376ndash382httpsdoiorg1021932005‐680
ThomasCDampLennonJJ (1999)Birdsextendtheirrangesnorth‐wardsNature399213httpsdoiorg10103820335
Trathan P N Garciacutea‐Borboroglu P Boersma D Bost C ACrawford R J CrossinG ThellipWienecke B (2015) Pollutionhabitatlossfishingandclimatechangeascriticalthreatstopen‐guins Conservation Biology 29 31ndash41 httpsdoiorg101111cobi12349
TravisJMJ(2003)Climatechangeandhabitatdestructionadeadlyanthropogenic cocktail Proceedings of the Royal Society of London Series B Biological Sciences 270(1514) 467ndash473 httpsdoiorg101098rspb20022246
UrbanMC (2015)Acceleratingextinction risk fromclimatechangeScience348571ndash573httpsdoiorg101126scienceaaa4984
WalshJWuebblesDHayhoeKKossinJKunkelKStephensGhellipSomervilleR(2014)OurchangingclimateInJMMelilloTCRichmondampGWYohe(Eds)Climate change impacts in the United States The Third National climate assessment(pp19ndash67)WashingtonDCUSGovernmentPrintingOffice
YackulicCBNichols JDReid JampDerR (2015)Topredict thenichemodelcolonizationandextinctionEcology9616ndash23httpsdoiorg10189014‐13611
Yalcin S amp Leroux S J (2018) An empirical test of the relative andcombinedeffectsofland‐coverandclimatechangeonlocalcoloni‐zationandextinctionGlobal Change Biology243849ndash3861httpsdoiorg101111gcb14169
ZhuKWoodallCWampClarkJS(2012)FailuretomigrateLackoftreerangeexpansioninresponsetoclimatechangeGlobal Change Biology181042ndash1052httpsdoiorg101111j1365‐2486201102571x
How to cite this articleClementMJNicholsJDCollazoJATerandoAJHinesJEWilliamsSGPartitioningglobalchangeAssessingtherelativeimportanceofchangesinclimateandlandcoverforchangesinaviandistributionEcol Evol 201991985ndash2003 httpsdoiorg101002ece34890

emspensp emsp | emsp1989CLEMENT ET aL
processes drive changes in distribution Our predictor variableswereourmeasuresof climate and land cover suitability for eachspeciesdescribedaboveWeuseddynamic correlated‐detectionoccupancymodels with detection heterogeneity to estimate therelationshipbetweenourpredictorsandbirddistributiondynam‐ics (Clement et al 2016HinesNichols amp Collazo 2014Hinesetal2010)Occupancymodelsareaclassofmodelsthatestimatetheprobabilityofpresencewhileaccountingforimperfectdetec‐tionofspeciesbyanalyzingreplicated(usuallytemporally)surveys(MacKenzieetal20022018)Dynamicoccupancymodelsmodelinterannual changes in occupancy by positing aMarkov processinwhich the probability of occurrence at time t is a function ofthe probability of occurrence at time tminus1 (MacKenzie NicholsHinesKnutsonampFranklin2003)IncontrasttostaticmodelsthisMarkovprocess recognizes that thedistributionof a species is afunctionofpreviousenvironmentalconditionsanddispersalcon‐straintsaswellascurrentenvironmentalconditions(Dormannetal2012KeacuteryGuillera‐ArroitaampLahoz‐Monfort2013)Dynamiccorrelated‐detectionoccupancymodels areamodelextension inwhichspatiallycorrelatedreplicatedsurveysareusedtoestimatedetectionprobabilities (Hinesetal20142010)Weselectedanoccupancymodelingapproachbecause failure toaccount for im‐perfect detection can bias estimates of colonization and extinc‐tionrates(Ruiz‐GutieacuterrezampZipkin2011)Weselectedadynamicapproachbecausedynamicmodelsoffermoreecological realismmoreaccurateprojectionsandgreatergeneralizabilitythanstaticmodels (Clementetal2016Yackulicetal2015)Weusedcor‐related‐detection models because the BBS generates spatiallyreplicatedsurveysandfailuretoaccountforspatialcorrelationofreplicatescanbiasoccupancyestimates(Hinesetal20142010)Finallywe used a finitemixturemodel to account for detectionheterogeneity because of the variation in habitat and focal spe‐ciesabundanceacrossthestudyarea (Clementetal2016)Afi‐nitemixturemodel approximates the heterogeneity of detectionprobabilitiesbypositingthatthepopulationconsistsofamixtureofrouteswhichhaveeitherarelativelyhighdetectionprobabilityorarelativelylowdetectionprobability
AsdetailedaboveBBSdata consistofnumerous routes eachcomposedofbirdobservationsat50 stopsA speciesmaybeab‐sentfromindividualstopswhenitispresentonaroutebutnotviceversaDetectionof a species is taken asproofof presence but anondetectionmayresultfromatrueabsenceorafalseabsenceForclarityweusethetermldquooccupiedrdquotoindicateaspeciesispresentonarouteandthetermldquoavailablerdquotoindicateaspeciesislocallypres‐entataspecificstopatthetimeofthesurvey(NicholsThomasampConn2009)Bythisterminologythedynamiccorrelated‐detectionoccupancymodelincludesthefollowingparameters
ψ=Pr(routeoccupiedduringthefirstseasonofsurveys)θ = Pr (species available at stop | route occupied and species unavailable
at previous stop)θprime= Pr (species available at stop | route occupied and species available
at previous stop)
p1 = Pr (detection at a stop for low‐detection routes | route occupied and species available at stop)
p2 = Pr (detection at a stop for high‐detection routes | route occupied and species available at stop)
π = Pr (probability that a route is a low‐detection route)εt = Pr (route is not occupied in season t + 1 | occupied in season t) andγt = Pr (route is occupied in season t + 1 | not occupied in season t)
Hinesetal(20142010)developedamodellikelihoodthatallowstheseparameterstobemodeledasfunctionsofroute‐andyear‐spe‐cificcovariateswhiledetectionparameterscanalsobeinfluencedbystop‐specificcovariatesParameterscanthenbeestimatedusingmax‐imum‐likelihoodestimationinprogramPRESENCE(Hines2006)
WedevelopedsixspecificmodelsofγandεforeachbirdspeciesWepartitionedspatialandtemporalvariationincovariatesbyparti‐tioningcovariatesintotheaveragevalueovertimeandtheannualdeviationfromthataverageInthiswaytheaveragedcovariatexiincludedspatialvariationonlywhilethedeviationcomponentΔxiyincludedtemporalvariationonlyUsingthesecovariatesourmodelsincluded(a)anullmodelinwhichγandεareconstantthroughtimeandspaceand(b)aglobal(mostgeneral)modelinwhichγandεvarythroughspaceaccordingtomeanclimateandlandcoversuitabilityandthroughtimeusingadummycovariate foreachyearWealsoconsideredintermediatemodelstoassesstheexplanatorypowerofourcovariates(c)γandεvaryspatiallywithmeanclimateandlandcover suitability but notwith time (d) γ and ε vary spatiallywithmeanclimateandlandcoversuitabilityandtemporallywithannualchangesinclimateonly(e)γandεvaryspatiallywithmeanclimateand land cover suitability and temporally with annual changes inlandcoveronlyand(f)γandεvaryspatiallywithmeanclimateandlandcoversuitabilityandtemporallywithannualchangesinclimateandlandcover
Prior to comparing these models we worked to achieve ade‐quatefitfortheothermodelparametersBecauseofthenumberofmodelparametersweconsideredthemsequentiallyInitiallywefittheglobalmodelforγandεandwemodeledθθprimeandψasfunctionsofclimateandlandcovercovariatesInthisinitialmodelweusedafinitemixturemodelonptoaccountforheterogeneitypresumablycausedbydifferencesamongroutesinhabitatobserversbirdabun‐danceandotherfeaturesthatwerenotexplicitlymodeled(Clementetal2016)Wealsomodeledpasa functionofclimateand landcover covariates and year Because bird activity often varieswithtimeofdaywealsomodeleddetectionasaquadratic functionofstopnumberwhichweconsidertobeaproxyfortimeofdayFromthis highly parameterizedmodelwe fit a reducedmodel by elim‐inating the covariatewith the smallest estimate‐to‐standard errorratioWecontinuedtoeliminatecovariatesinthiswayuntilAICin‐creasedandthenselectedthemodelwiththelowestAICWeusedthisparameterreductionprotocolwithψθθprimeandpinturnmain‐tainingthemodelstructureonγandεAfteridentifyingparsimoni‐ousmodelsforψθθprimeandpwefitthesetofsixmodelsforγandεdescribedaboveThesequentialapproachmaynotyieldidenticalresultsasasingleanalysisbutitisapragmaticapproachfordealing
1990emsp |emsp emspensp CLEMENT ET aL
with complexmodels (MacKenzie et al 2018) By beginningwithhighlyparameterizedmodelsandprogressingtowardreducedmod‐elswereducedtheriskofconfoundingtheoccupancyanddetectionprocesses (MacKenzieet al2018)Wealsochecked forpotentialmulti‐collinearityissuesbycalculatingPearsonscorrelationcoeffi‐cientforthedifferentcovariatesusedinthemodelsWetreatedanR2lt05asindicativeofalackofmulti‐collinearity
WealsoevaluatedthegoodnessoffitoftheglobalmodelforeachspeciesusingthenaiumlvecolonizationandextinctionratestocalculatetheteststatisticofaHosmerndashLemeshowtest(HosmerampLemeshow2000) Inthiscontext thenaiumlvecolonizationrate is thenumberofsitesthatchangedfromnondetectionsitesinyeart todetectionsitesinyear t+1while thenaiumlveextinction rate is thenumberof sitesthatchangedfromdetectionsitesinyeart tonondetectionsitesinyeart+1Wedividedtheroutesintodecilesbasedontheexpectedcolonizationandextinctionratesandcomparedthepredictedratesto theobservedrateswithchi‐squaretests (α=005) Ifnecessarywecombineddecilestoensurethatthepredictednumberofdetec‐tionsinadecilewasgt4AlthoughtheHosmerndashLemeshowteststatis‐ticisoftencalculatedfrompredictedpresencevitalratesseemedtobemoreappropriatemeasuresinthiscasebecausedynamicmodelsestimatechangesinpresenceratherthanpresence(Clementetal2016)Wenotethatthisuseofnaiumlveratesfocusesonproductsofmodelparameters(ieγεp ψ ϑ)andthereforewedonottestthefitofthefullyparameterizedmodelWefocusedonnaiumlveratesbe‐causethetruevital rates (ieγandε)cannotbedirectlyobservedwhendetectionisimperfectwhilethenaiumlveratescanbeobserved
We used an analysis of deviance approach (ANODEV egSkalski 1996) to evaluate the relative importance of climate andland cover to changes in bird distributions In this procedure wecompared thedeviance explainedbyour parameter of interest tothedevianceexplainedbythemostfullyparameterizedmodelinthemodelset
Dev(Mnull)isthedevianceofModel3whichaccountsforspatialbutnottemporalvariationincolonizationandextinctionprobabili‐tiesDev(Mfull)isthedevianceofModel2theglobalmodelwithbothspatialandtemporalvariationinγandεWethenevaluatedthedevi‐anceexplainedbyclimateandorlandcoverbyusingmodels45and6 forDev(Mcov)HighervaluesofR
2
Dev indicategreaterexplanatory
powerwith0indicatingnopowerand1indicatingpowerequaltothatofthefullyparameterizedmodelalthoughR2
Devcouldpotentially
exceed1becauseourcovariatemodelsarenotstrictlynestedwithinModel2ThereforewecalculatedandreportR2
Dev forbothclimate
and landcoverThis isnot theonlypossibleapproach toassessingrelativeimportanceofexplanatoryvariablesbutitissoundandhasbeenusedandrecommendedbyothers(Grosboisetal2008)
The general ANODEV approach to partitioning variation ledustofocusonhypothesistestingasameansofassessingtheneedforextramodelparameters (sourcesofvariation)Thesmallsetof
specificapriorihypothesesalsosets thisworkapart fromexplor‐atorystudiesinvestigatingtheadequacyofmanydifferentmodelsForthesereasonsweusedlikelihoodratiotestsasameansofcom‐paringdifferentmodelsalthoughwenotethatuseofmodelselec‐tioncriteria(egAICc)yieldedsimilarinferences
3emsp |emspRESULTS
Ourcovariatesindicatedasmalllossofhabitatandincreasedtem‐peraturesduringthestudyperioddespitetherelativelyshorttimescaleForEasternWoodPeweehabitatchangewasgt1on38ofrouteswiththemeanshareofsuitablehabitatnearroutesde‐cliningfrom416to409ForRed‐eyedVireohabitatchangewasgt1on23of routeswithmeansuitablehabitatdecliningfrom480to476Routetemperatureswererarelyabovetheheatstress thresholdbut theaveragetimeabovethis thresholdper route increased from 05hr per year from 1982 to 1997 to17hrperyearfrom1997to2012Wealsoobservedadeclineinperiodsofcoldstressfrom5450to5291hrperrouteperyearThePearsons correlation coefficient did not indicate anymulti‐collinearityproblemsamongourcovariates(R2lt02inallcases)
31emsp|emspEastern Wood Pewee
ForEasternWoodPeweethebest‐supportedmodelincludedbothclimateandhabitatmeasuresas covariatesofψθ andθprimeWeex‐cludedclimateandhabitatcovariatesfromthedetectionprobabilitymodelbecausesomeofthosemodelsdidnotconvergeThereforethedetectionmodelforeachspeciesincludedyearandstopnum‐ber as covariates As a result the global model included 85 pa‐rameters (Table1)TheHosmerndashLemeshowtest indicatedthattheglobalmodel fit thedataadequatelywithnoevidenceofdiscrep‐ancies between estimated and observed naiumlve colonization rates(χ2=441 df =8 p=082) and naiumlve extinction rates (χ2=1117df =8p=019)
EasternWoodPeweewerewidespreadinthestudyareawithanaverageprobabilityofoccupancyonBBSroutesin1997of090intheglobalmodel(Figure1)Occupancydeclinedslightlyto088by2012mirroringthedeclineincountsreportedbytheBBSInitialoccupancywaspositivelyrelatedtoavailablehabitatandhoursofcoldstress(Table1)Probabilityofdetectionatthefirstindividualstopduring1997waslowat013whileprobabilityofdetectionat the route level was much higher at 095 because birds weretypically available atmany stopswithina routeModel3whichincludedspatialvariationinclimateandhabitatinthecolonizationandextinctionmodelssignificantlyimprovedmodelfitrelativetoModel1(likelihoodratioχ2=4419df =6plt0001)Underthismodelestimatedcolonizationratesin1998rangedamongroutesfrom 008 to 028 (with amean of 018) while extinction ratesvariedfrom001to004(withameanof002Figure2)Model2differedfromModel3inthat itallowedcolonizationandextinc‐tionratestovaryeachyearModel2improvedmodelfitrelative
R2
Dev=Dev(Mnull)minusDev(Mcov)
Dev(Mnull)minusDev(Mfull)
emspensp emsp | emsp1991CLEMENT ET aL
TA B L E 1 emspEstimatedparametersforcorrelated‐detectiondynamicoccupancyModel2relatinghabitatandclimatecovariatestooccupancydynamicsofbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012Parameterestimatesindicatechangeinlogoddsofparametersinresponsetoz‐transformedcovariatesParametersψϑϑprimeγεp1p2andπdefinedintext
Parameters
Eastern Wood Pewee Red‐eyed Vireo
Estimate SE Estimate SE
Ψintercept 2367 0177 4136 0404
Ψhabitat 0556 0163 2315 0300
Ψheatstress minus0011 0198 minus0337 0281
Ψcoldstress 0598 0166 1020 0258
ϑintercept minus1821 0026 minus1385 0010
ϑhabitat minus0055 0026 0516 0013
ϑheatstress minus0094 0030 0017 0008
ϑcoldstress 0172 0040 0258 0013
ϑprimeintercept 2292 0045 0426 0020
ϑprimehabitat 0316 0036 1210 0018
ϑprimeheatstress minus0006 0038 0054 0013
ϑprimecoldstress minus0622 0064 0876 0019
γintercept minus0884 0353 0010 0424
γ 1999 minus0601 0585 minus0150 0532
γ 2000 minus1083 0633 0516 0464
γ 2001 minus0300 0484 0228 0483
γ 2002 minus1113 0577 0262 0511
γ 2003 minus1055 0564 minus0311 0551
γ2004 minus0656 0525 minus0179 0534
γ 2005 minus0677 0533 0468 0485
γ2006 minus1014 0620 minus0217 0579
γ2007 minus0162 0453 minus0093 0548
γ 2008 minus23545 70483373 minus0478 0607
γ 2009 minus0428 0455 minus0253 0566
γ 2010 minus0292 0478 minus0159 0517
γ 2011 minus1136 0607 0012 0490
γ 2012 minus0498 0474 0141 0497
γaveragehabitat 0095 0091 0801 0112
γaverageheatstress minus0024 0067 0005 0062
γaveragecoldstress 0227 0071 0358 0120
εintercept minus3957 0386 minus3767 0242
ε 1999 0365 0482 minus0189 0337
ε 2000 0049 0512 minus0921 0399
ε 2001 minus0266 0548 minus0532 0362
ε 2002 minus0576 0660 minus1156 0398
ε 2003 minus0298 0595 minus0692 0355
ε2004 0113 0509 minus0621 0343
ε 2005 0173 0482 minus1096 0429
ε2006 minus0145 0561 minus1173 0463
ε2007 0451 0458 minus0894 0366
ε 2008 0283 0480 minus0976 0424
ε 2009 0218 0496 minus0898 0381
(Continues)
1992emsp |emsp emspensp CLEMENT ET aL
Parameters
Eastern Wood Pewee Red‐eyed Vireo
Estimate SE Estimate SE
ε 2010 minus0397 0605 minus0348 0304
ε 2011 0643 0455 minus0999 0406
ε 2012 0153 0644 minus0803 0410
εaveragehabitat 0238 0090 minus1670 0122
εaverageheatstress minus0071 0092 minus0075 0074
εaveragecoldstress minus0307 0141 minus0904 0104
p1intercept minus2172 0043 1352 0038
p1 1998 minus0118 0048 minus0127 0058
p1 1999 minus0112 0049 minus0043 0058
p1 2000 minus0166 0050 minus0016 0059
p1 2001 minus0156 0049 0031 0060
p1 2002 minus0179 0049 0076 0061
p1 2003 minus0216 0051 0161 0064
p12004 minus0242 0051 0066 0060
p1 2005 minus0124 0050 0149 0060
p12006 minus0182 0051 0124 0059
p12007 minus0149 0050 0184 0061
p1 2008 minus0202 0052 0192 0063
p1 2009 minus0221 0052 0135 0061
p1 2010 minus0195 0051 0062 0059
p1 2011 minus0122 0051 0203 0062
p1 2012 minus0227 0055 0194 0062
p1stopnumber 0033 0009 minus0055 0011
p1stopnumbersquared minus0069 0010 minus0247 0013
p2intercept minus0921 0018 minus0557 0036
p2 1998 minus0138 0033 minus0106 0050
p2 1999 minus0137 0032 0000 0052
p2 2000 minus0164 0032 minus0027 0051
p2 2001 minus0077 0032 minus0053 0053
p2 2002 minus0101 0032 minus0019 0052
p2 2003 minus0199 0033 0086 0053
p22004 minus0186 0032 0124 0057
p2 2005 minus0146 0032 0076 0051
p22006 minus0155 0032 0128 0054
p22007 minus0153 0032 0120 0054
p2 2008 minus0168 0032 0072 0056
p2 2009 minus0077 0033 0101 0055
p2 2010 minus0114 0033 0085 0060
p2 2011 minus0079 0032 0124 0054
p2 2012 minus0186 0033 0036 0061
p2stopnumber 0045 0007 0165 0010
p2stopnumbersquared minus0105 0008 minus0183 0010
π 0533 0064 minus0505 0065
TA B L E 1 emsp (Continued)
emspensp emsp | emsp1993CLEMENT ET aL
toModel3althoughnotsignificantlyduetothenumberofaddi‐tional parameters (χ2=2679df =28p=053)UnderModel 2estimatedcolonizationratesvariedfrom000in2008to030in1998whileextinctionratesvariedfrom001 in2001ndash2003and2010to004in2011WeusedModels45and6toestimatethepercentof improvedmodel fit (measuredby thechange indevi‐ance)generatedbyModel2thatcouldbeaccountedforbytempo‐ralvariationinclimateandhabitatcovariates(Table2)Temporalvariationinhabitataccountedfor04ofthechangeindeviancewhile temporal variation in climate accounted for 196Model6whichaccountedfortemporalvariationinbothhabitatandcli‐mateindicatedthatin1998colonizationinthesouthwestportionofthestudyareawasabovethelocalaveragewhileextinctioninthesouthwestwasbelowthelocalaverageindicatingarelativelystrongoccupancy trend in thesouthwest in thatyear (Figure3)By2012thispatternhadreversedyieldingarelativelyweakoc‐cupancytrendinthesouthwest(Figure3)NoneofthesemodelsrepresentedasignificantimprovementoverModel3(pgt026)Insomecasesthesignoftheestimatedeffectoftemporalvariationinclimateandhabitatwasoppositeoftheexpectedeffectbutinnoinstancesweretheseestimatessignificant(Table3)
32emsp|emspRed‐eyed Vireo
The best‐supportedmodel for Red‐eyedVireo included the samecovariatesasforEasternWoodPeweeyieldingaglobalmodelwith85parameters (Table1)TheHosmerndashLemeshowtest indicatedfitwasadequate for theRed‐eyedVireoglobalmodel forbothnaiumlvecolonization rates (χ2=540df =4p=025) and naiumlve extinctionrates(χ2=100df =4p=091)
Red‐eyedVireowere similarlywidespread in the study areawith an average probability of occupancy of 091 in the global
model (Figure1)Occupancydecreasedslightlyto090by2012incontrasttothe increase incountsreportedbytheBBS Initialoccupancywas positively related to available habitat and hoursofcoldstress (Table1)Probabilityofdetectionat the first stopwasrelativelyhighat038whileprobabilityofdetectionattheroute levelnearlyperfectat098Model3which includedspa‐tialvariationinclimateandhabitattothecolonizationandextinc‐tionmodelsimprovedmodelfitdramatically(χ2=41884df =6plt0001)relativetothenullmodelInkeepingwiththegreaterchangeindevianceforRed‐eyedVireoModel3estimatedmorespatialvariationinvitalratescomparedtoEasternWoodPeweeIn 1998 estimated colonization rates varied across BBS routesfrom 010 to 088 (mean of 049)while extinction rates rangedfrom000to048(meanof004Figure4)Model2whichallowedcolonization and extinction rates to vary each year improvedmodelfitrelativetoModel3butnotsignificantlyduetothenum‐berofadditionalparameters(χ2=3427df =28p=019)UnderModel2estimatedcolonizationratesvariedfrom039in2008to060 in2000while estimatedextinction rates varied from003in2006to007in1998WeusedModels45and6toestimatethepercentofthedeviancereductioninModel2thatcouldbeac‐countedforbytemporalvariationinclimateandhabitatcovariates(Table2)Wefoundthattemporalvariation inhabitataccountedfor01of the change indeviancewhile climate accounted for56Model 6 which accounted for temporal variation in bothhabitatandclimateindicatedthatin1998bothcolonizationandextinctionwereaboveaverageindicatinghigherturnoverearlyinthestudyperiod(Figure5)By2012bothcolonizationandextinc‐tion rateshaddeclined indicatinggreater stability inoccupancy(Figure 5) None of these models represented a significant im‐provementoverModel3(pgt074)Someoftheestimatedeffects
F I G U R E 1 emspOccupancyprobabilityfor(a)EasternWoodPeweeand(b)Red‐eyedVireointheeasternUnitedStates1997underModel2Occupancyvariesbytheamountofsuitablehabitathoursofheatstressandhoursofcoldstressin1997
1994emsp |emsp emspensp CLEMENT ET aL
oftemporalvariationinclimateandhabitatdifferedfromexpecta‐tionsbutnotsignificantly(Table3)
4emsp |emspDISCUSSION
Wefound that forbothbirdspecies initialoccupancycoloniza‐tionratesandextinctionratescorrelatedwithspatialvariationinclimateandhabitatcovariatesHowevercolonizationandextinc‐tionrateswerenotcorrelatedwithtemporalvariation inclimateand habitat covariates Our results correlating initial occupancywith environmental covariates are broadly speaking consistentwithbiologicalexpectationandaraftofpreviousstudiesthathaveindicatedasignificantrelationshipbetweenspeciesdistributionsandhabitat(RobinsonWilsonampCrick2001ThogmartinSauerampKnutson 2007) climate (Barbet‐Massinamp Jetz 2014BellardBertelsmeier Leadley Thuiller amp Courchamp 2012) or both(Seoane Bustamante amp Diacuteaz‐Delgado 2004 Sohl 2014)Howeverwesuggestthatestimatedrelationshipsbetweenspatialvariationinenvironmentalcovariatesandcolonizationandextinc‐tion rates are of greater ecological interest than relatively phe‐nomenological species distribution models or climate envelopemodels(Clementetal2016)Theestimatedvitalratesofcoloni‐zationandextinctionbringusclosertounderstandingthedynamicprocessunderlying thedistributionof these speciesGivencon‐stantvital rateswecanalsoproject theequilibrium levelofoc‐cupancyatsites ( i
i+i
atsite i)Wecancomparetheseprojected
equilibria tocurrentoccupancytoproject trends for thespeciesunder current environmental conditions We can also use
projectedchangesinkeyenvironmentalvariablestodeveloppre‐dicted rangechanges in the futureWeargue that theseprojec‐tions are more robust than those from static climate envelopemodels which assume that species are currently in equilibrium(Yackulicetal2015)Incontraststaticmodelscanoverestimaterange changes when the equilibrium does not hold (Morin ampThuiller2009)
Given the importanceofglobal changeweweremotivated toinvestigate factors affecting temporal aswell as spatial variationin vital rates In this case spatial variation must be incorporatedintosuchanalysesinordertoaccommodatethedifferentdynamicsexpectedindifferentportionsofaspeciesrangeWeviewthere‐lationshipbetweentemporalvariationincovariatevaluesandvitalrates as the most relevant to understanding the effect of globalchangeontherecentandfuturedistributionsofourstudyspeciesFurthermore a correlation between vital rates and spatially vary‐ingcovariatesmaynot indicateasimilarcorrelationwithtemporalchangesinthosesamecovariatesForexampleaspecieswithstrongdispersal constraintsmight have a strong spatial relationshipwithacovariatebutaweaktemporalrelationshipduetoits immobility(Devictor et al 2008) Similarly an estimated spatial relationshipmaybepurelyphenomenologicalduetothenatureofspatialvari‐ablesWhen spatial variables followgradients it is relatively easytoobtainstatisticallysignificantcorrelationsabsentanycausalre‐lationship(Grosboisetal2008)ForexampleifcolonizationratesandtemperaturesbothfollownorthndashsouthgradientstheymaybespatiallycorrelatedevenifcolonizationisgovernedbysomeotherunidentifiedfactorInthiscasewewouldexpecttheweakcorrela‐tion between temporal changes in temperature and colonization
F I G U R E 2 emspProbabilityof(a)colonizationand(b)extinctionforEasternWoodPeweeintheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1995CLEMENT ET aL
ratestobemoreindicativeofthetruerelationshipthanthestrongcorrelationwithspatialvariationintemperature
BBS surveys indicate different abundance trajectories for ourstudyspecieswithcounts increasingforRed‐eyedVireosandde‐creasing for EasternWood Pewees (Sauer et al 2015) Becauseabundanceandoccupancytendtobelinked(Gastonetal2000)weexpectedsomedivergenceinoccupancydynamicsWefoundthatestimatedoccupancywashighandstableforbothspeciesalthoughtherewasgreaterturnoverforRed‐eyedVireosForexampleunderModel3 (spatialvariationonly incovariates)bothaveragecoloni‐zationrates(049)andaverageextinctionrates(004)wereatleasttwiceashighforRed‐eyedVireoscomparedtoEasternWoodPewee(018and002respectively)Red‐eyedVireosexhibitedgreaterspa‐tialvariationincolonizationandextinctionrates(Figure4)althoughour selected covariates explained little temporal variation for thisspeciesEasternWoodPeweecolonizationandextinctionratesex‐hibitedmoreofalatitudinalgradientreflectingaspatialcorrelationwith extreme temperature (Figure 2)Our results also hinted at agreatercorrelationbetweentemporalchanges in temperatureandoccupancyvitalratesforEasternWoodPeweewhichcouldplayaroleinthedeclineofthisspeciesalthoughtheresultswerenotsig‐nificant(Table3)Alternatively ithasbeensuggestedthatEasternWoodPeweemaybedecliningduetoa lossofforestedwinteringhabitat(RobbinsSauerGreenbergampDroege1989)orduetodeer‐mediatedchanges in foreststructureonbreedinggrounds (deCal‐esta 1994) illustrating the complexity of ascertaining cause andeffectusingobservationalstudiesinthesenaturalsystems
Althoughrelativelyrare(Siramietal2017)otherstudieshaveinvestigated both climate and land cover effects on bird distribu‐tions Inagreementwithour finding that temporalvariation incli‐mate explainedmuchmore of themodel deviance than temporalvariation in habitat climate‐only species distribution models hadbetter cross‐validation support than habitat‐only models for 409Europeanbirdspecies(Barbet‐Massinetal2012)SimilarlymodelsestimatingthenumberofnewlyoccupiedareasbyHoodedWarblers(Wilsonia citrina) inOntariousingclimatedatahadbetter informa‐tioncriterionsupportthanmodelsusingforestdata(MellesFortinLindsay amp Badzinski 2011) In contrast land‐use models outper‐formedclimatemodels for18 farmlandbird species in theUnitedKingdom(EglingtonampPearce‐Higgins2012)Similarlyvegetation‐based species distributionmodels had superior performance thanclimate‐based models for 79 Spanish bird species (Seoane et al2004)andforthreebirdspeciesinNewMexico(FriggensampFinch2015) Two studies focused on colonization and extinction ratesfoundmixed results ForGardenWarblers (Sylvia borin) inBritainvegetationexplainedmorevariationincolonizationwhiletempera‐turesexplainedmorevariationinlocalextinction(MustinAmarampRedpath2014)For122birdspeciesinOntariothebest‐supportedcovariates (land cover or climate change) varied among species(YalcinampLeroux2018)
Thedivergentresultsamongstudiesregardingtheimportanceofclimateandlandcovermaypartlybeduetothebiologyofindividualspecies (Yalcinamp Leroux 2018) For example farmland birdsmayTA
BLE
2emspR2 DevfordifferentmodelsofcolonizationandextinctionprobabilitiesforbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012
Mod
el
Cova
riate
s on
colo
niza
tion
and
extin
ctio
nEa
ster
n W
ood
Pew
eeRe
d‐ey
ed V
ireo
Inte
rcep
tAv
erag
e H
abita
tAv
erag
e Cl
imat
eH
abita
t Dev
iatio
nCl
imat
e D
evia
tion
Year
Eff
ect
Δ minus
2LL
R2 Dev
Δ minus
2LL
R2 Dev
2X
XX
X0
0010
00
0010
0
6X
XX
XX
minus2138
202
minus3230
57
4X
XX
Xminus2154
196
minus3234
56
5X
XX
Xminus2667
04
minus3422
01
3X
XX
minus2679
0minus3427
0
1X
minus7098
NA
minus45311
NA
Not
eNAnotapplicable
1996emsp |emsp emspensp CLEMENT ET aL
beparticularlyaffectedbychangesinagriculturalintensitybecauseoftheirextensiveuseoffarms(EglingtonampPearce‐Higgins2012)Alternativelyithasbeenarguedthatspatialscalemayplayaroleintherelativeimportanceofglobalchangefactorswithclimatemoreimportantatlargescales(PearsonampDawson2003)Wenotethattwostudiesthatfoundalargerroleforclimatebothreliedonlower‐resolution bird surveys (05deg atlas in Barbet‐Massin et al 2012
10km2atlasinMellesetal2011)IntheBBSdatathatweanalyzedsurveyswereconductedevery800mbutwecalculatedoccupancyacross each 394km transect In addition the cited studies useddifferentdata sources anddiverse analysismethods complicatinginterpretationofthedifferingresults
While we estimated that climate explained more model de‐viance than habitat these estimated relationships were not
F I G U R E 3 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionrates((t)minus (t)minus )forEasternWoodPeweeintheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
emspensp emsp | emsp1997CLEMENT ET aL
TA B L E 3 emspEstimatedparametersforcorrelated‐detectiondynamicoccupancyModel6relatinghabitatandclimatecovariatestooccupancydynamicsofbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012Parameterestimatesindicatechangeinlogoddsofparametersinresponsetoz‐transformedcovariatesParametersψϑϑprimeγε p1p2andπdefinedintextForkeyparameters(boldtext)expectedsignsincluded
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
Ψintercept 2367 0177 4087 0381
Ψhabitat 0556 0163 2323 0288
Ψheatstress minus0011 0198 minus0322 0275
Ψcoldstress 0598 0166 1054 0251
ϑintercept minus1821 0026 minus1385 0010
ϑhabitat minus0055 0026 0516 0013
ϑheatstress minus0094 0030 0017 0008
ϑcoldstress 0172 0040 0256 0013
ϑprimeintercept 2292 0045 0426 0020
ϑprimehabitat 0316 0036 1210 0018
ϑprimeheatstress minus0006 0038 0054 0013
ϑprimecoldstress minus0622 0064 0875 0019
γintercept minus0884 0353 0023 0162
γhabitatdeviation + minus0292 0478 minus0240 1768
γheatstressdeviation minus minus1136 0607 0064 0194
γcoldstressdeviation minus minus0498 0474 2398 2253
γaveragehabitat 0095 0091 0788 0111
γaverageheatstress minus0024 0067 0004 0062
γaveragecoldstress 0227 0071 0333 0119
εintercept minus3957 0386 minus4471 0141
εhabitatdeviation minus minus0397 0605 0121 1492
εheatstressdeviation + 0643 0455 minus0034 0182
εcoldstressdeviation + 0153 0644 2042 2072
εaveragehabitat 0238 0090 minus1664 0123
εaverageheatstress minus0071 0092 minus0080 0074
εaveragecoldstress minus0307 0141 minus0939 0104
p1intercept minus2172 0043 1351 0039
p1 1998 minus0118 0048 minus0128 0058
p1 1999 minus0112 0049 minus0043 0058
p1 2000 minus0166 0050 minus0017 0059
p1 2001 minus0156 0049 0031 0061
p1 2002 minus0179 0049 0076 0062
p1 2003 minus0216 0051 0160 0064
p12004 minus0242 0051 0066 0061
p1 2005 minus0124 0050 0149 0061
p12006 minus0182 0051 0124 0060
p12007 minus0149 0050 0183 0062
p1 2008 minus0202 0052 0191 0064
p1 2009 minus0221 0052 0134 0062
p1 2010 minus0195 0051 0062 0060
p1 2011 minus0122 0051 0202 0063
(Continues)
1998emsp |emsp emspensp CLEMENT ET aL
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
p1 2012 minus0227 0055 0193 0063
p1stopnumber 0033 0009 minus0055 0011
p1stopnumbersquared minus0069 0010 minus0247 0013
p2intercept minus0921 0018 minus0555 0037
p2 1998 minus0138 0033 minus0121 0050
p2 1999 minus0137 0032 minus0025 0052
p2 2000 minus0164 0032 minus0027 0051
p2 2001 minus0077 0032 minus0059 0053
p2 2002 minus0101 0032 minus0012 0052
p2 2003 minus0199 0033 0085 0053
p22004 minus0186 0032 0113 0059
p2 2005 minus0146 0032 0082 0052
p22006 minus0155 0032 0135 0054
p22007 minus0153 0032 0124 0055
p2 2008 minus0168 0032 0073 0057
p2 2009 minus0077 0033 0100 0056
p2 2010 minus0114 0033 0071 0062
p2 2011 minus0079 0032 0120 0055
p2 2012 minus0186 0033 0030 0063
p2stopnumber 0045 0007 0165 0010
p2stopnumbersquared minus0105 0008 minus0182 0011
π 0533 0064 minus0505 0065
TA B L E 3 emsp (Continued)
F I G U R E 4 emspProbabilityof(a)colonizationand(b)extinctionforRed‐eyedVireointheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1999CLEMENT ET aL
significantanddidnotexplainalargefractionofvariationThereareseveralpotentialexplanationsforwhywedidnotfindasub‐stantialresponsetochangesinhabitatandclimateFirstourdatawerelackingindynamiceventstoinformourmodelsThecovari‐ateschangedrelativelylittleoverthetimeperiodmakingitdiffi‐culttodetectaresponseThelackofchangewaspartlybecauseoftherelativelyshort(16‐year)timeperiodstudiedAlsoweusedamovingaverageforclimatedatawhichtendstoreducedifferences
overshorttimeperiods Inadditionthestudyregionhasexperi‐encedlesswarmingoverthepastcenturythanotherpartsoftheplanet (Walshetal2014)Thebirddistributionsalso lackeddy‐namismBecauseextinctionrateswerelowtherewerefewlocalextinctioneventstoinformthemodelAlthoughcolonizationrateswerehighthefactthatinitialoccupancywashighandextinctionrateswerelowmeanttherewerefewopportunitiesforcoloniza‐tionevents
F I G U R E 5 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionratesforRed‐eyedVireointheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
2000emsp |emsp emspensp CLEMENT ET aL
Second measurement error may have obscured responses toglobalchangeThePRISMclimatedataareestimatedbyinterpola‐tionratherthandirectmeasurementacrossthestudyarea(Dalyetal2008)AssuchtheremaybeartifactsoranomaliesthatimpededourestimationandincreaseduncertaintySimilarlyNLCDdataareremotelysensedandmay includesomemeasurementerrorwhichcan increase uncertainty in our estimates (Homer et al 2015)Additionallylandcoverwasonlymeasuredinthreeyearsrequiringus to imputevalues in13yearsAsa result all landcover changethatoccurredbetween2002and2006wasrecordedin2002whilechangesbetween2007and2011wererecorded in2007Thisap‐proximation isnot idealbutwasnecessarygiventhe5‐yearreso‐lutionofthedataThislowerresolutionofthelandcoverdatamayhavereducedthesensitivityofthemodel
Third wemay havemisspecified ourmodelWe tried to de‐velopbiologicallymeaningfulcovariatesbutwemightnotunder‐standthesystemaswellaswehopedItispossiblethatbirdvitalrates respond to climateon adifferent time scale (ie not a15‐yearmoving average) or to climateduringdifferent timesof theyearortodifferenttemperaturethresholdsortodifferentclimatecomponents (egprecipitation Illaacutenetal2014) It isalsopossi‐ble that thebirds respond to finerhabitat features than the landcovertypesweusedoratadifferentspatialscale(ienot400mPearsonampDawson2003)Alternatively theremaybe importantregionaldifferencesor interactionsamongcovariatesthatwedidnotcaptureOritispossiblethatchangesinclimateandhabitatintheirbreedingareassimplyhavelittleeffectonvitalratesforthesespecies relative to other factors such as competitionwith otherbirdspeciesorglobalchangeoccurringontheirwinteringgroundsinSouthAmerica(Dormann2007)Ofcoursethesemisspecifica‐tionissuesareinherenttoallobservationalstudiesandnotuniquetothisstudy
Itwould be possible for us to explore some of themisspec‐ification issues just mentioned For example we could rerunthe samemodelsusingnewcovariates representing a varietyoftemperature thresholds and habitat buffer sizes to search for astrongercorrelationExperiencesuggeststhatpersistencewouldeventuallyberewardedwithap‐valuelt005ora lowmodelse‐lection valueHowever one reason to avoid such exploration isthat it tends to lead toType Ierrorsoverfitmodels andbiasedparameterestimates(FiebergampJohnson2015)Furthermorewefind thecontrastbetweensignificant correlations for spatial co‐variatesandnonsignificantcorrelationsfortemporalcovariatesil‐luminatingIntypicalclimateenvelopestudieswewouldestimatethe relationshipbetweencovariatesand thecurrentdistributionofaspeciesmuchlikeourestimateofinitialoccupancy(Dormann2007)BecauseweobtainedsignificantparameterestimateswemightconcludethatwehaveidentifiedcovariatesthatdeterminetherangeofthisspeciesThenwemightusetheseresultstoproj‐ectthefuturerangeofthespeciesundervariousclimatescenariosbased on the (questionable) assumption that the covariatendashspe‐cies relationship will not change as climate and habitat change(Cuddington et al 2013 Elith et al 2010Gustafson 2013) In
this case the insignificant correlationwith the temporal covari‐atessuggeststhatouroccupancycovariatesmerelydescribetherangeofthesebirdswithoutdeterminingtherangeFurthermoreour results suggest that despite significant results in the staticportionof themodelwehaveyet to identify themost relevantcomponents of climate change shaping thedistributionof thesebirds although itmay be difficult to identify these componentsgiven the lackofobservedchangeduringa relatively short timeperiodFinallyourresultssuggestthatprojectionsbasedonthestaticportionofourmodelareunlikelytobereliabledespitetheirstatisticalsignificance
Ifourinterpretationiscorrectitsuggeststhatsomeprojectionsof futurerangeshiftsbasedonstaticmodelsrelyonunsupportedassumptionsandarenot reliableWehavedemonstratedadiffer‐entapproachthatdoesnotrequirethestrongassumptionsofstaticmodelingapproachesOurapproachwouldbemoreapttoidentifyfactorsthatguidespeciesrsquoresponsetoglobalchangeifboththespe‐ciesandthecovariatesexhibitedmoredynamismandifweanalyzedalongertimeperiodthatencompassedgreaterchange
ACKNOWLEDG MENTS
WethankKeithPardieckDaveZiolkowskiJrandthethousandsofhighlyskilledvolunteerswhohavecoordinatedandperformedtheNorthAmericanBreedingBirdSurvey for half a centuryWealsothanktheClimateResearchUnitattheUniversityofEastAngliaformakingvaluableclimatedataeasilyaccessibleMJCwasfinanciallysupported by the BBS and the USGS Northeast Climate ScienceCenterThecontentsofthisdocumentaretheresponsibilityoftheauthorsanddonotnecessarilyrepresenttheviewsoftheUSGSAnyuseoftradeproductorfirmnamesisfordescriptivepurposesonlyanddoesnotimplyendorsementbytheUSGovernment
DEDIC ATION
We dedicate this paper to the late Chandler S Robbins ldquoFatherrdquooftheNorthAmericanBreedingBirdSurvey (BBS)DatafromtheBBSformthebasisforthisspecificpaperandforhundredsofothersfocusedon topics ranging from local trends tocontinentalmacro‐ecologyChanwasauniquecombinationofbird‐loverandscientistwhosevisionandinquisitiveenthusiasmwereaninspirationtoallofusfortunateenoughtohaveknownhim
CONFLIC T OF INTERE S T
Nonedeclared
AUTHORSrsquo CONTRIBUTION
JACandJDNconceivedthestudyAJTandSWanalyzedhabitatandclimatedataMJCJEHandJDNperformedtheoccupancyanaly‐sisMJCledthewritingofthemanuscriptAllauthorsgavefinalap‐provalforpublication
emspensp emsp | emsp2001CLEMENT ET aL
DATA ACCE SSIBILIT Y
Raw data are available from the following sources Breeding BirdSurvey httpswwwpwrcusgsgovbbsrawdata PRISM climatedata httpwwwprismoregonstateedu NLCD land cover datahttpswwwmrlcgov Processed data will be archived with theDryadDigitalRepository
ORCID
Matthew J Clement httpsorcidorg0000‐0003‐4231‐7949
James D Nichols httpsorcidorg0000‐0002‐7631‐2890
Adam J Terando httpsorcidorg0000‐0002‐9280‐043X
James E Hines httpsorcidorg0000‐0001‐5478‐7230
Steven G Williams httpsorcidorg0000‐0003‐3760‐6818
R E FE R E N C E S
Anderson D R Burnham K P Gould W R amp Cherry S (2001)Concerns about finding effects that are actually spuriousWildlife Society Bulletin29311ndash316
Arauacutejo M B Cabeza M Thuiller W Hannah L amp WilliamsP H (2004) Would climate change drive species out of re‐serves An assessment of existing reserve‐selection meth‐ods Global Change Biology 10 1618ndash1626 httpsdoiorg101111j1365‐2486200400828x
Barbet‐MassinMampJetzW(2014)A40‐yearcontinent‐widemulti‐speciesassessmentofrelevantclimatepredictorsforspeciesdistri‐butionmodellingDiversity and Distributions201285ndash1295httpsdoiorg101111ddi12229
Barbet‐Massin M Thuiller W amp Jiguet F (2012) The fate ofEuropean breeding birds under climate land‐use and disper‐sal scenarios Global Change Biology 18 881ndash890 httpsdoiorg101111j1365‐2486201102552x
Bellard C Bertelsmeier C Leadley P Thuiller W ampCourchamp F (2012) Impacts of climate change on the fu‐ture of biodiversity Ecology Letters 15 365ndash377 httpsdoiorg101111j1461‐0248201101736x
Boumlhning‐Gaese K amp Bauer H G (1996) Changes in species abun‐dance distribution and diversity in a central European birdcommunity Conservation Biology 10 175ndash187 httpsdoiorg101046j1523‐1739199610010175x
BrommerJELehikoinenAampValkamaJ(2012)Thebreedingrangesof central European and Arctic bird speciesmove poleward PLoS ONE7e43648httpsdoiorg101371journalpone0043648
CalderWAampKingJR(1974)ThermalandcaloricrelationsofbirdsInDSFarnerampJRKing(Eds)Avian biology(Vol4pp259ndash413)NewYorkNYAcademicPress
CesaraccioCSpanoDDucePampSnyderRL(2001)Animprovedmodel for determining degree‐day values from daily temperaturedataInternational Journal of Biometeorology45161ndash169httpsdoiorg101007s004840100104
ChamberlinTC (1890)ThemethodofmultipleworkinghypothesesScience1592ndash96
ClementMJHinesJENicholsJDPardieckKLampZiolkowskiDJJr (2016)Estimating indicesofrangeshifts inbirdsusingdy‐namicmodelswhendetectionisimperfectGlobal Change Biology223273ndash3285httpsdoiorg101111gcb13283
Commission for Environmental Cooperation (1997) Ecological regionsofNorthAmericaTowardacommonperspectiveCommission for
EnvironmentalCooperationMontrealRetrievedfromftpftpepagovwedecoregionscec_naCEC_NAecopdf
Crowl TA Crist TO Parmenter R R BelovskyGamp LugoA E(2008)Thespreadofinvasivespeciesandinfectiousdiseaseasdriv‐ersofecosystemchangeFrontiers in Ecology and the Environment6238ndash246httpsdoiorg101890070151
Cuddington K FortinM J Gerber L RHastings A Liebhold AOConnorMampRayC(2013)Process‐basedmodelsarerequiredtomanageecologicalsystemsinachangingworldEcosphere41ndash12httpsdoiorg101890ES12‐001781
DalyCHalbleibMSmithJIGibsonWPDoggettMKTaylorGHhellipPasterisPP(2008)Physiographicallysensitivemappingofcli‐matologicaltemperatureandprecipitationacrosstheconterminousUnited States International Journal of Climatology 28 2031ndash2064httpsdoiorg101002joc1688
DawsonW (1958) Relation of oxygen consumption and evaporativewater losstotemperatureintheCardinalPhysiological Zoology3137ndash48httpsdoiorg101086physzool31130155377
DawsonWRampWhittowGC (2000)Regulationofbodytempera‐ture InG CWhittow (Ed)Sturkiersquos avian physiology (5th ed pp343ndash390)SanDiegoCAAcademicPress
DeCalestaDS(1994)Effectofwhite‐taileddeeronsongbirdswithinmanagedforestsinPennsylvaniaJournal of Wildlife Management58711ndash718httpsdoiorg1023073809685
DevictorVJulliardRCouvetDampJiguetF(2008)Birdsaretrack‐ing climatewarming but not fast enoughProceedings of the Royal Society of London B Biological Sciences2752743ndash2748httpsdoiorg101098rspb20080878
Dormann C F (2007) Promising the future Global change projec‐tionsofspeciesdistributionsBasic and Applied Ecology8387ndash397httpsdoiorg101016jbaae200611001
Dormann C F Schymanski S J Cabral J Chuine I Graham CHartigFhellipSingerA(2012)Correlationandprocessinspeciesdis‐tributionmodelsBridgingadichotomyJournal of Biogeography392119ndash2131httpsdoiorg101111j1365‐2699201102659x
EccelE(2010)WhatwecanasktohourlytemperaturerecordingPartIIHourlyinterpolationoftemperaturesforclimatologyandmodel‐lingItalian Journal of Agrometeorology1545ndash50
EccelEampCordanoE(2013)InterpolTHourlyinterpolationofmulti‐pletemperaturedailyseriesRpackageversion211RetrievedfromhttpsCRANR‐projectorgpackage=InterpolT
EglingtonSMampPearce‐HigginsJW(2012)Disentanglingtherela‐tiveimportanceofchangesinclimateandland‐useintensityindriv‐ingrecentbirdpopulationtrendsPLoS ONE7e30407httpsdoiorg101371journalpone0030407
Elith JKearneyMampPhillipsS (2010)Theartofmodelling range‐shifting species Methods in Ecology and Evolution 1 330ndash342httpsdoiorg101111j2041‐210X201000036x
FiebergJampJohnsonDH(2015)MMIMultimodelinferenceormod‐elswithmanagement implications Journal of Wildlife Management79708ndash718
FriggensMMampFinchDM (2015) Implicationsof climatechangeforbirdconservationinthesouthwesternUSunderthreealternativefuturesPLoS ONE10e0144089
FryJXianGZJinSDewitzJHomerCGYangLhellipWickhamJD (2011)Completionof the2006national landcoverdatabasefor theconterminousUnitedStatesPhotogrammetric Engineering amp Remote Sensing77858ndash864
Fuller R J Gregory R D Gibbons D W Marchant J HWilson J D Baillie S R amp Carter N (1995) Population de‐clines and range contractions among lowland farmland birdsin Britain Conservation Biology 9 1425ndash1441 httpsdoiorg101046j1523‐1739199509061425x
Gaston K J Blackburn T M Greenwood J J Gregory R DQuinn R M amp Lawton J H (2000) Abundancendashoccupancy
2002emsp |emsp emspensp CLEMENT ET aL
relationships Journal of Applied Ecology 37 39ndash59 httpsdoiorg101046j1365‐2664200000485x
GoldsteinRB(1974)RelationofmetabolismtoambienttemperatureintheverdinCondor76116ndash119httpsdoiorg1023071365995
GrinnellJ(1917)FieldtestsoftheoriesconcerningdistributionalcontrolAmerican Naturalist51115ndash128httpsdoiorg101086279591
GrosboisVGimenezOGaillardJMPradelRBarbraudCClobertJhellipWeimerskirchH(2008)Assessingtheimpactofclimatevari‐ation on survival in vertebrate populationsBiological Reviews 83357ndash399httpsdoiorg101111j1469‐185X200800047x
GustafsonEJ(2013)WhenrelationshipsestimatedinthepastcannotbeusedtopredictthefutureUsingmechanisticmodelstopredictlandscapeecologicaldynamicsinachangingworldLandscape Ecology281429ndash1437httpsdoiorg101007s10980‐013‐9927‐4
Hamel P B (1992) LandManagers guide to the birds of the SouthGeneralTechnicalReportSE‐22USDAForestServiceSoutheasternForestExperimentStationandTheNatureConservancyChapelHillNC
Hinds D S amp Calder W A (1973) Temperature regulation of thePyrrhuloxiaandtheArizonaCardinalPhysiological Zoology4655ndash71httpsdoiorg101086physzool46130152517
HinesJE(2006)PRESENCE Software to estimate patch occupancy and related parameters Laurel MD US Geological survey PatuxentWildlife Research Center Retrieved from httpwwwmbr‐pwrcusgsgovsoftwarepresencehtml
HinesJENicholsJDampCollazoJA(2014)Multiseasonoccupancymodels for correlated replicate surveys Methods in Ecology and Evolution5583ndash591httpsdoiorg1011112041‐210X12186
HinesJENicholsJDRoyleJAMacKenzieDIGopalaswamyAMKumarNampKaranthKU(2010)TigersontrailsOccupancymodeling for cluster sampling Ecological Applications 20 1456ndash1466httpsdoiorg10189009‐03211
HockeyPA SiramiCRidleyARMidgleyGFampBabikerHA(2011) Interrogating recent range changes in South African birdsConfounding signals from land use and climate change present achallenge for attribution Diversity and Distributions 17 254ndash261httpsdoiorg101111j1472‐4642201000741x
Hoegh‐Guldberg O Hughes L McIntyre S Lindenmayer D BParmesanCPossinghamHPampThomasCD (2008)AssistedcolonizationandrapidclimatechangeScience321345ndash346
Homer C Dewitz J Fry J Coan M Hossain N Larson C hellipWickham J (2007) Completion of the 2001 national land coverdatabase for the conterminous United States Photogrammetric Engineering amp Remote Sensing73337
HomerCDewitzJYangLJinSDanielsonPXianGhellipMegownK(2015)Completionofthe2011NationalLandCoverDatabasefortheconterminousUnitedStatesndashrepresentingadecadeoflandcoverchange informationPhotogrammetric Engineering amp Remote Sensing81345ndash354
HootenMBWikleCKDorazioRMampRoyleJA(2007)HierarchicalspatiotemporalmatrixmodelsforcharacterizinginvasionsBiometrics63558ndash567httpsdoiorg101111j1541‐0420200600725x
HosmerDWJrampLemeshowS(2000)Applied logistic regression(2nded)NewYorkNYJohnWileyampSons
Huntley B Collingham Y CGreen R EHiltonGM RahbekCamp Willis S G (2006) Potential impacts of climatic change upongeographical distributions of birds Ibis 148 8ndash28 httpsdoiorg101111j1474‐919X200600523x
IgleciaMNCollazoJAampMcKerrowAJ(2012)Useofoccupancymodels to evaluate expert knowledge‐based species‐habitat rela‐tionshipsAvian Conservation and Ecology7(2)13pRetrievedfromhttpwwwace‐ecoorgvol7iss2art5
IllaacutenJGThomasCDJonesJAWongWKShirleySMampBettsMG(2014)Precipitationandwintertemperaturepredictlong‐termrange‐scale abundancechanges inWesternNorthAmericanbirds
Global Change Biology 20 3351ndash3364 httpsdoiorg101111gcb12642
IoannidisJP (2005)WhymostpublishedresearchfindingsarefalsePLoS Medicine2e124
KeacuteryMGuillera‐ArroitaGampLahoz‐Monfort J J (2013)Analysingand mapping species range dynamics using occupancy modelsJournal of Biogeography 40 1463ndash1474 httpsdoiorg101111jbi12087
Khaliq I Hof C Prinzinger R Boumlhning‐Gaese K amp PfenningerM(2014) Global variation in thermal tolerances and vulnerability ofendotherms to climate change Proceedings of the Royal Society B Biological Sciences 281(1789) 20141097 httpsdoiorg101098rspb20141097
Laursen K amp Frikke J (2008) Hunting from motorboats displacesWadden Sea eiders Somateria mollissima from their favouredfeeding distribution Wildlife Biology 14 423ndash433 httpsdoiorg1029810909‐6396‐144423
LemoineNBauerHGPeintingerMampBoumlhning‐GaeseK (2007)Effectsof climate and land‐use changeon species abundance in acentralEuropeanbirdcommunityConservation Biology21495ndash503httpsdoiorg101111j1523‐1739200600633x
MacArthurRH(1972)Geographical ecology Patterns in the distribution of speciesNewYorkNYHarperampRow
MacKenzieDINicholsJDHinesJEKnutsonMGampFranklinAB(2003)Estimatingsiteoccupancycolonizationandlocalextinc‐tionwhenaspeciesisdetectedimperfectlyEcology842200ndash2207httpsdoiorg10189002‐3090
MacKenzieDINicholsJDLachmanGBDroegeSRoyleJAampLangtimmCA(2002)Estimatingsiteoccupancyrateswhendetec‐tionprobabilitiesarelessthanoneEcology832248ndash2255httpsdoiorg1018900012‐9658(2002)083[2248ESORWD]20CO2
MacKenzieDINicholsJDRoyleJAPollockKHBaileyLampHinesJE(2018)Occupancy estimation and modeling Inferring pat‐terns and dynamics of species occurrence (2nd ed) BurlingtonMAAcademicPress
MellesSJFortinMJLindsayKampBadzinskiD(2011)ExpandingnorthwardInfluenceofclimatechangeforestconnectivityandpop‐ulationprocessesonathreatenedspeciesrangeshiftGlobal Change Biology1717ndash31httpsdoiorg101111j1365‐2486201002214x
Morin X amp Thuiller W (2009) Comparing niche‐and process‐based models to reduce prediction uncertainty in species rangeshifts under climate change Ecology 90 1301ndash1313 httpsdoiorg10189008‐01341
MustinKAmarAampRedpathSM(2014)Colonizationandextinctiondynamicsofadecliningmigratorybirdareinfluencedbyclimateandhabitat degradation Ibis 156 788ndash798 httpsdoiorg101111ibi12173
NicholsJDThomasLampConnPB(2009)InferencesaboutlandbirdabundancefromcountdataRecentadvancesandfuturedirectionsInDLThomsonEJCoochampMJConroy(Eds)Modeling demo‐graphic processes in marked populations(pp201ndash235)NewYorkNYSpringer
OmernikJM(1995)EcoregionsAspatialframeworkforenvironmen‐talmanagement InWSDavisampTSimon(Eds)Biological assess‐ment and criteria Tools for water resource planning and decision making (pp49ndash62)BocaRatonFLLewisPublishers
OmernikJM(2004)Perspectivesonthenatureanddefinitionofeco‐logicalregionsEnvironmental Management34(Supplement1)S27ndashS38httpsdoiorg101007s00267‐003‐5197‐2
Omernik J M amp Griffith G E (2014) Ecoregions of the contermi‐nousUnited States Evolution of a hierarchical spatial frameworkEnvironmental Management541249ndash1266httpsdoiorg101007s00267‐014‐0364‐1
Pardieck K L ZiolkowskiD J JrampHudsonM‐A R (2015)NorthAmericanBreedingBirdSurveyDataset1966ndash2014version20140
emspensp emsp | emsp2003CLEMENT ET aL
USGeologicalSurveyPatuxentWildlifeResearchCenterRetrievedfromhttpswwwpwrcusgsgovbbsrawdata
ParmesanCampYoheG (2003)Agloballycoherentfingerprintofcli‐mate change impacts across natural systemsNature 421 37ndash42httpsdoiorg101038nature01286
PearsonRGampDawsonTP(2003)PredictingtheimpactsofclimatechangeonthedistributionofspeciesArebioclimateenvelopemod‐elsusefulGlobal Ecology and Biogeography12361ndash371
PlattJR(1964)StronginferenceScience146347ndash353PopperK(1959)The logic of scientific discoveryLondonUKHutchinsonRobbins C S Sauer J R Greenberg R S amp Droege S (1989)
Population declines in North American birds that migrate to theNeotropics Proceedings of the National Academy of Sciences 867658ndash7662httpsdoiorg101073pnas86197658
Robinson R A Wilson J D amp Crick H Q (2001) The impor‐tance of arable habitat for farmland birds in grassland land‐scapes Journal of Applied Ecology 38 1059ndash1069 httpsdoiorg101046j1365‐2664200100654x
Romesburg H C (1981) Wildlife science Gaining reliable knowl‐edge Journal of Wildlife Management 45 293ndash313 httpsdoiorg1023073807913
Root T (1988) Energy constraints on avian distributions and abun‐dancesEcology69330ndash339httpsdoiorg1023071940431
Ruiz‐GutieacuterrezVampZipkinEF (2011)Detectionbiasesyieldmis‐leadingpatternsof speciespersistence and colonization in frag‐mented landscapesEcosphere2 1ndash14 httpsdoiorg101890ES10‐002071
SauerJRHinesJEFallonJEPardieckKLZiolkowskiDJJrampLinkWA(2015)The North American breeding bird survey results and analysis 1966ndash2013 Version 01302015LaurelMDUSGSPatuxentWildlifeResearchCenterhttpswwwmbr‐pwrcusgsgovbbsbbshtml
Sauer J R LinkWA Fallon J E Pardieck K L amp ZiolkowskiDJJr(2013)TheNorthAmericanbreedingbirdsurvey1966ndash2011SummaryanalysisandspeciesaccountsNorth American Fauna791ndash32httpsdoiorg103996nafa790001
SeoaneJBustamanteJampDiacuteaz‐DelgadoR(2004)Competingrolesforlandscapevegetationtopographyandclimateinpredictivemod‐elsofbirddistributionEcological Modelling171209ndash222httpsdoiorg101016jecolmodel200308006
Sirami C Caplat P Popy S Clamens A Arlettaz R Jiguet F hellipMartin J‐L (2017) Impacts of global change on species distri‐butions Obstacles and solutions to integrate climate and landuse Global Ecology and Biogeography 26 385ndash394 httpsdoiorg101111geb12555
SkalskiJR(1996)Regressionofabundanceestimatesfrommarkrecap‐ture surveys against environmental covariatesCanadian Journal of
Fisheries and Aquatic Sciences53196ndash204httpsdoiorg101139f95‐169
SohlTL(2014)Therelativeimpactsofclimateandland‐usechangeonconterminousUnitedStatesbird species from2001 to2075PLoS ONE9e112251httpsdoiorg101371journalpone0112251
ThogmartinWESauerJRampKnutsonMG(2007)ModelingandmappingabundanceofAmericanwoodcockacrosstheMidwesternandNortheasternUnitedStatesJournal of Wildlife Management71376ndash382httpsdoiorg1021932005‐680
ThomasCDampLennonJJ (1999)Birdsextendtheirrangesnorth‐wardsNature399213httpsdoiorg10103820335
Trathan P N Garciacutea‐Borboroglu P Boersma D Bost C ACrawford R J CrossinG ThellipWienecke B (2015) Pollutionhabitatlossfishingandclimatechangeascriticalthreatstopen‐guins Conservation Biology 29 31ndash41 httpsdoiorg101111cobi12349
TravisJMJ(2003)Climatechangeandhabitatdestructionadeadlyanthropogenic cocktail Proceedings of the Royal Society of London Series B Biological Sciences 270(1514) 467ndash473 httpsdoiorg101098rspb20022246
UrbanMC (2015)Acceleratingextinction risk fromclimatechangeScience348571ndash573httpsdoiorg101126scienceaaa4984
WalshJWuebblesDHayhoeKKossinJKunkelKStephensGhellipSomervilleR(2014)OurchangingclimateInJMMelilloTCRichmondampGWYohe(Eds)Climate change impacts in the United States The Third National climate assessment(pp19ndash67)WashingtonDCUSGovernmentPrintingOffice
YackulicCBNichols JDReid JampDerR (2015)Topredict thenichemodelcolonizationandextinctionEcology9616ndash23httpsdoiorg10189014‐13611
Yalcin S amp Leroux S J (2018) An empirical test of the relative andcombinedeffectsofland‐coverandclimatechangeonlocalcoloni‐zationandextinctionGlobal Change Biology243849ndash3861httpsdoiorg101111gcb14169
ZhuKWoodallCWampClarkJS(2012)FailuretomigrateLackoftreerangeexpansioninresponsetoclimatechangeGlobal Change Biology181042ndash1052httpsdoiorg101111j1365‐2486201102571x
How to cite this articleClementMJNicholsJDCollazoJATerandoAJHinesJEWilliamsSGPartitioningglobalchangeAssessingtherelativeimportanceofchangesinclimateandlandcoverforchangesinaviandistributionEcol Evol 201991985ndash2003 httpsdoiorg101002ece34890

1990emsp |emsp emspensp CLEMENT ET aL
with complexmodels (MacKenzie et al 2018) By beginningwithhighlyparameterizedmodelsandprogressingtowardreducedmod‐elswereducedtheriskofconfoundingtheoccupancyanddetectionprocesses (MacKenzieet al2018)Wealsochecked forpotentialmulti‐collinearityissuesbycalculatingPearsonscorrelationcoeffi‐cientforthedifferentcovariatesusedinthemodelsWetreatedanR2lt05asindicativeofalackofmulti‐collinearity
WealsoevaluatedthegoodnessoffitoftheglobalmodelforeachspeciesusingthenaiumlvecolonizationandextinctionratestocalculatetheteststatisticofaHosmerndashLemeshowtest(HosmerampLemeshow2000) Inthiscontext thenaiumlvecolonizationrate is thenumberofsitesthatchangedfromnondetectionsitesinyeart todetectionsitesinyear t+1while thenaiumlveextinction rate is thenumberof sitesthatchangedfromdetectionsitesinyeart tonondetectionsitesinyeart+1Wedividedtheroutesintodecilesbasedontheexpectedcolonizationandextinctionratesandcomparedthepredictedratesto theobservedrateswithchi‐squaretests (α=005) Ifnecessarywecombineddecilestoensurethatthepredictednumberofdetec‐tionsinadecilewasgt4AlthoughtheHosmerndashLemeshowteststatis‐ticisoftencalculatedfrompredictedpresencevitalratesseemedtobemoreappropriatemeasuresinthiscasebecausedynamicmodelsestimatechangesinpresenceratherthanpresence(Clementetal2016)Wenotethatthisuseofnaiumlveratesfocusesonproductsofmodelparameters(ieγεp ψ ϑ)andthereforewedonottestthefitofthefullyparameterizedmodelWefocusedonnaiumlveratesbe‐causethetruevital rates (ieγandε)cannotbedirectlyobservedwhendetectionisimperfectwhilethenaiumlveratescanbeobserved
We used an analysis of deviance approach (ANODEV egSkalski 1996) to evaluate the relative importance of climate andland cover to changes in bird distributions In this procedure wecompared thedeviance explainedbyour parameter of interest tothedevianceexplainedbythemostfullyparameterizedmodelinthemodelset
Dev(Mnull)isthedevianceofModel3whichaccountsforspatialbutnottemporalvariationincolonizationandextinctionprobabili‐tiesDev(Mfull)isthedevianceofModel2theglobalmodelwithbothspatialandtemporalvariationinγandεWethenevaluatedthedevi‐anceexplainedbyclimateandorlandcoverbyusingmodels45and6 forDev(Mcov)HighervaluesofR
2
Dev indicategreaterexplanatory
powerwith0indicatingnopowerand1indicatingpowerequaltothatofthefullyparameterizedmodelalthoughR2
Devcouldpotentially
exceed1becauseourcovariatemodelsarenotstrictlynestedwithinModel2ThereforewecalculatedandreportR2
Dev forbothclimate
and landcoverThis isnot theonlypossibleapproach toassessingrelativeimportanceofexplanatoryvariablesbutitissoundandhasbeenusedandrecommendedbyothers(Grosboisetal2008)
The general ANODEV approach to partitioning variation ledustofocusonhypothesistestingasameansofassessingtheneedforextramodelparameters (sourcesofvariation)Thesmallsetof
specificapriorihypothesesalsosets thisworkapart fromexplor‐atorystudiesinvestigatingtheadequacyofmanydifferentmodelsForthesereasonsweusedlikelihoodratiotestsasameansofcom‐paringdifferentmodelsalthoughwenotethatuseofmodelselec‐tioncriteria(egAICc)yieldedsimilarinferences
3emsp |emspRESULTS
Ourcovariatesindicatedasmalllossofhabitatandincreasedtem‐peraturesduringthestudyperioddespitetherelativelyshorttimescaleForEasternWoodPeweehabitatchangewasgt1on38ofrouteswiththemeanshareofsuitablehabitatnearroutesde‐cliningfrom416to409ForRed‐eyedVireohabitatchangewasgt1on23of routeswithmeansuitablehabitatdecliningfrom480to476Routetemperatureswererarelyabovetheheatstress thresholdbut theaveragetimeabovethis thresholdper route increased from 05hr per year from 1982 to 1997 to17hrperyearfrom1997to2012Wealsoobservedadeclineinperiodsofcoldstressfrom5450to5291hrperrouteperyearThePearsons correlation coefficient did not indicate anymulti‐collinearityproblemsamongourcovariates(R2lt02inallcases)
31emsp|emspEastern Wood Pewee
ForEasternWoodPeweethebest‐supportedmodelincludedbothclimateandhabitatmeasuresas covariatesofψθ andθprimeWeex‐cludedclimateandhabitatcovariatesfromthedetectionprobabilitymodelbecausesomeofthosemodelsdidnotconvergeThereforethedetectionmodelforeachspeciesincludedyearandstopnum‐ber as covariates As a result the global model included 85 pa‐rameters (Table1)TheHosmerndashLemeshowtest indicatedthattheglobalmodel fit thedataadequatelywithnoevidenceofdiscrep‐ancies between estimated and observed naiumlve colonization rates(χ2=441 df =8 p=082) and naiumlve extinction rates (χ2=1117df =8p=019)
EasternWoodPeweewerewidespreadinthestudyareawithanaverageprobabilityofoccupancyonBBSroutesin1997of090intheglobalmodel(Figure1)Occupancydeclinedslightlyto088by2012mirroringthedeclineincountsreportedbytheBBSInitialoccupancywaspositivelyrelatedtoavailablehabitatandhoursofcoldstress(Table1)Probabilityofdetectionatthefirstindividualstopduring1997waslowat013whileprobabilityofdetectionat the route level was much higher at 095 because birds weretypically available atmany stopswithina routeModel3whichincludedspatialvariationinclimateandhabitatinthecolonizationandextinctionmodelssignificantlyimprovedmodelfitrelativetoModel1(likelihoodratioχ2=4419df =6plt0001)Underthismodelestimatedcolonizationratesin1998rangedamongroutesfrom 008 to 028 (with amean of 018) while extinction ratesvariedfrom001to004(withameanof002Figure2)Model2differedfromModel3inthat itallowedcolonizationandextinc‐tionratestovaryeachyearModel2improvedmodelfitrelative
R2
Dev=Dev(Mnull)minusDev(Mcov)
Dev(Mnull)minusDev(Mfull)
emspensp emsp | emsp1991CLEMENT ET aL
TA B L E 1 emspEstimatedparametersforcorrelated‐detectiondynamicoccupancyModel2relatinghabitatandclimatecovariatestooccupancydynamicsofbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012Parameterestimatesindicatechangeinlogoddsofparametersinresponsetoz‐transformedcovariatesParametersψϑϑprimeγεp1p2andπdefinedintext
Parameters
Eastern Wood Pewee Red‐eyed Vireo
Estimate SE Estimate SE
Ψintercept 2367 0177 4136 0404
Ψhabitat 0556 0163 2315 0300
Ψheatstress minus0011 0198 minus0337 0281
Ψcoldstress 0598 0166 1020 0258
ϑintercept minus1821 0026 minus1385 0010
ϑhabitat minus0055 0026 0516 0013
ϑheatstress minus0094 0030 0017 0008
ϑcoldstress 0172 0040 0258 0013
ϑprimeintercept 2292 0045 0426 0020
ϑprimehabitat 0316 0036 1210 0018
ϑprimeheatstress minus0006 0038 0054 0013
ϑprimecoldstress minus0622 0064 0876 0019
γintercept minus0884 0353 0010 0424
γ 1999 minus0601 0585 minus0150 0532
γ 2000 minus1083 0633 0516 0464
γ 2001 minus0300 0484 0228 0483
γ 2002 minus1113 0577 0262 0511
γ 2003 minus1055 0564 minus0311 0551
γ2004 minus0656 0525 minus0179 0534
γ 2005 minus0677 0533 0468 0485
γ2006 minus1014 0620 minus0217 0579
γ2007 minus0162 0453 minus0093 0548
γ 2008 minus23545 70483373 minus0478 0607
γ 2009 minus0428 0455 minus0253 0566
γ 2010 minus0292 0478 minus0159 0517
γ 2011 minus1136 0607 0012 0490
γ 2012 minus0498 0474 0141 0497
γaveragehabitat 0095 0091 0801 0112
γaverageheatstress minus0024 0067 0005 0062
γaveragecoldstress 0227 0071 0358 0120
εintercept minus3957 0386 minus3767 0242
ε 1999 0365 0482 minus0189 0337
ε 2000 0049 0512 minus0921 0399
ε 2001 minus0266 0548 minus0532 0362
ε 2002 minus0576 0660 minus1156 0398
ε 2003 minus0298 0595 minus0692 0355
ε2004 0113 0509 minus0621 0343
ε 2005 0173 0482 minus1096 0429
ε2006 minus0145 0561 minus1173 0463
ε2007 0451 0458 minus0894 0366
ε 2008 0283 0480 minus0976 0424
ε 2009 0218 0496 minus0898 0381
(Continues)
1992emsp |emsp emspensp CLEMENT ET aL
Parameters
Eastern Wood Pewee Red‐eyed Vireo
Estimate SE Estimate SE
ε 2010 minus0397 0605 minus0348 0304
ε 2011 0643 0455 minus0999 0406
ε 2012 0153 0644 minus0803 0410
εaveragehabitat 0238 0090 minus1670 0122
εaverageheatstress minus0071 0092 minus0075 0074
εaveragecoldstress minus0307 0141 minus0904 0104
p1intercept minus2172 0043 1352 0038
p1 1998 minus0118 0048 minus0127 0058
p1 1999 minus0112 0049 minus0043 0058
p1 2000 minus0166 0050 minus0016 0059
p1 2001 minus0156 0049 0031 0060
p1 2002 minus0179 0049 0076 0061
p1 2003 minus0216 0051 0161 0064
p12004 minus0242 0051 0066 0060
p1 2005 minus0124 0050 0149 0060
p12006 minus0182 0051 0124 0059
p12007 minus0149 0050 0184 0061
p1 2008 minus0202 0052 0192 0063
p1 2009 minus0221 0052 0135 0061
p1 2010 minus0195 0051 0062 0059
p1 2011 minus0122 0051 0203 0062
p1 2012 minus0227 0055 0194 0062
p1stopnumber 0033 0009 minus0055 0011
p1stopnumbersquared minus0069 0010 minus0247 0013
p2intercept minus0921 0018 minus0557 0036
p2 1998 minus0138 0033 minus0106 0050
p2 1999 minus0137 0032 0000 0052
p2 2000 minus0164 0032 minus0027 0051
p2 2001 minus0077 0032 minus0053 0053
p2 2002 minus0101 0032 minus0019 0052
p2 2003 minus0199 0033 0086 0053
p22004 minus0186 0032 0124 0057
p2 2005 minus0146 0032 0076 0051
p22006 minus0155 0032 0128 0054
p22007 minus0153 0032 0120 0054
p2 2008 minus0168 0032 0072 0056
p2 2009 minus0077 0033 0101 0055
p2 2010 minus0114 0033 0085 0060
p2 2011 minus0079 0032 0124 0054
p2 2012 minus0186 0033 0036 0061
p2stopnumber 0045 0007 0165 0010
p2stopnumbersquared minus0105 0008 minus0183 0010
π 0533 0064 minus0505 0065
TA B L E 1 emsp (Continued)
emspensp emsp | emsp1993CLEMENT ET aL
toModel3althoughnotsignificantlyduetothenumberofaddi‐tional parameters (χ2=2679df =28p=053)UnderModel 2estimatedcolonizationratesvariedfrom000in2008to030in1998whileextinctionratesvariedfrom001 in2001ndash2003and2010to004in2011WeusedModels45and6toestimatethepercentof improvedmodel fit (measuredby thechange indevi‐ance)generatedbyModel2thatcouldbeaccountedforbytempo‐ralvariationinclimateandhabitatcovariates(Table2)Temporalvariationinhabitataccountedfor04ofthechangeindeviancewhile temporal variation in climate accounted for 196Model6whichaccountedfortemporalvariationinbothhabitatandcli‐mateindicatedthatin1998colonizationinthesouthwestportionofthestudyareawasabovethelocalaveragewhileextinctioninthesouthwestwasbelowthelocalaverageindicatingarelativelystrongoccupancy trend in thesouthwest in thatyear (Figure3)By2012thispatternhadreversedyieldingarelativelyweakoc‐cupancytrendinthesouthwest(Figure3)NoneofthesemodelsrepresentedasignificantimprovementoverModel3(pgt026)Insomecasesthesignoftheestimatedeffectoftemporalvariationinclimateandhabitatwasoppositeoftheexpectedeffectbutinnoinstancesweretheseestimatessignificant(Table3)
32emsp|emspRed‐eyed Vireo
The best‐supportedmodel for Red‐eyedVireo included the samecovariatesasforEasternWoodPeweeyieldingaglobalmodelwith85parameters (Table1)TheHosmerndashLemeshowtest indicatedfitwasadequate for theRed‐eyedVireoglobalmodel forbothnaiumlvecolonization rates (χ2=540df =4p=025) and naiumlve extinctionrates(χ2=100df =4p=091)
Red‐eyedVireowere similarlywidespread in the study areawith an average probability of occupancy of 091 in the global
model (Figure1)Occupancydecreasedslightlyto090by2012incontrasttothe increase incountsreportedbytheBBS Initialoccupancywas positively related to available habitat and hoursofcoldstress (Table1)Probabilityofdetectionat the first stopwasrelativelyhighat038whileprobabilityofdetectionattheroute levelnearlyperfectat098Model3which includedspa‐tialvariationinclimateandhabitattothecolonizationandextinc‐tionmodelsimprovedmodelfitdramatically(χ2=41884df =6plt0001)relativetothenullmodelInkeepingwiththegreaterchangeindevianceforRed‐eyedVireoModel3estimatedmorespatialvariationinvitalratescomparedtoEasternWoodPeweeIn 1998 estimated colonization rates varied across BBS routesfrom 010 to 088 (mean of 049)while extinction rates rangedfrom000to048(meanof004Figure4)Model2whichallowedcolonization and extinction rates to vary each year improvedmodelfitrelativetoModel3butnotsignificantlyduetothenum‐berofadditionalparameters(χ2=3427df =28p=019)UnderModel2estimatedcolonizationratesvariedfrom039in2008to060 in2000while estimatedextinction rates varied from003in2006to007in1998WeusedModels45and6toestimatethepercentofthedeviancereductioninModel2thatcouldbeac‐countedforbytemporalvariationinclimateandhabitatcovariates(Table2)Wefoundthattemporalvariation inhabitataccountedfor01of the change indeviancewhile climate accounted for56Model 6 which accounted for temporal variation in bothhabitatandclimateindicatedthatin1998bothcolonizationandextinctionwereaboveaverageindicatinghigherturnoverearlyinthestudyperiod(Figure5)By2012bothcolonizationandextinc‐tion rateshaddeclined indicatinggreater stability inoccupancy(Figure 5) None of these models represented a significant im‐provementoverModel3(pgt074)Someoftheestimatedeffects
F I G U R E 1 emspOccupancyprobabilityfor(a)EasternWoodPeweeand(b)Red‐eyedVireointheeasternUnitedStates1997underModel2Occupancyvariesbytheamountofsuitablehabitathoursofheatstressandhoursofcoldstressin1997
1994emsp |emsp emspensp CLEMENT ET aL
oftemporalvariationinclimateandhabitatdifferedfromexpecta‐tionsbutnotsignificantly(Table3)
4emsp |emspDISCUSSION
Wefound that forbothbirdspecies initialoccupancycoloniza‐tionratesandextinctionratescorrelatedwithspatialvariationinclimateandhabitatcovariatesHowevercolonizationandextinc‐tionrateswerenotcorrelatedwithtemporalvariation inclimateand habitat covariates Our results correlating initial occupancywith environmental covariates are broadly speaking consistentwithbiologicalexpectationandaraftofpreviousstudiesthathaveindicatedasignificantrelationshipbetweenspeciesdistributionsandhabitat(RobinsonWilsonampCrick2001ThogmartinSauerampKnutson 2007) climate (Barbet‐Massinamp Jetz 2014BellardBertelsmeier Leadley Thuiller amp Courchamp 2012) or both(Seoane Bustamante amp Diacuteaz‐Delgado 2004 Sohl 2014)Howeverwesuggestthatestimatedrelationshipsbetweenspatialvariationinenvironmentalcovariatesandcolonizationandextinc‐tion rates are of greater ecological interest than relatively phe‐nomenological species distribution models or climate envelopemodels(Clementetal2016)Theestimatedvitalratesofcoloni‐zationandextinctionbringusclosertounderstandingthedynamicprocessunderlying thedistributionof these speciesGivencon‐stantvital rateswecanalsoproject theequilibrium levelofoc‐cupancyatsites ( i
i+i
atsite i)Wecancomparetheseprojected
equilibria tocurrentoccupancytoproject trends for thespeciesunder current environmental conditions We can also use
projectedchangesinkeyenvironmentalvariablestodeveloppre‐dicted rangechanges in the futureWeargue that theseprojec‐tions are more robust than those from static climate envelopemodels which assume that species are currently in equilibrium(Yackulicetal2015)Incontraststaticmodelscanoverestimaterange changes when the equilibrium does not hold (Morin ampThuiller2009)
Given the importanceofglobal changeweweremotivated toinvestigate factors affecting temporal aswell as spatial variationin vital rates In this case spatial variation must be incorporatedintosuchanalysesinordertoaccommodatethedifferentdynamicsexpectedindifferentportionsofaspeciesrangeWeviewthere‐lationshipbetweentemporalvariationincovariatevaluesandvitalrates as the most relevant to understanding the effect of globalchangeontherecentandfuturedistributionsofourstudyspeciesFurthermore a correlation between vital rates and spatially vary‐ingcovariatesmaynot indicateasimilarcorrelationwithtemporalchangesinthosesamecovariatesForexampleaspecieswithstrongdispersal constraintsmight have a strong spatial relationshipwithacovariatebutaweaktemporalrelationshipduetoits immobility(Devictor et al 2008) Similarly an estimated spatial relationshipmaybepurelyphenomenologicalduetothenatureofspatialvari‐ablesWhen spatial variables followgradients it is relatively easytoobtainstatisticallysignificantcorrelationsabsentanycausalre‐lationship(Grosboisetal2008)ForexampleifcolonizationratesandtemperaturesbothfollownorthndashsouthgradientstheymaybespatiallycorrelatedevenifcolonizationisgovernedbysomeotherunidentifiedfactorInthiscasewewouldexpecttheweakcorrela‐tion between temporal changes in temperature and colonization
F I G U R E 2 emspProbabilityof(a)colonizationand(b)extinctionforEasternWoodPeweeintheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1995CLEMENT ET aL
ratestobemoreindicativeofthetruerelationshipthanthestrongcorrelationwithspatialvariationintemperature
BBS surveys indicate different abundance trajectories for ourstudyspecieswithcounts increasingforRed‐eyedVireosandde‐creasing for EasternWood Pewees (Sauer et al 2015) Becauseabundanceandoccupancytendtobelinked(Gastonetal2000)weexpectedsomedivergenceinoccupancydynamicsWefoundthatestimatedoccupancywashighandstableforbothspeciesalthoughtherewasgreaterturnoverforRed‐eyedVireosForexampleunderModel3 (spatialvariationonly incovariates)bothaveragecoloni‐zationrates(049)andaverageextinctionrates(004)wereatleasttwiceashighforRed‐eyedVireoscomparedtoEasternWoodPewee(018and002respectively)Red‐eyedVireosexhibitedgreaterspa‐tialvariationincolonizationandextinctionrates(Figure4)althoughour selected covariates explained little temporal variation for thisspeciesEasternWoodPeweecolonizationandextinctionratesex‐hibitedmoreofalatitudinalgradientreflectingaspatialcorrelationwith extreme temperature (Figure 2)Our results also hinted at agreatercorrelationbetweentemporalchanges in temperatureandoccupancyvitalratesforEasternWoodPeweewhichcouldplayaroleinthedeclineofthisspeciesalthoughtheresultswerenotsig‐nificant(Table3)Alternatively ithasbeensuggestedthatEasternWoodPeweemaybedecliningduetoa lossofforestedwinteringhabitat(RobbinsSauerGreenbergampDroege1989)orduetodeer‐mediatedchanges in foreststructureonbreedinggrounds (deCal‐esta 1994) illustrating the complexity of ascertaining cause andeffectusingobservationalstudiesinthesenaturalsystems
Althoughrelativelyrare(Siramietal2017)otherstudieshaveinvestigated both climate and land cover effects on bird distribu‐tions Inagreementwithour finding that temporalvariation incli‐mate explainedmuchmore of themodel deviance than temporalvariation in habitat climate‐only species distribution models hadbetter cross‐validation support than habitat‐only models for 409Europeanbirdspecies(Barbet‐Massinetal2012)SimilarlymodelsestimatingthenumberofnewlyoccupiedareasbyHoodedWarblers(Wilsonia citrina) inOntariousingclimatedatahadbetter informa‐tioncriterionsupportthanmodelsusingforestdata(MellesFortinLindsay amp Badzinski 2011) In contrast land‐use models outper‐formedclimatemodels for18 farmlandbird species in theUnitedKingdom(EglingtonampPearce‐Higgins2012)Similarlyvegetation‐based species distributionmodels had superior performance thanclimate‐based models for 79 Spanish bird species (Seoane et al2004)andforthreebirdspeciesinNewMexico(FriggensampFinch2015) Two studies focused on colonization and extinction ratesfoundmixed results ForGardenWarblers (Sylvia borin) inBritainvegetationexplainedmorevariationincolonizationwhiletempera‐turesexplainedmorevariationinlocalextinction(MustinAmarampRedpath2014)For122birdspeciesinOntariothebest‐supportedcovariates (land cover or climate change) varied among species(YalcinampLeroux2018)
Thedivergentresultsamongstudiesregardingtheimportanceofclimateandlandcovermaypartlybeduetothebiologyofindividualspecies (Yalcinamp Leroux 2018) For example farmland birdsmayTA
BLE
2emspR2 DevfordifferentmodelsofcolonizationandextinctionprobabilitiesforbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012
Mod
el
Cova
riate
s on
colo
niza
tion
and
extin
ctio
nEa
ster
n W
ood
Pew
eeRe
d‐ey
ed V
ireo
Inte
rcep
tAv
erag
e H
abita
tAv
erag
e Cl
imat
eH
abita
t Dev
iatio
nCl
imat
e D
evia
tion
Year
Eff
ect
Δ minus
2LL
R2 Dev
Δ minus
2LL
R2 Dev
2X
XX
X0
0010
00
0010
0
6X
XX
XX
minus2138
202
minus3230
57
4X
XX
Xminus2154
196
minus3234
56
5X
XX
Xminus2667
04
minus3422
01
3X
XX
minus2679
0minus3427
0
1X
minus7098
NA
minus45311
NA
Not
eNAnotapplicable
1996emsp |emsp emspensp CLEMENT ET aL
beparticularlyaffectedbychangesinagriculturalintensitybecauseoftheirextensiveuseoffarms(EglingtonampPearce‐Higgins2012)Alternativelyithasbeenarguedthatspatialscalemayplayaroleintherelativeimportanceofglobalchangefactorswithclimatemoreimportantatlargescales(PearsonampDawson2003)Wenotethattwostudiesthatfoundalargerroleforclimatebothreliedonlower‐resolution bird surveys (05deg atlas in Barbet‐Massin et al 2012
10km2atlasinMellesetal2011)IntheBBSdatathatweanalyzedsurveyswereconductedevery800mbutwecalculatedoccupancyacross each 394km transect In addition the cited studies useddifferentdata sources anddiverse analysismethods complicatinginterpretationofthedifferingresults
While we estimated that climate explained more model de‐viance than habitat these estimated relationships were not
F I G U R E 3 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionrates((t)minus (t)minus )forEasternWoodPeweeintheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
emspensp emsp | emsp1997CLEMENT ET aL
TA B L E 3 emspEstimatedparametersforcorrelated‐detectiondynamicoccupancyModel6relatinghabitatandclimatecovariatestooccupancydynamicsofbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012Parameterestimatesindicatechangeinlogoddsofparametersinresponsetoz‐transformedcovariatesParametersψϑϑprimeγε p1p2andπdefinedintextForkeyparameters(boldtext)expectedsignsincluded
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
Ψintercept 2367 0177 4087 0381
Ψhabitat 0556 0163 2323 0288
Ψheatstress minus0011 0198 minus0322 0275
Ψcoldstress 0598 0166 1054 0251
ϑintercept minus1821 0026 minus1385 0010
ϑhabitat minus0055 0026 0516 0013
ϑheatstress minus0094 0030 0017 0008
ϑcoldstress 0172 0040 0256 0013
ϑprimeintercept 2292 0045 0426 0020
ϑprimehabitat 0316 0036 1210 0018
ϑprimeheatstress minus0006 0038 0054 0013
ϑprimecoldstress minus0622 0064 0875 0019
γintercept minus0884 0353 0023 0162
γhabitatdeviation + minus0292 0478 minus0240 1768
γheatstressdeviation minus minus1136 0607 0064 0194
γcoldstressdeviation minus minus0498 0474 2398 2253
γaveragehabitat 0095 0091 0788 0111
γaverageheatstress minus0024 0067 0004 0062
γaveragecoldstress 0227 0071 0333 0119
εintercept minus3957 0386 minus4471 0141
εhabitatdeviation minus minus0397 0605 0121 1492
εheatstressdeviation + 0643 0455 minus0034 0182
εcoldstressdeviation + 0153 0644 2042 2072
εaveragehabitat 0238 0090 minus1664 0123
εaverageheatstress minus0071 0092 minus0080 0074
εaveragecoldstress minus0307 0141 minus0939 0104
p1intercept minus2172 0043 1351 0039
p1 1998 minus0118 0048 minus0128 0058
p1 1999 minus0112 0049 minus0043 0058
p1 2000 minus0166 0050 minus0017 0059
p1 2001 minus0156 0049 0031 0061
p1 2002 minus0179 0049 0076 0062
p1 2003 minus0216 0051 0160 0064
p12004 minus0242 0051 0066 0061
p1 2005 minus0124 0050 0149 0061
p12006 minus0182 0051 0124 0060
p12007 minus0149 0050 0183 0062
p1 2008 minus0202 0052 0191 0064
p1 2009 minus0221 0052 0134 0062
p1 2010 minus0195 0051 0062 0060
p1 2011 minus0122 0051 0202 0063
(Continues)
1998emsp |emsp emspensp CLEMENT ET aL
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
p1 2012 minus0227 0055 0193 0063
p1stopnumber 0033 0009 minus0055 0011
p1stopnumbersquared minus0069 0010 minus0247 0013
p2intercept minus0921 0018 minus0555 0037
p2 1998 minus0138 0033 minus0121 0050
p2 1999 minus0137 0032 minus0025 0052
p2 2000 minus0164 0032 minus0027 0051
p2 2001 minus0077 0032 minus0059 0053
p2 2002 minus0101 0032 minus0012 0052
p2 2003 minus0199 0033 0085 0053
p22004 minus0186 0032 0113 0059
p2 2005 minus0146 0032 0082 0052
p22006 minus0155 0032 0135 0054
p22007 minus0153 0032 0124 0055
p2 2008 minus0168 0032 0073 0057
p2 2009 minus0077 0033 0100 0056
p2 2010 minus0114 0033 0071 0062
p2 2011 minus0079 0032 0120 0055
p2 2012 minus0186 0033 0030 0063
p2stopnumber 0045 0007 0165 0010
p2stopnumbersquared minus0105 0008 minus0182 0011
π 0533 0064 minus0505 0065
TA B L E 3 emsp (Continued)
F I G U R E 4 emspProbabilityof(a)colonizationand(b)extinctionforRed‐eyedVireointheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1999CLEMENT ET aL
significantanddidnotexplainalargefractionofvariationThereareseveralpotentialexplanationsforwhywedidnotfindasub‐stantialresponsetochangesinhabitatandclimateFirstourdatawerelackingindynamiceventstoinformourmodelsThecovari‐ateschangedrelativelylittleoverthetimeperiodmakingitdiffi‐culttodetectaresponseThelackofchangewaspartlybecauseoftherelativelyshort(16‐year)timeperiodstudiedAlsoweusedamovingaverageforclimatedatawhichtendstoreducedifferences
overshorttimeperiods Inadditionthestudyregionhasexperi‐encedlesswarmingoverthepastcenturythanotherpartsoftheplanet (Walshetal2014)Thebirddistributionsalso lackeddy‐namismBecauseextinctionrateswerelowtherewerefewlocalextinctioneventstoinformthemodelAlthoughcolonizationrateswerehighthefactthatinitialoccupancywashighandextinctionrateswerelowmeanttherewerefewopportunitiesforcoloniza‐tionevents
F I G U R E 5 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionratesforRed‐eyedVireointheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
2000emsp |emsp emspensp CLEMENT ET aL
Second measurement error may have obscured responses toglobalchangeThePRISMclimatedataareestimatedbyinterpola‐tionratherthandirectmeasurementacrossthestudyarea(Dalyetal2008)AssuchtheremaybeartifactsoranomaliesthatimpededourestimationandincreaseduncertaintySimilarlyNLCDdataareremotelysensedandmay includesomemeasurementerrorwhichcan increase uncertainty in our estimates (Homer et al 2015)Additionallylandcoverwasonlymeasuredinthreeyearsrequiringus to imputevalues in13yearsAsa result all landcover changethatoccurredbetween2002and2006wasrecordedin2002whilechangesbetween2007and2011wererecorded in2007Thisap‐proximation isnot idealbutwasnecessarygiventhe5‐yearreso‐lutionofthedataThislowerresolutionofthelandcoverdatamayhavereducedthesensitivityofthemodel
Third wemay havemisspecified ourmodelWe tried to de‐velopbiologicallymeaningfulcovariatesbutwemightnotunder‐standthesystemaswellaswehopedItispossiblethatbirdvitalrates respond to climateon adifferent time scale (ie not a15‐yearmoving average) or to climateduringdifferent timesof theyearortodifferenttemperaturethresholdsortodifferentclimatecomponents (egprecipitation Illaacutenetal2014) It isalsopossi‐ble that thebirds respond to finerhabitat features than the landcovertypesweusedoratadifferentspatialscale(ienot400mPearsonampDawson2003)Alternatively theremaybe importantregionaldifferencesor interactionsamongcovariatesthatwedidnotcaptureOritispossiblethatchangesinclimateandhabitatintheirbreedingareassimplyhavelittleeffectonvitalratesforthesespecies relative to other factors such as competitionwith otherbirdspeciesorglobalchangeoccurringontheirwinteringgroundsinSouthAmerica(Dormann2007)Ofcoursethesemisspecifica‐tionissuesareinherenttoallobservationalstudiesandnotuniquetothisstudy
Itwould be possible for us to explore some of themisspec‐ification issues just mentioned For example we could rerunthe samemodelsusingnewcovariates representing a varietyoftemperature thresholds and habitat buffer sizes to search for astrongercorrelationExperiencesuggeststhatpersistencewouldeventuallyberewardedwithap‐valuelt005ora lowmodelse‐lection valueHowever one reason to avoid such exploration isthat it tends to lead toType Ierrorsoverfitmodels andbiasedparameterestimates(FiebergampJohnson2015)Furthermorewefind thecontrastbetweensignificant correlations for spatial co‐variatesandnonsignificantcorrelationsfortemporalcovariatesil‐luminatingIntypicalclimateenvelopestudieswewouldestimatethe relationshipbetweencovariatesand thecurrentdistributionofaspeciesmuchlikeourestimateofinitialoccupancy(Dormann2007)BecauseweobtainedsignificantparameterestimateswemightconcludethatwehaveidentifiedcovariatesthatdeterminetherangeofthisspeciesThenwemightusetheseresultstoproj‐ectthefuturerangeofthespeciesundervariousclimatescenariosbased on the (questionable) assumption that the covariatendashspe‐cies relationship will not change as climate and habitat change(Cuddington et al 2013 Elith et al 2010Gustafson 2013) In
this case the insignificant correlationwith the temporal covari‐atessuggeststhatouroccupancycovariatesmerelydescribetherangeofthesebirdswithoutdeterminingtherangeFurthermoreour results suggest that despite significant results in the staticportionof themodelwehaveyet to identify themost relevantcomponents of climate change shaping thedistributionof thesebirds although itmay be difficult to identify these componentsgiven the lackofobservedchangeduringa relatively short timeperiodFinallyourresultssuggestthatprojectionsbasedonthestaticportionofourmodelareunlikelytobereliabledespitetheirstatisticalsignificance
Ifourinterpretationiscorrectitsuggeststhatsomeprojectionsof futurerangeshiftsbasedonstaticmodelsrelyonunsupportedassumptionsandarenot reliableWehavedemonstratedadiffer‐entapproachthatdoesnotrequirethestrongassumptionsofstaticmodelingapproachesOurapproachwouldbemoreapttoidentifyfactorsthatguidespeciesrsquoresponsetoglobalchangeifboththespe‐ciesandthecovariatesexhibitedmoredynamismandifweanalyzedalongertimeperiodthatencompassedgreaterchange
ACKNOWLEDG MENTS
WethankKeithPardieckDaveZiolkowskiJrandthethousandsofhighlyskilledvolunteerswhohavecoordinatedandperformedtheNorthAmericanBreedingBirdSurvey for half a centuryWealsothanktheClimateResearchUnitattheUniversityofEastAngliaformakingvaluableclimatedataeasilyaccessibleMJCwasfinanciallysupported by the BBS and the USGS Northeast Climate ScienceCenterThecontentsofthisdocumentaretheresponsibilityoftheauthorsanddonotnecessarilyrepresenttheviewsoftheUSGSAnyuseoftradeproductorfirmnamesisfordescriptivepurposesonlyanddoesnotimplyendorsementbytheUSGovernment
DEDIC ATION
We dedicate this paper to the late Chandler S Robbins ldquoFatherrdquooftheNorthAmericanBreedingBirdSurvey (BBS)DatafromtheBBSformthebasisforthisspecificpaperandforhundredsofothersfocusedon topics ranging from local trends tocontinentalmacro‐ecologyChanwasauniquecombinationofbird‐loverandscientistwhosevisionandinquisitiveenthusiasmwereaninspirationtoallofusfortunateenoughtohaveknownhim
CONFLIC T OF INTERE S T
Nonedeclared
AUTHORSrsquo CONTRIBUTION
JACandJDNconceivedthestudyAJTandSWanalyzedhabitatandclimatedataMJCJEHandJDNperformedtheoccupancyanaly‐sisMJCledthewritingofthemanuscriptAllauthorsgavefinalap‐provalforpublication
emspensp emsp | emsp2001CLEMENT ET aL
DATA ACCE SSIBILIT Y
Raw data are available from the following sources Breeding BirdSurvey httpswwwpwrcusgsgovbbsrawdata PRISM climatedata httpwwwprismoregonstateedu NLCD land cover datahttpswwwmrlcgov Processed data will be archived with theDryadDigitalRepository
ORCID
Matthew J Clement httpsorcidorg0000‐0003‐4231‐7949
James D Nichols httpsorcidorg0000‐0002‐7631‐2890
Adam J Terando httpsorcidorg0000‐0002‐9280‐043X
James E Hines httpsorcidorg0000‐0001‐5478‐7230
Steven G Williams httpsorcidorg0000‐0003‐3760‐6818
R E FE R E N C E S
Anderson D R Burnham K P Gould W R amp Cherry S (2001)Concerns about finding effects that are actually spuriousWildlife Society Bulletin29311ndash316
Arauacutejo M B Cabeza M Thuiller W Hannah L amp WilliamsP H (2004) Would climate change drive species out of re‐serves An assessment of existing reserve‐selection meth‐ods Global Change Biology 10 1618ndash1626 httpsdoiorg101111j1365‐2486200400828x
Barbet‐MassinMampJetzW(2014)A40‐yearcontinent‐widemulti‐speciesassessmentofrelevantclimatepredictorsforspeciesdistri‐butionmodellingDiversity and Distributions201285ndash1295httpsdoiorg101111ddi12229
Barbet‐Massin M Thuiller W amp Jiguet F (2012) The fate ofEuropean breeding birds under climate land‐use and disper‐sal scenarios Global Change Biology 18 881ndash890 httpsdoiorg101111j1365‐2486201102552x
Bellard C Bertelsmeier C Leadley P Thuiller W ampCourchamp F (2012) Impacts of climate change on the fu‐ture of biodiversity Ecology Letters 15 365ndash377 httpsdoiorg101111j1461‐0248201101736x
Boumlhning‐Gaese K amp Bauer H G (1996) Changes in species abun‐dance distribution and diversity in a central European birdcommunity Conservation Biology 10 175ndash187 httpsdoiorg101046j1523‐1739199610010175x
BrommerJELehikoinenAampValkamaJ(2012)Thebreedingrangesof central European and Arctic bird speciesmove poleward PLoS ONE7e43648httpsdoiorg101371journalpone0043648
CalderWAampKingJR(1974)ThermalandcaloricrelationsofbirdsInDSFarnerampJRKing(Eds)Avian biology(Vol4pp259ndash413)NewYorkNYAcademicPress
CesaraccioCSpanoDDucePampSnyderRL(2001)Animprovedmodel for determining degree‐day values from daily temperaturedataInternational Journal of Biometeorology45161ndash169httpsdoiorg101007s004840100104
ChamberlinTC (1890)ThemethodofmultipleworkinghypothesesScience1592ndash96
ClementMJHinesJENicholsJDPardieckKLampZiolkowskiDJJr (2016)Estimating indicesofrangeshifts inbirdsusingdy‐namicmodelswhendetectionisimperfectGlobal Change Biology223273ndash3285httpsdoiorg101111gcb13283
Commission for Environmental Cooperation (1997) Ecological regionsofNorthAmericaTowardacommonperspectiveCommission for
EnvironmentalCooperationMontrealRetrievedfromftpftpepagovwedecoregionscec_naCEC_NAecopdf
Crowl TA Crist TO Parmenter R R BelovskyGamp LugoA E(2008)Thespreadofinvasivespeciesandinfectiousdiseaseasdriv‐ersofecosystemchangeFrontiers in Ecology and the Environment6238ndash246httpsdoiorg101890070151
Cuddington K FortinM J Gerber L RHastings A Liebhold AOConnorMampRayC(2013)Process‐basedmodelsarerequiredtomanageecologicalsystemsinachangingworldEcosphere41ndash12httpsdoiorg101890ES12‐001781
DalyCHalbleibMSmithJIGibsonWPDoggettMKTaylorGHhellipPasterisPP(2008)Physiographicallysensitivemappingofcli‐matologicaltemperatureandprecipitationacrosstheconterminousUnited States International Journal of Climatology 28 2031ndash2064httpsdoiorg101002joc1688
DawsonW (1958) Relation of oxygen consumption and evaporativewater losstotemperatureintheCardinalPhysiological Zoology3137ndash48httpsdoiorg101086physzool31130155377
DawsonWRampWhittowGC (2000)Regulationofbodytempera‐ture InG CWhittow (Ed)Sturkiersquos avian physiology (5th ed pp343ndash390)SanDiegoCAAcademicPress
DeCalestaDS(1994)Effectofwhite‐taileddeeronsongbirdswithinmanagedforestsinPennsylvaniaJournal of Wildlife Management58711ndash718httpsdoiorg1023073809685
DevictorVJulliardRCouvetDampJiguetF(2008)Birdsaretrack‐ing climatewarming but not fast enoughProceedings of the Royal Society of London B Biological Sciences2752743ndash2748httpsdoiorg101098rspb20080878
Dormann C F (2007) Promising the future Global change projec‐tionsofspeciesdistributionsBasic and Applied Ecology8387ndash397httpsdoiorg101016jbaae200611001
Dormann C F Schymanski S J Cabral J Chuine I Graham CHartigFhellipSingerA(2012)Correlationandprocessinspeciesdis‐tributionmodelsBridgingadichotomyJournal of Biogeography392119ndash2131httpsdoiorg101111j1365‐2699201102659x
EccelE(2010)WhatwecanasktohourlytemperaturerecordingPartIIHourlyinterpolationoftemperaturesforclimatologyandmodel‐lingItalian Journal of Agrometeorology1545ndash50
EccelEampCordanoE(2013)InterpolTHourlyinterpolationofmulti‐pletemperaturedailyseriesRpackageversion211RetrievedfromhttpsCRANR‐projectorgpackage=InterpolT
EglingtonSMampPearce‐HigginsJW(2012)Disentanglingtherela‐tiveimportanceofchangesinclimateandland‐useintensityindriv‐ingrecentbirdpopulationtrendsPLoS ONE7e30407httpsdoiorg101371journalpone0030407
Elith JKearneyMampPhillipsS (2010)Theartofmodelling range‐shifting species Methods in Ecology and Evolution 1 330ndash342httpsdoiorg101111j2041‐210X201000036x
FiebergJampJohnsonDH(2015)MMIMultimodelinferenceormod‐elswithmanagement implications Journal of Wildlife Management79708ndash718
FriggensMMampFinchDM (2015) Implicationsof climatechangeforbirdconservationinthesouthwesternUSunderthreealternativefuturesPLoS ONE10e0144089
FryJXianGZJinSDewitzJHomerCGYangLhellipWickhamJD (2011)Completionof the2006national landcoverdatabasefor theconterminousUnitedStatesPhotogrammetric Engineering amp Remote Sensing77858ndash864
Fuller R J Gregory R D Gibbons D W Marchant J HWilson J D Baillie S R amp Carter N (1995) Population de‐clines and range contractions among lowland farmland birdsin Britain Conservation Biology 9 1425ndash1441 httpsdoiorg101046j1523‐1739199509061425x
Gaston K J Blackburn T M Greenwood J J Gregory R DQuinn R M amp Lawton J H (2000) Abundancendashoccupancy
2002emsp |emsp emspensp CLEMENT ET aL
relationships Journal of Applied Ecology 37 39ndash59 httpsdoiorg101046j1365‐2664200000485x
GoldsteinRB(1974)RelationofmetabolismtoambienttemperatureintheverdinCondor76116ndash119httpsdoiorg1023071365995
GrinnellJ(1917)FieldtestsoftheoriesconcerningdistributionalcontrolAmerican Naturalist51115ndash128httpsdoiorg101086279591
GrosboisVGimenezOGaillardJMPradelRBarbraudCClobertJhellipWeimerskirchH(2008)Assessingtheimpactofclimatevari‐ation on survival in vertebrate populationsBiological Reviews 83357ndash399httpsdoiorg101111j1469‐185X200800047x
GustafsonEJ(2013)WhenrelationshipsestimatedinthepastcannotbeusedtopredictthefutureUsingmechanisticmodelstopredictlandscapeecologicaldynamicsinachangingworldLandscape Ecology281429ndash1437httpsdoiorg101007s10980‐013‐9927‐4
Hamel P B (1992) LandManagers guide to the birds of the SouthGeneralTechnicalReportSE‐22USDAForestServiceSoutheasternForestExperimentStationandTheNatureConservancyChapelHillNC
Hinds D S amp Calder W A (1973) Temperature regulation of thePyrrhuloxiaandtheArizonaCardinalPhysiological Zoology4655ndash71httpsdoiorg101086physzool46130152517
HinesJE(2006)PRESENCE Software to estimate patch occupancy and related parameters Laurel MD US Geological survey PatuxentWildlife Research Center Retrieved from httpwwwmbr‐pwrcusgsgovsoftwarepresencehtml
HinesJENicholsJDampCollazoJA(2014)Multiseasonoccupancymodels for correlated replicate surveys Methods in Ecology and Evolution5583ndash591httpsdoiorg1011112041‐210X12186
HinesJENicholsJDRoyleJAMacKenzieDIGopalaswamyAMKumarNampKaranthKU(2010)TigersontrailsOccupancymodeling for cluster sampling Ecological Applications 20 1456ndash1466httpsdoiorg10189009‐03211
HockeyPA SiramiCRidleyARMidgleyGFampBabikerHA(2011) Interrogating recent range changes in South African birdsConfounding signals from land use and climate change present achallenge for attribution Diversity and Distributions 17 254ndash261httpsdoiorg101111j1472‐4642201000741x
Hoegh‐Guldberg O Hughes L McIntyre S Lindenmayer D BParmesanCPossinghamHPampThomasCD (2008)AssistedcolonizationandrapidclimatechangeScience321345ndash346
Homer C Dewitz J Fry J Coan M Hossain N Larson C hellipWickham J (2007) Completion of the 2001 national land coverdatabase for the conterminous United States Photogrammetric Engineering amp Remote Sensing73337
HomerCDewitzJYangLJinSDanielsonPXianGhellipMegownK(2015)Completionofthe2011NationalLandCoverDatabasefortheconterminousUnitedStatesndashrepresentingadecadeoflandcoverchange informationPhotogrammetric Engineering amp Remote Sensing81345ndash354
HootenMBWikleCKDorazioRMampRoyleJA(2007)HierarchicalspatiotemporalmatrixmodelsforcharacterizinginvasionsBiometrics63558ndash567httpsdoiorg101111j1541‐0420200600725x
HosmerDWJrampLemeshowS(2000)Applied logistic regression(2nded)NewYorkNYJohnWileyampSons
Huntley B Collingham Y CGreen R EHiltonGM RahbekCamp Willis S G (2006) Potential impacts of climatic change upongeographical distributions of birds Ibis 148 8ndash28 httpsdoiorg101111j1474‐919X200600523x
IgleciaMNCollazoJAampMcKerrowAJ(2012)Useofoccupancymodels to evaluate expert knowledge‐based species‐habitat rela‐tionshipsAvian Conservation and Ecology7(2)13pRetrievedfromhttpwwwace‐ecoorgvol7iss2art5
IllaacutenJGThomasCDJonesJAWongWKShirleySMampBettsMG(2014)Precipitationandwintertemperaturepredictlong‐termrange‐scale abundancechanges inWesternNorthAmericanbirds
Global Change Biology 20 3351ndash3364 httpsdoiorg101111gcb12642
IoannidisJP (2005)WhymostpublishedresearchfindingsarefalsePLoS Medicine2e124
KeacuteryMGuillera‐ArroitaGampLahoz‐Monfort J J (2013)Analysingand mapping species range dynamics using occupancy modelsJournal of Biogeography 40 1463ndash1474 httpsdoiorg101111jbi12087
Khaliq I Hof C Prinzinger R Boumlhning‐Gaese K amp PfenningerM(2014) Global variation in thermal tolerances and vulnerability ofendotherms to climate change Proceedings of the Royal Society B Biological Sciences 281(1789) 20141097 httpsdoiorg101098rspb20141097
Laursen K amp Frikke J (2008) Hunting from motorboats displacesWadden Sea eiders Somateria mollissima from their favouredfeeding distribution Wildlife Biology 14 423ndash433 httpsdoiorg1029810909‐6396‐144423
LemoineNBauerHGPeintingerMampBoumlhning‐GaeseK (2007)Effectsof climate and land‐use changeon species abundance in acentralEuropeanbirdcommunityConservation Biology21495ndash503httpsdoiorg101111j1523‐1739200600633x
MacArthurRH(1972)Geographical ecology Patterns in the distribution of speciesNewYorkNYHarperampRow
MacKenzieDINicholsJDHinesJEKnutsonMGampFranklinAB(2003)Estimatingsiteoccupancycolonizationandlocalextinc‐tionwhenaspeciesisdetectedimperfectlyEcology842200ndash2207httpsdoiorg10189002‐3090
MacKenzieDINicholsJDLachmanGBDroegeSRoyleJAampLangtimmCA(2002)Estimatingsiteoccupancyrateswhendetec‐tionprobabilitiesarelessthanoneEcology832248ndash2255httpsdoiorg1018900012‐9658(2002)083[2248ESORWD]20CO2
MacKenzieDINicholsJDRoyleJAPollockKHBaileyLampHinesJE(2018)Occupancy estimation and modeling Inferring pat‐terns and dynamics of species occurrence (2nd ed) BurlingtonMAAcademicPress
MellesSJFortinMJLindsayKampBadzinskiD(2011)ExpandingnorthwardInfluenceofclimatechangeforestconnectivityandpop‐ulationprocessesonathreatenedspeciesrangeshiftGlobal Change Biology1717ndash31httpsdoiorg101111j1365‐2486201002214x
Morin X amp Thuiller W (2009) Comparing niche‐and process‐based models to reduce prediction uncertainty in species rangeshifts under climate change Ecology 90 1301ndash1313 httpsdoiorg10189008‐01341
MustinKAmarAampRedpathSM(2014)Colonizationandextinctiondynamicsofadecliningmigratorybirdareinfluencedbyclimateandhabitat degradation Ibis 156 788ndash798 httpsdoiorg101111ibi12173
NicholsJDThomasLampConnPB(2009)InferencesaboutlandbirdabundancefromcountdataRecentadvancesandfuturedirectionsInDLThomsonEJCoochampMJConroy(Eds)Modeling demo‐graphic processes in marked populations(pp201ndash235)NewYorkNYSpringer
OmernikJM(1995)EcoregionsAspatialframeworkforenvironmen‐talmanagement InWSDavisampTSimon(Eds)Biological assess‐ment and criteria Tools for water resource planning and decision making (pp49ndash62)BocaRatonFLLewisPublishers
OmernikJM(2004)Perspectivesonthenatureanddefinitionofeco‐logicalregionsEnvironmental Management34(Supplement1)S27ndashS38httpsdoiorg101007s00267‐003‐5197‐2
Omernik J M amp Griffith G E (2014) Ecoregions of the contermi‐nousUnited States Evolution of a hierarchical spatial frameworkEnvironmental Management541249ndash1266httpsdoiorg101007s00267‐014‐0364‐1
Pardieck K L ZiolkowskiD J JrampHudsonM‐A R (2015)NorthAmericanBreedingBirdSurveyDataset1966ndash2014version20140
emspensp emsp | emsp2003CLEMENT ET aL
USGeologicalSurveyPatuxentWildlifeResearchCenterRetrievedfromhttpswwwpwrcusgsgovbbsrawdata
ParmesanCampYoheG (2003)Agloballycoherentfingerprintofcli‐mate change impacts across natural systemsNature 421 37ndash42httpsdoiorg101038nature01286
PearsonRGampDawsonTP(2003)PredictingtheimpactsofclimatechangeonthedistributionofspeciesArebioclimateenvelopemod‐elsusefulGlobal Ecology and Biogeography12361ndash371
PlattJR(1964)StronginferenceScience146347ndash353PopperK(1959)The logic of scientific discoveryLondonUKHutchinsonRobbins C S Sauer J R Greenberg R S amp Droege S (1989)
Population declines in North American birds that migrate to theNeotropics Proceedings of the National Academy of Sciences 867658ndash7662httpsdoiorg101073pnas86197658
Robinson R A Wilson J D amp Crick H Q (2001) The impor‐tance of arable habitat for farmland birds in grassland land‐scapes Journal of Applied Ecology 38 1059ndash1069 httpsdoiorg101046j1365‐2664200100654x
Romesburg H C (1981) Wildlife science Gaining reliable knowl‐edge Journal of Wildlife Management 45 293ndash313 httpsdoiorg1023073807913
Root T (1988) Energy constraints on avian distributions and abun‐dancesEcology69330ndash339httpsdoiorg1023071940431
Ruiz‐GutieacuterrezVampZipkinEF (2011)Detectionbiasesyieldmis‐leadingpatternsof speciespersistence and colonization in frag‐mented landscapesEcosphere2 1ndash14 httpsdoiorg101890ES10‐002071
SauerJRHinesJEFallonJEPardieckKLZiolkowskiDJJrampLinkWA(2015)The North American breeding bird survey results and analysis 1966ndash2013 Version 01302015LaurelMDUSGSPatuxentWildlifeResearchCenterhttpswwwmbr‐pwrcusgsgovbbsbbshtml
Sauer J R LinkWA Fallon J E Pardieck K L amp ZiolkowskiDJJr(2013)TheNorthAmericanbreedingbirdsurvey1966ndash2011SummaryanalysisandspeciesaccountsNorth American Fauna791ndash32httpsdoiorg103996nafa790001
SeoaneJBustamanteJampDiacuteaz‐DelgadoR(2004)Competingrolesforlandscapevegetationtopographyandclimateinpredictivemod‐elsofbirddistributionEcological Modelling171209ndash222httpsdoiorg101016jecolmodel200308006
Sirami C Caplat P Popy S Clamens A Arlettaz R Jiguet F hellipMartin J‐L (2017) Impacts of global change on species distri‐butions Obstacles and solutions to integrate climate and landuse Global Ecology and Biogeography 26 385ndash394 httpsdoiorg101111geb12555
SkalskiJR(1996)Regressionofabundanceestimatesfrommarkrecap‐ture surveys against environmental covariatesCanadian Journal of
Fisheries and Aquatic Sciences53196ndash204httpsdoiorg101139f95‐169
SohlTL(2014)Therelativeimpactsofclimateandland‐usechangeonconterminousUnitedStatesbird species from2001 to2075PLoS ONE9e112251httpsdoiorg101371journalpone0112251
ThogmartinWESauerJRampKnutsonMG(2007)ModelingandmappingabundanceofAmericanwoodcockacrosstheMidwesternandNortheasternUnitedStatesJournal of Wildlife Management71376ndash382httpsdoiorg1021932005‐680
ThomasCDampLennonJJ (1999)Birdsextendtheirrangesnorth‐wardsNature399213httpsdoiorg10103820335
Trathan P N Garciacutea‐Borboroglu P Boersma D Bost C ACrawford R J CrossinG ThellipWienecke B (2015) Pollutionhabitatlossfishingandclimatechangeascriticalthreatstopen‐guins Conservation Biology 29 31ndash41 httpsdoiorg101111cobi12349
TravisJMJ(2003)Climatechangeandhabitatdestructionadeadlyanthropogenic cocktail Proceedings of the Royal Society of London Series B Biological Sciences 270(1514) 467ndash473 httpsdoiorg101098rspb20022246
UrbanMC (2015)Acceleratingextinction risk fromclimatechangeScience348571ndash573httpsdoiorg101126scienceaaa4984
WalshJWuebblesDHayhoeKKossinJKunkelKStephensGhellipSomervilleR(2014)OurchangingclimateInJMMelilloTCRichmondampGWYohe(Eds)Climate change impacts in the United States The Third National climate assessment(pp19ndash67)WashingtonDCUSGovernmentPrintingOffice
YackulicCBNichols JDReid JampDerR (2015)Topredict thenichemodelcolonizationandextinctionEcology9616ndash23httpsdoiorg10189014‐13611
Yalcin S amp Leroux S J (2018) An empirical test of the relative andcombinedeffectsofland‐coverandclimatechangeonlocalcoloni‐zationandextinctionGlobal Change Biology243849ndash3861httpsdoiorg101111gcb14169
ZhuKWoodallCWampClarkJS(2012)FailuretomigrateLackoftreerangeexpansioninresponsetoclimatechangeGlobal Change Biology181042ndash1052httpsdoiorg101111j1365‐2486201102571x
How to cite this articleClementMJNicholsJDCollazoJATerandoAJHinesJEWilliamsSGPartitioningglobalchangeAssessingtherelativeimportanceofchangesinclimateandlandcoverforchangesinaviandistributionEcol Evol 201991985ndash2003 httpsdoiorg101002ece34890

emspensp emsp | emsp1991CLEMENT ET aL
TA B L E 1 emspEstimatedparametersforcorrelated‐detectiondynamicoccupancyModel2relatinghabitatandclimatecovariatestooccupancydynamicsofbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012Parameterestimatesindicatechangeinlogoddsofparametersinresponsetoz‐transformedcovariatesParametersψϑϑprimeγεp1p2andπdefinedintext
Parameters
Eastern Wood Pewee Red‐eyed Vireo
Estimate SE Estimate SE
Ψintercept 2367 0177 4136 0404
Ψhabitat 0556 0163 2315 0300
Ψheatstress minus0011 0198 minus0337 0281
Ψcoldstress 0598 0166 1020 0258
ϑintercept minus1821 0026 minus1385 0010
ϑhabitat minus0055 0026 0516 0013
ϑheatstress minus0094 0030 0017 0008
ϑcoldstress 0172 0040 0258 0013
ϑprimeintercept 2292 0045 0426 0020
ϑprimehabitat 0316 0036 1210 0018
ϑprimeheatstress minus0006 0038 0054 0013
ϑprimecoldstress minus0622 0064 0876 0019
γintercept minus0884 0353 0010 0424
γ 1999 minus0601 0585 minus0150 0532
γ 2000 minus1083 0633 0516 0464
γ 2001 minus0300 0484 0228 0483
γ 2002 minus1113 0577 0262 0511
γ 2003 minus1055 0564 minus0311 0551
γ2004 minus0656 0525 minus0179 0534
γ 2005 minus0677 0533 0468 0485
γ2006 minus1014 0620 minus0217 0579
γ2007 minus0162 0453 minus0093 0548
γ 2008 minus23545 70483373 minus0478 0607
γ 2009 minus0428 0455 minus0253 0566
γ 2010 minus0292 0478 minus0159 0517
γ 2011 minus1136 0607 0012 0490
γ 2012 minus0498 0474 0141 0497
γaveragehabitat 0095 0091 0801 0112
γaverageheatstress minus0024 0067 0005 0062
γaveragecoldstress 0227 0071 0358 0120
εintercept minus3957 0386 minus3767 0242
ε 1999 0365 0482 minus0189 0337
ε 2000 0049 0512 minus0921 0399
ε 2001 minus0266 0548 minus0532 0362
ε 2002 minus0576 0660 minus1156 0398
ε 2003 minus0298 0595 minus0692 0355
ε2004 0113 0509 minus0621 0343
ε 2005 0173 0482 minus1096 0429
ε2006 minus0145 0561 minus1173 0463
ε2007 0451 0458 minus0894 0366
ε 2008 0283 0480 minus0976 0424
ε 2009 0218 0496 minus0898 0381
(Continues)
1992emsp |emsp emspensp CLEMENT ET aL
Parameters
Eastern Wood Pewee Red‐eyed Vireo
Estimate SE Estimate SE
ε 2010 minus0397 0605 minus0348 0304
ε 2011 0643 0455 minus0999 0406
ε 2012 0153 0644 minus0803 0410
εaveragehabitat 0238 0090 minus1670 0122
εaverageheatstress minus0071 0092 minus0075 0074
εaveragecoldstress minus0307 0141 minus0904 0104
p1intercept minus2172 0043 1352 0038
p1 1998 minus0118 0048 minus0127 0058
p1 1999 minus0112 0049 minus0043 0058
p1 2000 minus0166 0050 minus0016 0059
p1 2001 minus0156 0049 0031 0060
p1 2002 minus0179 0049 0076 0061
p1 2003 minus0216 0051 0161 0064
p12004 minus0242 0051 0066 0060
p1 2005 minus0124 0050 0149 0060
p12006 minus0182 0051 0124 0059
p12007 minus0149 0050 0184 0061
p1 2008 minus0202 0052 0192 0063
p1 2009 minus0221 0052 0135 0061
p1 2010 minus0195 0051 0062 0059
p1 2011 minus0122 0051 0203 0062
p1 2012 minus0227 0055 0194 0062
p1stopnumber 0033 0009 minus0055 0011
p1stopnumbersquared minus0069 0010 minus0247 0013
p2intercept minus0921 0018 minus0557 0036
p2 1998 minus0138 0033 minus0106 0050
p2 1999 minus0137 0032 0000 0052
p2 2000 minus0164 0032 minus0027 0051
p2 2001 minus0077 0032 minus0053 0053
p2 2002 minus0101 0032 minus0019 0052
p2 2003 minus0199 0033 0086 0053
p22004 minus0186 0032 0124 0057
p2 2005 minus0146 0032 0076 0051
p22006 minus0155 0032 0128 0054
p22007 minus0153 0032 0120 0054
p2 2008 minus0168 0032 0072 0056
p2 2009 minus0077 0033 0101 0055
p2 2010 minus0114 0033 0085 0060
p2 2011 minus0079 0032 0124 0054
p2 2012 minus0186 0033 0036 0061
p2stopnumber 0045 0007 0165 0010
p2stopnumbersquared minus0105 0008 minus0183 0010
π 0533 0064 minus0505 0065
TA B L E 1 emsp (Continued)
emspensp emsp | emsp1993CLEMENT ET aL
toModel3althoughnotsignificantlyduetothenumberofaddi‐tional parameters (χ2=2679df =28p=053)UnderModel 2estimatedcolonizationratesvariedfrom000in2008to030in1998whileextinctionratesvariedfrom001 in2001ndash2003and2010to004in2011WeusedModels45and6toestimatethepercentof improvedmodel fit (measuredby thechange indevi‐ance)generatedbyModel2thatcouldbeaccountedforbytempo‐ralvariationinclimateandhabitatcovariates(Table2)Temporalvariationinhabitataccountedfor04ofthechangeindeviancewhile temporal variation in climate accounted for 196Model6whichaccountedfortemporalvariationinbothhabitatandcli‐mateindicatedthatin1998colonizationinthesouthwestportionofthestudyareawasabovethelocalaveragewhileextinctioninthesouthwestwasbelowthelocalaverageindicatingarelativelystrongoccupancy trend in thesouthwest in thatyear (Figure3)By2012thispatternhadreversedyieldingarelativelyweakoc‐cupancytrendinthesouthwest(Figure3)NoneofthesemodelsrepresentedasignificantimprovementoverModel3(pgt026)Insomecasesthesignoftheestimatedeffectoftemporalvariationinclimateandhabitatwasoppositeoftheexpectedeffectbutinnoinstancesweretheseestimatessignificant(Table3)
32emsp|emspRed‐eyed Vireo
The best‐supportedmodel for Red‐eyedVireo included the samecovariatesasforEasternWoodPeweeyieldingaglobalmodelwith85parameters (Table1)TheHosmerndashLemeshowtest indicatedfitwasadequate for theRed‐eyedVireoglobalmodel forbothnaiumlvecolonization rates (χ2=540df =4p=025) and naiumlve extinctionrates(χ2=100df =4p=091)
Red‐eyedVireowere similarlywidespread in the study areawith an average probability of occupancy of 091 in the global
model (Figure1)Occupancydecreasedslightlyto090by2012incontrasttothe increase incountsreportedbytheBBS Initialoccupancywas positively related to available habitat and hoursofcoldstress (Table1)Probabilityofdetectionat the first stopwasrelativelyhighat038whileprobabilityofdetectionattheroute levelnearlyperfectat098Model3which includedspa‐tialvariationinclimateandhabitattothecolonizationandextinc‐tionmodelsimprovedmodelfitdramatically(χ2=41884df =6plt0001)relativetothenullmodelInkeepingwiththegreaterchangeindevianceforRed‐eyedVireoModel3estimatedmorespatialvariationinvitalratescomparedtoEasternWoodPeweeIn 1998 estimated colonization rates varied across BBS routesfrom 010 to 088 (mean of 049)while extinction rates rangedfrom000to048(meanof004Figure4)Model2whichallowedcolonization and extinction rates to vary each year improvedmodelfitrelativetoModel3butnotsignificantlyduetothenum‐berofadditionalparameters(χ2=3427df =28p=019)UnderModel2estimatedcolonizationratesvariedfrom039in2008to060 in2000while estimatedextinction rates varied from003in2006to007in1998WeusedModels45and6toestimatethepercentofthedeviancereductioninModel2thatcouldbeac‐countedforbytemporalvariationinclimateandhabitatcovariates(Table2)Wefoundthattemporalvariation inhabitataccountedfor01of the change indeviancewhile climate accounted for56Model 6 which accounted for temporal variation in bothhabitatandclimateindicatedthatin1998bothcolonizationandextinctionwereaboveaverageindicatinghigherturnoverearlyinthestudyperiod(Figure5)By2012bothcolonizationandextinc‐tion rateshaddeclined indicatinggreater stability inoccupancy(Figure 5) None of these models represented a significant im‐provementoverModel3(pgt074)Someoftheestimatedeffects
F I G U R E 1 emspOccupancyprobabilityfor(a)EasternWoodPeweeand(b)Red‐eyedVireointheeasternUnitedStates1997underModel2Occupancyvariesbytheamountofsuitablehabitathoursofheatstressandhoursofcoldstressin1997
1994emsp |emsp emspensp CLEMENT ET aL
oftemporalvariationinclimateandhabitatdifferedfromexpecta‐tionsbutnotsignificantly(Table3)
4emsp |emspDISCUSSION
Wefound that forbothbirdspecies initialoccupancycoloniza‐tionratesandextinctionratescorrelatedwithspatialvariationinclimateandhabitatcovariatesHowevercolonizationandextinc‐tionrateswerenotcorrelatedwithtemporalvariation inclimateand habitat covariates Our results correlating initial occupancywith environmental covariates are broadly speaking consistentwithbiologicalexpectationandaraftofpreviousstudiesthathaveindicatedasignificantrelationshipbetweenspeciesdistributionsandhabitat(RobinsonWilsonampCrick2001ThogmartinSauerampKnutson 2007) climate (Barbet‐Massinamp Jetz 2014BellardBertelsmeier Leadley Thuiller amp Courchamp 2012) or both(Seoane Bustamante amp Diacuteaz‐Delgado 2004 Sohl 2014)Howeverwesuggestthatestimatedrelationshipsbetweenspatialvariationinenvironmentalcovariatesandcolonizationandextinc‐tion rates are of greater ecological interest than relatively phe‐nomenological species distribution models or climate envelopemodels(Clementetal2016)Theestimatedvitalratesofcoloni‐zationandextinctionbringusclosertounderstandingthedynamicprocessunderlying thedistributionof these speciesGivencon‐stantvital rateswecanalsoproject theequilibrium levelofoc‐cupancyatsites ( i
i+i
atsite i)Wecancomparetheseprojected
equilibria tocurrentoccupancytoproject trends for thespeciesunder current environmental conditions We can also use
projectedchangesinkeyenvironmentalvariablestodeveloppre‐dicted rangechanges in the futureWeargue that theseprojec‐tions are more robust than those from static climate envelopemodels which assume that species are currently in equilibrium(Yackulicetal2015)Incontraststaticmodelscanoverestimaterange changes when the equilibrium does not hold (Morin ampThuiller2009)
Given the importanceofglobal changeweweremotivated toinvestigate factors affecting temporal aswell as spatial variationin vital rates In this case spatial variation must be incorporatedintosuchanalysesinordertoaccommodatethedifferentdynamicsexpectedindifferentportionsofaspeciesrangeWeviewthere‐lationshipbetweentemporalvariationincovariatevaluesandvitalrates as the most relevant to understanding the effect of globalchangeontherecentandfuturedistributionsofourstudyspeciesFurthermore a correlation between vital rates and spatially vary‐ingcovariatesmaynot indicateasimilarcorrelationwithtemporalchangesinthosesamecovariatesForexampleaspecieswithstrongdispersal constraintsmight have a strong spatial relationshipwithacovariatebutaweaktemporalrelationshipduetoits immobility(Devictor et al 2008) Similarly an estimated spatial relationshipmaybepurelyphenomenologicalduetothenatureofspatialvari‐ablesWhen spatial variables followgradients it is relatively easytoobtainstatisticallysignificantcorrelationsabsentanycausalre‐lationship(Grosboisetal2008)ForexampleifcolonizationratesandtemperaturesbothfollownorthndashsouthgradientstheymaybespatiallycorrelatedevenifcolonizationisgovernedbysomeotherunidentifiedfactorInthiscasewewouldexpecttheweakcorrela‐tion between temporal changes in temperature and colonization
F I G U R E 2 emspProbabilityof(a)colonizationand(b)extinctionforEasternWoodPeweeintheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1995CLEMENT ET aL
ratestobemoreindicativeofthetruerelationshipthanthestrongcorrelationwithspatialvariationintemperature
BBS surveys indicate different abundance trajectories for ourstudyspecieswithcounts increasingforRed‐eyedVireosandde‐creasing for EasternWood Pewees (Sauer et al 2015) Becauseabundanceandoccupancytendtobelinked(Gastonetal2000)weexpectedsomedivergenceinoccupancydynamicsWefoundthatestimatedoccupancywashighandstableforbothspeciesalthoughtherewasgreaterturnoverforRed‐eyedVireosForexampleunderModel3 (spatialvariationonly incovariates)bothaveragecoloni‐zationrates(049)andaverageextinctionrates(004)wereatleasttwiceashighforRed‐eyedVireoscomparedtoEasternWoodPewee(018and002respectively)Red‐eyedVireosexhibitedgreaterspa‐tialvariationincolonizationandextinctionrates(Figure4)althoughour selected covariates explained little temporal variation for thisspeciesEasternWoodPeweecolonizationandextinctionratesex‐hibitedmoreofalatitudinalgradientreflectingaspatialcorrelationwith extreme temperature (Figure 2)Our results also hinted at agreatercorrelationbetweentemporalchanges in temperatureandoccupancyvitalratesforEasternWoodPeweewhichcouldplayaroleinthedeclineofthisspeciesalthoughtheresultswerenotsig‐nificant(Table3)Alternatively ithasbeensuggestedthatEasternWoodPeweemaybedecliningduetoa lossofforestedwinteringhabitat(RobbinsSauerGreenbergampDroege1989)orduetodeer‐mediatedchanges in foreststructureonbreedinggrounds (deCal‐esta 1994) illustrating the complexity of ascertaining cause andeffectusingobservationalstudiesinthesenaturalsystems
Althoughrelativelyrare(Siramietal2017)otherstudieshaveinvestigated both climate and land cover effects on bird distribu‐tions Inagreementwithour finding that temporalvariation incli‐mate explainedmuchmore of themodel deviance than temporalvariation in habitat climate‐only species distribution models hadbetter cross‐validation support than habitat‐only models for 409Europeanbirdspecies(Barbet‐Massinetal2012)SimilarlymodelsestimatingthenumberofnewlyoccupiedareasbyHoodedWarblers(Wilsonia citrina) inOntariousingclimatedatahadbetter informa‐tioncriterionsupportthanmodelsusingforestdata(MellesFortinLindsay amp Badzinski 2011) In contrast land‐use models outper‐formedclimatemodels for18 farmlandbird species in theUnitedKingdom(EglingtonampPearce‐Higgins2012)Similarlyvegetation‐based species distributionmodels had superior performance thanclimate‐based models for 79 Spanish bird species (Seoane et al2004)andforthreebirdspeciesinNewMexico(FriggensampFinch2015) Two studies focused on colonization and extinction ratesfoundmixed results ForGardenWarblers (Sylvia borin) inBritainvegetationexplainedmorevariationincolonizationwhiletempera‐turesexplainedmorevariationinlocalextinction(MustinAmarampRedpath2014)For122birdspeciesinOntariothebest‐supportedcovariates (land cover or climate change) varied among species(YalcinampLeroux2018)
Thedivergentresultsamongstudiesregardingtheimportanceofclimateandlandcovermaypartlybeduetothebiologyofindividualspecies (Yalcinamp Leroux 2018) For example farmland birdsmayTA
BLE
2emspR2 DevfordifferentmodelsofcolonizationandextinctionprobabilitiesforbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012
Mod
el
Cova
riate
s on
colo
niza
tion
and
extin
ctio
nEa
ster
n W
ood
Pew
eeRe
d‐ey
ed V
ireo
Inte
rcep
tAv
erag
e H
abita
tAv
erag
e Cl
imat
eH
abita
t Dev
iatio
nCl
imat
e D
evia
tion
Year
Eff
ect
Δ minus
2LL
R2 Dev
Δ minus
2LL
R2 Dev
2X
XX
X0
0010
00
0010
0
6X
XX
XX
minus2138
202
minus3230
57
4X
XX
Xminus2154
196
minus3234
56
5X
XX
Xminus2667
04
minus3422
01
3X
XX
minus2679
0minus3427
0
1X
minus7098
NA
minus45311
NA
Not
eNAnotapplicable
1996emsp |emsp emspensp CLEMENT ET aL
beparticularlyaffectedbychangesinagriculturalintensitybecauseoftheirextensiveuseoffarms(EglingtonampPearce‐Higgins2012)Alternativelyithasbeenarguedthatspatialscalemayplayaroleintherelativeimportanceofglobalchangefactorswithclimatemoreimportantatlargescales(PearsonampDawson2003)Wenotethattwostudiesthatfoundalargerroleforclimatebothreliedonlower‐resolution bird surveys (05deg atlas in Barbet‐Massin et al 2012
10km2atlasinMellesetal2011)IntheBBSdatathatweanalyzedsurveyswereconductedevery800mbutwecalculatedoccupancyacross each 394km transect In addition the cited studies useddifferentdata sources anddiverse analysismethods complicatinginterpretationofthedifferingresults
While we estimated that climate explained more model de‐viance than habitat these estimated relationships were not
F I G U R E 3 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionrates((t)minus (t)minus )forEasternWoodPeweeintheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
emspensp emsp | emsp1997CLEMENT ET aL
TA B L E 3 emspEstimatedparametersforcorrelated‐detectiondynamicoccupancyModel6relatinghabitatandclimatecovariatestooccupancydynamicsofbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012Parameterestimatesindicatechangeinlogoddsofparametersinresponsetoz‐transformedcovariatesParametersψϑϑprimeγε p1p2andπdefinedintextForkeyparameters(boldtext)expectedsignsincluded
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
Ψintercept 2367 0177 4087 0381
Ψhabitat 0556 0163 2323 0288
Ψheatstress minus0011 0198 minus0322 0275
Ψcoldstress 0598 0166 1054 0251
ϑintercept minus1821 0026 minus1385 0010
ϑhabitat minus0055 0026 0516 0013
ϑheatstress minus0094 0030 0017 0008
ϑcoldstress 0172 0040 0256 0013
ϑprimeintercept 2292 0045 0426 0020
ϑprimehabitat 0316 0036 1210 0018
ϑprimeheatstress minus0006 0038 0054 0013
ϑprimecoldstress minus0622 0064 0875 0019
γintercept minus0884 0353 0023 0162
γhabitatdeviation + minus0292 0478 minus0240 1768
γheatstressdeviation minus minus1136 0607 0064 0194
γcoldstressdeviation minus minus0498 0474 2398 2253
γaveragehabitat 0095 0091 0788 0111
γaverageheatstress minus0024 0067 0004 0062
γaveragecoldstress 0227 0071 0333 0119
εintercept minus3957 0386 minus4471 0141
εhabitatdeviation minus minus0397 0605 0121 1492
εheatstressdeviation + 0643 0455 minus0034 0182
εcoldstressdeviation + 0153 0644 2042 2072
εaveragehabitat 0238 0090 minus1664 0123
εaverageheatstress minus0071 0092 minus0080 0074
εaveragecoldstress minus0307 0141 minus0939 0104
p1intercept minus2172 0043 1351 0039
p1 1998 minus0118 0048 minus0128 0058
p1 1999 minus0112 0049 minus0043 0058
p1 2000 minus0166 0050 minus0017 0059
p1 2001 minus0156 0049 0031 0061
p1 2002 minus0179 0049 0076 0062
p1 2003 minus0216 0051 0160 0064
p12004 minus0242 0051 0066 0061
p1 2005 minus0124 0050 0149 0061
p12006 minus0182 0051 0124 0060
p12007 minus0149 0050 0183 0062
p1 2008 minus0202 0052 0191 0064
p1 2009 minus0221 0052 0134 0062
p1 2010 minus0195 0051 0062 0060
p1 2011 minus0122 0051 0202 0063
(Continues)
1998emsp |emsp emspensp CLEMENT ET aL
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
p1 2012 minus0227 0055 0193 0063
p1stopnumber 0033 0009 minus0055 0011
p1stopnumbersquared minus0069 0010 minus0247 0013
p2intercept minus0921 0018 minus0555 0037
p2 1998 minus0138 0033 minus0121 0050
p2 1999 minus0137 0032 minus0025 0052
p2 2000 minus0164 0032 minus0027 0051
p2 2001 minus0077 0032 minus0059 0053
p2 2002 minus0101 0032 minus0012 0052
p2 2003 minus0199 0033 0085 0053
p22004 minus0186 0032 0113 0059
p2 2005 minus0146 0032 0082 0052
p22006 minus0155 0032 0135 0054
p22007 minus0153 0032 0124 0055
p2 2008 minus0168 0032 0073 0057
p2 2009 minus0077 0033 0100 0056
p2 2010 minus0114 0033 0071 0062
p2 2011 minus0079 0032 0120 0055
p2 2012 minus0186 0033 0030 0063
p2stopnumber 0045 0007 0165 0010
p2stopnumbersquared minus0105 0008 minus0182 0011
π 0533 0064 minus0505 0065
TA B L E 3 emsp (Continued)
F I G U R E 4 emspProbabilityof(a)colonizationand(b)extinctionforRed‐eyedVireointheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1999CLEMENT ET aL
significantanddidnotexplainalargefractionofvariationThereareseveralpotentialexplanationsforwhywedidnotfindasub‐stantialresponsetochangesinhabitatandclimateFirstourdatawerelackingindynamiceventstoinformourmodelsThecovari‐ateschangedrelativelylittleoverthetimeperiodmakingitdiffi‐culttodetectaresponseThelackofchangewaspartlybecauseoftherelativelyshort(16‐year)timeperiodstudiedAlsoweusedamovingaverageforclimatedatawhichtendstoreducedifferences
overshorttimeperiods Inadditionthestudyregionhasexperi‐encedlesswarmingoverthepastcenturythanotherpartsoftheplanet (Walshetal2014)Thebirddistributionsalso lackeddy‐namismBecauseextinctionrateswerelowtherewerefewlocalextinctioneventstoinformthemodelAlthoughcolonizationrateswerehighthefactthatinitialoccupancywashighandextinctionrateswerelowmeanttherewerefewopportunitiesforcoloniza‐tionevents
F I G U R E 5 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionratesforRed‐eyedVireointheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
2000emsp |emsp emspensp CLEMENT ET aL
Second measurement error may have obscured responses toglobalchangeThePRISMclimatedataareestimatedbyinterpola‐tionratherthandirectmeasurementacrossthestudyarea(Dalyetal2008)AssuchtheremaybeartifactsoranomaliesthatimpededourestimationandincreaseduncertaintySimilarlyNLCDdataareremotelysensedandmay includesomemeasurementerrorwhichcan increase uncertainty in our estimates (Homer et al 2015)Additionallylandcoverwasonlymeasuredinthreeyearsrequiringus to imputevalues in13yearsAsa result all landcover changethatoccurredbetween2002and2006wasrecordedin2002whilechangesbetween2007and2011wererecorded in2007Thisap‐proximation isnot idealbutwasnecessarygiventhe5‐yearreso‐lutionofthedataThislowerresolutionofthelandcoverdatamayhavereducedthesensitivityofthemodel
Third wemay havemisspecified ourmodelWe tried to de‐velopbiologicallymeaningfulcovariatesbutwemightnotunder‐standthesystemaswellaswehopedItispossiblethatbirdvitalrates respond to climateon adifferent time scale (ie not a15‐yearmoving average) or to climateduringdifferent timesof theyearortodifferenttemperaturethresholdsortodifferentclimatecomponents (egprecipitation Illaacutenetal2014) It isalsopossi‐ble that thebirds respond to finerhabitat features than the landcovertypesweusedoratadifferentspatialscale(ienot400mPearsonampDawson2003)Alternatively theremaybe importantregionaldifferencesor interactionsamongcovariatesthatwedidnotcaptureOritispossiblethatchangesinclimateandhabitatintheirbreedingareassimplyhavelittleeffectonvitalratesforthesespecies relative to other factors such as competitionwith otherbirdspeciesorglobalchangeoccurringontheirwinteringgroundsinSouthAmerica(Dormann2007)Ofcoursethesemisspecifica‐tionissuesareinherenttoallobservationalstudiesandnotuniquetothisstudy
Itwould be possible for us to explore some of themisspec‐ification issues just mentioned For example we could rerunthe samemodelsusingnewcovariates representing a varietyoftemperature thresholds and habitat buffer sizes to search for astrongercorrelationExperiencesuggeststhatpersistencewouldeventuallyberewardedwithap‐valuelt005ora lowmodelse‐lection valueHowever one reason to avoid such exploration isthat it tends to lead toType Ierrorsoverfitmodels andbiasedparameterestimates(FiebergampJohnson2015)Furthermorewefind thecontrastbetweensignificant correlations for spatial co‐variatesandnonsignificantcorrelationsfortemporalcovariatesil‐luminatingIntypicalclimateenvelopestudieswewouldestimatethe relationshipbetweencovariatesand thecurrentdistributionofaspeciesmuchlikeourestimateofinitialoccupancy(Dormann2007)BecauseweobtainedsignificantparameterestimateswemightconcludethatwehaveidentifiedcovariatesthatdeterminetherangeofthisspeciesThenwemightusetheseresultstoproj‐ectthefuturerangeofthespeciesundervariousclimatescenariosbased on the (questionable) assumption that the covariatendashspe‐cies relationship will not change as climate and habitat change(Cuddington et al 2013 Elith et al 2010Gustafson 2013) In
this case the insignificant correlationwith the temporal covari‐atessuggeststhatouroccupancycovariatesmerelydescribetherangeofthesebirdswithoutdeterminingtherangeFurthermoreour results suggest that despite significant results in the staticportionof themodelwehaveyet to identify themost relevantcomponents of climate change shaping thedistributionof thesebirds although itmay be difficult to identify these componentsgiven the lackofobservedchangeduringa relatively short timeperiodFinallyourresultssuggestthatprojectionsbasedonthestaticportionofourmodelareunlikelytobereliabledespitetheirstatisticalsignificance
Ifourinterpretationiscorrectitsuggeststhatsomeprojectionsof futurerangeshiftsbasedonstaticmodelsrelyonunsupportedassumptionsandarenot reliableWehavedemonstratedadiffer‐entapproachthatdoesnotrequirethestrongassumptionsofstaticmodelingapproachesOurapproachwouldbemoreapttoidentifyfactorsthatguidespeciesrsquoresponsetoglobalchangeifboththespe‐ciesandthecovariatesexhibitedmoredynamismandifweanalyzedalongertimeperiodthatencompassedgreaterchange
ACKNOWLEDG MENTS
WethankKeithPardieckDaveZiolkowskiJrandthethousandsofhighlyskilledvolunteerswhohavecoordinatedandperformedtheNorthAmericanBreedingBirdSurvey for half a centuryWealsothanktheClimateResearchUnitattheUniversityofEastAngliaformakingvaluableclimatedataeasilyaccessibleMJCwasfinanciallysupported by the BBS and the USGS Northeast Climate ScienceCenterThecontentsofthisdocumentaretheresponsibilityoftheauthorsanddonotnecessarilyrepresenttheviewsoftheUSGSAnyuseoftradeproductorfirmnamesisfordescriptivepurposesonlyanddoesnotimplyendorsementbytheUSGovernment
DEDIC ATION
We dedicate this paper to the late Chandler S Robbins ldquoFatherrdquooftheNorthAmericanBreedingBirdSurvey (BBS)DatafromtheBBSformthebasisforthisspecificpaperandforhundredsofothersfocusedon topics ranging from local trends tocontinentalmacro‐ecologyChanwasauniquecombinationofbird‐loverandscientistwhosevisionandinquisitiveenthusiasmwereaninspirationtoallofusfortunateenoughtohaveknownhim
CONFLIC T OF INTERE S T
Nonedeclared
AUTHORSrsquo CONTRIBUTION
JACandJDNconceivedthestudyAJTandSWanalyzedhabitatandclimatedataMJCJEHandJDNperformedtheoccupancyanaly‐sisMJCledthewritingofthemanuscriptAllauthorsgavefinalap‐provalforpublication
emspensp emsp | emsp2001CLEMENT ET aL
DATA ACCE SSIBILIT Y
Raw data are available from the following sources Breeding BirdSurvey httpswwwpwrcusgsgovbbsrawdata PRISM climatedata httpwwwprismoregonstateedu NLCD land cover datahttpswwwmrlcgov Processed data will be archived with theDryadDigitalRepository
ORCID
Matthew J Clement httpsorcidorg0000‐0003‐4231‐7949
James D Nichols httpsorcidorg0000‐0002‐7631‐2890
Adam J Terando httpsorcidorg0000‐0002‐9280‐043X
James E Hines httpsorcidorg0000‐0001‐5478‐7230
Steven G Williams httpsorcidorg0000‐0003‐3760‐6818
R E FE R E N C E S
Anderson D R Burnham K P Gould W R amp Cherry S (2001)Concerns about finding effects that are actually spuriousWildlife Society Bulletin29311ndash316
Arauacutejo M B Cabeza M Thuiller W Hannah L amp WilliamsP H (2004) Would climate change drive species out of re‐serves An assessment of existing reserve‐selection meth‐ods Global Change Biology 10 1618ndash1626 httpsdoiorg101111j1365‐2486200400828x
Barbet‐MassinMampJetzW(2014)A40‐yearcontinent‐widemulti‐speciesassessmentofrelevantclimatepredictorsforspeciesdistri‐butionmodellingDiversity and Distributions201285ndash1295httpsdoiorg101111ddi12229
Barbet‐Massin M Thuiller W amp Jiguet F (2012) The fate ofEuropean breeding birds under climate land‐use and disper‐sal scenarios Global Change Biology 18 881ndash890 httpsdoiorg101111j1365‐2486201102552x
Bellard C Bertelsmeier C Leadley P Thuiller W ampCourchamp F (2012) Impacts of climate change on the fu‐ture of biodiversity Ecology Letters 15 365ndash377 httpsdoiorg101111j1461‐0248201101736x
Boumlhning‐Gaese K amp Bauer H G (1996) Changes in species abun‐dance distribution and diversity in a central European birdcommunity Conservation Biology 10 175ndash187 httpsdoiorg101046j1523‐1739199610010175x
BrommerJELehikoinenAampValkamaJ(2012)Thebreedingrangesof central European and Arctic bird speciesmove poleward PLoS ONE7e43648httpsdoiorg101371journalpone0043648
CalderWAampKingJR(1974)ThermalandcaloricrelationsofbirdsInDSFarnerampJRKing(Eds)Avian biology(Vol4pp259ndash413)NewYorkNYAcademicPress
CesaraccioCSpanoDDucePampSnyderRL(2001)Animprovedmodel for determining degree‐day values from daily temperaturedataInternational Journal of Biometeorology45161ndash169httpsdoiorg101007s004840100104
ChamberlinTC (1890)ThemethodofmultipleworkinghypothesesScience1592ndash96
ClementMJHinesJENicholsJDPardieckKLampZiolkowskiDJJr (2016)Estimating indicesofrangeshifts inbirdsusingdy‐namicmodelswhendetectionisimperfectGlobal Change Biology223273ndash3285httpsdoiorg101111gcb13283
Commission for Environmental Cooperation (1997) Ecological regionsofNorthAmericaTowardacommonperspectiveCommission for
EnvironmentalCooperationMontrealRetrievedfromftpftpepagovwedecoregionscec_naCEC_NAecopdf
Crowl TA Crist TO Parmenter R R BelovskyGamp LugoA E(2008)Thespreadofinvasivespeciesandinfectiousdiseaseasdriv‐ersofecosystemchangeFrontiers in Ecology and the Environment6238ndash246httpsdoiorg101890070151
Cuddington K FortinM J Gerber L RHastings A Liebhold AOConnorMampRayC(2013)Process‐basedmodelsarerequiredtomanageecologicalsystemsinachangingworldEcosphere41ndash12httpsdoiorg101890ES12‐001781
DalyCHalbleibMSmithJIGibsonWPDoggettMKTaylorGHhellipPasterisPP(2008)Physiographicallysensitivemappingofcli‐matologicaltemperatureandprecipitationacrosstheconterminousUnited States International Journal of Climatology 28 2031ndash2064httpsdoiorg101002joc1688
DawsonW (1958) Relation of oxygen consumption and evaporativewater losstotemperatureintheCardinalPhysiological Zoology3137ndash48httpsdoiorg101086physzool31130155377
DawsonWRampWhittowGC (2000)Regulationofbodytempera‐ture InG CWhittow (Ed)Sturkiersquos avian physiology (5th ed pp343ndash390)SanDiegoCAAcademicPress
DeCalestaDS(1994)Effectofwhite‐taileddeeronsongbirdswithinmanagedforestsinPennsylvaniaJournal of Wildlife Management58711ndash718httpsdoiorg1023073809685
DevictorVJulliardRCouvetDampJiguetF(2008)Birdsaretrack‐ing climatewarming but not fast enoughProceedings of the Royal Society of London B Biological Sciences2752743ndash2748httpsdoiorg101098rspb20080878
Dormann C F (2007) Promising the future Global change projec‐tionsofspeciesdistributionsBasic and Applied Ecology8387ndash397httpsdoiorg101016jbaae200611001
Dormann C F Schymanski S J Cabral J Chuine I Graham CHartigFhellipSingerA(2012)Correlationandprocessinspeciesdis‐tributionmodelsBridgingadichotomyJournal of Biogeography392119ndash2131httpsdoiorg101111j1365‐2699201102659x
EccelE(2010)WhatwecanasktohourlytemperaturerecordingPartIIHourlyinterpolationoftemperaturesforclimatologyandmodel‐lingItalian Journal of Agrometeorology1545ndash50
EccelEampCordanoE(2013)InterpolTHourlyinterpolationofmulti‐pletemperaturedailyseriesRpackageversion211RetrievedfromhttpsCRANR‐projectorgpackage=InterpolT
EglingtonSMampPearce‐HigginsJW(2012)Disentanglingtherela‐tiveimportanceofchangesinclimateandland‐useintensityindriv‐ingrecentbirdpopulationtrendsPLoS ONE7e30407httpsdoiorg101371journalpone0030407
Elith JKearneyMampPhillipsS (2010)Theartofmodelling range‐shifting species Methods in Ecology and Evolution 1 330ndash342httpsdoiorg101111j2041‐210X201000036x
FiebergJampJohnsonDH(2015)MMIMultimodelinferenceormod‐elswithmanagement implications Journal of Wildlife Management79708ndash718
FriggensMMampFinchDM (2015) Implicationsof climatechangeforbirdconservationinthesouthwesternUSunderthreealternativefuturesPLoS ONE10e0144089
FryJXianGZJinSDewitzJHomerCGYangLhellipWickhamJD (2011)Completionof the2006national landcoverdatabasefor theconterminousUnitedStatesPhotogrammetric Engineering amp Remote Sensing77858ndash864
Fuller R J Gregory R D Gibbons D W Marchant J HWilson J D Baillie S R amp Carter N (1995) Population de‐clines and range contractions among lowland farmland birdsin Britain Conservation Biology 9 1425ndash1441 httpsdoiorg101046j1523‐1739199509061425x
Gaston K J Blackburn T M Greenwood J J Gregory R DQuinn R M amp Lawton J H (2000) Abundancendashoccupancy
2002emsp |emsp emspensp CLEMENT ET aL
relationships Journal of Applied Ecology 37 39ndash59 httpsdoiorg101046j1365‐2664200000485x
GoldsteinRB(1974)RelationofmetabolismtoambienttemperatureintheverdinCondor76116ndash119httpsdoiorg1023071365995
GrinnellJ(1917)FieldtestsoftheoriesconcerningdistributionalcontrolAmerican Naturalist51115ndash128httpsdoiorg101086279591
GrosboisVGimenezOGaillardJMPradelRBarbraudCClobertJhellipWeimerskirchH(2008)Assessingtheimpactofclimatevari‐ation on survival in vertebrate populationsBiological Reviews 83357ndash399httpsdoiorg101111j1469‐185X200800047x
GustafsonEJ(2013)WhenrelationshipsestimatedinthepastcannotbeusedtopredictthefutureUsingmechanisticmodelstopredictlandscapeecologicaldynamicsinachangingworldLandscape Ecology281429ndash1437httpsdoiorg101007s10980‐013‐9927‐4
Hamel P B (1992) LandManagers guide to the birds of the SouthGeneralTechnicalReportSE‐22USDAForestServiceSoutheasternForestExperimentStationandTheNatureConservancyChapelHillNC
Hinds D S amp Calder W A (1973) Temperature regulation of thePyrrhuloxiaandtheArizonaCardinalPhysiological Zoology4655ndash71httpsdoiorg101086physzool46130152517
HinesJE(2006)PRESENCE Software to estimate patch occupancy and related parameters Laurel MD US Geological survey PatuxentWildlife Research Center Retrieved from httpwwwmbr‐pwrcusgsgovsoftwarepresencehtml
HinesJENicholsJDampCollazoJA(2014)Multiseasonoccupancymodels for correlated replicate surveys Methods in Ecology and Evolution5583ndash591httpsdoiorg1011112041‐210X12186
HinesJENicholsJDRoyleJAMacKenzieDIGopalaswamyAMKumarNampKaranthKU(2010)TigersontrailsOccupancymodeling for cluster sampling Ecological Applications 20 1456ndash1466httpsdoiorg10189009‐03211
HockeyPA SiramiCRidleyARMidgleyGFampBabikerHA(2011) Interrogating recent range changes in South African birdsConfounding signals from land use and climate change present achallenge for attribution Diversity and Distributions 17 254ndash261httpsdoiorg101111j1472‐4642201000741x
Hoegh‐Guldberg O Hughes L McIntyre S Lindenmayer D BParmesanCPossinghamHPampThomasCD (2008)AssistedcolonizationandrapidclimatechangeScience321345ndash346
Homer C Dewitz J Fry J Coan M Hossain N Larson C hellipWickham J (2007) Completion of the 2001 national land coverdatabase for the conterminous United States Photogrammetric Engineering amp Remote Sensing73337
HomerCDewitzJYangLJinSDanielsonPXianGhellipMegownK(2015)Completionofthe2011NationalLandCoverDatabasefortheconterminousUnitedStatesndashrepresentingadecadeoflandcoverchange informationPhotogrammetric Engineering amp Remote Sensing81345ndash354
HootenMBWikleCKDorazioRMampRoyleJA(2007)HierarchicalspatiotemporalmatrixmodelsforcharacterizinginvasionsBiometrics63558ndash567httpsdoiorg101111j1541‐0420200600725x
HosmerDWJrampLemeshowS(2000)Applied logistic regression(2nded)NewYorkNYJohnWileyampSons
Huntley B Collingham Y CGreen R EHiltonGM RahbekCamp Willis S G (2006) Potential impacts of climatic change upongeographical distributions of birds Ibis 148 8ndash28 httpsdoiorg101111j1474‐919X200600523x
IgleciaMNCollazoJAampMcKerrowAJ(2012)Useofoccupancymodels to evaluate expert knowledge‐based species‐habitat rela‐tionshipsAvian Conservation and Ecology7(2)13pRetrievedfromhttpwwwace‐ecoorgvol7iss2art5
IllaacutenJGThomasCDJonesJAWongWKShirleySMampBettsMG(2014)Precipitationandwintertemperaturepredictlong‐termrange‐scale abundancechanges inWesternNorthAmericanbirds
Global Change Biology 20 3351ndash3364 httpsdoiorg101111gcb12642
IoannidisJP (2005)WhymostpublishedresearchfindingsarefalsePLoS Medicine2e124
KeacuteryMGuillera‐ArroitaGampLahoz‐Monfort J J (2013)Analysingand mapping species range dynamics using occupancy modelsJournal of Biogeography 40 1463ndash1474 httpsdoiorg101111jbi12087
Khaliq I Hof C Prinzinger R Boumlhning‐Gaese K amp PfenningerM(2014) Global variation in thermal tolerances and vulnerability ofendotherms to climate change Proceedings of the Royal Society B Biological Sciences 281(1789) 20141097 httpsdoiorg101098rspb20141097
Laursen K amp Frikke J (2008) Hunting from motorboats displacesWadden Sea eiders Somateria mollissima from their favouredfeeding distribution Wildlife Biology 14 423ndash433 httpsdoiorg1029810909‐6396‐144423
LemoineNBauerHGPeintingerMampBoumlhning‐GaeseK (2007)Effectsof climate and land‐use changeon species abundance in acentralEuropeanbirdcommunityConservation Biology21495ndash503httpsdoiorg101111j1523‐1739200600633x
MacArthurRH(1972)Geographical ecology Patterns in the distribution of speciesNewYorkNYHarperampRow
MacKenzieDINicholsJDHinesJEKnutsonMGampFranklinAB(2003)Estimatingsiteoccupancycolonizationandlocalextinc‐tionwhenaspeciesisdetectedimperfectlyEcology842200ndash2207httpsdoiorg10189002‐3090
MacKenzieDINicholsJDLachmanGBDroegeSRoyleJAampLangtimmCA(2002)Estimatingsiteoccupancyrateswhendetec‐tionprobabilitiesarelessthanoneEcology832248ndash2255httpsdoiorg1018900012‐9658(2002)083[2248ESORWD]20CO2
MacKenzieDINicholsJDRoyleJAPollockKHBaileyLampHinesJE(2018)Occupancy estimation and modeling Inferring pat‐terns and dynamics of species occurrence (2nd ed) BurlingtonMAAcademicPress
MellesSJFortinMJLindsayKampBadzinskiD(2011)ExpandingnorthwardInfluenceofclimatechangeforestconnectivityandpop‐ulationprocessesonathreatenedspeciesrangeshiftGlobal Change Biology1717ndash31httpsdoiorg101111j1365‐2486201002214x
Morin X amp Thuiller W (2009) Comparing niche‐and process‐based models to reduce prediction uncertainty in species rangeshifts under climate change Ecology 90 1301ndash1313 httpsdoiorg10189008‐01341
MustinKAmarAampRedpathSM(2014)Colonizationandextinctiondynamicsofadecliningmigratorybirdareinfluencedbyclimateandhabitat degradation Ibis 156 788ndash798 httpsdoiorg101111ibi12173
NicholsJDThomasLampConnPB(2009)InferencesaboutlandbirdabundancefromcountdataRecentadvancesandfuturedirectionsInDLThomsonEJCoochampMJConroy(Eds)Modeling demo‐graphic processes in marked populations(pp201ndash235)NewYorkNYSpringer
OmernikJM(1995)EcoregionsAspatialframeworkforenvironmen‐talmanagement InWSDavisampTSimon(Eds)Biological assess‐ment and criteria Tools for water resource planning and decision making (pp49ndash62)BocaRatonFLLewisPublishers
OmernikJM(2004)Perspectivesonthenatureanddefinitionofeco‐logicalregionsEnvironmental Management34(Supplement1)S27ndashS38httpsdoiorg101007s00267‐003‐5197‐2
Omernik J M amp Griffith G E (2014) Ecoregions of the contermi‐nousUnited States Evolution of a hierarchical spatial frameworkEnvironmental Management541249ndash1266httpsdoiorg101007s00267‐014‐0364‐1
Pardieck K L ZiolkowskiD J JrampHudsonM‐A R (2015)NorthAmericanBreedingBirdSurveyDataset1966ndash2014version20140
emspensp emsp | emsp2003CLEMENT ET aL
USGeologicalSurveyPatuxentWildlifeResearchCenterRetrievedfromhttpswwwpwrcusgsgovbbsrawdata
ParmesanCampYoheG (2003)Agloballycoherentfingerprintofcli‐mate change impacts across natural systemsNature 421 37ndash42httpsdoiorg101038nature01286
PearsonRGampDawsonTP(2003)PredictingtheimpactsofclimatechangeonthedistributionofspeciesArebioclimateenvelopemod‐elsusefulGlobal Ecology and Biogeography12361ndash371
PlattJR(1964)StronginferenceScience146347ndash353PopperK(1959)The logic of scientific discoveryLondonUKHutchinsonRobbins C S Sauer J R Greenberg R S amp Droege S (1989)
Population declines in North American birds that migrate to theNeotropics Proceedings of the National Academy of Sciences 867658ndash7662httpsdoiorg101073pnas86197658
Robinson R A Wilson J D amp Crick H Q (2001) The impor‐tance of arable habitat for farmland birds in grassland land‐scapes Journal of Applied Ecology 38 1059ndash1069 httpsdoiorg101046j1365‐2664200100654x
Romesburg H C (1981) Wildlife science Gaining reliable knowl‐edge Journal of Wildlife Management 45 293ndash313 httpsdoiorg1023073807913
Root T (1988) Energy constraints on avian distributions and abun‐dancesEcology69330ndash339httpsdoiorg1023071940431
Ruiz‐GutieacuterrezVampZipkinEF (2011)Detectionbiasesyieldmis‐leadingpatternsof speciespersistence and colonization in frag‐mented landscapesEcosphere2 1ndash14 httpsdoiorg101890ES10‐002071
SauerJRHinesJEFallonJEPardieckKLZiolkowskiDJJrampLinkWA(2015)The North American breeding bird survey results and analysis 1966ndash2013 Version 01302015LaurelMDUSGSPatuxentWildlifeResearchCenterhttpswwwmbr‐pwrcusgsgovbbsbbshtml
Sauer J R LinkWA Fallon J E Pardieck K L amp ZiolkowskiDJJr(2013)TheNorthAmericanbreedingbirdsurvey1966ndash2011SummaryanalysisandspeciesaccountsNorth American Fauna791ndash32httpsdoiorg103996nafa790001
SeoaneJBustamanteJampDiacuteaz‐DelgadoR(2004)Competingrolesforlandscapevegetationtopographyandclimateinpredictivemod‐elsofbirddistributionEcological Modelling171209ndash222httpsdoiorg101016jecolmodel200308006
Sirami C Caplat P Popy S Clamens A Arlettaz R Jiguet F hellipMartin J‐L (2017) Impacts of global change on species distri‐butions Obstacles and solutions to integrate climate and landuse Global Ecology and Biogeography 26 385ndash394 httpsdoiorg101111geb12555
SkalskiJR(1996)Regressionofabundanceestimatesfrommarkrecap‐ture surveys against environmental covariatesCanadian Journal of
Fisheries and Aquatic Sciences53196ndash204httpsdoiorg101139f95‐169
SohlTL(2014)Therelativeimpactsofclimateandland‐usechangeonconterminousUnitedStatesbird species from2001 to2075PLoS ONE9e112251httpsdoiorg101371journalpone0112251
ThogmartinWESauerJRampKnutsonMG(2007)ModelingandmappingabundanceofAmericanwoodcockacrosstheMidwesternandNortheasternUnitedStatesJournal of Wildlife Management71376ndash382httpsdoiorg1021932005‐680
ThomasCDampLennonJJ (1999)Birdsextendtheirrangesnorth‐wardsNature399213httpsdoiorg10103820335
Trathan P N Garciacutea‐Borboroglu P Boersma D Bost C ACrawford R J CrossinG ThellipWienecke B (2015) Pollutionhabitatlossfishingandclimatechangeascriticalthreatstopen‐guins Conservation Biology 29 31ndash41 httpsdoiorg101111cobi12349
TravisJMJ(2003)Climatechangeandhabitatdestructionadeadlyanthropogenic cocktail Proceedings of the Royal Society of London Series B Biological Sciences 270(1514) 467ndash473 httpsdoiorg101098rspb20022246
UrbanMC (2015)Acceleratingextinction risk fromclimatechangeScience348571ndash573httpsdoiorg101126scienceaaa4984
WalshJWuebblesDHayhoeKKossinJKunkelKStephensGhellipSomervilleR(2014)OurchangingclimateInJMMelilloTCRichmondampGWYohe(Eds)Climate change impacts in the United States The Third National climate assessment(pp19ndash67)WashingtonDCUSGovernmentPrintingOffice
YackulicCBNichols JDReid JampDerR (2015)Topredict thenichemodelcolonizationandextinctionEcology9616ndash23httpsdoiorg10189014‐13611
Yalcin S amp Leroux S J (2018) An empirical test of the relative andcombinedeffectsofland‐coverandclimatechangeonlocalcoloni‐zationandextinctionGlobal Change Biology243849ndash3861httpsdoiorg101111gcb14169
ZhuKWoodallCWampClarkJS(2012)FailuretomigrateLackoftreerangeexpansioninresponsetoclimatechangeGlobal Change Biology181042ndash1052httpsdoiorg101111j1365‐2486201102571x
How to cite this articleClementMJNicholsJDCollazoJATerandoAJHinesJEWilliamsSGPartitioningglobalchangeAssessingtherelativeimportanceofchangesinclimateandlandcoverforchangesinaviandistributionEcol Evol 201991985ndash2003 httpsdoiorg101002ece34890

1992emsp |emsp emspensp CLEMENT ET aL
Parameters
Eastern Wood Pewee Red‐eyed Vireo
Estimate SE Estimate SE
ε 2010 minus0397 0605 minus0348 0304
ε 2011 0643 0455 minus0999 0406
ε 2012 0153 0644 minus0803 0410
εaveragehabitat 0238 0090 minus1670 0122
εaverageheatstress minus0071 0092 minus0075 0074
εaveragecoldstress minus0307 0141 minus0904 0104
p1intercept minus2172 0043 1352 0038
p1 1998 minus0118 0048 minus0127 0058
p1 1999 minus0112 0049 minus0043 0058
p1 2000 minus0166 0050 minus0016 0059
p1 2001 minus0156 0049 0031 0060
p1 2002 minus0179 0049 0076 0061
p1 2003 minus0216 0051 0161 0064
p12004 minus0242 0051 0066 0060
p1 2005 minus0124 0050 0149 0060
p12006 minus0182 0051 0124 0059
p12007 minus0149 0050 0184 0061
p1 2008 minus0202 0052 0192 0063
p1 2009 minus0221 0052 0135 0061
p1 2010 minus0195 0051 0062 0059
p1 2011 minus0122 0051 0203 0062
p1 2012 minus0227 0055 0194 0062
p1stopnumber 0033 0009 minus0055 0011
p1stopnumbersquared minus0069 0010 minus0247 0013
p2intercept minus0921 0018 minus0557 0036
p2 1998 minus0138 0033 minus0106 0050
p2 1999 minus0137 0032 0000 0052
p2 2000 minus0164 0032 minus0027 0051
p2 2001 minus0077 0032 minus0053 0053
p2 2002 minus0101 0032 minus0019 0052
p2 2003 minus0199 0033 0086 0053
p22004 minus0186 0032 0124 0057
p2 2005 minus0146 0032 0076 0051
p22006 minus0155 0032 0128 0054
p22007 minus0153 0032 0120 0054
p2 2008 minus0168 0032 0072 0056
p2 2009 minus0077 0033 0101 0055
p2 2010 minus0114 0033 0085 0060
p2 2011 minus0079 0032 0124 0054
p2 2012 minus0186 0033 0036 0061
p2stopnumber 0045 0007 0165 0010
p2stopnumbersquared minus0105 0008 minus0183 0010
π 0533 0064 minus0505 0065
TA B L E 1 emsp (Continued)
emspensp emsp | emsp1993CLEMENT ET aL
toModel3althoughnotsignificantlyduetothenumberofaddi‐tional parameters (χ2=2679df =28p=053)UnderModel 2estimatedcolonizationratesvariedfrom000in2008to030in1998whileextinctionratesvariedfrom001 in2001ndash2003and2010to004in2011WeusedModels45and6toestimatethepercentof improvedmodel fit (measuredby thechange indevi‐ance)generatedbyModel2thatcouldbeaccountedforbytempo‐ralvariationinclimateandhabitatcovariates(Table2)Temporalvariationinhabitataccountedfor04ofthechangeindeviancewhile temporal variation in climate accounted for 196Model6whichaccountedfortemporalvariationinbothhabitatandcli‐mateindicatedthatin1998colonizationinthesouthwestportionofthestudyareawasabovethelocalaveragewhileextinctioninthesouthwestwasbelowthelocalaverageindicatingarelativelystrongoccupancy trend in thesouthwest in thatyear (Figure3)By2012thispatternhadreversedyieldingarelativelyweakoc‐cupancytrendinthesouthwest(Figure3)NoneofthesemodelsrepresentedasignificantimprovementoverModel3(pgt026)Insomecasesthesignoftheestimatedeffectoftemporalvariationinclimateandhabitatwasoppositeoftheexpectedeffectbutinnoinstancesweretheseestimatessignificant(Table3)
32emsp|emspRed‐eyed Vireo
The best‐supportedmodel for Red‐eyedVireo included the samecovariatesasforEasternWoodPeweeyieldingaglobalmodelwith85parameters (Table1)TheHosmerndashLemeshowtest indicatedfitwasadequate for theRed‐eyedVireoglobalmodel forbothnaiumlvecolonization rates (χ2=540df =4p=025) and naiumlve extinctionrates(χ2=100df =4p=091)
Red‐eyedVireowere similarlywidespread in the study areawith an average probability of occupancy of 091 in the global
model (Figure1)Occupancydecreasedslightlyto090by2012incontrasttothe increase incountsreportedbytheBBS Initialoccupancywas positively related to available habitat and hoursofcoldstress (Table1)Probabilityofdetectionat the first stopwasrelativelyhighat038whileprobabilityofdetectionattheroute levelnearlyperfectat098Model3which includedspa‐tialvariationinclimateandhabitattothecolonizationandextinc‐tionmodelsimprovedmodelfitdramatically(χ2=41884df =6plt0001)relativetothenullmodelInkeepingwiththegreaterchangeindevianceforRed‐eyedVireoModel3estimatedmorespatialvariationinvitalratescomparedtoEasternWoodPeweeIn 1998 estimated colonization rates varied across BBS routesfrom 010 to 088 (mean of 049)while extinction rates rangedfrom000to048(meanof004Figure4)Model2whichallowedcolonization and extinction rates to vary each year improvedmodelfitrelativetoModel3butnotsignificantlyduetothenum‐berofadditionalparameters(χ2=3427df =28p=019)UnderModel2estimatedcolonizationratesvariedfrom039in2008to060 in2000while estimatedextinction rates varied from003in2006to007in1998WeusedModels45and6toestimatethepercentofthedeviancereductioninModel2thatcouldbeac‐countedforbytemporalvariationinclimateandhabitatcovariates(Table2)Wefoundthattemporalvariation inhabitataccountedfor01of the change indeviancewhile climate accounted for56Model 6 which accounted for temporal variation in bothhabitatandclimateindicatedthatin1998bothcolonizationandextinctionwereaboveaverageindicatinghigherturnoverearlyinthestudyperiod(Figure5)By2012bothcolonizationandextinc‐tion rateshaddeclined indicatinggreater stability inoccupancy(Figure 5) None of these models represented a significant im‐provementoverModel3(pgt074)Someoftheestimatedeffects
F I G U R E 1 emspOccupancyprobabilityfor(a)EasternWoodPeweeand(b)Red‐eyedVireointheeasternUnitedStates1997underModel2Occupancyvariesbytheamountofsuitablehabitathoursofheatstressandhoursofcoldstressin1997
1994emsp |emsp emspensp CLEMENT ET aL
oftemporalvariationinclimateandhabitatdifferedfromexpecta‐tionsbutnotsignificantly(Table3)
4emsp |emspDISCUSSION
Wefound that forbothbirdspecies initialoccupancycoloniza‐tionratesandextinctionratescorrelatedwithspatialvariationinclimateandhabitatcovariatesHowevercolonizationandextinc‐tionrateswerenotcorrelatedwithtemporalvariation inclimateand habitat covariates Our results correlating initial occupancywith environmental covariates are broadly speaking consistentwithbiologicalexpectationandaraftofpreviousstudiesthathaveindicatedasignificantrelationshipbetweenspeciesdistributionsandhabitat(RobinsonWilsonampCrick2001ThogmartinSauerampKnutson 2007) climate (Barbet‐Massinamp Jetz 2014BellardBertelsmeier Leadley Thuiller amp Courchamp 2012) or both(Seoane Bustamante amp Diacuteaz‐Delgado 2004 Sohl 2014)Howeverwesuggestthatestimatedrelationshipsbetweenspatialvariationinenvironmentalcovariatesandcolonizationandextinc‐tion rates are of greater ecological interest than relatively phe‐nomenological species distribution models or climate envelopemodels(Clementetal2016)Theestimatedvitalratesofcoloni‐zationandextinctionbringusclosertounderstandingthedynamicprocessunderlying thedistributionof these speciesGivencon‐stantvital rateswecanalsoproject theequilibrium levelofoc‐cupancyatsites ( i
i+i
atsite i)Wecancomparetheseprojected
equilibria tocurrentoccupancytoproject trends for thespeciesunder current environmental conditions We can also use
projectedchangesinkeyenvironmentalvariablestodeveloppre‐dicted rangechanges in the futureWeargue that theseprojec‐tions are more robust than those from static climate envelopemodels which assume that species are currently in equilibrium(Yackulicetal2015)Incontraststaticmodelscanoverestimaterange changes when the equilibrium does not hold (Morin ampThuiller2009)
Given the importanceofglobal changeweweremotivated toinvestigate factors affecting temporal aswell as spatial variationin vital rates In this case spatial variation must be incorporatedintosuchanalysesinordertoaccommodatethedifferentdynamicsexpectedindifferentportionsofaspeciesrangeWeviewthere‐lationshipbetweentemporalvariationincovariatevaluesandvitalrates as the most relevant to understanding the effect of globalchangeontherecentandfuturedistributionsofourstudyspeciesFurthermore a correlation between vital rates and spatially vary‐ingcovariatesmaynot indicateasimilarcorrelationwithtemporalchangesinthosesamecovariatesForexampleaspecieswithstrongdispersal constraintsmight have a strong spatial relationshipwithacovariatebutaweaktemporalrelationshipduetoits immobility(Devictor et al 2008) Similarly an estimated spatial relationshipmaybepurelyphenomenologicalduetothenatureofspatialvari‐ablesWhen spatial variables followgradients it is relatively easytoobtainstatisticallysignificantcorrelationsabsentanycausalre‐lationship(Grosboisetal2008)ForexampleifcolonizationratesandtemperaturesbothfollownorthndashsouthgradientstheymaybespatiallycorrelatedevenifcolonizationisgovernedbysomeotherunidentifiedfactorInthiscasewewouldexpecttheweakcorrela‐tion between temporal changes in temperature and colonization
F I G U R E 2 emspProbabilityof(a)colonizationand(b)extinctionforEasternWoodPeweeintheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1995CLEMENT ET aL
ratestobemoreindicativeofthetruerelationshipthanthestrongcorrelationwithspatialvariationintemperature
BBS surveys indicate different abundance trajectories for ourstudyspecieswithcounts increasingforRed‐eyedVireosandde‐creasing for EasternWood Pewees (Sauer et al 2015) Becauseabundanceandoccupancytendtobelinked(Gastonetal2000)weexpectedsomedivergenceinoccupancydynamicsWefoundthatestimatedoccupancywashighandstableforbothspeciesalthoughtherewasgreaterturnoverforRed‐eyedVireosForexampleunderModel3 (spatialvariationonly incovariates)bothaveragecoloni‐zationrates(049)andaverageextinctionrates(004)wereatleasttwiceashighforRed‐eyedVireoscomparedtoEasternWoodPewee(018and002respectively)Red‐eyedVireosexhibitedgreaterspa‐tialvariationincolonizationandextinctionrates(Figure4)althoughour selected covariates explained little temporal variation for thisspeciesEasternWoodPeweecolonizationandextinctionratesex‐hibitedmoreofalatitudinalgradientreflectingaspatialcorrelationwith extreme temperature (Figure 2)Our results also hinted at agreatercorrelationbetweentemporalchanges in temperatureandoccupancyvitalratesforEasternWoodPeweewhichcouldplayaroleinthedeclineofthisspeciesalthoughtheresultswerenotsig‐nificant(Table3)Alternatively ithasbeensuggestedthatEasternWoodPeweemaybedecliningduetoa lossofforestedwinteringhabitat(RobbinsSauerGreenbergampDroege1989)orduetodeer‐mediatedchanges in foreststructureonbreedinggrounds (deCal‐esta 1994) illustrating the complexity of ascertaining cause andeffectusingobservationalstudiesinthesenaturalsystems
Althoughrelativelyrare(Siramietal2017)otherstudieshaveinvestigated both climate and land cover effects on bird distribu‐tions Inagreementwithour finding that temporalvariation incli‐mate explainedmuchmore of themodel deviance than temporalvariation in habitat climate‐only species distribution models hadbetter cross‐validation support than habitat‐only models for 409Europeanbirdspecies(Barbet‐Massinetal2012)SimilarlymodelsestimatingthenumberofnewlyoccupiedareasbyHoodedWarblers(Wilsonia citrina) inOntariousingclimatedatahadbetter informa‐tioncriterionsupportthanmodelsusingforestdata(MellesFortinLindsay amp Badzinski 2011) In contrast land‐use models outper‐formedclimatemodels for18 farmlandbird species in theUnitedKingdom(EglingtonampPearce‐Higgins2012)Similarlyvegetation‐based species distributionmodels had superior performance thanclimate‐based models for 79 Spanish bird species (Seoane et al2004)andforthreebirdspeciesinNewMexico(FriggensampFinch2015) Two studies focused on colonization and extinction ratesfoundmixed results ForGardenWarblers (Sylvia borin) inBritainvegetationexplainedmorevariationincolonizationwhiletempera‐turesexplainedmorevariationinlocalextinction(MustinAmarampRedpath2014)For122birdspeciesinOntariothebest‐supportedcovariates (land cover or climate change) varied among species(YalcinampLeroux2018)
Thedivergentresultsamongstudiesregardingtheimportanceofclimateandlandcovermaypartlybeduetothebiologyofindividualspecies (Yalcinamp Leroux 2018) For example farmland birdsmayTA
BLE
2emspR2 DevfordifferentmodelsofcolonizationandextinctionprobabilitiesforbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012
Mod
el
Cova
riate
s on
colo
niza
tion
and
extin
ctio
nEa
ster
n W
ood
Pew
eeRe
d‐ey
ed V
ireo
Inte
rcep
tAv
erag
e H
abita
tAv
erag
e Cl
imat
eH
abita
t Dev
iatio
nCl
imat
e D
evia
tion
Year
Eff
ect
Δ minus
2LL
R2 Dev
Δ minus
2LL
R2 Dev
2X
XX
X0
0010
00
0010
0
6X
XX
XX
minus2138
202
minus3230
57
4X
XX
Xminus2154
196
minus3234
56
5X
XX
Xminus2667
04
minus3422
01
3X
XX
minus2679
0minus3427
0
1X
minus7098
NA
minus45311
NA
Not
eNAnotapplicable
1996emsp |emsp emspensp CLEMENT ET aL
beparticularlyaffectedbychangesinagriculturalintensitybecauseoftheirextensiveuseoffarms(EglingtonampPearce‐Higgins2012)Alternativelyithasbeenarguedthatspatialscalemayplayaroleintherelativeimportanceofglobalchangefactorswithclimatemoreimportantatlargescales(PearsonampDawson2003)Wenotethattwostudiesthatfoundalargerroleforclimatebothreliedonlower‐resolution bird surveys (05deg atlas in Barbet‐Massin et al 2012
10km2atlasinMellesetal2011)IntheBBSdatathatweanalyzedsurveyswereconductedevery800mbutwecalculatedoccupancyacross each 394km transect In addition the cited studies useddifferentdata sources anddiverse analysismethods complicatinginterpretationofthedifferingresults
While we estimated that climate explained more model de‐viance than habitat these estimated relationships were not
F I G U R E 3 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionrates((t)minus (t)minus )forEasternWoodPeweeintheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
emspensp emsp | emsp1997CLEMENT ET aL
TA B L E 3 emspEstimatedparametersforcorrelated‐detectiondynamicoccupancyModel6relatinghabitatandclimatecovariatestooccupancydynamicsofbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012Parameterestimatesindicatechangeinlogoddsofparametersinresponsetoz‐transformedcovariatesParametersψϑϑprimeγε p1p2andπdefinedintextForkeyparameters(boldtext)expectedsignsincluded
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
Ψintercept 2367 0177 4087 0381
Ψhabitat 0556 0163 2323 0288
Ψheatstress minus0011 0198 minus0322 0275
Ψcoldstress 0598 0166 1054 0251
ϑintercept minus1821 0026 minus1385 0010
ϑhabitat minus0055 0026 0516 0013
ϑheatstress minus0094 0030 0017 0008
ϑcoldstress 0172 0040 0256 0013
ϑprimeintercept 2292 0045 0426 0020
ϑprimehabitat 0316 0036 1210 0018
ϑprimeheatstress minus0006 0038 0054 0013
ϑprimecoldstress minus0622 0064 0875 0019
γintercept minus0884 0353 0023 0162
γhabitatdeviation + minus0292 0478 minus0240 1768
γheatstressdeviation minus minus1136 0607 0064 0194
γcoldstressdeviation minus minus0498 0474 2398 2253
γaveragehabitat 0095 0091 0788 0111
γaverageheatstress minus0024 0067 0004 0062
γaveragecoldstress 0227 0071 0333 0119
εintercept minus3957 0386 minus4471 0141
εhabitatdeviation minus minus0397 0605 0121 1492
εheatstressdeviation + 0643 0455 minus0034 0182
εcoldstressdeviation + 0153 0644 2042 2072
εaveragehabitat 0238 0090 minus1664 0123
εaverageheatstress minus0071 0092 minus0080 0074
εaveragecoldstress minus0307 0141 minus0939 0104
p1intercept minus2172 0043 1351 0039
p1 1998 minus0118 0048 minus0128 0058
p1 1999 minus0112 0049 minus0043 0058
p1 2000 minus0166 0050 minus0017 0059
p1 2001 minus0156 0049 0031 0061
p1 2002 minus0179 0049 0076 0062
p1 2003 minus0216 0051 0160 0064
p12004 minus0242 0051 0066 0061
p1 2005 minus0124 0050 0149 0061
p12006 minus0182 0051 0124 0060
p12007 minus0149 0050 0183 0062
p1 2008 minus0202 0052 0191 0064
p1 2009 minus0221 0052 0134 0062
p1 2010 minus0195 0051 0062 0060
p1 2011 minus0122 0051 0202 0063
(Continues)
1998emsp |emsp emspensp CLEMENT ET aL
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
p1 2012 minus0227 0055 0193 0063
p1stopnumber 0033 0009 minus0055 0011
p1stopnumbersquared minus0069 0010 minus0247 0013
p2intercept minus0921 0018 minus0555 0037
p2 1998 minus0138 0033 minus0121 0050
p2 1999 minus0137 0032 minus0025 0052
p2 2000 minus0164 0032 minus0027 0051
p2 2001 minus0077 0032 minus0059 0053
p2 2002 minus0101 0032 minus0012 0052
p2 2003 minus0199 0033 0085 0053
p22004 minus0186 0032 0113 0059
p2 2005 minus0146 0032 0082 0052
p22006 minus0155 0032 0135 0054
p22007 minus0153 0032 0124 0055
p2 2008 minus0168 0032 0073 0057
p2 2009 minus0077 0033 0100 0056
p2 2010 minus0114 0033 0071 0062
p2 2011 minus0079 0032 0120 0055
p2 2012 minus0186 0033 0030 0063
p2stopnumber 0045 0007 0165 0010
p2stopnumbersquared minus0105 0008 minus0182 0011
π 0533 0064 minus0505 0065
TA B L E 3 emsp (Continued)
F I G U R E 4 emspProbabilityof(a)colonizationand(b)extinctionforRed‐eyedVireointheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1999CLEMENT ET aL
significantanddidnotexplainalargefractionofvariationThereareseveralpotentialexplanationsforwhywedidnotfindasub‐stantialresponsetochangesinhabitatandclimateFirstourdatawerelackingindynamiceventstoinformourmodelsThecovari‐ateschangedrelativelylittleoverthetimeperiodmakingitdiffi‐culttodetectaresponseThelackofchangewaspartlybecauseoftherelativelyshort(16‐year)timeperiodstudiedAlsoweusedamovingaverageforclimatedatawhichtendstoreducedifferences
overshorttimeperiods Inadditionthestudyregionhasexperi‐encedlesswarmingoverthepastcenturythanotherpartsoftheplanet (Walshetal2014)Thebirddistributionsalso lackeddy‐namismBecauseextinctionrateswerelowtherewerefewlocalextinctioneventstoinformthemodelAlthoughcolonizationrateswerehighthefactthatinitialoccupancywashighandextinctionrateswerelowmeanttherewerefewopportunitiesforcoloniza‐tionevents
F I G U R E 5 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionratesforRed‐eyedVireointheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
2000emsp |emsp emspensp CLEMENT ET aL
Second measurement error may have obscured responses toglobalchangeThePRISMclimatedataareestimatedbyinterpola‐tionratherthandirectmeasurementacrossthestudyarea(Dalyetal2008)AssuchtheremaybeartifactsoranomaliesthatimpededourestimationandincreaseduncertaintySimilarlyNLCDdataareremotelysensedandmay includesomemeasurementerrorwhichcan increase uncertainty in our estimates (Homer et al 2015)Additionallylandcoverwasonlymeasuredinthreeyearsrequiringus to imputevalues in13yearsAsa result all landcover changethatoccurredbetween2002and2006wasrecordedin2002whilechangesbetween2007and2011wererecorded in2007Thisap‐proximation isnot idealbutwasnecessarygiventhe5‐yearreso‐lutionofthedataThislowerresolutionofthelandcoverdatamayhavereducedthesensitivityofthemodel
Third wemay havemisspecified ourmodelWe tried to de‐velopbiologicallymeaningfulcovariatesbutwemightnotunder‐standthesystemaswellaswehopedItispossiblethatbirdvitalrates respond to climateon adifferent time scale (ie not a15‐yearmoving average) or to climateduringdifferent timesof theyearortodifferenttemperaturethresholdsortodifferentclimatecomponents (egprecipitation Illaacutenetal2014) It isalsopossi‐ble that thebirds respond to finerhabitat features than the landcovertypesweusedoratadifferentspatialscale(ienot400mPearsonampDawson2003)Alternatively theremaybe importantregionaldifferencesor interactionsamongcovariatesthatwedidnotcaptureOritispossiblethatchangesinclimateandhabitatintheirbreedingareassimplyhavelittleeffectonvitalratesforthesespecies relative to other factors such as competitionwith otherbirdspeciesorglobalchangeoccurringontheirwinteringgroundsinSouthAmerica(Dormann2007)Ofcoursethesemisspecifica‐tionissuesareinherenttoallobservationalstudiesandnotuniquetothisstudy
Itwould be possible for us to explore some of themisspec‐ification issues just mentioned For example we could rerunthe samemodelsusingnewcovariates representing a varietyoftemperature thresholds and habitat buffer sizes to search for astrongercorrelationExperiencesuggeststhatpersistencewouldeventuallyberewardedwithap‐valuelt005ora lowmodelse‐lection valueHowever one reason to avoid such exploration isthat it tends to lead toType Ierrorsoverfitmodels andbiasedparameterestimates(FiebergampJohnson2015)Furthermorewefind thecontrastbetweensignificant correlations for spatial co‐variatesandnonsignificantcorrelationsfortemporalcovariatesil‐luminatingIntypicalclimateenvelopestudieswewouldestimatethe relationshipbetweencovariatesand thecurrentdistributionofaspeciesmuchlikeourestimateofinitialoccupancy(Dormann2007)BecauseweobtainedsignificantparameterestimateswemightconcludethatwehaveidentifiedcovariatesthatdeterminetherangeofthisspeciesThenwemightusetheseresultstoproj‐ectthefuturerangeofthespeciesundervariousclimatescenariosbased on the (questionable) assumption that the covariatendashspe‐cies relationship will not change as climate and habitat change(Cuddington et al 2013 Elith et al 2010Gustafson 2013) In
this case the insignificant correlationwith the temporal covari‐atessuggeststhatouroccupancycovariatesmerelydescribetherangeofthesebirdswithoutdeterminingtherangeFurthermoreour results suggest that despite significant results in the staticportionof themodelwehaveyet to identify themost relevantcomponents of climate change shaping thedistributionof thesebirds although itmay be difficult to identify these componentsgiven the lackofobservedchangeduringa relatively short timeperiodFinallyourresultssuggestthatprojectionsbasedonthestaticportionofourmodelareunlikelytobereliabledespitetheirstatisticalsignificance
Ifourinterpretationiscorrectitsuggeststhatsomeprojectionsof futurerangeshiftsbasedonstaticmodelsrelyonunsupportedassumptionsandarenot reliableWehavedemonstratedadiffer‐entapproachthatdoesnotrequirethestrongassumptionsofstaticmodelingapproachesOurapproachwouldbemoreapttoidentifyfactorsthatguidespeciesrsquoresponsetoglobalchangeifboththespe‐ciesandthecovariatesexhibitedmoredynamismandifweanalyzedalongertimeperiodthatencompassedgreaterchange
ACKNOWLEDG MENTS
WethankKeithPardieckDaveZiolkowskiJrandthethousandsofhighlyskilledvolunteerswhohavecoordinatedandperformedtheNorthAmericanBreedingBirdSurvey for half a centuryWealsothanktheClimateResearchUnitattheUniversityofEastAngliaformakingvaluableclimatedataeasilyaccessibleMJCwasfinanciallysupported by the BBS and the USGS Northeast Climate ScienceCenterThecontentsofthisdocumentaretheresponsibilityoftheauthorsanddonotnecessarilyrepresenttheviewsoftheUSGSAnyuseoftradeproductorfirmnamesisfordescriptivepurposesonlyanddoesnotimplyendorsementbytheUSGovernment
DEDIC ATION
We dedicate this paper to the late Chandler S Robbins ldquoFatherrdquooftheNorthAmericanBreedingBirdSurvey (BBS)DatafromtheBBSformthebasisforthisspecificpaperandforhundredsofothersfocusedon topics ranging from local trends tocontinentalmacro‐ecologyChanwasauniquecombinationofbird‐loverandscientistwhosevisionandinquisitiveenthusiasmwereaninspirationtoallofusfortunateenoughtohaveknownhim
CONFLIC T OF INTERE S T
Nonedeclared
AUTHORSrsquo CONTRIBUTION
JACandJDNconceivedthestudyAJTandSWanalyzedhabitatandclimatedataMJCJEHandJDNperformedtheoccupancyanaly‐sisMJCledthewritingofthemanuscriptAllauthorsgavefinalap‐provalforpublication
emspensp emsp | emsp2001CLEMENT ET aL
DATA ACCE SSIBILIT Y
Raw data are available from the following sources Breeding BirdSurvey httpswwwpwrcusgsgovbbsrawdata PRISM climatedata httpwwwprismoregonstateedu NLCD land cover datahttpswwwmrlcgov Processed data will be archived with theDryadDigitalRepository
ORCID
Matthew J Clement httpsorcidorg0000‐0003‐4231‐7949
James D Nichols httpsorcidorg0000‐0002‐7631‐2890
Adam J Terando httpsorcidorg0000‐0002‐9280‐043X
James E Hines httpsorcidorg0000‐0001‐5478‐7230
Steven G Williams httpsorcidorg0000‐0003‐3760‐6818
R E FE R E N C E S
Anderson D R Burnham K P Gould W R amp Cherry S (2001)Concerns about finding effects that are actually spuriousWildlife Society Bulletin29311ndash316
Arauacutejo M B Cabeza M Thuiller W Hannah L amp WilliamsP H (2004) Would climate change drive species out of re‐serves An assessment of existing reserve‐selection meth‐ods Global Change Biology 10 1618ndash1626 httpsdoiorg101111j1365‐2486200400828x
Barbet‐MassinMampJetzW(2014)A40‐yearcontinent‐widemulti‐speciesassessmentofrelevantclimatepredictorsforspeciesdistri‐butionmodellingDiversity and Distributions201285ndash1295httpsdoiorg101111ddi12229
Barbet‐Massin M Thuiller W amp Jiguet F (2012) The fate ofEuropean breeding birds under climate land‐use and disper‐sal scenarios Global Change Biology 18 881ndash890 httpsdoiorg101111j1365‐2486201102552x
Bellard C Bertelsmeier C Leadley P Thuiller W ampCourchamp F (2012) Impacts of climate change on the fu‐ture of biodiversity Ecology Letters 15 365ndash377 httpsdoiorg101111j1461‐0248201101736x
Boumlhning‐Gaese K amp Bauer H G (1996) Changes in species abun‐dance distribution and diversity in a central European birdcommunity Conservation Biology 10 175ndash187 httpsdoiorg101046j1523‐1739199610010175x
BrommerJELehikoinenAampValkamaJ(2012)Thebreedingrangesof central European and Arctic bird speciesmove poleward PLoS ONE7e43648httpsdoiorg101371journalpone0043648
CalderWAampKingJR(1974)ThermalandcaloricrelationsofbirdsInDSFarnerampJRKing(Eds)Avian biology(Vol4pp259ndash413)NewYorkNYAcademicPress
CesaraccioCSpanoDDucePampSnyderRL(2001)Animprovedmodel for determining degree‐day values from daily temperaturedataInternational Journal of Biometeorology45161ndash169httpsdoiorg101007s004840100104
ChamberlinTC (1890)ThemethodofmultipleworkinghypothesesScience1592ndash96
ClementMJHinesJENicholsJDPardieckKLampZiolkowskiDJJr (2016)Estimating indicesofrangeshifts inbirdsusingdy‐namicmodelswhendetectionisimperfectGlobal Change Biology223273ndash3285httpsdoiorg101111gcb13283
Commission for Environmental Cooperation (1997) Ecological regionsofNorthAmericaTowardacommonperspectiveCommission for
EnvironmentalCooperationMontrealRetrievedfromftpftpepagovwedecoregionscec_naCEC_NAecopdf
Crowl TA Crist TO Parmenter R R BelovskyGamp LugoA E(2008)Thespreadofinvasivespeciesandinfectiousdiseaseasdriv‐ersofecosystemchangeFrontiers in Ecology and the Environment6238ndash246httpsdoiorg101890070151
Cuddington K FortinM J Gerber L RHastings A Liebhold AOConnorMampRayC(2013)Process‐basedmodelsarerequiredtomanageecologicalsystemsinachangingworldEcosphere41ndash12httpsdoiorg101890ES12‐001781
DalyCHalbleibMSmithJIGibsonWPDoggettMKTaylorGHhellipPasterisPP(2008)Physiographicallysensitivemappingofcli‐matologicaltemperatureandprecipitationacrosstheconterminousUnited States International Journal of Climatology 28 2031ndash2064httpsdoiorg101002joc1688
DawsonW (1958) Relation of oxygen consumption and evaporativewater losstotemperatureintheCardinalPhysiological Zoology3137ndash48httpsdoiorg101086physzool31130155377
DawsonWRampWhittowGC (2000)Regulationofbodytempera‐ture InG CWhittow (Ed)Sturkiersquos avian physiology (5th ed pp343ndash390)SanDiegoCAAcademicPress
DeCalestaDS(1994)Effectofwhite‐taileddeeronsongbirdswithinmanagedforestsinPennsylvaniaJournal of Wildlife Management58711ndash718httpsdoiorg1023073809685
DevictorVJulliardRCouvetDampJiguetF(2008)Birdsaretrack‐ing climatewarming but not fast enoughProceedings of the Royal Society of London B Biological Sciences2752743ndash2748httpsdoiorg101098rspb20080878
Dormann C F (2007) Promising the future Global change projec‐tionsofspeciesdistributionsBasic and Applied Ecology8387ndash397httpsdoiorg101016jbaae200611001
Dormann C F Schymanski S J Cabral J Chuine I Graham CHartigFhellipSingerA(2012)Correlationandprocessinspeciesdis‐tributionmodelsBridgingadichotomyJournal of Biogeography392119ndash2131httpsdoiorg101111j1365‐2699201102659x
EccelE(2010)WhatwecanasktohourlytemperaturerecordingPartIIHourlyinterpolationoftemperaturesforclimatologyandmodel‐lingItalian Journal of Agrometeorology1545ndash50
EccelEampCordanoE(2013)InterpolTHourlyinterpolationofmulti‐pletemperaturedailyseriesRpackageversion211RetrievedfromhttpsCRANR‐projectorgpackage=InterpolT
EglingtonSMampPearce‐HigginsJW(2012)Disentanglingtherela‐tiveimportanceofchangesinclimateandland‐useintensityindriv‐ingrecentbirdpopulationtrendsPLoS ONE7e30407httpsdoiorg101371journalpone0030407
Elith JKearneyMampPhillipsS (2010)Theartofmodelling range‐shifting species Methods in Ecology and Evolution 1 330ndash342httpsdoiorg101111j2041‐210X201000036x
FiebergJampJohnsonDH(2015)MMIMultimodelinferenceormod‐elswithmanagement implications Journal of Wildlife Management79708ndash718
FriggensMMampFinchDM (2015) Implicationsof climatechangeforbirdconservationinthesouthwesternUSunderthreealternativefuturesPLoS ONE10e0144089
FryJXianGZJinSDewitzJHomerCGYangLhellipWickhamJD (2011)Completionof the2006national landcoverdatabasefor theconterminousUnitedStatesPhotogrammetric Engineering amp Remote Sensing77858ndash864
Fuller R J Gregory R D Gibbons D W Marchant J HWilson J D Baillie S R amp Carter N (1995) Population de‐clines and range contractions among lowland farmland birdsin Britain Conservation Biology 9 1425ndash1441 httpsdoiorg101046j1523‐1739199509061425x
Gaston K J Blackburn T M Greenwood J J Gregory R DQuinn R M amp Lawton J H (2000) Abundancendashoccupancy
2002emsp |emsp emspensp CLEMENT ET aL
relationships Journal of Applied Ecology 37 39ndash59 httpsdoiorg101046j1365‐2664200000485x
GoldsteinRB(1974)RelationofmetabolismtoambienttemperatureintheverdinCondor76116ndash119httpsdoiorg1023071365995
GrinnellJ(1917)FieldtestsoftheoriesconcerningdistributionalcontrolAmerican Naturalist51115ndash128httpsdoiorg101086279591
GrosboisVGimenezOGaillardJMPradelRBarbraudCClobertJhellipWeimerskirchH(2008)Assessingtheimpactofclimatevari‐ation on survival in vertebrate populationsBiological Reviews 83357ndash399httpsdoiorg101111j1469‐185X200800047x
GustafsonEJ(2013)WhenrelationshipsestimatedinthepastcannotbeusedtopredictthefutureUsingmechanisticmodelstopredictlandscapeecologicaldynamicsinachangingworldLandscape Ecology281429ndash1437httpsdoiorg101007s10980‐013‐9927‐4
Hamel P B (1992) LandManagers guide to the birds of the SouthGeneralTechnicalReportSE‐22USDAForestServiceSoutheasternForestExperimentStationandTheNatureConservancyChapelHillNC
Hinds D S amp Calder W A (1973) Temperature regulation of thePyrrhuloxiaandtheArizonaCardinalPhysiological Zoology4655ndash71httpsdoiorg101086physzool46130152517
HinesJE(2006)PRESENCE Software to estimate patch occupancy and related parameters Laurel MD US Geological survey PatuxentWildlife Research Center Retrieved from httpwwwmbr‐pwrcusgsgovsoftwarepresencehtml
HinesJENicholsJDampCollazoJA(2014)Multiseasonoccupancymodels for correlated replicate surveys Methods in Ecology and Evolution5583ndash591httpsdoiorg1011112041‐210X12186
HinesJENicholsJDRoyleJAMacKenzieDIGopalaswamyAMKumarNampKaranthKU(2010)TigersontrailsOccupancymodeling for cluster sampling Ecological Applications 20 1456ndash1466httpsdoiorg10189009‐03211
HockeyPA SiramiCRidleyARMidgleyGFampBabikerHA(2011) Interrogating recent range changes in South African birdsConfounding signals from land use and climate change present achallenge for attribution Diversity and Distributions 17 254ndash261httpsdoiorg101111j1472‐4642201000741x
Hoegh‐Guldberg O Hughes L McIntyre S Lindenmayer D BParmesanCPossinghamHPampThomasCD (2008)AssistedcolonizationandrapidclimatechangeScience321345ndash346
Homer C Dewitz J Fry J Coan M Hossain N Larson C hellipWickham J (2007) Completion of the 2001 national land coverdatabase for the conterminous United States Photogrammetric Engineering amp Remote Sensing73337
HomerCDewitzJYangLJinSDanielsonPXianGhellipMegownK(2015)Completionofthe2011NationalLandCoverDatabasefortheconterminousUnitedStatesndashrepresentingadecadeoflandcoverchange informationPhotogrammetric Engineering amp Remote Sensing81345ndash354
HootenMBWikleCKDorazioRMampRoyleJA(2007)HierarchicalspatiotemporalmatrixmodelsforcharacterizinginvasionsBiometrics63558ndash567httpsdoiorg101111j1541‐0420200600725x
HosmerDWJrampLemeshowS(2000)Applied logistic regression(2nded)NewYorkNYJohnWileyampSons
Huntley B Collingham Y CGreen R EHiltonGM RahbekCamp Willis S G (2006) Potential impacts of climatic change upongeographical distributions of birds Ibis 148 8ndash28 httpsdoiorg101111j1474‐919X200600523x
IgleciaMNCollazoJAampMcKerrowAJ(2012)Useofoccupancymodels to evaluate expert knowledge‐based species‐habitat rela‐tionshipsAvian Conservation and Ecology7(2)13pRetrievedfromhttpwwwace‐ecoorgvol7iss2art5
IllaacutenJGThomasCDJonesJAWongWKShirleySMampBettsMG(2014)Precipitationandwintertemperaturepredictlong‐termrange‐scale abundancechanges inWesternNorthAmericanbirds
Global Change Biology 20 3351ndash3364 httpsdoiorg101111gcb12642
IoannidisJP (2005)WhymostpublishedresearchfindingsarefalsePLoS Medicine2e124
KeacuteryMGuillera‐ArroitaGampLahoz‐Monfort J J (2013)Analysingand mapping species range dynamics using occupancy modelsJournal of Biogeography 40 1463ndash1474 httpsdoiorg101111jbi12087
Khaliq I Hof C Prinzinger R Boumlhning‐Gaese K amp PfenningerM(2014) Global variation in thermal tolerances and vulnerability ofendotherms to climate change Proceedings of the Royal Society B Biological Sciences 281(1789) 20141097 httpsdoiorg101098rspb20141097
Laursen K amp Frikke J (2008) Hunting from motorboats displacesWadden Sea eiders Somateria mollissima from their favouredfeeding distribution Wildlife Biology 14 423ndash433 httpsdoiorg1029810909‐6396‐144423
LemoineNBauerHGPeintingerMampBoumlhning‐GaeseK (2007)Effectsof climate and land‐use changeon species abundance in acentralEuropeanbirdcommunityConservation Biology21495ndash503httpsdoiorg101111j1523‐1739200600633x
MacArthurRH(1972)Geographical ecology Patterns in the distribution of speciesNewYorkNYHarperampRow
MacKenzieDINicholsJDHinesJEKnutsonMGampFranklinAB(2003)Estimatingsiteoccupancycolonizationandlocalextinc‐tionwhenaspeciesisdetectedimperfectlyEcology842200ndash2207httpsdoiorg10189002‐3090
MacKenzieDINicholsJDLachmanGBDroegeSRoyleJAampLangtimmCA(2002)Estimatingsiteoccupancyrateswhendetec‐tionprobabilitiesarelessthanoneEcology832248ndash2255httpsdoiorg1018900012‐9658(2002)083[2248ESORWD]20CO2
MacKenzieDINicholsJDRoyleJAPollockKHBaileyLampHinesJE(2018)Occupancy estimation and modeling Inferring pat‐terns and dynamics of species occurrence (2nd ed) BurlingtonMAAcademicPress
MellesSJFortinMJLindsayKampBadzinskiD(2011)ExpandingnorthwardInfluenceofclimatechangeforestconnectivityandpop‐ulationprocessesonathreatenedspeciesrangeshiftGlobal Change Biology1717ndash31httpsdoiorg101111j1365‐2486201002214x
Morin X amp Thuiller W (2009) Comparing niche‐and process‐based models to reduce prediction uncertainty in species rangeshifts under climate change Ecology 90 1301ndash1313 httpsdoiorg10189008‐01341
MustinKAmarAampRedpathSM(2014)Colonizationandextinctiondynamicsofadecliningmigratorybirdareinfluencedbyclimateandhabitat degradation Ibis 156 788ndash798 httpsdoiorg101111ibi12173
NicholsJDThomasLampConnPB(2009)InferencesaboutlandbirdabundancefromcountdataRecentadvancesandfuturedirectionsInDLThomsonEJCoochampMJConroy(Eds)Modeling demo‐graphic processes in marked populations(pp201ndash235)NewYorkNYSpringer
OmernikJM(1995)EcoregionsAspatialframeworkforenvironmen‐talmanagement InWSDavisampTSimon(Eds)Biological assess‐ment and criteria Tools for water resource planning and decision making (pp49ndash62)BocaRatonFLLewisPublishers
OmernikJM(2004)Perspectivesonthenatureanddefinitionofeco‐logicalregionsEnvironmental Management34(Supplement1)S27ndashS38httpsdoiorg101007s00267‐003‐5197‐2
Omernik J M amp Griffith G E (2014) Ecoregions of the contermi‐nousUnited States Evolution of a hierarchical spatial frameworkEnvironmental Management541249ndash1266httpsdoiorg101007s00267‐014‐0364‐1
Pardieck K L ZiolkowskiD J JrampHudsonM‐A R (2015)NorthAmericanBreedingBirdSurveyDataset1966ndash2014version20140
emspensp emsp | emsp2003CLEMENT ET aL
USGeologicalSurveyPatuxentWildlifeResearchCenterRetrievedfromhttpswwwpwrcusgsgovbbsrawdata
ParmesanCampYoheG (2003)Agloballycoherentfingerprintofcli‐mate change impacts across natural systemsNature 421 37ndash42httpsdoiorg101038nature01286
PearsonRGampDawsonTP(2003)PredictingtheimpactsofclimatechangeonthedistributionofspeciesArebioclimateenvelopemod‐elsusefulGlobal Ecology and Biogeography12361ndash371
PlattJR(1964)StronginferenceScience146347ndash353PopperK(1959)The logic of scientific discoveryLondonUKHutchinsonRobbins C S Sauer J R Greenberg R S amp Droege S (1989)
Population declines in North American birds that migrate to theNeotropics Proceedings of the National Academy of Sciences 867658ndash7662httpsdoiorg101073pnas86197658
Robinson R A Wilson J D amp Crick H Q (2001) The impor‐tance of arable habitat for farmland birds in grassland land‐scapes Journal of Applied Ecology 38 1059ndash1069 httpsdoiorg101046j1365‐2664200100654x
Romesburg H C (1981) Wildlife science Gaining reliable knowl‐edge Journal of Wildlife Management 45 293ndash313 httpsdoiorg1023073807913
Root T (1988) Energy constraints on avian distributions and abun‐dancesEcology69330ndash339httpsdoiorg1023071940431
Ruiz‐GutieacuterrezVampZipkinEF (2011)Detectionbiasesyieldmis‐leadingpatternsof speciespersistence and colonization in frag‐mented landscapesEcosphere2 1ndash14 httpsdoiorg101890ES10‐002071
SauerJRHinesJEFallonJEPardieckKLZiolkowskiDJJrampLinkWA(2015)The North American breeding bird survey results and analysis 1966ndash2013 Version 01302015LaurelMDUSGSPatuxentWildlifeResearchCenterhttpswwwmbr‐pwrcusgsgovbbsbbshtml
Sauer J R LinkWA Fallon J E Pardieck K L amp ZiolkowskiDJJr(2013)TheNorthAmericanbreedingbirdsurvey1966ndash2011SummaryanalysisandspeciesaccountsNorth American Fauna791ndash32httpsdoiorg103996nafa790001
SeoaneJBustamanteJampDiacuteaz‐DelgadoR(2004)Competingrolesforlandscapevegetationtopographyandclimateinpredictivemod‐elsofbirddistributionEcological Modelling171209ndash222httpsdoiorg101016jecolmodel200308006
Sirami C Caplat P Popy S Clamens A Arlettaz R Jiguet F hellipMartin J‐L (2017) Impacts of global change on species distri‐butions Obstacles and solutions to integrate climate and landuse Global Ecology and Biogeography 26 385ndash394 httpsdoiorg101111geb12555
SkalskiJR(1996)Regressionofabundanceestimatesfrommarkrecap‐ture surveys against environmental covariatesCanadian Journal of
Fisheries and Aquatic Sciences53196ndash204httpsdoiorg101139f95‐169
SohlTL(2014)Therelativeimpactsofclimateandland‐usechangeonconterminousUnitedStatesbird species from2001 to2075PLoS ONE9e112251httpsdoiorg101371journalpone0112251
ThogmartinWESauerJRampKnutsonMG(2007)ModelingandmappingabundanceofAmericanwoodcockacrosstheMidwesternandNortheasternUnitedStatesJournal of Wildlife Management71376ndash382httpsdoiorg1021932005‐680
ThomasCDampLennonJJ (1999)Birdsextendtheirrangesnorth‐wardsNature399213httpsdoiorg10103820335
Trathan P N Garciacutea‐Borboroglu P Boersma D Bost C ACrawford R J CrossinG ThellipWienecke B (2015) Pollutionhabitatlossfishingandclimatechangeascriticalthreatstopen‐guins Conservation Biology 29 31ndash41 httpsdoiorg101111cobi12349
TravisJMJ(2003)Climatechangeandhabitatdestructionadeadlyanthropogenic cocktail Proceedings of the Royal Society of London Series B Biological Sciences 270(1514) 467ndash473 httpsdoiorg101098rspb20022246
UrbanMC (2015)Acceleratingextinction risk fromclimatechangeScience348571ndash573httpsdoiorg101126scienceaaa4984
WalshJWuebblesDHayhoeKKossinJKunkelKStephensGhellipSomervilleR(2014)OurchangingclimateInJMMelilloTCRichmondampGWYohe(Eds)Climate change impacts in the United States The Third National climate assessment(pp19ndash67)WashingtonDCUSGovernmentPrintingOffice
YackulicCBNichols JDReid JampDerR (2015)Topredict thenichemodelcolonizationandextinctionEcology9616ndash23httpsdoiorg10189014‐13611
Yalcin S amp Leroux S J (2018) An empirical test of the relative andcombinedeffectsofland‐coverandclimatechangeonlocalcoloni‐zationandextinctionGlobal Change Biology243849ndash3861httpsdoiorg101111gcb14169
ZhuKWoodallCWampClarkJS(2012)FailuretomigrateLackoftreerangeexpansioninresponsetoclimatechangeGlobal Change Biology181042ndash1052httpsdoiorg101111j1365‐2486201102571x
How to cite this articleClementMJNicholsJDCollazoJATerandoAJHinesJEWilliamsSGPartitioningglobalchangeAssessingtherelativeimportanceofchangesinclimateandlandcoverforchangesinaviandistributionEcol Evol 201991985ndash2003 httpsdoiorg101002ece34890

emspensp emsp | emsp1993CLEMENT ET aL
toModel3althoughnotsignificantlyduetothenumberofaddi‐tional parameters (χ2=2679df =28p=053)UnderModel 2estimatedcolonizationratesvariedfrom000in2008to030in1998whileextinctionratesvariedfrom001 in2001ndash2003and2010to004in2011WeusedModels45and6toestimatethepercentof improvedmodel fit (measuredby thechange indevi‐ance)generatedbyModel2thatcouldbeaccountedforbytempo‐ralvariationinclimateandhabitatcovariates(Table2)Temporalvariationinhabitataccountedfor04ofthechangeindeviancewhile temporal variation in climate accounted for 196Model6whichaccountedfortemporalvariationinbothhabitatandcli‐mateindicatedthatin1998colonizationinthesouthwestportionofthestudyareawasabovethelocalaveragewhileextinctioninthesouthwestwasbelowthelocalaverageindicatingarelativelystrongoccupancy trend in thesouthwest in thatyear (Figure3)By2012thispatternhadreversedyieldingarelativelyweakoc‐cupancytrendinthesouthwest(Figure3)NoneofthesemodelsrepresentedasignificantimprovementoverModel3(pgt026)Insomecasesthesignoftheestimatedeffectoftemporalvariationinclimateandhabitatwasoppositeoftheexpectedeffectbutinnoinstancesweretheseestimatessignificant(Table3)
32emsp|emspRed‐eyed Vireo
The best‐supportedmodel for Red‐eyedVireo included the samecovariatesasforEasternWoodPeweeyieldingaglobalmodelwith85parameters (Table1)TheHosmerndashLemeshowtest indicatedfitwasadequate for theRed‐eyedVireoglobalmodel forbothnaiumlvecolonization rates (χ2=540df =4p=025) and naiumlve extinctionrates(χ2=100df =4p=091)
Red‐eyedVireowere similarlywidespread in the study areawith an average probability of occupancy of 091 in the global
model (Figure1)Occupancydecreasedslightlyto090by2012incontrasttothe increase incountsreportedbytheBBS Initialoccupancywas positively related to available habitat and hoursofcoldstress (Table1)Probabilityofdetectionat the first stopwasrelativelyhighat038whileprobabilityofdetectionattheroute levelnearlyperfectat098Model3which includedspa‐tialvariationinclimateandhabitattothecolonizationandextinc‐tionmodelsimprovedmodelfitdramatically(χ2=41884df =6plt0001)relativetothenullmodelInkeepingwiththegreaterchangeindevianceforRed‐eyedVireoModel3estimatedmorespatialvariationinvitalratescomparedtoEasternWoodPeweeIn 1998 estimated colonization rates varied across BBS routesfrom 010 to 088 (mean of 049)while extinction rates rangedfrom000to048(meanof004Figure4)Model2whichallowedcolonization and extinction rates to vary each year improvedmodelfitrelativetoModel3butnotsignificantlyduetothenum‐berofadditionalparameters(χ2=3427df =28p=019)UnderModel2estimatedcolonizationratesvariedfrom039in2008to060 in2000while estimatedextinction rates varied from003in2006to007in1998WeusedModels45and6toestimatethepercentofthedeviancereductioninModel2thatcouldbeac‐countedforbytemporalvariationinclimateandhabitatcovariates(Table2)Wefoundthattemporalvariation inhabitataccountedfor01of the change indeviancewhile climate accounted for56Model 6 which accounted for temporal variation in bothhabitatandclimateindicatedthatin1998bothcolonizationandextinctionwereaboveaverageindicatinghigherturnoverearlyinthestudyperiod(Figure5)By2012bothcolonizationandextinc‐tion rateshaddeclined indicatinggreater stability inoccupancy(Figure 5) None of these models represented a significant im‐provementoverModel3(pgt074)Someoftheestimatedeffects
F I G U R E 1 emspOccupancyprobabilityfor(a)EasternWoodPeweeand(b)Red‐eyedVireointheeasternUnitedStates1997underModel2Occupancyvariesbytheamountofsuitablehabitathoursofheatstressandhoursofcoldstressin1997
1994emsp |emsp emspensp CLEMENT ET aL
oftemporalvariationinclimateandhabitatdifferedfromexpecta‐tionsbutnotsignificantly(Table3)
4emsp |emspDISCUSSION
Wefound that forbothbirdspecies initialoccupancycoloniza‐tionratesandextinctionratescorrelatedwithspatialvariationinclimateandhabitatcovariatesHowevercolonizationandextinc‐tionrateswerenotcorrelatedwithtemporalvariation inclimateand habitat covariates Our results correlating initial occupancywith environmental covariates are broadly speaking consistentwithbiologicalexpectationandaraftofpreviousstudiesthathaveindicatedasignificantrelationshipbetweenspeciesdistributionsandhabitat(RobinsonWilsonampCrick2001ThogmartinSauerampKnutson 2007) climate (Barbet‐Massinamp Jetz 2014BellardBertelsmeier Leadley Thuiller amp Courchamp 2012) or both(Seoane Bustamante amp Diacuteaz‐Delgado 2004 Sohl 2014)Howeverwesuggestthatestimatedrelationshipsbetweenspatialvariationinenvironmentalcovariatesandcolonizationandextinc‐tion rates are of greater ecological interest than relatively phe‐nomenological species distribution models or climate envelopemodels(Clementetal2016)Theestimatedvitalratesofcoloni‐zationandextinctionbringusclosertounderstandingthedynamicprocessunderlying thedistributionof these speciesGivencon‐stantvital rateswecanalsoproject theequilibrium levelofoc‐cupancyatsites ( i
i+i
atsite i)Wecancomparetheseprojected
equilibria tocurrentoccupancytoproject trends for thespeciesunder current environmental conditions We can also use
projectedchangesinkeyenvironmentalvariablestodeveloppre‐dicted rangechanges in the futureWeargue that theseprojec‐tions are more robust than those from static climate envelopemodels which assume that species are currently in equilibrium(Yackulicetal2015)Incontraststaticmodelscanoverestimaterange changes when the equilibrium does not hold (Morin ampThuiller2009)
Given the importanceofglobal changeweweremotivated toinvestigate factors affecting temporal aswell as spatial variationin vital rates In this case spatial variation must be incorporatedintosuchanalysesinordertoaccommodatethedifferentdynamicsexpectedindifferentportionsofaspeciesrangeWeviewthere‐lationshipbetweentemporalvariationincovariatevaluesandvitalrates as the most relevant to understanding the effect of globalchangeontherecentandfuturedistributionsofourstudyspeciesFurthermore a correlation between vital rates and spatially vary‐ingcovariatesmaynot indicateasimilarcorrelationwithtemporalchangesinthosesamecovariatesForexampleaspecieswithstrongdispersal constraintsmight have a strong spatial relationshipwithacovariatebutaweaktemporalrelationshipduetoits immobility(Devictor et al 2008) Similarly an estimated spatial relationshipmaybepurelyphenomenologicalduetothenatureofspatialvari‐ablesWhen spatial variables followgradients it is relatively easytoobtainstatisticallysignificantcorrelationsabsentanycausalre‐lationship(Grosboisetal2008)ForexampleifcolonizationratesandtemperaturesbothfollownorthndashsouthgradientstheymaybespatiallycorrelatedevenifcolonizationisgovernedbysomeotherunidentifiedfactorInthiscasewewouldexpecttheweakcorrela‐tion between temporal changes in temperature and colonization
F I G U R E 2 emspProbabilityof(a)colonizationand(b)extinctionforEasternWoodPeweeintheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1995CLEMENT ET aL
ratestobemoreindicativeofthetruerelationshipthanthestrongcorrelationwithspatialvariationintemperature
BBS surveys indicate different abundance trajectories for ourstudyspecieswithcounts increasingforRed‐eyedVireosandde‐creasing for EasternWood Pewees (Sauer et al 2015) Becauseabundanceandoccupancytendtobelinked(Gastonetal2000)weexpectedsomedivergenceinoccupancydynamicsWefoundthatestimatedoccupancywashighandstableforbothspeciesalthoughtherewasgreaterturnoverforRed‐eyedVireosForexampleunderModel3 (spatialvariationonly incovariates)bothaveragecoloni‐zationrates(049)andaverageextinctionrates(004)wereatleasttwiceashighforRed‐eyedVireoscomparedtoEasternWoodPewee(018and002respectively)Red‐eyedVireosexhibitedgreaterspa‐tialvariationincolonizationandextinctionrates(Figure4)althoughour selected covariates explained little temporal variation for thisspeciesEasternWoodPeweecolonizationandextinctionratesex‐hibitedmoreofalatitudinalgradientreflectingaspatialcorrelationwith extreme temperature (Figure 2)Our results also hinted at agreatercorrelationbetweentemporalchanges in temperatureandoccupancyvitalratesforEasternWoodPeweewhichcouldplayaroleinthedeclineofthisspeciesalthoughtheresultswerenotsig‐nificant(Table3)Alternatively ithasbeensuggestedthatEasternWoodPeweemaybedecliningduetoa lossofforestedwinteringhabitat(RobbinsSauerGreenbergampDroege1989)orduetodeer‐mediatedchanges in foreststructureonbreedinggrounds (deCal‐esta 1994) illustrating the complexity of ascertaining cause andeffectusingobservationalstudiesinthesenaturalsystems
Althoughrelativelyrare(Siramietal2017)otherstudieshaveinvestigated both climate and land cover effects on bird distribu‐tions Inagreementwithour finding that temporalvariation incli‐mate explainedmuchmore of themodel deviance than temporalvariation in habitat climate‐only species distribution models hadbetter cross‐validation support than habitat‐only models for 409Europeanbirdspecies(Barbet‐Massinetal2012)SimilarlymodelsestimatingthenumberofnewlyoccupiedareasbyHoodedWarblers(Wilsonia citrina) inOntariousingclimatedatahadbetter informa‐tioncriterionsupportthanmodelsusingforestdata(MellesFortinLindsay amp Badzinski 2011) In contrast land‐use models outper‐formedclimatemodels for18 farmlandbird species in theUnitedKingdom(EglingtonampPearce‐Higgins2012)Similarlyvegetation‐based species distributionmodels had superior performance thanclimate‐based models for 79 Spanish bird species (Seoane et al2004)andforthreebirdspeciesinNewMexico(FriggensampFinch2015) Two studies focused on colonization and extinction ratesfoundmixed results ForGardenWarblers (Sylvia borin) inBritainvegetationexplainedmorevariationincolonizationwhiletempera‐turesexplainedmorevariationinlocalextinction(MustinAmarampRedpath2014)For122birdspeciesinOntariothebest‐supportedcovariates (land cover or climate change) varied among species(YalcinampLeroux2018)
Thedivergentresultsamongstudiesregardingtheimportanceofclimateandlandcovermaypartlybeduetothebiologyofindividualspecies (Yalcinamp Leroux 2018) For example farmland birdsmayTA
BLE
2emspR2 DevfordifferentmodelsofcolonizationandextinctionprobabilitiesforbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012
Mod
el
Cova
riate
s on
colo
niza
tion
and
extin
ctio
nEa
ster
n W
ood
Pew
eeRe
d‐ey
ed V
ireo
Inte
rcep
tAv
erag
e H
abita
tAv
erag
e Cl
imat
eH
abita
t Dev
iatio
nCl
imat
e D
evia
tion
Year
Eff
ect
Δ minus
2LL
R2 Dev
Δ minus
2LL
R2 Dev
2X
XX
X0
0010
00
0010
0
6X
XX
XX
minus2138
202
minus3230
57
4X
XX
Xminus2154
196
minus3234
56
5X
XX
Xminus2667
04
minus3422
01
3X
XX
minus2679
0minus3427
0
1X
minus7098
NA
minus45311
NA
Not
eNAnotapplicable
1996emsp |emsp emspensp CLEMENT ET aL
beparticularlyaffectedbychangesinagriculturalintensitybecauseoftheirextensiveuseoffarms(EglingtonampPearce‐Higgins2012)Alternativelyithasbeenarguedthatspatialscalemayplayaroleintherelativeimportanceofglobalchangefactorswithclimatemoreimportantatlargescales(PearsonampDawson2003)Wenotethattwostudiesthatfoundalargerroleforclimatebothreliedonlower‐resolution bird surveys (05deg atlas in Barbet‐Massin et al 2012
10km2atlasinMellesetal2011)IntheBBSdatathatweanalyzedsurveyswereconductedevery800mbutwecalculatedoccupancyacross each 394km transect In addition the cited studies useddifferentdata sources anddiverse analysismethods complicatinginterpretationofthedifferingresults
While we estimated that climate explained more model de‐viance than habitat these estimated relationships were not
F I G U R E 3 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionrates((t)minus (t)minus )forEasternWoodPeweeintheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
emspensp emsp | emsp1997CLEMENT ET aL
TA B L E 3 emspEstimatedparametersforcorrelated‐detectiondynamicoccupancyModel6relatinghabitatandclimatecovariatestooccupancydynamicsofbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012Parameterestimatesindicatechangeinlogoddsofparametersinresponsetoz‐transformedcovariatesParametersψϑϑprimeγε p1p2andπdefinedintextForkeyparameters(boldtext)expectedsignsincluded
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
Ψintercept 2367 0177 4087 0381
Ψhabitat 0556 0163 2323 0288
Ψheatstress minus0011 0198 minus0322 0275
Ψcoldstress 0598 0166 1054 0251
ϑintercept minus1821 0026 minus1385 0010
ϑhabitat minus0055 0026 0516 0013
ϑheatstress minus0094 0030 0017 0008
ϑcoldstress 0172 0040 0256 0013
ϑprimeintercept 2292 0045 0426 0020
ϑprimehabitat 0316 0036 1210 0018
ϑprimeheatstress minus0006 0038 0054 0013
ϑprimecoldstress minus0622 0064 0875 0019
γintercept minus0884 0353 0023 0162
γhabitatdeviation + minus0292 0478 minus0240 1768
γheatstressdeviation minus minus1136 0607 0064 0194
γcoldstressdeviation minus minus0498 0474 2398 2253
γaveragehabitat 0095 0091 0788 0111
γaverageheatstress minus0024 0067 0004 0062
γaveragecoldstress 0227 0071 0333 0119
εintercept minus3957 0386 minus4471 0141
εhabitatdeviation minus minus0397 0605 0121 1492
εheatstressdeviation + 0643 0455 minus0034 0182
εcoldstressdeviation + 0153 0644 2042 2072
εaveragehabitat 0238 0090 minus1664 0123
εaverageheatstress minus0071 0092 minus0080 0074
εaveragecoldstress minus0307 0141 minus0939 0104
p1intercept minus2172 0043 1351 0039
p1 1998 minus0118 0048 minus0128 0058
p1 1999 minus0112 0049 minus0043 0058
p1 2000 minus0166 0050 minus0017 0059
p1 2001 minus0156 0049 0031 0061
p1 2002 minus0179 0049 0076 0062
p1 2003 minus0216 0051 0160 0064
p12004 minus0242 0051 0066 0061
p1 2005 minus0124 0050 0149 0061
p12006 minus0182 0051 0124 0060
p12007 minus0149 0050 0183 0062
p1 2008 minus0202 0052 0191 0064
p1 2009 minus0221 0052 0134 0062
p1 2010 minus0195 0051 0062 0060
p1 2011 minus0122 0051 0202 0063
(Continues)
1998emsp |emsp emspensp CLEMENT ET aL
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
p1 2012 minus0227 0055 0193 0063
p1stopnumber 0033 0009 minus0055 0011
p1stopnumbersquared minus0069 0010 minus0247 0013
p2intercept minus0921 0018 minus0555 0037
p2 1998 minus0138 0033 minus0121 0050
p2 1999 minus0137 0032 minus0025 0052
p2 2000 minus0164 0032 minus0027 0051
p2 2001 minus0077 0032 minus0059 0053
p2 2002 minus0101 0032 minus0012 0052
p2 2003 minus0199 0033 0085 0053
p22004 minus0186 0032 0113 0059
p2 2005 minus0146 0032 0082 0052
p22006 minus0155 0032 0135 0054
p22007 minus0153 0032 0124 0055
p2 2008 minus0168 0032 0073 0057
p2 2009 minus0077 0033 0100 0056
p2 2010 minus0114 0033 0071 0062
p2 2011 minus0079 0032 0120 0055
p2 2012 minus0186 0033 0030 0063
p2stopnumber 0045 0007 0165 0010
p2stopnumbersquared minus0105 0008 minus0182 0011
π 0533 0064 minus0505 0065
TA B L E 3 emsp (Continued)
F I G U R E 4 emspProbabilityof(a)colonizationand(b)extinctionforRed‐eyedVireointheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1999CLEMENT ET aL
significantanddidnotexplainalargefractionofvariationThereareseveralpotentialexplanationsforwhywedidnotfindasub‐stantialresponsetochangesinhabitatandclimateFirstourdatawerelackingindynamiceventstoinformourmodelsThecovari‐ateschangedrelativelylittleoverthetimeperiodmakingitdiffi‐culttodetectaresponseThelackofchangewaspartlybecauseoftherelativelyshort(16‐year)timeperiodstudiedAlsoweusedamovingaverageforclimatedatawhichtendstoreducedifferences
overshorttimeperiods Inadditionthestudyregionhasexperi‐encedlesswarmingoverthepastcenturythanotherpartsoftheplanet (Walshetal2014)Thebirddistributionsalso lackeddy‐namismBecauseextinctionrateswerelowtherewerefewlocalextinctioneventstoinformthemodelAlthoughcolonizationrateswerehighthefactthatinitialoccupancywashighandextinctionrateswerelowmeanttherewerefewopportunitiesforcoloniza‐tionevents
F I G U R E 5 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionratesforRed‐eyedVireointheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
2000emsp |emsp emspensp CLEMENT ET aL
Second measurement error may have obscured responses toglobalchangeThePRISMclimatedataareestimatedbyinterpola‐tionratherthandirectmeasurementacrossthestudyarea(Dalyetal2008)AssuchtheremaybeartifactsoranomaliesthatimpededourestimationandincreaseduncertaintySimilarlyNLCDdataareremotelysensedandmay includesomemeasurementerrorwhichcan increase uncertainty in our estimates (Homer et al 2015)Additionallylandcoverwasonlymeasuredinthreeyearsrequiringus to imputevalues in13yearsAsa result all landcover changethatoccurredbetween2002and2006wasrecordedin2002whilechangesbetween2007and2011wererecorded in2007Thisap‐proximation isnot idealbutwasnecessarygiventhe5‐yearreso‐lutionofthedataThislowerresolutionofthelandcoverdatamayhavereducedthesensitivityofthemodel
Third wemay havemisspecified ourmodelWe tried to de‐velopbiologicallymeaningfulcovariatesbutwemightnotunder‐standthesystemaswellaswehopedItispossiblethatbirdvitalrates respond to climateon adifferent time scale (ie not a15‐yearmoving average) or to climateduringdifferent timesof theyearortodifferenttemperaturethresholdsortodifferentclimatecomponents (egprecipitation Illaacutenetal2014) It isalsopossi‐ble that thebirds respond to finerhabitat features than the landcovertypesweusedoratadifferentspatialscale(ienot400mPearsonampDawson2003)Alternatively theremaybe importantregionaldifferencesor interactionsamongcovariatesthatwedidnotcaptureOritispossiblethatchangesinclimateandhabitatintheirbreedingareassimplyhavelittleeffectonvitalratesforthesespecies relative to other factors such as competitionwith otherbirdspeciesorglobalchangeoccurringontheirwinteringgroundsinSouthAmerica(Dormann2007)Ofcoursethesemisspecifica‐tionissuesareinherenttoallobservationalstudiesandnotuniquetothisstudy
Itwould be possible for us to explore some of themisspec‐ification issues just mentioned For example we could rerunthe samemodelsusingnewcovariates representing a varietyoftemperature thresholds and habitat buffer sizes to search for astrongercorrelationExperiencesuggeststhatpersistencewouldeventuallyberewardedwithap‐valuelt005ora lowmodelse‐lection valueHowever one reason to avoid such exploration isthat it tends to lead toType Ierrorsoverfitmodels andbiasedparameterestimates(FiebergampJohnson2015)Furthermorewefind thecontrastbetweensignificant correlations for spatial co‐variatesandnonsignificantcorrelationsfortemporalcovariatesil‐luminatingIntypicalclimateenvelopestudieswewouldestimatethe relationshipbetweencovariatesand thecurrentdistributionofaspeciesmuchlikeourestimateofinitialoccupancy(Dormann2007)BecauseweobtainedsignificantparameterestimateswemightconcludethatwehaveidentifiedcovariatesthatdeterminetherangeofthisspeciesThenwemightusetheseresultstoproj‐ectthefuturerangeofthespeciesundervariousclimatescenariosbased on the (questionable) assumption that the covariatendashspe‐cies relationship will not change as climate and habitat change(Cuddington et al 2013 Elith et al 2010Gustafson 2013) In
this case the insignificant correlationwith the temporal covari‐atessuggeststhatouroccupancycovariatesmerelydescribetherangeofthesebirdswithoutdeterminingtherangeFurthermoreour results suggest that despite significant results in the staticportionof themodelwehaveyet to identify themost relevantcomponents of climate change shaping thedistributionof thesebirds although itmay be difficult to identify these componentsgiven the lackofobservedchangeduringa relatively short timeperiodFinallyourresultssuggestthatprojectionsbasedonthestaticportionofourmodelareunlikelytobereliabledespitetheirstatisticalsignificance
Ifourinterpretationiscorrectitsuggeststhatsomeprojectionsof futurerangeshiftsbasedonstaticmodelsrelyonunsupportedassumptionsandarenot reliableWehavedemonstratedadiffer‐entapproachthatdoesnotrequirethestrongassumptionsofstaticmodelingapproachesOurapproachwouldbemoreapttoidentifyfactorsthatguidespeciesrsquoresponsetoglobalchangeifboththespe‐ciesandthecovariatesexhibitedmoredynamismandifweanalyzedalongertimeperiodthatencompassedgreaterchange
ACKNOWLEDG MENTS
WethankKeithPardieckDaveZiolkowskiJrandthethousandsofhighlyskilledvolunteerswhohavecoordinatedandperformedtheNorthAmericanBreedingBirdSurvey for half a centuryWealsothanktheClimateResearchUnitattheUniversityofEastAngliaformakingvaluableclimatedataeasilyaccessibleMJCwasfinanciallysupported by the BBS and the USGS Northeast Climate ScienceCenterThecontentsofthisdocumentaretheresponsibilityoftheauthorsanddonotnecessarilyrepresenttheviewsoftheUSGSAnyuseoftradeproductorfirmnamesisfordescriptivepurposesonlyanddoesnotimplyendorsementbytheUSGovernment
DEDIC ATION
We dedicate this paper to the late Chandler S Robbins ldquoFatherrdquooftheNorthAmericanBreedingBirdSurvey (BBS)DatafromtheBBSformthebasisforthisspecificpaperandforhundredsofothersfocusedon topics ranging from local trends tocontinentalmacro‐ecologyChanwasauniquecombinationofbird‐loverandscientistwhosevisionandinquisitiveenthusiasmwereaninspirationtoallofusfortunateenoughtohaveknownhim
CONFLIC T OF INTERE S T
Nonedeclared
AUTHORSrsquo CONTRIBUTION
JACandJDNconceivedthestudyAJTandSWanalyzedhabitatandclimatedataMJCJEHandJDNperformedtheoccupancyanaly‐sisMJCledthewritingofthemanuscriptAllauthorsgavefinalap‐provalforpublication
emspensp emsp | emsp2001CLEMENT ET aL
DATA ACCE SSIBILIT Y
Raw data are available from the following sources Breeding BirdSurvey httpswwwpwrcusgsgovbbsrawdata PRISM climatedata httpwwwprismoregonstateedu NLCD land cover datahttpswwwmrlcgov Processed data will be archived with theDryadDigitalRepository
ORCID
Matthew J Clement httpsorcidorg0000‐0003‐4231‐7949
James D Nichols httpsorcidorg0000‐0002‐7631‐2890
Adam J Terando httpsorcidorg0000‐0002‐9280‐043X
James E Hines httpsorcidorg0000‐0001‐5478‐7230
Steven G Williams httpsorcidorg0000‐0003‐3760‐6818
R E FE R E N C E S
Anderson D R Burnham K P Gould W R amp Cherry S (2001)Concerns about finding effects that are actually spuriousWildlife Society Bulletin29311ndash316
Arauacutejo M B Cabeza M Thuiller W Hannah L amp WilliamsP H (2004) Would climate change drive species out of re‐serves An assessment of existing reserve‐selection meth‐ods Global Change Biology 10 1618ndash1626 httpsdoiorg101111j1365‐2486200400828x
Barbet‐MassinMampJetzW(2014)A40‐yearcontinent‐widemulti‐speciesassessmentofrelevantclimatepredictorsforspeciesdistri‐butionmodellingDiversity and Distributions201285ndash1295httpsdoiorg101111ddi12229
Barbet‐Massin M Thuiller W amp Jiguet F (2012) The fate ofEuropean breeding birds under climate land‐use and disper‐sal scenarios Global Change Biology 18 881ndash890 httpsdoiorg101111j1365‐2486201102552x
Bellard C Bertelsmeier C Leadley P Thuiller W ampCourchamp F (2012) Impacts of climate change on the fu‐ture of biodiversity Ecology Letters 15 365ndash377 httpsdoiorg101111j1461‐0248201101736x
Boumlhning‐Gaese K amp Bauer H G (1996) Changes in species abun‐dance distribution and diversity in a central European birdcommunity Conservation Biology 10 175ndash187 httpsdoiorg101046j1523‐1739199610010175x
BrommerJELehikoinenAampValkamaJ(2012)Thebreedingrangesof central European and Arctic bird speciesmove poleward PLoS ONE7e43648httpsdoiorg101371journalpone0043648
CalderWAampKingJR(1974)ThermalandcaloricrelationsofbirdsInDSFarnerampJRKing(Eds)Avian biology(Vol4pp259ndash413)NewYorkNYAcademicPress
CesaraccioCSpanoDDucePampSnyderRL(2001)Animprovedmodel for determining degree‐day values from daily temperaturedataInternational Journal of Biometeorology45161ndash169httpsdoiorg101007s004840100104
ChamberlinTC (1890)ThemethodofmultipleworkinghypothesesScience1592ndash96
ClementMJHinesJENicholsJDPardieckKLampZiolkowskiDJJr (2016)Estimating indicesofrangeshifts inbirdsusingdy‐namicmodelswhendetectionisimperfectGlobal Change Biology223273ndash3285httpsdoiorg101111gcb13283
Commission for Environmental Cooperation (1997) Ecological regionsofNorthAmericaTowardacommonperspectiveCommission for
EnvironmentalCooperationMontrealRetrievedfromftpftpepagovwedecoregionscec_naCEC_NAecopdf
Crowl TA Crist TO Parmenter R R BelovskyGamp LugoA E(2008)Thespreadofinvasivespeciesandinfectiousdiseaseasdriv‐ersofecosystemchangeFrontiers in Ecology and the Environment6238ndash246httpsdoiorg101890070151
Cuddington K FortinM J Gerber L RHastings A Liebhold AOConnorMampRayC(2013)Process‐basedmodelsarerequiredtomanageecologicalsystemsinachangingworldEcosphere41ndash12httpsdoiorg101890ES12‐001781
DalyCHalbleibMSmithJIGibsonWPDoggettMKTaylorGHhellipPasterisPP(2008)Physiographicallysensitivemappingofcli‐matologicaltemperatureandprecipitationacrosstheconterminousUnited States International Journal of Climatology 28 2031ndash2064httpsdoiorg101002joc1688
DawsonW (1958) Relation of oxygen consumption and evaporativewater losstotemperatureintheCardinalPhysiological Zoology3137ndash48httpsdoiorg101086physzool31130155377
DawsonWRampWhittowGC (2000)Regulationofbodytempera‐ture InG CWhittow (Ed)Sturkiersquos avian physiology (5th ed pp343ndash390)SanDiegoCAAcademicPress
DeCalestaDS(1994)Effectofwhite‐taileddeeronsongbirdswithinmanagedforestsinPennsylvaniaJournal of Wildlife Management58711ndash718httpsdoiorg1023073809685
DevictorVJulliardRCouvetDampJiguetF(2008)Birdsaretrack‐ing climatewarming but not fast enoughProceedings of the Royal Society of London B Biological Sciences2752743ndash2748httpsdoiorg101098rspb20080878
Dormann C F (2007) Promising the future Global change projec‐tionsofspeciesdistributionsBasic and Applied Ecology8387ndash397httpsdoiorg101016jbaae200611001
Dormann C F Schymanski S J Cabral J Chuine I Graham CHartigFhellipSingerA(2012)Correlationandprocessinspeciesdis‐tributionmodelsBridgingadichotomyJournal of Biogeography392119ndash2131httpsdoiorg101111j1365‐2699201102659x
EccelE(2010)WhatwecanasktohourlytemperaturerecordingPartIIHourlyinterpolationoftemperaturesforclimatologyandmodel‐lingItalian Journal of Agrometeorology1545ndash50
EccelEampCordanoE(2013)InterpolTHourlyinterpolationofmulti‐pletemperaturedailyseriesRpackageversion211RetrievedfromhttpsCRANR‐projectorgpackage=InterpolT
EglingtonSMampPearce‐HigginsJW(2012)Disentanglingtherela‐tiveimportanceofchangesinclimateandland‐useintensityindriv‐ingrecentbirdpopulationtrendsPLoS ONE7e30407httpsdoiorg101371journalpone0030407
Elith JKearneyMampPhillipsS (2010)Theartofmodelling range‐shifting species Methods in Ecology and Evolution 1 330ndash342httpsdoiorg101111j2041‐210X201000036x
FiebergJampJohnsonDH(2015)MMIMultimodelinferenceormod‐elswithmanagement implications Journal of Wildlife Management79708ndash718
FriggensMMampFinchDM (2015) Implicationsof climatechangeforbirdconservationinthesouthwesternUSunderthreealternativefuturesPLoS ONE10e0144089
FryJXianGZJinSDewitzJHomerCGYangLhellipWickhamJD (2011)Completionof the2006national landcoverdatabasefor theconterminousUnitedStatesPhotogrammetric Engineering amp Remote Sensing77858ndash864
Fuller R J Gregory R D Gibbons D W Marchant J HWilson J D Baillie S R amp Carter N (1995) Population de‐clines and range contractions among lowland farmland birdsin Britain Conservation Biology 9 1425ndash1441 httpsdoiorg101046j1523‐1739199509061425x
Gaston K J Blackburn T M Greenwood J J Gregory R DQuinn R M amp Lawton J H (2000) Abundancendashoccupancy
2002emsp |emsp emspensp CLEMENT ET aL
relationships Journal of Applied Ecology 37 39ndash59 httpsdoiorg101046j1365‐2664200000485x
GoldsteinRB(1974)RelationofmetabolismtoambienttemperatureintheverdinCondor76116ndash119httpsdoiorg1023071365995
GrinnellJ(1917)FieldtestsoftheoriesconcerningdistributionalcontrolAmerican Naturalist51115ndash128httpsdoiorg101086279591
GrosboisVGimenezOGaillardJMPradelRBarbraudCClobertJhellipWeimerskirchH(2008)Assessingtheimpactofclimatevari‐ation on survival in vertebrate populationsBiological Reviews 83357ndash399httpsdoiorg101111j1469‐185X200800047x
GustafsonEJ(2013)WhenrelationshipsestimatedinthepastcannotbeusedtopredictthefutureUsingmechanisticmodelstopredictlandscapeecologicaldynamicsinachangingworldLandscape Ecology281429ndash1437httpsdoiorg101007s10980‐013‐9927‐4
Hamel P B (1992) LandManagers guide to the birds of the SouthGeneralTechnicalReportSE‐22USDAForestServiceSoutheasternForestExperimentStationandTheNatureConservancyChapelHillNC
Hinds D S amp Calder W A (1973) Temperature regulation of thePyrrhuloxiaandtheArizonaCardinalPhysiological Zoology4655ndash71httpsdoiorg101086physzool46130152517
HinesJE(2006)PRESENCE Software to estimate patch occupancy and related parameters Laurel MD US Geological survey PatuxentWildlife Research Center Retrieved from httpwwwmbr‐pwrcusgsgovsoftwarepresencehtml
HinesJENicholsJDampCollazoJA(2014)Multiseasonoccupancymodels for correlated replicate surveys Methods in Ecology and Evolution5583ndash591httpsdoiorg1011112041‐210X12186
HinesJENicholsJDRoyleJAMacKenzieDIGopalaswamyAMKumarNampKaranthKU(2010)TigersontrailsOccupancymodeling for cluster sampling Ecological Applications 20 1456ndash1466httpsdoiorg10189009‐03211
HockeyPA SiramiCRidleyARMidgleyGFampBabikerHA(2011) Interrogating recent range changes in South African birdsConfounding signals from land use and climate change present achallenge for attribution Diversity and Distributions 17 254ndash261httpsdoiorg101111j1472‐4642201000741x
Hoegh‐Guldberg O Hughes L McIntyre S Lindenmayer D BParmesanCPossinghamHPampThomasCD (2008)AssistedcolonizationandrapidclimatechangeScience321345ndash346
Homer C Dewitz J Fry J Coan M Hossain N Larson C hellipWickham J (2007) Completion of the 2001 national land coverdatabase for the conterminous United States Photogrammetric Engineering amp Remote Sensing73337
HomerCDewitzJYangLJinSDanielsonPXianGhellipMegownK(2015)Completionofthe2011NationalLandCoverDatabasefortheconterminousUnitedStatesndashrepresentingadecadeoflandcoverchange informationPhotogrammetric Engineering amp Remote Sensing81345ndash354
HootenMBWikleCKDorazioRMampRoyleJA(2007)HierarchicalspatiotemporalmatrixmodelsforcharacterizinginvasionsBiometrics63558ndash567httpsdoiorg101111j1541‐0420200600725x
HosmerDWJrampLemeshowS(2000)Applied logistic regression(2nded)NewYorkNYJohnWileyampSons
Huntley B Collingham Y CGreen R EHiltonGM RahbekCamp Willis S G (2006) Potential impacts of climatic change upongeographical distributions of birds Ibis 148 8ndash28 httpsdoiorg101111j1474‐919X200600523x
IgleciaMNCollazoJAampMcKerrowAJ(2012)Useofoccupancymodels to evaluate expert knowledge‐based species‐habitat rela‐tionshipsAvian Conservation and Ecology7(2)13pRetrievedfromhttpwwwace‐ecoorgvol7iss2art5
IllaacutenJGThomasCDJonesJAWongWKShirleySMampBettsMG(2014)Precipitationandwintertemperaturepredictlong‐termrange‐scale abundancechanges inWesternNorthAmericanbirds
Global Change Biology 20 3351ndash3364 httpsdoiorg101111gcb12642
IoannidisJP (2005)WhymostpublishedresearchfindingsarefalsePLoS Medicine2e124
KeacuteryMGuillera‐ArroitaGampLahoz‐Monfort J J (2013)Analysingand mapping species range dynamics using occupancy modelsJournal of Biogeography 40 1463ndash1474 httpsdoiorg101111jbi12087
Khaliq I Hof C Prinzinger R Boumlhning‐Gaese K amp PfenningerM(2014) Global variation in thermal tolerances and vulnerability ofendotherms to climate change Proceedings of the Royal Society B Biological Sciences 281(1789) 20141097 httpsdoiorg101098rspb20141097
Laursen K amp Frikke J (2008) Hunting from motorboats displacesWadden Sea eiders Somateria mollissima from their favouredfeeding distribution Wildlife Biology 14 423ndash433 httpsdoiorg1029810909‐6396‐144423
LemoineNBauerHGPeintingerMampBoumlhning‐GaeseK (2007)Effectsof climate and land‐use changeon species abundance in acentralEuropeanbirdcommunityConservation Biology21495ndash503httpsdoiorg101111j1523‐1739200600633x
MacArthurRH(1972)Geographical ecology Patterns in the distribution of speciesNewYorkNYHarperampRow
MacKenzieDINicholsJDHinesJEKnutsonMGampFranklinAB(2003)Estimatingsiteoccupancycolonizationandlocalextinc‐tionwhenaspeciesisdetectedimperfectlyEcology842200ndash2207httpsdoiorg10189002‐3090
MacKenzieDINicholsJDLachmanGBDroegeSRoyleJAampLangtimmCA(2002)Estimatingsiteoccupancyrateswhendetec‐tionprobabilitiesarelessthanoneEcology832248ndash2255httpsdoiorg1018900012‐9658(2002)083[2248ESORWD]20CO2
MacKenzieDINicholsJDRoyleJAPollockKHBaileyLampHinesJE(2018)Occupancy estimation and modeling Inferring pat‐terns and dynamics of species occurrence (2nd ed) BurlingtonMAAcademicPress
MellesSJFortinMJLindsayKampBadzinskiD(2011)ExpandingnorthwardInfluenceofclimatechangeforestconnectivityandpop‐ulationprocessesonathreatenedspeciesrangeshiftGlobal Change Biology1717ndash31httpsdoiorg101111j1365‐2486201002214x
Morin X amp Thuiller W (2009) Comparing niche‐and process‐based models to reduce prediction uncertainty in species rangeshifts under climate change Ecology 90 1301ndash1313 httpsdoiorg10189008‐01341
MustinKAmarAampRedpathSM(2014)Colonizationandextinctiondynamicsofadecliningmigratorybirdareinfluencedbyclimateandhabitat degradation Ibis 156 788ndash798 httpsdoiorg101111ibi12173
NicholsJDThomasLampConnPB(2009)InferencesaboutlandbirdabundancefromcountdataRecentadvancesandfuturedirectionsInDLThomsonEJCoochampMJConroy(Eds)Modeling demo‐graphic processes in marked populations(pp201ndash235)NewYorkNYSpringer
OmernikJM(1995)EcoregionsAspatialframeworkforenvironmen‐talmanagement InWSDavisampTSimon(Eds)Biological assess‐ment and criteria Tools for water resource planning and decision making (pp49ndash62)BocaRatonFLLewisPublishers
OmernikJM(2004)Perspectivesonthenatureanddefinitionofeco‐logicalregionsEnvironmental Management34(Supplement1)S27ndashS38httpsdoiorg101007s00267‐003‐5197‐2
Omernik J M amp Griffith G E (2014) Ecoregions of the contermi‐nousUnited States Evolution of a hierarchical spatial frameworkEnvironmental Management541249ndash1266httpsdoiorg101007s00267‐014‐0364‐1
Pardieck K L ZiolkowskiD J JrampHudsonM‐A R (2015)NorthAmericanBreedingBirdSurveyDataset1966ndash2014version20140
emspensp emsp | emsp2003CLEMENT ET aL
USGeologicalSurveyPatuxentWildlifeResearchCenterRetrievedfromhttpswwwpwrcusgsgovbbsrawdata
ParmesanCampYoheG (2003)Agloballycoherentfingerprintofcli‐mate change impacts across natural systemsNature 421 37ndash42httpsdoiorg101038nature01286
PearsonRGampDawsonTP(2003)PredictingtheimpactsofclimatechangeonthedistributionofspeciesArebioclimateenvelopemod‐elsusefulGlobal Ecology and Biogeography12361ndash371
PlattJR(1964)StronginferenceScience146347ndash353PopperK(1959)The logic of scientific discoveryLondonUKHutchinsonRobbins C S Sauer J R Greenberg R S amp Droege S (1989)
Population declines in North American birds that migrate to theNeotropics Proceedings of the National Academy of Sciences 867658ndash7662httpsdoiorg101073pnas86197658
Robinson R A Wilson J D amp Crick H Q (2001) The impor‐tance of arable habitat for farmland birds in grassland land‐scapes Journal of Applied Ecology 38 1059ndash1069 httpsdoiorg101046j1365‐2664200100654x
Romesburg H C (1981) Wildlife science Gaining reliable knowl‐edge Journal of Wildlife Management 45 293ndash313 httpsdoiorg1023073807913
Root T (1988) Energy constraints on avian distributions and abun‐dancesEcology69330ndash339httpsdoiorg1023071940431
Ruiz‐GutieacuterrezVampZipkinEF (2011)Detectionbiasesyieldmis‐leadingpatternsof speciespersistence and colonization in frag‐mented landscapesEcosphere2 1ndash14 httpsdoiorg101890ES10‐002071
SauerJRHinesJEFallonJEPardieckKLZiolkowskiDJJrampLinkWA(2015)The North American breeding bird survey results and analysis 1966ndash2013 Version 01302015LaurelMDUSGSPatuxentWildlifeResearchCenterhttpswwwmbr‐pwrcusgsgovbbsbbshtml
Sauer J R LinkWA Fallon J E Pardieck K L amp ZiolkowskiDJJr(2013)TheNorthAmericanbreedingbirdsurvey1966ndash2011SummaryanalysisandspeciesaccountsNorth American Fauna791ndash32httpsdoiorg103996nafa790001
SeoaneJBustamanteJampDiacuteaz‐DelgadoR(2004)Competingrolesforlandscapevegetationtopographyandclimateinpredictivemod‐elsofbirddistributionEcological Modelling171209ndash222httpsdoiorg101016jecolmodel200308006
Sirami C Caplat P Popy S Clamens A Arlettaz R Jiguet F hellipMartin J‐L (2017) Impacts of global change on species distri‐butions Obstacles and solutions to integrate climate and landuse Global Ecology and Biogeography 26 385ndash394 httpsdoiorg101111geb12555
SkalskiJR(1996)Regressionofabundanceestimatesfrommarkrecap‐ture surveys against environmental covariatesCanadian Journal of
Fisheries and Aquatic Sciences53196ndash204httpsdoiorg101139f95‐169
SohlTL(2014)Therelativeimpactsofclimateandland‐usechangeonconterminousUnitedStatesbird species from2001 to2075PLoS ONE9e112251httpsdoiorg101371journalpone0112251
ThogmartinWESauerJRampKnutsonMG(2007)ModelingandmappingabundanceofAmericanwoodcockacrosstheMidwesternandNortheasternUnitedStatesJournal of Wildlife Management71376ndash382httpsdoiorg1021932005‐680
ThomasCDampLennonJJ (1999)Birdsextendtheirrangesnorth‐wardsNature399213httpsdoiorg10103820335
Trathan P N Garciacutea‐Borboroglu P Boersma D Bost C ACrawford R J CrossinG ThellipWienecke B (2015) Pollutionhabitatlossfishingandclimatechangeascriticalthreatstopen‐guins Conservation Biology 29 31ndash41 httpsdoiorg101111cobi12349
TravisJMJ(2003)Climatechangeandhabitatdestructionadeadlyanthropogenic cocktail Proceedings of the Royal Society of London Series B Biological Sciences 270(1514) 467ndash473 httpsdoiorg101098rspb20022246
UrbanMC (2015)Acceleratingextinction risk fromclimatechangeScience348571ndash573httpsdoiorg101126scienceaaa4984
WalshJWuebblesDHayhoeKKossinJKunkelKStephensGhellipSomervilleR(2014)OurchangingclimateInJMMelilloTCRichmondampGWYohe(Eds)Climate change impacts in the United States The Third National climate assessment(pp19ndash67)WashingtonDCUSGovernmentPrintingOffice
YackulicCBNichols JDReid JampDerR (2015)Topredict thenichemodelcolonizationandextinctionEcology9616ndash23httpsdoiorg10189014‐13611
Yalcin S amp Leroux S J (2018) An empirical test of the relative andcombinedeffectsofland‐coverandclimatechangeonlocalcoloni‐zationandextinctionGlobal Change Biology243849ndash3861httpsdoiorg101111gcb14169
ZhuKWoodallCWampClarkJS(2012)FailuretomigrateLackoftreerangeexpansioninresponsetoclimatechangeGlobal Change Biology181042ndash1052httpsdoiorg101111j1365‐2486201102571x
How to cite this articleClementMJNicholsJDCollazoJATerandoAJHinesJEWilliamsSGPartitioningglobalchangeAssessingtherelativeimportanceofchangesinclimateandlandcoverforchangesinaviandistributionEcol Evol 201991985ndash2003 httpsdoiorg101002ece34890

1994emsp |emsp emspensp CLEMENT ET aL
oftemporalvariationinclimateandhabitatdifferedfromexpecta‐tionsbutnotsignificantly(Table3)
4emsp |emspDISCUSSION
Wefound that forbothbirdspecies initialoccupancycoloniza‐tionratesandextinctionratescorrelatedwithspatialvariationinclimateandhabitatcovariatesHowevercolonizationandextinc‐tionrateswerenotcorrelatedwithtemporalvariation inclimateand habitat covariates Our results correlating initial occupancywith environmental covariates are broadly speaking consistentwithbiologicalexpectationandaraftofpreviousstudiesthathaveindicatedasignificantrelationshipbetweenspeciesdistributionsandhabitat(RobinsonWilsonampCrick2001ThogmartinSauerampKnutson 2007) climate (Barbet‐Massinamp Jetz 2014BellardBertelsmeier Leadley Thuiller amp Courchamp 2012) or both(Seoane Bustamante amp Diacuteaz‐Delgado 2004 Sohl 2014)Howeverwesuggestthatestimatedrelationshipsbetweenspatialvariationinenvironmentalcovariatesandcolonizationandextinc‐tion rates are of greater ecological interest than relatively phe‐nomenological species distribution models or climate envelopemodels(Clementetal2016)Theestimatedvitalratesofcoloni‐zationandextinctionbringusclosertounderstandingthedynamicprocessunderlying thedistributionof these speciesGivencon‐stantvital rateswecanalsoproject theequilibrium levelofoc‐cupancyatsites ( i
i+i
atsite i)Wecancomparetheseprojected
equilibria tocurrentoccupancytoproject trends for thespeciesunder current environmental conditions We can also use
projectedchangesinkeyenvironmentalvariablestodeveloppre‐dicted rangechanges in the futureWeargue that theseprojec‐tions are more robust than those from static climate envelopemodels which assume that species are currently in equilibrium(Yackulicetal2015)Incontraststaticmodelscanoverestimaterange changes when the equilibrium does not hold (Morin ampThuiller2009)
Given the importanceofglobal changeweweremotivated toinvestigate factors affecting temporal aswell as spatial variationin vital rates In this case spatial variation must be incorporatedintosuchanalysesinordertoaccommodatethedifferentdynamicsexpectedindifferentportionsofaspeciesrangeWeviewthere‐lationshipbetweentemporalvariationincovariatevaluesandvitalrates as the most relevant to understanding the effect of globalchangeontherecentandfuturedistributionsofourstudyspeciesFurthermore a correlation between vital rates and spatially vary‐ingcovariatesmaynot indicateasimilarcorrelationwithtemporalchangesinthosesamecovariatesForexampleaspecieswithstrongdispersal constraintsmight have a strong spatial relationshipwithacovariatebutaweaktemporalrelationshipduetoits immobility(Devictor et al 2008) Similarly an estimated spatial relationshipmaybepurelyphenomenologicalduetothenatureofspatialvari‐ablesWhen spatial variables followgradients it is relatively easytoobtainstatisticallysignificantcorrelationsabsentanycausalre‐lationship(Grosboisetal2008)ForexampleifcolonizationratesandtemperaturesbothfollownorthndashsouthgradientstheymaybespatiallycorrelatedevenifcolonizationisgovernedbysomeotherunidentifiedfactorInthiscasewewouldexpecttheweakcorrela‐tion between temporal changes in temperature and colonization
F I G U R E 2 emspProbabilityof(a)colonizationand(b)extinctionforEasternWoodPeweeintheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1995CLEMENT ET aL
ratestobemoreindicativeofthetruerelationshipthanthestrongcorrelationwithspatialvariationintemperature
BBS surveys indicate different abundance trajectories for ourstudyspecieswithcounts increasingforRed‐eyedVireosandde‐creasing for EasternWood Pewees (Sauer et al 2015) Becauseabundanceandoccupancytendtobelinked(Gastonetal2000)weexpectedsomedivergenceinoccupancydynamicsWefoundthatestimatedoccupancywashighandstableforbothspeciesalthoughtherewasgreaterturnoverforRed‐eyedVireosForexampleunderModel3 (spatialvariationonly incovariates)bothaveragecoloni‐zationrates(049)andaverageextinctionrates(004)wereatleasttwiceashighforRed‐eyedVireoscomparedtoEasternWoodPewee(018and002respectively)Red‐eyedVireosexhibitedgreaterspa‐tialvariationincolonizationandextinctionrates(Figure4)althoughour selected covariates explained little temporal variation for thisspeciesEasternWoodPeweecolonizationandextinctionratesex‐hibitedmoreofalatitudinalgradientreflectingaspatialcorrelationwith extreme temperature (Figure 2)Our results also hinted at agreatercorrelationbetweentemporalchanges in temperatureandoccupancyvitalratesforEasternWoodPeweewhichcouldplayaroleinthedeclineofthisspeciesalthoughtheresultswerenotsig‐nificant(Table3)Alternatively ithasbeensuggestedthatEasternWoodPeweemaybedecliningduetoa lossofforestedwinteringhabitat(RobbinsSauerGreenbergampDroege1989)orduetodeer‐mediatedchanges in foreststructureonbreedinggrounds (deCal‐esta 1994) illustrating the complexity of ascertaining cause andeffectusingobservationalstudiesinthesenaturalsystems
Althoughrelativelyrare(Siramietal2017)otherstudieshaveinvestigated both climate and land cover effects on bird distribu‐tions Inagreementwithour finding that temporalvariation incli‐mate explainedmuchmore of themodel deviance than temporalvariation in habitat climate‐only species distribution models hadbetter cross‐validation support than habitat‐only models for 409Europeanbirdspecies(Barbet‐Massinetal2012)SimilarlymodelsestimatingthenumberofnewlyoccupiedareasbyHoodedWarblers(Wilsonia citrina) inOntariousingclimatedatahadbetter informa‐tioncriterionsupportthanmodelsusingforestdata(MellesFortinLindsay amp Badzinski 2011) In contrast land‐use models outper‐formedclimatemodels for18 farmlandbird species in theUnitedKingdom(EglingtonampPearce‐Higgins2012)Similarlyvegetation‐based species distributionmodels had superior performance thanclimate‐based models for 79 Spanish bird species (Seoane et al2004)andforthreebirdspeciesinNewMexico(FriggensampFinch2015) Two studies focused on colonization and extinction ratesfoundmixed results ForGardenWarblers (Sylvia borin) inBritainvegetationexplainedmorevariationincolonizationwhiletempera‐turesexplainedmorevariationinlocalextinction(MustinAmarampRedpath2014)For122birdspeciesinOntariothebest‐supportedcovariates (land cover or climate change) varied among species(YalcinampLeroux2018)
Thedivergentresultsamongstudiesregardingtheimportanceofclimateandlandcovermaypartlybeduetothebiologyofindividualspecies (Yalcinamp Leroux 2018) For example farmland birdsmayTA
BLE
2emspR2 DevfordifferentmodelsofcolonizationandextinctionprobabilitiesforbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012
Mod
el
Cova
riate
s on
colo
niza
tion
and
extin
ctio
nEa
ster
n W
ood
Pew
eeRe
d‐ey
ed V
ireo
Inte
rcep
tAv
erag
e H
abita
tAv
erag
e Cl
imat
eH
abita
t Dev
iatio
nCl
imat
e D
evia
tion
Year
Eff
ect
Δ minus
2LL
R2 Dev
Δ minus
2LL
R2 Dev
2X
XX
X0
0010
00
0010
0
6X
XX
XX
minus2138
202
minus3230
57
4X
XX
Xminus2154
196
minus3234
56
5X
XX
Xminus2667
04
minus3422
01
3X
XX
minus2679
0minus3427
0
1X
minus7098
NA
minus45311
NA
Not
eNAnotapplicable
1996emsp |emsp emspensp CLEMENT ET aL
beparticularlyaffectedbychangesinagriculturalintensitybecauseoftheirextensiveuseoffarms(EglingtonampPearce‐Higgins2012)Alternativelyithasbeenarguedthatspatialscalemayplayaroleintherelativeimportanceofglobalchangefactorswithclimatemoreimportantatlargescales(PearsonampDawson2003)Wenotethattwostudiesthatfoundalargerroleforclimatebothreliedonlower‐resolution bird surveys (05deg atlas in Barbet‐Massin et al 2012
10km2atlasinMellesetal2011)IntheBBSdatathatweanalyzedsurveyswereconductedevery800mbutwecalculatedoccupancyacross each 394km transect In addition the cited studies useddifferentdata sources anddiverse analysismethods complicatinginterpretationofthedifferingresults
While we estimated that climate explained more model de‐viance than habitat these estimated relationships were not
F I G U R E 3 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionrates((t)minus (t)minus )forEasternWoodPeweeintheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
emspensp emsp | emsp1997CLEMENT ET aL
TA B L E 3 emspEstimatedparametersforcorrelated‐detectiondynamicoccupancyModel6relatinghabitatandclimatecovariatestooccupancydynamicsofbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012Parameterestimatesindicatechangeinlogoddsofparametersinresponsetoz‐transformedcovariatesParametersψϑϑprimeγε p1p2andπdefinedintextForkeyparameters(boldtext)expectedsignsincluded
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
Ψintercept 2367 0177 4087 0381
Ψhabitat 0556 0163 2323 0288
Ψheatstress minus0011 0198 minus0322 0275
Ψcoldstress 0598 0166 1054 0251
ϑintercept minus1821 0026 minus1385 0010
ϑhabitat minus0055 0026 0516 0013
ϑheatstress minus0094 0030 0017 0008
ϑcoldstress 0172 0040 0256 0013
ϑprimeintercept 2292 0045 0426 0020
ϑprimehabitat 0316 0036 1210 0018
ϑprimeheatstress minus0006 0038 0054 0013
ϑprimecoldstress minus0622 0064 0875 0019
γintercept minus0884 0353 0023 0162
γhabitatdeviation + minus0292 0478 minus0240 1768
γheatstressdeviation minus minus1136 0607 0064 0194
γcoldstressdeviation minus minus0498 0474 2398 2253
γaveragehabitat 0095 0091 0788 0111
γaverageheatstress minus0024 0067 0004 0062
γaveragecoldstress 0227 0071 0333 0119
εintercept minus3957 0386 minus4471 0141
εhabitatdeviation minus minus0397 0605 0121 1492
εheatstressdeviation + 0643 0455 minus0034 0182
εcoldstressdeviation + 0153 0644 2042 2072
εaveragehabitat 0238 0090 minus1664 0123
εaverageheatstress minus0071 0092 minus0080 0074
εaveragecoldstress minus0307 0141 minus0939 0104
p1intercept minus2172 0043 1351 0039
p1 1998 minus0118 0048 minus0128 0058
p1 1999 minus0112 0049 minus0043 0058
p1 2000 minus0166 0050 minus0017 0059
p1 2001 minus0156 0049 0031 0061
p1 2002 minus0179 0049 0076 0062
p1 2003 minus0216 0051 0160 0064
p12004 minus0242 0051 0066 0061
p1 2005 minus0124 0050 0149 0061
p12006 minus0182 0051 0124 0060
p12007 minus0149 0050 0183 0062
p1 2008 minus0202 0052 0191 0064
p1 2009 minus0221 0052 0134 0062
p1 2010 minus0195 0051 0062 0060
p1 2011 minus0122 0051 0202 0063
(Continues)
1998emsp |emsp emspensp CLEMENT ET aL
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
p1 2012 minus0227 0055 0193 0063
p1stopnumber 0033 0009 minus0055 0011
p1stopnumbersquared minus0069 0010 minus0247 0013
p2intercept minus0921 0018 minus0555 0037
p2 1998 minus0138 0033 minus0121 0050
p2 1999 minus0137 0032 minus0025 0052
p2 2000 minus0164 0032 minus0027 0051
p2 2001 minus0077 0032 minus0059 0053
p2 2002 minus0101 0032 minus0012 0052
p2 2003 minus0199 0033 0085 0053
p22004 minus0186 0032 0113 0059
p2 2005 minus0146 0032 0082 0052
p22006 minus0155 0032 0135 0054
p22007 minus0153 0032 0124 0055
p2 2008 minus0168 0032 0073 0057
p2 2009 minus0077 0033 0100 0056
p2 2010 minus0114 0033 0071 0062
p2 2011 minus0079 0032 0120 0055
p2 2012 minus0186 0033 0030 0063
p2stopnumber 0045 0007 0165 0010
p2stopnumbersquared minus0105 0008 minus0182 0011
π 0533 0064 minus0505 0065
TA B L E 3 emsp (Continued)
F I G U R E 4 emspProbabilityof(a)colonizationand(b)extinctionforRed‐eyedVireointheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1999CLEMENT ET aL
significantanddidnotexplainalargefractionofvariationThereareseveralpotentialexplanationsforwhywedidnotfindasub‐stantialresponsetochangesinhabitatandclimateFirstourdatawerelackingindynamiceventstoinformourmodelsThecovari‐ateschangedrelativelylittleoverthetimeperiodmakingitdiffi‐culttodetectaresponseThelackofchangewaspartlybecauseoftherelativelyshort(16‐year)timeperiodstudiedAlsoweusedamovingaverageforclimatedatawhichtendstoreducedifferences
overshorttimeperiods Inadditionthestudyregionhasexperi‐encedlesswarmingoverthepastcenturythanotherpartsoftheplanet (Walshetal2014)Thebirddistributionsalso lackeddy‐namismBecauseextinctionrateswerelowtherewerefewlocalextinctioneventstoinformthemodelAlthoughcolonizationrateswerehighthefactthatinitialoccupancywashighandextinctionrateswerelowmeanttherewerefewopportunitiesforcoloniza‐tionevents
F I G U R E 5 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionratesforRed‐eyedVireointheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
2000emsp |emsp emspensp CLEMENT ET aL
Second measurement error may have obscured responses toglobalchangeThePRISMclimatedataareestimatedbyinterpola‐tionratherthandirectmeasurementacrossthestudyarea(Dalyetal2008)AssuchtheremaybeartifactsoranomaliesthatimpededourestimationandincreaseduncertaintySimilarlyNLCDdataareremotelysensedandmay includesomemeasurementerrorwhichcan increase uncertainty in our estimates (Homer et al 2015)Additionallylandcoverwasonlymeasuredinthreeyearsrequiringus to imputevalues in13yearsAsa result all landcover changethatoccurredbetween2002and2006wasrecordedin2002whilechangesbetween2007and2011wererecorded in2007Thisap‐proximation isnot idealbutwasnecessarygiventhe5‐yearreso‐lutionofthedataThislowerresolutionofthelandcoverdatamayhavereducedthesensitivityofthemodel
Third wemay havemisspecified ourmodelWe tried to de‐velopbiologicallymeaningfulcovariatesbutwemightnotunder‐standthesystemaswellaswehopedItispossiblethatbirdvitalrates respond to climateon adifferent time scale (ie not a15‐yearmoving average) or to climateduringdifferent timesof theyearortodifferenttemperaturethresholdsortodifferentclimatecomponents (egprecipitation Illaacutenetal2014) It isalsopossi‐ble that thebirds respond to finerhabitat features than the landcovertypesweusedoratadifferentspatialscale(ienot400mPearsonampDawson2003)Alternatively theremaybe importantregionaldifferencesor interactionsamongcovariatesthatwedidnotcaptureOritispossiblethatchangesinclimateandhabitatintheirbreedingareassimplyhavelittleeffectonvitalratesforthesespecies relative to other factors such as competitionwith otherbirdspeciesorglobalchangeoccurringontheirwinteringgroundsinSouthAmerica(Dormann2007)Ofcoursethesemisspecifica‐tionissuesareinherenttoallobservationalstudiesandnotuniquetothisstudy
Itwould be possible for us to explore some of themisspec‐ification issues just mentioned For example we could rerunthe samemodelsusingnewcovariates representing a varietyoftemperature thresholds and habitat buffer sizes to search for astrongercorrelationExperiencesuggeststhatpersistencewouldeventuallyberewardedwithap‐valuelt005ora lowmodelse‐lection valueHowever one reason to avoid such exploration isthat it tends to lead toType Ierrorsoverfitmodels andbiasedparameterestimates(FiebergampJohnson2015)Furthermorewefind thecontrastbetweensignificant correlations for spatial co‐variatesandnonsignificantcorrelationsfortemporalcovariatesil‐luminatingIntypicalclimateenvelopestudieswewouldestimatethe relationshipbetweencovariatesand thecurrentdistributionofaspeciesmuchlikeourestimateofinitialoccupancy(Dormann2007)BecauseweobtainedsignificantparameterestimateswemightconcludethatwehaveidentifiedcovariatesthatdeterminetherangeofthisspeciesThenwemightusetheseresultstoproj‐ectthefuturerangeofthespeciesundervariousclimatescenariosbased on the (questionable) assumption that the covariatendashspe‐cies relationship will not change as climate and habitat change(Cuddington et al 2013 Elith et al 2010Gustafson 2013) In
this case the insignificant correlationwith the temporal covari‐atessuggeststhatouroccupancycovariatesmerelydescribetherangeofthesebirdswithoutdeterminingtherangeFurthermoreour results suggest that despite significant results in the staticportionof themodelwehaveyet to identify themost relevantcomponents of climate change shaping thedistributionof thesebirds although itmay be difficult to identify these componentsgiven the lackofobservedchangeduringa relatively short timeperiodFinallyourresultssuggestthatprojectionsbasedonthestaticportionofourmodelareunlikelytobereliabledespitetheirstatisticalsignificance
Ifourinterpretationiscorrectitsuggeststhatsomeprojectionsof futurerangeshiftsbasedonstaticmodelsrelyonunsupportedassumptionsandarenot reliableWehavedemonstratedadiffer‐entapproachthatdoesnotrequirethestrongassumptionsofstaticmodelingapproachesOurapproachwouldbemoreapttoidentifyfactorsthatguidespeciesrsquoresponsetoglobalchangeifboththespe‐ciesandthecovariatesexhibitedmoredynamismandifweanalyzedalongertimeperiodthatencompassedgreaterchange
ACKNOWLEDG MENTS
WethankKeithPardieckDaveZiolkowskiJrandthethousandsofhighlyskilledvolunteerswhohavecoordinatedandperformedtheNorthAmericanBreedingBirdSurvey for half a centuryWealsothanktheClimateResearchUnitattheUniversityofEastAngliaformakingvaluableclimatedataeasilyaccessibleMJCwasfinanciallysupported by the BBS and the USGS Northeast Climate ScienceCenterThecontentsofthisdocumentaretheresponsibilityoftheauthorsanddonotnecessarilyrepresenttheviewsoftheUSGSAnyuseoftradeproductorfirmnamesisfordescriptivepurposesonlyanddoesnotimplyendorsementbytheUSGovernment
DEDIC ATION
We dedicate this paper to the late Chandler S Robbins ldquoFatherrdquooftheNorthAmericanBreedingBirdSurvey (BBS)DatafromtheBBSformthebasisforthisspecificpaperandforhundredsofothersfocusedon topics ranging from local trends tocontinentalmacro‐ecologyChanwasauniquecombinationofbird‐loverandscientistwhosevisionandinquisitiveenthusiasmwereaninspirationtoallofusfortunateenoughtohaveknownhim
CONFLIC T OF INTERE S T
Nonedeclared
AUTHORSrsquo CONTRIBUTION
JACandJDNconceivedthestudyAJTandSWanalyzedhabitatandclimatedataMJCJEHandJDNperformedtheoccupancyanaly‐sisMJCledthewritingofthemanuscriptAllauthorsgavefinalap‐provalforpublication
emspensp emsp | emsp2001CLEMENT ET aL
DATA ACCE SSIBILIT Y
Raw data are available from the following sources Breeding BirdSurvey httpswwwpwrcusgsgovbbsrawdata PRISM climatedata httpwwwprismoregonstateedu NLCD land cover datahttpswwwmrlcgov Processed data will be archived with theDryadDigitalRepository
ORCID
Matthew J Clement httpsorcidorg0000‐0003‐4231‐7949
James D Nichols httpsorcidorg0000‐0002‐7631‐2890
Adam J Terando httpsorcidorg0000‐0002‐9280‐043X
James E Hines httpsorcidorg0000‐0001‐5478‐7230
Steven G Williams httpsorcidorg0000‐0003‐3760‐6818
R E FE R E N C E S
Anderson D R Burnham K P Gould W R amp Cherry S (2001)Concerns about finding effects that are actually spuriousWildlife Society Bulletin29311ndash316
Arauacutejo M B Cabeza M Thuiller W Hannah L amp WilliamsP H (2004) Would climate change drive species out of re‐serves An assessment of existing reserve‐selection meth‐ods Global Change Biology 10 1618ndash1626 httpsdoiorg101111j1365‐2486200400828x
Barbet‐MassinMampJetzW(2014)A40‐yearcontinent‐widemulti‐speciesassessmentofrelevantclimatepredictorsforspeciesdistri‐butionmodellingDiversity and Distributions201285ndash1295httpsdoiorg101111ddi12229
Barbet‐Massin M Thuiller W amp Jiguet F (2012) The fate ofEuropean breeding birds under climate land‐use and disper‐sal scenarios Global Change Biology 18 881ndash890 httpsdoiorg101111j1365‐2486201102552x
Bellard C Bertelsmeier C Leadley P Thuiller W ampCourchamp F (2012) Impacts of climate change on the fu‐ture of biodiversity Ecology Letters 15 365ndash377 httpsdoiorg101111j1461‐0248201101736x
Boumlhning‐Gaese K amp Bauer H G (1996) Changes in species abun‐dance distribution and diversity in a central European birdcommunity Conservation Biology 10 175ndash187 httpsdoiorg101046j1523‐1739199610010175x
BrommerJELehikoinenAampValkamaJ(2012)Thebreedingrangesof central European and Arctic bird speciesmove poleward PLoS ONE7e43648httpsdoiorg101371journalpone0043648
CalderWAampKingJR(1974)ThermalandcaloricrelationsofbirdsInDSFarnerampJRKing(Eds)Avian biology(Vol4pp259ndash413)NewYorkNYAcademicPress
CesaraccioCSpanoDDucePampSnyderRL(2001)Animprovedmodel for determining degree‐day values from daily temperaturedataInternational Journal of Biometeorology45161ndash169httpsdoiorg101007s004840100104
ChamberlinTC (1890)ThemethodofmultipleworkinghypothesesScience1592ndash96
ClementMJHinesJENicholsJDPardieckKLampZiolkowskiDJJr (2016)Estimating indicesofrangeshifts inbirdsusingdy‐namicmodelswhendetectionisimperfectGlobal Change Biology223273ndash3285httpsdoiorg101111gcb13283
Commission for Environmental Cooperation (1997) Ecological regionsofNorthAmericaTowardacommonperspectiveCommission for
EnvironmentalCooperationMontrealRetrievedfromftpftpepagovwedecoregionscec_naCEC_NAecopdf
Crowl TA Crist TO Parmenter R R BelovskyGamp LugoA E(2008)Thespreadofinvasivespeciesandinfectiousdiseaseasdriv‐ersofecosystemchangeFrontiers in Ecology and the Environment6238ndash246httpsdoiorg101890070151
Cuddington K FortinM J Gerber L RHastings A Liebhold AOConnorMampRayC(2013)Process‐basedmodelsarerequiredtomanageecologicalsystemsinachangingworldEcosphere41ndash12httpsdoiorg101890ES12‐001781
DalyCHalbleibMSmithJIGibsonWPDoggettMKTaylorGHhellipPasterisPP(2008)Physiographicallysensitivemappingofcli‐matologicaltemperatureandprecipitationacrosstheconterminousUnited States International Journal of Climatology 28 2031ndash2064httpsdoiorg101002joc1688
DawsonW (1958) Relation of oxygen consumption and evaporativewater losstotemperatureintheCardinalPhysiological Zoology3137ndash48httpsdoiorg101086physzool31130155377
DawsonWRampWhittowGC (2000)Regulationofbodytempera‐ture InG CWhittow (Ed)Sturkiersquos avian physiology (5th ed pp343ndash390)SanDiegoCAAcademicPress
DeCalestaDS(1994)Effectofwhite‐taileddeeronsongbirdswithinmanagedforestsinPennsylvaniaJournal of Wildlife Management58711ndash718httpsdoiorg1023073809685
DevictorVJulliardRCouvetDampJiguetF(2008)Birdsaretrack‐ing climatewarming but not fast enoughProceedings of the Royal Society of London B Biological Sciences2752743ndash2748httpsdoiorg101098rspb20080878
Dormann C F (2007) Promising the future Global change projec‐tionsofspeciesdistributionsBasic and Applied Ecology8387ndash397httpsdoiorg101016jbaae200611001
Dormann C F Schymanski S J Cabral J Chuine I Graham CHartigFhellipSingerA(2012)Correlationandprocessinspeciesdis‐tributionmodelsBridgingadichotomyJournal of Biogeography392119ndash2131httpsdoiorg101111j1365‐2699201102659x
EccelE(2010)WhatwecanasktohourlytemperaturerecordingPartIIHourlyinterpolationoftemperaturesforclimatologyandmodel‐lingItalian Journal of Agrometeorology1545ndash50
EccelEampCordanoE(2013)InterpolTHourlyinterpolationofmulti‐pletemperaturedailyseriesRpackageversion211RetrievedfromhttpsCRANR‐projectorgpackage=InterpolT
EglingtonSMampPearce‐HigginsJW(2012)Disentanglingtherela‐tiveimportanceofchangesinclimateandland‐useintensityindriv‐ingrecentbirdpopulationtrendsPLoS ONE7e30407httpsdoiorg101371journalpone0030407
Elith JKearneyMampPhillipsS (2010)Theartofmodelling range‐shifting species Methods in Ecology and Evolution 1 330ndash342httpsdoiorg101111j2041‐210X201000036x
FiebergJampJohnsonDH(2015)MMIMultimodelinferenceormod‐elswithmanagement implications Journal of Wildlife Management79708ndash718
FriggensMMampFinchDM (2015) Implicationsof climatechangeforbirdconservationinthesouthwesternUSunderthreealternativefuturesPLoS ONE10e0144089
FryJXianGZJinSDewitzJHomerCGYangLhellipWickhamJD (2011)Completionof the2006national landcoverdatabasefor theconterminousUnitedStatesPhotogrammetric Engineering amp Remote Sensing77858ndash864
Fuller R J Gregory R D Gibbons D W Marchant J HWilson J D Baillie S R amp Carter N (1995) Population de‐clines and range contractions among lowland farmland birdsin Britain Conservation Biology 9 1425ndash1441 httpsdoiorg101046j1523‐1739199509061425x
Gaston K J Blackburn T M Greenwood J J Gregory R DQuinn R M amp Lawton J H (2000) Abundancendashoccupancy
2002emsp |emsp emspensp CLEMENT ET aL
relationships Journal of Applied Ecology 37 39ndash59 httpsdoiorg101046j1365‐2664200000485x
GoldsteinRB(1974)RelationofmetabolismtoambienttemperatureintheverdinCondor76116ndash119httpsdoiorg1023071365995
GrinnellJ(1917)FieldtestsoftheoriesconcerningdistributionalcontrolAmerican Naturalist51115ndash128httpsdoiorg101086279591
GrosboisVGimenezOGaillardJMPradelRBarbraudCClobertJhellipWeimerskirchH(2008)Assessingtheimpactofclimatevari‐ation on survival in vertebrate populationsBiological Reviews 83357ndash399httpsdoiorg101111j1469‐185X200800047x
GustafsonEJ(2013)WhenrelationshipsestimatedinthepastcannotbeusedtopredictthefutureUsingmechanisticmodelstopredictlandscapeecologicaldynamicsinachangingworldLandscape Ecology281429ndash1437httpsdoiorg101007s10980‐013‐9927‐4
Hamel P B (1992) LandManagers guide to the birds of the SouthGeneralTechnicalReportSE‐22USDAForestServiceSoutheasternForestExperimentStationandTheNatureConservancyChapelHillNC
Hinds D S amp Calder W A (1973) Temperature regulation of thePyrrhuloxiaandtheArizonaCardinalPhysiological Zoology4655ndash71httpsdoiorg101086physzool46130152517
HinesJE(2006)PRESENCE Software to estimate patch occupancy and related parameters Laurel MD US Geological survey PatuxentWildlife Research Center Retrieved from httpwwwmbr‐pwrcusgsgovsoftwarepresencehtml
HinesJENicholsJDampCollazoJA(2014)Multiseasonoccupancymodels for correlated replicate surveys Methods in Ecology and Evolution5583ndash591httpsdoiorg1011112041‐210X12186
HinesJENicholsJDRoyleJAMacKenzieDIGopalaswamyAMKumarNampKaranthKU(2010)TigersontrailsOccupancymodeling for cluster sampling Ecological Applications 20 1456ndash1466httpsdoiorg10189009‐03211
HockeyPA SiramiCRidleyARMidgleyGFampBabikerHA(2011) Interrogating recent range changes in South African birdsConfounding signals from land use and climate change present achallenge for attribution Diversity and Distributions 17 254ndash261httpsdoiorg101111j1472‐4642201000741x
Hoegh‐Guldberg O Hughes L McIntyre S Lindenmayer D BParmesanCPossinghamHPampThomasCD (2008)AssistedcolonizationandrapidclimatechangeScience321345ndash346
Homer C Dewitz J Fry J Coan M Hossain N Larson C hellipWickham J (2007) Completion of the 2001 national land coverdatabase for the conterminous United States Photogrammetric Engineering amp Remote Sensing73337
HomerCDewitzJYangLJinSDanielsonPXianGhellipMegownK(2015)Completionofthe2011NationalLandCoverDatabasefortheconterminousUnitedStatesndashrepresentingadecadeoflandcoverchange informationPhotogrammetric Engineering amp Remote Sensing81345ndash354
HootenMBWikleCKDorazioRMampRoyleJA(2007)HierarchicalspatiotemporalmatrixmodelsforcharacterizinginvasionsBiometrics63558ndash567httpsdoiorg101111j1541‐0420200600725x
HosmerDWJrampLemeshowS(2000)Applied logistic regression(2nded)NewYorkNYJohnWileyampSons
Huntley B Collingham Y CGreen R EHiltonGM RahbekCamp Willis S G (2006) Potential impacts of climatic change upongeographical distributions of birds Ibis 148 8ndash28 httpsdoiorg101111j1474‐919X200600523x
IgleciaMNCollazoJAampMcKerrowAJ(2012)Useofoccupancymodels to evaluate expert knowledge‐based species‐habitat rela‐tionshipsAvian Conservation and Ecology7(2)13pRetrievedfromhttpwwwace‐ecoorgvol7iss2art5
IllaacutenJGThomasCDJonesJAWongWKShirleySMampBettsMG(2014)Precipitationandwintertemperaturepredictlong‐termrange‐scale abundancechanges inWesternNorthAmericanbirds
Global Change Biology 20 3351ndash3364 httpsdoiorg101111gcb12642
IoannidisJP (2005)WhymostpublishedresearchfindingsarefalsePLoS Medicine2e124
KeacuteryMGuillera‐ArroitaGampLahoz‐Monfort J J (2013)Analysingand mapping species range dynamics using occupancy modelsJournal of Biogeography 40 1463ndash1474 httpsdoiorg101111jbi12087
Khaliq I Hof C Prinzinger R Boumlhning‐Gaese K amp PfenningerM(2014) Global variation in thermal tolerances and vulnerability ofendotherms to climate change Proceedings of the Royal Society B Biological Sciences 281(1789) 20141097 httpsdoiorg101098rspb20141097
Laursen K amp Frikke J (2008) Hunting from motorboats displacesWadden Sea eiders Somateria mollissima from their favouredfeeding distribution Wildlife Biology 14 423ndash433 httpsdoiorg1029810909‐6396‐144423
LemoineNBauerHGPeintingerMampBoumlhning‐GaeseK (2007)Effectsof climate and land‐use changeon species abundance in acentralEuropeanbirdcommunityConservation Biology21495ndash503httpsdoiorg101111j1523‐1739200600633x
MacArthurRH(1972)Geographical ecology Patterns in the distribution of speciesNewYorkNYHarperampRow
MacKenzieDINicholsJDHinesJEKnutsonMGampFranklinAB(2003)Estimatingsiteoccupancycolonizationandlocalextinc‐tionwhenaspeciesisdetectedimperfectlyEcology842200ndash2207httpsdoiorg10189002‐3090
MacKenzieDINicholsJDLachmanGBDroegeSRoyleJAampLangtimmCA(2002)Estimatingsiteoccupancyrateswhendetec‐tionprobabilitiesarelessthanoneEcology832248ndash2255httpsdoiorg1018900012‐9658(2002)083[2248ESORWD]20CO2
MacKenzieDINicholsJDRoyleJAPollockKHBaileyLampHinesJE(2018)Occupancy estimation and modeling Inferring pat‐terns and dynamics of species occurrence (2nd ed) BurlingtonMAAcademicPress
MellesSJFortinMJLindsayKampBadzinskiD(2011)ExpandingnorthwardInfluenceofclimatechangeforestconnectivityandpop‐ulationprocessesonathreatenedspeciesrangeshiftGlobal Change Biology1717ndash31httpsdoiorg101111j1365‐2486201002214x
Morin X amp Thuiller W (2009) Comparing niche‐and process‐based models to reduce prediction uncertainty in species rangeshifts under climate change Ecology 90 1301ndash1313 httpsdoiorg10189008‐01341
MustinKAmarAampRedpathSM(2014)Colonizationandextinctiondynamicsofadecliningmigratorybirdareinfluencedbyclimateandhabitat degradation Ibis 156 788ndash798 httpsdoiorg101111ibi12173
NicholsJDThomasLampConnPB(2009)InferencesaboutlandbirdabundancefromcountdataRecentadvancesandfuturedirectionsInDLThomsonEJCoochampMJConroy(Eds)Modeling demo‐graphic processes in marked populations(pp201ndash235)NewYorkNYSpringer
OmernikJM(1995)EcoregionsAspatialframeworkforenvironmen‐talmanagement InWSDavisampTSimon(Eds)Biological assess‐ment and criteria Tools for water resource planning and decision making (pp49ndash62)BocaRatonFLLewisPublishers
OmernikJM(2004)Perspectivesonthenatureanddefinitionofeco‐logicalregionsEnvironmental Management34(Supplement1)S27ndashS38httpsdoiorg101007s00267‐003‐5197‐2
Omernik J M amp Griffith G E (2014) Ecoregions of the contermi‐nousUnited States Evolution of a hierarchical spatial frameworkEnvironmental Management541249ndash1266httpsdoiorg101007s00267‐014‐0364‐1
Pardieck K L ZiolkowskiD J JrampHudsonM‐A R (2015)NorthAmericanBreedingBirdSurveyDataset1966ndash2014version20140
emspensp emsp | emsp2003CLEMENT ET aL
USGeologicalSurveyPatuxentWildlifeResearchCenterRetrievedfromhttpswwwpwrcusgsgovbbsrawdata
ParmesanCampYoheG (2003)Agloballycoherentfingerprintofcli‐mate change impacts across natural systemsNature 421 37ndash42httpsdoiorg101038nature01286
PearsonRGampDawsonTP(2003)PredictingtheimpactsofclimatechangeonthedistributionofspeciesArebioclimateenvelopemod‐elsusefulGlobal Ecology and Biogeography12361ndash371
PlattJR(1964)StronginferenceScience146347ndash353PopperK(1959)The logic of scientific discoveryLondonUKHutchinsonRobbins C S Sauer J R Greenberg R S amp Droege S (1989)
Population declines in North American birds that migrate to theNeotropics Proceedings of the National Academy of Sciences 867658ndash7662httpsdoiorg101073pnas86197658
Robinson R A Wilson J D amp Crick H Q (2001) The impor‐tance of arable habitat for farmland birds in grassland land‐scapes Journal of Applied Ecology 38 1059ndash1069 httpsdoiorg101046j1365‐2664200100654x
Romesburg H C (1981) Wildlife science Gaining reliable knowl‐edge Journal of Wildlife Management 45 293ndash313 httpsdoiorg1023073807913
Root T (1988) Energy constraints on avian distributions and abun‐dancesEcology69330ndash339httpsdoiorg1023071940431
Ruiz‐GutieacuterrezVampZipkinEF (2011)Detectionbiasesyieldmis‐leadingpatternsof speciespersistence and colonization in frag‐mented landscapesEcosphere2 1ndash14 httpsdoiorg101890ES10‐002071
SauerJRHinesJEFallonJEPardieckKLZiolkowskiDJJrampLinkWA(2015)The North American breeding bird survey results and analysis 1966ndash2013 Version 01302015LaurelMDUSGSPatuxentWildlifeResearchCenterhttpswwwmbr‐pwrcusgsgovbbsbbshtml
Sauer J R LinkWA Fallon J E Pardieck K L amp ZiolkowskiDJJr(2013)TheNorthAmericanbreedingbirdsurvey1966ndash2011SummaryanalysisandspeciesaccountsNorth American Fauna791ndash32httpsdoiorg103996nafa790001
SeoaneJBustamanteJampDiacuteaz‐DelgadoR(2004)Competingrolesforlandscapevegetationtopographyandclimateinpredictivemod‐elsofbirddistributionEcological Modelling171209ndash222httpsdoiorg101016jecolmodel200308006
Sirami C Caplat P Popy S Clamens A Arlettaz R Jiguet F hellipMartin J‐L (2017) Impacts of global change on species distri‐butions Obstacles and solutions to integrate climate and landuse Global Ecology and Biogeography 26 385ndash394 httpsdoiorg101111geb12555
SkalskiJR(1996)Regressionofabundanceestimatesfrommarkrecap‐ture surveys against environmental covariatesCanadian Journal of
Fisheries and Aquatic Sciences53196ndash204httpsdoiorg101139f95‐169
SohlTL(2014)Therelativeimpactsofclimateandland‐usechangeonconterminousUnitedStatesbird species from2001 to2075PLoS ONE9e112251httpsdoiorg101371journalpone0112251
ThogmartinWESauerJRampKnutsonMG(2007)ModelingandmappingabundanceofAmericanwoodcockacrosstheMidwesternandNortheasternUnitedStatesJournal of Wildlife Management71376ndash382httpsdoiorg1021932005‐680
ThomasCDampLennonJJ (1999)Birdsextendtheirrangesnorth‐wardsNature399213httpsdoiorg10103820335
Trathan P N Garciacutea‐Borboroglu P Boersma D Bost C ACrawford R J CrossinG ThellipWienecke B (2015) Pollutionhabitatlossfishingandclimatechangeascriticalthreatstopen‐guins Conservation Biology 29 31ndash41 httpsdoiorg101111cobi12349
TravisJMJ(2003)Climatechangeandhabitatdestructionadeadlyanthropogenic cocktail Proceedings of the Royal Society of London Series B Biological Sciences 270(1514) 467ndash473 httpsdoiorg101098rspb20022246
UrbanMC (2015)Acceleratingextinction risk fromclimatechangeScience348571ndash573httpsdoiorg101126scienceaaa4984
WalshJWuebblesDHayhoeKKossinJKunkelKStephensGhellipSomervilleR(2014)OurchangingclimateInJMMelilloTCRichmondampGWYohe(Eds)Climate change impacts in the United States The Third National climate assessment(pp19ndash67)WashingtonDCUSGovernmentPrintingOffice
YackulicCBNichols JDReid JampDerR (2015)Topredict thenichemodelcolonizationandextinctionEcology9616ndash23httpsdoiorg10189014‐13611
Yalcin S amp Leroux S J (2018) An empirical test of the relative andcombinedeffectsofland‐coverandclimatechangeonlocalcoloni‐zationandextinctionGlobal Change Biology243849ndash3861httpsdoiorg101111gcb14169
ZhuKWoodallCWampClarkJS(2012)FailuretomigrateLackoftreerangeexpansioninresponsetoclimatechangeGlobal Change Biology181042ndash1052httpsdoiorg101111j1365‐2486201102571x
How to cite this articleClementMJNicholsJDCollazoJATerandoAJHinesJEWilliamsSGPartitioningglobalchangeAssessingtherelativeimportanceofchangesinclimateandlandcoverforchangesinaviandistributionEcol Evol 201991985ndash2003 httpsdoiorg101002ece34890

emspensp emsp | emsp1995CLEMENT ET aL
ratestobemoreindicativeofthetruerelationshipthanthestrongcorrelationwithspatialvariationintemperature
BBS surveys indicate different abundance trajectories for ourstudyspecieswithcounts increasingforRed‐eyedVireosandde‐creasing for EasternWood Pewees (Sauer et al 2015) Becauseabundanceandoccupancytendtobelinked(Gastonetal2000)weexpectedsomedivergenceinoccupancydynamicsWefoundthatestimatedoccupancywashighandstableforbothspeciesalthoughtherewasgreaterturnoverforRed‐eyedVireosForexampleunderModel3 (spatialvariationonly incovariates)bothaveragecoloni‐zationrates(049)andaverageextinctionrates(004)wereatleasttwiceashighforRed‐eyedVireoscomparedtoEasternWoodPewee(018and002respectively)Red‐eyedVireosexhibitedgreaterspa‐tialvariationincolonizationandextinctionrates(Figure4)althoughour selected covariates explained little temporal variation for thisspeciesEasternWoodPeweecolonizationandextinctionratesex‐hibitedmoreofalatitudinalgradientreflectingaspatialcorrelationwith extreme temperature (Figure 2)Our results also hinted at agreatercorrelationbetweentemporalchanges in temperatureandoccupancyvitalratesforEasternWoodPeweewhichcouldplayaroleinthedeclineofthisspeciesalthoughtheresultswerenotsig‐nificant(Table3)Alternatively ithasbeensuggestedthatEasternWoodPeweemaybedecliningduetoa lossofforestedwinteringhabitat(RobbinsSauerGreenbergampDroege1989)orduetodeer‐mediatedchanges in foreststructureonbreedinggrounds (deCal‐esta 1994) illustrating the complexity of ascertaining cause andeffectusingobservationalstudiesinthesenaturalsystems
Althoughrelativelyrare(Siramietal2017)otherstudieshaveinvestigated both climate and land cover effects on bird distribu‐tions Inagreementwithour finding that temporalvariation incli‐mate explainedmuchmore of themodel deviance than temporalvariation in habitat climate‐only species distribution models hadbetter cross‐validation support than habitat‐only models for 409Europeanbirdspecies(Barbet‐Massinetal2012)SimilarlymodelsestimatingthenumberofnewlyoccupiedareasbyHoodedWarblers(Wilsonia citrina) inOntariousingclimatedatahadbetter informa‐tioncriterionsupportthanmodelsusingforestdata(MellesFortinLindsay amp Badzinski 2011) In contrast land‐use models outper‐formedclimatemodels for18 farmlandbird species in theUnitedKingdom(EglingtonampPearce‐Higgins2012)Similarlyvegetation‐based species distributionmodels had superior performance thanclimate‐based models for 79 Spanish bird species (Seoane et al2004)andforthreebirdspeciesinNewMexico(FriggensampFinch2015) Two studies focused on colonization and extinction ratesfoundmixed results ForGardenWarblers (Sylvia borin) inBritainvegetationexplainedmorevariationincolonizationwhiletempera‐turesexplainedmorevariationinlocalextinction(MustinAmarampRedpath2014)For122birdspeciesinOntariothebest‐supportedcovariates (land cover or climate change) varied among species(YalcinampLeroux2018)
Thedivergentresultsamongstudiesregardingtheimportanceofclimateandlandcovermaypartlybeduetothebiologyofindividualspecies (Yalcinamp Leroux 2018) For example farmland birdsmayTA
BLE
2emspR2 DevfordifferentmodelsofcolonizationandextinctionprobabilitiesforbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012
Mod
el
Cova
riate
s on
colo
niza
tion
and
extin
ctio
nEa
ster
n W
ood
Pew
eeRe
d‐ey
ed V
ireo
Inte
rcep
tAv
erag
e H
abita
tAv
erag
e Cl
imat
eH
abita
t Dev
iatio
nCl
imat
e D
evia
tion
Year
Eff
ect
Δ minus
2LL
R2 Dev
Δ minus
2LL
R2 Dev
2X
XX
X0
0010
00
0010
0
6X
XX
XX
minus2138
202
minus3230
57
4X
XX
Xminus2154
196
minus3234
56
5X
XX
Xminus2667
04
minus3422
01
3X
XX
minus2679
0minus3427
0
1X
minus7098
NA
minus45311
NA
Not
eNAnotapplicable
1996emsp |emsp emspensp CLEMENT ET aL
beparticularlyaffectedbychangesinagriculturalintensitybecauseoftheirextensiveuseoffarms(EglingtonampPearce‐Higgins2012)Alternativelyithasbeenarguedthatspatialscalemayplayaroleintherelativeimportanceofglobalchangefactorswithclimatemoreimportantatlargescales(PearsonampDawson2003)Wenotethattwostudiesthatfoundalargerroleforclimatebothreliedonlower‐resolution bird surveys (05deg atlas in Barbet‐Massin et al 2012
10km2atlasinMellesetal2011)IntheBBSdatathatweanalyzedsurveyswereconductedevery800mbutwecalculatedoccupancyacross each 394km transect In addition the cited studies useddifferentdata sources anddiverse analysismethods complicatinginterpretationofthedifferingresults
While we estimated that climate explained more model de‐viance than habitat these estimated relationships were not
F I G U R E 3 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionrates((t)minus (t)minus )forEasternWoodPeweeintheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
emspensp emsp | emsp1997CLEMENT ET aL
TA B L E 3 emspEstimatedparametersforcorrelated‐detectiondynamicoccupancyModel6relatinghabitatandclimatecovariatestooccupancydynamicsofbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012Parameterestimatesindicatechangeinlogoddsofparametersinresponsetoz‐transformedcovariatesParametersψϑϑprimeγε p1p2andπdefinedintextForkeyparameters(boldtext)expectedsignsincluded
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
Ψintercept 2367 0177 4087 0381
Ψhabitat 0556 0163 2323 0288
Ψheatstress minus0011 0198 minus0322 0275
Ψcoldstress 0598 0166 1054 0251
ϑintercept minus1821 0026 minus1385 0010
ϑhabitat minus0055 0026 0516 0013
ϑheatstress minus0094 0030 0017 0008
ϑcoldstress 0172 0040 0256 0013
ϑprimeintercept 2292 0045 0426 0020
ϑprimehabitat 0316 0036 1210 0018
ϑprimeheatstress minus0006 0038 0054 0013
ϑprimecoldstress minus0622 0064 0875 0019
γintercept minus0884 0353 0023 0162
γhabitatdeviation + minus0292 0478 minus0240 1768
γheatstressdeviation minus minus1136 0607 0064 0194
γcoldstressdeviation minus minus0498 0474 2398 2253
γaveragehabitat 0095 0091 0788 0111
γaverageheatstress minus0024 0067 0004 0062
γaveragecoldstress 0227 0071 0333 0119
εintercept minus3957 0386 minus4471 0141
εhabitatdeviation minus minus0397 0605 0121 1492
εheatstressdeviation + 0643 0455 minus0034 0182
εcoldstressdeviation + 0153 0644 2042 2072
εaveragehabitat 0238 0090 minus1664 0123
εaverageheatstress minus0071 0092 minus0080 0074
εaveragecoldstress minus0307 0141 minus0939 0104
p1intercept minus2172 0043 1351 0039
p1 1998 minus0118 0048 minus0128 0058
p1 1999 minus0112 0049 minus0043 0058
p1 2000 minus0166 0050 minus0017 0059
p1 2001 minus0156 0049 0031 0061
p1 2002 minus0179 0049 0076 0062
p1 2003 minus0216 0051 0160 0064
p12004 minus0242 0051 0066 0061
p1 2005 minus0124 0050 0149 0061
p12006 minus0182 0051 0124 0060
p12007 minus0149 0050 0183 0062
p1 2008 minus0202 0052 0191 0064
p1 2009 minus0221 0052 0134 0062
p1 2010 minus0195 0051 0062 0060
p1 2011 minus0122 0051 0202 0063
(Continues)
1998emsp |emsp emspensp CLEMENT ET aL
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
p1 2012 minus0227 0055 0193 0063
p1stopnumber 0033 0009 minus0055 0011
p1stopnumbersquared minus0069 0010 minus0247 0013
p2intercept minus0921 0018 minus0555 0037
p2 1998 minus0138 0033 minus0121 0050
p2 1999 minus0137 0032 minus0025 0052
p2 2000 minus0164 0032 minus0027 0051
p2 2001 minus0077 0032 minus0059 0053
p2 2002 minus0101 0032 minus0012 0052
p2 2003 minus0199 0033 0085 0053
p22004 minus0186 0032 0113 0059
p2 2005 minus0146 0032 0082 0052
p22006 minus0155 0032 0135 0054
p22007 minus0153 0032 0124 0055
p2 2008 minus0168 0032 0073 0057
p2 2009 minus0077 0033 0100 0056
p2 2010 minus0114 0033 0071 0062
p2 2011 minus0079 0032 0120 0055
p2 2012 minus0186 0033 0030 0063
p2stopnumber 0045 0007 0165 0010
p2stopnumbersquared minus0105 0008 minus0182 0011
π 0533 0064 minus0505 0065
TA B L E 3 emsp (Continued)
F I G U R E 4 emspProbabilityof(a)colonizationand(b)extinctionforRed‐eyedVireointheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1999CLEMENT ET aL
significantanddidnotexplainalargefractionofvariationThereareseveralpotentialexplanationsforwhywedidnotfindasub‐stantialresponsetochangesinhabitatandclimateFirstourdatawerelackingindynamiceventstoinformourmodelsThecovari‐ateschangedrelativelylittleoverthetimeperiodmakingitdiffi‐culttodetectaresponseThelackofchangewaspartlybecauseoftherelativelyshort(16‐year)timeperiodstudiedAlsoweusedamovingaverageforclimatedatawhichtendstoreducedifferences
overshorttimeperiods Inadditionthestudyregionhasexperi‐encedlesswarmingoverthepastcenturythanotherpartsoftheplanet (Walshetal2014)Thebirddistributionsalso lackeddy‐namismBecauseextinctionrateswerelowtherewerefewlocalextinctioneventstoinformthemodelAlthoughcolonizationrateswerehighthefactthatinitialoccupancywashighandextinctionrateswerelowmeanttherewerefewopportunitiesforcoloniza‐tionevents
F I G U R E 5 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionratesforRed‐eyedVireointheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
2000emsp |emsp emspensp CLEMENT ET aL
Second measurement error may have obscured responses toglobalchangeThePRISMclimatedataareestimatedbyinterpola‐tionratherthandirectmeasurementacrossthestudyarea(Dalyetal2008)AssuchtheremaybeartifactsoranomaliesthatimpededourestimationandincreaseduncertaintySimilarlyNLCDdataareremotelysensedandmay includesomemeasurementerrorwhichcan increase uncertainty in our estimates (Homer et al 2015)Additionallylandcoverwasonlymeasuredinthreeyearsrequiringus to imputevalues in13yearsAsa result all landcover changethatoccurredbetween2002and2006wasrecordedin2002whilechangesbetween2007and2011wererecorded in2007Thisap‐proximation isnot idealbutwasnecessarygiventhe5‐yearreso‐lutionofthedataThislowerresolutionofthelandcoverdatamayhavereducedthesensitivityofthemodel
Third wemay havemisspecified ourmodelWe tried to de‐velopbiologicallymeaningfulcovariatesbutwemightnotunder‐standthesystemaswellaswehopedItispossiblethatbirdvitalrates respond to climateon adifferent time scale (ie not a15‐yearmoving average) or to climateduringdifferent timesof theyearortodifferenttemperaturethresholdsortodifferentclimatecomponents (egprecipitation Illaacutenetal2014) It isalsopossi‐ble that thebirds respond to finerhabitat features than the landcovertypesweusedoratadifferentspatialscale(ienot400mPearsonampDawson2003)Alternatively theremaybe importantregionaldifferencesor interactionsamongcovariatesthatwedidnotcaptureOritispossiblethatchangesinclimateandhabitatintheirbreedingareassimplyhavelittleeffectonvitalratesforthesespecies relative to other factors such as competitionwith otherbirdspeciesorglobalchangeoccurringontheirwinteringgroundsinSouthAmerica(Dormann2007)Ofcoursethesemisspecifica‐tionissuesareinherenttoallobservationalstudiesandnotuniquetothisstudy
Itwould be possible for us to explore some of themisspec‐ification issues just mentioned For example we could rerunthe samemodelsusingnewcovariates representing a varietyoftemperature thresholds and habitat buffer sizes to search for astrongercorrelationExperiencesuggeststhatpersistencewouldeventuallyberewardedwithap‐valuelt005ora lowmodelse‐lection valueHowever one reason to avoid such exploration isthat it tends to lead toType Ierrorsoverfitmodels andbiasedparameterestimates(FiebergampJohnson2015)Furthermorewefind thecontrastbetweensignificant correlations for spatial co‐variatesandnonsignificantcorrelationsfortemporalcovariatesil‐luminatingIntypicalclimateenvelopestudieswewouldestimatethe relationshipbetweencovariatesand thecurrentdistributionofaspeciesmuchlikeourestimateofinitialoccupancy(Dormann2007)BecauseweobtainedsignificantparameterestimateswemightconcludethatwehaveidentifiedcovariatesthatdeterminetherangeofthisspeciesThenwemightusetheseresultstoproj‐ectthefuturerangeofthespeciesundervariousclimatescenariosbased on the (questionable) assumption that the covariatendashspe‐cies relationship will not change as climate and habitat change(Cuddington et al 2013 Elith et al 2010Gustafson 2013) In
this case the insignificant correlationwith the temporal covari‐atessuggeststhatouroccupancycovariatesmerelydescribetherangeofthesebirdswithoutdeterminingtherangeFurthermoreour results suggest that despite significant results in the staticportionof themodelwehaveyet to identify themost relevantcomponents of climate change shaping thedistributionof thesebirds although itmay be difficult to identify these componentsgiven the lackofobservedchangeduringa relatively short timeperiodFinallyourresultssuggestthatprojectionsbasedonthestaticportionofourmodelareunlikelytobereliabledespitetheirstatisticalsignificance
Ifourinterpretationiscorrectitsuggeststhatsomeprojectionsof futurerangeshiftsbasedonstaticmodelsrelyonunsupportedassumptionsandarenot reliableWehavedemonstratedadiffer‐entapproachthatdoesnotrequirethestrongassumptionsofstaticmodelingapproachesOurapproachwouldbemoreapttoidentifyfactorsthatguidespeciesrsquoresponsetoglobalchangeifboththespe‐ciesandthecovariatesexhibitedmoredynamismandifweanalyzedalongertimeperiodthatencompassedgreaterchange
ACKNOWLEDG MENTS
WethankKeithPardieckDaveZiolkowskiJrandthethousandsofhighlyskilledvolunteerswhohavecoordinatedandperformedtheNorthAmericanBreedingBirdSurvey for half a centuryWealsothanktheClimateResearchUnitattheUniversityofEastAngliaformakingvaluableclimatedataeasilyaccessibleMJCwasfinanciallysupported by the BBS and the USGS Northeast Climate ScienceCenterThecontentsofthisdocumentaretheresponsibilityoftheauthorsanddonotnecessarilyrepresenttheviewsoftheUSGSAnyuseoftradeproductorfirmnamesisfordescriptivepurposesonlyanddoesnotimplyendorsementbytheUSGovernment
DEDIC ATION
We dedicate this paper to the late Chandler S Robbins ldquoFatherrdquooftheNorthAmericanBreedingBirdSurvey (BBS)DatafromtheBBSformthebasisforthisspecificpaperandforhundredsofothersfocusedon topics ranging from local trends tocontinentalmacro‐ecologyChanwasauniquecombinationofbird‐loverandscientistwhosevisionandinquisitiveenthusiasmwereaninspirationtoallofusfortunateenoughtohaveknownhim
CONFLIC T OF INTERE S T
Nonedeclared
AUTHORSrsquo CONTRIBUTION
JACandJDNconceivedthestudyAJTandSWanalyzedhabitatandclimatedataMJCJEHandJDNperformedtheoccupancyanaly‐sisMJCledthewritingofthemanuscriptAllauthorsgavefinalap‐provalforpublication
emspensp emsp | emsp2001CLEMENT ET aL
DATA ACCE SSIBILIT Y
Raw data are available from the following sources Breeding BirdSurvey httpswwwpwrcusgsgovbbsrawdata PRISM climatedata httpwwwprismoregonstateedu NLCD land cover datahttpswwwmrlcgov Processed data will be archived with theDryadDigitalRepository
ORCID
Matthew J Clement httpsorcidorg0000‐0003‐4231‐7949
James D Nichols httpsorcidorg0000‐0002‐7631‐2890
Adam J Terando httpsorcidorg0000‐0002‐9280‐043X
James E Hines httpsorcidorg0000‐0001‐5478‐7230
Steven G Williams httpsorcidorg0000‐0003‐3760‐6818
R E FE R E N C E S
Anderson D R Burnham K P Gould W R amp Cherry S (2001)Concerns about finding effects that are actually spuriousWildlife Society Bulletin29311ndash316
Arauacutejo M B Cabeza M Thuiller W Hannah L amp WilliamsP H (2004) Would climate change drive species out of re‐serves An assessment of existing reserve‐selection meth‐ods Global Change Biology 10 1618ndash1626 httpsdoiorg101111j1365‐2486200400828x
Barbet‐MassinMampJetzW(2014)A40‐yearcontinent‐widemulti‐speciesassessmentofrelevantclimatepredictorsforspeciesdistri‐butionmodellingDiversity and Distributions201285ndash1295httpsdoiorg101111ddi12229
Barbet‐Massin M Thuiller W amp Jiguet F (2012) The fate ofEuropean breeding birds under climate land‐use and disper‐sal scenarios Global Change Biology 18 881ndash890 httpsdoiorg101111j1365‐2486201102552x
Bellard C Bertelsmeier C Leadley P Thuiller W ampCourchamp F (2012) Impacts of climate change on the fu‐ture of biodiversity Ecology Letters 15 365ndash377 httpsdoiorg101111j1461‐0248201101736x
Boumlhning‐Gaese K amp Bauer H G (1996) Changes in species abun‐dance distribution and diversity in a central European birdcommunity Conservation Biology 10 175ndash187 httpsdoiorg101046j1523‐1739199610010175x
BrommerJELehikoinenAampValkamaJ(2012)Thebreedingrangesof central European and Arctic bird speciesmove poleward PLoS ONE7e43648httpsdoiorg101371journalpone0043648
CalderWAampKingJR(1974)ThermalandcaloricrelationsofbirdsInDSFarnerampJRKing(Eds)Avian biology(Vol4pp259ndash413)NewYorkNYAcademicPress
CesaraccioCSpanoDDucePampSnyderRL(2001)Animprovedmodel for determining degree‐day values from daily temperaturedataInternational Journal of Biometeorology45161ndash169httpsdoiorg101007s004840100104
ChamberlinTC (1890)ThemethodofmultipleworkinghypothesesScience1592ndash96
ClementMJHinesJENicholsJDPardieckKLampZiolkowskiDJJr (2016)Estimating indicesofrangeshifts inbirdsusingdy‐namicmodelswhendetectionisimperfectGlobal Change Biology223273ndash3285httpsdoiorg101111gcb13283
Commission for Environmental Cooperation (1997) Ecological regionsofNorthAmericaTowardacommonperspectiveCommission for
EnvironmentalCooperationMontrealRetrievedfromftpftpepagovwedecoregionscec_naCEC_NAecopdf
Crowl TA Crist TO Parmenter R R BelovskyGamp LugoA E(2008)Thespreadofinvasivespeciesandinfectiousdiseaseasdriv‐ersofecosystemchangeFrontiers in Ecology and the Environment6238ndash246httpsdoiorg101890070151
Cuddington K FortinM J Gerber L RHastings A Liebhold AOConnorMampRayC(2013)Process‐basedmodelsarerequiredtomanageecologicalsystemsinachangingworldEcosphere41ndash12httpsdoiorg101890ES12‐001781
DalyCHalbleibMSmithJIGibsonWPDoggettMKTaylorGHhellipPasterisPP(2008)Physiographicallysensitivemappingofcli‐matologicaltemperatureandprecipitationacrosstheconterminousUnited States International Journal of Climatology 28 2031ndash2064httpsdoiorg101002joc1688
DawsonW (1958) Relation of oxygen consumption and evaporativewater losstotemperatureintheCardinalPhysiological Zoology3137ndash48httpsdoiorg101086physzool31130155377
DawsonWRampWhittowGC (2000)Regulationofbodytempera‐ture InG CWhittow (Ed)Sturkiersquos avian physiology (5th ed pp343ndash390)SanDiegoCAAcademicPress
DeCalestaDS(1994)Effectofwhite‐taileddeeronsongbirdswithinmanagedforestsinPennsylvaniaJournal of Wildlife Management58711ndash718httpsdoiorg1023073809685
DevictorVJulliardRCouvetDampJiguetF(2008)Birdsaretrack‐ing climatewarming but not fast enoughProceedings of the Royal Society of London B Biological Sciences2752743ndash2748httpsdoiorg101098rspb20080878
Dormann C F (2007) Promising the future Global change projec‐tionsofspeciesdistributionsBasic and Applied Ecology8387ndash397httpsdoiorg101016jbaae200611001
Dormann C F Schymanski S J Cabral J Chuine I Graham CHartigFhellipSingerA(2012)Correlationandprocessinspeciesdis‐tributionmodelsBridgingadichotomyJournal of Biogeography392119ndash2131httpsdoiorg101111j1365‐2699201102659x
EccelE(2010)WhatwecanasktohourlytemperaturerecordingPartIIHourlyinterpolationoftemperaturesforclimatologyandmodel‐lingItalian Journal of Agrometeorology1545ndash50
EccelEampCordanoE(2013)InterpolTHourlyinterpolationofmulti‐pletemperaturedailyseriesRpackageversion211RetrievedfromhttpsCRANR‐projectorgpackage=InterpolT
EglingtonSMampPearce‐HigginsJW(2012)Disentanglingtherela‐tiveimportanceofchangesinclimateandland‐useintensityindriv‐ingrecentbirdpopulationtrendsPLoS ONE7e30407httpsdoiorg101371journalpone0030407
Elith JKearneyMampPhillipsS (2010)Theartofmodelling range‐shifting species Methods in Ecology and Evolution 1 330ndash342httpsdoiorg101111j2041‐210X201000036x
FiebergJampJohnsonDH(2015)MMIMultimodelinferenceormod‐elswithmanagement implications Journal of Wildlife Management79708ndash718
FriggensMMampFinchDM (2015) Implicationsof climatechangeforbirdconservationinthesouthwesternUSunderthreealternativefuturesPLoS ONE10e0144089
FryJXianGZJinSDewitzJHomerCGYangLhellipWickhamJD (2011)Completionof the2006national landcoverdatabasefor theconterminousUnitedStatesPhotogrammetric Engineering amp Remote Sensing77858ndash864
Fuller R J Gregory R D Gibbons D W Marchant J HWilson J D Baillie S R amp Carter N (1995) Population de‐clines and range contractions among lowland farmland birdsin Britain Conservation Biology 9 1425ndash1441 httpsdoiorg101046j1523‐1739199509061425x
Gaston K J Blackburn T M Greenwood J J Gregory R DQuinn R M amp Lawton J H (2000) Abundancendashoccupancy
2002emsp |emsp emspensp CLEMENT ET aL
relationships Journal of Applied Ecology 37 39ndash59 httpsdoiorg101046j1365‐2664200000485x
GoldsteinRB(1974)RelationofmetabolismtoambienttemperatureintheverdinCondor76116ndash119httpsdoiorg1023071365995
GrinnellJ(1917)FieldtestsoftheoriesconcerningdistributionalcontrolAmerican Naturalist51115ndash128httpsdoiorg101086279591
GrosboisVGimenezOGaillardJMPradelRBarbraudCClobertJhellipWeimerskirchH(2008)Assessingtheimpactofclimatevari‐ation on survival in vertebrate populationsBiological Reviews 83357ndash399httpsdoiorg101111j1469‐185X200800047x
GustafsonEJ(2013)WhenrelationshipsestimatedinthepastcannotbeusedtopredictthefutureUsingmechanisticmodelstopredictlandscapeecologicaldynamicsinachangingworldLandscape Ecology281429ndash1437httpsdoiorg101007s10980‐013‐9927‐4
Hamel P B (1992) LandManagers guide to the birds of the SouthGeneralTechnicalReportSE‐22USDAForestServiceSoutheasternForestExperimentStationandTheNatureConservancyChapelHillNC
Hinds D S amp Calder W A (1973) Temperature regulation of thePyrrhuloxiaandtheArizonaCardinalPhysiological Zoology4655ndash71httpsdoiorg101086physzool46130152517
HinesJE(2006)PRESENCE Software to estimate patch occupancy and related parameters Laurel MD US Geological survey PatuxentWildlife Research Center Retrieved from httpwwwmbr‐pwrcusgsgovsoftwarepresencehtml
HinesJENicholsJDampCollazoJA(2014)Multiseasonoccupancymodels for correlated replicate surveys Methods in Ecology and Evolution5583ndash591httpsdoiorg1011112041‐210X12186
HinesJENicholsJDRoyleJAMacKenzieDIGopalaswamyAMKumarNampKaranthKU(2010)TigersontrailsOccupancymodeling for cluster sampling Ecological Applications 20 1456ndash1466httpsdoiorg10189009‐03211
HockeyPA SiramiCRidleyARMidgleyGFampBabikerHA(2011) Interrogating recent range changes in South African birdsConfounding signals from land use and climate change present achallenge for attribution Diversity and Distributions 17 254ndash261httpsdoiorg101111j1472‐4642201000741x
Hoegh‐Guldberg O Hughes L McIntyre S Lindenmayer D BParmesanCPossinghamHPampThomasCD (2008)AssistedcolonizationandrapidclimatechangeScience321345ndash346
Homer C Dewitz J Fry J Coan M Hossain N Larson C hellipWickham J (2007) Completion of the 2001 national land coverdatabase for the conterminous United States Photogrammetric Engineering amp Remote Sensing73337
HomerCDewitzJYangLJinSDanielsonPXianGhellipMegownK(2015)Completionofthe2011NationalLandCoverDatabasefortheconterminousUnitedStatesndashrepresentingadecadeoflandcoverchange informationPhotogrammetric Engineering amp Remote Sensing81345ndash354
HootenMBWikleCKDorazioRMampRoyleJA(2007)HierarchicalspatiotemporalmatrixmodelsforcharacterizinginvasionsBiometrics63558ndash567httpsdoiorg101111j1541‐0420200600725x
HosmerDWJrampLemeshowS(2000)Applied logistic regression(2nded)NewYorkNYJohnWileyampSons
Huntley B Collingham Y CGreen R EHiltonGM RahbekCamp Willis S G (2006) Potential impacts of climatic change upongeographical distributions of birds Ibis 148 8ndash28 httpsdoiorg101111j1474‐919X200600523x
IgleciaMNCollazoJAampMcKerrowAJ(2012)Useofoccupancymodels to evaluate expert knowledge‐based species‐habitat rela‐tionshipsAvian Conservation and Ecology7(2)13pRetrievedfromhttpwwwace‐ecoorgvol7iss2art5
IllaacutenJGThomasCDJonesJAWongWKShirleySMampBettsMG(2014)Precipitationandwintertemperaturepredictlong‐termrange‐scale abundancechanges inWesternNorthAmericanbirds
Global Change Biology 20 3351ndash3364 httpsdoiorg101111gcb12642
IoannidisJP (2005)WhymostpublishedresearchfindingsarefalsePLoS Medicine2e124
KeacuteryMGuillera‐ArroitaGampLahoz‐Monfort J J (2013)Analysingand mapping species range dynamics using occupancy modelsJournal of Biogeography 40 1463ndash1474 httpsdoiorg101111jbi12087
Khaliq I Hof C Prinzinger R Boumlhning‐Gaese K amp PfenningerM(2014) Global variation in thermal tolerances and vulnerability ofendotherms to climate change Proceedings of the Royal Society B Biological Sciences 281(1789) 20141097 httpsdoiorg101098rspb20141097
Laursen K amp Frikke J (2008) Hunting from motorboats displacesWadden Sea eiders Somateria mollissima from their favouredfeeding distribution Wildlife Biology 14 423ndash433 httpsdoiorg1029810909‐6396‐144423
LemoineNBauerHGPeintingerMampBoumlhning‐GaeseK (2007)Effectsof climate and land‐use changeon species abundance in acentralEuropeanbirdcommunityConservation Biology21495ndash503httpsdoiorg101111j1523‐1739200600633x
MacArthurRH(1972)Geographical ecology Patterns in the distribution of speciesNewYorkNYHarperampRow
MacKenzieDINicholsJDHinesJEKnutsonMGampFranklinAB(2003)Estimatingsiteoccupancycolonizationandlocalextinc‐tionwhenaspeciesisdetectedimperfectlyEcology842200ndash2207httpsdoiorg10189002‐3090
MacKenzieDINicholsJDLachmanGBDroegeSRoyleJAampLangtimmCA(2002)Estimatingsiteoccupancyrateswhendetec‐tionprobabilitiesarelessthanoneEcology832248ndash2255httpsdoiorg1018900012‐9658(2002)083[2248ESORWD]20CO2
MacKenzieDINicholsJDRoyleJAPollockKHBaileyLampHinesJE(2018)Occupancy estimation and modeling Inferring pat‐terns and dynamics of species occurrence (2nd ed) BurlingtonMAAcademicPress
MellesSJFortinMJLindsayKampBadzinskiD(2011)ExpandingnorthwardInfluenceofclimatechangeforestconnectivityandpop‐ulationprocessesonathreatenedspeciesrangeshiftGlobal Change Biology1717ndash31httpsdoiorg101111j1365‐2486201002214x
Morin X amp Thuiller W (2009) Comparing niche‐and process‐based models to reduce prediction uncertainty in species rangeshifts under climate change Ecology 90 1301ndash1313 httpsdoiorg10189008‐01341
MustinKAmarAampRedpathSM(2014)Colonizationandextinctiondynamicsofadecliningmigratorybirdareinfluencedbyclimateandhabitat degradation Ibis 156 788ndash798 httpsdoiorg101111ibi12173
NicholsJDThomasLampConnPB(2009)InferencesaboutlandbirdabundancefromcountdataRecentadvancesandfuturedirectionsInDLThomsonEJCoochampMJConroy(Eds)Modeling demo‐graphic processes in marked populations(pp201ndash235)NewYorkNYSpringer
OmernikJM(1995)EcoregionsAspatialframeworkforenvironmen‐talmanagement InWSDavisampTSimon(Eds)Biological assess‐ment and criteria Tools for water resource planning and decision making (pp49ndash62)BocaRatonFLLewisPublishers
OmernikJM(2004)Perspectivesonthenatureanddefinitionofeco‐logicalregionsEnvironmental Management34(Supplement1)S27ndashS38httpsdoiorg101007s00267‐003‐5197‐2
Omernik J M amp Griffith G E (2014) Ecoregions of the contermi‐nousUnited States Evolution of a hierarchical spatial frameworkEnvironmental Management541249ndash1266httpsdoiorg101007s00267‐014‐0364‐1
Pardieck K L ZiolkowskiD J JrampHudsonM‐A R (2015)NorthAmericanBreedingBirdSurveyDataset1966ndash2014version20140
emspensp emsp | emsp2003CLEMENT ET aL
USGeologicalSurveyPatuxentWildlifeResearchCenterRetrievedfromhttpswwwpwrcusgsgovbbsrawdata
ParmesanCampYoheG (2003)Agloballycoherentfingerprintofcli‐mate change impacts across natural systemsNature 421 37ndash42httpsdoiorg101038nature01286
PearsonRGampDawsonTP(2003)PredictingtheimpactsofclimatechangeonthedistributionofspeciesArebioclimateenvelopemod‐elsusefulGlobal Ecology and Biogeography12361ndash371
PlattJR(1964)StronginferenceScience146347ndash353PopperK(1959)The logic of scientific discoveryLondonUKHutchinsonRobbins C S Sauer J R Greenberg R S amp Droege S (1989)
Population declines in North American birds that migrate to theNeotropics Proceedings of the National Academy of Sciences 867658ndash7662httpsdoiorg101073pnas86197658
Robinson R A Wilson J D amp Crick H Q (2001) The impor‐tance of arable habitat for farmland birds in grassland land‐scapes Journal of Applied Ecology 38 1059ndash1069 httpsdoiorg101046j1365‐2664200100654x
Romesburg H C (1981) Wildlife science Gaining reliable knowl‐edge Journal of Wildlife Management 45 293ndash313 httpsdoiorg1023073807913
Root T (1988) Energy constraints on avian distributions and abun‐dancesEcology69330ndash339httpsdoiorg1023071940431
Ruiz‐GutieacuterrezVampZipkinEF (2011)Detectionbiasesyieldmis‐leadingpatternsof speciespersistence and colonization in frag‐mented landscapesEcosphere2 1ndash14 httpsdoiorg101890ES10‐002071
SauerJRHinesJEFallonJEPardieckKLZiolkowskiDJJrampLinkWA(2015)The North American breeding bird survey results and analysis 1966ndash2013 Version 01302015LaurelMDUSGSPatuxentWildlifeResearchCenterhttpswwwmbr‐pwrcusgsgovbbsbbshtml
Sauer J R LinkWA Fallon J E Pardieck K L amp ZiolkowskiDJJr(2013)TheNorthAmericanbreedingbirdsurvey1966ndash2011SummaryanalysisandspeciesaccountsNorth American Fauna791ndash32httpsdoiorg103996nafa790001
SeoaneJBustamanteJampDiacuteaz‐DelgadoR(2004)Competingrolesforlandscapevegetationtopographyandclimateinpredictivemod‐elsofbirddistributionEcological Modelling171209ndash222httpsdoiorg101016jecolmodel200308006
Sirami C Caplat P Popy S Clamens A Arlettaz R Jiguet F hellipMartin J‐L (2017) Impacts of global change on species distri‐butions Obstacles and solutions to integrate climate and landuse Global Ecology and Biogeography 26 385ndash394 httpsdoiorg101111geb12555
SkalskiJR(1996)Regressionofabundanceestimatesfrommarkrecap‐ture surveys against environmental covariatesCanadian Journal of
Fisheries and Aquatic Sciences53196ndash204httpsdoiorg101139f95‐169
SohlTL(2014)Therelativeimpactsofclimateandland‐usechangeonconterminousUnitedStatesbird species from2001 to2075PLoS ONE9e112251httpsdoiorg101371journalpone0112251
ThogmartinWESauerJRampKnutsonMG(2007)ModelingandmappingabundanceofAmericanwoodcockacrosstheMidwesternandNortheasternUnitedStatesJournal of Wildlife Management71376ndash382httpsdoiorg1021932005‐680
ThomasCDampLennonJJ (1999)Birdsextendtheirrangesnorth‐wardsNature399213httpsdoiorg10103820335
Trathan P N Garciacutea‐Borboroglu P Boersma D Bost C ACrawford R J CrossinG ThellipWienecke B (2015) Pollutionhabitatlossfishingandclimatechangeascriticalthreatstopen‐guins Conservation Biology 29 31ndash41 httpsdoiorg101111cobi12349
TravisJMJ(2003)Climatechangeandhabitatdestructionadeadlyanthropogenic cocktail Proceedings of the Royal Society of London Series B Biological Sciences 270(1514) 467ndash473 httpsdoiorg101098rspb20022246
UrbanMC (2015)Acceleratingextinction risk fromclimatechangeScience348571ndash573httpsdoiorg101126scienceaaa4984
WalshJWuebblesDHayhoeKKossinJKunkelKStephensGhellipSomervilleR(2014)OurchangingclimateInJMMelilloTCRichmondampGWYohe(Eds)Climate change impacts in the United States The Third National climate assessment(pp19ndash67)WashingtonDCUSGovernmentPrintingOffice
YackulicCBNichols JDReid JampDerR (2015)Topredict thenichemodelcolonizationandextinctionEcology9616ndash23httpsdoiorg10189014‐13611
Yalcin S amp Leroux S J (2018) An empirical test of the relative andcombinedeffectsofland‐coverandclimatechangeonlocalcoloni‐zationandextinctionGlobal Change Biology243849ndash3861httpsdoiorg101111gcb14169
ZhuKWoodallCWampClarkJS(2012)FailuretomigrateLackoftreerangeexpansioninresponsetoclimatechangeGlobal Change Biology181042ndash1052httpsdoiorg101111j1365‐2486201102571x
How to cite this articleClementMJNicholsJDCollazoJATerandoAJHinesJEWilliamsSGPartitioningglobalchangeAssessingtherelativeimportanceofchangesinclimateandlandcoverforchangesinaviandistributionEcol Evol 201991985ndash2003 httpsdoiorg101002ece34890
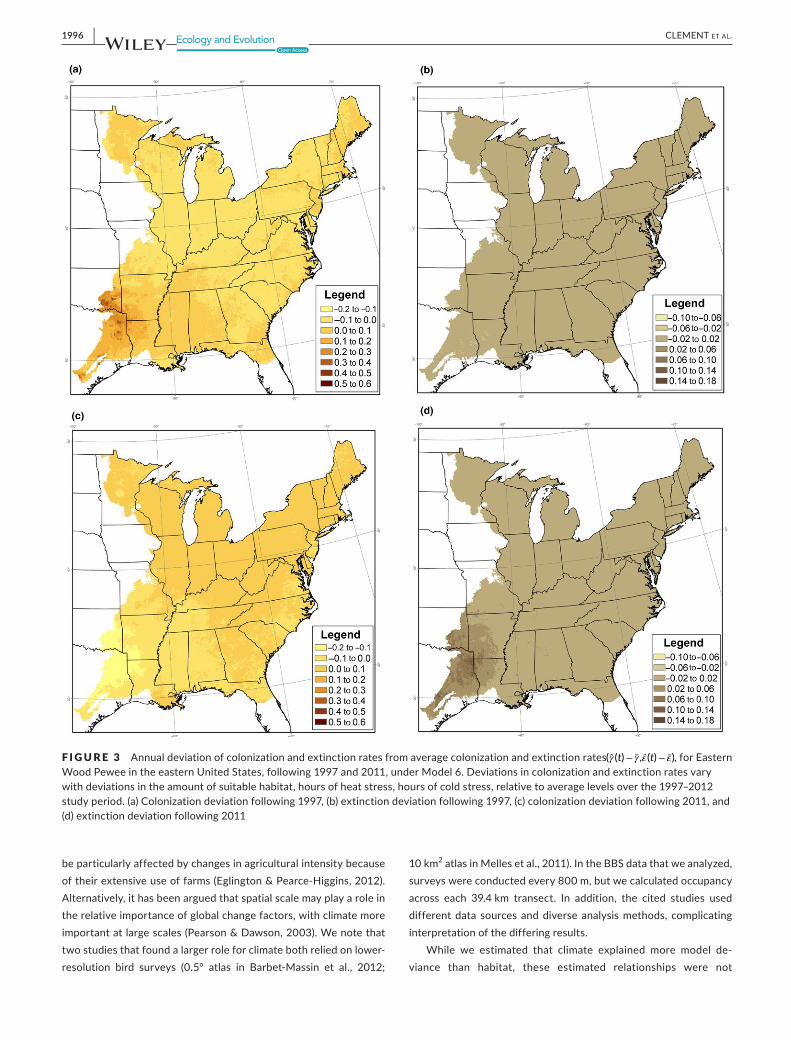
1996emsp |emsp emspensp CLEMENT ET aL
beparticularlyaffectedbychangesinagriculturalintensitybecauseoftheirextensiveuseoffarms(EglingtonampPearce‐Higgins2012)Alternativelyithasbeenarguedthatspatialscalemayplayaroleintherelativeimportanceofglobalchangefactorswithclimatemoreimportantatlargescales(PearsonampDawson2003)Wenotethattwostudiesthatfoundalargerroleforclimatebothreliedonlower‐resolution bird surveys (05deg atlas in Barbet‐Massin et al 2012
10km2atlasinMellesetal2011)IntheBBSdatathatweanalyzedsurveyswereconductedevery800mbutwecalculatedoccupancyacross each 394km transect In addition the cited studies useddifferentdata sources anddiverse analysismethods complicatinginterpretationofthedifferingresults
While we estimated that climate explained more model de‐viance than habitat these estimated relationships were not
F I G U R E 3 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionrates((t)minus (t)minus )forEasternWoodPeweeintheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
emspensp emsp | emsp1997CLEMENT ET aL
TA B L E 3 emspEstimatedparametersforcorrelated‐detectiondynamicoccupancyModel6relatinghabitatandclimatecovariatestooccupancydynamicsofbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012Parameterestimatesindicatechangeinlogoddsofparametersinresponsetoz‐transformedcovariatesParametersψϑϑprimeγε p1p2andπdefinedintextForkeyparameters(boldtext)expectedsignsincluded
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
Ψintercept 2367 0177 4087 0381
Ψhabitat 0556 0163 2323 0288
Ψheatstress minus0011 0198 minus0322 0275
Ψcoldstress 0598 0166 1054 0251
ϑintercept minus1821 0026 minus1385 0010
ϑhabitat minus0055 0026 0516 0013
ϑheatstress minus0094 0030 0017 0008
ϑcoldstress 0172 0040 0256 0013
ϑprimeintercept 2292 0045 0426 0020
ϑprimehabitat 0316 0036 1210 0018
ϑprimeheatstress minus0006 0038 0054 0013
ϑprimecoldstress minus0622 0064 0875 0019
γintercept minus0884 0353 0023 0162
γhabitatdeviation + minus0292 0478 minus0240 1768
γheatstressdeviation minus minus1136 0607 0064 0194
γcoldstressdeviation minus minus0498 0474 2398 2253
γaveragehabitat 0095 0091 0788 0111
γaverageheatstress minus0024 0067 0004 0062
γaveragecoldstress 0227 0071 0333 0119
εintercept minus3957 0386 minus4471 0141
εhabitatdeviation minus minus0397 0605 0121 1492
εheatstressdeviation + 0643 0455 minus0034 0182
εcoldstressdeviation + 0153 0644 2042 2072
εaveragehabitat 0238 0090 minus1664 0123
εaverageheatstress minus0071 0092 minus0080 0074
εaveragecoldstress minus0307 0141 minus0939 0104
p1intercept minus2172 0043 1351 0039
p1 1998 minus0118 0048 minus0128 0058
p1 1999 minus0112 0049 minus0043 0058
p1 2000 minus0166 0050 minus0017 0059
p1 2001 minus0156 0049 0031 0061
p1 2002 minus0179 0049 0076 0062
p1 2003 minus0216 0051 0160 0064
p12004 minus0242 0051 0066 0061
p1 2005 minus0124 0050 0149 0061
p12006 minus0182 0051 0124 0060
p12007 minus0149 0050 0183 0062
p1 2008 minus0202 0052 0191 0064
p1 2009 minus0221 0052 0134 0062
p1 2010 minus0195 0051 0062 0060
p1 2011 minus0122 0051 0202 0063
(Continues)
1998emsp |emsp emspensp CLEMENT ET aL
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
p1 2012 minus0227 0055 0193 0063
p1stopnumber 0033 0009 minus0055 0011
p1stopnumbersquared minus0069 0010 minus0247 0013
p2intercept minus0921 0018 minus0555 0037
p2 1998 minus0138 0033 minus0121 0050
p2 1999 minus0137 0032 minus0025 0052
p2 2000 minus0164 0032 minus0027 0051
p2 2001 minus0077 0032 minus0059 0053
p2 2002 minus0101 0032 minus0012 0052
p2 2003 minus0199 0033 0085 0053
p22004 minus0186 0032 0113 0059
p2 2005 minus0146 0032 0082 0052
p22006 minus0155 0032 0135 0054
p22007 minus0153 0032 0124 0055
p2 2008 minus0168 0032 0073 0057
p2 2009 minus0077 0033 0100 0056
p2 2010 minus0114 0033 0071 0062
p2 2011 minus0079 0032 0120 0055
p2 2012 minus0186 0033 0030 0063
p2stopnumber 0045 0007 0165 0010
p2stopnumbersquared minus0105 0008 minus0182 0011
π 0533 0064 minus0505 0065
TA B L E 3 emsp (Continued)
F I G U R E 4 emspProbabilityof(a)colonizationand(b)extinctionforRed‐eyedVireointheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1999CLEMENT ET aL
significantanddidnotexplainalargefractionofvariationThereareseveralpotentialexplanationsforwhywedidnotfindasub‐stantialresponsetochangesinhabitatandclimateFirstourdatawerelackingindynamiceventstoinformourmodelsThecovari‐ateschangedrelativelylittleoverthetimeperiodmakingitdiffi‐culttodetectaresponseThelackofchangewaspartlybecauseoftherelativelyshort(16‐year)timeperiodstudiedAlsoweusedamovingaverageforclimatedatawhichtendstoreducedifferences
overshorttimeperiods Inadditionthestudyregionhasexperi‐encedlesswarmingoverthepastcenturythanotherpartsoftheplanet (Walshetal2014)Thebirddistributionsalso lackeddy‐namismBecauseextinctionrateswerelowtherewerefewlocalextinctioneventstoinformthemodelAlthoughcolonizationrateswerehighthefactthatinitialoccupancywashighandextinctionrateswerelowmeanttherewerefewopportunitiesforcoloniza‐tionevents
F I G U R E 5 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionratesforRed‐eyedVireointheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
2000emsp |emsp emspensp CLEMENT ET aL
Second measurement error may have obscured responses toglobalchangeThePRISMclimatedataareestimatedbyinterpola‐tionratherthandirectmeasurementacrossthestudyarea(Dalyetal2008)AssuchtheremaybeartifactsoranomaliesthatimpededourestimationandincreaseduncertaintySimilarlyNLCDdataareremotelysensedandmay includesomemeasurementerrorwhichcan increase uncertainty in our estimates (Homer et al 2015)Additionallylandcoverwasonlymeasuredinthreeyearsrequiringus to imputevalues in13yearsAsa result all landcover changethatoccurredbetween2002and2006wasrecordedin2002whilechangesbetween2007and2011wererecorded in2007Thisap‐proximation isnot idealbutwasnecessarygiventhe5‐yearreso‐lutionofthedataThislowerresolutionofthelandcoverdatamayhavereducedthesensitivityofthemodel
Third wemay havemisspecified ourmodelWe tried to de‐velopbiologicallymeaningfulcovariatesbutwemightnotunder‐standthesystemaswellaswehopedItispossiblethatbirdvitalrates respond to climateon adifferent time scale (ie not a15‐yearmoving average) or to climateduringdifferent timesof theyearortodifferenttemperaturethresholdsortodifferentclimatecomponents (egprecipitation Illaacutenetal2014) It isalsopossi‐ble that thebirds respond to finerhabitat features than the landcovertypesweusedoratadifferentspatialscale(ienot400mPearsonampDawson2003)Alternatively theremaybe importantregionaldifferencesor interactionsamongcovariatesthatwedidnotcaptureOritispossiblethatchangesinclimateandhabitatintheirbreedingareassimplyhavelittleeffectonvitalratesforthesespecies relative to other factors such as competitionwith otherbirdspeciesorglobalchangeoccurringontheirwinteringgroundsinSouthAmerica(Dormann2007)Ofcoursethesemisspecifica‐tionissuesareinherenttoallobservationalstudiesandnotuniquetothisstudy
Itwould be possible for us to explore some of themisspec‐ification issues just mentioned For example we could rerunthe samemodelsusingnewcovariates representing a varietyoftemperature thresholds and habitat buffer sizes to search for astrongercorrelationExperiencesuggeststhatpersistencewouldeventuallyberewardedwithap‐valuelt005ora lowmodelse‐lection valueHowever one reason to avoid such exploration isthat it tends to lead toType Ierrorsoverfitmodels andbiasedparameterestimates(FiebergampJohnson2015)Furthermorewefind thecontrastbetweensignificant correlations for spatial co‐variatesandnonsignificantcorrelationsfortemporalcovariatesil‐luminatingIntypicalclimateenvelopestudieswewouldestimatethe relationshipbetweencovariatesand thecurrentdistributionofaspeciesmuchlikeourestimateofinitialoccupancy(Dormann2007)BecauseweobtainedsignificantparameterestimateswemightconcludethatwehaveidentifiedcovariatesthatdeterminetherangeofthisspeciesThenwemightusetheseresultstoproj‐ectthefuturerangeofthespeciesundervariousclimatescenariosbased on the (questionable) assumption that the covariatendashspe‐cies relationship will not change as climate and habitat change(Cuddington et al 2013 Elith et al 2010Gustafson 2013) In
this case the insignificant correlationwith the temporal covari‐atessuggeststhatouroccupancycovariatesmerelydescribetherangeofthesebirdswithoutdeterminingtherangeFurthermoreour results suggest that despite significant results in the staticportionof themodelwehaveyet to identify themost relevantcomponents of climate change shaping thedistributionof thesebirds although itmay be difficult to identify these componentsgiven the lackofobservedchangeduringa relatively short timeperiodFinallyourresultssuggestthatprojectionsbasedonthestaticportionofourmodelareunlikelytobereliabledespitetheirstatisticalsignificance
Ifourinterpretationiscorrectitsuggeststhatsomeprojectionsof futurerangeshiftsbasedonstaticmodelsrelyonunsupportedassumptionsandarenot reliableWehavedemonstratedadiffer‐entapproachthatdoesnotrequirethestrongassumptionsofstaticmodelingapproachesOurapproachwouldbemoreapttoidentifyfactorsthatguidespeciesrsquoresponsetoglobalchangeifboththespe‐ciesandthecovariatesexhibitedmoredynamismandifweanalyzedalongertimeperiodthatencompassedgreaterchange
ACKNOWLEDG MENTS
WethankKeithPardieckDaveZiolkowskiJrandthethousandsofhighlyskilledvolunteerswhohavecoordinatedandperformedtheNorthAmericanBreedingBirdSurvey for half a centuryWealsothanktheClimateResearchUnitattheUniversityofEastAngliaformakingvaluableclimatedataeasilyaccessibleMJCwasfinanciallysupported by the BBS and the USGS Northeast Climate ScienceCenterThecontentsofthisdocumentaretheresponsibilityoftheauthorsanddonotnecessarilyrepresenttheviewsoftheUSGSAnyuseoftradeproductorfirmnamesisfordescriptivepurposesonlyanddoesnotimplyendorsementbytheUSGovernment
DEDIC ATION
We dedicate this paper to the late Chandler S Robbins ldquoFatherrdquooftheNorthAmericanBreedingBirdSurvey (BBS)DatafromtheBBSformthebasisforthisspecificpaperandforhundredsofothersfocusedon topics ranging from local trends tocontinentalmacro‐ecologyChanwasauniquecombinationofbird‐loverandscientistwhosevisionandinquisitiveenthusiasmwereaninspirationtoallofusfortunateenoughtohaveknownhim
CONFLIC T OF INTERE S T
Nonedeclared
AUTHORSrsquo CONTRIBUTION
JACandJDNconceivedthestudyAJTandSWanalyzedhabitatandclimatedataMJCJEHandJDNperformedtheoccupancyanaly‐sisMJCledthewritingofthemanuscriptAllauthorsgavefinalap‐provalforpublication
emspensp emsp | emsp2001CLEMENT ET aL
DATA ACCE SSIBILIT Y
Raw data are available from the following sources Breeding BirdSurvey httpswwwpwrcusgsgovbbsrawdata PRISM climatedata httpwwwprismoregonstateedu NLCD land cover datahttpswwwmrlcgov Processed data will be archived with theDryadDigitalRepository
ORCID
Matthew J Clement httpsorcidorg0000‐0003‐4231‐7949
James D Nichols httpsorcidorg0000‐0002‐7631‐2890
Adam J Terando httpsorcidorg0000‐0002‐9280‐043X
James E Hines httpsorcidorg0000‐0001‐5478‐7230
Steven G Williams httpsorcidorg0000‐0003‐3760‐6818
R E FE R E N C E S
Anderson D R Burnham K P Gould W R amp Cherry S (2001)Concerns about finding effects that are actually spuriousWildlife Society Bulletin29311ndash316
Arauacutejo M B Cabeza M Thuiller W Hannah L amp WilliamsP H (2004) Would climate change drive species out of re‐serves An assessment of existing reserve‐selection meth‐ods Global Change Biology 10 1618ndash1626 httpsdoiorg101111j1365‐2486200400828x
Barbet‐MassinMampJetzW(2014)A40‐yearcontinent‐widemulti‐speciesassessmentofrelevantclimatepredictorsforspeciesdistri‐butionmodellingDiversity and Distributions201285ndash1295httpsdoiorg101111ddi12229
Barbet‐Massin M Thuiller W amp Jiguet F (2012) The fate ofEuropean breeding birds under climate land‐use and disper‐sal scenarios Global Change Biology 18 881ndash890 httpsdoiorg101111j1365‐2486201102552x
Bellard C Bertelsmeier C Leadley P Thuiller W ampCourchamp F (2012) Impacts of climate change on the fu‐ture of biodiversity Ecology Letters 15 365ndash377 httpsdoiorg101111j1461‐0248201101736x
Boumlhning‐Gaese K amp Bauer H G (1996) Changes in species abun‐dance distribution and diversity in a central European birdcommunity Conservation Biology 10 175ndash187 httpsdoiorg101046j1523‐1739199610010175x
BrommerJELehikoinenAampValkamaJ(2012)Thebreedingrangesof central European and Arctic bird speciesmove poleward PLoS ONE7e43648httpsdoiorg101371journalpone0043648
CalderWAampKingJR(1974)ThermalandcaloricrelationsofbirdsInDSFarnerampJRKing(Eds)Avian biology(Vol4pp259ndash413)NewYorkNYAcademicPress
CesaraccioCSpanoDDucePampSnyderRL(2001)Animprovedmodel for determining degree‐day values from daily temperaturedataInternational Journal of Biometeorology45161ndash169httpsdoiorg101007s004840100104
ChamberlinTC (1890)ThemethodofmultipleworkinghypothesesScience1592ndash96
ClementMJHinesJENicholsJDPardieckKLampZiolkowskiDJJr (2016)Estimating indicesofrangeshifts inbirdsusingdy‐namicmodelswhendetectionisimperfectGlobal Change Biology223273ndash3285httpsdoiorg101111gcb13283
Commission for Environmental Cooperation (1997) Ecological regionsofNorthAmericaTowardacommonperspectiveCommission for
EnvironmentalCooperationMontrealRetrievedfromftpftpepagovwedecoregionscec_naCEC_NAecopdf
Crowl TA Crist TO Parmenter R R BelovskyGamp LugoA E(2008)Thespreadofinvasivespeciesandinfectiousdiseaseasdriv‐ersofecosystemchangeFrontiers in Ecology and the Environment6238ndash246httpsdoiorg101890070151
Cuddington K FortinM J Gerber L RHastings A Liebhold AOConnorMampRayC(2013)Process‐basedmodelsarerequiredtomanageecologicalsystemsinachangingworldEcosphere41ndash12httpsdoiorg101890ES12‐001781
DalyCHalbleibMSmithJIGibsonWPDoggettMKTaylorGHhellipPasterisPP(2008)Physiographicallysensitivemappingofcli‐matologicaltemperatureandprecipitationacrosstheconterminousUnited States International Journal of Climatology 28 2031ndash2064httpsdoiorg101002joc1688
DawsonW (1958) Relation of oxygen consumption and evaporativewater losstotemperatureintheCardinalPhysiological Zoology3137ndash48httpsdoiorg101086physzool31130155377
DawsonWRampWhittowGC (2000)Regulationofbodytempera‐ture InG CWhittow (Ed)Sturkiersquos avian physiology (5th ed pp343ndash390)SanDiegoCAAcademicPress
DeCalestaDS(1994)Effectofwhite‐taileddeeronsongbirdswithinmanagedforestsinPennsylvaniaJournal of Wildlife Management58711ndash718httpsdoiorg1023073809685
DevictorVJulliardRCouvetDampJiguetF(2008)Birdsaretrack‐ing climatewarming but not fast enoughProceedings of the Royal Society of London B Biological Sciences2752743ndash2748httpsdoiorg101098rspb20080878
Dormann C F (2007) Promising the future Global change projec‐tionsofspeciesdistributionsBasic and Applied Ecology8387ndash397httpsdoiorg101016jbaae200611001
Dormann C F Schymanski S J Cabral J Chuine I Graham CHartigFhellipSingerA(2012)Correlationandprocessinspeciesdis‐tributionmodelsBridgingadichotomyJournal of Biogeography392119ndash2131httpsdoiorg101111j1365‐2699201102659x
EccelE(2010)WhatwecanasktohourlytemperaturerecordingPartIIHourlyinterpolationoftemperaturesforclimatologyandmodel‐lingItalian Journal of Agrometeorology1545ndash50
EccelEampCordanoE(2013)InterpolTHourlyinterpolationofmulti‐pletemperaturedailyseriesRpackageversion211RetrievedfromhttpsCRANR‐projectorgpackage=InterpolT
EglingtonSMampPearce‐HigginsJW(2012)Disentanglingtherela‐tiveimportanceofchangesinclimateandland‐useintensityindriv‐ingrecentbirdpopulationtrendsPLoS ONE7e30407httpsdoiorg101371journalpone0030407
Elith JKearneyMampPhillipsS (2010)Theartofmodelling range‐shifting species Methods in Ecology and Evolution 1 330ndash342httpsdoiorg101111j2041‐210X201000036x
FiebergJampJohnsonDH(2015)MMIMultimodelinferenceormod‐elswithmanagement implications Journal of Wildlife Management79708ndash718
FriggensMMampFinchDM (2015) Implicationsof climatechangeforbirdconservationinthesouthwesternUSunderthreealternativefuturesPLoS ONE10e0144089
FryJXianGZJinSDewitzJHomerCGYangLhellipWickhamJD (2011)Completionof the2006national landcoverdatabasefor theconterminousUnitedStatesPhotogrammetric Engineering amp Remote Sensing77858ndash864
Fuller R J Gregory R D Gibbons D W Marchant J HWilson J D Baillie S R amp Carter N (1995) Population de‐clines and range contractions among lowland farmland birdsin Britain Conservation Biology 9 1425ndash1441 httpsdoiorg101046j1523‐1739199509061425x
Gaston K J Blackburn T M Greenwood J J Gregory R DQuinn R M amp Lawton J H (2000) Abundancendashoccupancy
2002emsp |emsp emspensp CLEMENT ET aL
relationships Journal of Applied Ecology 37 39ndash59 httpsdoiorg101046j1365‐2664200000485x
GoldsteinRB(1974)RelationofmetabolismtoambienttemperatureintheverdinCondor76116ndash119httpsdoiorg1023071365995
GrinnellJ(1917)FieldtestsoftheoriesconcerningdistributionalcontrolAmerican Naturalist51115ndash128httpsdoiorg101086279591
GrosboisVGimenezOGaillardJMPradelRBarbraudCClobertJhellipWeimerskirchH(2008)Assessingtheimpactofclimatevari‐ation on survival in vertebrate populationsBiological Reviews 83357ndash399httpsdoiorg101111j1469‐185X200800047x
GustafsonEJ(2013)WhenrelationshipsestimatedinthepastcannotbeusedtopredictthefutureUsingmechanisticmodelstopredictlandscapeecologicaldynamicsinachangingworldLandscape Ecology281429ndash1437httpsdoiorg101007s10980‐013‐9927‐4
Hamel P B (1992) LandManagers guide to the birds of the SouthGeneralTechnicalReportSE‐22USDAForestServiceSoutheasternForestExperimentStationandTheNatureConservancyChapelHillNC
Hinds D S amp Calder W A (1973) Temperature regulation of thePyrrhuloxiaandtheArizonaCardinalPhysiological Zoology4655ndash71httpsdoiorg101086physzool46130152517
HinesJE(2006)PRESENCE Software to estimate patch occupancy and related parameters Laurel MD US Geological survey PatuxentWildlife Research Center Retrieved from httpwwwmbr‐pwrcusgsgovsoftwarepresencehtml
HinesJENicholsJDampCollazoJA(2014)Multiseasonoccupancymodels for correlated replicate surveys Methods in Ecology and Evolution5583ndash591httpsdoiorg1011112041‐210X12186
HinesJENicholsJDRoyleJAMacKenzieDIGopalaswamyAMKumarNampKaranthKU(2010)TigersontrailsOccupancymodeling for cluster sampling Ecological Applications 20 1456ndash1466httpsdoiorg10189009‐03211
HockeyPA SiramiCRidleyARMidgleyGFampBabikerHA(2011) Interrogating recent range changes in South African birdsConfounding signals from land use and climate change present achallenge for attribution Diversity and Distributions 17 254ndash261httpsdoiorg101111j1472‐4642201000741x
Hoegh‐Guldberg O Hughes L McIntyre S Lindenmayer D BParmesanCPossinghamHPampThomasCD (2008)AssistedcolonizationandrapidclimatechangeScience321345ndash346
Homer C Dewitz J Fry J Coan M Hossain N Larson C hellipWickham J (2007) Completion of the 2001 national land coverdatabase for the conterminous United States Photogrammetric Engineering amp Remote Sensing73337
HomerCDewitzJYangLJinSDanielsonPXianGhellipMegownK(2015)Completionofthe2011NationalLandCoverDatabasefortheconterminousUnitedStatesndashrepresentingadecadeoflandcoverchange informationPhotogrammetric Engineering amp Remote Sensing81345ndash354
HootenMBWikleCKDorazioRMampRoyleJA(2007)HierarchicalspatiotemporalmatrixmodelsforcharacterizinginvasionsBiometrics63558ndash567httpsdoiorg101111j1541‐0420200600725x
HosmerDWJrampLemeshowS(2000)Applied logistic regression(2nded)NewYorkNYJohnWileyampSons
Huntley B Collingham Y CGreen R EHiltonGM RahbekCamp Willis S G (2006) Potential impacts of climatic change upongeographical distributions of birds Ibis 148 8ndash28 httpsdoiorg101111j1474‐919X200600523x
IgleciaMNCollazoJAampMcKerrowAJ(2012)Useofoccupancymodels to evaluate expert knowledge‐based species‐habitat rela‐tionshipsAvian Conservation and Ecology7(2)13pRetrievedfromhttpwwwace‐ecoorgvol7iss2art5
IllaacutenJGThomasCDJonesJAWongWKShirleySMampBettsMG(2014)Precipitationandwintertemperaturepredictlong‐termrange‐scale abundancechanges inWesternNorthAmericanbirds
Global Change Biology 20 3351ndash3364 httpsdoiorg101111gcb12642
IoannidisJP (2005)WhymostpublishedresearchfindingsarefalsePLoS Medicine2e124
KeacuteryMGuillera‐ArroitaGampLahoz‐Monfort J J (2013)Analysingand mapping species range dynamics using occupancy modelsJournal of Biogeography 40 1463ndash1474 httpsdoiorg101111jbi12087
Khaliq I Hof C Prinzinger R Boumlhning‐Gaese K amp PfenningerM(2014) Global variation in thermal tolerances and vulnerability ofendotherms to climate change Proceedings of the Royal Society B Biological Sciences 281(1789) 20141097 httpsdoiorg101098rspb20141097
Laursen K amp Frikke J (2008) Hunting from motorboats displacesWadden Sea eiders Somateria mollissima from their favouredfeeding distribution Wildlife Biology 14 423ndash433 httpsdoiorg1029810909‐6396‐144423
LemoineNBauerHGPeintingerMampBoumlhning‐GaeseK (2007)Effectsof climate and land‐use changeon species abundance in acentralEuropeanbirdcommunityConservation Biology21495ndash503httpsdoiorg101111j1523‐1739200600633x
MacArthurRH(1972)Geographical ecology Patterns in the distribution of speciesNewYorkNYHarperampRow
MacKenzieDINicholsJDHinesJEKnutsonMGampFranklinAB(2003)Estimatingsiteoccupancycolonizationandlocalextinc‐tionwhenaspeciesisdetectedimperfectlyEcology842200ndash2207httpsdoiorg10189002‐3090
MacKenzieDINicholsJDLachmanGBDroegeSRoyleJAampLangtimmCA(2002)Estimatingsiteoccupancyrateswhendetec‐tionprobabilitiesarelessthanoneEcology832248ndash2255httpsdoiorg1018900012‐9658(2002)083[2248ESORWD]20CO2
MacKenzieDINicholsJDRoyleJAPollockKHBaileyLampHinesJE(2018)Occupancy estimation and modeling Inferring pat‐terns and dynamics of species occurrence (2nd ed) BurlingtonMAAcademicPress
MellesSJFortinMJLindsayKampBadzinskiD(2011)ExpandingnorthwardInfluenceofclimatechangeforestconnectivityandpop‐ulationprocessesonathreatenedspeciesrangeshiftGlobal Change Biology1717ndash31httpsdoiorg101111j1365‐2486201002214x
Morin X amp Thuiller W (2009) Comparing niche‐and process‐based models to reduce prediction uncertainty in species rangeshifts under climate change Ecology 90 1301ndash1313 httpsdoiorg10189008‐01341
MustinKAmarAampRedpathSM(2014)Colonizationandextinctiondynamicsofadecliningmigratorybirdareinfluencedbyclimateandhabitat degradation Ibis 156 788ndash798 httpsdoiorg101111ibi12173
NicholsJDThomasLampConnPB(2009)InferencesaboutlandbirdabundancefromcountdataRecentadvancesandfuturedirectionsInDLThomsonEJCoochampMJConroy(Eds)Modeling demo‐graphic processes in marked populations(pp201ndash235)NewYorkNYSpringer
OmernikJM(1995)EcoregionsAspatialframeworkforenvironmen‐talmanagement InWSDavisampTSimon(Eds)Biological assess‐ment and criteria Tools for water resource planning and decision making (pp49ndash62)BocaRatonFLLewisPublishers
OmernikJM(2004)Perspectivesonthenatureanddefinitionofeco‐logicalregionsEnvironmental Management34(Supplement1)S27ndashS38httpsdoiorg101007s00267‐003‐5197‐2
Omernik J M amp Griffith G E (2014) Ecoregions of the contermi‐nousUnited States Evolution of a hierarchical spatial frameworkEnvironmental Management541249ndash1266httpsdoiorg101007s00267‐014‐0364‐1
Pardieck K L ZiolkowskiD J JrampHudsonM‐A R (2015)NorthAmericanBreedingBirdSurveyDataset1966ndash2014version20140
emspensp emsp | emsp2003CLEMENT ET aL
USGeologicalSurveyPatuxentWildlifeResearchCenterRetrievedfromhttpswwwpwrcusgsgovbbsrawdata
ParmesanCampYoheG (2003)Agloballycoherentfingerprintofcli‐mate change impacts across natural systemsNature 421 37ndash42httpsdoiorg101038nature01286
PearsonRGampDawsonTP(2003)PredictingtheimpactsofclimatechangeonthedistributionofspeciesArebioclimateenvelopemod‐elsusefulGlobal Ecology and Biogeography12361ndash371
PlattJR(1964)StronginferenceScience146347ndash353PopperK(1959)The logic of scientific discoveryLondonUKHutchinsonRobbins C S Sauer J R Greenberg R S amp Droege S (1989)
Population declines in North American birds that migrate to theNeotropics Proceedings of the National Academy of Sciences 867658ndash7662httpsdoiorg101073pnas86197658
Robinson R A Wilson J D amp Crick H Q (2001) The impor‐tance of arable habitat for farmland birds in grassland land‐scapes Journal of Applied Ecology 38 1059ndash1069 httpsdoiorg101046j1365‐2664200100654x
Romesburg H C (1981) Wildlife science Gaining reliable knowl‐edge Journal of Wildlife Management 45 293ndash313 httpsdoiorg1023073807913
Root T (1988) Energy constraints on avian distributions and abun‐dancesEcology69330ndash339httpsdoiorg1023071940431
Ruiz‐GutieacuterrezVampZipkinEF (2011)Detectionbiasesyieldmis‐leadingpatternsof speciespersistence and colonization in frag‐mented landscapesEcosphere2 1ndash14 httpsdoiorg101890ES10‐002071
SauerJRHinesJEFallonJEPardieckKLZiolkowskiDJJrampLinkWA(2015)The North American breeding bird survey results and analysis 1966ndash2013 Version 01302015LaurelMDUSGSPatuxentWildlifeResearchCenterhttpswwwmbr‐pwrcusgsgovbbsbbshtml
Sauer J R LinkWA Fallon J E Pardieck K L amp ZiolkowskiDJJr(2013)TheNorthAmericanbreedingbirdsurvey1966ndash2011SummaryanalysisandspeciesaccountsNorth American Fauna791ndash32httpsdoiorg103996nafa790001
SeoaneJBustamanteJampDiacuteaz‐DelgadoR(2004)Competingrolesforlandscapevegetationtopographyandclimateinpredictivemod‐elsofbirddistributionEcological Modelling171209ndash222httpsdoiorg101016jecolmodel200308006
Sirami C Caplat P Popy S Clamens A Arlettaz R Jiguet F hellipMartin J‐L (2017) Impacts of global change on species distri‐butions Obstacles and solutions to integrate climate and landuse Global Ecology and Biogeography 26 385ndash394 httpsdoiorg101111geb12555
SkalskiJR(1996)Regressionofabundanceestimatesfrommarkrecap‐ture surveys against environmental covariatesCanadian Journal of
Fisheries and Aquatic Sciences53196ndash204httpsdoiorg101139f95‐169
SohlTL(2014)Therelativeimpactsofclimateandland‐usechangeonconterminousUnitedStatesbird species from2001 to2075PLoS ONE9e112251httpsdoiorg101371journalpone0112251
ThogmartinWESauerJRampKnutsonMG(2007)ModelingandmappingabundanceofAmericanwoodcockacrosstheMidwesternandNortheasternUnitedStatesJournal of Wildlife Management71376ndash382httpsdoiorg1021932005‐680
ThomasCDampLennonJJ (1999)Birdsextendtheirrangesnorth‐wardsNature399213httpsdoiorg10103820335
Trathan P N Garciacutea‐Borboroglu P Boersma D Bost C ACrawford R J CrossinG ThellipWienecke B (2015) Pollutionhabitatlossfishingandclimatechangeascriticalthreatstopen‐guins Conservation Biology 29 31ndash41 httpsdoiorg101111cobi12349
TravisJMJ(2003)Climatechangeandhabitatdestructionadeadlyanthropogenic cocktail Proceedings of the Royal Society of London Series B Biological Sciences 270(1514) 467ndash473 httpsdoiorg101098rspb20022246
UrbanMC (2015)Acceleratingextinction risk fromclimatechangeScience348571ndash573httpsdoiorg101126scienceaaa4984
WalshJWuebblesDHayhoeKKossinJKunkelKStephensGhellipSomervilleR(2014)OurchangingclimateInJMMelilloTCRichmondampGWYohe(Eds)Climate change impacts in the United States The Third National climate assessment(pp19ndash67)WashingtonDCUSGovernmentPrintingOffice
YackulicCBNichols JDReid JampDerR (2015)Topredict thenichemodelcolonizationandextinctionEcology9616ndash23httpsdoiorg10189014‐13611
Yalcin S amp Leroux S J (2018) An empirical test of the relative andcombinedeffectsofland‐coverandclimatechangeonlocalcoloni‐zationandextinctionGlobal Change Biology243849ndash3861httpsdoiorg101111gcb14169
ZhuKWoodallCWampClarkJS(2012)FailuretomigrateLackoftreerangeexpansioninresponsetoclimatechangeGlobal Change Biology181042ndash1052httpsdoiorg101111j1365‐2486201102571x
How to cite this articleClementMJNicholsJDCollazoJATerandoAJHinesJEWilliamsSGPartitioningglobalchangeAssessingtherelativeimportanceofchangesinclimateandlandcoverforchangesinaviandistributionEcol Evol 201991985ndash2003 httpsdoiorg101002ece34890
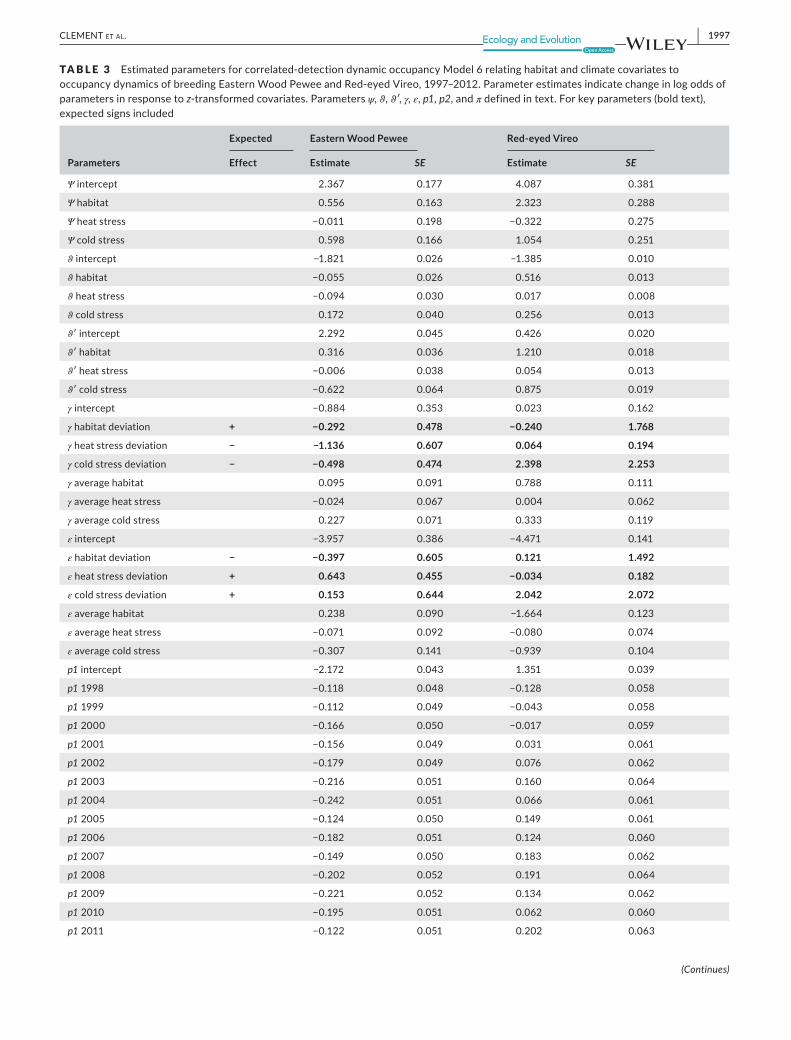
emspensp emsp | emsp1997CLEMENT ET aL
TA B L E 3 emspEstimatedparametersforcorrelated‐detectiondynamicoccupancyModel6relatinghabitatandclimatecovariatestooccupancydynamicsofbreedingEasternWoodPeweeandRed‐eyedVireo1997ndash2012Parameterestimatesindicatechangeinlogoddsofparametersinresponsetoz‐transformedcovariatesParametersψϑϑprimeγε p1p2andπdefinedintextForkeyparameters(boldtext)expectedsignsincluded
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
Ψintercept 2367 0177 4087 0381
Ψhabitat 0556 0163 2323 0288
Ψheatstress minus0011 0198 minus0322 0275
Ψcoldstress 0598 0166 1054 0251
ϑintercept minus1821 0026 minus1385 0010
ϑhabitat minus0055 0026 0516 0013
ϑheatstress minus0094 0030 0017 0008
ϑcoldstress 0172 0040 0256 0013
ϑprimeintercept 2292 0045 0426 0020
ϑprimehabitat 0316 0036 1210 0018
ϑprimeheatstress minus0006 0038 0054 0013
ϑprimecoldstress minus0622 0064 0875 0019
γintercept minus0884 0353 0023 0162
γhabitatdeviation + minus0292 0478 minus0240 1768
γheatstressdeviation minus minus1136 0607 0064 0194
γcoldstressdeviation minus minus0498 0474 2398 2253
γaveragehabitat 0095 0091 0788 0111
γaverageheatstress minus0024 0067 0004 0062
γaveragecoldstress 0227 0071 0333 0119
εintercept minus3957 0386 minus4471 0141
εhabitatdeviation minus minus0397 0605 0121 1492
εheatstressdeviation + 0643 0455 minus0034 0182
εcoldstressdeviation + 0153 0644 2042 2072
εaveragehabitat 0238 0090 minus1664 0123
εaverageheatstress minus0071 0092 minus0080 0074
εaveragecoldstress minus0307 0141 minus0939 0104
p1intercept minus2172 0043 1351 0039
p1 1998 minus0118 0048 minus0128 0058
p1 1999 minus0112 0049 minus0043 0058
p1 2000 minus0166 0050 minus0017 0059
p1 2001 minus0156 0049 0031 0061
p1 2002 minus0179 0049 0076 0062
p1 2003 minus0216 0051 0160 0064
p12004 minus0242 0051 0066 0061
p1 2005 minus0124 0050 0149 0061
p12006 minus0182 0051 0124 0060
p12007 minus0149 0050 0183 0062
p1 2008 minus0202 0052 0191 0064
p1 2009 minus0221 0052 0134 0062
p1 2010 minus0195 0051 0062 0060
p1 2011 minus0122 0051 0202 0063
(Continues)
1998emsp |emsp emspensp CLEMENT ET aL
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
p1 2012 minus0227 0055 0193 0063
p1stopnumber 0033 0009 minus0055 0011
p1stopnumbersquared minus0069 0010 minus0247 0013
p2intercept minus0921 0018 minus0555 0037
p2 1998 minus0138 0033 minus0121 0050
p2 1999 minus0137 0032 minus0025 0052
p2 2000 minus0164 0032 minus0027 0051
p2 2001 minus0077 0032 minus0059 0053
p2 2002 minus0101 0032 minus0012 0052
p2 2003 minus0199 0033 0085 0053
p22004 minus0186 0032 0113 0059
p2 2005 minus0146 0032 0082 0052
p22006 minus0155 0032 0135 0054
p22007 minus0153 0032 0124 0055
p2 2008 minus0168 0032 0073 0057
p2 2009 minus0077 0033 0100 0056
p2 2010 minus0114 0033 0071 0062
p2 2011 minus0079 0032 0120 0055
p2 2012 minus0186 0033 0030 0063
p2stopnumber 0045 0007 0165 0010
p2stopnumbersquared minus0105 0008 minus0182 0011
π 0533 0064 minus0505 0065
TA B L E 3 emsp (Continued)
F I G U R E 4 emspProbabilityof(a)colonizationand(b)extinctionforRed‐eyedVireointheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1999CLEMENT ET aL
significantanddidnotexplainalargefractionofvariationThereareseveralpotentialexplanationsforwhywedidnotfindasub‐stantialresponsetochangesinhabitatandclimateFirstourdatawerelackingindynamiceventstoinformourmodelsThecovari‐ateschangedrelativelylittleoverthetimeperiodmakingitdiffi‐culttodetectaresponseThelackofchangewaspartlybecauseoftherelativelyshort(16‐year)timeperiodstudiedAlsoweusedamovingaverageforclimatedatawhichtendstoreducedifferences
overshorttimeperiods Inadditionthestudyregionhasexperi‐encedlesswarmingoverthepastcenturythanotherpartsoftheplanet (Walshetal2014)Thebirddistributionsalso lackeddy‐namismBecauseextinctionrateswerelowtherewerefewlocalextinctioneventstoinformthemodelAlthoughcolonizationrateswerehighthefactthatinitialoccupancywashighandextinctionrateswerelowmeanttherewerefewopportunitiesforcoloniza‐tionevents
F I G U R E 5 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionratesforRed‐eyedVireointheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
2000emsp |emsp emspensp CLEMENT ET aL
Second measurement error may have obscured responses toglobalchangeThePRISMclimatedataareestimatedbyinterpola‐tionratherthandirectmeasurementacrossthestudyarea(Dalyetal2008)AssuchtheremaybeartifactsoranomaliesthatimpededourestimationandincreaseduncertaintySimilarlyNLCDdataareremotelysensedandmay includesomemeasurementerrorwhichcan increase uncertainty in our estimates (Homer et al 2015)Additionallylandcoverwasonlymeasuredinthreeyearsrequiringus to imputevalues in13yearsAsa result all landcover changethatoccurredbetween2002and2006wasrecordedin2002whilechangesbetween2007and2011wererecorded in2007Thisap‐proximation isnot idealbutwasnecessarygiventhe5‐yearreso‐lutionofthedataThislowerresolutionofthelandcoverdatamayhavereducedthesensitivityofthemodel
Third wemay havemisspecified ourmodelWe tried to de‐velopbiologicallymeaningfulcovariatesbutwemightnotunder‐standthesystemaswellaswehopedItispossiblethatbirdvitalrates respond to climateon adifferent time scale (ie not a15‐yearmoving average) or to climateduringdifferent timesof theyearortodifferenttemperaturethresholdsortodifferentclimatecomponents (egprecipitation Illaacutenetal2014) It isalsopossi‐ble that thebirds respond to finerhabitat features than the landcovertypesweusedoratadifferentspatialscale(ienot400mPearsonampDawson2003)Alternatively theremaybe importantregionaldifferencesor interactionsamongcovariatesthatwedidnotcaptureOritispossiblethatchangesinclimateandhabitatintheirbreedingareassimplyhavelittleeffectonvitalratesforthesespecies relative to other factors such as competitionwith otherbirdspeciesorglobalchangeoccurringontheirwinteringgroundsinSouthAmerica(Dormann2007)Ofcoursethesemisspecifica‐tionissuesareinherenttoallobservationalstudiesandnotuniquetothisstudy
Itwould be possible for us to explore some of themisspec‐ification issues just mentioned For example we could rerunthe samemodelsusingnewcovariates representing a varietyoftemperature thresholds and habitat buffer sizes to search for astrongercorrelationExperiencesuggeststhatpersistencewouldeventuallyberewardedwithap‐valuelt005ora lowmodelse‐lection valueHowever one reason to avoid such exploration isthat it tends to lead toType Ierrorsoverfitmodels andbiasedparameterestimates(FiebergampJohnson2015)Furthermorewefind thecontrastbetweensignificant correlations for spatial co‐variatesandnonsignificantcorrelationsfortemporalcovariatesil‐luminatingIntypicalclimateenvelopestudieswewouldestimatethe relationshipbetweencovariatesand thecurrentdistributionofaspeciesmuchlikeourestimateofinitialoccupancy(Dormann2007)BecauseweobtainedsignificantparameterestimateswemightconcludethatwehaveidentifiedcovariatesthatdeterminetherangeofthisspeciesThenwemightusetheseresultstoproj‐ectthefuturerangeofthespeciesundervariousclimatescenariosbased on the (questionable) assumption that the covariatendashspe‐cies relationship will not change as climate and habitat change(Cuddington et al 2013 Elith et al 2010Gustafson 2013) In
this case the insignificant correlationwith the temporal covari‐atessuggeststhatouroccupancycovariatesmerelydescribetherangeofthesebirdswithoutdeterminingtherangeFurthermoreour results suggest that despite significant results in the staticportionof themodelwehaveyet to identify themost relevantcomponents of climate change shaping thedistributionof thesebirds although itmay be difficult to identify these componentsgiven the lackofobservedchangeduringa relatively short timeperiodFinallyourresultssuggestthatprojectionsbasedonthestaticportionofourmodelareunlikelytobereliabledespitetheirstatisticalsignificance
Ifourinterpretationiscorrectitsuggeststhatsomeprojectionsof futurerangeshiftsbasedonstaticmodelsrelyonunsupportedassumptionsandarenot reliableWehavedemonstratedadiffer‐entapproachthatdoesnotrequirethestrongassumptionsofstaticmodelingapproachesOurapproachwouldbemoreapttoidentifyfactorsthatguidespeciesrsquoresponsetoglobalchangeifboththespe‐ciesandthecovariatesexhibitedmoredynamismandifweanalyzedalongertimeperiodthatencompassedgreaterchange
ACKNOWLEDG MENTS
WethankKeithPardieckDaveZiolkowskiJrandthethousandsofhighlyskilledvolunteerswhohavecoordinatedandperformedtheNorthAmericanBreedingBirdSurvey for half a centuryWealsothanktheClimateResearchUnitattheUniversityofEastAngliaformakingvaluableclimatedataeasilyaccessibleMJCwasfinanciallysupported by the BBS and the USGS Northeast Climate ScienceCenterThecontentsofthisdocumentaretheresponsibilityoftheauthorsanddonotnecessarilyrepresenttheviewsoftheUSGSAnyuseoftradeproductorfirmnamesisfordescriptivepurposesonlyanddoesnotimplyendorsementbytheUSGovernment
DEDIC ATION
We dedicate this paper to the late Chandler S Robbins ldquoFatherrdquooftheNorthAmericanBreedingBirdSurvey (BBS)DatafromtheBBSformthebasisforthisspecificpaperandforhundredsofothersfocusedon topics ranging from local trends tocontinentalmacro‐ecologyChanwasauniquecombinationofbird‐loverandscientistwhosevisionandinquisitiveenthusiasmwereaninspirationtoallofusfortunateenoughtohaveknownhim
CONFLIC T OF INTERE S T
Nonedeclared
AUTHORSrsquo CONTRIBUTION
JACandJDNconceivedthestudyAJTandSWanalyzedhabitatandclimatedataMJCJEHandJDNperformedtheoccupancyanaly‐sisMJCledthewritingofthemanuscriptAllauthorsgavefinalap‐provalforpublication
emspensp emsp | emsp2001CLEMENT ET aL
DATA ACCE SSIBILIT Y
Raw data are available from the following sources Breeding BirdSurvey httpswwwpwrcusgsgovbbsrawdata PRISM climatedata httpwwwprismoregonstateedu NLCD land cover datahttpswwwmrlcgov Processed data will be archived with theDryadDigitalRepository
ORCID
Matthew J Clement httpsorcidorg0000‐0003‐4231‐7949
James D Nichols httpsorcidorg0000‐0002‐7631‐2890
Adam J Terando httpsorcidorg0000‐0002‐9280‐043X
James E Hines httpsorcidorg0000‐0001‐5478‐7230
Steven G Williams httpsorcidorg0000‐0003‐3760‐6818
R E FE R E N C E S
Anderson D R Burnham K P Gould W R amp Cherry S (2001)Concerns about finding effects that are actually spuriousWildlife Society Bulletin29311ndash316
Arauacutejo M B Cabeza M Thuiller W Hannah L amp WilliamsP H (2004) Would climate change drive species out of re‐serves An assessment of existing reserve‐selection meth‐ods Global Change Biology 10 1618ndash1626 httpsdoiorg101111j1365‐2486200400828x
Barbet‐MassinMampJetzW(2014)A40‐yearcontinent‐widemulti‐speciesassessmentofrelevantclimatepredictorsforspeciesdistri‐butionmodellingDiversity and Distributions201285ndash1295httpsdoiorg101111ddi12229
Barbet‐Massin M Thuiller W amp Jiguet F (2012) The fate ofEuropean breeding birds under climate land‐use and disper‐sal scenarios Global Change Biology 18 881ndash890 httpsdoiorg101111j1365‐2486201102552x
Bellard C Bertelsmeier C Leadley P Thuiller W ampCourchamp F (2012) Impacts of climate change on the fu‐ture of biodiversity Ecology Letters 15 365ndash377 httpsdoiorg101111j1461‐0248201101736x
Boumlhning‐Gaese K amp Bauer H G (1996) Changes in species abun‐dance distribution and diversity in a central European birdcommunity Conservation Biology 10 175ndash187 httpsdoiorg101046j1523‐1739199610010175x
BrommerJELehikoinenAampValkamaJ(2012)Thebreedingrangesof central European and Arctic bird speciesmove poleward PLoS ONE7e43648httpsdoiorg101371journalpone0043648
CalderWAampKingJR(1974)ThermalandcaloricrelationsofbirdsInDSFarnerampJRKing(Eds)Avian biology(Vol4pp259ndash413)NewYorkNYAcademicPress
CesaraccioCSpanoDDucePampSnyderRL(2001)Animprovedmodel for determining degree‐day values from daily temperaturedataInternational Journal of Biometeorology45161ndash169httpsdoiorg101007s004840100104
ChamberlinTC (1890)ThemethodofmultipleworkinghypothesesScience1592ndash96
ClementMJHinesJENicholsJDPardieckKLampZiolkowskiDJJr (2016)Estimating indicesofrangeshifts inbirdsusingdy‐namicmodelswhendetectionisimperfectGlobal Change Biology223273ndash3285httpsdoiorg101111gcb13283
Commission for Environmental Cooperation (1997) Ecological regionsofNorthAmericaTowardacommonperspectiveCommission for
EnvironmentalCooperationMontrealRetrievedfromftpftpepagovwedecoregionscec_naCEC_NAecopdf
Crowl TA Crist TO Parmenter R R BelovskyGamp LugoA E(2008)Thespreadofinvasivespeciesandinfectiousdiseaseasdriv‐ersofecosystemchangeFrontiers in Ecology and the Environment6238ndash246httpsdoiorg101890070151
Cuddington K FortinM J Gerber L RHastings A Liebhold AOConnorMampRayC(2013)Process‐basedmodelsarerequiredtomanageecologicalsystemsinachangingworldEcosphere41ndash12httpsdoiorg101890ES12‐001781
DalyCHalbleibMSmithJIGibsonWPDoggettMKTaylorGHhellipPasterisPP(2008)Physiographicallysensitivemappingofcli‐matologicaltemperatureandprecipitationacrosstheconterminousUnited States International Journal of Climatology 28 2031ndash2064httpsdoiorg101002joc1688
DawsonW (1958) Relation of oxygen consumption and evaporativewater losstotemperatureintheCardinalPhysiological Zoology3137ndash48httpsdoiorg101086physzool31130155377
DawsonWRampWhittowGC (2000)Regulationofbodytempera‐ture InG CWhittow (Ed)Sturkiersquos avian physiology (5th ed pp343ndash390)SanDiegoCAAcademicPress
DeCalestaDS(1994)Effectofwhite‐taileddeeronsongbirdswithinmanagedforestsinPennsylvaniaJournal of Wildlife Management58711ndash718httpsdoiorg1023073809685
DevictorVJulliardRCouvetDampJiguetF(2008)Birdsaretrack‐ing climatewarming but not fast enoughProceedings of the Royal Society of London B Biological Sciences2752743ndash2748httpsdoiorg101098rspb20080878
Dormann C F (2007) Promising the future Global change projec‐tionsofspeciesdistributionsBasic and Applied Ecology8387ndash397httpsdoiorg101016jbaae200611001
Dormann C F Schymanski S J Cabral J Chuine I Graham CHartigFhellipSingerA(2012)Correlationandprocessinspeciesdis‐tributionmodelsBridgingadichotomyJournal of Biogeography392119ndash2131httpsdoiorg101111j1365‐2699201102659x
EccelE(2010)WhatwecanasktohourlytemperaturerecordingPartIIHourlyinterpolationoftemperaturesforclimatologyandmodel‐lingItalian Journal of Agrometeorology1545ndash50
EccelEampCordanoE(2013)InterpolTHourlyinterpolationofmulti‐pletemperaturedailyseriesRpackageversion211RetrievedfromhttpsCRANR‐projectorgpackage=InterpolT
EglingtonSMampPearce‐HigginsJW(2012)Disentanglingtherela‐tiveimportanceofchangesinclimateandland‐useintensityindriv‐ingrecentbirdpopulationtrendsPLoS ONE7e30407httpsdoiorg101371journalpone0030407
Elith JKearneyMampPhillipsS (2010)Theartofmodelling range‐shifting species Methods in Ecology and Evolution 1 330ndash342httpsdoiorg101111j2041‐210X201000036x
FiebergJampJohnsonDH(2015)MMIMultimodelinferenceormod‐elswithmanagement implications Journal of Wildlife Management79708ndash718
FriggensMMampFinchDM (2015) Implicationsof climatechangeforbirdconservationinthesouthwesternUSunderthreealternativefuturesPLoS ONE10e0144089
FryJXianGZJinSDewitzJHomerCGYangLhellipWickhamJD (2011)Completionof the2006national landcoverdatabasefor theconterminousUnitedStatesPhotogrammetric Engineering amp Remote Sensing77858ndash864
Fuller R J Gregory R D Gibbons D W Marchant J HWilson J D Baillie S R amp Carter N (1995) Population de‐clines and range contractions among lowland farmland birdsin Britain Conservation Biology 9 1425ndash1441 httpsdoiorg101046j1523‐1739199509061425x
Gaston K J Blackburn T M Greenwood J J Gregory R DQuinn R M amp Lawton J H (2000) Abundancendashoccupancy
2002emsp |emsp emspensp CLEMENT ET aL
relationships Journal of Applied Ecology 37 39ndash59 httpsdoiorg101046j1365‐2664200000485x
GoldsteinRB(1974)RelationofmetabolismtoambienttemperatureintheverdinCondor76116ndash119httpsdoiorg1023071365995
GrinnellJ(1917)FieldtestsoftheoriesconcerningdistributionalcontrolAmerican Naturalist51115ndash128httpsdoiorg101086279591
GrosboisVGimenezOGaillardJMPradelRBarbraudCClobertJhellipWeimerskirchH(2008)Assessingtheimpactofclimatevari‐ation on survival in vertebrate populationsBiological Reviews 83357ndash399httpsdoiorg101111j1469‐185X200800047x
GustafsonEJ(2013)WhenrelationshipsestimatedinthepastcannotbeusedtopredictthefutureUsingmechanisticmodelstopredictlandscapeecologicaldynamicsinachangingworldLandscape Ecology281429ndash1437httpsdoiorg101007s10980‐013‐9927‐4
Hamel P B (1992) LandManagers guide to the birds of the SouthGeneralTechnicalReportSE‐22USDAForestServiceSoutheasternForestExperimentStationandTheNatureConservancyChapelHillNC
Hinds D S amp Calder W A (1973) Temperature regulation of thePyrrhuloxiaandtheArizonaCardinalPhysiological Zoology4655ndash71httpsdoiorg101086physzool46130152517
HinesJE(2006)PRESENCE Software to estimate patch occupancy and related parameters Laurel MD US Geological survey PatuxentWildlife Research Center Retrieved from httpwwwmbr‐pwrcusgsgovsoftwarepresencehtml
HinesJENicholsJDampCollazoJA(2014)Multiseasonoccupancymodels for correlated replicate surveys Methods in Ecology and Evolution5583ndash591httpsdoiorg1011112041‐210X12186
HinesJENicholsJDRoyleJAMacKenzieDIGopalaswamyAMKumarNampKaranthKU(2010)TigersontrailsOccupancymodeling for cluster sampling Ecological Applications 20 1456ndash1466httpsdoiorg10189009‐03211
HockeyPA SiramiCRidleyARMidgleyGFampBabikerHA(2011) Interrogating recent range changes in South African birdsConfounding signals from land use and climate change present achallenge for attribution Diversity and Distributions 17 254ndash261httpsdoiorg101111j1472‐4642201000741x
Hoegh‐Guldberg O Hughes L McIntyre S Lindenmayer D BParmesanCPossinghamHPampThomasCD (2008)AssistedcolonizationandrapidclimatechangeScience321345ndash346
Homer C Dewitz J Fry J Coan M Hossain N Larson C hellipWickham J (2007) Completion of the 2001 national land coverdatabase for the conterminous United States Photogrammetric Engineering amp Remote Sensing73337
HomerCDewitzJYangLJinSDanielsonPXianGhellipMegownK(2015)Completionofthe2011NationalLandCoverDatabasefortheconterminousUnitedStatesndashrepresentingadecadeoflandcoverchange informationPhotogrammetric Engineering amp Remote Sensing81345ndash354
HootenMBWikleCKDorazioRMampRoyleJA(2007)HierarchicalspatiotemporalmatrixmodelsforcharacterizinginvasionsBiometrics63558ndash567httpsdoiorg101111j1541‐0420200600725x
HosmerDWJrampLemeshowS(2000)Applied logistic regression(2nded)NewYorkNYJohnWileyampSons
Huntley B Collingham Y CGreen R EHiltonGM RahbekCamp Willis S G (2006) Potential impacts of climatic change upongeographical distributions of birds Ibis 148 8ndash28 httpsdoiorg101111j1474‐919X200600523x
IgleciaMNCollazoJAampMcKerrowAJ(2012)Useofoccupancymodels to evaluate expert knowledge‐based species‐habitat rela‐tionshipsAvian Conservation and Ecology7(2)13pRetrievedfromhttpwwwace‐ecoorgvol7iss2art5
IllaacutenJGThomasCDJonesJAWongWKShirleySMampBettsMG(2014)Precipitationandwintertemperaturepredictlong‐termrange‐scale abundancechanges inWesternNorthAmericanbirds
Global Change Biology 20 3351ndash3364 httpsdoiorg101111gcb12642
IoannidisJP (2005)WhymostpublishedresearchfindingsarefalsePLoS Medicine2e124
KeacuteryMGuillera‐ArroitaGampLahoz‐Monfort J J (2013)Analysingand mapping species range dynamics using occupancy modelsJournal of Biogeography 40 1463ndash1474 httpsdoiorg101111jbi12087
Khaliq I Hof C Prinzinger R Boumlhning‐Gaese K amp PfenningerM(2014) Global variation in thermal tolerances and vulnerability ofendotherms to climate change Proceedings of the Royal Society B Biological Sciences 281(1789) 20141097 httpsdoiorg101098rspb20141097
Laursen K amp Frikke J (2008) Hunting from motorboats displacesWadden Sea eiders Somateria mollissima from their favouredfeeding distribution Wildlife Biology 14 423ndash433 httpsdoiorg1029810909‐6396‐144423
LemoineNBauerHGPeintingerMampBoumlhning‐GaeseK (2007)Effectsof climate and land‐use changeon species abundance in acentralEuropeanbirdcommunityConservation Biology21495ndash503httpsdoiorg101111j1523‐1739200600633x
MacArthurRH(1972)Geographical ecology Patterns in the distribution of speciesNewYorkNYHarperampRow
MacKenzieDINicholsJDHinesJEKnutsonMGampFranklinAB(2003)Estimatingsiteoccupancycolonizationandlocalextinc‐tionwhenaspeciesisdetectedimperfectlyEcology842200ndash2207httpsdoiorg10189002‐3090
MacKenzieDINicholsJDLachmanGBDroegeSRoyleJAampLangtimmCA(2002)Estimatingsiteoccupancyrateswhendetec‐tionprobabilitiesarelessthanoneEcology832248ndash2255httpsdoiorg1018900012‐9658(2002)083[2248ESORWD]20CO2
MacKenzieDINicholsJDRoyleJAPollockKHBaileyLampHinesJE(2018)Occupancy estimation and modeling Inferring pat‐terns and dynamics of species occurrence (2nd ed) BurlingtonMAAcademicPress
MellesSJFortinMJLindsayKampBadzinskiD(2011)ExpandingnorthwardInfluenceofclimatechangeforestconnectivityandpop‐ulationprocessesonathreatenedspeciesrangeshiftGlobal Change Biology1717ndash31httpsdoiorg101111j1365‐2486201002214x
Morin X amp Thuiller W (2009) Comparing niche‐and process‐based models to reduce prediction uncertainty in species rangeshifts under climate change Ecology 90 1301ndash1313 httpsdoiorg10189008‐01341
MustinKAmarAampRedpathSM(2014)Colonizationandextinctiondynamicsofadecliningmigratorybirdareinfluencedbyclimateandhabitat degradation Ibis 156 788ndash798 httpsdoiorg101111ibi12173
NicholsJDThomasLampConnPB(2009)InferencesaboutlandbirdabundancefromcountdataRecentadvancesandfuturedirectionsInDLThomsonEJCoochampMJConroy(Eds)Modeling demo‐graphic processes in marked populations(pp201ndash235)NewYorkNYSpringer
OmernikJM(1995)EcoregionsAspatialframeworkforenvironmen‐talmanagement InWSDavisampTSimon(Eds)Biological assess‐ment and criteria Tools for water resource planning and decision making (pp49ndash62)BocaRatonFLLewisPublishers
OmernikJM(2004)Perspectivesonthenatureanddefinitionofeco‐logicalregionsEnvironmental Management34(Supplement1)S27ndashS38httpsdoiorg101007s00267‐003‐5197‐2
Omernik J M amp Griffith G E (2014) Ecoregions of the contermi‐nousUnited States Evolution of a hierarchical spatial frameworkEnvironmental Management541249ndash1266httpsdoiorg101007s00267‐014‐0364‐1
Pardieck K L ZiolkowskiD J JrampHudsonM‐A R (2015)NorthAmericanBreedingBirdSurveyDataset1966ndash2014version20140
emspensp emsp | emsp2003CLEMENT ET aL
USGeologicalSurveyPatuxentWildlifeResearchCenterRetrievedfromhttpswwwpwrcusgsgovbbsrawdata
ParmesanCampYoheG (2003)Agloballycoherentfingerprintofcli‐mate change impacts across natural systemsNature 421 37ndash42httpsdoiorg101038nature01286
PearsonRGampDawsonTP(2003)PredictingtheimpactsofclimatechangeonthedistributionofspeciesArebioclimateenvelopemod‐elsusefulGlobal Ecology and Biogeography12361ndash371
PlattJR(1964)StronginferenceScience146347ndash353PopperK(1959)The logic of scientific discoveryLondonUKHutchinsonRobbins C S Sauer J R Greenberg R S amp Droege S (1989)
Population declines in North American birds that migrate to theNeotropics Proceedings of the National Academy of Sciences 867658ndash7662httpsdoiorg101073pnas86197658
Robinson R A Wilson J D amp Crick H Q (2001) The impor‐tance of arable habitat for farmland birds in grassland land‐scapes Journal of Applied Ecology 38 1059ndash1069 httpsdoiorg101046j1365‐2664200100654x
Romesburg H C (1981) Wildlife science Gaining reliable knowl‐edge Journal of Wildlife Management 45 293ndash313 httpsdoiorg1023073807913
Root T (1988) Energy constraints on avian distributions and abun‐dancesEcology69330ndash339httpsdoiorg1023071940431
Ruiz‐GutieacuterrezVampZipkinEF (2011)Detectionbiasesyieldmis‐leadingpatternsof speciespersistence and colonization in frag‐mented landscapesEcosphere2 1ndash14 httpsdoiorg101890ES10‐002071
SauerJRHinesJEFallonJEPardieckKLZiolkowskiDJJrampLinkWA(2015)The North American breeding bird survey results and analysis 1966ndash2013 Version 01302015LaurelMDUSGSPatuxentWildlifeResearchCenterhttpswwwmbr‐pwrcusgsgovbbsbbshtml
Sauer J R LinkWA Fallon J E Pardieck K L amp ZiolkowskiDJJr(2013)TheNorthAmericanbreedingbirdsurvey1966ndash2011SummaryanalysisandspeciesaccountsNorth American Fauna791ndash32httpsdoiorg103996nafa790001
SeoaneJBustamanteJampDiacuteaz‐DelgadoR(2004)Competingrolesforlandscapevegetationtopographyandclimateinpredictivemod‐elsofbirddistributionEcological Modelling171209ndash222httpsdoiorg101016jecolmodel200308006
Sirami C Caplat P Popy S Clamens A Arlettaz R Jiguet F hellipMartin J‐L (2017) Impacts of global change on species distri‐butions Obstacles and solutions to integrate climate and landuse Global Ecology and Biogeography 26 385ndash394 httpsdoiorg101111geb12555
SkalskiJR(1996)Regressionofabundanceestimatesfrommarkrecap‐ture surveys against environmental covariatesCanadian Journal of
Fisheries and Aquatic Sciences53196ndash204httpsdoiorg101139f95‐169
SohlTL(2014)Therelativeimpactsofclimateandland‐usechangeonconterminousUnitedStatesbird species from2001 to2075PLoS ONE9e112251httpsdoiorg101371journalpone0112251
ThogmartinWESauerJRampKnutsonMG(2007)ModelingandmappingabundanceofAmericanwoodcockacrosstheMidwesternandNortheasternUnitedStatesJournal of Wildlife Management71376ndash382httpsdoiorg1021932005‐680
ThomasCDampLennonJJ (1999)Birdsextendtheirrangesnorth‐wardsNature399213httpsdoiorg10103820335
Trathan P N Garciacutea‐Borboroglu P Boersma D Bost C ACrawford R J CrossinG ThellipWienecke B (2015) Pollutionhabitatlossfishingandclimatechangeascriticalthreatstopen‐guins Conservation Biology 29 31ndash41 httpsdoiorg101111cobi12349
TravisJMJ(2003)Climatechangeandhabitatdestructionadeadlyanthropogenic cocktail Proceedings of the Royal Society of London Series B Biological Sciences 270(1514) 467ndash473 httpsdoiorg101098rspb20022246
UrbanMC (2015)Acceleratingextinction risk fromclimatechangeScience348571ndash573httpsdoiorg101126scienceaaa4984
WalshJWuebblesDHayhoeKKossinJKunkelKStephensGhellipSomervilleR(2014)OurchangingclimateInJMMelilloTCRichmondampGWYohe(Eds)Climate change impacts in the United States The Third National climate assessment(pp19ndash67)WashingtonDCUSGovernmentPrintingOffice
YackulicCBNichols JDReid JampDerR (2015)Topredict thenichemodelcolonizationandextinctionEcology9616ndash23httpsdoiorg10189014‐13611
Yalcin S amp Leroux S J (2018) An empirical test of the relative andcombinedeffectsofland‐coverandclimatechangeonlocalcoloni‐zationandextinctionGlobal Change Biology243849ndash3861httpsdoiorg101111gcb14169
ZhuKWoodallCWampClarkJS(2012)FailuretomigrateLackoftreerangeexpansioninresponsetoclimatechangeGlobal Change Biology181042ndash1052httpsdoiorg101111j1365‐2486201102571x
How to cite this articleClementMJNicholsJDCollazoJATerandoAJHinesJEWilliamsSGPartitioningglobalchangeAssessingtherelativeimportanceofchangesinclimateandlandcoverforchangesinaviandistributionEcol Evol 201991985ndash2003 httpsdoiorg101002ece34890

1998emsp |emsp emspensp CLEMENT ET aL
Parameters
Expected Eastern Wood Pewee Red‐eyed Vireo
Effect Estimate SE Estimate SE
p1 2012 minus0227 0055 0193 0063
p1stopnumber 0033 0009 minus0055 0011
p1stopnumbersquared minus0069 0010 minus0247 0013
p2intercept minus0921 0018 minus0555 0037
p2 1998 minus0138 0033 minus0121 0050
p2 1999 minus0137 0032 minus0025 0052
p2 2000 minus0164 0032 minus0027 0051
p2 2001 minus0077 0032 minus0059 0053
p2 2002 minus0101 0032 minus0012 0052
p2 2003 minus0199 0033 0085 0053
p22004 minus0186 0032 0113 0059
p2 2005 minus0146 0032 0082 0052
p22006 minus0155 0032 0135 0054
p22007 minus0153 0032 0124 0055
p2 2008 minus0168 0032 0073 0057
p2 2009 minus0077 0033 0100 0056
p2 2010 minus0114 0033 0071 0062
p2 2011 minus0079 0032 0120 0055
p2 2012 minus0186 0033 0030 0063
p2stopnumber 0045 0007 0165 0010
p2stopnumbersquared minus0105 0008 minus0182 0011
π 0533 0064 minus0505 0065
TA B L E 3 emsp (Continued)
F I G U R E 4 emspProbabilityof(a)colonizationand(b)extinctionforRed‐eyedVireointheeasternUnitedStatesfollowing1997underModel3Colonizationandextinctionvarybytheamountofsuitablehabitathoursofheatstresshoursofcoldstressallaveragedoverthe1997ndash2012studyperiod
emspensp emsp | emsp1999CLEMENT ET aL
significantanddidnotexplainalargefractionofvariationThereareseveralpotentialexplanationsforwhywedidnotfindasub‐stantialresponsetochangesinhabitatandclimateFirstourdatawerelackingindynamiceventstoinformourmodelsThecovari‐ateschangedrelativelylittleoverthetimeperiodmakingitdiffi‐culttodetectaresponseThelackofchangewaspartlybecauseoftherelativelyshort(16‐year)timeperiodstudiedAlsoweusedamovingaverageforclimatedatawhichtendstoreducedifferences
overshorttimeperiods Inadditionthestudyregionhasexperi‐encedlesswarmingoverthepastcenturythanotherpartsoftheplanet (Walshetal2014)Thebirddistributionsalso lackeddy‐namismBecauseextinctionrateswerelowtherewerefewlocalextinctioneventstoinformthemodelAlthoughcolonizationrateswerehighthefactthatinitialoccupancywashighandextinctionrateswerelowmeanttherewerefewopportunitiesforcoloniza‐tionevents
F I G U R E 5 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionratesforRed‐eyedVireointheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
2000emsp |emsp emspensp CLEMENT ET aL
Second measurement error may have obscured responses toglobalchangeThePRISMclimatedataareestimatedbyinterpola‐tionratherthandirectmeasurementacrossthestudyarea(Dalyetal2008)AssuchtheremaybeartifactsoranomaliesthatimpededourestimationandincreaseduncertaintySimilarlyNLCDdataareremotelysensedandmay includesomemeasurementerrorwhichcan increase uncertainty in our estimates (Homer et al 2015)Additionallylandcoverwasonlymeasuredinthreeyearsrequiringus to imputevalues in13yearsAsa result all landcover changethatoccurredbetween2002and2006wasrecordedin2002whilechangesbetween2007and2011wererecorded in2007Thisap‐proximation isnot idealbutwasnecessarygiventhe5‐yearreso‐lutionofthedataThislowerresolutionofthelandcoverdatamayhavereducedthesensitivityofthemodel
Third wemay havemisspecified ourmodelWe tried to de‐velopbiologicallymeaningfulcovariatesbutwemightnotunder‐standthesystemaswellaswehopedItispossiblethatbirdvitalrates respond to climateon adifferent time scale (ie not a15‐yearmoving average) or to climateduringdifferent timesof theyearortodifferenttemperaturethresholdsortodifferentclimatecomponents (egprecipitation Illaacutenetal2014) It isalsopossi‐ble that thebirds respond to finerhabitat features than the landcovertypesweusedoratadifferentspatialscale(ienot400mPearsonampDawson2003)Alternatively theremaybe importantregionaldifferencesor interactionsamongcovariatesthatwedidnotcaptureOritispossiblethatchangesinclimateandhabitatintheirbreedingareassimplyhavelittleeffectonvitalratesforthesespecies relative to other factors such as competitionwith otherbirdspeciesorglobalchangeoccurringontheirwinteringgroundsinSouthAmerica(Dormann2007)Ofcoursethesemisspecifica‐tionissuesareinherenttoallobservationalstudiesandnotuniquetothisstudy
Itwould be possible for us to explore some of themisspec‐ification issues just mentioned For example we could rerunthe samemodelsusingnewcovariates representing a varietyoftemperature thresholds and habitat buffer sizes to search for astrongercorrelationExperiencesuggeststhatpersistencewouldeventuallyberewardedwithap‐valuelt005ora lowmodelse‐lection valueHowever one reason to avoid such exploration isthat it tends to lead toType Ierrorsoverfitmodels andbiasedparameterestimates(FiebergampJohnson2015)Furthermorewefind thecontrastbetweensignificant correlations for spatial co‐variatesandnonsignificantcorrelationsfortemporalcovariatesil‐luminatingIntypicalclimateenvelopestudieswewouldestimatethe relationshipbetweencovariatesand thecurrentdistributionofaspeciesmuchlikeourestimateofinitialoccupancy(Dormann2007)BecauseweobtainedsignificantparameterestimateswemightconcludethatwehaveidentifiedcovariatesthatdeterminetherangeofthisspeciesThenwemightusetheseresultstoproj‐ectthefuturerangeofthespeciesundervariousclimatescenariosbased on the (questionable) assumption that the covariatendashspe‐cies relationship will not change as climate and habitat change(Cuddington et al 2013 Elith et al 2010Gustafson 2013) In
this case the insignificant correlationwith the temporal covari‐atessuggeststhatouroccupancycovariatesmerelydescribetherangeofthesebirdswithoutdeterminingtherangeFurthermoreour results suggest that despite significant results in the staticportionof themodelwehaveyet to identify themost relevantcomponents of climate change shaping thedistributionof thesebirds although itmay be difficult to identify these componentsgiven the lackofobservedchangeduringa relatively short timeperiodFinallyourresultssuggestthatprojectionsbasedonthestaticportionofourmodelareunlikelytobereliabledespitetheirstatisticalsignificance
Ifourinterpretationiscorrectitsuggeststhatsomeprojectionsof futurerangeshiftsbasedonstaticmodelsrelyonunsupportedassumptionsandarenot reliableWehavedemonstratedadiffer‐entapproachthatdoesnotrequirethestrongassumptionsofstaticmodelingapproachesOurapproachwouldbemoreapttoidentifyfactorsthatguidespeciesrsquoresponsetoglobalchangeifboththespe‐ciesandthecovariatesexhibitedmoredynamismandifweanalyzedalongertimeperiodthatencompassedgreaterchange
ACKNOWLEDG MENTS
WethankKeithPardieckDaveZiolkowskiJrandthethousandsofhighlyskilledvolunteerswhohavecoordinatedandperformedtheNorthAmericanBreedingBirdSurvey for half a centuryWealsothanktheClimateResearchUnitattheUniversityofEastAngliaformakingvaluableclimatedataeasilyaccessibleMJCwasfinanciallysupported by the BBS and the USGS Northeast Climate ScienceCenterThecontentsofthisdocumentaretheresponsibilityoftheauthorsanddonotnecessarilyrepresenttheviewsoftheUSGSAnyuseoftradeproductorfirmnamesisfordescriptivepurposesonlyanddoesnotimplyendorsementbytheUSGovernment
DEDIC ATION
We dedicate this paper to the late Chandler S Robbins ldquoFatherrdquooftheNorthAmericanBreedingBirdSurvey (BBS)DatafromtheBBSformthebasisforthisspecificpaperandforhundredsofothersfocusedon topics ranging from local trends tocontinentalmacro‐ecologyChanwasauniquecombinationofbird‐loverandscientistwhosevisionandinquisitiveenthusiasmwereaninspirationtoallofusfortunateenoughtohaveknownhim
CONFLIC T OF INTERE S T
Nonedeclared
AUTHORSrsquo CONTRIBUTION
JACandJDNconceivedthestudyAJTandSWanalyzedhabitatandclimatedataMJCJEHandJDNperformedtheoccupancyanaly‐sisMJCledthewritingofthemanuscriptAllauthorsgavefinalap‐provalforpublication
emspensp emsp | emsp2001CLEMENT ET aL
DATA ACCE SSIBILIT Y
Raw data are available from the following sources Breeding BirdSurvey httpswwwpwrcusgsgovbbsrawdata PRISM climatedata httpwwwprismoregonstateedu NLCD land cover datahttpswwwmrlcgov Processed data will be archived with theDryadDigitalRepository
ORCID
Matthew J Clement httpsorcidorg0000‐0003‐4231‐7949
James D Nichols httpsorcidorg0000‐0002‐7631‐2890
Adam J Terando httpsorcidorg0000‐0002‐9280‐043X
James E Hines httpsorcidorg0000‐0001‐5478‐7230
Steven G Williams httpsorcidorg0000‐0003‐3760‐6818
R E FE R E N C E S
Anderson D R Burnham K P Gould W R amp Cherry S (2001)Concerns about finding effects that are actually spuriousWildlife Society Bulletin29311ndash316
Arauacutejo M B Cabeza M Thuiller W Hannah L amp WilliamsP H (2004) Would climate change drive species out of re‐serves An assessment of existing reserve‐selection meth‐ods Global Change Biology 10 1618ndash1626 httpsdoiorg101111j1365‐2486200400828x
Barbet‐MassinMampJetzW(2014)A40‐yearcontinent‐widemulti‐speciesassessmentofrelevantclimatepredictorsforspeciesdistri‐butionmodellingDiversity and Distributions201285ndash1295httpsdoiorg101111ddi12229
Barbet‐Massin M Thuiller W amp Jiguet F (2012) The fate ofEuropean breeding birds under climate land‐use and disper‐sal scenarios Global Change Biology 18 881ndash890 httpsdoiorg101111j1365‐2486201102552x
Bellard C Bertelsmeier C Leadley P Thuiller W ampCourchamp F (2012) Impacts of climate change on the fu‐ture of biodiversity Ecology Letters 15 365ndash377 httpsdoiorg101111j1461‐0248201101736x
Boumlhning‐Gaese K amp Bauer H G (1996) Changes in species abun‐dance distribution and diversity in a central European birdcommunity Conservation Biology 10 175ndash187 httpsdoiorg101046j1523‐1739199610010175x
BrommerJELehikoinenAampValkamaJ(2012)Thebreedingrangesof central European and Arctic bird speciesmove poleward PLoS ONE7e43648httpsdoiorg101371journalpone0043648
CalderWAampKingJR(1974)ThermalandcaloricrelationsofbirdsInDSFarnerampJRKing(Eds)Avian biology(Vol4pp259ndash413)NewYorkNYAcademicPress
CesaraccioCSpanoDDucePampSnyderRL(2001)Animprovedmodel for determining degree‐day values from daily temperaturedataInternational Journal of Biometeorology45161ndash169httpsdoiorg101007s004840100104
ChamberlinTC (1890)ThemethodofmultipleworkinghypothesesScience1592ndash96
ClementMJHinesJENicholsJDPardieckKLampZiolkowskiDJJr (2016)Estimating indicesofrangeshifts inbirdsusingdy‐namicmodelswhendetectionisimperfectGlobal Change Biology223273ndash3285httpsdoiorg101111gcb13283
Commission for Environmental Cooperation (1997) Ecological regionsofNorthAmericaTowardacommonperspectiveCommission for
EnvironmentalCooperationMontrealRetrievedfromftpftpepagovwedecoregionscec_naCEC_NAecopdf
Crowl TA Crist TO Parmenter R R BelovskyGamp LugoA E(2008)Thespreadofinvasivespeciesandinfectiousdiseaseasdriv‐ersofecosystemchangeFrontiers in Ecology and the Environment6238ndash246httpsdoiorg101890070151
Cuddington K FortinM J Gerber L RHastings A Liebhold AOConnorMampRayC(2013)Process‐basedmodelsarerequiredtomanageecologicalsystemsinachangingworldEcosphere41ndash12httpsdoiorg101890ES12‐001781
DalyCHalbleibMSmithJIGibsonWPDoggettMKTaylorGHhellipPasterisPP(2008)Physiographicallysensitivemappingofcli‐matologicaltemperatureandprecipitationacrosstheconterminousUnited States International Journal of Climatology 28 2031ndash2064httpsdoiorg101002joc1688
DawsonW (1958) Relation of oxygen consumption and evaporativewater losstotemperatureintheCardinalPhysiological Zoology3137ndash48httpsdoiorg101086physzool31130155377
DawsonWRampWhittowGC (2000)Regulationofbodytempera‐ture InG CWhittow (Ed)Sturkiersquos avian physiology (5th ed pp343ndash390)SanDiegoCAAcademicPress
DeCalestaDS(1994)Effectofwhite‐taileddeeronsongbirdswithinmanagedforestsinPennsylvaniaJournal of Wildlife Management58711ndash718httpsdoiorg1023073809685
DevictorVJulliardRCouvetDampJiguetF(2008)Birdsaretrack‐ing climatewarming but not fast enoughProceedings of the Royal Society of London B Biological Sciences2752743ndash2748httpsdoiorg101098rspb20080878
Dormann C F (2007) Promising the future Global change projec‐tionsofspeciesdistributionsBasic and Applied Ecology8387ndash397httpsdoiorg101016jbaae200611001
Dormann C F Schymanski S J Cabral J Chuine I Graham CHartigFhellipSingerA(2012)Correlationandprocessinspeciesdis‐tributionmodelsBridgingadichotomyJournal of Biogeography392119ndash2131httpsdoiorg101111j1365‐2699201102659x
EccelE(2010)WhatwecanasktohourlytemperaturerecordingPartIIHourlyinterpolationoftemperaturesforclimatologyandmodel‐lingItalian Journal of Agrometeorology1545ndash50
EccelEampCordanoE(2013)InterpolTHourlyinterpolationofmulti‐pletemperaturedailyseriesRpackageversion211RetrievedfromhttpsCRANR‐projectorgpackage=InterpolT
EglingtonSMampPearce‐HigginsJW(2012)Disentanglingtherela‐tiveimportanceofchangesinclimateandland‐useintensityindriv‐ingrecentbirdpopulationtrendsPLoS ONE7e30407httpsdoiorg101371journalpone0030407
Elith JKearneyMampPhillipsS (2010)Theartofmodelling range‐shifting species Methods in Ecology and Evolution 1 330ndash342httpsdoiorg101111j2041‐210X201000036x
FiebergJampJohnsonDH(2015)MMIMultimodelinferenceormod‐elswithmanagement implications Journal of Wildlife Management79708ndash718
FriggensMMampFinchDM (2015) Implicationsof climatechangeforbirdconservationinthesouthwesternUSunderthreealternativefuturesPLoS ONE10e0144089
FryJXianGZJinSDewitzJHomerCGYangLhellipWickhamJD (2011)Completionof the2006national landcoverdatabasefor theconterminousUnitedStatesPhotogrammetric Engineering amp Remote Sensing77858ndash864
Fuller R J Gregory R D Gibbons D W Marchant J HWilson J D Baillie S R amp Carter N (1995) Population de‐clines and range contractions among lowland farmland birdsin Britain Conservation Biology 9 1425ndash1441 httpsdoiorg101046j1523‐1739199509061425x
Gaston K J Blackburn T M Greenwood J J Gregory R DQuinn R M amp Lawton J H (2000) Abundancendashoccupancy
2002emsp |emsp emspensp CLEMENT ET aL
relationships Journal of Applied Ecology 37 39ndash59 httpsdoiorg101046j1365‐2664200000485x
GoldsteinRB(1974)RelationofmetabolismtoambienttemperatureintheverdinCondor76116ndash119httpsdoiorg1023071365995
GrinnellJ(1917)FieldtestsoftheoriesconcerningdistributionalcontrolAmerican Naturalist51115ndash128httpsdoiorg101086279591
GrosboisVGimenezOGaillardJMPradelRBarbraudCClobertJhellipWeimerskirchH(2008)Assessingtheimpactofclimatevari‐ation on survival in vertebrate populationsBiological Reviews 83357ndash399httpsdoiorg101111j1469‐185X200800047x
GustafsonEJ(2013)WhenrelationshipsestimatedinthepastcannotbeusedtopredictthefutureUsingmechanisticmodelstopredictlandscapeecologicaldynamicsinachangingworldLandscape Ecology281429ndash1437httpsdoiorg101007s10980‐013‐9927‐4
Hamel P B (1992) LandManagers guide to the birds of the SouthGeneralTechnicalReportSE‐22USDAForestServiceSoutheasternForestExperimentStationandTheNatureConservancyChapelHillNC
Hinds D S amp Calder W A (1973) Temperature regulation of thePyrrhuloxiaandtheArizonaCardinalPhysiological Zoology4655ndash71httpsdoiorg101086physzool46130152517
HinesJE(2006)PRESENCE Software to estimate patch occupancy and related parameters Laurel MD US Geological survey PatuxentWildlife Research Center Retrieved from httpwwwmbr‐pwrcusgsgovsoftwarepresencehtml
HinesJENicholsJDampCollazoJA(2014)Multiseasonoccupancymodels for correlated replicate surveys Methods in Ecology and Evolution5583ndash591httpsdoiorg1011112041‐210X12186
HinesJENicholsJDRoyleJAMacKenzieDIGopalaswamyAMKumarNampKaranthKU(2010)TigersontrailsOccupancymodeling for cluster sampling Ecological Applications 20 1456ndash1466httpsdoiorg10189009‐03211
HockeyPA SiramiCRidleyARMidgleyGFampBabikerHA(2011) Interrogating recent range changes in South African birdsConfounding signals from land use and climate change present achallenge for attribution Diversity and Distributions 17 254ndash261httpsdoiorg101111j1472‐4642201000741x
Hoegh‐Guldberg O Hughes L McIntyre S Lindenmayer D BParmesanCPossinghamHPampThomasCD (2008)AssistedcolonizationandrapidclimatechangeScience321345ndash346
Homer C Dewitz J Fry J Coan M Hossain N Larson C hellipWickham J (2007) Completion of the 2001 national land coverdatabase for the conterminous United States Photogrammetric Engineering amp Remote Sensing73337
HomerCDewitzJYangLJinSDanielsonPXianGhellipMegownK(2015)Completionofthe2011NationalLandCoverDatabasefortheconterminousUnitedStatesndashrepresentingadecadeoflandcoverchange informationPhotogrammetric Engineering amp Remote Sensing81345ndash354
HootenMBWikleCKDorazioRMampRoyleJA(2007)HierarchicalspatiotemporalmatrixmodelsforcharacterizinginvasionsBiometrics63558ndash567httpsdoiorg101111j1541‐0420200600725x
HosmerDWJrampLemeshowS(2000)Applied logistic regression(2nded)NewYorkNYJohnWileyampSons
Huntley B Collingham Y CGreen R EHiltonGM RahbekCamp Willis S G (2006) Potential impacts of climatic change upongeographical distributions of birds Ibis 148 8ndash28 httpsdoiorg101111j1474‐919X200600523x
IgleciaMNCollazoJAampMcKerrowAJ(2012)Useofoccupancymodels to evaluate expert knowledge‐based species‐habitat rela‐tionshipsAvian Conservation and Ecology7(2)13pRetrievedfromhttpwwwace‐ecoorgvol7iss2art5
IllaacutenJGThomasCDJonesJAWongWKShirleySMampBettsMG(2014)Precipitationandwintertemperaturepredictlong‐termrange‐scale abundancechanges inWesternNorthAmericanbirds
Global Change Biology 20 3351ndash3364 httpsdoiorg101111gcb12642
IoannidisJP (2005)WhymostpublishedresearchfindingsarefalsePLoS Medicine2e124
KeacuteryMGuillera‐ArroitaGampLahoz‐Monfort J J (2013)Analysingand mapping species range dynamics using occupancy modelsJournal of Biogeography 40 1463ndash1474 httpsdoiorg101111jbi12087
Khaliq I Hof C Prinzinger R Boumlhning‐Gaese K amp PfenningerM(2014) Global variation in thermal tolerances and vulnerability ofendotherms to climate change Proceedings of the Royal Society B Biological Sciences 281(1789) 20141097 httpsdoiorg101098rspb20141097
Laursen K amp Frikke J (2008) Hunting from motorboats displacesWadden Sea eiders Somateria mollissima from their favouredfeeding distribution Wildlife Biology 14 423ndash433 httpsdoiorg1029810909‐6396‐144423
LemoineNBauerHGPeintingerMampBoumlhning‐GaeseK (2007)Effectsof climate and land‐use changeon species abundance in acentralEuropeanbirdcommunityConservation Biology21495ndash503httpsdoiorg101111j1523‐1739200600633x
MacArthurRH(1972)Geographical ecology Patterns in the distribution of speciesNewYorkNYHarperampRow
MacKenzieDINicholsJDHinesJEKnutsonMGampFranklinAB(2003)Estimatingsiteoccupancycolonizationandlocalextinc‐tionwhenaspeciesisdetectedimperfectlyEcology842200ndash2207httpsdoiorg10189002‐3090
MacKenzieDINicholsJDLachmanGBDroegeSRoyleJAampLangtimmCA(2002)Estimatingsiteoccupancyrateswhendetec‐tionprobabilitiesarelessthanoneEcology832248ndash2255httpsdoiorg1018900012‐9658(2002)083[2248ESORWD]20CO2
MacKenzieDINicholsJDRoyleJAPollockKHBaileyLampHinesJE(2018)Occupancy estimation and modeling Inferring pat‐terns and dynamics of species occurrence (2nd ed) BurlingtonMAAcademicPress
MellesSJFortinMJLindsayKampBadzinskiD(2011)ExpandingnorthwardInfluenceofclimatechangeforestconnectivityandpop‐ulationprocessesonathreatenedspeciesrangeshiftGlobal Change Biology1717ndash31httpsdoiorg101111j1365‐2486201002214x
Morin X amp Thuiller W (2009) Comparing niche‐and process‐based models to reduce prediction uncertainty in species rangeshifts under climate change Ecology 90 1301ndash1313 httpsdoiorg10189008‐01341
MustinKAmarAampRedpathSM(2014)Colonizationandextinctiondynamicsofadecliningmigratorybirdareinfluencedbyclimateandhabitat degradation Ibis 156 788ndash798 httpsdoiorg101111ibi12173
NicholsJDThomasLampConnPB(2009)InferencesaboutlandbirdabundancefromcountdataRecentadvancesandfuturedirectionsInDLThomsonEJCoochampMJConroy(Eds)Modeling demo‐graphic processes in marked populations(pp201ndash235)NewYorkNYSpringer
OmernikJM(1995)EcoregionsAspatialframeworkforenvironmen‐talmanagement InWSDavisampTSimon(Eds)Biological assess‐ment and criteria Tools for water resource planning and decision making (pp49ndash62)BocaRatonFLLewisPublishers
OmernikJM(2004)Perspectivesonthenatureanddefinitionofeco‐logicalregionsEnvironmental Management34(Supplement1)S27ndashS38httpsdoiorg101007s00267‐003‐5197‐2
Omernik J M amp Griffith G E (2014) Ecoregions of the contermi‐nousUnited States Evolution of a hierarchical spatial frameworkEnvironmental Management541249ndash1266httpsdoiorg101007s00267‐014‐0364‐1
Pardieck K L ZiolkowskiD J JrampHudsonM‐A R (2015)NorthAmericanBreedingBirdSurveyDataset1966ndash2014version20140
emspensp emsp | emsp2003CLEMENT ET aL
USGeologicalSurveyPatuxentWildlifeResearchCenterRetrievedfromhttpswwwpwrcusgsgovbbsrawdata
ParmesanCampYoheG (2003)Agloballycoherentfingerprintofcli‐mate change impacts across natural systemsNature 421 37ndash42httpsdoiorg101038nature01286
PearsonRGampDawsonTP(2003)PredictingtheimpactsofclimatechangeonthedistributionofspeciesArebioclimateenvelopemod‐elsusefulGlobal Ecology and Biogeography12361ndash371
PlattJR(1964)StronginferenceScience146347ndash353PopperK(1959)The logic of scientific discoveryLondonUKHutchinsonRobbins C S Sauer J R Greenberg R S amp Droege S (1989)
Population declines in North American birds that migrate to theNeotropics Proceedings of the National Academy of Sciences 867658ndash7662httpsdoiorg101073pnas86197658
Robinson R A Wilson J D amp Crick H Q (2001) The impor‐tance of arable habitat for farmland birds in grassland land‐scapes Journal of Applied Ecology 38 1059ndash1069 httpsdoiorg101046j1365‐2664200100654x
Romesburg H C (1981) Wildlife science Gaining reliable knowl‐edge Journal of Wildlife Management 45 293ndash313 httpsdoiorg1023073807913
Root T (1988) Energy constraints on avian distributions and abun‐dancesEcology69330ndash339httpsdoiorg1023071940431
Ruiz‐GutieacuterrezVampZipkinEF (2011)Detectionbiasesyieldmis‐leadingpatternsof speciespersistence and colonization in frag‐mented landscapesEcosphere2 1ndash14 httpsdoiorg101890ES10‐002071
SauerJRHinesJEFallonJEPardieckKLZiolkowskiDJJrampLinkWA(2015)The North American breeding bird survey results and analysis 1966ndash2013 Version 01302015LaurelMDUSGSPatuxentWildlifeResearchCenterhttpswwwmbr‐pwrcusgsgovbbsbbshtml
Sauer J R LinkWA Fallon J E Pardieck K L amp ZiolkowskiDJJr(2013)TheNorthAmericanbreedingbirdsurvey1966ndash2011SummaryanalysisandspeciesaccountsNorth American Fauna791ndash32httpsdoiorg103996nafa790001
SeoaneJBustamanteJampDiacuteaz‐DelgadoR(2004)Competingrolesforlandscapevegetationtopographyandclimateinpredictivemod‐elsofbirddistributionEcological Modelling171209ndash222httpsdoiorg101016jecolmodel200308006
Sirami C Caplat P Popy S Clamens A Arlettaz R Jiguet F hellipMartin J‐L (2017) Impacts of global change on species distri‐butions Obstacles and solutions to integrate climate and landuse Global Ecology and Biogeography 26 385ndash394 httpsdoiorg101111geb12555
SkalskiJR(1996)Regressionofabundanceestimatesfrommarkrecap‐ture surveys against environmental covariatesCanadian Journal of
Fisheries and Aquatic Sciences53196ndash204httpsdoiorg101139f95‐169
SohlTL(2014)Therelativeimpactsofclimateandland‐usechangeonconterminousUnitedStatesbird species from2001 to2075PLoS ONE9e112251httpsdoiorg101371journalpone0112251
ThogmartinWESauerJRampKnutsonMG(2007)ModelingandmappingabundanceofAmericanwoodcockacrosstheMidwesternandNortheasternUnitedStatesJournal of Wildlife Management71376ndash382httpsdoiorg1021932005‐680
ThomasCDampLennonJJ (1999)Birdsextendtheirrangesnorth‐wardsNature399213httpsdoiorg10103820335
Trathan P N Garciacutea‐Borboroglu P Boersma D Bost C ACrawford R J CrossinG ThellipWienecke B (2015) Pollutionhabitatlossfishingandclimatechangeascriticalthreatstopen‐guins Conservation Biology 29 31ndash41 httpsdoiorg101111cobi12349
TravisJMJ(2003)Climatechangeandhabitatdestructionadeadlyanthropogenic cocktail Proceedings of the Royal Society of London Series B Biological Sciences 270(1514) 467ndash473 httpsdoiorg101098rspb20022246
UrbanMC (2015)Acceleratingextinction risk fromclimatechangeScience348571ndash573httpsdoiorg101126scienceaaa4984
WalshJWuebblesDHayhoeKKossinJKunkelKStephensGhellipSomervilleR(2014)OurchangingclimateInJMMelilloTCRichmondampGWYohe(Eds)Climate change impacts in the United States The Third National climate assessment(pp19ndash67)WashingtonDCUSGovernmentPrintingOffice
YackulicCBNichols JDReid JampDerR (2015)Topredict thenichemodelcolonizationandextinctionEcology9616ndash23httpsdoiorg10189014‐13611
Yalcin S amp Leroux S J (2018) An empirical test of the relative andcombinedeffectsofland‐coverandclimatechangeonlocalcoloni‐zationandextinctionGlobal Change Biology243849ndash3861httpsdoiorg101111gcb14169
ZhuKWoodallCWampClarkJS(2012)FailuretomigrateLackoftreerangeexpansioninresponsetoclimatechangeGlobal Change Biology181042ndash1052httpsdoiorg101111j1365‐2486201102571x
How to cite this articleClementMJNicholsJDCollazoJATerandoAJHinesJEWilliamsSGPartitioningglobalchangeAssessingtherelativeimportanceofchangesinclimateandlandcoverforchangesinaviandistributionEcol Evol 201991985ndash2003 httpsdoiorg101002ece34890

emspensp emsp | emsp1999CLEMENT ET aL
significantanddidnotexplainalargefractionofvariationThereareseveralpotentialexplanationsforwhywedidnotfindasub‐stantialresponsetochangesinhabitatandclimateFirstourdatawerelackingindynamiceventstoinformourmodelsThecovari‐ateschangedrelativelylittleoverthetimeperiodmakingitdiffi‐culttodetectaresponseThelackofchangewaspartlybecauseoftherelativelyshort(16‐year)timeperiodstudiedAlsoweusedamovingaverageforclimatedatawhichtendstoreducedifferences
overshorttimeperiods Inadditionthestudyregionhasexperi‐encedlesswarmingoverthepastcenturythanotherpartsoftheplanet (Walshetal2014)Thebirddistributionsalso lackeddy‐namismBecauseextinctionrateswerelowtherewerefewlocalextinctioneventstoinformthemodelAlthoughcolonizationrateswerehighthefactthatinitialoccupancywashighandextinctionrateswerelowmeanttherewerefewopportunitiesforcoloniza‐tionevents
F I G U R E 5 emspAnnualdeviationofcolonizationandextinctionratesfromaveragecolonizationandextinctionratesforRed‐eyedVireointheeasternUnitedStatesfollowing1997and2011underModel6Deviationsincolonizationandextinctionratesvarywithdeviationsintheamountofsuitablehabitathoursofheatstresshoursofcoldstressrelativetoaveragelevelsoverthe1997ndash2012studyperiod(a)Colonizationdeviationfollowing1997(b)extinctiondeviationfollowing1997(c)colonizationdeviationfollowing2011and(d)extinctiondeviationfollowing2011
2000emsp |emsp emspensp CLEMENT ET aL
Second measurement error may have obscured responses toglobalchangeThePRISMclimatedataareestimatedbyinterpola‐tionratherthandirectmeasurementacrossthestudyarea(Dalyetal2008)AssuchtheremaybeartifactsoranomaliesthatimpededourestimationandincreaseduncertaintySimilarlyNLCDdataareremotelysensedandmay includesomemeasurementerrorwhichcan increase uncertainty in our estimates (Homer et al 2015)Additionallylandcoverwasonlymeasuredinthreeyearsrequiringus to imputevalues in13yearsAsa result all landcover changethatoccurredbetween2002and2006wasrecordedin2002whilechangesbetween2007and2011wererecorded in2007Thisap‐proximation isnot idealbutwasnecessarygiventhe5‐yearreso‐lutionofthedataThislowerresolutionofthelandcoverdatamayhavereducedthesensitivityofthemodel
Third wemay havemisspecified ourmodelWe tried to de‐velopbiologicallymeaningfulcovariatesbutwemightnotunder‐standthesystemaswellaswehopedItispossiblethatbirdvitalrates respond to climateon adifferent time scale (ie not a15‐yearmoving average) or to climateduringdifferent timesof theyearortodifferenttemperaturethresholdsortodifferentclimatecomponents (egprecipitation Illaacutenetal2014) It isalsopossi‐ble that thebirds respond to finerhabitat features than the landcovertypesweusedoratadifferentspatialscale(ienot400mPearsonampDawson2003)Alternatively theremaybe importantregionaldifferencesor interactionsamongcovariatesthatwedidnotcaptureOritispossiblethatchangesinclimateandhabitatintheirbreedingareassimplyhavelittleeffectonvitalratesforthesespecies relative to other factors such as competitionwith otherbirdspeciesorglobalchangeoccurringontheirwinteringgroundsinSouthAmerica(Dormann2007)Ofcoursethesemisspecifica‐tionissuesareinherenttoallobservationalstudiesandnotuniquetothisstudy
Itwould be possible for us to explore some of themisspec‐ification issues just mentioned For example we could rerunthe samemodelsusingnewcovariates representing a varietyoftemperature thresholds and habitat buffer sizes to search for astrongercorrelationExperiencesuggeststhatpersistencewouldeventuallyberewardedwithap‐valuelt005ora lowmodelse‐lection valueHowever one reason to avoid such exploration isthat it tends to lead toType Ierrorsoverfitmodels andbiasedparameterestimates(FiebergampJohnson2015)Furthermorewefind thecontrastbetweensignificant correlations for spatial co‐variatesandnonsignificantcorrelationsfortemporalcovariatesil‐luminatingIntypicalclimateenvelopestudieswewouldestimatethe relationshipbetweencovariatesand thecurrentdistributionofaspeciesmuchlikeourestimateofinitialoccupancy(Dormann2007)BecauseweobtainedsignificantparameterestimateswemightconcludethatwehaveidentifiedcovariatesthatdeterminetherangeofthisspeciesThenwemightusetheseresultstoproj‐ectthefuturerangeofthespeciesundervariousclimatescenariosbased on the (questionable) assumption that the covariatendashspe‐cies relationship will not change as climate and habitat change(Cuddington et al 2013 Elith et al 2010Gustafson 2013) In
this case the insignificant correlationwith the temporal covari‐atessuggeststhatouroccupancycovariatesmerelydescribetherangeofthesebirdswithoutdeterminingtherangeFurthermoreour results suggest that despite significant results in the staticportionof themodelwehaveyet to identify themost relevantcomponents of climate change shaping thedistributionof thesebirds although itmay be difficult to identify these componentsgiven the lackofobservedchangeduringa relatively short timeperiodFinallyourresultssuggestthatprojectionsbasedonthestaticportionofourmodelareunlikelytobereliabledespitetheirstatisticalsignificance
Ifourinterpretationiscorrectitsuggeststhatsomeprojectionsof futurerangeshiftsbasedonstaticmodelsrelyonunsupportedassumptionsandarenot reliableWehavedemonstratedadiffer‐entapproachthatdoesnotrequirethestrongassumptionsofstaticmodelingapproachesOurapproachwouldbemoreapttoidentifyfactorsthatguidespeciesrsquoresponsetoglobalchangeifboththespe‐ciesandthecovariatesexhibitedmoredynamismandifweanalyzedalongertimeperiodthatencompassedgreaterchange
ACKNOWLEDG MENTS
WethankKeithPardieckDaveZiolkowskiJrandthethousandsofhighlyskilledvolunteerswhohavecoordinatedandperformedtheNorthAmericanBreedingBirdSurvey for half a centuryWealsothanktheClimateResearchUnitattheUniversityofEastAngliaformakingvaluableclimatedataeasilyaccessibleMJCwasfinanciallysupported by the BBS and the USGS Northeast Climate ScienceCenterThecontentsofthisdocumentaretheresponsibilityoftheauthorsanddonotnecessarilyrepresenttheviewsoftheUSGSAnyuseoftradeproductorfirmnamesisfordescriptivepurposesonlyanddoesnotimplyendorsementbytheUSGovernment
DEDIC ATION
We dedicate this paper to the late Chandler S Robbins ldquoFatherrdquooftheNorthAmericanBreedingBirdSurvey (BBS)DatafromtheBBSformthebasisforthisspecificpaperandforhundredsofothersfocusedon topics ranging from local trends tocontinentalmacro‐ecologyChanwasauniquecombinationofbird‐loverandscientistwhosevisionandinquisitiveenthusiasmwereaninspirationtoallofusfortunateenoughtohaveknownhim
CONFLIC T OF INTERE S T
Nonedeclared
AUTHORSrsquo CONTRIBUTION
JACandJDNconceivedthestudyAJTandSWanalyzedhabitatandclimatedataMJCJEHandJDNperformedtheoccupancyanaly‐sisMJCledthewritingofthemanuscriptAllauthorsgavefinalap‐provalforpublication
emspensp emsp | emsp2001CLEMENT ET aL
DATA ACCE SSIBILIT Y
Raw data are available from the following sources Breeding BirdSurvey httpswwwpwrcusgsgovbbsrawdata PRISM climatedata httpwwwprismoregonstateedu NLCD land cover datahttpswwwmrlcgov Processed data will be archived with theDryadDigitalRepository
ORCID
Matthew J Clement httpsorcidorg0000‐0003‐4231‐7949
James D Nichols httpsorcidorg0000‐0002‐7631‐2890
Adam J Terando httpsorcidorg0000‐0002‐9280‐043X
James E Hines httpsorcidorg0000‐0001‐5478‐7230
Steven G Williams httpsorcidorg0000‐0003‐3760‐6818
R E FE R E N C E S
Anderson D R Burnham K P Gould W R amp Cherry S (2001)Concerns about finding effects that are actually spuriousWildlife Society Bulletin29311ndash316
Arauacutejo M B Cabeza M Thuiller W Hannah L amp WilliamsP H (2004) Would climate change drive species out of re‐serves An assessment of existing reserve‐selection meth‐ods Global Change Biology 10 1618ndash1626 httpsdoiorg101111j1365‐2486200400828x
Barbet‐MassinMampJetzW(2014)A40‐yearcontinent‐widemulti‐speciesassessmentofrelevantclimatepredictorsforspeciesdistri‐butionmodellingDiversity and Distributions201285ndash1295httpsdoiorg101111ddi12229
Barbet‐Massin M Thuiller W amp Jiguet F (2012) The fate ofEuropean breeding birds under climate land‐use and disper‐sal scenarios Global Change Biology 18 881ndash890 httpsdoiorg101111j1365‐2486201102552x
Bellard C Bertelsmeier C Leadley P Thuiller W ampCourchamp F (2012) Impacts of climate change on the fu‐ture of biodiversity Ecology Letters 15 365ndash377 httpsdoiorg101111j1461‐0248201101736x
Boumlhning‐Gaese K amp Bauer H G (1996) Changes in species abun‐dance distribution and diversity in a central European birdcommunity Conservation Biology 10 175ndash187 httpsdoiorg101046j1523‐1739199610010175x
BrommerJELehikoinenAampValkamaJ(2012)Thebreedingrangesof central European and Arctic bird speciesmove poleward PLoS ONE7e43648httpsdoiorg101371journalpone0043648
CalderWAampKingJR(1974)ThermalandcaloricrelationsofbirdsInDSFarnerampJRKing(Eds)Avian biology(Vol4pp259ndash413)NewYorkNYAcademicPress
CesaraccioCSpanoDDucePampSnyderRL(2001)Animprovedmodel for determining degree‐day values from daily temperaturedataInternational Journal of Biometeorology45161ndash169httpsdoiorg101007s004840100104
ChamberlinTC (1890)ThemethodofmultipleworkinghypothesesScience1592ndash96
ClementMJHinesJENicholsJDPardieckKLampZiolkowskiDJJr (2016)Estimating indicesofrangeshifts inbirdsusingdy‐namicmodelswhendetectionisimperfectGlobal Change Biology223273ndash3285httpsdoiorg101111gcb13283
Commission for Environmental Cooperation (1997) Ecological regionsofNorthAmericaTowardacommonperspectiveCommission for
EnvironmentalCooperationMontrealRetrievedfromftpftpepagovwedecoregionscec_naCEC_NAecopdf
Crowl TA Crist TO Parmenter R R BelovskyGamp LugoA E(2008)Thespreadofinvasivespeciesandinfectiousdiseaseasdriv‐ersofecosystemchangeFrontiers in Ecology and the Environment6238ndash246httpsdoiorg101890070151
Cuddington K FortinM J Gerber L RHastings A Liebhold AOConnorMampRayC(2013)Process‐basedmodelsarerequiredtomanageecologicalsystemsinachangingworldEcosphere41ndash12httpsdoiorg101890ES12‐001781
DalyCHalbleibMSmithJIGibsonWPDoggettMKTaylorGHhellipPasterisPP(2008)Physiographicallysensitivemappingofcli‐matologicaltemperatureandprecipitationacrosstheconterminousUnited States International Journal of Climatology 28 2031ndash2064httpsdoiorg101002joc1688
DawsonW (1958) Relation of oxygen consumption and evaporativewater losstotemperatureintheCardinalPhysiological Zoology3137ndash48httpsdoiorg101086physzool31130155377
DawsonWRampWhittowGC (2000)Regulationofbodytempera‐ture InG CWhittow (Ed)Sturkiersquos avian physiology (5th ed pp343ndash390)SanDiegoCAAcademicPress
DeCalestaDS(1994)Effectofwhite‐taileddeeronsongbirdswithinmanagedforestsinPennsylvaniaJournal of Wildlife Management58711ndash718httpsdoiorg1023073809685
DevictorVJulliardRCouvetDampJiguetF(2008)Birdsaretrack‐ing climatewarming but not fast enoughProceedings of the Royal Society of London B Biological Sciences2752743ndash2748httpsdoiorg101098rspb20080878
Dormann C F (2007) Promising the future Global change projec‐tionsofspeciesdistributionsBasic and Applied Ecology8387ndash397httpsdoiorg101016jbaae200611001
Dormann C F Schymanski S J Cabral J Chuine I Graham CHartigFhellipSingerA(2012)Correlationandprocessinspeciesdis‐tributionmodelsBridgingadichotomyJournal of Biogeography392119ndash2131httpsdoiorg101111j1365‐2699201102659x
EccelE(2010)WhatwecanasktohourlytemperaturerecordingPartIIHourlyinterpolationoftemperaturesforclimatologyandmodel‐lingItalian Journal of Agrometeorology1545ndash50
EccelEampCordanoE(2013)InterpolTHourlyinterpolationofmulti‐pletemperaturedailyseriesRpackageversion211RetrievedfromhttpsCRANR‐projectorgpackage=InterpolT
EglingtonSMampPearce‐HigginsJW(2012)Disentanglingtherela‐tiveimportanceofchangesinclimateandland‐useintensityindriv‐ingrecentbirdpopulationtrendsPLoS ONE7e30407httpsdoiorg101371journalpone0030407
Elith JKearneyMampPhillipsS (2010)Theartofmodelling range‐shifting species Methods in Ecology and Evolution 1 330ndash342httpsdoiorg101111j2041‐210X201000036x
FiebergJampJohnsonDH(2015)MMIMultimodelinferenceormod‐elswithmanagement implications Journal of Wildlife Management79708ndash718
FriggensMMampFinchDM (2015) Implicationsof climatechangeforbirdconservationinthesouthwesternUSunderthreealternativefuturesPLoS ONE10e0144089
FryJXianGZJinSDewitzJHomerCGYangLhellipWickhamJD (2011)Completionof the2006national landcoverdatabasefor theconterminousUnitedStatesPhotogrammetric Engineering amp Remote Sensing77858ndash864
Fuller R J Gregory R D Gibbons D W Marchant J HWilson J D Baillie S R amp Carter N (1995) Population de‐clines and range contractions among lowland farmland birdsin Britain Conservation Biology 9 1425ndash1441 httpsdoiorg101046j1523‐1739199509061425x
Gaston K J Blackburn T M Greenwood J J Gregory R DQuinn R M amp Lawton J H (2000) Abundancendashoccupancy
2002emsp |emsp emspensp CLEMENT ET aL
relationships Journal of Applied Ecology 37 39ndash59 httpsdoiorg101046j1365‐2664200000485x
GoldsteinRB(1974)RelationofmetabolismtoambienttemperatureintheverdinCondor76116ndash119httpsdoiorg1023071365995
GrinnellJ(1917)FieldtestsoftheoriesconcerningdistributionalcontrolAmerican Naturalist51115ndash128httpsdoiorg101086279591
GrosboisVGimenezOGaillardJMPradelRBarbraudCClobertJhellipWeimerskirchH(2008)Assessingtheimpactofclimatevari‐ation on survival in vertebrate populationsBiological Reviews 83357ndash399httpsdoiorg101111j1469‐185X200800047x
GustafsonEJ(2013)WhenrelationshipsestimatedinthepastcannotbeusedtopredictthefutureUsingmechanisticmodelstopredictlandscapeecologicaldynamicsinachangingworldLandscape Ecology281429ndash1437httpsdoiorg101007s10980‐013‐9927‐4
Hamel P B (1992) LandManagers guide to the birds of the SouthGeneralTechnicalReportSE‐22USDAForestServiceSoutheasternForestExperimentStationandTheNatureConservancyChapelHillNC
Hinds D S amp Calder W A (1973) Temperature regulation of thePyrrhuloxiaandtheArizonaCardinalPhysiological Zoology4655ndash71httpsdoiorg101086physzool46130152517
HinesJE(2006)PRESENCE Software to estimate patch occupancy and related parameters Laurel MD US Geological survey PatuxentWildlife Research Center Retrieved from httpwwwmbr‐pwrcusgsgovsoftwarepresencehtml
HinesJENicholsJDampCollazoJA(2014)Multiseasonoccupancymodels for correlated replicate surveys Methods in Ecology and Evolution5583ndash591httpsdoiorg1011112041‐210X12186
HinesJENicholsJDRoyleJAMacKenzieDIGopalaswamyAMKumarNampKaranthKU(2010)TigersontrailsOccupancymodeling for cluster sampling Ecological Applications 20 1456ndash1466httpsdoiorg10189009‐03211
HockeyPA SiramiCRidleyARMidgleyGFampBabikerHA(2011) Interrogating recent range changes in South African birdsConfounding signals from land use and climate change present achallenge for attribution Diversity and Distributions 17 254ndash261httpsdoiorg101111j1472‐4642201000741x
Hoegh‐Guldberg O Hughes L McIntyre S Lindenmayer D BParmesanCPossinghamHPampThomasCD (2008)AssistedcolonizationandrapidclimatechangeScience321345ndash346
Homer C Dewitz J Fry J Coan M Hossain N Larson C hellipWickham J (2007) Completion of the 2001 national land coverdatabase for the conterminous United States Photogrammetric Engineering amp Remote Sensing73337
HomerCDewitzJYangLJinSDanielsonPXianGhellipMegownK(2015)Completionofthe2011NationalLandCoverDatabasefortheconterminousUnitedStatesndashrepresentingadecadeoflandcoverchange informationPhotogrammetric Engineering amp Remote Sensing81345ndash354
HootenMBWikleCKDorazioRMampRoyleJA(2007)HierarchicalspatiotemporalmatrixmodelsforcharacterizinginvasionsBiometrics63558ndash567httpsdoiorg101111j1541‐0420200600725x
HosmerDWJrampLemeshowS(2000)Applied logistic regression(2nded)NewYorkNYJohnWileyampSons
Huntley B Collingham Y CGreen R EHiltonGM RahbekCamp Willis S G (2006) Potential impacts of climatic change upongeographical distributions of birds Ibis 148 8ndash28 httpsdoiorg101111j1474‐919X200600523x
IgleciaMNCollazoJAampMcKerrowAJ(2012)Useofoccupancymodels to evaluate expert knowledge‐based species‐habitat rela‐tionshipsAvian Conservation and Ecology7(2)13pRetrievedfromhttpwwwace‐ecoorgvol7iss2art5
IllaacutenJGThomasCDJonesJAWongWKShirleySMampBettsMG(2014)Precipitationandwintertemperaturepredictlong‐termrange‐scale abundancechanges inWesternNorthAmericanbirds
Global Change Biology 20 3351ndash3364 httpsdoiorg101111gcb12642
IoannidisJP (2005)WhymostpublishedresearchfindingsarefalsePLoS Medicine2e124
KeacuteryMGuillera‐ArroitaGampLahoz‐Monfort J J (2013)Analysingand mapping species range dynamics using occupancy modelsJournal of Biogeography 40 1463ndash1474 httpsdoiorg101111jbi12087
Khaliq I Hof C Prinzinger R Boumlhning‐Gaese K amp PfenningerM(2014) Global variation in thermal tolerances and vulnerability ofendotherms to climate change Proceedings of the Royal Society B Biological Sciences 281(1789) 20141097 httpsdoiorg101098rspb20141097
Laursen K amp Frikke J (2008) Hunting from motorboats displacesWadden Sea eiders Somateria mollissima from their favouredfeeding distribution Wildlife Biology 14 423ndash433 httpsdoiorg1029810909‐6396‐144423
LemoineNBauerHGPeintingerMampBoumlhning‐GaeseK (2007)Effectsof climate and land‐use changeon species abundance in acentralEuropeanbirdcommunityConservation Biology21495ndash503httpsdoiorg101111j1523‐1739200600633x
MacArthurRH(1972)Geographical ecology Patterns in the distribution of speciesNewYorkNYHarperampRow
MacKenzieDINicholsJDHinesJEKnutsonMGampFranklinAB(2003)Estimatingsiteoccupancycolonizationandlocalextinc‐tionwhenaspeciesisdetectedimperfectlyEcology842200ndash2207httpsdoiorg10189002‐3090
MacKenzieDINicholsJDLachmanGBDroegeSRoyleJAampLangtimmCA(2002)Estimatingsiteoccupancyrateswhendetec‐tionprobabilitiesarelessthanoneEcology832248ndash2255httpsdoiorg1018900012‐9658(2002)083[2248ESORWD]20CO2
MacKenzieDINicholsJDRoyleJAPollockKHBaileyLampHinesJE(2018)Occupancy estimation and modeling Inferring pat‐terns and dynamics of species occurrence (2nd ed) BurlingtonMAAcademicPress
MellesSJFortinMJLindsayKampBadzinskiD(2011)ExpandingnorthwardInfluenceofclimatechangeforestconnectivityandpop‐ulationprocessesonathreatenedspeciesrangeshiftGlobal Change Biology1717ndash31httpsdoiorg101111j1365‐2486201002214x
Morin X amp Thuiller W (2009) Comparing niche‐and process‐based models to reduce prediction uncertainty in species rangeshifts under climate change Ecology 90 1301ndash1313 httpsdoiorg10189008‐01341
MustinKAmarAampRedpathSM(2014)Colonizationandextinctiondynamicsofadecliningmigratorybirdareinfluencedbyclimateandhabitat degradation Ibis 156 788ndash798 httpsdoiorg101111ibi12173
NicholsJDThomasLampConnPB(2009)InferencesaboutlandbirdabundancefromcountdataRecentadvancesandfuturedirectionsInDLThomsonEJCoochampMJConroy(Eds)Modeling demo‐graphic processes in marked populations(pp201ndash235)NewYorkNYSpringer
OmernikJM(1995)EcoregionsAspatialframeworkforenvironmen‐talmanagement InWSDavisampTSimon(Eds)Biological assess‐ment and criteria Tools for water resource planning and decision making (pp49ndash62)BocaRatonFLLewisPublishers
OmernikJM(2004)Perspectivesonthenatureanddefinitionofeco‐logicalregionsEnvironmental Management34(Supplement1)S27ndashS38httpsdoiorg101007s00267‐003‐5197‐2
Omernik J M amp Griffith G E (2014) Ecoregions of the contermi‐nousUnited States Evolution of a hierarchical spatial frameworkEnvironmental Management541249ndash1266httpsdoiorg101007s00267‐014‐0364‐1
Pardieck K L ZiolkowskiD J JrampHudsonM‐A R (2015)NorthAmericanBreedingBirdSurveyDataset1966ndash2014version20140
emspensp emsp | emsp2003CLEMENT ET aL
USGeologicalSurveyPatuxentWildlifeResearchCenterRetrievedfromhttpswwwpwrcusgsgovbbsrawdata
ParmesanCampYoheG (2003)Agloballycoherentfingerprintofcli‐mate change impacts across natural systemsNature 421 37ndash42httpsdoiorg101038nature01286
PearsonRGampDawsonTP(2003)PredictingtheimpactsofclimatechangeonthedistributionofspeciesArebioclimateenvelopemod‐elsusefulGlobal Ecology and Biogeography12361ndash371
PlattJR(1964)StronginferenceScience146347ndash353PopperK(1959)The logic of scientific discoveryLondonUKHutchinsonRobbins C S Sauer J R Greenberg R S amp Droege S (1989)
Population declines in North American birds that migrate to theNeotropics Proceedings of the National Academy of Sciences 867658ndash7662httpsdoiorg101073pnas86197658
Robinson R A Wilson J D amp Crick H Q (2001) The impor‐tance of arable habitat for farmland birds in grassland land‐scapes Journal of Applied Ecology 38 1059ndash1069 httpsdoiorg101046j1365‐2664200100654x
Romesburg H C (1981) Wildlife science Gaining reliable knowl‐edge Journal of Wildlife Management 45 293ndash313 httpsdoiorg1023073807913
Root T (1988) Energy constraints on avian distributions and abun‐dancesEcology69330ndash339httpsdoiorg1023071940431
Ruiz‐GutieacuterrezVampZipkinEF (2011)Detectionbiasesyieldmis‐leadingpatternsof speciespersistence and colonization in frag‐mented landscapesEcosphere2 1ndash14 httpsdoiorg101890ES10‐002071
SauerJRHinesJEFallonJEPardieckKLZiolkowskiDJJrampLinkWA(2015)The North American breeding bird survey results and analysis 1966ndash2013 Version 01302015LaurelMDUSGSPatuxentWildlifeResearchCenterhttpswwwmbr‐pwrcusgsgovbbsbbshtml
Sauer J R LinkWA Fallon J E Pardieck K L amp ZiolkowskiDJJr(2013)TheNorthAmericanbreedingbirdsurvey1966ndash2011SummaryanalysisandspeciesaccountsNorth American Fauna791ndash32httpsdoiorg103996nafa790001
SeoaneJBustamanteJampDiacuteaz‐DelgadoR(2004)Competingrolesforlandscapevegetationtopographyandclimateinpredictivemod‐elsofbirddistributionEcological Modelling171209ndash222httpsdoiorg101016jecolmodel200308006
Sirami C Caplat P Popy S Clamens A Arlettaz R Jiguet F hellipMartin J‐L (2017) Impacts of global change on species distri‐butions Obstacles and solutions to integrate climate and landuse Global Ecology and Biogeography 26 385ndash394 httpsdoiorg101111geb12555
SkalskiJR(1996)Regressionofabundanceestimatesfrommarkrecap‐ture surveys against environmental covariatesCanadian Journal of
Fisheries and Aquatic Sciences53196ndash204httpsdoiorg101139f95‐169
SohlTL(2014)Therelativeimpactsofclimateandland‐usechangeonconterminousUnitedStatesbird species from2001 to2075PLoS ONE9e112251httpsdoiorg101371journalpone0112251
ThogmartinWESauerJRampKnutsonMG(2007)ModelingandmappingabundanceofAmericanwoodcockacrosstheMidwesternandNortheasternUnitedStatesJournal of Wildlife Management71376ndash382httpsdoiorg1021932005‐680
ThomasCDampLennonJJ (1999)Birdsextendtheirrangesnorth‐wardsNature399213httpsdoiorg10103820335
Trathan P N Garciacutea‐Borboroglu P Boersma D Bost C ACrawford R J CrossinG ThellipWienecke B (2015) Pollutionhabitatlossfishingandclimatechangeascriticalthreatstopen‐guins Conservation Biology 29 31ndash41 httpsdoiorg101111cobi12349
TravisJMJ(2003)Climatechangeandhabitatdestructionadeadlyanthropogenic cocktail Proceedings of the Royal Society of London Series B Biological Sciences 270(1514) 467ndash473 httpsdoiorg101098rspb20022246
UrbanMC (2015)Acceleratingextinction risk fromclimatechangeScience348571ndash573httpsdoiorg101126scienceaaa4984
WalshJWuebblesDHayhoeKKossinJKunkelKStephensGhellipSomervilleR(2014)OurchangingclimateInJMMelilloTCRichmondampGWYohe(Eds)Climate change impacts in the United States The Third National climate assessment(pp19ndash67)WashingtonDCUSGovernmentPrintingOffice
YackulicCBNichols JDReid JampDerR (2015)Topredict thenichemodelcolonizationandextinctionEcology9616ndash23httpsdoiorg10189014‐13611
Yalcin S amp Leroux S J (2018) An empirical test of the relative andcombinedeffectsofland‐coverandclimatechangeonlocalcoloni‐zationandextinctionGlobal Change Biology243849ndash3861httpsdoiorg101111gcb14169
ZhuKWoodallCWampClarkJS(2012)FailuretomigrateLackoftreerangeexpansioninresponsetoclimatechangeGlobal Change Biology181042ndash1052httpsdoiorg101111j1365‐2486201102571x
How to cite this articleClementMJNicholsJDCollazoJATerandoAJHinesJEWilliamsSGPartitioningglobalchangeAssessingtherelativeimportanceofchangesinclimateandlandcoverforchangesinaviandistributionEcol Evol 201991985ndash2003 httpsdoiorg101002ece34890

2000emsp |emsp emspensp CLEMENT ET aL
Second measurement error may have obscured responses toglobalchangeThePRISMclimatedataareestimatedbyinterpola‐tionratherthandirectmeasurementacrossthestudyarea(Dalyetal2008)AssuchtheremaybeartifactsoranomaliesthatimpededourestimationandincreaseduncertaintySimilarlyNLCDdataareremotelysensedandmay includesomemeasurementerrorwhichcan increase uncertainty in our estimates (Homer et al 2015)Additionallylandcoverwasonlymeasuredinthreeyearsrequiringus to imputevalues in13yearsAsa result all landcover changethatoccurredbetween2002and2006wasrecordedin2002whilechangesbetween2007and2011wererecorded in2007Thisap‐proximation isnot idealbutwasnecessarygiventhe5‐yearreso‐lutionofthedataThislowerresolutionofthelandcoverdatamayhavereducedthesensitivityofthemodel
Third wemay havemisspecified ourmodelWe tried to de‐velopbiologicallymeaningfulcovariatesbutwemightnotunder‐standthesystemaswellaswehopedItispossiblethatbirdvitalrates respond to climateon adifferent time scale (ie not a15‐yearmoving average) or to climateduringdifferent timesof theyearortodifferenttemperaturethresholdsortodifferentclimatecomponents (egprecipitation Illaacutenetal2014) It isalsopossi‐ble that thebirds respond to finerhabitat features than the landcovertypesweusedoratadifferentspatialscale(ienot400mPearsonampDawson2003)Alternatively theremaybe importantregionaldifferencesor interactionsamongcovariatesthatwedidnotcaptureOritispossiblethatchangesinclimateandhabitatintheirbreedingareassimplyhavelittleeffectonvitalratesforthesespecies relative to other factors such as competitionwith otherbirdspeciesorglobalchangeoccurringontheirwinteringgroundsinSouthAmerica(Dormann2007)Ofcoursethesemisspecifica‐tionissuesareinherenttoallobservationalstudiesandnotuniquetothisstudy
Itwould be possible for us to explore some of themisspec‐ification issues just mentioned For example we could rerunthe samemodelsusingnewcovariates representing a varietyoftemperature thresholds and habitat buffer sizes to search for astrongercorrelationExperiencesuggeststhatpersistencewouldeventuallyberewardedwithap‐valuelt005ora lowmodelse‐lection valueHowever one reason to avoid such exploration isthat it tends to lead toType Ierrorsoverfitmodels andbiasedparameterestimates(FiebergampJohnson2015)Furthermorewefind thecontrastbetweensignificant correlations for spatial co‐variatesandnonsignificantcorrelationsfortemporalcovariatesil‐luminatingIntypicalclimateenvelopestudieswewouldestimatethe relationshipbetweencovariatesand thecurrentdistributionofaspeciesmuchlikeourestimateofinitialoccupancy(Dormann2007)BecauseweobtainedsignificantparameterestimateswemightconcludethatwehaveidentifiedcovariatesthatdeterminetherangeofthisspeciesThenwemightusetheseresultstoproj‐ectthefuturerangeofthespeciesundervariousclimatescenariosbased on the (questionable) assumption that the covariatendashspe‐cies relationship will not change as climate and habitat change(Cuddington et al 2013 Elith et al 2010Gustafson 2013) In
this case the insignificant correlationwith the temporal covari‐atessuggeststhatouroccupancycovariatesmerelydescribetherangeofthesebirdswithoutdeterminingtherangeFurthermoreour results suggest that despite significant results in the staticportionof themodelwehaveyet to identify themost relevantcomponents of climate change shaping thedistributionof thesebirds although itmay be difficult to identify these componentsgiven the lackofobservedchangeduringa relatively short timeperiodFinallyourresultssuggestthatprojectionsbasedonthestaticportionofourmodelareunlikelytobereliabledespitetheirstatisticalsignificance
Ifourinterpretationiscorrectitsuggeststhatsomeprojectionsof futurerangeshiftsbasedonstaticmodelsrelyonunsupportedassumptionsandarenot reliableWehavedemonstratedadiffer‐entapproachthatdoesnotrequirethestrongassumptionsofstaticmodelingapproachesOurapproachwouldbemoreapttoidentifyfactorsthatguidespeciesrsquoresponsetoglobalchangeifboththespe‐ciesandthecovariatesexhibitedmoredynamismandifweanalyzedalongertimeperiodthatencompassedgreaterchange
ACKNOWLEDG MENTS
WethankKeithPardieckDaveZiolkowskiJrandthethousandsofhighlyskilledvolunteerswhohavecoordinatedandperformedtheNorthAmericanBreedingBirdSurvey for half a centuryWealsothanktheClimateResearchUnitattheUniversityofEastAngliaformakingvaluableclimatedataeasilyaccessibleMJCwasfinanciallysupported by the BBS and the USGS Northeast Climate ScienceCenterThecontentsofthisdocumentaretheresponsibilityoftheauthorsanddonotnecessarilyrepresenttheviewsoftheUSGSAnyuseoftradeproductorfirmnamesisfordescriptivepurposesonlyanddoesnotimplyendorsementbytheUSGovernment
DEDIC ATION
We dedicate this paper to the late Chandler S Robbins ldquoFatherrdquooftheNorthAmericanBreedingBirdSurvey (BBS)DatafromtheBBSformthebasisforthisspecificpaperandforhundredsofothersfocusedon topics ranging from local trends tocontinentalmacro‐ecologyChanwasauniquecombinationofbird‐loverandscientistwhosevisionandinquisitiveenthusiasmwereaninspirationtoallofusfortunateenoughtohaveknownhim
CONFLIC T OF INTERE S T
Nonedeclared
AUTHORSrsquo CONTRIBUTION
JACandJDNconceivedthestudyAJTandSWanalyzedhabitatandclimatedataMJCJEHandJDNperformedtheoccupancyanaly‐sisMJCledthewritingofthemanuscriptAllauthorsgavefinalap‐provalforpublication
emspensp emsp | emsp2001CLEMENT ET aL
DATA ACCE SSIBILIT Y
Raw data are available from the following sources Breeding BirdSurvey httpswwwpwrcusgsgovbbsrawdata PRISM climatedata httpwwwprismoregonstateedu NLCD land cover datahttpswwwmrlcgov Processed data will be archived with theDryadDigitalRepository
ORCID
Matthew J Clement httpsorcidorg0000‐0003‐4231‐7949
James D Nichols httpsorcidorg0000‐0002‐7631‐2890
Adam J Terando httpsorcidorg0000‐0002‐9280‐043X
James E Hines httpsorcidorg0000‐0001‐5478‐7230
Steven G Williams httpsorcidorg0000‐0003‐3760‐6818
R E FE R E N C E S
Anderson D R Burnham K P Gould W R amp Cherry S (2001)Concerns about finding effects that are actually spuriousWildlife Society Bulletin29311ndash316
Arauacutejo M B Cabeza M Thuiller W Hannah L amp WilliamsP H (2004) Would climate change drive species out of re‐serves An assessment of existing reserve‐selection meth‐ods Global Change Biology 10 1618ndash1626 httpsdoiorg101111j1365‐2486200400828x
Barbet‐MassinMampJetzW(2014)A40‐yearcontinent‐widemulti‐speciesassessmentofrelevantclimatepredictorsforspeciesdistri‐butionmodellingDiversity and Distributions201285ndash1295httpsdoiorg101111ddi12229
Barbet‐Massin M Thuiller W amp Jiguet F (2012) The fate ofEuropean breeding birds under climate land‐use and disper‐sal scenarios Global Change Biology 18 881ndash890 httpsdoiorg101111j1365‐2486201102552x
Bellard C Bertelsmeier C Leadley P Thuiller W ampCourchamp F (2012) Impacts of climate change on the fu‐ture of biodiversity Ecology Letters 15 365ndash377 httpsdoiorg101111j1461‐0248201101736x
Boumlhning‐Gaese K amp Bauer H G (1996) Changes in species abun‐dance distribution and diversity in a central European birdcommunity Conservation Biology 10 175ndash187 httpsdoiorg101046j1523‐1739199610010175x
BrommerJELehikoinenAampValkamaJ(2012)Thebreedingrangesof central European and Arctic bird speciesmove poleward PLoS ONE7e43648httpsdoiorg101371journalpone0043648
CalderWAampKingJR(1974)ThermalandcaloricrelationsofbirdsInDSFarnerampJRKing(Eds)Avian biology(Vol4pp259ndash413)NewYorkNYAcademicPress
CesaraccioCSpanoDDucePampSnyderRL(2001)Animprovedmodel for determining degree‐day values from daily temperaturedataInternational Journal of Biometeorology45161ndash169httpsdoiorg101007s004840100104
ChamberlinTC (1890)ThemethodofmultipleworkinghypothesesScience1592ndash96
ClementMJHinesJENicholsJDPardieckKLampZiolkowskiDJJr (2016)Estimating indicesofrangeshifts inbirdsusingdy‐namicmodelswhendetectionisimperfectGlobal Change Biology223273ndash3285httpsdoiorg101111gcb13283
Commission for Environmental Cooperation (1997) Ecological regionsofNorthAmericaTowardacommonperspectiveCommission for
EnvironmentalCooperationMontrealRetrievedfromftpftpepagovwedecoregionscec_naCEC_NAecopdf
Crowl TA Crist TO Parmenter R R BelovskyGamp LugoA E(2008)Thespreadofinvasivespeciesandinfectiousdiseaseasdriv‐ersofecosystemchangeFrontiers in Ecology and the Environment6238ndash246httpsdoiorg101890070151
Cuddington K FortinM J Gerber L RHastings A Liebhold AOConnorMampRayC(2013)Process‐basedmodelsarerequiredtomanageecologicalsystemsinachangingworldEcosphere41ndash12httpsdoiorg101890ES12‐001781
DalyCHalbleibMSmithJIGibsonWPDoggettMKTaylorGHhellipPasterisPP(2008)Physiographicallysensitivemappingofcli‐matologicaltemperatureandprecipitationacrosstheconterminousUnited States International Journal of Climatology 28 2031ndash2064httpsdoiorg101002joc1688
DawsonW (1958) Relation of oxygen consumption and evaporativewater losstotemperatureintheCardinalPhysiological Zoology3137ndash48httpsdoiorg101086physzool31130155377
DawsonWRampWhittowGC (2000)Regulationofbodytempera‐ture InG CWhittow (Ed)Sturkiersquos avian physiology (5th ed pp343ndash390)SanDiegoCAAcademicPress
DeCalestaDS(1994)Effectofwhite‐taileddeeronsongbirdswithinmanagedforestsinPennsylvaniaJournal of Wildlife Management58711ndash718httpsdoiorg1023073809685
DevictorVJulliardRCouvetDampJiguetF(2008)Birdsaretrack‐ing climatewarming but not fast enoughProceedings of the Royal Society of London B Biological Sciences2752743ndash2748httpsdoiorg101098rspb20080878
Dormann C F (2007) Promising the future Global change projec‐tionsofspeciesdistributionsBasic and Applied Ecology8387ndash397httpsdoiorg101016jbaae200611001
Dormann C F Schymanski S J Cabral J Chuine I Graham CHartigFhellipSingerA(2012)Correlationandprocessinspeciesdis‐tributionmodelsBridgingadichotomyJournal of Biogeography392119ndash2131httpsdoiorg101111j1365‐2699201102659x
EccelE(2010)WhatwecanasktohourlytemperaturerecordingPartIIHourlyinterpolationoftemperaturesforclimatologyandmodel‐lingItalian Journal of Agrometeorology1545ndash50
EccelEampCordanoE(2013)InterpolTHourlyinterpolationofmulti‐pletemperaturedailyseriesRpackageversion211RetrievedfromhttpsCRANR‐projectorgpackage=InterpolT
EglingtonSMampPearce‐HigginsJW(2012)Disentanglingtherela‐tiveimportanceofchangesinclimateandland‐useintensityindriv‐ingrecentbirdpopulationtrendsPLoS ONE7e30407httpsdoiorg101371journalpone0030407
Elith JKearneyMampPhillipsS (2010)Theartofmodelling range‐shifting species Methods in Ecology and Evolution 1 330ndash342httpsdoiorg101111j2041‐210X201000036x
FiebergJampJohnsonDH(2015)MMIMultimodelinferenceormod‐elswithmanagement implications Journal of Wildlife Management79708ndash718
FriggensMMampFinchDM (2015) Implicationsof climatechangeforbirdconservationinthesouthwesternUSunderthreealternativefuturesPLoS ONE10e0144089
FryJXianGZJinSDewitzJHomerCGYangLhellipWickhamJD (2011)Completionof the2006national landcoverdatabasefor theconterminousUnitedStatesPhotogrammetric Engineering amp Remote Sensing77858ndash864
Fuller R J Gregory R D Gibbons D W Marchant J HWilson J D Baillie S R amp Carter N (1995) Population de‐clines and range contractions among lowland farmland birdsin Britain Conservation Biology 9 1425ndash1441 httpsdoiorg101046j1523‐1739199509061425x
Gaston K J Blackburn T M Greenwood J J Gregory R DQuinn R M amp Lawton J H (2000) Abundancendashoccupancy
2002emsp |emsp emspensp CLEMENT ET aL
relationships Journal of Applied Ecology 37 39ndash59 httpsdoiorg101046j1365‐2664200000485x
GoldsteinRB(1974)RelationofmetabolismtoambienttemperatureintheverdinCondor76116ndash119httpsdoiorg1023071365995
GrinnellJ(1917)FieldtestsoftheoriesconcerningdistributionalcontrolAmerican Naturalist51115ndash128httpsdoiorg101086279591
GrosboisVGimenezOGaillardJMPradelRBarbraudCClobertJhellipWeimerskirchH(2008)Assessingtheimpactofclimatevari‐ation on survival in vertebrate populationsBiological Reviews 83357ndash399httpsdoiorg101111j1469‐185X200800047x
GustafsonEJ(2013)WhenrelationshipsestimatedinthepastcannotbeusedtopredictthefutureUsingmechanisticmodelstopredictlandscapeecologicaldynamicsinachangingworldLandscape Ecology281429ndash1437httpsdoiorg101007s10980‐013‐9927‐4
Hamel P B (1992) LandManagers guide to the birds of the SouthGeneralTechnicalReportSE‐22USDAForestServiceSoutheasternForestExperimentStationandTheNatureConservancyChapelHillNC
Hinds D S amp Calder W A (1973) Temperature regulation of thePyrrhuloxiaandtheArizonaCardinalPhysiological Zoology4655ndash71httpsdoiorg101086physzool46130152517
HinesJE(2006)PRESENCE Software to estimate patch occupancy and related parameters Laurel MD US Geological survey PatuxentWildlife Research Center Retrieved from httpwwwmbr‐pwrcusgsgovsoftwarepresencehtml
HinesJENicholsJDampCollazoJA(2014)Multiseasonoccupancymodels for correlated replicate surveys Methods in Ecology and Evolution5583ndash591httpsdoiorg1011112041‐210X12186
HinesJENicholsJDRoyleJAMacKenzieDIGopalaswamyAMKumarNampKaranthKU(2010)TigersontrailsOccupancymodeling for cluster sampling Ecological Applications 20 1456ndash1466httpsdoiorg10189009‐03211
HockeyPA SiramiCRidleyARMidgleyGFampBabikerHA(2011) Interrogating recent range changes in South African birdsConfounding signals from land use and climate change present achallenge for attribution Diversity and Distributions 17 254ndash261httpsdoiorg101111j1472‐4642201000741x
Hoegh‐Guldberg O Hughes L McIntyre S Lindenmayer D BParmesanCPossinghamHPampThomasCD (2008)AssistedcolonizationandrapidclimatechangeScience321345ndash346
Homer C Dewitz J Fry J Coan M Hossain N Larson C hellipWickham J (2007) Completion of the 2001 national land coverdatabase for the conterminous United States Photogrammetric Engineering amp Remote Sensing73337
HomerCDewitzJYangLJinSDanielsonPXianGhellipMegownK(2015)Completionofthe2011NationalLandCoverDatabasefortheconterminousUnitedStatesndashrepresentingadecadeoflandcoverchange informationPhotogrammetric Engineering amp Remote Sensing81345ndash354
HootenMBWikleCKDorazioRMampRoyleJA(2007)HierarchicalspatiotemporalmatrixmodelsforcharacterizinginvasionsBiometrics63558ndash567httpsdoiorg101111j1541‐0420200600725x
HosmerDWJrampLemeshowS(2000)Applied logistic regression(2nded)NewYorkNYJohnWileyampSons
Huntley B Collingham Y CGreen R EHiltonGM RahbekCamp Willis S G (2006) Potential impacts of climatic change upongeographical distributions of birds Ibis 148 8ndash28 httpsdoiorg101111j1474‐919X200600523x
IgleciaMNCollazoJAampMcKerrowAJ(2012)Useofoccupancymodels to evaluate expert knowledge‐based species‐habitat rela‐tionshipsAvian Conservation and Ecology7(2)13pRetrievedfromhttpwwwace‐ecoorgvol7iss2art5
IllaacutenJGThomasCDJonesJAWongWKShirleySMampBettsMG(2014)Precipitationandwintertemperaturepredictlong‐termrange‐scale abundancechanges inWesternNorthAmericanbirds
Global Change Biology 20 3351ndash3364 httpsdoiorg101111gcb12642
IoannidisJP (2005)WhymostpublishedresearchfindingsarefalsePLoS Medicine2e124
KeacuteryMGuillera‐ArroitaGampLahoz‐Monfort J J (2013)Analysingand mapping species range dynamics using occupancy modelsJournal of Biogeography 40 1463ndash1474 httpsdoiorg101111jbi12087
Khaliq I Hof C Prinzinger R Boumlhning‐Gaese K amp PfenningerM(2014) Global variation in thermal tolerances and vulnerability ofendotherms to climate change Proceedings of the Royal Society B Biological Sciences 281(1789) 20141097 httpsdoiorg101098rspb20141097
Laursen K amp Frikke J (2008) Hunting from motorboats displacesWadden Sea eiders Somateria mollissima from their favouredfeeding distribution Wildlife Biology 14 423ndash433 httpsdoiorg1029810909‐6396‐144423
LemoineNBauerHGPeintingerMampBoumlhning‐GaeseK (2007)Effectsof climate and land‐use changeon species abundance in acentralEuropeanbirdcommunityConservation Biology21495ndash503httpsdoiorg101111j1523‐1739200600633x
MacArthurRH(1972)Geographical ecology Patterns in the distribution of speciesNewYorkNYHarperampRow
MacKenzieDINicholsJDHinesJEKnutsonMGampFranklinAB(2003)Estimatingsiteoccupancycolonizationandlocalextinc‐tionwhenaspeciesisdetectedimperfectlyEcology842200ndash2207httpsdoiorg10189002‐3090
MacKenzieDINicholsJDLachmanGBDroegeSRoyleJAampLangtimmCA(2002)Estimatingsiteoccupancyrateswhendetec‐tionprobabilitiesarelessthanoneEcology832248ndash2255httpsdoiorg1018900012‐9658(2002)083[2248ESORWD]20CO2
MacKenzieDINicholsJDRoyleJAPollockKHBaileyLampHinesJE(2018)Occupancy estimation and modeling Inferring pat‐terns and dynamics of species occurrence (2nd ed) BurlingtonMAAcademicPress
MellesSJFortinMJLindsayKampBadzinskiD(2011)ExpandingnorthwardInfluenceofclimatechangeforestconnectivityandpop‐ulationprocessesonathreatenedspeciesrangeshiftGlobal Change Biology1717ndash31httpsdoiorg101111j1365‐2486201002214x
Morin X amp Thuiller W (2009) Comparing niche‐and process‐based models to reduce prediction uncertainty in species rangeshifts under climate change Ecology 90 1301ndash1313 httpsdoiorg10189008‐01341
MustinKAmarAampRedpathSM(2014)Colonizationandextinctiondynamicsofadecliningmigratorybirdareinfluencedbyclimateandhabitat degradation Ibis 156 788ndash798 httpsdoiorg101111ibi12173
NicholsJDThomasLampConnPB(2009)InferencesaboutlandbirdabundancefromcountdataRecentadvancesandfuturedirectionsInDLThomsonEJCoochampMJConroy(Eds)Modeling demo‐graphic processes in marked populations(pp201ndash235)NewYorkNYSpringer
OmernikJM(1995)EcoregionsAspatialframeworkforenvironmen‐talmanagement InWSDavisampTSimon(Eds)Biological assess‐ment and criteria Tools for water resource planning and decision making (pp49ndash62)BocaRatonFLLewisPublishers
OmernikJM(2004)Perspectivesonthenatureanddefinitionofeco‐logicalregionsEnvironmental Management34(Supplement1)S27ndashS38httpsdoiorg101007s00267‐003‐5197‐2
Omernik J M amp Griffith G E (2014) Ecoregions of the contermi‐nousUnited States Evolution of a hierarchical spatial frameworkEnvironmental Management541249ndash1266httpsdoiorg101007s00267‐014‐0364‐1
Pardieck K L ZiolkowskiD J JrampHudsonM‐A R (2015)NorthAmericanBreedingBirdSurveyDataset1966ndash2014version20140
emspensp emsp | emsp2003CLEMENT ET aL
USGeologicalSurveyPatuxentWildlifeResearchCenterRetrievedfromhttpswwwpwrcusgsgovbbsrawdata
ParmesanCampYoheG (2003)Agloballycoherentfingerprintofcli‐mate change impacts across natural systemsNature 421 37ndash42httpsdoiorg101038nature01286
PearsonRGampDawsonTP(2003)PredictingtheimpactsofclimatechangeonthedistributionofspeciesArebioclimateenvelopemod‐elsusefulGlobal Ecology and Biogeography12361ndash371
PlattJR(1964)StronginferenceScience146347ndash353PopperK(1959)The logic of scientific discoveryLondonUKHutchinsonRobbins C S Sauer J R Greenberg R S amp Droege S (1989)
Population declines in North American birds that migrate to theNeotropics Proceedings of the National Academy of Sciences 867658ndash7662httpsdoiorg101073pnas86197658
Robinson R A Wilson J D amp Crick H Q (2001) The impor‐tance of arable habitat for farmland birds in grassland land‐scapes Journal of Applied Ecology 38 1059ndash1069 httpsdoiorg101046j1365‐2664200100654x
Romesburg H C (1981) Wildlife science Gaining reliable knowl‐edge Journal of Wildlife Management 45 293ndash313 httpsdoiorg1023073807913
Root T (1988) Energy constraints on avian distributions and abun‐dancesEcology69330ndash339httpsdoiorg1023071940431
Ruiz‐GutieacuterrezVampZipkinEF (2011)Detectionbiasesyieldmis‐leadingpatternsof speciespersistence and colonization in frag‐mented landscapesEcosphere2 1ndash14 httpsdoiorg101890ES10‐002071
SauerJRHinesJEFallonJEPardieckKLZiolkowskiDJJrampLinkWA(2015)The North American breeding bird survey results and analysis 1966ndash2013 Version 01302015LaurelMDUSGSPatuxentWildlifeResearchCenterhttpswwwmbr‐pwrcusgsgovbbsbbshtml
Sauer J R LinkWA Fallon J E Pardieck K L amp ZiolkowskiDJJr(2013)TheNorthAmericanbreedingbirdsurvey1966ndash2011SummaryanalysisandspeciesaccountsNorth American Fauna791ndash32httpsdoiorg103996nafa790001
SeoaneJBustamanteJampDiacuteaz‐DelgadoR(2004)Competingrolesforlandscapevegetationtopographyandclimateinpredictivemod‐elsofbirddistributionEcological Modelling171209ndash222httpsdoiorg101016jecolmodel200308006
Sirami C Caplat P Popy S Clamens A Arlettaz R Jiguet F hellipMartin J‐L (2017) Impacts of global change on species distri‐butions Obstacles and solutions to integrate climate and landuse Global Ecology and Biogeography 26 385ndash394 httpsdoiorg101111geb12555
SkalskiJR(1996)Regressionofabundanceestimatesfrommarkrecap‐ture surveys against environmental covariatesCanadian Journal of
Fisheries and Aquatic Sciences53196ndash204httpsdoiorg101139f95‐169
SohlTL(2014)Therelativeimpactsofclimateandland‐usechangeonconterminousUnitedStatesbird species from2001 to2075PLoS ONE9e112251httpsdoiorg101371journalpone0112251
ThogmartinWESauerJRampKnutsonMG(2007)ModelingandmappingabundanceofAmericanwoodcockacrosstheMidwesternandNortheasternUnitedStatesJournal of Wildlife Management71376ndash382httpsdoiorg1021932005‐680
ThomasCDampLennonJJ (1999)Birdsextendtheirrangesnorth‐wardsNature399213httpsdoiorg10103820335
Trathan P N Garciacutea‐Borboroglu P Boersma D Bost C ACrawford R J CrossinG ThellipWienecke B (2015) Pollutionhabitatlossfishingandclimatechangeascriticalthreatstopen‐guins Conservation Biology 29 31ndash41 httpsdoiorg101111cobi12349
TravisJMJ(2003)Climatechangeandhabitatdestructionadeadlyanthropogenic cocktail Proceedings of the Royal Society of London Series B Biological Sciences 270(1514) 467ndash473 httpsdoiorg101098rspb20022246
UrbanMC (2015)Acceleratingextinction risk fromclimatechangeScience348571ndash573httpsdoiorg101126scienceaaa4984
WalshJWuebblesDHayhoeKKossinJKunkelKStephensGhellipSomervilleR(2014)OurchangingclimateInJMMelilloTCRichmondampGWYohe(Eds)Climate change impacts in the United States The Third National climate assessment(pp19ndash67)WashingtonDCUSGovernmentPrintingOffice
YackulicCBNichols JDReid JampDerR (2015)Topredict thenichemodelcolonizationandextinctionEcology9616ndash23httpsdoiorg10189014‐13611
Yalcin S amp Leroux S J (2018) An empirical test of the relative andcombinedeffectsofland‐coverandclimatechangeonlocalcoloni‐zationandextinctionGlobal Change Biology243849ndash3861httpsdoiorg101111gcb14169
ZhuKWoodallCWampClarkJS(2012)FailuretomigrateLackoftreerangeexpansioninresponsetoclimatechangeGlobal Change Biology181042ndash1052httpsdoiorg101111j1365‐2486201102571x
How to cite this articleClementMJNicholsJDCollazoJATerandoAJHinesJEWilliamsSGPartitioningglobalchangeAssessingtherelativeimportanceofchangesinclimateandlandcoverforchangesinaviandistributionEcol Evol 201991985ndash2003 httpsdoiorg101002ece34890

emspensp emsp | emsp2001CLEMENT ET aL
DATA ACCE SSIBILIT Y
Raw data are available from the following sources Breeding BirdSurvey httpswwwpwrcusgsgovbbsrawdata PRISM climatedata httpwwwprismoregonstateedu NLCD land cover datahttpswwwmrlcgov Processed data will be archived with theDryadDigitalRepository
ORCID
Matthew J Clement httpsorcidorg0000‐0003‐4231‐7949
James D Nichols httpsorcidorg0000‐0002‐7631‐2890
Adam J Terando httpsorcidorg0000‐0002‐9280‐043X
James E Hines httpsorcidorg0000‐0001‐5478‐7230
Steven G Williams httpsorcidorg0000‐0003‐3760‐6818
R E FE R E N C E S
Anderson D R Burnham K P Gould W R amp Cherry S (2001)Concerns about finding effects that are actually spuriousWildlife Society Bulletin29311ndash316
Arauacutejo M B Cabeza M Thuiller W Hannah L amp WilliamsP H (2004) Would climate change drive species out of re‐serves An assessment of existing reserve‐selection meth‐ods Global Change Biology 10 1618ndash1626 httpsdoiorg101111j1365‐2486200400828x
Barbet‐MassinMampJetzW(2014)A40‐yearcontinent‐widemulti‐speciesassessmentofrelevantclimatepredictorsforspeciesdistri‐butionmodellingDiversity and Distributions201285ndash1295httpsdoiorg101111ddi12229
Barbet‐Massin M Thuiller W amp Jiguet F (2012) The fate ofEuropean breeding birds under climate land‐use and disper‐sal scenarios Global Change Biology 18 881ndash890 httpsdoiorg101111j1365‐2486201102552x
Bellard C Bertelsmeier C Leadley P Thuiller W ampCourchamp F (2012) Impacts of climate change on the fu‐ture of biodiversity Ecology Letters 15 365ndash377 httpsdoiorg101111j1461‐0248201101736x
Boumlhning‐Gaese K amp Bauer H G (1996) Changes in species abun‐dance distribution and diversity in a central European birdcommunity Conservation Biology 10 175ndash187 httpsdoiorg101046j1523‐1739199610010175x
BrommerJELehikoinenAampValkamaJ(2012)Thebreedingrangesof central European and Arctic bird speciesmove poleward PLoS ONE7e43648httpsdoiorg101371journalpone0043648
CalderWAampKingJR(1974)ThermalandcaloricrelationsofbirdsInDSFarnerampJRKing(Eds)Avian biology(Vol4pp259ndash413)NewYorkNYAcademicPress
CesaraccioCSpanoDDucePampSnyderRL(2001)Animprovedmodel for determining degree‐day values from daily temperaturedataInternational Journal of Biometeorology45161ndash169httpsdoiorg101007s004840100104
ChamberlinTC (1890)ThemethodofmultipleworkinghypothesesScience1592ndash96
ClementMJHinesJENicholsJDPardieckKLampZiolkowskiDJJr (2016)Estimating indicesofrangeshifts inbirdsusingdy‐namicmodelswhendetectionisimperfectGlobal Change Biology223273ndash3285httpsdoiorg101111gcb13283
Commission for Environmental Cooperation (1997) Ecological regionsofNorthAmericaTowardacommonperspectiveCommission for
EnvironmentalCooperationMontrealRetrievedfromftpftpepagovwedecoregionscec_naCEC_NAecopdf
Crowl TA Crist TO Parmenter R R BelovskyGamp LugoA E(2008)Thespreadofinvasivespeciesandinfectiousdiseaseasdriv‐ersofecosystemchangeFrontiers in Ecology and the Environment6238ndash246httpsdoiorg101890070151
Cuddington K FortinM J Gerber L RHastings A Liebhold AOConnorMampRayC(2013)Process‐basedmodelsarerequiredtomanageecologicalsystemsinachangingworldEcosphere41ndash12httpsdoiorg101890ES12‐001781
DalyCHalbleibMSmithJIGibsonWPDoggettMKTaylorGHhellipPasterisPP(2008)Physiographicallysensitivemappingofcli‐matologicaltemperatureandprecipitationacrosstheconterminousUnited States International Journal of Climatology 28 2031ndash2064httpsdoiorg101002joc1688
DawsonW (1958) Relation of oxygen consumption and evaporativewater losstotemperatureintheCardinalPhysiological Zoology3137ndash48httpsdoiorg101086physzool31130155377
DawsonWRampWhittowGC (2000)Regulationofbodytempera‐ture InG CWhittow (Ed)Sturkiersquos avian physiology (5th ed pp343ndash390)SanDiegoCAAcademicPress
DeCalestaDS(1994)Effectofwhite‐taileddeeronsongbirdswithinmanagedforestsinPennsylvaniaJournal of Wildlife Management58711ndash718httpsdoiorg1023073809685
DevictorVJulliardRCouvetDampJiguetF(2008)Birdsaretrack‐ing climatewarming but not fast enoughProceedings of the Royal Society of London B Biological Sciences2752743ndash2748httpsdoiorg101098rspb20080878
Dormann C F (2007) Promising the future Global change projec‐tionsofspeciesdistributionsBasic and Applied Ecology8387ndash397httpsdoiorg101016jbaae200611001
Dormann C F Schymanski S J Cabral J Chuine I Graham CHartigFhellipSingerA(2012)Correlationandprocessinspeciesdis‐tributionmodelsBridgingadichotomyJournal of Biogeography392119ndash2131httpsdoiorg101111j1365‐2699201102659x
EccelE(2010)WhatwecanasktohourlytemperaturerecordingPartIIHourlyinterpolationoftemperaturesforclimatologyandmodel‐lingItalian Journal of Agrometeorology1545ndash50
EccelEampCordanoE(2013)InterpolTHourlyinterpolationofmulti‐pletemperaturedailyseriesRpackageversion211RetrievedfromhttpsCRANR‐projectorgpackage=InterpolT
EglingtonSMampPearce‐HigginsJW(2012)Disentanglingtherela‐tiveimportanceofchangesinclimateandland‐useintensityindriv‐ingrecentbirdpopulationtrendsPLoS ONE7e30407httpsdoiorg101371journalpone0030407
Elith JKearneyMampPhillipsS (2010)Theartofmodelling range‐shifting species Methods in Ecology and Evolution 1 330ndash342httpsdoiorg101111j2041‐210X201000036x
FiebergJampJohnsonDH(2015)MMIMultimodelinferenceormod‐elswithmanagement implications Journal of Wildlife Management79708ndash718
FriggensMMampFinchDM (2015) Implicationsof climatechangeforbirdconservationinthesouthwesternUSunderthreealternativefuturesPLoS ONE10e0144089
FryJXianGZJinSDewitzJHomerCGYangLhellipWickhamJD (2011)Completionof the2006national landcoverdatabasefor theconterminousUnitedStatesPhotogrammetric Engineering amp Remote Sensing77858ndash864
Fuller R J Gregory R D Gibbons D W Marchant J HWilson J D Baillie S R amp Carter N (1995) Population de‐clines and range contractions among lowland farmland birdsin Britain Conservation Biology 9 1425ndash1441 httpsdoiorg101046j1523‐1739199509061425x
Gaston K J Blackburn T M Greenwood J J Gregory R DQuinn R M amp Lawton J H (2000) Abundancendashoccupancy
2002emsp |emsp emspensp CLEMENT ET aL
relationships Journal of Applied Ecology 37 39ndash59 httpsdoiorg101046j1365‐2664200000485x
GoldsteinRB(1974)RelationofmetabolismtoambienttemperatureintheverdinCondor76116ndash119httpsdoiorg1023071365995
GrinnellJ(1917)FieldtestsoftheoriesconcerningdistributionalcontrolAmerican Naturalist51115ndash128httpsdoiorg101086279591
GrosboisVGimenezOGaillardJMPradelRBarbraudCClobertJhellipWeimerskirchH(2008)Assessingtheimpactofclimatevari‐ation on survival in vertebrate populationsBiological Reviews 83357ndash399httpsdoiorg101111j1469‐185X200800047x
GustafsonEJ(2013)WhenrelationshipsestimatedinthepastcannotbeusedtopredictthefutureUsingmechanisticmodelstopredictlandscapeecologicaldynamicsinachangingworldLandscape Ecology281429ndash1437httpsdoiorg101007s10980‐013‐9927‐4
Hamel P B (1992) LandManagers guide to the birds of the SouthGeneralTechnicalReportSE‐22USDAForestServiceSoutheasternForestExperimentStationandTheNatureConservancyChapelHillNC
Hinds D S amp Calder W A (1973) Temperature regulation of thePyrrhuloxiaandtheArizonaCardinalPhysiological Zoology4655ndash71httpsdoiorg101086physzool46130152517
HinesJE(2006)PRESENCE Software to estimate patch occupancy and related parameters Laurel MD US Geological survey PatuxentWildlife Research Center Retrieved from httpwwwmbr‐pwrcusgsgovsoftwarepresencehtml
HinesJENicholsJDampCollazoJA(2014)Multiseasonoccupancymodels for correlated replicate surveys Methods in Ecology and Evolution5583ndash591httpsdoiorg1011112041‐210X12186
HinesJENicholsJDRoyleJAMacKenzieDIGopalaswamyAMKumarNampKaranthKU(2010)TigersontrailsOccupancymodeling for cluster sampling Ecological Applications 20 1456ndash1466httpsdoiorg10189009‐03211
HockeyPA SiramiCRidleyARMidgleyGFampBabikerHA(2011) Interrogating recent range changes in South African birdsConfounding signals from land use and climate change present achallenge for attribution Diversity and Distributions 17 254ndash261httpsdoiorg101111j1472‐4642201000741x
Hoegh‐Guldberg O Hughes L McIntyre S Lindenmayer D BParmesanCPossinghamHPampThomasCD (2008)AssistedcolonizationandrapidclimatechangeScience321345ndash346
Homer C Dewitz J Fry J Coan M Hossain N Larson C hellipWickham J (2007) Completion of the 2001 national land coverdatabase for the conterminous United States Photogrammetric Engineering amp Remote Sensing73337
HomerCDewitzJYangLJinSDanielsonPXianGhellipMegownK(2015)Completionofthe2011NationalLandCoverDatabasefortheconterminousUnitedStatesndashrepresentingadecadeoflandcoverchange informationPhotogrammetric Engineering amp Remote Sensing81345ndash354
HootenMBWikleCKDorazioRMampRoyleJA(2007)HierarchicalspatiotemporalmatrixmodelsforcharacterizinginvasionsBiometrics63558ndash567httpsdoiorg101111j1541‐0420200600725x
HosmerDWJrampLemeshowS(2000)Applied logistic regression(2nded)NewYorkNYJohnWileyampSons
Huntley B Collingham Y CGreen R EHiltonGM RahbekCamp Willis S G (2006) Potential impacts of climatic change upongeographical distributions of birds Ibis 148 8ndash28 httpsdoiorg101111j1474‐919X200600523x
IgleciaMNCollazoJAampMcKerrowAJ(2012)Useofoccupancymodels to evaluate expert knowledge‐based species‐habitat rela‐tionshipsAvian Conservation and Ecology7(2)13pRetrievedfromhttpwwwace‐ecoorgvol7iss2art5
IllaacutenJGThomasCDJonesJAWongWKShirleySMampBettsMG(2014)Precipitationandwintertemperaturepredictlong‐termrange‐scale abundancechanges inWesternNorthAmericanbirds
Global Change Biology 20 3351ndash3364 httpsdoiorg101111gcb12642
IoannidisJP (2005)WhymostpublishedresearchfindingsarefalsePLoS Medicine2e124
KeacuteryMGuillera‐ArroitaGampLahoz‐Monfort J J (2013)Analysingand mapping species range dynamics using occupancy modelsJournal of Biogeography 40 1463ndash1474 httpsdoiorg101111jbi12087
Khaliq I Hof C Prinzinger R Boumlhning‐Gaese K amp PfenningerM(2014) Global variation in thermal tolerances and vulnerability ofendotherms to climate change Proceedings of the Royal Society B Biological Sciences 281(1789) 20141097 httpsdoiorg101098rspb20141097
Laursen K amp Frikke J (2008) Hunting from motorboats displacesWadden Sea eiders Somateria mollissima from their favouredfeeding distribution Wildlife Biology 14 423ndash433 httpsdoiorg1029810909‐6396‐144423
LemoineNBauerHGPeintingerMampBoumlhning‐GaeseK (2007)Effectsof climate and land‐use changeon species abundance in acentralEuropeanbirdcommunityConservation Biology21495ndash503httpsdoiorg101111j1523‐1739200600633x
MacArthurRH(1972)Geographical ecology Patterns in the distribution of speciesNewYorkNYHarperampRow
MacKenzieDINicholsJDHinesJEKnutsonMGampFranklinAB(2003)Estimatingsiteoccupancycolonizationandlocalextinc‐tionwhenaspeciesisdetectedimperfectlyEcology842200ndash2207httpsdoiorg10189002‐3090
MacKenzieDINicholsJDLachmanGBDroegeSRoyleJAampLangtimmCA(2002)Estimatingsiteoccupancyrateswhendetec‐tionprobabilitiesarelessthanoneEcology832248ndash2255httpsdoiorg1018900012‐9658(2002)083[2248ESORWD]20CO2
MacKenzieDINicholsJDRoyleJAPollockKHBaileyLampHinesJE(2018)Occupancy estimation and modeling Inferring pat‐terns and dynamics of species occurrence (2nd ed) BurlingtonMAAcademicPress
MellesSJFortinMJLindsayKampBadzinskiD(2011)ExpandingnorthwardInfluenceofclimatechangeforestconnectivityandpop‐ulationprocessesonathreatenedspeciesrangeshiftGlobal Change Biology1717ndash31httpsdoiorg101111j1365‐2486201002214x
Morin X amp Thuiller W (2009) Comparing niche‐and process‐based models to reduce prediction uncertainty in species rangeshifts under climate change Ecology 90 1301ndash1313 httpsdoiorg10189008‐01341
MustinKAmarAampRedpathSM(2014)Colonizationandextinctiondynamicsofadecliningmigratorybirdareinfluencedbyclimateandhabitat degradation Ibis 156 788ndash798 httpsdoiorg101111ibi12173
NicholsJDThomasLampConnPB(2009)InferencesaboutlandbirdabundancefromcountdataRecentadvancesandfuturedirectionsInDLThomsonEJCoochampMJConroy(Eds)Modeling demo‐graphic processes in marked populations(pp201ndash235)NewYorkNYSpringer
OmernikJM(1995)EcoregionsAspatialframeworkforenvironmen‐talmanagement InWSDavisampTSimon(Eds)Biological assess‐ment and criteria Tools for water resource planning and decision making (pp49ndash62)BocaRatonFLLewisPublishers
OmernikJM(2004)Perspectivesonthenatureanddefinitionofeco‐logicalregionsEnvironmental Management34(Supplement1)S27ndashS38httpsdoiorg101007s00267‐003‐5197‐2
Omernik J M amp Griffith G E (2014) Ecoregions of the contermi‐nousUnited States Evolution of a hierarchical spatial frameworkEnvironmental Management541249ndash1266httpsdoiorg101007s00267‐014‐0364‐1
Pardieck K L ZiolkowskiD J JrampHudsonM‐A R (2015)NorthAmericanBreedingBirdSurveyDataset1966ndash2014version20140
emspensp emsp | emsp2003CLEMENT ET aL
USGeologicalSurveyPatuxentWildlifeResearchCenterRetrievedfromhttpswwwpwrcusgsgovbbsrawdata
ParmesanCampYoheG (2003)Agloballycoherentfingerprintofcli‐mate change impacts across natural systemsNature 421 37ndash42httpsdoiorg101038nature01286
PearsonRGampDawsonTP(2003)PredictingtheimpactsofclimatechangeonthedistributionofspeciesArebioclimateenvelopemod‐elsusefulGlobal Ecology and Biogeography12361ndash371
PlattJR(1964)StronginferenceScience146347ndash353PopperK(1959)The logic of scientific discoveryLondonUKHutchinsonRobbins C S Sauer J R Greenberg R S amp Droege S (1989)
Population declines in North American birds that migrate to theNeotropics Proceedings of the National Academy of Sciences 867658ndash7662httpsdoiorg101073pnas86197658
Robinson R A Wilson J D amp Crick H Q (2001) The impor‐tance of arable habitat for farmland birds in grassland land‐scapes Journal of Applied Ecology 38 1059ndash1069 httpsdoiorg101046j1365‐2664200100654x
Romesburg H C (1981) Wildlife science Gaining reliable knowl‐edge Journal of Wildlife Management 45 293ndash313 httpsdoiorg1023073807913
Root T (1988) Energy constraints on avian distributions and abun‐dancesEcology69330ndash339httpsdoiorg1023071940431
Ruiz‐GutieacuterrezVampZipkinEF (2011)Detectionbiasesyieldmis‐leadingpatternsof speciespersistence and colonization in frag‐mented landscapesEcosphere2 1ndash14 httpsdoiorg101890ES10‐002071
SauerJRHinesJEFallonJEPardieckKLZiolkowskiDJJrampLinkWA(2015)The North American breeding bird survey results and analysis 1966ndash2013 Version 01302015LaurelMDUSGSPatuxentWildlifeResearchCenterhttpswwwmbr‐pwrcusgsgovbbsbbshtml
Sauer J R LinkWA Fallon J E Pardieck K L amp ZiolkowskiDJJr(2013)TheNorthAmericanbreedingbirdsurvey1966ndash2011SummaryanalysisandspeciesaccountsNorth American Fauna791ndash32httpsdoiorg103996nafa790001
SeoaneJBustamanteJampDiacuteaz‐DelgadoR(2004)Competingrolesforlandscapevegetationtopographyandclimateinpredictivemod‐elsofbirddistributionEcological Modelling171209ndash222httpsdoiorg101016jecolmodel200308006
Sirami C Caplat P Popy S Clamens A Arlettaz R Jiguet F hellipMartin J‐L (2017) Impacts of global change on species distri‐butions Obstacles and solutions to integrate climate and landuse Global Ecology and Biogeography 26 385ndash394 httpsdoiorg101111geb12555
SkalskiJR(1996)Regressionofabundanceestimatesfrommarkrecap‐ture surveys against environmental covariatesCanadian Journal of
Fisheries and Aquatic Sciences53196ndash204httpsdoiorg101139f95‐169
SohlTL(2014)Therelativeimpactsofclimateandland‐usechangeonconterminousUnitedStatesbird species from2001 to2075PLoS ONE9e112251httpsdoiorg101371journalpone0112251
ThogmartinWESauerJRampKnutsonMG(2007)ModelingandmappingabundanceofAmericanwoodcockacrosstheMidwesternandNortheasternUnitedStatesJournal of Wildlife Management71376ndash382httpsdoiorg1021932005‐680
ThomasCDampLennonJJ (1999)Birdsextendtheirrangesnorth‐wardsNature399213httpsdoiorg10103820335
Trathan P N Garciacutea‐Borboroglu P Boersma D Bost C ACrawford R J CrossinG ThellipWienecke B (2015) Pollutionhabitatlossfishingandclimatechangeascriticalthreatstopen‐guins Conservation Biology 29 31ndash41 httpsdoiorg101111cobi12349
TravisJMJ(2003)Climatechangeandhabitatdestructionadeadlyanthropogenic cocktail Proceedings of the Royal Society of London Series B Biological Sciences 270(1514) 467ndash473 httpsdoiorg101098rspb20022246
UrbanMC (2015)Acceleratingextinction risk fromclimatechangeScience348571ndash573httpsdoiorg101126scienceaaa4984
WalshJWuebblesDHayhoeKKossinJKunkelKStephensGhellipSomervilleR(2014)OurchangingclimateInJMMelilloTCRichmondampGWYohe(Eds)Climate change impacts in the United States The Third National climate assessment(pp19ndash67)WashingtonDCUSGovernmentPrintingOffice
YackulicCBNichols JDReid JampDerR (2015)Topredict thenichemodelcolonizationandextinctionEcology9616ndash23httpsdoiorg10189014‐13611
Yalcin S amp Leroux S J (2018) An empirical test of the relative andcombinedeffectsofland‐coverandclimatechangeonlocalcoloni‐zationandextinctionGlobal Change Biology243849ndash3861httpsdoiorg101111gcb14169
ZhuKWoodallCWampClarkJS(2012)FailuretomigrateLackoftreerangeexpansioninresponsetoclimatechangeGlobal Change Biology181042ndash1052httpsdoiorg101111j1365‐2486201102571x
How to cite this articleClementMJNicholsJDCollazoJATerandoAJHinesJEWilliamsSGPartitioningglobalchangeAssessingtherelativeimportanceofchangesinclimateandlandcoverforchangesinaviandistributionEcol Evol 201991985ndash2003 httpsdoiorg101002ece34890

2002emsp |emsp emspensp CLEMENT ET aL
relationships Journal of Applied Ecology 37 39ndash59 httpsdoiorg101046j1365‐2664200000485x
GoldsteinRB(1974)RelationofmetabolismtoambienttemperatureintheverdinCondor76116ndash119httpsdoiorg1023071365995
GrinnellJ(1917)FieldtestsoftheoriesconcerningdistributionalcontrolAmerican Naturalist51115ndash128httpsdoiorg101086279591
GrosboisVGimenezOGaillardJMPradelRBarbraudCClobertJhellipWeimerskirchH(2008)Assessingtheimpactofclimatevari‐ation on survival in vertebrate populationsBiological Reviews 83357ndash399httpsdoiorg101111j1469‐185X200800047x
GustafsonEJ(2013)WhenrelationshipsestimatedinthepastcannotbeusedtopredictthefutureUsingmechanisticmodelstopredictlandscapeecologicaldynamicsinachangingworldLandscape Ecology281429ndash1437httpsdoiorg101007s10980‐013‐9927‐4
Hamel P B (1992) LandManagers guide to the birds of the SouthGeneralTechnicalReportSE‐22USDAForestServiceSoutheasternForestExperimentStationandTheNatureConservancyChapelHillNC
Hinds D S amp Calder W A (1973) Temperature regulation of thePyrrhuloxiaandtheArizonaCardinalPhysiological Zoology4655ndash71httpsdoiorg101086physzool46130152517
HinesJE(2006)PRESENCE Software to estimate patch occupancy and related parameters Laurel MD US Geological survey PatuxentWildlife Research Center Retrieved from httpwwwmbr‐pwrcusgsgovsoftwarepresencehtml
HinesJENicholsJDampCollazoJA(2014)Multiseasonoccupancymodels for correlated replicate surveys Methods in Ecology and Evolution5583ndash591httpsdoiorg1011112041‐210X12186
HinesJENicholsJDRoyleJAMacKenzieDIGopalaswamyAMKumarNampKaranthKU(2010)TigersontrailsOccupancymodeling for cluster sampling Ecological Applications 20 1456ndash1466httpsdoiorg10189009‐03211
HockeyPA SiramiCRidleyARMidgleyGFampBabikerHA(2011) Interrogating recent range changes in South African birdsConfounding signals from land use and climate change present achallenge for attribution Diversity and Distributions 17 254ndash261httpsdoiorg101111j1472‐4642201000741x
Hoegh‐Guldberg O Hughes L McIntyre S Lindenmayer D BParmesanCPossinghamHPampThomasCD (2008)AssistedcolonizationandrapidclimatechangeScience321345ndash346
Homer C Dewitz J Fry J Coan M Hossain N Larson C hellipWickham J (2007) Completion of the 2001 national land coverdatabase for the conterminous United States Photogrammetric Engineering amp Remote Sensing73337
HomerCDewitzJYangLJinSDanielsonPXianGhellipMegownK(2015)Completionofthe2011NationalLandCoverDatabasefortheconterminousUnitedStatesndashrepresentingadecadeoflandcoverchange informationPhotogrammetric Engineering amp Remote Sensing81345ndash354
HootenMBWikleCKDorazioRMampRoyleJA(2007)HierarchicalspatiotemporalmatrixmodelsforcharacterizinginvasionsBiometrics63558ndash567httpsdoiorg101111j1541‐0420200600725x
HosmerDWJrampLemeshowS(2000)Applied logistic regression(2nded)NewYorkNYJohnWileyampSons
Huntley B Collingham Y CGreen R EHiltonGM RahbekCamp Willis S G (2006) Potential impacts of climatic change upongeographical distributions of birds Ibis 148 8ndash28 httpsdoiorg101111j1474‐919X200600523x
IgleciaMNCollazoJAampMcKerrowAJ(2012)Useofoccupancymodels to evaluate expert knowledge‐based species‐habitat rela‐tionshipsAvian Conservation and Ecology7(2)13pRetrievedfromhttpwwwace‐ecoorgvol7iss2art5
IllaacutenJGThomasCDJonesJAWongWKShirleySMampBettsMG(2014)Precipitationandwintertemperaturepredictlong‐termrange‐scale abundancechanges inWesternNorthAmericanbirds
Global Change Biology 20 3351ndash3364 httpsdoiorg101111gcb12642
IoannidisJP (2005)WhymostpublishedresearchfindingsarefalsePLoS Medicine2e124
KeacuteryMGuillera‐ArroitaGampLahoz‐Monfort J J (2013)Analysingand mapping species range dynamics using occupancy modelsJournal of Biogeography 40 1463ndash1474 httpsdoiorg101111jbi12087
Khaliq I Hof C Prinzinger R Boumlhning‐Gaese K amp PfenningerM(2014) Global variation in thermal tolerances and vulnerability ofendotherms to climate change Proceedings of the Royal Society B Biological Sciences 281(1789) 20141097 httpsdoiorg101098rspb20141097
Laursen K amp Frikke J (2008) Hunting from motorboats displacesWadden Sea eiders Somateria mollissima from their favouredfeeding distribution Wildlife Biology 14 423ndash433 httpsdoiorg1029810909‐6396‐144423
LemoineNBauerHGPeintingerMampBoumlhning‐GaeseK (2007)Effectsof climate and land‐use changeon species abundance in acentralEuropeanbirdcommunityConservation Biology21495ndash503httpsdoiorg101111j1523‐1739200600633x
MacArthurRH(1972)Geographical ecology Patterns in the distribution of speciesNewYorkNYHarperampRow
MacKenzieDINicholsJDHinesJEKnutsonMGampFranklinAB(2003)Estimatingsiteoccupancycolonizationandlocalextinc‐tionwhenaspeciesisdetectedimperfectlyEcology842200ndash2207httpsdoiorg10189002‐3090
MacKenzieDINicholsJDLachmanGBDroegeSRoyleJAampLangtimmCA(2002)Estimatingsiteoccupancyrateswhendetec‐tionprobabilitiesarelessthanoneEcology832248ndash2255httpsdoiorg1018900012‐9658(2002)083[2248ESORWD]20CO2
MacKenzieDINicholsJDRoyleJAPollockKHBaileyLampHinesJE(2018)Occupancy estimation and modeling Inferring pat‐terns and dynamics of species occurrence (2nd ed) BurlingtonMAAcademicPress
MellesSJFortinMJLindsayKampBadzinskiD(2011)ExpandingnorthwardInfluenceofclimatechangeforestconnectivityandpop‐ulationprocessesonathreatenedspeciesrangeshiftGlobal Change Biology1717ndash31httpsdoiorg101111j1365‐2486201002214x
Morin X amp Thuiller W (2009) Comparing niche‐and process‐based models to reduce prediction uncertainty in species rangeshifts under climate change Ecology 90 1301ndash1313 httpsdoiorg10189008‐01341
MustinKAmarAampRedpathSM(2014)Colonizationandextinctiondynamicsofadecliningmigratorybirdareinfluencedbyclimateandhabitat degradation Ibis 156 788ndash798 httpsdoiorg101111ibi12173
NicholsJDThomasLampConnPB(2009)InferencesaboutlandbirdabundancefromcountdataRecentadvancesandfuturedirectionsInDLThomsonEJCoochampMJConroy(Eds)Modeling demo‐graphic processes in marked populations(pp201ndash235)NewYorkNYSpringer
OmernikJM(1995)EcoregionsAspatialframeworkforenvironmen‐talmanagement InWSDavisampTSimon(Eds)Biological assess‐ment and criteria Tools for water resource planning and decision making (pp49ndash62)BocaRatonFLLewisPublishers
OmernikJM(2004)Perspectivesonthenatureanddefinitionofeco‐logicalregionsEnvironmental Management34(Supplement1)S27ndashS38httpsdoiorg101007s00267‐003‐5197‐2
Omernik J M amp Griffith G E (2014) Ecoregions of the contermi‐nousUnited States Evolution of a hierarchical spatial frameworkEnvironmental Management541249ndash1266httpsdoiorg101007s00267‐014‐0364‐1
Pardieck K L ZiolkowskiD J JrampHudsonM‐A R (2015)NorthAmericanBreedingBirdSurveyDataset1966ndash2014version20140
emspensp emsp | emsp2003CLEMENT ET aL
USGeologicalSurveyPatuxentWildlifeResearchCenterRetrievedfromhttpswwwpwrcusgsgovbbsrawdata
ParmesanCampYoheG (2003)Agloballycoherentfingerprintofcli‐mate change impacts across natural systemsNature 421 37ndash42httpsdoiorg101038nature01286
PearsonRGampDawsonTP(2003)PredictingtheimpactsofclimatechangeonthedistributionofspeciesArebioclimateenvelopemod‐elsusefulGlobal Ecology and Biogeography12361ndash371
PlattJR(1964)StronginferenceScience146347ndash353PopperK(1959)The logic of scientific discoveryLondonUKHutchinsonRobbins C S Sauer J R Greenberg R S amp Droege S (1989)
Population declines in North American birds that migrate to theNeotropics Proceedings of the National Academy of Sciences 867658ndash7662httpsdoiorg101073pnas86197658
Robinson R A Wilson J D amp Crick H Q (2001) The impor‐tance of arable habitat for farmland birds in grassland land‐scapes Journal of Applied Ecology 38 1059ndash1069 httpsdoiorg101046j1365‐2664200100654x
Romesburg H C (1981) Wildlife science Gaining reliable knowl‐edge Journal of Wildlife Management 45 293ndash313 httpsdoiorg1023073807913
Root T (1988) Energy constraints on avian distributions and abun‐dancesEcology69330ndash339httpsdoiorg1023071940431
Ruiz‐GutieacuterrezVampZipkinEF (2011)Detectionbiasesyieldmis‐leadingpatternsof speciespersistence and colonization in frag‐mented landscapesEcosphere2 1ndash14 httpsdoiorg101890ES10‐002071
SauerJRHinesJEFallonJEPardieckKLZiolkowskiDJJrampLinkWA(2015)The North American breeding bird survey results and analysis 1966ndash2013 Version 01302015LaurelMDUSGSPatuxentWildlifeResearchCenterhttpswwwmbr‐pwrcusgsgovbbsbbshtml
Sauer J R LinkWA Fallon J E Pardieck K L amp ZiolkowskiDJJr(2013)TheNorthAmericanbreedingbirdsurvey1966ndash2011SummaryanalysisandspeciesaccountsNorth American Fauna791ndash32httpsdoiorg103996nafa790001
SeoaneJBustamanteJampDiacuteaz‐DelgadoR(2004)Competingrolesforlandscapevegetationtopographyandclimateinpredictivemod‐elsofbirddistributionEcological Modelling171209ndash222httpsdoiorg101016jecolmodel200308006
Sirami C Caplat P Popy S Clamens A Arlettaz R Jiguet F hellipMartin J‐L (2017) Impacts of global change on species distri‐butions Obstacles and solutions to integrate climate and landuse Global Ecology and Biogeography 26 385ndash394 httpsdoiorg101111geb12555
SkalskiJR(1996)Regressionofabundanceestimatesfrommarkrecap‐ture surveys against environmental covariatesCanadian Journal of
Fisheries and Aquatic Sciences53196ndash204httpsdoiorg101139f95‐169
SohlTL(2014)Therelativeimpactsofclimateandland‐usechangeonconterminousUnitedStatesbird species from2001 to2075PLoS ONE9e112251httpsdoiorg101371journalpone0112251
ThogmartinWESauerJRampKnutsonMG(2007)ModelingandmappingabundanceofAmericanwoodcockacrosstheMidwesternandNortheasternUnitedStatesJournal of Wildlife Management71376ndash382httpsdoiorg1021932005‐680
ThomasCDampLennonJJ (1999)Birdsextendtheirrangesnorth‐wardsNature399213httpsdoiorg10103820335
Trathan P N Garciacutea‐Borboroglu P Boersma D Bost C ACrawford R J CrossinG ThellipWienecke B (2015) Pollutionhabitatlossfishingandclimatechangeascriticalthreatstopen‐guins Conservation Biology 29 31ndash41 httpsdoiorg101111cobi12349
TravisJMJ(2003)Climatechangeandhabitatdestructionadeadlyanthropogenic cocktail Proceedings of the Royal Society of London Series B Biological Sciences 270(1514) 467ndash473 httpsdoiorg101098rspb20022246
UrbanMC (2015)Acceleratingextinction risk fromclimatechangeScience348571ndash573httpsdoiorg101126scienceaaa4984
WalshJWuebblesDHayhoeKKossinJKunkelKStephensGhellipSomervilleR(2014)OurchangingclimateInJMMelilloTCRichmondampGWYohe(Eds)Climate change impacts in the United States The Third National climate assessment(pp19ndash67)WashingtonDCUSGovernmentPrintingOffice
YackulicCBNichols JDReid JampDerR (2015)Topredict thenichemodelcolonizationandextinctionEcology9616ndash23httpsdoiorg10189014‐13611
Yalcin S amp Leroux S J (2018) An empirical test of the relative andcombinedeffectsofland‐coverandclimatechangeonlocalcoloni‐zationandextinctionGlobal Change Biology243849ndash3861httpsdoiorg101111gcb14169
ZhuKWoodallCWampClarkJS(2012)FailuretomigrateLackoftreerangeexpansioninresponsetoclimatechangeGlobal Change Biology181042ndash1052httpsdoiorg101111j1365‐2486201102571x
How to cite this articleClementMJNicholsJDCollazoJATerandoAJHinesJEWilliamsSGPartitioningglobalchangeAssessingtherelativeimportanceofchangesinclimateandlandcoverforchangesinaviandistributionEcol Evol 201991985ndash2003 httpsdoiorg101002ece34890

emspensp emsp | emsp2003CLEMENT ET aL
USGeologicalSurveyPatuxentWildlifeResearchCenterRetrievedfromhttpswwwpwrcusgsgovbbsrawdata
ParmesanCampYoheG (2003)Agloballycoherentfingerprintofcli‐mate change impacts across natural systemsNature 421 37ndash42httpsdoiorg101038nature01286
PearsonRGampDawsonTP(2003)PredictingtheimpactsofclimatechangeonthedistributionofspeciesArebioclimateenvelopemod‐elsusefulGlobal Ecology and Biogeography12361ndash371
PlattJR(1964)StronginferenceScience146347ndash353PopperK(1959)The logic of scientific discoveryLondonUKHutchinsonRobbins C S Sauer J R Greenberg R S amp Droege S (1989)
Population declines in North American birds that migrate to theNeotropics Proceedings of the National Academy of Sciences 867658ndash7662httpsdoiorg101073pnas86197658
Robinson R A Wilson J D amp Crick H Q (2001) The impor‐tance of arable habitat for farmland birds in grassland land‐scapes Journal of Applied Ecology 38 1059ndash1069 httpsdoiorg101046j1365‐2664200100654x
Romesburg H C (1981) Wildlife science Gaining reliable knowl‐edge Journal of Wildlife Management 45 293ndash313 httpsdoiorg1023073807913
Root T (1988) Energy constraints on avian distributions and abun‐dancesEcology69330ndash339httpsdoiorg1023071940431
Ruiz‐GutieacuterrezVampZipkinEF (2011)Detectionbiasesyieldmis‐leadingpatternsof speciespersistence and colonization in frag‐mented landscapesEcosphere2 1ndash14 httpsdoiorg101890ES10‐002071
SauerJRHinesJEFallonJEPardieckKLZiolkowskiDJJrampLinkWA(2015)The North American breeding bird survey results and analysis 1966ndash2013 Version 01302015LaurelMDUSGSPatuxentWildlifeResearchCenterhttpswwwmbr‐pwrcusgsgovbbsbbshtml
Sauer J R LinkWA Fallon J E Pardieck K L amp ZiolkowskiDJJr(2013)TheNorthAmericanbreedingbirdsurvey1966ndash2011SummaryanalysisandspeciesaccountsNorth American Fauna791ndash32httpsdoiorg103996nafa790001
SeoaneJBustamanteJampDiacuteaz‐DelgadoR(2004)Competingrolesforlandscapevegetationtopographyandclimateinpredictivemod‐elsofbirddistributionEcological Modelling171209ndash222httpsdoiorg101016jecolmodel200308006
Sirami C Caplat P Popy S Clamens A Arlettaz R Jiguet F hellipMartin J‐L (2017) Impacts of global change on species distri‐butions Obstacles and solutions to integrate climate and landuse Global Ecology and Biogeography 26 385ndash394 httpsdoiorg101111geb12555
SkalskiJR(1996)Regressionofabundanceestimatesfrommarkrecap‐ture surveys against environmental covariatesCanadian Journal of
Fisheries and Aquatic Sciences53196ndash204httpsdoiorg101139f95‐169
SohlTL(2014)Therelativeimpactsofclimateandland‐usechangeonconterminousUnitedStatesbird species from2001 to2075PLoS ONE9e112251httpsdoiorg101371journalpone0112251
ThogmartinWESauerJRampKnutsonMG(2007)ModelingandmappingabundanceofAmericanwoodcockacrosstheMidwesternandNortheasternUnitedStatesJournal of Wildlife Management71376ndash382httpsdoiorg1021932005‐680
ThomasCDampLennonJJ (1999)Birdsextendtheirrangesnorth‐wardsNature399213httpsdoiorg10103820335
Trathan P N Garciacutea‐Borboroglu P Boersma D Bost C ACrawford R J CrossinG ThellipWienecke B (2015) Pollutionhabitatlossfishingandclimatechangeascriticalthreatstopen‐guins Conservation Biology 29 31ndash41 httpsdoiorg101111cobi12349
TravisJMJ(2003)Climatechangeandhabitatdestructionadeadlyanthropogenic cocktail Proceedings of the Royal Society of London Series B Biological Sciences 270(1514) 467ndash473 httpsdoiorg101098rspb20022246
UrbanMC (2015)Acceleratingextinction risk fromclimatechangeScience348571ndash573httpsdoiorg101126scienceaaa4984
WalshJWuebblesDHayhoeKKossinJKunkelKStephensGhellipSomervilleR(2014)OurchangingclimateInJMMelilloTCRichmondampGWYohe(Eds)Climate change impacts in the United States The Third National climate assessment(pp19ndash67)WashingtonDCUSGovernmentPrintingOffice
YackulicCBNichols JDReid JampDerR (2015)Topredict thenichemodelcolonizationandextinctionEcology9616ndash23httpsdoiorg10189014‐13611
Yalcin S amp Leroux S J (2018) An empirical test of the relative andcombinedeffectsofland‐coverandclimatechangeonlocalcoloni‐zationandextinctionGlobal Change Biology243849ndash3861httpsdoiorg101111gcb14169
ZhuKWoodallCWampClarkJS(2012)FailuretomigrateLackoftreerangeexpansioninresponsetoclimatechangeGlobal Change Biology181042ndash1052httpsdoiorg101111j1365‐2486201102571x
How to cite this articleClementMJNicholsJDCollazoJATerandoAJHinesJEWilliamsSGPartitioningglobalchangeAssessingtherelativeimportanceofchangesinclimateandlandcoverforchangesinaviandistributionEcol Evol 201991985ndash2003 httpsdoiorg101002ece34890





























