Embed Size (px)
Citation preview

Part A
Fundamentals of Odor Sensing
Handbook of Machine Olfaction: Electronic Nose Technology.Edited by T.C. Pearce, S.S. Schiffman, H.T. Nagle, J.W. GardnerCopyright ª 2003 WILEY-VCH Verlag GmbH Co. KGaA, WeinheimISBN: 3-527-30358-8

1
Introduction to Olfaction: Perception, Anatomy, Physiology,
and Molecular Biology
Susan S. Schiffman, Tim C. Pearce
Abstract
Odors are sensations that occur when compounds (called odorants) stimulate recep-tors located in the olfactory epithelium at the roof of the nasal cavity. Odorants arehydrophobic, volatile compounds with a molecular weight of less than 300 daltons.Humans can recognize and distinguish up to 10 000 different substances on the basisof their odor quality. Odorant receptors (ORs) in the nasal cavity detect and discrimi-nate among these thousands of diverse chemical ligands. An individual odorant canbind to multiple receptor types, and structurally different odorants can bind to a singlereceptor. Specific patterns of activation generate signals that allow us to discriminatebetween the vast number of distinct smells. The physicochemical attributes of odor-ants that induce specific odor sensations are not well understood. The genes that codefor ORs have been cloned, and results from cloning studies indicate that ORs aremembers of a superfamily of hundreds of different G-protein-coupled receptorsthat possess seven transmembrane domains. A complete knowledge of structure-odor relationships in olfaction awaits the three-dimensional analysis of this large fa-mily of ORs. Ultimately, simultaneous knowledge of the three-dimensional structureof ORs as well as odorants will allow us to develop a pattern recognition paradigm thatcan predict odor quality.
1.1
Introduction to Olfaction
All living organisms from simple bacteria to complex mammals including humansrespond to chemicals in their environment. Chemical signals play a major role infeeding (e.g. nutrients), territorial recognition, sexual behavior, and detection of po-tentially harmful conditions such as fire, gas, and rancid food. In higher organisms,special chemical sensing systems (smell and taste) have developed that are distin-guished anatomically by the location of their receptors in the nasal and oral cav-ities, respectively. This chapter will focus on the nature of odors (sensations) and odor-ants (odorous molecules) that are relevant to human smell perception. The physiology
Handbook of Machine Olfaction: Electronic Nose Technology.Edited by T.C. Pearce, S.S. Schiffman, H.T. Nagle, J.W. GardnerCopyright ª 2003 WILEY-VCH Verlag GmbH & Co. KGaA, WeinheimISBN: 3-527-30358-8
11
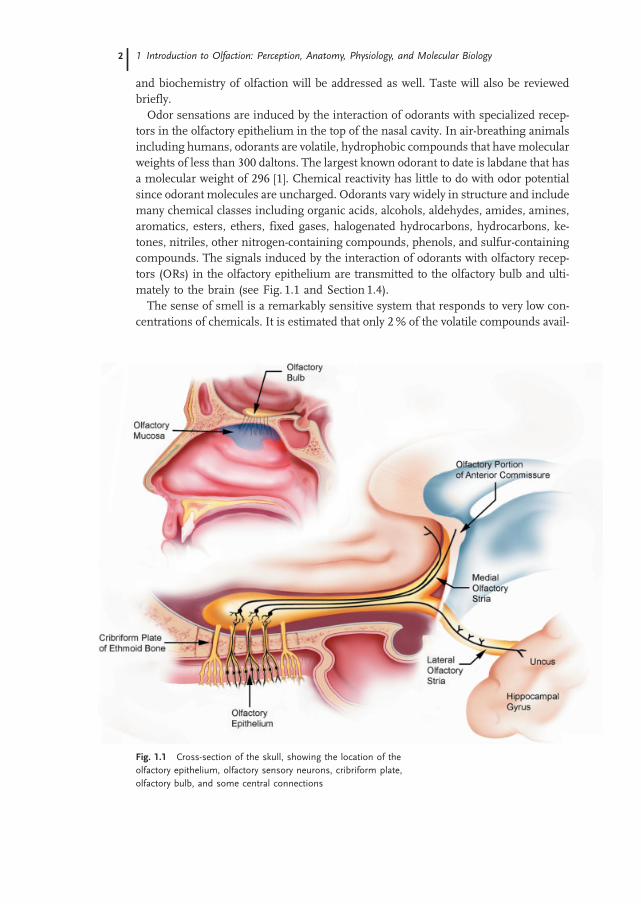
and biochemistry of olfaction will be addressed as well. Taste will also be reviewedbriefly.Odor sensations are induced by the interaction of odorants with specialized recep-
tors in the olfactory epithelium in the top of the nasal cavity. In air-breathing animalsincluding humans, odorants are volatile, hydrophobic compounds that have molecularweights of less than 300 daltons. The largest known odorant to date is labdane that hasa molecular weight of 296 [1]. Chemical reactivity has little to do with odor potentialsince odorant molecules are uncharged. Odorants vary widely in structure and includemany chemical classes including organic acids, alcohols, aldehydes, amides, amines,aromatics, esters, ethers, fixed gases, halogenated hydrocarbons, hydrocarbons, ke-tones, nitriles, other nitrogen-containing compounds, phenols, and sulfur-containingcompounds. The signals induced by the interaction of odorants with olfactory recep-tors (ORs) in the olfactory epithelium are transmitted to the olfactory bulb and ulti-mately to the brain (see Fig. 1.1 and Section 1.4).The sense of smell is a remarkably sensitive system that responds to very low con-
centrations of chemicals. It is estimated that only 2% of the volatile compounds avail-
Fig. 1.1 Cross-section of the skull, showing the location of the
olfactory epithelium, olfactory sensory neurons, cribriform plate,
olfactory bulb, and some central connections
1 Introduction to Olfaction: Perception, Anatomy, Physiology, and Molecular Biology2

able in a single sniff will reach the olfactory receptors, and as few as 40 molecules ofsome mercaptans are sufficient to perceive an odor [2, 3]. The exquisite sensitivity ofthe smell system is illustrated by the human detection thresholds given in Table 1.1(data from ref. [4]). It can be seen that these compounds can be detected at concentra-tions in the low parts-per-billion (ppb) and even low parts-per-trillion (ppt) range as inthe case of thiophenol, thiocresol, and propyl mercaptan. Over the course of a day,odorants have enormous opportunities to reach olfactory receptors during the processof inhalation and exhalation. An average person breathes 15 times per minute (or21 600 times per day) moving an average of 0.5 liters of air per breath (or 10 800 litersof air per day).Most odor sensations are produced by mixtures of hundreds of odorants rather than
by a single compound. Individual components tend to harmonize or blend together inmixtures leading to perceptual fusion. Humans have limited capacity to identify singleodorants in mixtures with three to four components being maximum [5]. This limita-tion in the ability to identify the individual components of mixtures appears to be aninherent property of olfaction since it is unrelated to the experience of the subjects orthe type of odorants.Odor sensations are characterized by general descriptors, such as sulfurous, fruity,
floral, and earthy, or by their source such as banana or orange. The range of distinctiveodor sensations is enormous, and a skilled perfume chemist can recognize and dis-tinguish 8000 to 10 000 different substances on the basis of their odor quality [6, 7] andeven respond to chemicals never before encountered in our environment. The olfac-tory system detects and discriminates among this immense number of odorant typesdue to the broad repertoire of olfactory receptor proteins that are encoded by a largeolfactory gene family [8–10] (see Section 1.5). Humans have several hundred distinctgenes that encode olfactory receptor proteins and rodents have upwards of 500 to 1000separate genes, that is, as much as 1% of the genome [9, 10]. This extremely broadrange of receptor types permits the detection of odor sources comprised of unpredict-able mixtures of molecular species, and even allows detection of newly synthesizedcompounds with no known function.
Table 1.1 Odor thresholds of representative sulfur compounds [44].
Compound Characteristic odor Odor Threshold
Allyl mercaptan Garlic-coffee 0.05 ppb
Amyl mercaptan Unpleasant strong 0.3 ppb
Benzyl mercaptan Unpleasant strong 0.19 ppb
Crotyl mercaptan Skunk-like 0.029 ppb
Dimethyl sulfide Decayed vegetables 0.1 ppb
Ethyl mercaptan Decayed cabbage 0.19 ppb
Hydrogen sulfide Rotten eggs 1.1 ppb
Methyl mercaptan Decayed cabbage 1.1 ppb
Propyl mercaptan Unpleasant 0.075 ppb
t-butyl mercaptan Skunk, unpleasant 0.08 ppb
Thiocresol Skunk, rancid 0.062 ppb
Thiophenol Putrid, garlic-like 0.062 ppb
1.1 Introduction to Olfaction 33

1.2
Odor Classification Schemes Based on Adjective Descriptors
Classification systems based on adjective descriptors have been used historically toorganize the many thousands of different odor sensations into a limited numberof groups. Table 1.2 shows some of the early schemes for classification of odor sensa-tions. Modern olfactory specialists including perfumers who work with fragrances,however, find the small number of categories of early classification schemes to beinadequate for describing the odors that they encounter in their work. Over thelast half century there has been a movement away from trying to classify odors ina few limited classes but rather to develop an extensive vocabulary that is relevantfor use with the odor stimuli being examined. Hence, modern odor classificationmethods are based on an extensive array of adjective descriptors selected for theirrelevance for specific applications.Modern descriptive classificationmethods can be general (e.g. for the broad range of
odors encountered in everyday life) or specific (e.g. relevant to particular applicationsin the food or fragrance industry). In the food industry, the odors of chemical com-pounds are often categorized by the identity of the edible material of which they arereminiscent. Sample odor classes for foods include caramel, honey, vanilla, citrus, andbutter. Fragrance odors are often classified by floral and herbal groupings, such asjasmine, rose, balsam, or pine. Table 1.3 presents a series of 146 adjective descriptorsdeveloped by the American Society for Testing and Materials [20] for general classi-fication of odors commonly encountered in everyday life. Table 1.4 gives a more spe-cific list of descriptors used by the fragrance industry [21]. Other odor descriptors canbe found in flavor and fragrance catalogs (Aldrich [22], for example) as well as ontechnical web sites (for example, ref. [23]).
Table 1.2 Descriptive categories proposed historically for smell
sensations.
Number of Categories Category Classification Reference
6 Sweet, acid (sour), harsh, rich/fat, astringent, fetid 11
7 Aromatic, fragrant, ambrosial (musk-like), alliaceous
(garlic-like), hircine (goat-like), foul, nauseating
12
9 Aromatic, ethereal, fragrant, ambrosial, empyreumatic
(burnt), alliaceous, hircine, repulsive, nauseous
13
6 Flowery, fruity, spicy, resinous, burnt, putrid 14
8 Flowery, fruity, herbaceous (green), animal/ambrosial/
human flesh aura, spicy/minty/camphoric, earthy/
fungoid, woody/balsamic/nut-like, Disagreeable: acrid/
phenolic/burnt/nauseating
15
7 Ethereal, floral, pepperminty, camphoraceous, musky,
pungent, putrid
16–18
9 Etherish, fragrant, sweet, spicy, oily, burnt, sulfurous,
rancid, metallic
19
1 Introduction to Olfaction: Perception, Anatomy, Physiology, and Molecular Biology4

Table 1.3 ASTM descriptive categories used for general odor quality
characterizations.a)
(01) Fragrant (50) Vanilla-like (99) Alcohol-like
(02) Sweaty (51) Fecal (like manure) (100) Dill-like
(03) Almond-like (52) Floral (101) Chemical
(04) Burnt, smoky (53) Yeasty (102) Creosote
(05) Herbal, green, cut grass (54) Cheesy (103) Green pepper
(06) Etherish, anesthetic (55) Honey-like (104) Household gas
(07) Sour, acid, vinegar (56) Anise (licorice) (105) Peanut butter
(08) Like blood, raw meat (57) Turpentine (pine oil) (106) Violets
(09) Dry, powdery (58) Fresh green vegetables (107) Tea-leaves-like
(10) Like ammonia (59) Medicinal (108) Strawberry-like
(11) Disinfectant, carbolic (60) Orange (fruit) (109) Stale
(12) Aromatic (61) Buttery (110) Cork-like
(13) Meaty (cooked) (62) Like burnt paper (111) Lavender
(14) Sickening (63) Cologne (112) Cat-urine-like
(15) Mushy, earthy, moldy (64) Caraway (113) Pineapple (fruit)
(16) Sharp, pungent, acid (65) Bark-like, birch bark (114) Fresh tobacco smoke
(17) Camphor-like (66) Rose-like (115) Nutty
(18) Light (67) Celery (116) Fried fat
(19) Heavy (68) Burnt candle (117) Wet paper-like
(20) Cool, cooling (69) Mushroom-like (118) Coffee-like
(21) Warm (70) Wet wool, wet dog (119) Peach (fruit)
(22) Metallic (71) Chalky (120) Laurel leaves
(23) Perfumery (72) Leather-like (121) Scorched milk
(24) Malty (73) Pear (fruit) (122) Sewer odor
(25) Cinnamon (74) Stale tobacco smoke (123) Sooty
(26) Popcorn (75) Raw cucumber-like (124) Crushed weeds
(27) Incense (76) Raw potato-like (125) Rubbery (new rubber)
(28) Cantalope, honey dew melon (77) Mouse-like (126) Bakery, fresh bread
(29) Tar-like (78) Black pepper-like (127) Oak wood, cognac-like
(30) Eucalyptus (79) Bean-like (128) Grapefruit
(31) Oily, fatty (80) Banana-like (129) Grape-juice-like
(32) Like mothballs (81) Burnt rubber-like (130) Eggy (fresh eggs)
(33) Like gasoline, solvent (82) Geranium leaves (131) Bitter
(34) Cooked vegetables (83) Urine-like (132) Cadaverous, dead animal
(35) Sweet (84) Beery (beer-like) (133) Maple (syrup)
(36) Fishy (85) Cedar wood-like (134) Seasoning (for meat)
(37) Spicy (86) Coconut-like (135) Apple (fruit)
(38) Paint-like (87) Rope-like (136) Soupy
(39) Rancid (88) Seminal, sperm-like (137) Grainy (as grain)
(40) Minty, peppermint (89) Like cleaning fluid (138) Clove-like
(41) Sulphidic (90) Cardboard-like (139) Raisins
(42) Fruity (citrus) (91) Lemon (fruit) (140) Hay
(43) Fruity (other) (92) Dirty linen-like (141) Kerosene
(44) Putrid, foul, decayed (93) Kippery (smoked fish) (142) Nail polish remover
(45) Woody, resinous (94) Caramel (143) Fermented fruit
(46) Musk-like (95) Sauerkraut-like (144) Cherry (berry)
(47) Soapy (96) Crushed grass (145) Varnish
(48) Garlic, onion (97) Chocolate (146) Sour milk
(49) Animal (98) Molasses
a) Odor Quality characterizations. Each sample israted on 146 adjectives using a five-point scale
where 0 indicates no odor and 5 indicates extre-mely strong odor.
1.2 Odor Classification Schemes Based on Adjective Descriptors 55

Table 1.4 Odor descriptors in Allured’s Perfumer’s Compendium [21]
used by perfumers and flavorists.
(1) Agrumen (50) Gardenia (99) Ozone (fresh air, marine)
(2) Aldehydic (51) Geranium (100) Patchouli
(3) Almond (52) Ginger (101) Peach
(4) Amber (53) Grape (102) Pear
(5) Ambergris (54) Grapefruit (103) Pepper
(6) Animal (55) Grass (104) Peppermint
(7) Anisic (56) Green (105) Petal
(8) Apple Blossom (57) Hawthorne (106) Petitgrain
(9) Apple Fruity (58) Hay (107) Pimento
(10) Armoise (59) Herbal (108) Pine
(11) Balsamic (60) Honey (109) Pineapple
(12) Banana (61) Honeysuckle (110) Plum
(13) Basil (62) Hyacinth (111) Powdery
(14) Bay (63) Incense (112) Raspberry
(15) Bergamot (64) Jasmin (113) Rooty
(16) Camphoraceous (65) Juicy (114) Rose
(17) Cardamom (66) Juniper (115) Sage
(18) Carnation (67) Kiwi (116) Sandalwood
(19) Cassie (68) Labdanum (117) Sappy-green wood
(20) Cassis (69) Lactonic (118) Smokey
(21) Castoreum (70) Lavender (119) Spicy
(22) Cedar (71) Leafy (120) Strawberry
(23) Celery (72) Leather (121) Styrax
(24) Chamomile (73) Lemon (122) Sweet
(25) Cherry (74) Lilac (123) Sweet pea
(26) Chocolate (75) Lime (124) Tagette
(27) Chrysanthemum (76) Mandarin (125) Tangerine
(28) Cinnamon (77) Medicated (126) Tea
(29) Citrus (78) Melon (127) Thyme
(30) Civet (79) Metallic (128) Tobacco
(31) Clary sage (80) Mimosa (129) Tolu
(32) Clove (81) Minty (130) Tonka
(33) Coconut (82) Moss (131) Tuberose
(34) Cognac (83) Muguet (132) Vanilla
(35) Coriander (84) Mushroom (133) Verbena
(36) Costus (85) Musk (134) Vetivert
(37) Cumin (86) Myrrh (135) Violet
(38) Dry (87) Narcisse (136) Waxy
(39) Earthy (88) Nasturtium (137) Wintergreen
(40) Eucalyptus (89) Neroli (138) Woody
(41) Fatty (90) Nutmeg (139) Ylang
(42) Fecal-animal (91) Nutty (140) Zesty, peely (citrus)
(43) Fig (92) Oily
(44) Floral (93) Olibanum
(45) Floral bouquet (94) Opoponax
(46) Fougere (95) Orange flower
(47) Freesia (96) Orange fruit
(48) Fruity (97) Oriental
(49) Galbanum (98) Orris
1 Introduction to Olfaction: Perception, Anatomy, Physiology, and Molecular Biology6

Classification schemes for odor quality are beset, however, by a variety of limitations.First there are inherent interindividual differences in the emotional and hedonic prop-erties of odors. Labels such as pleasant, delightful, disgusting, and revolting are com-mon associations with odors, and these subjective evaluations can influence the choiceof descriptors of odor quality. Emotional responses to odors probably derive from thefact that olfaction is a primal sense that is used in the animal kingdom to identify food,mates, predators, and warnings of danger. Second, there are individual differences inthe actual perception of odor based on genetic differences [24–26]. Third, there areindividual differences in the use of odor descriptors even among trained panelists.Fourth, the vocabulary of most languages lacks words that describe the full rangeof odor sensations. For this last reason, measures of similarity rather than adjectivedescriptors have been used to quantify odor quality by arranging odor sensations inmultidimensional spaces (to be described in the next section).
1.3
Odor Classification Based on Chemical Properties
Although much progress has been made in our knowledge of olfactory physiology andbiochemistry, the fundamental relationship between odor quality and molecular prop-erties is still poorly understood. Even slight alterations in the chemical structure of anodorant can induce profound changes in odor quality. Current structure-activity mod-els in olfaction are, for the most part, simply collections of disparate facts with nounifying theme; furthermore, they have inadequate predictive accuracy [27]. As a con-sequence, the basic logic necessary to develop a comprehensive odor classificationscheme based on particular features of molecules remains elusive.There are several reasons for the lack of progress in classifying odors on the basis of
chemical properties. First, it is not yet possible to model odorant-receptor interactionsbecause the three-dimensional (3D) protein structures of the receptor sites are notknown. Second, unlike structure-activity counterparts in pharmacology, there arevast numbers of agonist types (thousands of odorant structures) as well as thousandsof different odor sensations. Third, identical molecules may activate different receptortypes depending on the orientation of the molecule at the receptor. Beets [6] empha-sized that identical molecules arrive near receptor sites at different orientations andwith different conformations. Thus, a given odorant would be expected to interact witha variety of receptor types, and odor quality must be encoded by a pattern of informa-tion frommultiple receptors (rather than activation of a single receptor type). A fourthproblem is that there are no standard methods for quantifying odor quality for use instructure-activity studies.
1.3 Odor Classification Based on Chemical Properties 77

1.3.1
History of Structure-activity Studies of Olfaction
In spite of these limitations, a variety of attempts have been made to relate odors tophysicochemical parameters. Amoore [16] proposed that the shape and size of a mo-lecule are the physicochemical parameters that determine odor quality, i.e. odorants fitinto receptor sites in a ‘lock and key’ fashion with molecules of similar size and shapeexpected to have similar odor quality. In support of this theory, Amoore and Venstrom[28] reported significant correlations between odor quality of 107 odorants and a hand-calculated index of molecular size and shape for five classes of odors (ethereal, cam-phoraceous, musky, floral, and minty). Amoore [29] also reported a correlation of 0.90between odor quality and a computer-generated molecular shape index when 25 sub-stances were compared with benzaldehyde (almond odor).Wright [30] challenged Amoore’s results, indicating that it is inappropriate to repre-
sent a complex 3Dmolecular shape by an index consisting of a single number becausemany different 3D profiles could share the same molecular shape index. Wright [31]suggested that the mechanism for stimulation of olfactory receptors is low-energymolecular vibrations, and that molecules with similar vibrational frequency patternsshould have similar odor quality. Wright and Robson [32] supported their hypothesiswith the finding of similarity between the pattern of frequencies in the far infraredspectra for odorants with a bitter almond odor.Dravnieks and Laffort [33] suggested that four factors related to intermolecular
interaction forces (an apolar factor, a proton receptor factor, an electron factor, and aproton donor factor) could predict both quantitative and qualitative odor discrimina-tion in human beings. In spite of many attempts in addition to those just described, nogeneral structure-activity model or theory has yet been proposed that accurately pre-dicts odor quality of molecules a priori from physicochemical parameters[1, 6, 27, 34, 35].
1.3.2
Odor Structures Associated with Specific Odor Classes Based on Qualitative Descriptors
Figures 1.2 to 1.6 provide examples of chemical structures for compounds classified byexperienced odor specialists as having musk, ambergris, muguet, green, and bitteralmond odors. Each figure gives the structure of representative chemicals withineach specific odor quality. These figures illustrate that compounds with widely vary-ing chemical structures can have similar odor qualities. Musk is an odor category thatis used in fragrance with its original source being the glandular secretions of the malemusk deer. Molecules with this odor quality are very diverse in structure as shown inFig. 1.2; they include steroidal, linear, macrocyclic, nitro, as well as bi- and tricycliccompounds. Ambergris is an odor quality used in fragrance that originally derivedfrom the sperm or cachalot whale. Muguet is a lily-of-valley odor. Green is theodor of natural green vegetable products. Bitter almond is an odor quality of an es-sential oil obtained by stem distillation of kernals from bitter almond (P. amygda-lus). The types ofmolecules within each odor quality can vary considerably in structure.
1 Introduction to Olfaction: Perception, Anatomy, Physiology, and Molecular Biology8

Studies of enantiomers have also been used to gain insight into the relationshipbetween physicochemical properties and odor quality. These studies reveal that enan-tiomers of chiral odorants may or may not show differences in odor quality[1, 6, 27, 34, 35]. There are significant differences in the two enantiomers of carvonewith R-(�)-carvone having an odor of spearmint oil and S-(þ)-carvone having an odorof caraway oil (see Fig. 1.7). Significant differences in the odor quality of enantiomersof nootkatone have also been reported. However, enantiomers of 2-octanol and carbi-naol were not found to differ in odor quality.Overall, Figs. 1.2–1.7 demonstrate that structurally unrelated chemicals can yield
similar odor qualities. Furthermore, differences in the odor quality of certain enan-tiomers indicate that very subtle differences in structure are capable of producing verydifferent and distinct odors. A better understanding of the physicochemical para-
Fig. 1.2 Compounds with musk odor: a) androst-16-en-3b-ol,b) ethyl citronellyl oxalate, c) cyclopentadecanolide, d) musk ketone,
e) Traesolide�, f) Galaxolide
1.3 Odor Classification Based on Chemical Properties 99
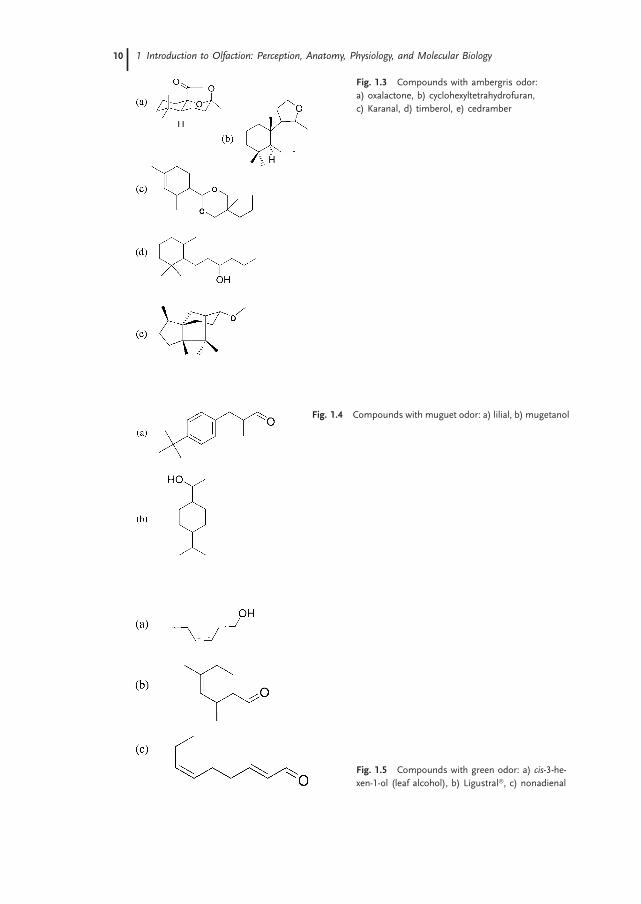
Fig. 1.3 Compounds with ambergris odor:
a) oxalactone, b) cyclohexyltetrahydrofuran,
c) Karanal, d) timberol, e) cedramber
Fig. 1.4 Compounds with muguet odor: a) lilial, b) mugetanol
Fig. 1.5 Compounds with green odor: a) cis-3-he-
xen-1-ol (leaf alcohol), b) Ligustral�, c) nonadienal
1 Introduction to Olfaction: Perception, Anatomy, Physiology, and Molecular Biology10

meters responsible for specific odor qualities requires more knowledge about the 3Dstructure of ORs.
1.3.3
Relationship of Physicochemical Parameters to Classifications of Odor Based on SimilarityMeasures
Themethodology of multidimensional scaling has also been used to better understandthe relationship between odor quality and physicochemical variables [36, 37]. Multi-dimensional scaling (MDS) procedures represent odor sensations in spatial maps.The input for multidimensional scaling procedures consists of quantitative measuresof similarity between pairs of odors. For example, if two odors are judged by humansubjects to have similar odor quality, they will be positioned near each other in themultidimensional quality space. Stimuli judged to be dissimilar are located distantfrom one another. Two examples of studies that relate physicochemical propertiesto odor quality as defined by multidimensional maps are given below. The mathema-
Fig. 1.6 Compounds with bitter almond odor: a) benzaldehyde,
b) hydrogen cyanide
Fig. 1.7 Enantiomers of carvone. a) R-(�) carvone which has a
spearmint-like odor, b) S-(þ) carvone which has a caraway-like odor
1.3 Odor Classification Based on Chemical Properties 1111

tical procedures used to relate the physicochemical properties to themultidimensionalmaps are given in the Appendix.
1.3.3.1 Study 1: Broad Range of Unrelated Odorants
A group of 50 odorants (5 of which were duplications) that ranged widely in quality andstructure were arranged in a two-dimensional (2D) space by MDS on the basis of odorsimilarity [38]. The 2D space that is shown in Fig. 1.8 accounts for 91% of the humansimilarity data. The odor stimuli were roughly positioned by MDS into two groups; thelarger subset on the left is affectively more pleasant than the one on the right.Because the spatial arrangement could not be accounted for by a single physico-
chemical variable (such as chemical structure, molecular weight, number of doublebonds, or dipole moment), a series of physicochemical variables were weighted in anattempt to regenerate the space. A mathematical technique generated weights for aseries of physicochemical variables such that the distances and thus the spatial ar-rangements among the stimuli in Fig. 1.8 were regenerated.The mathematical procedure used to maximize the configurational similarity of the
psychologically determined space in Fig. 1.8, with a space generated by weighted phy-sicochemical parameters was based on a least-squares method (see Appendix). Thephysicochemical parameters that were weighted to reconstruct the 2D space inFig. 1.8 as well as the means for these physicochemical variables are shown in Ta-ble 1.5. Functional groups were coded according to their number in a particular mo-lecule; for example, benzaldehyde has one aldehyde group and the mean number of
Fig. 1.8 Two-dimensional solution for a broad range of odor stimuli.
Compounds with similar odor qualities are located near each other in
space. The more pleasant stimuli are located in the subset to the left,
and the more unpleasant stimuli are in the subset on the right
(modified after Schiffman [38])
1 Introduction to Olfaction: Perception, Anatomy, Physiology, and Molecular Biology12
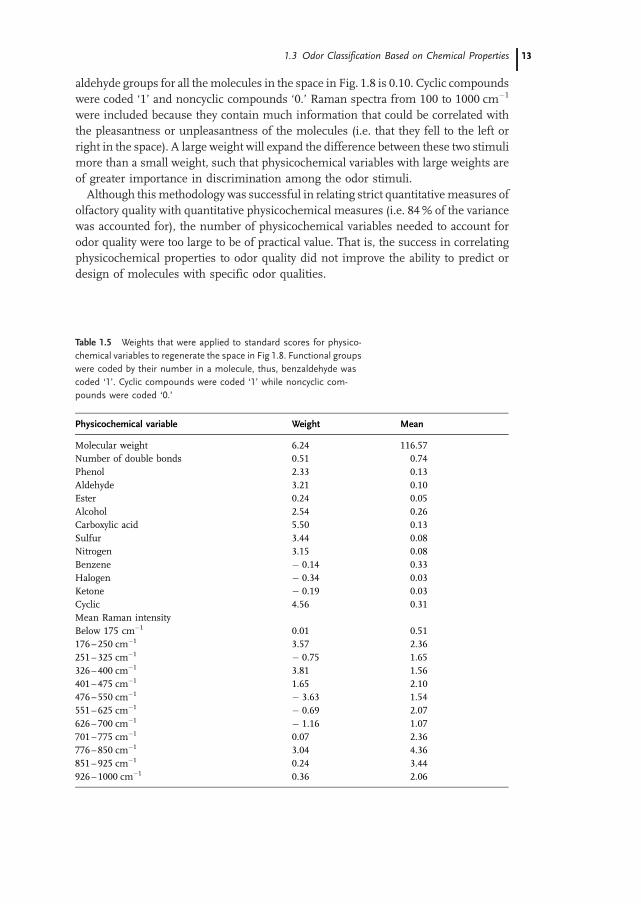
aldehyde groups for all themolecules in the space in Fig. 1.8 is 0.10. Cyclic compoundswere coded ‘1’ and noncyclic compounds ‘0.’ Raman spectra from 100 to 1000 cm�1
were included because they contain much information that could be correlated withthe pleasantness or unpleasantness of the molecules (i.e. that they fell to the left orright in the space). A large weight will expand the difference between these two stimulimore than a small weight, such that physicochemical variables with large weights areof greater importance in discrimination among the odor stimuli.Although thismethodology was successful in relating strict quantitative measures of
olfactory quality with quantitative physicochemical measures (i.e. 84% of the variancewas accounted for), the number of physicochemical variables needed to account forodor quality were too large to be of practical value. That is, the success in correlatingphysicochemical properties to odor quality did not improve the ability to predict ordesign of molecules with specific odor qualities.
Table 1.5 Weights that were applied to standard scores for physico-
chemical variables to regenerate the space in Fig 1.8. Functional groups
were coded by their number in a molecule, thus, benzaldehyde was
coded ‘1’. Cyclic compounds were coded ‘1’ while noncyclic com-
pounds were coded ‘0.’
Physicochemical variable Weight Mean
Molecular weight 6.24 116.57
Number of double bonds 0.51 0.74
Phenol 2.33 0.13
Aldehyde 3.21 0.10
Ester 0.24 0.05
Alcohol 2.54 0.26
Carboxylic acid 5.50 0.13
Sulfur 3.44 0.08
Nitrogen 3.15 0.08
Benzene � 0.14 0.33
Halogen � 0.34 0.03
Ketone � 0.19 0.03
Cyclic 4.56 0.31
Mean Raman intensity
Below 175 cm�1 0.01 0.51
176–250 cm�1 3.57 2.36
251–325 cm�1 � 0.75 1.65
326–400 cm�1 3.81 1.56
401–475 cm�1 1.65 2.10
476–550 cm�1 � 3.63 1.54
551–625 cm�1 � 0.69 2.07
626–700 cm�1 � 1.16 1.07
701–775 cm�1 0.07 2.36
776–850 cm�1 3.04 4.36
851–925 cm�1 0.24 3.44
926–1000 cm�1 0.36 2.06
1.3 Odor Classification Based on Chemical Properties 1313

1.3.3.2 Study 2: Pyrazines
Multiple physicochemical parameters were also necessary to account for an odor spacerepresenting the similarity among related compounds (pyrazines). The pyrazines wereordered in the 3D space in Fig. 1.9 on the basis of similarity of odor quality [39]. Next, aset of descriptors (see Table 1.6) was generated by the automated data analysis andpattern recognition toolkit (ADAPT), a computer system for automated data analysesby pattern recognition techniques [40, 41]. The substructures used to generate theenvironment descriptors are given in Fig. 1.10.Canonical regression, a common statistical technique [42], was used to relate the
descriptors in Table 1.6 to the 3D arrangement in Fig. 1.9. Canonical analysis extendsmultiple regression analysis from one criterion variable to a set of criterion variables.For simple multiple regression, the relationship of a set of predictors to a single cri-terion variable is analyzed. In the current application, canonical regression was used todetermine the relationships between two sets of variables, that is, the stimulus coor-
Table 1.6 Descriptors generated by ADAPT [40] for analysis of
pyrazines.
1 Number of atoms except hydrogen
2 Number of carbon atoms
3 Number of oxygen atoms
4 Number of bonds
5 Number of single bonds
6 Number of double bonds
7 Molecular weight
8 Path 1 molecular connectivity for all bonds in the structure
9 Path 1 molecular connectivity corrected for rings
10 Path 1molecular connectivity calculated using the valences of heteroatoms and corrected for rings
11 Path 2 molecular connectivity
12 Path 3 molecular connectivity
13 Path 4 molecular connectivity
14 Molecular volume
15 Number of substructure 1 (see Fig. 1.11)
16 Environment-substructure 1 (calculates connectivity for substructure 1 and nearest neighbors)
17 Number of substructure 2
18 Environment-substructure 2
19 Number of substructure 3
20 Environment-substructure 3
21 Number of substructure 4
22 Environment-substructure 4
23 Number of substructure 5
24 Environment-substructure 5
25 Number of substructure 6
26 Environment-substructure 6
27 Number of substructure 7
28 Environment-substructure 7
29 Number of substructure 8
30 Environment-substructure 8
1 Introduction to Olfaction: Perception, Anatomy, Physiology, and Molecular Biology14
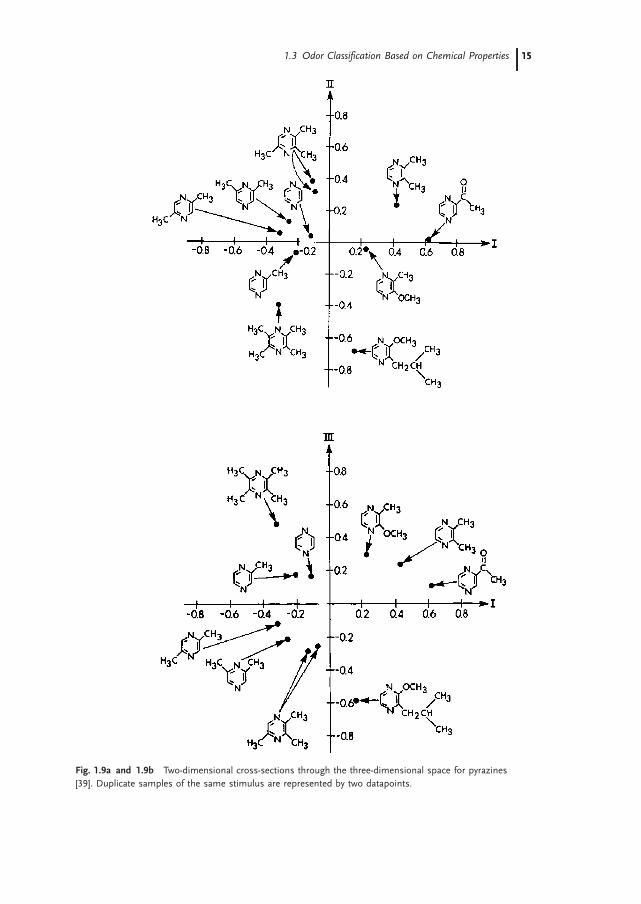
Fig. 1.9a and 1.9b Two-dimensional cross-sections through the three-dimensional space for pyrazines
[39]. Duplicate samples of the same stimulus are represented by two datapoints.
1.3 Odor Classification Based on Chemical Properties 1515

dinates of the 3D MDS space and the physicochemical parameters in Table 1.6. Theequations for canonical correlation used are given in the Appendix.Small subsets of the physicochemical parameters were used in the tests because in
canonical correlation analysis, the number of stimuli should be greater than the num-ber of dimensions and physicochemical parameters combined. The analysis revealedthat a linear combination of two ADAPT parameters in Table 1.6 (number of oxygenatoms and chemical environment of substructure 7) in addition to a concentrationvariable accounted for 63% of the arrangement of the pyrazine odor space inFig. 1.9. This study, along with Study 1, again illustrates the difficulty in relating quan-titative physicochemical parameters with odor quality.
1.3.4
Molecular Parameters and Odor Thresholds
In addition to odor quality, attempts have been made to determine the relationshipbetween odor thresholds (or suprathreshold intensity) and molecular parameters.Variables that have been related to thresholds and intensities include molecularweight, cross-sectional area, adsorption constants at an oil-water interface, hydropho-bicity, molar volume, pKa, saturated vapor pressure, polarizability, hydrogen bondingability, air/water partition coefficients, log P (octanol-water partition coefficient), para-meters derived from gas chromatograpy, Taft polar constants, and various steric para-meters [34]). Like structure-activity studies of odor quality, there appear to be no rulesthat can be generalized for the entire range of odorous compounds.
1.3.5
Conclusions Regarding Physicochemical Parameters and Odor Quality
Although it is possible to develop techniques that weight a series of parameters topredict odor quality, this is of little practical use in understanding the physiological
Fig. 1.10 The substructures utilized by ADAPT for generating
environment descriptors for analysis of pyrazines
1 Introduction to Olfaction: Perception, Anatomy, Physiology, and Molecular Biology16

basis of odor quality. A more complete understanding of structure-activity relation-ships in olfaction will occur when the molecular structure of the odorant receptor(including the stereoelectronic arrangements of binding sites) is brought into theequation along with the structure of the odorant.
1.4
Physiology and Anatomy of Olfaction
1.4.1
Basic Anatomy
The functional organization of the olfactory system is similar to other sensory systems(e.g. vision) but, in this case, the sensory input is provided bymolecules (i.e. odorants).Odorants are recognized by specific receptor proteins situated on the ciliary mem-branes of olfactory sensory neurons located in the olfactory epithelium at the topof the nasal cavity (see Fig. 1.1). The olfactory epithelium is comprised of three celltypes as shown in Fig. 1.1: the bipolar olfactory sensory neurons (primary sensoryneurons) with dendritic cilia projecting from their terminal ends in a thin mucuslayer (10–100 lM thick); supporting or sustentacular cells (a type of glial cell) thatterminate in microvilli; and basal cells (like stem cells) which make new olfactoryreceptor cells.The olfactory epithelium is a thin tissue in the nasal cavity that is easily distinguish-
able bilaterally in rats and dogs due to its yellowish color. In humans, however, the twosmall patches (about 2 square inches or 6.5 square centimeters) are more difficult tovisualize because their pinkish hue blends with the respiratory epithelium that lines
Fig. 1.11 Olfactory epithelium showing three cell
types: olfactory sensory neurons (also called receptor
cells), supporting (or sustenacular cells), and basal
cells
1.4 Physiology and Anatomy of Olfaction 1717

the rest of the nasal cavity. Odorants can reach the olfactory receptors via orthonasaltransport through the nares (e.g. when sniffing) or via retronasal transport from theoral cavity (e.g. when chewing food). In orthonasal transport, the turbinates (bones inthe nose) create turbulent airflow patterns that direct volatile compounds to the olfac-tory receptor cells in the top of the nasal cavity. Inhaled odorants have been shown toreach the blood and brain after three hours of exposure [43], and as a consequence,olfactory receptors can also respond to blood-borne odorants [44].There are vast numbers of olfactory sensory neurons with estimates between 106 and
108 in man [45, 46]. These olfactory neurons turn over continuously with an averagetime for replacement of approximately 30 days. This neurogenesis is active throughoutthe lifespan, and arises from basal cells deep in the epithelium [47]. Olfactory sensoryneurons consist structurally of a soma (cell body), a peripheral dendritic knob withfine, long cilia that project into the watery mucus that protects the nasal cavity,and an unmyelinated axon that projects centrally from the soma and propagates actionpotentials to the olfactory bulb. Specific receptor subtypes are expressed in subsets ofolfactory sensory neurons spatially distributed in distinct zones of the olfactory epithe-lium, and only one odorant receptor type is expressed on the vast majority of individualolfactory sensory neurons [9, 10]. Yet, single olfactory cells respond to a range of com-pounds with a variety of olfactory qualities because individual olfactory receptors haverelatively broad molecular receptive ranges [48].Axons of the bipolar olfactory sensory neurons fasiculate together and coarse
through tiny holes in the cribriform plate of the ethmoid bone to the olfactorybulb where they make their first synapses with second-order neurons in intricate sphe-rical masses of neuropil called glomeruli (see Fig. 1.12). The axons of the bipolar cellsconstitute the fibers of the olfactory nerve. The neuropil of glomeruli consists of theaxons of incoming olfactory sensory neurons and the dendrites of the mitral cells onwhich they synapse. Olfactory sensory neurons that express a specific odor receptortype converge upon a common glomerulus in the olfactory bulb [9, 10, 49]. In humans,axons from thousands of olfactory sensory neurons expressing a single odorant recep-tor type are thought to converge onto two or three glomeruli in the olfactory bulb, witheach glomerulus receiving input from a single type of olfactory receptor. Local neu-ronal circuits in the bulb provide the first tier of central processing of odors with ol-factory signals sharpened via lateral inhibition among glomerular modules [50]. As aresult of this neural processing, mitral cells have narrower molecular receptive rangesthan olfactory receptor neurons [48]. Because individual olfactory sensory neurons canrespond to multiple odorants, it follows that the pattern across multiple glomeruliprovides the basis for discrimination of olfactory quality. The distinct spatial patternsof glomerular activation by specific odorants can be visualized using optical imagingtechniques [51, 52].Olfactory information from the olfactory bulb is next transmitted by the olfactory
tract to the anterior olfactory nucleus, the olfactory tubercle, the prepyriform cor-tex, and the amygdala, and ultimately to higher brain centers that process the olfactorysignals. The prepyriform cortex and the amygdala are brain structures that are part ofthe limbic system, which processes emotions and memories in addition to olfactorysignals. Olfactory information is ultimately transmitted to the hypothalamus (which
1 Introduction to Olfaction: Perception, Anatomy, Physiology, and Molecular Biology18

mediates food intake) and to the neocortex. Non-invasive imaging techniques such aselectroencephalography, positron emission tomography, and functional magnetic re-sonance spectrometry have found that the degree of activation of the pyriform cortex,orbitofrontal areas, and parts of the parietal and temporal cortices is dependent on theodor quality and pleasantness of the stimuli (for example see refs. [53] and [54]). Age-related losses occur in the olfactory epithelium, olfactory bulb and nerves, hippocam-pus and amygdaloid complex, and hypothalamus, and these changes parallel percep-tual losses in the olfactory system during the aging process.At elevated concentrations, odorants can also stimulate free nerve endings of the
trigeminal nerve in the nose. Trigeminal stimulation by odorous chemicals inducessensations such as irritation, tickling, burning, stinging, scratching, prickling, anditching [55, 56]. Sensory information transmitted by the trigeminal nerve is not con-sidered an ‘odor’ because the trigeminal nerve is not directly stimulated by electricalsignals from olfactory receptor neurons; rather trigeminal stimulation involves a dif-ferent sense called chemesthesis which is related to nociception (e.g. pain).
Fig. 1.12 Cross-section of the
olfactory bulb. A.C. indicates
anterior commissure.
1.4 Physiology and Anatomy of Olfaction 1919

1.4.2
Transduction and Adaptation of Olfactory Signals
Odorants first traverse the aqueous interphase that lines the surface of the olfactoryepithelium in order to interact with the olfactory receptors in the ciliary membranes.This process is facilitated by soluble odorant binding proteins that ‘shuttle’ the hydro-phobic odorants through the aqueous mucus layer towards specific odorant receptors.Odorant receptors are members of a superfamily of up to 1000 different G-protein-coupled receptors that possess seven transmembrane (7TM) domains. The location ofodorant binding is thought to be a hydrophobic pocket in transmembrane regions 3, 4,and 5 of the seven-membrane-spanning receptor. Olfactory signaling is initiated by theinteraction of an odorant molecule with a protein receptor on the ciliary surface. Thisligand binding triggersmultistep intracellular reaction cascades that open an ion chan-nel in the cell membrane leading to depolarization by a few tens of millivolts [8, 57].Figure 1.13 illustrates the binding of an odorant to a G-protein-coupled receptor inolfactory neurons that results in activation of cAMP. Odorant binding involves a sig-naling pathway that includes a Gs-like protein (Golf) that activates a specific adenylylcyclase leading to generation of cyclic AMP (cAMP). cAMP binds directly to a cyclicnucleotide-gated (CNG) ion channel in the cell membrane that increases the probabil-ity of positive ions flowing into the cell, leading to depolarization and action potentials.
Figure 1.13 a. and b. In most olfactory neurons, an odorant binds to
an odorant receptor (OR) leading to an exchange of GTP (guanosine
triphosphate) for GDP (guanosine diphosphate) on the heterotrimeric
G-protein (Golf ). c. The a subunit of Golf activates adenylyl cyclase
leading to generation of cAMP. d. Cyclic AMP binds directly to a cyclic
nucleotide-gated (CNG) ion channel in the cell membrane that in-
creases the probability of positive ions flowing into the cell. This causes
depolarization of the cell membrane and transmission of a signal along
the axon to the bulb. CNG channels are nonselective and permeable to
cations including Naþ and Ca2þ
1 Introduction to Olfaction: Perception, Anatomy, Physiology, and Molecular Biology20

Another intracellular second messenger, inositol triphosphate may also mediatechanges in the conductance in some olfactory neurons, leading to depolarizationof olfactory cells in response to odorant-receptor binding. Olfactory signaling is ter-minated when receptors are phosphorylated via a negative feedback reaction catalyzedby two types of kinases [57].The large family of G-protein-coupled 7TM receptors just described may not be the
only odorant receptors. An alternate signaling pathway for olfactory transduction hasrecently been proposed by Gibson and Garbers [58]. They have found a large family ofolfactory neuron-specific guanylyl cyclases that are membrane-bound and contain ex-tracellular domains that may constitute a second family of odorant receptors. Activa-tion of guanylyl cyclase elevates cyclic GMP (cGMP) that converges on the same CNGchannels as cAMP to generate action potentials.Repeated stimulation of olfactory receptor neurons leads to decrements in the neur-
al responses, i.e. adaptation. Three forms of olfactory adaptation can take place inolfactory receptor neurons: two rapid forms and one persistent form. These threedifferent adaptation phenomena are controlled, at least in part, by separate molecularmechanisms. These mechanisms involve Ca2þ entry through CNG channels, Ca2þ-dependent CNG channel modulation, Ca2þ/calmodulin kinase II-dependent attenua-tion of adenylyl cyclase, and the activity of the carbon monoxide/cyclic GMP secondmessenger system [59].
1.5
Molecular Biology Of Olfaction
The molecular era of olfaction began in 1991 with the discovery by Buck and Axel of amultigene family of G-protein-coupled ORs with a 7TM-spanning typology. Buck andAxel [8] obtained complementary DNA (cDNA) utilizing olfactory epithelial RNA fromrat in conjunction with an amplification process called the polymerase chain reaction(PCR). (Complementary DNA is a copy of a messenger RNA). They found a PCRproduct (PCR 13) that contained multiple species of DNA that are representativeof a multiple gene family that encodes transmembrane domain proteins that are re-stricted to the olfactory epithelium. Further work has shown that there is a conserva-tion of certain amino acidmotifs within OR gene sequences that distinguish ORs fromother 7TM proteins [8, 60, 61]. There are also hypervariable regions within certainmembrane regions of ORs (i.e. TMs 3, 4, and 5) that provide a diversity of ligand-binding pockets [61]. A single amino acid substitution in the hypervariable regioncan change ligand-binding specificity [62]. This diversity in ligand-binding domainsis necessary to accommodate the enormous number of structurally diverse volatilechemicals that can activate the olfactory sensory neurons.Early estimates suggested that there are approximately 500 to 750 genes that encode
ORs in humans with an estimated 1000 genes in mouse and rat [7, 9, 10, 63]. However,there appears to be a high frequency of pseudogenes (genes with defects that are in-compatible with receptor function) in the human but not rat OR repertoire; between38% to 76% of the human sequences do not encode full-length polypeptides
1.5 Molecular Biology Of Olfaction 2121

[9, 10, 64, 65]. One recent report claims to have identified and physically cloned 347human OR genes that they believe represent the complete repertoire of functionalhuman ORs [66]. This reduction in the functional fraction of olfactory receptor genesin humans compared to rats implies that olfaction may have become less importantduring the course of evolution, perhaps due to relaxed selective constraints [65]. Thedecrease in viable odorant genesmay also be the cause of specific anosmias in humans(inabilities to smell a particular odorant).OR genes are typically organized in clusters of ten ormore and are distributed across
numerous chromosomes [9, 10, 66–68]. The 347 olfactory genes identified by Zozulyaet al. [66] were located on all human chromosomes, except for 2, 4, 18, 20, 21, and Y,with themajority (155 ORs) on chromosome 11 followed in frequency by chromosome1 (42ORs), 9 (26ORs) and 6 (24ORs). The average human OR is approximately 315amino acids long. In general, only one OR gene is expressed in a single olfactorysensory neuron [69], and olfactory sensory neurons (OSNs) that express a singleOR converge on the same glomerulus in the olfactory bulb. Thus for the adult mousewhich has � 1000OR types and � 1800 glomeruli [70], each OR may be associatedwith only two specific glomeruli. However, it should be noted that one recent studyreported that there may be a subset of OSNs that expresses two distinct receptor types[71].Knowledge of the physiological functioning of specific ORs is still in its infancy.
That is, we know very little about the range of ligands that interact with each ofthe particular odorant receptors. This is due in part to the large number of odorantreceptors and the enormous repertoire (many thousands) of odorous compounds.Experimental approaches in which ORs are functionally expressed in olfactory sen-sory neurons are necessary to determine the tuning of a specific OR. Functional ex-pression of a specific ORs is achieved experimentally when a given receptor type isinserted into the plasma membrane, couples with the second messenger system,and produces a measurable response to an odorant ligand.Direct functional proof that the 7TM receptors cloned by Buck and Axel [8] were
actually odor receptors was obtained by Zhao et al. [72] who inserted a gene discoveredby Buck and Axel into the rat olfactory system, producing electrical activity in olfactoryneurons to specific odorant chemicals. Zhao et al. functionally expressed an OR inolfactory sensory neurons of rat in vivo using an adenovirus-mediated gene transferof a cloned OR, I7 (see ref. 8 for nomenclature). They inserted the I7 genes into anadenovirus vector linked to a gene for green fluorescent protein (GFP) that is used tomark genetically altered cells. (Disabled adenovirus vectors are used as a tool to trans-fer genes into mammalian cells. A viral gene can be replaced with another gene thatencodes an OR protein.) Cells that carried the rat I7 gene also carried the GFP gene,and thus could be visualized because they glowed bright green when exposed to bluelight. Extracellular transepithelial potential recordings from summed activity of manyolfactory neurons (called an electro-olfactogram) in the infected epithelium were ele-vated to heptaldehyde (C7), octyl aldehyde (C8), nonyl aldehyde (C9), and decyl aldehyde(C10) when compared with uninfected epithelium [72]. However, electro-olfactogramamplitudes were not elevated for hexaldehyde (C6) or undecylic aldehyde (C11). Thesefindings suggested that the response profile of the 17 receptor is relatively specific for
1 Introduction to Olfaction: Perception, Anatomy, Physiology, and Molecular Biology22

C7 to C10 saturated aliphatic aldehydes at least within the limited set of 74 odorants thatwas tested. Heptaldehyde (C7), octyl aldehyde (C8), nonyl aldehyde (C9), and decyl al-dehyde (C10) can be differentiated on the basis of odor quality so that a single receptortype does not code for a specific odor quality.Malnic et al. [69] used a combination of calcium imaging and single-cell RT-PCR
(PCR with reverse transcription) to identify ORs for odorants with related structuresbut varied odors. Their results indicate that one OR recognizes multiple odorants, oneodorant is recognized by multiple ORs, but that different odorants are recognized bydifferent combinations of ORs. They concluded that the olfactory system uses a com-binatorial receptor coding scheme to encode odor identities.
1.6
Taste
A brief overview of taste will also be given here because some of the sensors describedin this book are ‘taste sensors.’
1.6.1
Taste Classification Schemes Based on Sensory Properties
Historically, the taste literature often suggests that there are only four (or possibly five)basic taste qualities (sweet, sour, salty, and bitter, and possibly ‘umami’ which is thetaste of glutamate salts). All other tastes have been presumed to be combinations ofthese basic tastes. However, data are now accumulating that the range of taste sensa-tions is much broader and includes qualities such as astringency, metallic, fatty, andcalcium-like (e.g. chalky) [73–78].
1.6.2
Physiology and Anatomy of Taste
The receptor cells for taste are neuroepithelial cells that are clustered into buds anddistributed on the dorsal surface of the tongue, tongue cheek margin, base of thetongue near ducts of the sublingual glands, the soft palate, pharynx, larynx, epiglot-tis, uvula, and first third of the esophagus (see Schiffman and Warwick [79] for anoverview of anatomy). Taste sensations are induced by the interaction of chemicals(e.g. from food) with taste-buds during ingestion, chewing, and swallowing. Indivi-dual taste cells generally respond to more than one type of taste. Taste buds consistof approximately 50–100 cells that arranged in an onion-like structure (see Fig. 1.14).Individual cells in a taste-bud undergo continuous renewal every 10 to 10 1/2 days.Taste-buds on the tongue are positioned on specialized epithelial projections termedpapillae. There are three different kinds of lingual papillae that contain taste-buds:fungiform papillae (which are shaped somewhat like mushrooms), foliate papillae
1.6 Taste 2323

(which consist of linear depressions or vertical folds), and circumvallate papillae(which are surrounded by deep moats). The entire tongue is sensitive to all taste qua-lities but there are regional differences in sensitivity; for example, buds on fungiformpapillae are more sensitive to sodium salts, foliate papillae to acids, and circumvallateto bitter compounds.
Fig. 1.14 Taste bud
Fig. 1.15 Anatomy of taste showing the cranial nerves and nucleus of the solitary tract
1 Introduction to Olfaction: Perception, Anatomy, Physiology, and Molecular Biology24

Taste bud cells form direct neural connections called synapses with three cranialnerves: the facial nerve (VII), glossopharyngeal nerve (IX), and vagus nerve (X). Thesethree cranial nerves relay signals from taste receptor cells to the rostral portion of thenucleus of the solitary tract located in the medulla in the brain stem (see Fig. 1.15).Signals are ultimately transmitted to the thalamus and gustatory cortex. Electrophy-siological studies indicate that individual taste neurons have broad, overlapping re-sponse patterns (i.e. they are broadly tuned) so that an individual fiber is non-specificbut collectively the pattern of activity across multiple neurons is unique for a givenstimulus [77, 80].
1.6.3
Transduction of Taste Signals
Taste stimuli interact with taste proteins (e.g. taste receptors) or with ion channels onthe surface of taste cells, which induces electrical signals that ultimately reach thebrain to register a taste. The salty taste of sodium salts is produced when Naþ ionstraverse sodium channels in the membranes of taste cells [81]. The taste of potassiumsalts, like sodium salts, involves conductance of Kþ cations through taste cell mem-branes [82] Most studies indicate that the detection of bitter and sweet by tastantsreceptor cells involves G-protein-coupled receptors. Some but not all sweet com-pounds appear to bind to 7TM-spanning cell-surface receptors that activate the ade-nylate cyclase second messenger cascade [83]. At least two pathways are involved inbitter taste transduction: 1) the phosphatidylinositol secondmessenger cascade, and 2)the alpha-gustducin/phosphodiesterase pathway [86].
1.6.4
Molecular Biology of Taste
At current writing, two families of G-protein-coupled receptors designated as T1R(taste receptor family 1) and T2R (taste receptor family 2) are known to be selectivelyexpressed in subsets of taste receptor cells. In 1999, Hoon et al. [84] cloned and char-acterized two novel 7TM domain proteins T1R1 and T1R2 (taste receptor family 1,members 1 and 2) that are expressed in topographically distinct subpopulations oftaste receptor cells and taste buds. The receptors were localized to the taste pore.The following year, a novel family of receptors T2R were identified [85–87], andlike T1R1 and T1R2, the T2R genes were selectively expressed in taste receptorcells. The T2R family consists of 40-80 proteins that appear to code specifically forbitter tastants. A candidate sweet receptor gene, called T1R3 (taste receptor family1, member 3) was also been identified [88–91]. Further research has shown that re-ceptors T1R2 and T1R3 combine by dimerization producing heterodimers (T1R2 þ 3)to recognize sweet-tasting molecules with different structures such as sucrose andsaccharin [92]. Receptors T1R1 and T1R3 combine by dimerization producing hetero-dimers (T1R1 þ 3) that are broadly tuned to recognize L-amino acids [93]. A receptor
1.6 Taste 2525

that binds the amino acid L-glutamate called mGluR4 has also been cloned and char-acterized [94].Cells expressing T1R2 þ 3 are found predominantly on the posterior tongue, which
is innervated by the glossopharyngeal nerve [92]. Cells expressing T1R2 þ 3 are alsolocated on the palate. Cells expressing T1R1 þ 3 are found predominantly on the frontof the tongue, which is innervated by the chorda tympani nerve. Even though thesedifferent taste receptor types appear to be segregated anatomically, electrophysiologi-cal experiments indicate that individual taste cells and nerve fibers respond to stimulihaving multiple taste qualities [77, 80]. Thus, further research is needed clarify the fullrange of taste receptors as well as elucidate how this taste information is coded by thenervous system.
1.7
Final Comment
The biological chemosensory systems just described share many analogies to exam-ples of machine olfaction described in this book. For example, both the human olfac-tory system andmachines havemechanisms for sample handing. In humans, a sniff isinitiated when the diaphragm creates a relative negative pressure in the lungs andforces an air sample to be drawn through the nostrils and directed by the curved tur-binates onto the sensory layer of the olfactory epithelium. In a typical electronic nose, avacuum pump produces a negative relative pressure to draw the air sample through atube (plastic or stainless steel) in a small chamber housing the electronic sensor array.Both biological systems and machines have far fewer sensors than the thousands ofknown odorants. Humans have several hundred different receptor types while theelectronic nose typically has only 5 to 32 sensors. Both biological and machinessend their responses into multilevel neural networks that identify and characterizethe odor being produced by the odorant sample. Future advances in the molecularbiology of smell and taste will undoubtedly impact the development of new electronicnose and electronic tongue devices.
References
1 G. Ohloff. Chemistry of odor stimuli. Ex-perientia 1986, 42, 271–279.
2 M. Stuiver. Biophysics of the sense of smell.PhD Thesis. Groningen. 1958.
3 H. DeVries, M. Stuiver. The absolute sen-sitivity of the human sense of smell. inSensory Communication (Ed.: W. A. Rosen-blith), JohnWiley and Sons, New York, 1961,Chapter 9, pp. 159–167.
4 USEPA (United States Environmental Pro-tection Agency). Odor and corrosion control insanitary sewerage systems and treatment plants.USEPA, Cincinnati, OH EPA/625/1-85/018,1985.
5 A. Jinks, D. G. Laing. The analysis of odormixtures: evidence for a configurationalprocess. Physiol. Behav. 2001, 72, 51–63.
6 M. G. J. Beets. Structure-activity relationshipsin human chemoreception. Applied SciencePublishers Ltd., London, 1978.
7 R. Axel. The molecular logic of smell.Sci. Am. 1995, 273, 154–159.
8 L. Buck, R. Axel. A novel multigene familymay encode odorant receptors: a molecularbasis for odor recognition. Cell 1991, 65,175–87.
1 Introduction to Olfaction: Perception, Anatomy, Physiology, and Molecular Biology26

9 P. Mombaerts. Molecular biology of odorantreceptors in vertebrates. Annu. Rev. Neurosci.1999, 22, 487–509.
10 P. Mombaerts. Seven-transmembraneproteins as odorant and chemosensoryreceptors. Science 1999, 286, 707–711.
11 Aristotle, De Anima, translated byW. S. Hett, Heinemann, London, Revisedand reprinted, 1957.
12 C. Linnaeus (C. Von Linne) Odores medica-mentorum in Amoenitates Academicae, vol. 3,Lars Salvius: Stockholm, 1752, p. 183.
13 H. Zwaardemaker. Die Physiologie desGeruchs, translated from Dutch by A. J.von Langegg, W. Engelmann: Leipzig, 1895,p. 324.
14 H. Henning. Der Geruch I, Z. Psychol.Physiol. Sinnesorgane 1915, 73, 161–257.
15 S. Klein. Primary odour element classifica-tion. Amer. Perfum. Essent. Oil Rev. 1947, 50,453–454.
16 J. E. Amoore. The stereochemical specifici-ties of human olfactory receptors. Perfum.Essent. Oil Rec. 1952, 43, 321–323, and 330.
17 J. E. Amoore. The stereochemical theory ofolfaction. 1. Identification of seven primaryodours. Proc. Sci. Sect. Toilet Goods Assoc.New York 1962, 37, S1–S12.
18 J. E. Amoore. The stereochemical theory ofolfaction. 2. Elucidation of the stereoche-mical properties of the olfactory receptorsites. Proc. Sci. Sect. Toilet Goods Assoc.New York 1962, 37, S13–S23.
19 H. G. Schutz. A matching standards methodfor characterising odour qualities. Ann.N. Y. Acad. Sci. 1964, 116, 517–526.
20 American Society for Testing and Materials(ASTM), Atlas Of Odor Character Profiles. DS61. ASTM, Philadelphia, 1992.
21 www.AlluredCompendium.com. AlluredPublishing Corp., 362 S. Schmale Road,Carol Stream, IL USA 60188-2787, 2001.
22 Aldrich Flavors and Fragrances http://www.sigma-aldrich.com. Aldrich,1001 St. Paul Avenue, Milwaukee,WI 53233 USA.
23 http://www.leffingwell.com.24 D. Whissell-Buechy, J. E. Amoore. Odour-
blindness to musk: simple recessive in-heritance. Nature 1973, 242, 271–273.
25 C. J. Wysocki, G. K. Beauchamp. Abilityto smell androstenone is genetically deter-mined. Proc. Natl. Acad. Sci. USA 1984, 81,4899–4902.
26 J. E. Amoore, Specific anosmia and theconcept of primary odors. Chem. SensesFlavor 1977, 2, 267–281.
27 L. Turin, F. Yoshii. Structure-odor relations:A modern perspective. in Handbook of Ol-faction and Gustation (Ed.: R. L. Doty),Marcel Dekker, New York, 2002.
28 J. E. Amoore, D. Venstrom. Correlationsbetween sterochemical assessment and or-ganoleptic analysis of odorous compounds.In Olfaction and Taste, vol. 2 (Ed.: T.Hayashi), Pergamon, Oxford, 1967, p. 3–17.
29 J. E. Amoore. Stereochemical and vibrationaltheories of odour. Nature 1971, 233,270–271.
30 R. H. Wright. Stereochemical and vibratio-nal theories of odour. Nature 1972, 239, 226.
31 R. H. Wright. Odour and molecular vibra-tion. Nature 1966, 209, 571–572.
32 R. H. Wright, A. Robson. Basis of odourspecificity: homologues of benzaldehyde andnitrobenzene. Nature 1969, 222, 290–292.
33 A. Dravnieks, P. Laffort. Physico-chemicalbasis of quantitative and qualitative odordiscrimination in humans. in Olfaction andTaste, vol. 4, (Ed.: D. Schneider), Wissen-schaftliche Verlagsgesellschaft, Stuttgart,1972, pp. 142–148.
34 M. Chastrette. Trends in structure-odorrelationships. SARQSAR Environ. Res. 1997,6, 215–254.
35 K. J. Rossiter. Structure-Odor Relationships.Chem. Rev. 1996, 96, 3201–3240.
36 S. S. Schiffman, M. L. Reynolds, F. W.Young. Introduction to multidimensional sca-ling: Theory, methods, and applications. NewYork: Academic Press, New York, 1981.
37 S. S. Schiffman. Future design of flavourmolecules by computer. Chem Ind. 1983, 3,39–42.
38 S. S. Schiffman. Physicochemical correlatesof olfactory quality. Science 1974, 185,112–117.
39 S. S. Schiffman, J. C. Leffingwell. Perceptionof odors of simple pyrazines by youngand elderly subjects: A multidimensionalanalysis. Pharmacol. Biochem. Behav. 1981,14, 787–798.
40 A. J. Stuper, P. C. Jurs. ADAPT: A computersystem for automated data analysis usingpattern recognition techniques. J. Chem. Inf.Comp. Sci. 1976, 16, 99–105.
41 http://zeus.chem.psu.edu/ADAPT.html
1.7 Final Comment 2727

42 J. Cohen, P. Cohen. Applied multiple regres-sion/correlation analysis for the behavioral sci-ences. Lawrence Erlbaum, Hillsdale NJ, 1983.
43 V. A. Benignus, K. E. Muller, J. A. Graham,C. N. Barton. Toluene levels in blood andbrain of rats as a function of toluene level ininspired air. Environ. Res. 1984, 33, 39–46.
44 J. A. Maruniak, W. L. Silver, D. G. Moulton.Olfactory receptors respond to blood-borneodorants. Brain Res. 1983, 265, 312-316.
45 D. G.Moulton. Dynamics of cell populationsin the olfactory epithelium. Ann. N Y Acad.Sci. 1974, 237, 52–61.
46 D. Lancet. Olfaction. The strong scent ofsuccess. Nature 1991, 351, 275–276.
47 E. E. Morrison. Morphology and plasticiy ofthe vertebrate olfactory epithelium. inScience of olfaction (Ed.: M. J. Serby,K. L. Chobor) Springer-Verlag, New York,1992, p. 31–50.
48 K. Mori, G. M. Shepherd. Emergingprinciples of molecular signal processingby mitral/tufted cells in the olfactory bulb.Semin. Cell Biol. 1994, 5, 65–74.
49 K. J. Ressler, S. L. Sullivan, L. B. Buck.A zonal organization of odorant receptorgene expression in the olfactory epithelium.Cell 1993, 73, 597–609.
50 K. Mori , H. Nagao, Y. Yoshihara. Theolfactory bulb: coding and processing ofodor molecule information. Science 1999,286, 711–715.
51 B. D. Rubin, L. C. Katz. Optical imaging ofodorant representations in the mammalianolfactory bulb. Neuron 1999, 23, 499–511.
52 L. Belluscio, L. C. Katz. Symmetry, stereo-typy, and topography of odorant represen-tations in mouse olfactory bulbs. J. Neurosci.2001, 21, 2113–2122.
53 B. Kettenmann, C. Hummel, H. Stefan,G. Kobal. Multichannel magnetoencephalo-graphical recordings: separation of corticalresponses to different chemical stimulationin man. Electroencephalogr. Clin. Neurophy-siol. Suppl. 1996, 46, 271–274.
54 D. H. Zald, J. V. Pardo. Emotion, olfactionand the human amygdala: Amygdalaactivation during aversive olfactorystimulation. Proc. Natl. Acad. Sci. USA 1997,94, 4119–4124.
55 J. E. Cometto-Muniz, W. S. Cain. Sensoryirritation. Relation to indoor air pollution.Ann. N. Y. Acad. Sci. 1992, 641, 137–151.
56 J. E. Cometto-Muniz, W. S. Cain. Sensoryreactions of nasal pungency and odor tovolatile organic compounds: the alkylben-zenes. AIHAJ 1994, 55, 811–817.
57 H. Breer. Odor recognition and secondmessenger signaling in olfactory receptorneurons. Semin. Cell Biol. 1994, 5, 25–32.
58 A. D. Gibson, D. L. Garbers. Guanylylcyclases as a family of putative odorantreceptors. Ann. Rev. Neurosci. 2000, 23,417–439.
59 F. Zufall, T. Leinders-Zufall. The cellularand molecular basis of odor adaptation.Chem. Senses 2000, 25, 473–481.
60 W. C. Probst, L. A. Snyder, D. I. Schuster,J. Brosius, S. C. Sealfon. Sequencealignment of the G-protein coupled receptorsuperfamily. DNA Cell Biol. 1992, 11, 1–20.
61 Y. Pilpel, D. Lancet. The variable andconserved interfaces of modeled olfactoryreceptor proteins. Protein Sci. 1999, 8,969–977.
62 D. Krautwurst, K. W. Yau, R. R. Reed.Identification of ligands for olfactory recep-tors by functional expression of a receptorlibrary. Cell 1998, 95, 917–926.
63 L. B. Buck. Information coding in the ver-tebrate olfactory system.Annu. Rev. Neurosci.1996, 19, 517–544.
64 S. Rouqier, C. Friedman, C. Delettre,G. van den Engh, A. Blancher,B. Crouau-Roy, B. J. Trask, D. Giorgi.A gene recently inactivated in humandefines a new olfactory receptor familyin mammals. Hum. Mol. Gen. 1998, 7,1337–1345.
65 S. Rouquier, A. Blancher, D. Giorgi.The olfactory receptor gene repertoire inprimates and mouse: evidence for reductionof the functional fraction in primates. Proc.Natl. Acad. Sci. USA 2000, 97, 2870–2874.
66 S. Zozulya, F. Echeverri, T. Nguyen. Thehuman olfactory receptor repertoire.GenomeBiol. 2001, 2(6), research0018.1-0018.12,http://genomebiology.com/2001/2/6/research/0018.
67 N. Ben-Arie N, D. Lancet, C. Taylor,M. Khen, N. Walker, D. H. Ledbetter,R. Carrozzo, K. Patel, D. Sheer, H. Lehrach.Olfactory receptor gene cluster on humanchromosome 17: Possible duplication of anancestral receptor repertoire. Hum. Mol.Genet. 1994, 3, 229–235.
1 Introduction to Olfaction: Perception, Anatomy, Physiology, and Molecular Biology28

68 B. J. Trask, C. Friedman, A.Martin-Gallardo,L. Rowen, C. Akinbami, J. Blankenship,C. Collins, D. Giorgi, S. Iadonato,F. Johnson, W. L. Kuo, H. Massa,T. Morrish, S. Naylor, O. T. Nguyen,S. Rouquier, T. Smith, D. J. Wong, J.Youngblom, G. van den Engh. Members ofthe olfactory receptor gene family are con-tained in large blocks of DNA duplicatedpolymorphically near the ends of humanchromosomes. Hum. Mol. Genet. 1998, 7,13–26.
69 B. Malnic, J. Hirono, T. Sato, L. B. Buck.Combinatorial receptor codes for odors. Cell1999, 96, 713–723.
70 J. P. Royet, C. Souchier, F. Jourdan,H. Ploye. Morphometric study of the glo-merular population in the mouse olfactorybulb: numerical density and size distributionalong the rostrocaudal axis. J. Comp Neurol.1988, 270, 559–568.
71 N. E. Rawson, J. Eberwine, R. Dotson,J. Jackson, P. Ulrich, D. Restrepo. Expres-sion of mRNAs encoding for two differentolfactory receptors in a subset of olfactoryreceptor neurons. J. Neurochem. 2000, 75,185–195.
72 H. Zhao, L. Ivic, J. M. Otaki , M. Hashimoto,K. Mikoshiba, S. Firestein. Functionalexpression of a mammalian odorant recep-tor. Science 1998, 279, 237–242.
73 R. D. Mattes. The taste of fat elevates post-prandial triacylglycerol. Physiol. Behav. 2001,74, 343–348.
74 S. S. Schiffman, M. S. Suggs, A. L. Sostman,S. A. Simon. Chorda tympani and lingualnerve responses to astringent compounds inrodents. Physiol. Behav. 1992, 51, 55–63.
75 S. S. Schiffman, B. G. Graham, E. A. Sattely-Miller, Z. S. Warwick. Orosensory percep-tion of dietary fat. Curr. Dir. Psychol. Sci.1998, 7, 137–143.
76 M. G. Tordoff. Calcium: Taste, intake, andappetite. Physiol. Rev. 2001, 81, 1567–1597.
77 S. S. Schiffman. Taste quality and neuralcoding: implications from psychophysicsand neurophysiology. Physiol. Behav. 2000,69, 147–159.
78 T. A. Gilbertson. Gustatory mechanismsfor the detection of fat.Curr. Opin. Neurobiol.1998, 8, 447–452.
79 S. S. Schiffman, Z. S. Warwick. The biologyof taste and food intake. in The Science OfFood Regulation: Food Intake, Taste, Nutrient
Partitioning, And Energy Expenditure,Pennington Center Nutrition Series, Volume 2(Ed.: G. A. Bray, D. H. Ryan), LouisianaState University Press, Baton Rouge, 1992,p. 293–312.
80 T. A.Gilbertson, J. D. Boughter Jr,H. Zhang,D. V. Smith. Distribution of gustatorysensitivities in rat taste cells: whole-cellresponses to apical chemical stimulation.J. Neurosci. 2001, 21, 4931–4941.
81 S. S. Schiffman, E. Lockhead, F. W. Maes.Amiloride reduces the taste intensity ofNaþ and Liþ salts and sweeteners. Proc. Natl.Acad. Sci. 1983, 80, 6136–6140.
82 M. Kim, C. M. Mistretta. 4-aminopyridinereduces chorda tympani nerve tasteresponses to potassium and alkali saltsin rat. Brain Res. 1993, 612, 96–103.
83 B. Lindemann. Chemoreception: tastingthe sweet and the bitter. Curr. Biol. 1996, 6,1234–1237.
84 M. A. Hoon, E. Adler, J. Lindemeier,J. F. Battey, N. J. Ryba, C. S. Zuker. Putativemammalian taste receptors: a class of taste-specific GPCRs with distinct topographicselectivity. Cell 1999, 96, 541–551.
85 E. Adler, M. A. Hoon, K. L. Mueller,J. Chandrashekar, N. J. Ryba, C. S. Zuker.A novel family of mammalian taste recep-tors. Cell 2000, 100, 693–702.
86 H. Matsunami, J. P. Montmayeur,L. B. Buck. A family of candidate tastereceptors in human and mouse. Nature2000, 404, 601–604.
87 J. Chandrashekar, K. L. Mueller,M. A. Hoon, E. Adler, L. Feng, W. Guo,C. S. Zuker, N. J. Ryba. T2Rs functionas bitter taste receptors. Cell 2000, 100,703–711.
88 J. P. Montmayeur, S. D. Liberles,H. Matsunami, L. B. Buck. A candidatetaste receptor gene near a sweet taste locus.Nat. Neurosci. 2001, 4, 492–498.
89 M. Max, Y. G. Shanker, L. Huang, M. Rong,Z. Liu, F. Campagne, H. Weinstein,S. Damak , R. F. Margolskee. Tas1r3,encoding a new candidate taste receptor,is allelic to the sweet responsiveness locusSac. Nat. Genet. 2001, 28, 58–63.
90 E. Sainz, J. N. Korley, J. F. Battey, S. L.Sullivan. Identification of a novel memberof the T1R family of putative taste receptors.J. Neurochem. 2001, 77, 896–903.
1.7 Final Comment 2929

91 X. Li, M. Inoue, D. R. Reed, T. Huque,R. B. Puchalski, M. G. Tordoff, Y. Ninomiya ,G. K. Beauchamp, A. A Bachmanov. High-resolution genetic mapping of the saccharinpreference locus (Sac) and the putative sweettaste receptor (T1R1) gene (Gpr70) to mousedistal Chromosome 4.Mamm.Genome 2001,12, 13–16.
92 G. Nelson, M. A. Hoon, J. Chandrashekar,Y. Zhang, N. J. Ryba, C. S. Zuker.Mammalian sweet taste receptors. Cell 2001,106, 381–390.
93 G. Nelson, J. Chandrashekar, M. A. Hoon„L. Feng, G Zhao, N. J. Ryba, C. S. Zuker.An amino-acid receptor. Nature onlinepublication, 24 February 2002 (DOI10.1038/nature726).
94 N. Chaudhari, A. M. Landin, S. D. Roper.A metabotropic glutamate receptor variantfunctions as a taste receptor. Nat. Neurosci.2000, 3, 113–119.
Appendix
The basic matrix equations used by Schiffman [38] to maximize the configurationalsimilarity of the psychologically determined space in Fig. 1.8 with a space generated byweighted physicochemical parameters are:
P ¼ P þ EP ¼ DQP ¼ DQ þ E
where P is an (n)(n�1)/2 column vector whose elements pij represent all the inter-stimulus distances between stimulus i and stimulus j and where n is the total numberof stimuli; P is an (n)(n�1)/2 column vector representing the proximity measuresbased on weighted physicochemical parameters; D is an [(n)(n�2)/2] by k scalar di-stance matrix whose elements d2ðijÞk are the squared differences between stimulus i andstimulus j for each physicochemical parameter k; Q is a k element column vector ofweights for the k physicochemical parameters; and E is an (n)(n�1)/2 column vectorrepresenting the error between the subjective proximities and the proximities based onphysicochemical measures.
The error to be minimized is
@E 0E=@Qk ¼ 0
leading to the least squares solution
Q ¼ ðD 0DÞ�1D 0P
The equations for canonical correlation used to relate the descriptors in Table 1.6 to thethree dimensional arrangement in Fig. 1.9 are given below.
yyki ¼ ako þ ak1ðyi1Þ þ ::::: þ akrðyirÞ
xxki ¼ bko þ bk1ðxi1Þ þ :::::þ bkrðxirÞ
1 Introduction to Olfaction: Perception, Anatomy, Physiology, and Molecular Biology30

where xil, xi2, etc. are the values of stimulus i on dimensions 1 and 2 of the MDS spacejust as in multiple regression equations, and yi1, yi2, etc. are ratings of stimulus i onseveral physicochemical parameters. The intercepts and weights are solved to maxi-mize the correlation between yykiand xxki.
1.7 Final Comment 3131





























