Embed Size (px)
Citation preview

Comparative Biochemistry and Physiology, Part A 154 (2009) 551–556
Contents lists available at ScienceDirect
Comparative Biochemistry and Physiology, Part A
j ourna l homepage: www.e lsev ie r.com/ locate /cbpa
Osmoregulatory traits of broad-toothed field mouse (Apodemus mystacinus)populations from different habitats
Michael Scantlebury a,b,⁎, Uri Shanas a,c, Keren Or-Chen d, Abraham Haim a,c
a Department of Biology, University of Haifa—Oranim, Tivon, 36006 Israelb Quercus, School of Biological Sciences, Queen's University Belfast, Belfast BT9 7BL, Northern Ireland, United Kingdomc Department of Evolutionary and Environmental Biology, University of Haifa, Mount Carmel, Haifa, Israeld School of Social Work, Faculty of Social welfare and Health Studies, University of Haifa, Mount Carmel, Haifa, Israel
⁎ Corresponding author. Quercus, School of BiologicaBelfast. Belfast BT9 7BL, UK. Tel.: +44 28 9097 2278; fa
E-mail address: [email protected] (M. Scantl
1095-6433/$ – see front matter © 2009 Elsevier Inc. Aldoi:10.1016/j.cbpa.2009.08.021
a b s t r a c t
a r t i c l e i n f oArticle history:Received 14 July 2009Received in revised form 29 August 2009Accepted 31 August 2009Available online 9 September 2009
Keywords:PhotoperiodSalinityUrineEnvironmental challengeRodentEvolutionThermoregulation
One mechanism for physiological adjustment of small mammals to different habitats and different seasons is byseasonal acclimatization of their osmoregulatory system. We examined the abilities of broad-toothed field mice(Apodemus mystacinus) from different ecosystems (‘sub-alpine’ and ‘Mediterranean’) to copewith salinity stressunder short day (SD) and long day (LD) photoperiod regimes. We compared urine volume, osmolarity, urea andelectrolyte (sodium, potassium and chloride) concentrations. Significant differenceswere noted in the abilities ofmice fromthe twoecosystems todealwith salinity load; inparticular sub-alpinemiceproduced less concentratedurine than Mediterranean mice with SD− sub-alpine mice seeming to produce particularly dilute urine. Ureaconcentration generally decreased with increasing salinity, whereas sodium and potassium levels increased,however SD− sub-alpine mice behaved differently and appeared not to be able to excrete electrolytes as effec-tively as the other groups of mice. Differences observed provide an insight into the kinds of variability that arepresent within populations inhabiting different ecosystems, thus how populations may be able to respond topotential changes in their environment. Physiological data pertaining to adaptation to increased xeric conditions,as modelled by A. mystacinus, provides valuable information as to how other species may cope with potentialclimatic challenges.
l Sciences, Queen's Universityx: +44 28 9097 5877.ebury).
l rights reserved.
© 2009 Elsevier Inc. All rights reserved.
1. Introduction
Small mammals inhabiting different ecosystems have evolvedvarying physiological capabilities which are necessary for survival intheir particular area (Haim and Izhaki,1993; Haim et al., 2005a;Wilmeret al., 2005). Outside of the tropics, photoperiod changes are an im-portant initial cue for seasonal acclimatization of physiological systemsin many species of small rodent (Haim, 1982; Haim and Levi, 1990).Much attention has been given to understanding the processes resultingfrom photoperiod manipulations involving reproductive and thermo-regulatory systems (Bronson, 1989; Haim et al., 2005a). In general, forseasonal acclimatizationof the thermoregulatory system, short day (SD)acclimation is an initial signal for winter acclimatizationwhilst long day(LD) acclimation is an initial signal for summer acclimatization (Haimand Fourie, 1980; Heldmaier et al., 1982; Haim, 1982; Haim and Yahav,1982). Differences in thermoregulatory and reproductive physiologies(e.g. age at reproduction, lifespan) are notable between individuals frompopulations inhabiting different environments, such as those fromdifferent altitudes, latitudes or from areas differing in aridity (Bronson,
1989). Other studies have highlighted the importance of photoperiodto behavioural and physiological responses. For example, acclimatiza-tion of the reproductive system to periods of light interference of one orfive min of light during the scotophase to SD-acclimated Siberianhamsters (Phodopus sangorus) resulted in activation of the malereproductive system (Hoffmann,1979).More recently, light interferenceto SD-acclimated social voles (Microtus socialis) was shown to result inthe loss of winter acclimatization of the thermoregulatory system—aphenomenon termed “seasons out of time” (Haim et al., 2005b). Thesestudies suggest that providing opposing seasonality signals may‘confuse’ the animal and result in an inadequate response to what waspreviously the major signal for seasonal acclimatization.
Environmental adjustments of small mammals to different habitatsand different seasons can be achieved by seasonal acclimatization of theosmoregulatory system (Ron and Haim, 2001; Shanas et al., 2002;Bozinovic et al., 2003). However, to our knowledge no studies have yetexamined the seasonality of kidney function in individuals from dif-ferent habitats in relation to manipulations of conflicting signals (e.g.short photoperiod as a signal for the rainy season and water stress as asignal for xeric conditions that occur during long days). Such a studywould be advantageous because it would demonstrate how differencesin the sensitivity to these signals would show population-level dif-ferences in physiological characteristics. Moreover, this would help

552 M. Scantlebury et al. / Comparative Biochemistry and Physiology, Part A 154 (2009) 551–556
elucidate site-specific differences in the sensitivities to a particularsignal (e.g. photoperiod versus water stress) in populations adapted todifferent ecosystems.
The Levant sub-region (e.g. northern Israel) is characterized byasymmetrical seasonality with a relatively short cold wet winter and along hot dry summer. Therefore, changes in kidney function, which canresult in increased urine osmolarity, are likely to be an essential adap-tation for rodents inhabiting this ecosystem (Shanas et al., 2003).Waterstress can be expressed by an increased particulate content of thewatersource (Scantlebury et al., 2002). In free-living animals, water stress isgreater during the summer (LD-conditions). By comparison, during thewinter (SD-conditions) water is not a limiting factor. The responses ofrodents to combined signals of photoperiod and water stress can there-fore occur under two different laboratory scenarios: (1) when bothsignals are in the same direction (i.e. LD and high salinity or SD and lowsalinity), or (2)when signals are in opposing directions (i.e. SD and highsalinity or LD and low salinity). These combinations were examinedin twodifferent populations of the broad-toothedfieldmouseApodemusmystacinus originating fromdifferent areas, which have previously beenshown to vary in their physiological characteristics (Haim et al., 1993,1996). As a non-communal species, distributed at different altitudes andhabitats in the Levant (Harrison and Bates, 1991), it is a good model forstudying physiological adaptations to environmental challenges. In thecurrent study we compared two populations of A. mystacinus, fromdifferent altitudes that are separated geographically by the AfricanGreat Rift Valley: mice from Mount Carmel (‘Mediterranean’ mice—lower altitude Mediterranean ecosystem) and mice from Mount Her-mon (‘sub-alpine’ mice—high altitude sub-Alpine ecosystem). In theformer region, the long hot and dry summers (8–9 months) have beensuggested to present the most problematic climatic challenge for thisspecies (Haim et al., 1993; Haim and Rubal; 1994). During this season,the vegetation is expected to have an increased particle (or osmotic)concentration due to the high evaporative water loss (Grammatikopou-los, 1999). By comparison, the cold winters and presence of snow arepresumed to present the most demanding conditions for mice inhab-iting the latter (Haim et al., 1993). Bearing in mind the differences be-tween the two ecosystems,we predicted that the greater environmentalchallenges sub-alpinemice face with regards to cold will be manifest assensitivity to changes in photoperiod in their osmoregulatory abilities.By comparison, the greater challenges Mediterranean mice face withrespect to xeric conditions will be manifest as sensitivity to changes inwater stress.
2. Materials and methods
2.1. Animals
A. mystacinus were captured from a site on the slopes of MountHermon representing a sub-alpine ecosystem (35°00′ E 33°00′ N;1650 m elevation) (n=16; 9 males and 7 females) in December 2001and June 2002 (‘sub-alpine’mice) and from the Har Horsham region inthe Carmel representing aMediterranean ecosystem (32°43′ E 34°58′N;100 m) (n=14; 8males and 6 females) inMarch2002 (‘Mediterranean’mice). Mice were taken to the laboratory at the University of Haifa—Oranim and maintained individually in cages (35×25×15 cm) wheretheywere providedwith dry sawdust and tissue paper as bedding. Theywere offered rodent chow (21% crude protein, 4% crude fat, 4% cellulose,13% moisture, 7% ash, 17.4 kJ/g gross energy; Koffolk, Israel) and dicedcarrots ad libitum for at least one month prior to any experiments. Nofemales were found to be pregnant.
For the experiments, agar gel (20 g of agar gel dissolved in 1000mLof water) was substituted as a water source. Mice were initially ac-climated to a short photoperiod (short day; ‘SD’) (16D:8 L) for threeweeks and then urine was collected at three different salinity levels(below). They were then acclimated to a long photoperiod (long day;‘LD’) (8D:16 L) for three weeks and all measurements were repeated.
The intensity of the photophase was 450lx, which was provided bywhitefluorescent lighting. Adim red lightwason continuously (<25lx),which allowed experimenters to work in the ‘dark’ periods. Ambienttemperature was kept constant at 25±0.1 °C throughout the entire ex-perimental period. Animalswere kept under licence in the animal houseat the University of Haifa—Oranim and experimental protocols wereassessed and approved by the University of Haifa ethics committee.
2.2. Salinity changes
To simulate changes of osmotic load experienced in thewild acrossthe seasons, animals were exposed to different levels of water stressby altering the level of salt (NaCl) dissolved in the agar provided astheir water source. This has been found to affect the thermoregulatoryand osmoregulatory capabilities of rodents in the Mediterraneanecosystem (e.g. Acomys cahirinus: Scantlebury et al., 2002; Shanaset al., 2003). In the current study we exposed both SD- and LD-acclimated mice to salt concentrations in the agar of 0.9% to 1.4% and1.8%. Mice were weighed at the end of each two-week acclimationperiod, prior to urine collection. Experiments were repeated for eachof the six different treatment conditions (i.e. for the two photoperiodsand the three salinities) for mice from both ecosystems (below).
2.3. Urine collection
After acclimation to a specific salinity, mice were transferred tometabolic cages (19.5×11.5×9 cm) with mesh floors above sheets ofParafilm® for 24 h. Theywere providedwith rodent chow ad libitum andthe particular agar gel as the water source. Every two hours the urinewas collected from the Parafilm® with a Pasteur pipette and stored inEppendorf tubes at 4 °C until analysis.
2.4. Urine volume, osmolarity, electrolytes and urea analysis
Within 24 h of collection, the urine volume was measured (±1µL,Pipetman, Gilson) and the urine osmolarity was determined using avapor pressure osmometer (Wescor 5500). Samples were stored at 4 °Cin sealed plastic tubes. Sodium and potassium concentrations weredetermined within 2 weeks using a flame photometer (Sherwood 410).Chloride concentrations were determined by titration on a chlorideanalyzer (Corning 925). Urea concentrations were determined using acommercial test kit (Sigma 640) on a spectrophotometer (Milton RoySpectronic 401).
2.5. Data analysis
We used a mixed design analysis (Page et al., 2003; Keppel andWickens, 2004). Groups were set as between-subject effects; season(long and short photoperiod acclimations) and salinity were set aswithin-subject effects.Means are presented±S.E. Significant effects arerepresented at different levels of p<0.001, p<0.01 and p<0.05,marginal effects between p>0.05 and p<0.1 and non significant effectsby p>0.1.
3. Results
3.1. Urine volume and body mass
For all groups of mice, urine volume and body mass (Wb) did notchange in response to increasing dietary salt content (F2,62=1.03,p>0.1 and F2,62=0.33, p>0.1, respectively). There was no interactionbetween urine volume or Wb and habitat (F1,62=0.12, p>0.1 andF1,62=0.65, p>0.1, respectively) or photoperiod (F1,62=0.60,p>0.1and F1,62=0.05, p>0.1 respectively).
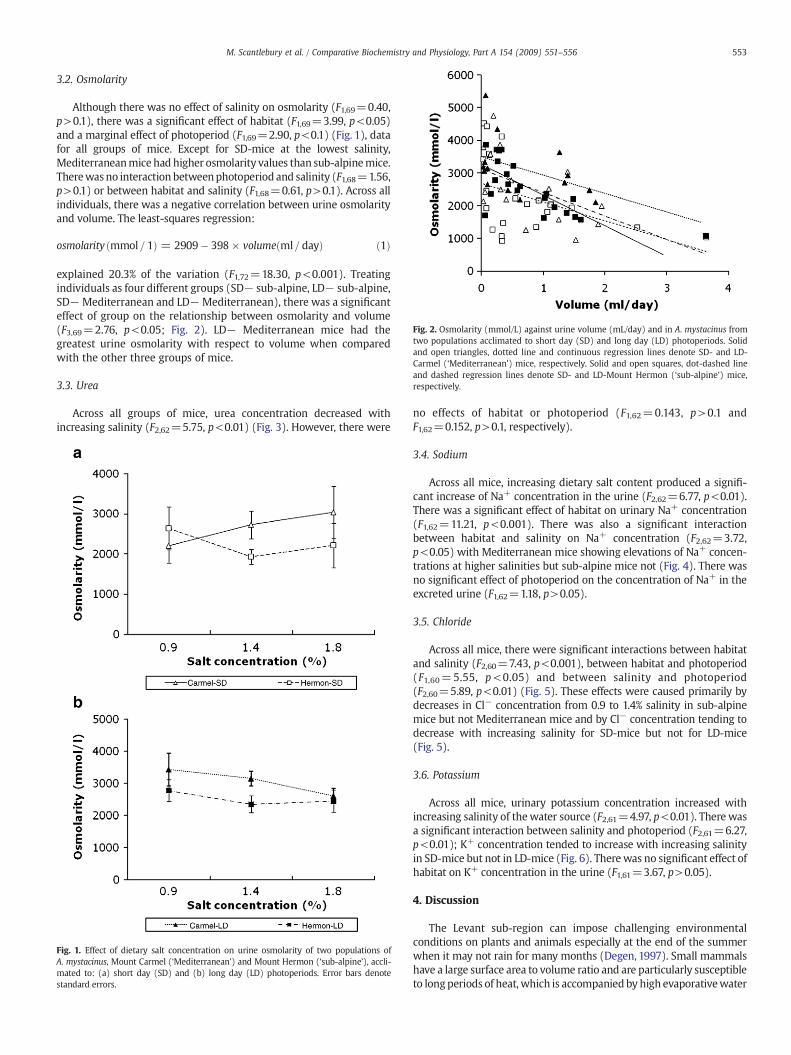
Fig. 2. Osmolarity (mmol/L) against urine volume (mL/day) and in A. mystacinus fromtwo populations acclimated to short day (SD) and long day (LD) photoperiods. Solidand open triangles, dotted line and continuous regression lines denote SD- and LD-Carmel (‘Mediterranean’) mice, respectively. Solid and open squares, dot-dashed lineand dashed regression lines denote SD- and LD-Mount Hermon (‘sub-alpine’) mice,respectively.
553M. Scantlebury et al. / Comparative Biochemistry and Physiology, Part A 154 (2009) 551–556
3.2. Osmolarity
Although there was no effect of salinity on osmolarity (F1,69=0.40,p>0.1), there was a significant effect of habitat (F1,69=3.99, p<0.05)and a marginal effect of photoperiod (F1,69=2.90, p<0.1) (Fig. 1), datafor all groups of mice. Except for SD-mice at the lowest salinity,Mediterraneanmice had higher osmolarity values than sub-alpinemice.Therewasno interaction betweenphotoperiod and salinity (F1,68=1.56,p>0.1) or between habitat and salinity (F1,68=0.61, p>0.1). Across allindividuals, there was a negative correlation between urine osmolarityand volume. The least-squares regression:
osmolarity ðmmol = 1Þ = 2909� 398 × volumeðml= dayÞ ð1Þ
explained 20.3% of the variation (F1,72=18.30, p<0.001). Treatingindividuals as four different groups (SD− sub-alpine, LD− sub-alpine,SD−Mediterranean and LD−Mediterranean), there was a significanteffect of group on the relationship between osmolarity and volume(F3,69=2.76, p<0.05; Fig. 2). LD− Mediterranean mice had thegreatest urine osmolarity with respect to volume when comparedwith the other three groups of mice.
3.3. Urea
Across all groups of mice, urea concentration decreased withincreasing salinity (F2,62=5.75, p<0.01) (Fig. 3). However, there were
Fig. 1. Effect of dietary salt concentration on urine osmolarity of two populations ofA. mystacinus, Mount Carmel (‘Mediterranean’) and Mount Hermon (‘sub-alpine’), accli-mated to: (a) short day (SD) and (b) long day (LD) photoperiods. Error bars denotestandard errors.
no effects of habitat or photoperiod (F1,62=0.143, p>0.1 andF1,62=0.152, p>0.1, respectively).
3.4. Sodium
Across all mice, increasing dietary salt content produced a signifi-cant increase of Na+ concentration in the urine (F2,62=6.77, p<0.01).There was a significant effect of habitat on urinary Na+ concentration(F1,62=11.21, p<0.001). There was also a significant interactionbetween habitat and salinity on Na+ concentration (F2,62=3.72,p<0.05) with Mediterranean mice showing elevations of Na+ concen-trations at higher salinities but sub-alpine mice not (Fig. 4). There wasno significant effect of photoperiod on the concentration of Na+ in theexcreted urine (F1,62=1.18, p>0.05).
3.5. Chloride
Across all mice, there were significant interactions between habitatand salinity (F2,60=7.43, p<0.001), between habitat and photoperiod(F1,60=5.55, p<0.05) and between salinity and photoperiod(F2,60=5.89, p<0.01) (Fig. 5). These effects were caused primarily bydecreases in Cl− concentration from 0.9 to 1.4% salinity in sub-alpinemice but not Mediterranean mice and by Cl− concentration tending todecrease with increasing salinity for SD-mice but not for LD-mice(Fig. 5).
3.6. Potassium
Across all mice, urinary potassium concentration increased withincreasing salinity of thewater source (F2,61=4.97, p<0.01). There wasa significant interaction between salinity and photoperiod (F2,61=6.27,p<0.01); K+ concentration tended to increase with increasing salinityin SD-mice but not in LD-mice (Fig. 6). Therewas no significant effect ofhabitat on K+ concentration in the urine (F1,61=3.67, p>0.05).
4. Discussion
The Levant sub-region can impose challenging environmentalconditions on plants and animals especially at the end of the summerwhen it may not rain for many months (Degen, 1997). Small mammalshave a large surface area to volume ratio and are particularly susceptibleto long periods of heat, which is accompanied by high evaporativewater

Fig. 3. Effect of dietary salt concentration on urea concentration (mg/dL) of twopopulations of A. mystacinus, Mount Carmel (‘Mediterranean’) and Mount Hermon(‘subalpine’), acclimated to: (a) short day (SD) and (b) long day (LD). Open triangles andopen squares denote SD− Mount Carmel and SD− Mount Hermon mice, respectively.Solid triangles and solid squares denote LD− Mount Carmel and LD− Mount Hermonmice, respectively. Error bars denote standard errors. (It pertains to Figs. 3–6).
Fig. 4. Effect of dietary salt concentration on Na+ urine concentration (mmol/L).
554 M. Scantlebury et al. / Comparative Biochemistry and Physiology, Part A 154 (2009) 551–556
loss. They have therefore developed mechanisms of decreasing waterloss through a combination of behavioural and physiological means(Schmidt-Nielsen, 1964; Peinke and Brown, 1999). Indeed, closelyrelated species show differences in their physiological traits; which aresubsequently presumed to illustrate adaptation to environments whichmay vary on spatial and temporal scales (Shkolnik and Borut, 1969;Goyal et al., 1988). In some cases, physiological differences are apparentin populations within the same species inwhich the distribution coversdiverse habitats (Haim and Borut, 1981; Haim et al., 1993; Weissenbergand Shkolnik, 1994). Scantlebury et al. (2003, 2005) and Shanas et al.(2002, 2003) showed that populations of the same species within thesame locality, but occupying different micro-habitats may exhibit sig-nificant differences in their physiological capabilities including waterand energy use. These latter experiments were held in two opposingslopes of the same canyon (“Evolution Canyon” Nevo et al., 1998).
In the current study,we comparedphysiological differences betweenpopulations of A. mystacinus from a Mediterranean and a sub-alpineecosystem. Previous studies have demonstrated that these two popu-lations differ in their thermoregulatory capabilities (Haim et al., 1996)and in their daily variation in noradrenaline-induced thermogenesis(Spiegel and Haim, 2004). We used two environmental signals—
photoperiod and water stress (drinking water salinity) that affect theosmoregulatory system in different ways. Thus, our results can be con-sidered in twodifferentways: (1)whenboth signalswere conveying thesame season (e.g. ‘summer’—high water stress and LD or ‘winter’—lowwater stress and SD), and (2) where signals conveyed conflictinginformation (e.g. high water stress and SD or low water stress and LD).
We discovered no change in the volume of urine produced in re-sponse to salinity as in other studies (Sasidharan et al., 1990). Althoughprevious studies have shown that changes in photoperiod regimespromote thermoregulatory responses in A. mystacinus (Spiegel andHaim, 2004), it is possible that the purely photoperiod-simulatedseasonality in this study was not sufficient to elicit a response in urineproduction. Therefore, photoperiod manipulations may need to be ac-companied by other changes in the habitat such as water conditions,ambient temperature and/or humidity to produce a change. Mediter-ranean mice did however appear to behave differently from sub-alpinemice and present higher osmolarity values with increasing dietarysalinity (Fig.1).Moreover, althoughallmice responded as expectedwithan inverse relationship between urine volume and urine osmolarity(Ntshotsho et al., 2004), LD-Mediterraneanmicehadpersistently higherosmolarity values per given volume than other groups of mice. Thissuggests that LD-Mediterranean mice are able to generate more con-centrated urine and thus better able to deal with increases in waterstress than the other groups of mice.
In xeric environments the expectation is that urinevolumedecreasesand urea concentration increases with increasing water deprivation, as

Fig. 5. Effect of dietary salt concentration on Cl− urine concentration (mmol/L).
555M. Scantlebury et al. / Comparative Biochemistry and Physiology, Part A 154 (2009) 551–556
the urea production helps to build the osmotic gradient in the kidney(Frean et al., 1998; Peinke and Brown, 1999; Shanas et al., 2003). How-ever, we found that urea concentrations decreased with the increasingdietary salt concentration (Fig. 3). One possibility for this observedrelationship is that as A. mystacinus from the current study populationsare less xeric-adapted than rodents in other cited studies, theymay lackthe ability to increase urea production upon exposure towater stress. Inaddition, decreased urea concentration should be viewed in relation to
Fig. 6. Effect of dietary salt concentration on K+ urine concentration (mmol/L).
the overall electrolyte concentration in the urine. An inverse correlationbetween urea and electrolyte concentration has been demonstrated inseveral species; which is logical given that the kidneys have a finiteability to produce urine to a certain osmolarity (Schmidt-Nielsen et al.,1961).
Finally, we found differences between the populations in the sodium,potassium and chloride urinary concentrations when mice werechallenged with photoperiod and salinity. Mediterranean mice and LD−sub-alpine mice increased their urinary sodium concentration withincreasingdietary salinity but SD− sub-alpinemicedidnot (Fig. 4). This ispossibly explained by the fact that sub-alpine mice may not be asproficient in excreting high concentrations of electrolytes as Mediterra-neanmice;with SD− sub-alpinemice beingparticularly inept. In termsofconflicting signals, sub-alpine mice may therefore not have been‘expecting’ a high salinity load, especially under a SD (winter) photo-period. Concerning the differences in urinary chloride output with in-creasing dietary salinity, Cl− tended todecreasewith increasing salinity inSD-mice but not in LD-mice; this decrease was especially notable in SD−sub-alpine mice. Therefore, SD-mice, and in particular SD− sub-alpinemice, were less able to excrete Cl− under increasing salinity load than LD-or Mediterranean mice. This suggests again that sub-alpine mice may beless able to deal with water stress than Mediterranean mice, particularlyunder SD photoperiod. In contrast to these results, potassium urinaryconcentration increased in SD-mice in response to increasing dietarysalinity but not in LD-mice. The implications of this result are unclear butmight suggest some form of electrolyte compensation occurring in thekidneys; i.e. if osmolarity remains relatively unchanged, a reduction in theelimination rate of one electrolyte (Na+) might incur an increase in theelimination of another (K+) (Speakman and Weldy, 2001).
Overall, our findings show that two populations of A. mystacinusexhibit differences in osmoregulatory function as evidenced by theirdiffering abilities to concentrate urinary solutes under varying levels ofsalt load. In particular, sub-alpine mice were less adept at producingconcentrated urine thanMediterraneanmicewith SD− sub-alpinemiceseeming to be particularly inept. These differences can be understood interms of the different physiological capabilities of mice from the twohabitats to deal with water stress and also of the inability of somemiceto deal with water stress during a SD photoperiod. Variation in phy-siological characteristics between populations inhabiting differentecosystems canprovide an indication of species adaptability. Differencesobserved here provide an insight into the kinds of variability that arepresent within populations inhabiting vastly different ecosystems, thushow populations may be able to respond to potential changes in theirenvironment. Mediterranean habitats are expected to be especiallyvulnerable to increasing aridity (Thuiller, 2007). Physiological data per-taining to adaptation to xeric conditions, as modelled by A. mystacinus,provides valuable information as tohow this andother speciesmaycopewith these challenges, which may be particularly important in thecurrent era of global climatic change.
Acknowledgements
We would like to thank Dmitri Rozenband for his assistance incaring for the animals and Laura Buckley with help in the laboratory.This project was funded in part by the University of Haifa ResearchAuthorities to AH and a Royal Society Post Doctoral Visiting Fellowshipto MS.
References
Bozinovic, F., Gallardo, P.A., Visser, R.H., Cortés, A., 2003. Seasonal acclimatization inwaterflux rate, urine osmolality and kidney water channels in free-living degus: molecularmechanisms, physiological processes and ecological implications. J. Exp. Biol. 206,2959–2966.
Bronson, F.H., 1989. Mammalian Reproductive Biology. University of Chicago press,Chicago.
Degen, A.A., 1997. Ecophysiology of Small Desert Mammals. Springer-Verlag, Berlin.

556 M. Scantlebury et al. / Comparative Biochemistry and Physiology, Part A 154 (2009) 551–556
Frean, J.L., Downs, C.T., Lovegrove, B.G., 1998. Osmoregulatory capacity of an arborealrodent Thallomys nigricauda. J. Arid. Environ. 40, 97–107.
Goyal, S.P., Ghosh, P.K., Sasidharan, T.O., Chand, P.,1988. Bodywater relations in two speciesof gerbil (Tatera indica indica andMeriones hurrianae). J. Comp. Physiol. B 158,127–134.
Grammatikopoulos, G., 1999. Mechanisms for drought tolerance in two Mediterraneanseasonal dimorphic shrubs. Aust. J. Plant Physiol. 26, 587–593.
Haim, A.,1982. Effects of long scotophase and cold acclamation on heat production in twodiurnal rodents. J. Comp. Physiol. B 148, 77–81.
Haim, A., Fourie, F., 1980. Heat production in cold and long scotophase acclimated andwinter acclimatized rodents. Int. J. Biometeorol. 24, 231–235.
Haim, A., Borut, A., 1981. Heat production and dissipation in golden spiny mouse Acomysrussatus from two extreme habitats. J. Comp. Physiol. B 142, 445–450.
Haim,A., Levi, G.,1990. Role of body temperature in seasonal acclimatization: photoperiod-induced rhythms and heat production inMeriones crassus. J. Exp. Biol. 256, 237–241.
Haim, A., Izhaki, I., 1993. The ecological significance of metabolic rate and nonshiveringthermogenesis of rodents. J. Therm. Biol. 18, 71–81.
Haim, A., Rubal, A.,1994. Seasonal acclimatization of daily rhythms of body temperature intwo rodent species of different origins inhabitingMediterraneanwoodlands. Pol. Ecol.Stud. 20, 357–363.
Haim, A., Yahav, S., 1982. Non-shivering thermogenesis in winter acclimatized and longscotophase and cold acclimated Apodemus mystacinus (Rodentia). J. Therm. Biol. 7,193–195.
Haim, A., Rubal, A., Harari, J.,1993. Comparative thermoregulatoryadaptations offieldmiceof the genus Apodemus to habitat challenges. J. Comp. Physiol. B 163, 602–607.
Haim, A., Plaut, I., Zobedat, S., 1996. Physiological diversity within and among wood mice(Apodemus) species in Israel. Isr. J. Zool. 42, 347–351.
Haim, A., Shanas, U., Scantlebury, M., 2005a. Comparative physiology of heat production inrodents under increasing salinity: effects of habitat and habits. Belg. J. Zool.135, 55–59.
Haim, A., Shanas, U., El Salam Zubidad, A., Scantlebury, M., 2005b. Seasonality and seasonsout of time—the thermoregulatory effects of light interference. Chronobiol. Int. 22,59–66.
Harrison, D.L., Bates, P.J.J., 1991. The Mammals of Arabia. Harrison Zoological Museum,Sevenoaks, Kent.
Heldmaier, G., Steinlechner, S., Rafael, J., Latteier, B., 1982. Photoperiodic and ambienttemperature as environmental cues for seasonal thermogenic adaptation in theDjungarian hamster Phodopus sungorus. Int. J. Biometeorol. 26, 339–345.
Hoffmann, K., 1979. Photoperiodic effects in the Djungarian hamster: one minute of lightduring darktime mimics influence of long photoperiods on testicular recrudescence,body weight and pelage colour. Cell. Mol. Life Sci. 35, 1529–1530.
Keppel, G., Wickens, T.D., 2004. Design and Analysis: A Researcher's Handbook. PrenticeHall, Upper Saddle River, NJ.
Nevo, E., Filippucci, G.M., Pavlicek, T., Gorlova, O., Shenbrot, G., Ivanitskaya, E., Beiles, A.,1998. Genotypic and phenotypic divergence of rodents (Acomys cahirinus and Apo-demus mystacinus) at “Evolution Canyon”: Micro- and macroscale parallelism. ActaTheriol. Suppl. 5, 9–34.
Ntshotsho, P., Van Aarde, R.J., Nicolson, S.W., Jackson, T.P., 2004. Renal physiology of twosouthern AfricanMastomys species (Rodentia; Muridae): a salt-loading experimentto assess concentrating ability. Comp. Biochem. Physiol. 139, 441–447.
Page, M.C., Braver, S.L., MacKinnon, D.P., 2003. Levine's Guide to SPSS for Analysis ofVariance. Eribaum, Nahwah, NJ.
Peinke, D.M., Brown, C.R., 1999. Osmoregulation and water balance in the springhare(Pedetes capensis). J. Comp. Physiol. B 169, 1–10.
Ron,U.,Haim,A., 2001.Howdehydration affects the thermoregulatoryandosmoregulatoryabilities of the golden spiny mouse Acomys russatus. Isr. J. Zool. 47, 15–20.
Sasidharan, T.O.,Goyal, S.P., Chand, P.,Ghosh, P.K.,1990.Theeffects ofwaterdeprivation andsalt loadonwater conservation efficiency in two Indian desert gerbils. J. Comp. Physiol.B 160, 413–422.
Scantlebury, M., Afik, D., Shanas, U., Shanas, U., Haim, A., 2002. Comparative non-shiveringthermogenesis in adjacent populations of the common spiny mouse (Acomyscahirinus) from opposite slopes: the effects of increasing salinity. J. Comp. Physiol. A172, 1–5.
Scantlebury, M., Shanas, U., Speakman, J.R., Kupshtein, H., Afik, D., Haim, A., 2003. Ener-getics and water economy of common spiny mice Acomys cahirinus from north- andsouth-facing slopes of a Mediterranean valley. Func. Ecol. 17, 178–185.
Scantlebury, M., Shanas, U., Afik, D., Haim, A., 2005. Comparative seasonal acclimatizationof food and energy consumption in adjacent populations of common spiny mice(Acomys cahirinus). J. Zool. (Lond.) 267, 323–328.
Schmidt-Nielsen, B., O'Dell, R., Osaki, H., 1961. Interdependence of urea and electrolytes inproduction of a concentrated urine. Am. J. Physiol. 200, 1125–1132.
Schmidt-Nielsen, K., 1964. Desert Animals: Physiological Problems of Heat and Water.Clarendon Press, Oxford.
Shanas, U., Afik, D., Scantlebury, M., Haim, A., 2002. The effects of season and dietary saltcontent on body temperature daily rhythms of common spiny mice from differentmicro-habitats. Comp. Biochem. Physiol., Part A Mol. Integr. Physiol. 132, 287–295.
Shanas,U., Afik,D., Scantlebury,M., Haim,A., 2003.Differential osmoregulatorycapabilitiesof commonspinymice (Acomys cahirinus) fromadjacentmicrohabitats. J. Zool. (Lond.)261, 7–13.
Shkolnik, A., Borut, A., 1969. Temperature and water relations in two species of spinymice(Acomys). J. Mammal. 50, 245–255.
Speakman, E., Weldy, N.J., 2001. Body Fluids and Electrolytes, 8th ed. Mosby Incorporated,London.
Spiegel,M., Haim,A., 2004.Daily rhythms of nonshivering thermogenesis and responses tophotoperiodmanipulations in Apodemusmystacinus from two different ecosystems. J.Therm. Biol. 29, 635–640.
Thuiller, W., 2007. Climate change and the ecologist. Nature 448, 550–552.Weissenberg, S., Shkolnik, A., 1994. Metabolic rate and water economy in the desert and
Mediterranean populations of the common spiny mouse (Acomys cahirinus) in Israel.Isr. J. Zool. (Lond.) 40, 135–143.
Wilmer, P., Stone, G., Johnston, I., 2005. Environ. Physiol. Anim. Blackwell, Oxford.





























