Embed Size (px)
Citation preview

Neurobiology Research ReviewAn Overview of Recent Publications Featuring Illumina Technology

3 An Overview of Publications Featuring Illumina® Technology
CONTENTS
4 IntroductionDiseases
Schizophrenia
Autism Spectrum Disorder
Alzheimer’s Disease
Parkinson’s Disease
23 Genetic MechanismsCopy-Number Variants
Alternative Splicing
Epigenetic Modifications
Small RNAs
Genetic Variants
37 Model Systems Animal Models
43 Stem-Cell Models
45 BiologyImmunity
47 Metagenomics
49 Environmental Factors
50 Single Cells
52 Bibliography

4 Neurobiology Research Review
INTRODUCTION
Neurological disorders are complex diseases caused by a combination of
genetic, environmental, and lifestyle factors. Most neurological diseases—such
as schizophrenia, autism, and Alzheimer’s and Parkinson’s disorders—have been
described many decades ago. However, it is only recently, with the use of next-
generation sequencing (NGS), that their full complexity is being revealed.1,2 There
is an increasing awareness that disease development is driven through a complex
interplay between somatic (non-inherited) mutations, inherited mutations, and
epigenetic modifications.3,4,5,6 This complexity results in extremely weak genotype-
phenotype correlations, and the same disease can present with a variety of
pathological phenotypes in different individuals. Not surprisingly, different neurological
diseases present a spectrum of similar symptomatic profiles (such as dementia, e.g.,
Parkinson’s disease and dementia with Lewy bodies).7 These overlapping symptoms
could indicate the involvement of the same underlying molecular mechanisms. A
diagnosis based on these underlying molecular processes, as well as the observed
phenotype, promises more objective and accurate diagnosis and treatment in
the future.8
The increased information obtained from NGS, along with a variety of library
preparation methods, offers an impressive armamentarium of tools to unravel the
genomic and epigenomic aspects of psychiatric and neurodegenerative diseases.
The improved understanding also could translate ultimately into the development of
new, effective therapies. Additionally, whole-genome sequencing (WGS) potentially
could detect predisposition to these disorders, to allow preventative care and
early intervention. A recently announced massive 100,000 Genomes project,
initiated by Genomics England, signifies the importance of genomic diagnostics for
tomorrow’s medicine.
Diagnostic yield for patients with severe intellectual disability (IQ < 50), specified by technology: genomic microarrays, whole-exome sequencing (WES), and WGS. Percentages indicate the number of patients in whom a conclusive cause was identified using the specified technique.9
1. Bras J., Guerreiro R. and Hardy J. (2012) Use of next-generation sequencing and other whole-genome strategies to dissect neurologi-cal disease. Nat Rev Neurosci 13: 453-464
2. Koboldt D. C., Steinberg K. M., Larson D. E., Wilson R. K. and Mardis E. R. (2013) The next-generation sequencing revolution and its impact on genomics. Cell 155: 27-38
3. Yu T. W., Chahrour M. H., Coulter M. E., Jiralerspong S., Okamura-Ikeda K., et al. (2013) Using whole-exome sequencing to identify inherited causes of autism. Neuron 77: 259-273
4. Fromer M., Pocklington A. J., Kavanagh D. H., Williams H. J., Dwyer S., et al. (2014) De novo mutations in schizophrenia implicate synaptic networks. Nature 506: 179-184
5. De Jager P. L., Srivastava G., Lunnon K., Burgess J., Schalkwyk L. C., et al. (2014) Alzheimer's disease: early alterations in brain DNA methylation at ANK1, BIN1, RHBDF2 and other loci. Nat Neurosci 17: 1156-1163
6. Chung S. J., Armasu S. M., Anderson K. J., Biernacka J. M., Lesnick T. G., et al. (2013) Genetic susceptibility loci, environmental expo-sures, and Parkinson's disease: a case-con-trol study of gene-environment interactions. Parkinsonism Relat Disord 19: 595-599
7. McCarthy S. E., Gillis J., Kramer M., Lihm J., Yoon S., et al. (2014) De novo mutations in schizophrenia implicate chromatin remodeling and support a genetic overlap with autism and intellectual disability. Mol Psychiatry 19: 652-658
8. Fromer M., Pocklington A. J., Kavanagh D. H., Williams H. J., Dwyer S., et al. (2014) De novo mutations in schizophrenia implicate synaptic networks. Nature 506: 179-184
9. Gilissen C., Hehir-Kwa J. Y., Thung D. T., van de Vorst M., van Bon B. W., et al. (2014) Genome sequencing identifies major causes of severe intellectual disability. Nature 511: 344-347

5 An Overview of Publications Featuring Illumina® Technology
Diseases
Neurological diseases represent a daunting spectrum of complex multifactorial
pathologies, ranging from subtle to life-threatening. To illustrate the most recent
research and the use of genomics, this review focuses on schizophrenia and
autism as examples of complex neurodevelopmental disorders, and Alzheimer’s
and Parkinson’s as examples of neurodegenerative diseases. The approaches
and techniques used in these studies can be applied to a wide variety of
neurological diseases.
Schizophrenia
Schizophrenia is one of the most complex psychiatric diseases, and it affects as
much as 1% of the global adult population.10 The most common symptoms include
irrational thinking, auditory hallucinations, false beliefs, and reduced social activity.
The disease typically develops between 12 and 25 years of age and is a highly
heritable, polygenic disorder. Recent genomic analysis studies have shown that
schizophrenia is attributed to over a thousand gene loci, many of which appear
in non-coding parts of the genome.11-15 A significant subset of risk alleles for
schizophrenia is also implicated across other diseases in this diagnostic category,
such as bipolar disorder, autism, and depression.16,17,18 All these diseases are truly
spectrum disorders, which has complicated understanding of their genetic causes
and the development of targeted therapies. However, the advent of high-throughput
gene sequencing technology provides a tool for deeper analysis of the genetic basis
of these diseases, and it holds promise for unraveling the complex interplay between
multiple genetic and epigenetic modifications.
Genotype-phenotype correlations in schizophrenia are extremely weak, and the same
disease can present with a variety of pathological phenotypes in different patients.
Studies with twins revealed that over 80% of the risk of developing schizophrenia
comes from genetic predisposition, but exposure to environmental risk factors can
play a significant role.20 The first genetic mutations detected in schizophrenia were
rare variants, including copy-number variants (CNVs).21 Altogether, these variants
account for almost 20% of disease cases.22 The remaining genetic hits are most likely
represented by common variants.23 Presumably, the effects of individual common
variants are mild; however, in combination, they may be sufficient to trigger the onset
of schizophrenia.24,25
“Schizophrenia liability is being mapped to hundreds, perhaps ultimately more than a thousand, genetic loci, each contributing a small increment of risk.” Hyman 2014
10. Elert E. (2014) Aetiology: Searching for schizo-phrenia's roots. Nature 508: S2-3
11. Ripke S., O'Dushlaine C., Chambert K., Moran J. L., Kahler A. K., et al. (2013) Genome-wide association analysis identifies 13 new risk loci for schizophrenia. Nat Genet 45: 1150-1159
12. Purcell S. M., Moran J. L., Fromer M., Ruderfer D., Solovieff N., et al. (2014) A polygenic bur-den of rare disruptive mutations in schizophre-nia. Nature 506: 185-190
13. Hyman S. E. (2014) Perspective: Revealing molecular secrets. Nature 508: S20
14. Fromer M., Pocklington A. J., Kavanagh D. H., Williams H. J., Dwyer S., et al. (2014) De novo mutations in schizophrenia implicate synaptic networks. Nature 506: 179-184
15. McCarroll S. A. and Hyman S. E. (2013) Progress in the genetics of polygenic brain disorders: significant new challenges for neu-robiology. Neuron 80: 578-587
16. Kirov G., Rees E., Walters J. T., Escott-Price V., Georgieva L., et al. (2014) The penetrance of copy number variations for schizophrenia and developmental delay. Biol Psychiatry 75: 378-385
17. Cross-Disorder Group of the Psychiatric Ge-nomics C., Lee S. H., Ripke S., Neale B. M., Faraone S. V., et al. (2013) Genetic relationship between five psychiatric disorders estimated from genome-wide SNPs. Nat Genet 45: 984-994
18. Cukier H. N., Dueker N. D., Slifer S. H., Lee J. M., Whitehead P. L., et al. (2014) Exome sequencing of extended families with autism reveals genes shared across neurodevelop-mental and neuropsychiatric disorders. Mol Autism 5: 1
19. Wright J. (2014) Genetics: Unravelling com-plexity. Nature 508: S6-7
20. Hyman S. E. (2014) Perspective: Revealing molecular secrets. Nature 508: S20
21. Wright J. (2014) Genetics: Unravelling complexity. Nature 508: S6-7
22. Wright J. (2014) Genetics: Unravelling complexity. Nature 508: S6-7
23. Wright J. (2014) Genetics: Unravelling complexity. Nature 508: S6-7
24. Wright J. (2014) Genetics: Unravelling complexity. Nature 508: S6-7
25. Gaugler T., Klei L., Sanders S. J., Bodea C. A., Goldberg A. P., et al. (2014) Most genetic risk for autism resides with common variation. Nat Genet 46: 881-885

6 Neurobiology Research Review
Along with determining the individual mutations leading to schizophrenia, the power
of genetic analysis is expected to give answers to some other important questions,
such as why the development of schizophrenia is associated with accelerated
ageing26 or why schizophrenic patients suffer from heart, lung, and metabolic
diseases at young ages and at a much higher rate than the general population.29
These effects could be due to additional lifestyle risks, such as substance abuse and
smoking. Approximately 50% of patients with chronic schizophrenia have substance-
use disorder, and their risk to develop this disorder is 4.6 times higher than for the
general population. Over 80% of schizophrenic patients in the U.S. are also heavy
smokers,30 as nicotine serves as an agonist of the nicotine acetylcholine receptor and
presumably can attenuate some cognitive impairment associated
with schizophrenia.31
Smoking and substance abuse are common among schizophrenia patients. Over 80% of schizophrenic patients in the U.S. are heavy smokers,32 as nicotine presumably can attenuate some of the cognitive impairment associated with schizophrenia.
Early, accurate, and objective diagnosis of the underlying molecular mechanisms
of the disease will allow a better control of the disease symptoms with available
therapies. In the more distant future, an improved understanding of the disease
should lead to the development of more effective, targeted, and
personalized therapies.
ReviewsAnthes E. (2014) Ageing: Live faster, die younger. Nature 508: S16-17
Brody H. (2014) Schizophrenia. Nature 508: S1
Dolgin E. (2014) Therapeutics: Negative feedback. Nature 508: S10-11
Elert E. (2014) Aetiology: Searching for schizophrenia's roots. Nature 508: S2-3
Horvath S. and Mirnics K. (2014) Immune system disturbances in schizophrenia. Biol Psychiatry 75: 316-323
Hyman S. E. (2014) Perspective: Revealing molecular secrets. Nature 508: S20
Singh S., Kumar A., Agarwal S., Phadke S. R. and Jaiswal Y. (2014) Genetic insight of schizophrenia: past and future perspectives. Gene 535: 97-100
Schizophrenia Working Group of the Psychiatric Genomics C. (2014) Biological insights from 108 schizophrenia-associated genetic loci. Nature 511: 421-427
Wright J. (2014) Genetics: Unravelling complexity. Nature 508: S6-7
Malhotra D. and Sebat J. (2013) CNVs: harbingers of a rare variant revolution in psychiatric genetics. Cell 148: 1223-1241
Schreiber M., Dorschner M. and Tsuang D. (2013) Next-generation sequencing in schizophrenia and other neuropsychiatric disorders. Am J Med Genet B Neuropsychiatr Genet 162B: 671-678
26. Brody H. (2014) Schizophrenia. Nature 508: S1
27. Anthes E. (2014) Ageing: Live faster, die younger. Nature 508: S16-17
28. Hyman S. E. (2014) Perspective: Revealing molecular secrets. Nature 508: S20
29. Dixon L. (1999) Dual diagnosis of substance abuse in schizophrenia: prevalence and impact on outcomes. Schizophr Res 35 Suppl: S93-100
30. de Leon J. and Diaz F. J. (2005) A meta-anal-ysis of worldwide studies demonstrates an association between schizophrenia and tobacco smoking behaviors. Schizophr Res 76: 135-157
31. Ng E., McGirr A., Wong A. H. and Roder J. C. (2013) Using rodents to model schizophrenia and substance use comorbidity. Neurosci Biobehav Rev 37: 896-910
32. de Leon J. and Diaz F. J. (2005) A meta-anal-ysis of worldwide studies demonstrates an association between schizophrenia and tobacco smoking behaviors. Schizophr Res 76: 135-157

7 An Overview of Publications Featuring Illumina® Technology
ReferencesFromer M., Pocklington A. J., Kavanagh D. H., Williams H. J., Dwyer S., et al. (2014) De novo mutations in schizophrenia implicate synaptic networks. Nature 506: 179-184Of the known risk alleles for schizophrenia, the only ones definitively shown to confer considerable increments in risk are rare chromosomal CNVs that involve deletion or duplication of thousands of bases of DNA. This study examined the effect of small de novo mutations affecting one or a few nucleotides. By Illumina HiSeq WES of 623 schizophrenia trios, the authors assessed de novo mutation rates and shared genetic etiology for schizophrenia, intellectual disability, and autism-spectrum disorders (ASDs). They found several insights to suggest a common etiological mechanism.
Illumina Technology: HiSeq for exome sequencing
Karayannis T., Au E., Patel J. C., Kruglikov I., Markx S., et al. (2014) Cntnap4 differentially contributes to GABAergic and dopaminergic synaptic transmission. Nature 511: 236-240In an effort to understand the pathological development of neurological disorders, genetic effects have been studied in the context of proteins that are expressed in neural cells. In this study, the authors characterized the effect of CNTNAP4 knockouts on mouse behavior and development, and relate these results to the findings of CNVs in humans across a region including the CNTNAP2 gene. The authors found that CNTNAP4 is localized presynaptically, and its loss leads to a reduction in the output of cortical parvalbumin (PV)-positive GABAergic basket cells. In addition, CNTNAP4-mutant mice showed defects in these neuronal populations and exhibited sensory-motor gating and grooming endophenotypes.
Illumina Technology: HumanHap550, HumanOmni1-Quad
Purcell S. M., Moran J. L., Fromer M., Ruderfer D., Solovieff N., et al. (2014) A polygenic burden of rare disruptive mutations in schizophrenia. Nature 506: 185-190Identifying gene associations for complex genetic diseases remains challenging, with small sample sizes being a hindrance for finding significant effects. In this study of schizophrenia, the authors performed WES, based on Illumina technology, of 2,536 schizophrenia cases and 2,543 controls. They identified disruptive mutations distributed across many genes; however, no individual gene-based test for low frequency and moderately large effect achieves significance after correction for multiple testing.
Illumina Technology: HiSeq 2000, Genome AnalyzerIIx
Schizophrenia Working Group of the Psychiatric Genomics C. (2014) Biological insights from 108 schizophrenia-associated genetic loci. Nature 511: 421-427Schizophrenia is a highly heritable disorder, but the heritability is not found in a single gene effect. In this largest genome-wide association study (GWAS) for schizophrenia to date, the authors used single-nucleotide polymorphism (SNP) arrays for 36,989 cases and 113,075 controls to determine genetic risk factors for the disorder. The authors found that the significant genetic associations were not randomly spread across the genome, but enriched among genes expressed in brain and genes that have been associated with typical co-morbidity diagnoses, such as ASD and intellectual disability. Interestingly, links were also enriched within genes related to immunity, which fits the existing hypothesis of immune dysregulation in schizophrenia.
Illumina Technology: Human1M, HumanOmni2.5, HumanOmniExpress, HumanHap550, Human610, HumanHap650Y, HumanHap300
Stefansson H., Meyer-Lindenberg A., Steinberg S., Magnusdottir B., Morgen K., et al. (2014) CNVs conferring risk of autism or schizophrenia affect cognition in controls. Nature 505: 361-366Certain CNVs contribute to the pathogenesis of schizophrenia and autism. In this study, the authors investigated the influence of these CNVs on phenotypes separate from those of the mentioned diseases. In a big population-wide study of nearly a third of the Icelandic population (n = 101,655), the authors used Illumina SNP microarrays to test for associations of CNVs with cognitive deficits, dyslexia, dyscalculia, and brain structure changes. The authors found that the 15q11.2(BP1-BP2) deletion affects brain structure in a pattern consistent with first-episode psychosis in both schizophrenia and dyslexia.
Illumina Technology: HumanHap300, HumanCNV370-Duo, HumanHap650Y, Human1M, HumanOmni2.5, HumanOmniExpress, HumanOmni1S

8 Neurobiology Research Review
Yoon K. J., Nguyen H. N., Ursini G., Zhang F., Kim N. S., et al. (2014) Modeling a genetic risk for schizophrenia in iPSCs and mice reveals neural stem cell deficits associated with adherens junctions and polarity. Cell Stem Cell 15: 79-91Defects in brain development can contribute to the onset of neuropsychiatric disorders. This study set out to identify the functional role of the 15q11.2 deletion on neural development using induced pluripotent stem cell (iPSC)-derived human neural precursor cells (hNPCs) by RNA-Seq and SNP-genotyping arrays. They found that haploinsufficiency of CYFIP1, a gene within 15q11.2, affects radial glial cells, leading to their ectopic localization outside of the ventricular zone.
Illumina Technology: HumanOmni2.5S
Stoll G., Pietilainen O. P., Linder B., Suvisaari J., Brosi C., et al. (2013) Deletion of TOP3beta, a component of FMRP-containing mRNPs, contributes to neurodevelopmental disorders. Nat Neurosci 16: 1228-1237Genetic studies, including studies of mRNA-binding proteins, have shed new light on the connection of mRNA metabolism to disease. In this study, the authors found that deletion of the TOP3b gene was associated with neurodevelopmental disorders in the Northern Finnish population. Combining the genotyping with photoactivatable ribonucleoside–enhanced crosslinking and immunoprecipitation (PAR-CLIP), the authors found that the recruitment of TOP3b to cytosolic messenger ribonucleoproteins (mRNPs) was coupled to the co-recruitment of FMRP, the disease gene involved in Fragile X syndrome.
Illumina Technology: Human Gene Expression, Human610-Quad, HumanHap300, HumanCNV370-Duo
RNase T1 digestion
UV 365 nm
cDNA
Proteinase K
RNA-protein complex
RNA extraction and reverse transcription
Incorporate 4-thiouridine (4SU) into transcripts of cultured cells
Photoactivatable ribonucleoside–enhanced crosslinking and immunoprecipitation (PAR-CLIP) maps RNA-binding proteins (RBPs).33 This approach is similar to high-throughput sequencing of RNA isolated by crosslinking immunoprecipitation (HITS-CLIP) and cross-linking immunoprecipitation sequencing (CLIP-Seq), but uses much more efficient crosslinking to stabilize the protein-RNA complexes. The requirement for a photoactivatable ribonucleoside limits this approach to cell culture and in vitro systems. In this method, 4-thiouridine (4-SU) and 6-thioguanosine (6-SG) are incorporated into transcripts of cultured cells. Ultraviolet irradiation crosslinks 4-SU/6-SG–labeled transcripts to interacting RBPs. The targeted complexes are immunoprecipitated and digested with RNase T1, followed by Proteinase K, before RNA extraction. The RNA is reverse-transcribed to cDNA and sequenced. Deep sequencing of cDNA accurately maps RBPs interacting with labeled transcripts. (For more methods, see: http://applications.illumina.com/applications/sequencing/ngs-library-prep/library-prep-methods.ilmn)
Ionita-Laza I., Xu B., Makarov V., Buxbaum J. D., Roos J. L., et al. (2014) Scan statistic-based analysis of exome sequencing data identifies FAN1 at 15q13.3 as a susceptibility gene for schizophrenia and autism. Proc Natl Acad Sci U S A 111: 343-348
McCarthy S. E., Gillis J., Kramer M., Lihm J., Yoon S., et al. (2014) De novo mutations in schizophrenia implicate chromatin remodeling and support a genetic overlap with autism and intellectual disability. Mol Psychiatry 19: 652-658
Todarello G., Feng N., Kolachana B. S., Li C., Vakkalanka R., et al. (2014) Incomplete penetrance of NRXN1 deletions in families with schizophrenia. Schizophr Res 155: 1-7
Ripke S., O'Dushlaine C., Chambert K., Moran J. L., Kahler A. K., et al. (2013) Genome-wide association analysis identifies 13 new risk loci for schizophrenia. Nat Genet 45: 1150-1159
33. Hafner M., Landgraf P., Ludwig J., Rice A., Ojo T., et al. (2008) Identification of microRNAs and other small regulatory RNAs using cDNA library sequencing. Methods 44: 3-12

9 An Overview of Publications Featuring Illumina® Technology
Autism Spectrum Disorder
Autism spectrum disorder (ASD) comprises a group of polygenic, multi-locus
disorders,34 often accompanied by symptoms of other disorders, such as
developmental disability/intellectual disability (DD/ID; over 40% of ASD cases),
attention deficit/hyperactivity disorder (ADHD; 59%–75%), obsessive-compulsive
disorder (OCD; 60%), epilepsy (7%–46%), and other neurological and behavioral
patterns.35,36 Autism rates have been increasing rapidly, from 0.7% of the population
in 2000 to 1.1% in early 2010.37 In the U.S., 1 in 68 children has been diagnosed
as autistic. This trend can be partially attributed to the improved diagnosis of the
disease. Improved understanding of the genetic causes of ASD is anticipated to
facilitate development of palliative or therapeutic care for affected individuals. It is
also expected to be instrumental in providing a more accurate method to assess the
mental condition of at-risk populations, such as criminals and individuals with other
psychiatric pathologies.38,39,40
Autism is primarily a genetic disease, with 15–30 times increased risk of disease
development in siblings of autistic children.41 Heritability of this disease has
been estimated as high as 90%–96%, suggesting yet unidentified non-genetic
causes.42,43,44 A more accurate, systematic approach is still needed to improve
the distinction between essential autism and complex (syndromic, sporadic)
autism (Table 1). Genomic approaches promise to be efficient and reliable tools for
distinguishing between various types of autism.45
Table 1: Types of autism
Type of DiseasePercentage
of CasesDisease Characteristics
Essential autism46w 75% Higher male to female ratioLack of dysmorphic featuresHigher sibling recurrence riskPositive family historyCommon gene variants
Complex (syndromic, sporadic) autism47
25% Large number of highly penetrant rare mutations
Similar to schizophrenia, ASD is a heterogenic disorder.48 This heterogeneity may be
observed not only across individuals, but even across different sections of brain.
Gene expression may change based on the timing of the analysis, as shown in
experiments with mice.49
“Two years from now, researchers will need a larger T-shirt to flaunt their findings.” Wright 2014
34. Losh M., Childress D., Lam K. and Piven J. (2008) Defining key features of the broad autism phenotype: a comparison across parents of multiple- and single-incidence autism families. Am J Med Genet B Neuropsychiatr Genet 147B: 424-433
35. Buxbaum J. (2013) The Neuroscience of Autism Spectrum Disorders.
36. Lee H., Lin M. C., Kornblum H. I., Papazian D. M. and Nelson S. F. (2014) Exome sequencing identifies de novo gain of function missense mutation in KCND2 in identical twins with autism and seizures that slows potassium channel inactivation. Hum Mol Genet 23: 3481-3489
37. Gilbert J. A., Krajmalnik-Brown R., Porazinska D. L., Weiss S. J. and Knight R. (2013) Toward effective probiotics for autism and other neuro-developmental disorders. Cell 155: 1446-1448
38. King C. and Murphy G. H. (2014) A Systematic Review of People with Autism Spectrum Disor-der and the Criminal Justice System. J Autism Dev Disord
39. Gadow K. D. (2013) Association of schizophre-nia spectrum and autism spectrum disorder (ASD) symptoms in children with ASD and clinic controls. Res Dev Disabil 34: 1289-1299
40. Smith K. R. and Matson J. L. (2010) Social skills: differences among adults with intellectual disabil-ities, co-morbid autism spectrum disorders and epilepsy. Res Dev Disabil 31: 1366-1372
41. Szatmari P. (1999) Heterogeneity and the genet-ics of autism. J Psychiatry Neurosci 24: 159-165
42. Rosti R. O., Sadek A. A., Vaux K. K. and Gleeson J. G. (2014) The genetic landscape of autism spectrum disorders. Dev Med Child Neurol 56: 12-18
43. Buxbaum J. (2013) The Neuroscience of Autism Spectrum Disorders.
44. Nava C., Keren B., Mignot C., Rastetter A., Chantot-Bastaraud S., et al. (2014) Prospective diagnostic analysis of copy number variants using SNP microarrays in individuals with autism spectrum disorders. Eur J Hum Genet 22: 71-78
45. Rosti R. O., Sadek A. A., Vaux K. K. and Gleeson J. G. (2014) The genetic landscape of autism spectrum disorders. Dev Med Child Neurol 56: 12-18
46. Rosti R. O., Sadek A. A., Vaux K. K. and Gleeson J. G. (2014) The genetic landscape of autism spectrum disorders. Dev Med Child Neurol 56: 12-18
47. Ghosh A., Michalon A., Lindemann L., Fontoura P. and Santarelli L. (2013) Drug discovery for autism spectrum disorder: challenges and op-portunities. Nat Rev Drug Discov 12: 777-790
48. Willsey A. J., Sanders S. J., Li M., Dong S., Tebbenkamp A. T., et al. (2013) Coexpression networks implicate human midfetal deep cortical projection neurons in the pathogenesis of autism. Cell 155: 997-1007
49. Willsey A. J., Sanders S. J., Li M., Dong S., Tebbenkamp A. T., et al. (2013) Coexpression networks implicate human midfetal deep cortical projection neurons in the pathogenesis of autism. Cell 155: 997-1007

10 Neurobiology Research Review
CNVs were the first type of mutation associated with autism.50,51 The de novo rate of
CNVs is three to seven times higher than in controls. The gene groups most affected
by autism-associated CNVs are GTPase/Ras, ubiquitin degradation genes, and
genes involved in synapse development, axon targeting, and neuron motility.52,53,54
Large CNVs are present in 5%–10% of ASD patients, primarily in those with a
syndromic ASD phenotype.55,56,57 Private CNVs in autistic individuals often affect
genes encoding synaptic proteins and neuronal cell-adhesion proteins.58
(see Copy-Number Variants)
Furthermore, ASD can also be caused by rare mutations, deletions, duplications,59
and larger chromosomal abnormalities, which may be inherited or arise de novo.60
Monogenic mutations known to date contribute to 2%–5% of syndromic cases, with
Fragile X chromosome, PTEN macrocephaly, and tuberous sclerosis being the most
common abnormalities.61,62 PTEN mutations are also strongly associated with
tumor syndromes.63
Twin studies have become a standard model in the research of psychiatric diseases. They allow assessment of the contribution of genes and the environment to disease risk.
Recent WES and WGS studies have identified multiple, high-confidence ASD
genes.64 Of two large-scale WGS projects, one was initiated by the U.K. government
in collaboration with Illumina and the Wellcome Trust (100,000 Genomes project),
and the second was initiated by Beijing Genomics Institute (BGI) in collaboration with
Autism Speaks (Autism Genome 10K project). The pilot results of the latter study
have been published by Jiang et al.65
50. Iafrate A. J., Feuk L., Rivera M. N., Listewnik M. L., Donahoe P. K., et al. (2004) Detection of large-scale variation in the human genome. Nat Genet 36: 949-951
51. Redon R., Ishikawa S., Fitch K. R., Feuk L., Perry G. H., et al. (2006) Global variation in copy number in the human genome. Nature 444: 444-454
52. Glessner J. T., Wang K., Cai G., Korvatska O., Kim C. E., et al. (2009) Autism genome-wide copy number variation reveals ubiquitin and neuronal genes. Nature 459: 569-573
53. Gilman S. R., Iossifov I., Levy D., Ronemus M., Wigler M., et al. (2011) Rare de novo variants as-sociated with autism implicate a large functional network of genes involved in formation and function of synapses. Neuron 70: 898-907
54. Sakai Y., Shaw C. A., Dawson B. C., Dugas D. V., Al-Mohtaseb Z., et al. (2011) Protein interac-tome reveals converging molecular pathways among autism disorders. Sci Transl Med 3: 86ra49
55. Sanders S. J., Ercan-Sencicek A. G., Hus V., Luo R., Murtha M. T., et al. (2011) Multiple recur-rent de novo CNVs, including duplications of the 7q11.23 Williams syndrome region, are strongly associated with autism. Neuron 70: 863-885
56. Levy D., Ronemus M., Yamrom B., Lee Y. H., Leotta A., et al. (2011) Rare de novo and transmitted copy-number variation in autistic spectrum disorders. Neuron 70: 886-897
57. Guilmatre A., Dubourg C., Mosca A. L., Legallic S., Goldenberg A., et al. (2009) Recurrent rearrangements in synaptic and neurodevelop-mental genes and shared biologic pathways in schizophrenia, autism, and mental retardation. Arch Gen Psychiatry 66: 947-956
58. Nava C., Keren B., Mignot C., Rastetter A., Chantot-Bastaraud S., et al. (2014) Prospective diagnostic analysis of copy number variants using SNP microarrays in individuals with autism spectrum disorders. Eur J Hum Genet 22: 71-78.
59. Banerjee S., Riordan M. and Bhat M. A. (2014) Genetic aspects of autism spectrum disorders: insights from animal models. Front Cell Neurosci 8: 58
60. Nava C., Keren B., Mignot C., Rastetter A., Chantot-Bastaraud S., et al. (2014) Prospective diagnostic analysis of copy number variants using SNP microarrays in individuals with autism spectrum disorders. Eur J Hum Genet 22: 71-78
61. Miles J. H. (2011) Autism spectrum disorders--a genetics review. Genet Med 13: 278-294
62. Schaaf C. P. and Zoghbi H. Y. (2011) Solving the autism puzzle a few pieces at a time. Neuron 70: 806-808
63. Rosti R. O., Sadek A. A., Vaux K. K. and Gleeson J. G. (2014) The genetic landscape of autism spectrum disorders. Dev Med Child Neurol 56: 12-18
64. Willsey A. J., Sanders S. J., Li M., Dong S., Tebbenkamp A. T., et al. (2013) Coexpression networks implicate human midfetal deep cortical projection neurons in the pathogenesis of autism. Cell 155: 997-1007
65. Jiang Y. H., Yuen R. K., Jin X., Wang M., Chen N., et al. (2013) Detection of clinically relevant genetic variants in autism spectrum disorder by whole-genome sequencing. Am J Hum Genet 93: 249-263

11 An Overview of Publications Featuring Illumina® Technology
EpigeneticsSusceptibility to ASD can arise at both the genetic and epigenetic levels.66 Several
groups have independently identified multiple differentially methylated regions (DMRs)
in post-mortem samples of autistic individuals. These biologically diverse gene
regions include DNase hypersensitive sites and an alternative transcript termination
site.67,68,69 These studies provide an additional level of evidence for the role of
epigenetics in complex diseases, such as ASD.
ReviewsBanerjee S., Riordan M. and Bhat M. A. (2014) Genetic aspects of autism spectrum disorders: insights from animal models. Front Cell Neurosci 8: 58
Rosti R. O., Sadek A. A., Vaux K. K. and Gleeson J. G. (2014) The genetic landscape of autism spectrum disorders. Dev Med Child Neurol 56: 12-18
Buxbaum J. (2013) The Neuroscience of Autism Spectrum Disorders.
Ebert D. H. and Greenberg M. E. (2013) Activity-dependent neuronal signalling and autism spectrum disorder. Nature 493: 327-337
Gadad B. S., Hewitson L., Young K. A. and German D. C. (2013) Neuropathology and animal models of autism: genetic and environmental factors. Autism Res Treat 2013: 731935
Ghosh A., Michalon A., Lindemann L., Fontoura P. and Santarelli L. (2013) Drug discovery for autism spectrum disorder: challenges and opportunities. Nat Rev Drug Discov 12: 777-790
Persico A. M. and Napolioni V. (2013) Autism genetics. Behav Brain Res 251: 95-112
Wohr M. and Scattoni M. L. (2013) Behavioural methods used in rodent models of autism spectrum disorders: current standards and new developments. Behav Brain Res 251: 5-17
Malhotra D. and Sebat J. (2012) CNVs: harbingers of a rare variant revolution in psychiatric genetics. Cell 148: 1223-1241
ReferencesGaugler T., Klei L., Sanders S. J., Bodea C. A., Goldberg A. P., et al. (2014) Most genetic risk for autism resides with common variation. Nat Genet 46: 881-885Although ASDs have been studied widely, the proportion and nature of genetic heritability is uncertain. This study analyzed the largest autism cohort data to date, including 1.6 million Swedish families with at least two children and ~5,700 individuals with strict autism diagnoses. Using Illumina SNP arrays, the authors investigated the contribution of rare versus common genetic variants to the disease. They conclude that the heritability is ~52.4%, with common variation as the biggest contributor. Rare, de novo mutations contribute substantially to individual liability, yet their contribution to variance in liability is modest at 2.6%.
Illumina Technology: OmniExpress and Exome
Lee H., Lin M. C., Kornblum H. I., Papazian D. M. and Nelson S. F. (2014) Exome sequencing identifies de novo gain of function missense mutation in KCND2 in identical twins with autism and seizures that slows potassium channel inactivation. Hum Mol Genet 23: 3481-3489Numerous studies have reported comorbidity of autism and epilepsy, but the relationship between the two disorders is unknown. In this study, identical twins, affected by both autism and severe intractable seizures, were studied using exome sequencing. A novel variant in the KCND2 gene was observed in both twins. The de novo mutation is located in the protein coding the Kv4.2 potassium channel, and the authors expressed the mutant protein in Xenopus oocytes to observe functional effects. Expression analysis showed that the mutation dominantly impairs the closed-state inactivation of the potassium channel, strongly supporting KCND2 as the causal gene for epilepsy in this family.
Illumina Technology: HiSeq 2000, Illumina Paired-End Sequencing Library Prep Protocol
66. Wong C. C., Meaburn E. L., Ronald A., Price T. S., Jeffries A. R., et al. (2014) Methylomic analysis of monozygotic twins discordant for autism spectrum disorder and related be-havioural traits. Mol Psychiatry 19: 495-503
67. Ladd-Acosta C., Hansen K. D., Briem E., Fallin M. D., Kaufmann W. E., et al. (2013) Common DNA methylation alterations in multiple brain regions in autism. Mol Psychiatry
68. Ladd-Acosta C., Hansen K. D., Briem E., Fallin M. D., Kaufmann W. E., et al. (2013) Common DNA methylation alterations in multiple brain regions in autism. Mol Psychiatry
69. Wong C. C., Meaburn E. L., Ronald A., Price T. S., Jeffries A. R., et al. (2014) Methylomic analysis of monozygotic twins discordant for autism spectrum disorder and related be-havioural traits. Mol Psychiatry 19: 495-503

12 Neurobiology Research Review
King I. F., Yandava C. N., Mabb A. M., Hsiao J. S., Huang H. S., et al. (2013) Topoisomerases facilitate transcription of long genes linked to autism. Nature 501: 58-62Topoisomerases are expressed throughout the developing and adult brain, and are mutated in some individuals with ASD. However, the mechanism by which topoisomerases impact ASD is unknown. By transcriptome sequencing, in combination with genome-wide mapping of RNA polymerase II density in neurons, the authors found that expression of long genes was reduced after knockdown of topoisomerase in neurons. The authors noted that many high-confidence ASD candidate genes are exceptionally long and were reduced in expression after TOP1 inhibition. This observation suggests that defective topoisomerases could contribute to ASD.
Illumina Technology: HiSeq 2000, TruSeq RNA, TruSeq for ChIP-Seq
Ladd-Acosta C., Hansen K. D., Briem E., Fallin M. D., Kaufmann W. E., et al. (2013) Common DNA methylation alterations in multiple brain regions in autism. Mol Psychiatry Genetic as well as environmental factors are responsible for ASDs. In this study, the authors measured over 485,000 CpG loci and identified 4 genome-wide significantly different DMRs from 19 autism cases in human brain tissue. This study highlights a new selected set of affected genes.
Illumina Technology: HumanMethylation450
Willsey A. J., Sanders S. J., Li M., Dong S., Tebbenkamp A. T., et al. (2013) Coexpression networks implicate human midfetal deep cortical projection neurons in the pathogenesis of autism. Cell 155: 997-1007Recent studies employing WES and WGS have identified nine high-confidence ASD genes. This study examined the contribution of these nine genes to the common phenotype by combining Illumina WES and RNA-Seq data into co-expression networks. The authors explain how these networks will guide future ASD research by indicating which genes are most likely to have overlapping molecular, cellular or circuit-level phenotypes.
Illumina Technology: HiSeq 2000, Genome Analyzer
Yu T. W., Chahrour M. H., Coulter M. E., Jiralerspong S., Okamura-Ikeda K., et al. (2013) Using whole-exome sequencing to identify inherited causes of autism. Neuron 77: 259-273Steel syndrome is a developmental structural disorder first described in 1993 in 23 Hispanic children from Puerto Rico. This paper presents the genomic analysis of a family with two affected siblings. The authors used whole-exome sequencing using the Baylor College of Medicine Human Genome Sequencing Center (BCM-HGSC) core design followed by sequencing of both affected siblings, parents, and an affected cousin and her unaffected parents. By filtering the detected genetic variants by segregation, the authors discovered a single homozygous missense variant segregating with the disorder. The variant disrupts the collagen gene COL27A1, which codes for a protein expressed in developing cartilage.
Illumina Technology: HiSeq 2000, HumanOmniExpress
Lionel A. C., Tammimies K., Vaags A. K., Rosenfeld J. A., Ahn J. W., et al. (2014) Disruption of the ASTN2/TRIM32 locus at 9q33.1 is a risk factor in males for autism spectrum disorders, ADHD and other neurodevelopmental phenotypes. Hum Mol Genet 23: 2752-2768
McCarthy S. E., Gillis J., Kramer M., Lihm J., Yoon S., et al. (2014) De novo mutations in schizophrenia implicate chromatin remodeling and support a genetic overlap with autism and intellectual disability. Mol Psychiatry 19: 652-658
Nava C., Keren B., Mignot C., Rastetter A., Chantot-Bastaraud S., et al. (2014) Prospective diagnostic analysis of copy number variants using SNP microarrays in individuals with autism spectrum disorders. Eur J Hum Genet 22: 71-78
Wong C. C., Meaburn E. L., Ronald A., Price T. S., Jeffries A. R., et al. (2014) Methylomic analysis of monozygotic twins discordant for autism spectrum disorder and related behavioural traits. Mol Psychiatry 19: 495-503

13 An Overview of Publications Featuring Illumina® Technology
Alzheimer’s Disease
Alzheimer’s disease (AD) is the most common form of dementia, affecting over 40
million people world-wide, and the incidence may double by the year 2050.70 Eleven
percent of people 65 or older and 32% of people 85 or older are affected by this fatal
neurodegenerative disorder.71 Early symptoms of AD include gradually worsening
ability to memorize new information,72 followed by confusion, irritability, trouble with
language, and long-term memory loss. The disease destroys memory and cognitive
skills through the accumulation of two types of abnormal insoluble aggregates in the
brain: extracellular ß-amyloid plaques and intracellular neurofibrillary tangles. The
aggregates disrupt the intricate interplay between brain neurons, which eventually
cause the death of the neurons with consequent significant shrinkage of the
brain volume.73
Pathogenic ß-amyloid protein belongs to a group of prion proteins, which kick-start
a chain reaction of destructive processes by engaging new ß-amyloid “seeds” into
formation of insoluble oligomers and spreading across the brain. However, unlike
“mad cow disease” (bovine spongiform encephalopathy or BSE) prions, AD-related
amyloidosis is not infectious and cannot be transmitted between individuals.74
Neurofibrillary tangles are formed intracellularly by oligomerization of the hyper-
phosphorylated form of Tau protein, which is normally abundant in axons and is
responsible for maintaining the structure of microtubules. During the disease, tau
protein is misfolded and mislocalized to the neuronal soma.75
Beta-amyloid protein is the major component of amyloid plaques.
Alzheimer’s disease can be present in one of two forms: early-onset AD (EOAD,
30–60 years old) and late-onset AD (LOAD) (Table 2). EOAD is mostly a genetic
disease, whereas LOAD is a sporadic disease, associated with a complex interplay
among different mutations.
70. Brookmeyer R., Johnson E., Ziegler-Graham K. and Arrighi H. M. (2007) Forecasting the global burden of Alzheimer's disease. Alzhei-mers Dement 3: 186-191
71. Lu H., Liu X., Deng Y. and Qing H. (2013) DNA methylation, a hand behind neurodegenerative diseases. Front Aging Neurosci 5: 85
72. Lu H., Liu X., Deng Y. and Qing H. (2013) DNA methylation, a hand behind neurodegenerative diseases. Front Aging Neurosci 5: 85
73. Huang Y. and Mucke L. (2012) Alzheimer mechanisms and therapeutic strategies. Cell 148: 1204-1222
74. Aguzzi A. and Rajendran L. (2009) The trans-cellular spread of cytosolic amyloids, prions, and prionoids. Neuron 64: 783-790
75. Huang Y. and Mucke L. (2012) Alzheimer mechanisms and therapeutic strategies. Cell 148: 1204-1222

14 Neurobiology Research Review
Table 2: Types of AD
Type of DiseaseAge of Onset
Percentage of Cases
Disease Characteristics
Early-onset AD (EOAD)
30–60 years 2%–5% Mostly genetic76
Late-onset AD (LOAD)
>60 years 95%–98%77 Strong role for epigenetic markers, such as DNA methylation78
EOAD is associated with mutations in three genes: amyloid precursor protein
(APP, integral Type I membrane glycoprotein) and presenelins PSEN1 and PSEN2.
However, these mutations reportedly account for fewer than 2% of all AD cases.79,80
Mutations in these genes deregulate the APP pathway and cause accumulation of
plaques built of ß-amyloid protein.81 Interestingly, some mutations in APP can be
protective against AD,82,83 highlighting the importance of deep genetic analysis and
correlative studies of this disease.
LOAD had, until recently, only one known genetic risk factor: the E4 variant of the
apolipoprotein E gene (APOE).84 The risk of developing LOAD in individuals with
two copies of APOE4 (2% of population) is as high as 60% at 85 years old, and
in individuals with one copy of this gene (25% of the population), it is 30%.85 Each
additional copy of APOE4 increases the risk of developing AD by a factor of three
or more.86 It is possible that low-frequency variants with large influence on LOAD
pathogeneses may have been missed by traditional GWAS. Sequence-based
association studies may be able to identify risk alleles in complex diseases, and it is
anticipated that these studies will elucidate low-frequency variants with large
effect sizes.87
Extracellular amyloid plaques stick to each other, as well as to the neuron. They disrupt neuronal networks, causing death of neurons and impairment of brain activity.
An interesting experimental approach proposes that WES be run on a thoroughly
selected subgroup of individuals at increased risk of AD, and this analysis can be
followed by a combination of genotyping and resequencing assays.88
76. Guerreiro R. J., Gustafson D. R. and Hardy J. (2012) The genetic architecture of Alzheimer's disease: beyond APP, PSENs and APOE. Neurobiol Aging 33: 437-456
77. Coppede F. (2014) The potential of epigenetic therapies in neurodegenerative diseases. Front Genet 5: 220
78. Mitsui J. and Tsuji S. (2014) Genomic Aspects of Sporadic Neurodegenerative Diseases. Biochem Biophys Res Commun 452: 221-225
79. Lu H., Liu X., Deng Y. and Qing H. (2013) DNA methylation, a hand behind neurodegenerative diseases. Front Aging Neurosci 5: 85
80. Guerreiro R. J., Gustafson D. R. and Hardy J. (2012) The genetic architecture of Alzheimer's disease: beyond APP, PSENs and APOE. Neurobiol Aging 33: 437-456
81. Lu H., Liu X., Deng Y. and Qing H. (2013) DNA methylation, a hand behind neurodegenerative diseases. Front Aging Neurosci 5: 85
82. Jonsson T., Atwal J. K., Steinberg S., Snaedal J., Jonsson P. V., et al. (2012) A mutation in APP protects against Alzheimer's disease and age-related cognitive decline. Nature 488: 96-99
83. De Strooper B. and Voet T. (2012) Alzheimer's disease: A protective mutation. Nature 488: 38-39
84. Mitsui J. and Tsuji S. (2014) Genomic Aspects of Sporadic Neurodegenerative Diseases. Biochem Biophys Res Commun 452: 221-225
85. Genin E., Hannequin D., Wallon D., Sleegers K., Hiltunen M., et al. (2011) APOE and Alzhei-mer disease: a major gene with semi-dominant inheritance. Mol Psychiatry 16: 903-907
86. Medland S. E., Jahanshad N., Neale B. M. and Thompson P. M. (2014) Whole-genome analyses of whole-brain data: working within an expanded search space. Nat Neurosci 17: 791-800
87. Mitsui J. and Tsuji S. (2014) Genomic Aspects of Sporadic Neurodegenerative Diseases. Biochem Biophys Res Commun 452: 221-225
88. Cruchaga C., Karch C. M., Jin S. C., Benitez B. A., Cai Y., et al. (2014) Rare coding variants in the phospholipase D3 gene confer risk for Alzheimer's disease. Nature 505: 550-554

15 An Overview of Publications Featuring Illumina® Technology
Aging and AD are associated with a spectrum of epigenetic changes, such as
abnormal DNA methylation and histone modification.89 These changes can occur
under certain environmental conditions, such as stroke,90 hypertension, type II
diabetes, obesity,91 exposure to heavy metals,92,93 head injury, immunological
proteins,94,95,96 and maternal viral infections.97-100 (see Epigenetic Modifications)
An analysis of the two largest GWAS available for AD has shown that there is a
significant overlap between disease-associated genes in pathways associated with
AD, cholesterol metabolism, and immune response.101 Neurodegeneration is often
accompanied by the accumulation of microglia and monocytes around amyloid
plaques and dying neurons.102 Furthermore, neurons express some molecules
normally attributed to the immune system, thus hinting at an intricate interplay
between neuronal and immune systems.103 (see Biology: Immunity)
ReviewsGuo J. L. and Lee V. M. (2014) Cell-to-cell transmission of pathogenic proteins in neurodegenerative diseases. Nat Med 20: 130-138
Medland S. E., Jahanshad N., Neale B. M. and Thompson P. M. (2014) Whole-genome analyses of whole-brain data: working within an expanded search space. Nat Neurosci 17: 791-800
Mitsui J. and Tsuji S. (2014) Genomic Aspects of Sporadic Neurodegenerative Diseases. Biochem Biophys Res Commun 452: 221-225
Jucker M. and Walker L. C. (2013) Self-propagation of pathogenic protein aggregates in neurodegenerative diseases. Nature 501: 45-51
Bras J., Guerreiro R. and Hardy J. (2012) Use of next-generation sequencing and other whole-genome strategies to dissect neurological disease. Nat Rev Neurosci 13: 453-464
Eisenberg D. and Jucker M. (2012) The amyloid state of proteins in human diseases. Cell 148: 1188-1203
Guerreiro R. J., Gustafson D. R. and Hardy J. (2012) The genetic architecture of Alzheimer's disease: beyond APP, PSENs and APOE. Neurobiol Aging 33: 437-456
Huang Y. and Mucke L. (2012) Alzheimer mechanisms and therapeutic strategies. Cell 148: 1204-1222
Mills J. D. and Janitz M. (2012) Alternative splicing of mRNA in the molecular pathology of neurodegenerative diseases. Neurobiol Aging 33: 1012 e1011-1024
Selkoe D. J. (2012) Preventing Alzheimer's disease. Science 337: 1488-1492
ReferencesCruchaga C., Karch C. M., Jin S. C., Benitez B. A., Cai Y., et al. (2014) Rare coding variants in the phospholipase D3 gene confer risk for Alzheimer's disease. Nature 505: 550-554Research into LOAD has identified several genetic risk variants, but generally with small effects. To identify low-frequency coding variants with large effects, this study used WES on Illumina HiSeq in 14 large LOAD families and case-control data sets. The authors found a rare variant in PLD3 segregating with disease status in two independent families and doubling the risk for AD in seven independent case-control series. The authors conducted follow-up functional assays to determine the effect of PLD3 and found that it influences APP processing.
Illumina Technology: GoldenGate, HiSeq 2000
“Twin studies support the notion that epigenetic mechanisms modulate AD risk.” Huang and Mucke 2012
89. Chouliaras L., Rutten B. P., Kenis G., Peer-booms O., Visser P. J., et al. (2010) Epigenetic regulation in the pathophysiology of Alzhei-mer's disease. Prog Neurobiol 90: 498-510
90. Savva G. M., Stephan B. C. and Alzheimer's Society Vascular Dementia Systematic Review G. (2010) Epidemiological studies of the effect of stroke on incident dementia: a systematic review. Stroke 41: e41-46
91. Huang Y. and Mucke L. (2012) Alzheimer mechanisms and therapeutic strategies. Cell 148: 1204-1222
92. Bakulski K. M., Rozek L. S., Dolinoy D. C., Paulson H. L. and Hu H. (2012) Alzheimer's disease and environmental exposure to lead: the epidemiologic evidence and potential role of epigenetics. Curr Alzheimer Res 9: 563-573
93. Basha M. R., Wei W., Bakheet S. A., Benitez N., Siddiqi H. K., et al. (2005) The fetal basis of amyloidogenesis: exposure to lead and latent overexpression of amyloid precursor protein and beta-amyloid in the aging brain. J Neurosci 25: 823-829
94. Gadad B. S., Hewitson L., Young K. A. and German D. C. (2013) Neuropathology and animal models of autism: genetic and environmental factors. Autism Res Treat 2013: 731935
95. Durkin M. S., Maenner M. J., Newschaffer C. J., Lee L. C., Cunniff C. M., et al. (2008) Advanced parental age and the risk of autism spectrum disorder. Am J Epidemiol 168: 1268-1276
96. Chung S. J., Armasu S. M., Anderson K. J., Biernacka J. M., Lesnick T. G., et al. (2013) Genetic susceptibility loci, environmental expo-sures, and Parkinson's disease: a case-con-trol study of gene-environment interactions. Parkinsonism Relat Disord 19: 595-599
97. Sun Y., Christensen J. and Olsen J. (2013) Childhood epilepsy and maternal antibodies to microbial and tissue antigens during pregnan-cy. Epilepsy Res 107: 61-74
98. Brown A. S. and Derkits E. J. (2010) Prenatal infection and schizophrenia: a review of epidemiologic and translational studies. Am J Psychiatry 167: 261-280
99. Torrey E. F. and Yolken R. H. (2003) Toxoplas-ma gondii and schizophrenia. Emerg Infect Dis 9: 1375-1380
100. Borglum A. D., Demontis D., Grove J., Pallesen J., Hollegaard M. V., et al. (2014) Genome-wide study of association and interaction with maternal cytomegalovirus infection suggests new schizophrenia loci. Mol Psychiatry 19: 325-333
101. Jones L., Harold D. and Williams J. (2010) Genetic evidence for the involvement of lipid metabolism in Alzheimer's disease. Biochim Biophys Acta 1801: 754-761
102. Lynch M. A. and Mills K. H. (2012) Immunol-ogy meets neuroscience--opportunities for immune intervention in neurodegenerative diseases. Brain Behav Immun 26: 1-10
103. Boulanger L. M. (2009) Immune proteins in brain development and synaptic plasticity. Neuron 64: 93-109

16 Neurobiology Research Review
De Jager P. L., Srivastava G., Lunnon K., Burgess J., Schalkwyk L. C., et al. (2014) Alzheimer's disease: early alterations in brain DNA methylation at ANK1, BIN1, RHBDF2 and other loci. Nat Neurosci 17: 1156-1163DNA methylation is a genetic mechanism that may affect gene expression and, as such, may be implicated in disease susceptibility. The epigenomic influence on AD onset and progression was examined in this study using Illumina HumanMethylation450 arrays and bisulfite sequencing on Illumina HiSeq 2000. The authors found several replicated, functionally validated associations between altered DNA methylation and the pre-symptomatic accumulation of AD pathology. They hypothesized that the observed DNA methylation changes may be involved in the onset of AD.
Illumina Technology: HumanMethylation450, HiSeq 2000
Lunnon K., Smith R., Hannon E., De Jager P. L., Srivastava G., et al. (2014) Methylomic profiling implicates cortical deregulation of ANK1 in Alzheimer's disease. Nat Neurosci 17: 1164-1170In this study of AD methylomic variation, the authors used Illumina HumanMethylation450k arrays to characterize the genome-wide DNA methylation state across multiple tissues. Based on the results from 122 donor samples, the authors compared the methylation state in four brain regions and whole blood where available. The authors identified evidence for cortex-specific hypermethylation at CpG sites in the ANK1 gene associated with AD neuropathology.
Illumina Technology: HumanMethylation450, Human Gene Expression BeadArray
Raj T., Ryan K. J., Replogle J. M., Chibnik L. B., Rosenkrantz L., et al. (2014) CD33: increased inclusion of exon 2 implicates the Ig V-set domain in Alzheimer's disease susceptibility. Hum Mol Genet 23: 2729-2736The identification of possible therapeutic targets for disease requires a functional identification follow-up to identified genetic disease variants. In this study, a previously identified risk allele for AD was scrutinized in detail using expression data on population cohorts stratified by their genetic risk variant identified by Illumina Infinium arrays. The authors found that the risk allele rs3865444(C) results in a higher surface density of CD33 on monocytes. The risk allele is strongly associated with greater expression of CD33 exon 2, which is likely to be the functional consequence of the risk variant.
Illumina Technology: Human OmniExpress
Bai B., Hales C. M., Chen P. C., Gozal Y., Dammer E. B., et al. (2013) U1 small nuclear ribonucleoprotein complex and RNA splicing alterations in Alzheimer's disease. Proc Natl Acad Sci U S A 110: 16562-16567Many neurodegenerative diseases are characterized by deposition of insoluble protein aggregates. The universal presence of ß-amyloid and tau proteins in AD has facilitated advancement of the amyloid cascade and tau hypotheses that have dominated AD pathogenesis research and therapeutic development. This study investigated the human brain-insoluble proteome in AD by mass spectrometry and transcriptome sequencing. The authors identified 36 proteins that accumulate in the disease and found similarities with protein aggregates in mild cognitive impairment.
Illumina Technology: HiSeq 2000 (mRNA sequencing)
Cruchaga C., Kauwe J. S., Harari O., Jin S. C., Cai Y., et al. (2013) GWAS of cerebrospinal fluid tau levels identifies risk variants for Alzheimer's disease. Neuron 78: 256-268The progress of AD can be monitored by the tau phosphorylated threonine 181 (ptau) in cerebrospinal fluid (CSF). To identify the genetic mechanism associated with elevated ptau, the authors performed the largest GWAS to date, enrolling 1,269 participants. The participants were genotyped using Illumina OmniExpress arrays and tau/ptau levels measured. The authors identified three genome-wide significant loci for CSF tau and ptau; one of these showed a strong association with AD risk in independent data sets.
Illumina Technology: Human610-Quad, HumanOmniExpress
Lambert J. C., Ibrahim-Verbaas C. A., Harold D., Naj A. C., Sims R., et al. (2013) Meta-analysis of 74,046 individuals identifies 11 new susceptibility loci for Alzheimer's disease. Nat Genet 45: 1452-1458AD is a progressive neurological disorder, primarily affecting the elderly. Previous analyses have identified eleven genomic loci associated with LOAD. To search for additional risk loci, the authors conducted a large GWAS meta-analysis using published datasets of Illumina iSelect genotype data from ~17,000 AD cases and ~37,000 controls. The analysis resulted in 19 significant associated loci, of which 11 loci have not been associated previously with LOAD.
Illumina Technology: iSelect

17 An Overview of Publications Featuring Illumina® Technology
Zhang B., Gaiteri C., Bodea L. G., Wang Z., McElwee J., et al. (2013) Integrated systems approach identifies genetic nodes and networks in late-onset Alzheimer's disease. Cell 153: 707-720Despite decades of intensive research, the causal chain of mechanisms behind LOAD remains elusive. This study characterized the molecular systems associated with LOAD by RNA-Seq on Illumina HiSeq for both brain specimens and cell line samples. The authors built a rank-ordered molecular interaction network by LOAD pathology and identified an immune- and microglia-specific module that is dominated by genes involved in pathogen phagocytosis. The authors recommend the causal network structure as a useful predictor of response to gene perturbations and a framework to test models of disease mechanisms underlying LOAD.
Illumina Technology: HiSeq 2000 (mRNA sequencing), HT-12 Expression BeadChip, HumanHap 650Y
Sherva R., Tripodis Y., Bennett D. A., Chibnik L. B., Crane P. K., et al. (2014) Genome-wide association study of the rate of cognitive decline in Alzheimer's disease. Alzheimers Dement 10: 45-52
Guffanti G., Torri F., Rasmussen J., Clark A. P., Lakatos A., et al. (2013) Increased CNV-region deletions in mild cognitive impairment (MCI) and Alzheimer's disease (AD) subjects in the ADNI sample. Genomics 102: 112-122
Jonsson T., Atwal J. K., Steinberg S., Snaedal J., Jonsson P. V., et al. (2012) A mutation in APP protects against Alzheimer's disease and age-related cognitive decline. Nature 488: 96-99

18 Neurobiology Research Review
Parkinson’s Disease
Parkinson’s disease (PD) is the second most common neurodegenerative
disorder after AD.104 Approximately 1 million people in the U.S., and over 4 million
people world-wide, develop this pathological condition. The prevalence of PD in
industrialized countries is estimated at 1%–2% in people over 60 years of age, and
3%–5% in people over 85 years old. Only 1% of PD cases are familial; the rest are
sporadic.105 Typical symptoms include muscle rigidity, bradykinesia (slow movement),
tremors, and postural instability.106 As the disease progresses, memory loss can
occur and the symptoms may become very similar to those of AD.107 Persons
with PD may develop various neuropsychiatric disorders, such as anxiety, apathy,
depression, hallucinations, and delusions.108
The diagnosis of PD is based on symptoms, and no molecular tests are used at this
time.109 A majority of neurons exhibit degenerative dysfunction or are lost before the
onset of visible symptoms of PD,110 so early detection could improve the prognosis
substantially. Recently, a few groups have reported on the development of minimally
invasive diagnostic systems for detection of AD111,112,113 and PD114 in whole peripheral
blood, plasmacytoid bone marrow–derived cells (PBMCs), or CSF. Examples of
the target molecules for diagnostics include eukaryotic initiation factor 2 (EIF2),115
epidermal growth factor (EGF),116,117 and amyloid ß1-42.118,119
104. Lu H., Liu X., Deng Y. and Qing H. (2013) DNA methylation, a hand behind neurodegenerative diseases. Front Aging Neurosci 5: 85
105. Mills J. D. and Janitz M. (2012) Alternative splicing of mRNA in the molecular pathology of neurodegenerative diseases. Neurobiol Aging 33: 1012 e1011-1024
106. de Lau L. M. and Breteler M. M. (2006) Epide-miology of Parkinson's disease. Lancet Neurol 5: 525-535
107. Mills J. D. and Janitz M. (2012) Alternative splicing of mRNA in the molecular pathology of neurodegenerative diseases. Neurobiol Aging 33: 1012 e1011-1024
108. Emre M., Aarsland D., Brown R., Burn D. J., Duyckaerts C., et al. (2007) Clinical diagnostic criteria for dementia associated with Parkin-son's disease. Mov Disord 22: 1689-1707; quiz 1837
109. Fan H. C., Chen S. J., Harn H. J. and Lin S. Z. (2013) Parkinson's disease: from genetics to treatments. Cell Transplant 22: 639-652
110. Fan H. C., Chen S. J., Harn H. J. and Lin S. Z. (2013) Parkinson's disease: from genetics to treatments. Cell Transplant 22: 639-652
111. Booij B. B., Lindahl T., Wetterberg P., Skaane N. V., Saebo S., et al. (2011) A gene expres-sion pattern in blood for the early detection of Alzheimer's disease. J Alzheimers Dis 23: 109-119
112. Rye P. D., Booij B. B., Grave G., Lindahl T., Kristiansen L., et al. (2011) A novel blood test for the early detection of Alzheimer's disease. J Alzheimers Dis 23: 121-129
113. Fehlbaum-Beurdeley P., Sol O., Desire L., Touchon J., Dantoine T., et al. (2012) Validation of AclarusDx, a blood-based transcriptomic signature for the diagnosis of Alzheimer's disease. J Alzheimers Dis 32: 169-181
114. Karlsson M. K., Sharma P., Aasly J., Toft M., Skogar O., et al. (2013) Found in transcription: accurate Parkinson's disease classification in peripheral blood. J Parkinsons Dis 3: 19-29
115. Mutez E., Nkiliza A., Belarbi K., de Broucker A., Vanbesien-Mailliot C., et al. (2014) Involve-ment of the immune system, endocytosis and EIF2 signaling in both genetically determined and sporadic forms of Parkinson's disease. Neurobiol Dis 63: 165-170
116. Chen-Plotkin A. S., Hu W. T., Siderowf A., Weintraub D., Goldmann Gross R., et al. (2011) Plasma epidermal growth factor levels predict cognitive decline in Parkinson disease. Ann Neurol 69: 655-663
117. Irwin D. J., Lee V. M. and Trojanowski J. Q. (2013) Parkinson's disease dementia: conver-gence of alpha-synuclein, tau and amyloid-be-ta pathologies. Nat Rev Neurosci 14: 626-636
118. Siderowf A., Xie S. X., Hurtig H., Weintraub D., Duda J., et al. (2010) CSF amyloid {beta} 1-42 predicts cognitive decline in Parkinson disease. Neurology 75: 1055-1061
119. Irwin D. J., Lee V. M. and Trojanowski J. Q. (2013) Parkinson's disease dementia: conver-gence of alpha-synuclein, tau and amyloid-be-ta pathologies. Nat Rev Neurosci 14: 626-636
19 An Overview of Publications Featuring Illumina® Technology
The biological causes of PD include progressive loss of substantia nigra
dopaminergic neurons and striatal projections.120 The major pathological indicator of
PD is the accumulation of Lewy bodies, which are predominantly formed by self-
assembling small protein -synuclein expressed in multiple brain segments.121 Similar
to plaques in AD, Lewy bodies spread to other compartments of brain (e.g., limbic
and neocortical areas) as the disease progresses and cause neuronal death.122
Age is a major risk factor for development of PD123. Over 80% of PD patients will
eventually develop dementia, a condition termed Parkinson’s disease dementia
(PDD). It is believed that the major cause of dementia of this kind is the spread
of fibrillar α-synuclein from the brain stem to limbic and neocortical structures.124
Additionally, over 50% of PDD patients develop amyloid- ß plaques and neurofibrillary
tangles, typical for AD.125 Dual pathology of PDD and AD increases malignancy of the
disease and significantly worsens prognosis.
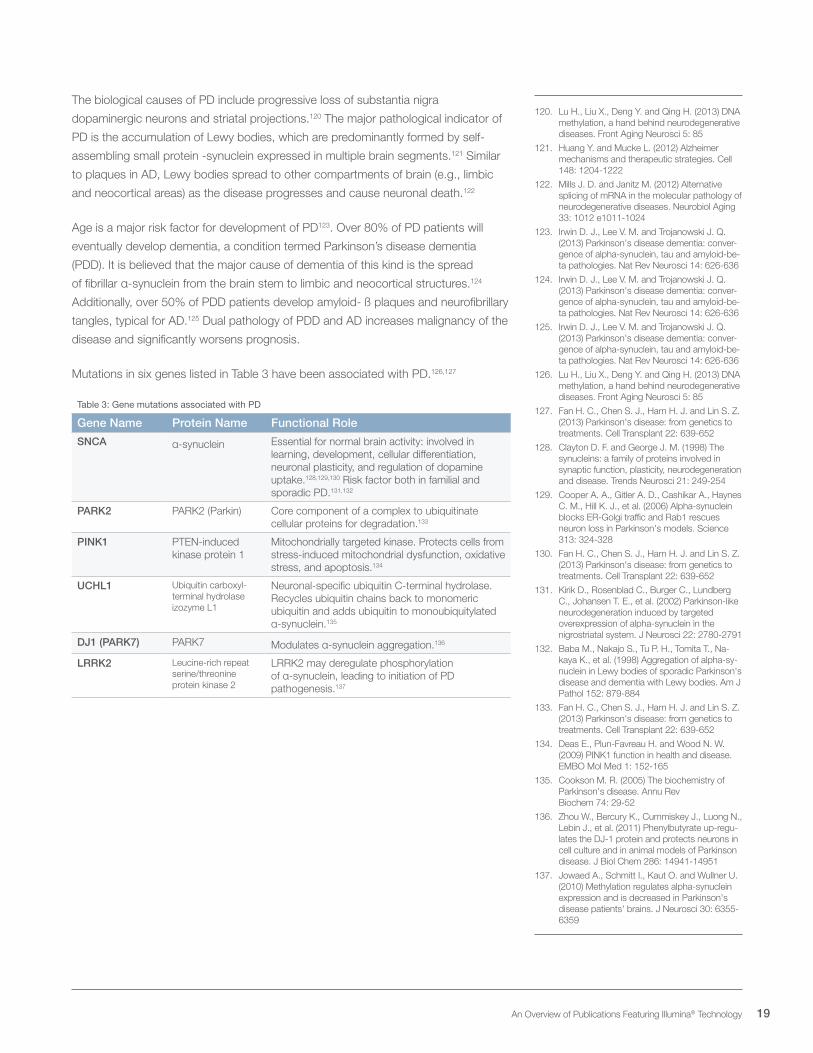
Mutations in six genes listed in Table 3 have been associated with PD.126,127
Table 3: Gene mutations associated with PD
Gene Name Protein Name Functional Role
SNCA α-synuclein Essential for normal brain activity: involved in learning, development, cellular differentiation, neuronal plasticity, and regulation of dopamine uptake.128,129,130 Risk factor both in familial and sporadic PD.131,132
PARK2 PARK2 (Parkin) Core component of a complex to ubiquitinate cellular proteins for degradation.133
PINK1 PTEN-induced kinase protein 1
Mitochondrially targeted kinase. Protects cells from stress-induced mitochondrial dysfunction, oxidative stress, and apoptosis.134
UCHL1 Ubiquitin carboxyl-terminal hydrolase izozyme L1
Neuronal-specific ubiquitin C-terminal hydrolase. Recycles ubiquitin chains back to monomeric ubiquitin and adds ubiquitin to monoubiquitylated α-synuclein.135
DJ1 (PARK7) PARK7 Modulates α-synuclein aggregation.136
LRRK2 Leucine-rich repeat serine/threonine protein kinase 2
LRRK2 may deregulate phosphorylation of α-synuclein, leading to initiation of PD pathogenesis.137
120. Lu H., Liu X., Deng Y. and Qing H. (2013) DNA methylation, a hand behind neurodegenerative diseases. Front Aging Neurosci 5: 85
121. Huang Y. and Mucke L. (2012) Alzheimer mechanisms and therapeutic strategies. Cell 148: 1204-1222
122. Mills J. D. and Janitz M. (2012) Alternative splicing of mRNA in the molecular pathology of neurodegenerative diseases. Neurobiol Aging 33: 1012 e1011-1024
123. Irwin D. J., Lee V. M. and Trojanowski J. Q. (2013) Parkinson's disease dementia: conver-gence of alpha-synuclein, tau and amyloid-be-ta pathologies. Nat Rev Neurosci 14: 626-636
124. Irwin D. J., Lee V. M. and Trojanowski J. Q. (2013) Parkinson's disease dementia: conver-gence of alpha-synuclein, tau and amyloid-be-ta pathologies. Nat Rev Neurosci 14: 626-636
125. Irwin D. J., Lee V. M. and Trojanowski J. Q. (2013) Parkinson's disease dementia: conver-gence of alpha-synuclein, tau and amyloid-be-ta pathologies. Nat Rev Neurosci 14: 626-636
126. Lu H., Liu X., Deng Y. and Qing H. (2013) DNA methylation, a hand behind neurodegenerative diseases. Front Aging Neurosci 5: 85
127. Fan H. C., Chen S. J., Harn H. J. and Lin S. Z. (2013) Parkinson's disease: from genetics to treatments. Cell Transplant 22: 639-652
128. Clayton D. F. and George J. M. (1998) The synucleins: a family of proteins involved in synaptic function, plasticity, neurodegeneration and disease. Trends Neurosci 21: 249-254
129. Cooper A. A., Gitler A. D., Cashikar A., Haynes C. M., Hill K. J., et al. (2006) Alpha-synuclein blocks ER-Golgi traffic and Rab1 rescues neuron loss in Parkinson's models. Science 313: 324-328
130. Fan H. C., Chen S. J., Harn H. J. and Lin S. Z. (2013) Parkinson's disease: from genetics to treatments. Cell Transplant 22: 639-652
131. Kirik D., Rosenblad C., Burger C., Lundberg C., Johansen T. E., et al. (2002) Parkinson-like neurodegeneration induced by targeted overexpression of alpha-synuclein in the nigrostriatal system. J Neurosci 22: 2780-2791
132. Baba M., Nakajo S., Tu P. H., Tomita T., Na-kaya K., et al. (1998) Aggregation of alpha-sy-nuclein in Lewy bodies of sporadic Parkinson's disease and dementia with Lewy bodies. Am J Pathol 152: 879-884
133. Fan H. C., Chen S. J., Harn H. J. and Lin S. Z. (2013) Parkinson's disease: from genetics to treatments. Cell Transplant 22: 639-652
134. Deas E., Plun-Favreau H. and Wood N. W. (2009) PINK1 function in health and disease. EMBO Mol Med 1: 152-165
135. Cookson M. R. (2005) The biochemistry of Parkinson's disease. Annu Rev Biochem 74: 29-52
136. Zhou W., Bercury K., Cummiskey J., Luong N., Lebin J., et al. (2011) Phenylbutyrate up-regu-lates the DJ-1 protein and protects neurons in cell culture and in animal models of Parkinson disease. J Biol Chem 286: 14941-14951
137. Jowaed A., Schmitt I., Kaut O. and Wullner U. (2010) Methylation regulates alpha-synuclein expression and is decreased in Parkinson's disease patients' brains. J Neurosci 30: 6355-6359

20 Neurobiology Research Review
The familial type of PD can be inherited in an autosomal dominant or autosomal
recessive manner. The former is associated with α-synuclein and LRRK2 and the
latter with Parkin, PINK1, DJ1, and ATP13A2. Improved understanding of the role
of these genes in PD will facilitate the development of genomic analysis as an early
diagnostic tool for familial PD.138
Currently, the major approach to controlling clinical symptoms of patients with
PD is pharmacological replacement of dopamine with L-DOPA, carbidopa, and
monoamine oxidase-B inhibitors.139 However, the effect of such treatment is transient,
and patients develop resistance to these therapies, leaving no further options for
treatment. Genomic analysis of PD is anticipated to improve treatment of PD. One
of the new therapeutic approaches made possible by NGS is based on the use
of mirtrons: miRNA relying on splicing, rather on Dicer and RNA-induced silencing
complex (RISC) to generate precursors for targeting disease-specific mRNA via an
RNA interference pathway.140 With this approach, researchers attained up to 85%
silencing of α-synuclein and LRRK2.
Methylation of α-synuclein was reduced in DNA from substantia nigra, cortex, and
putamen of patients with sporadic PD.141 Six risk loci have been associated with
proximal gene expression or DNA methylation142 (see Epigenetic Modifications).
ReviewsGuo J. L. and Lee V. M. (2014) Cell-to-cell transmission of pathogenic proteins in neurodegenerative diseases. Nat Med 20: 130-138
Sekiyama K., Takamatsu Y., Waragai M. and Hashimoto M. (2014) Role of genomics in translational research for Parkinson's disease. Biochem Biophys Res Commun 452: 226-235
Sharma M., Kruger R. and Gasser T. (2014) From genome-wide association studies to next-generation sequencing: lessons from the past and planning for the future. JAMA Neurol 71: 5-6
Fan H. C., Chen S. J., Harn H. J. and Lin S. Z. (2013) Parkinson's disease: from genetics to treatments. Cell Transplant 22: 639-652
Irwin D. J., Lee V. M. and Trojanowski J. Q. (2013) Parkinson's disease dementia: convergence of alpha-synuclein, tau and amyloid-beta pathologies. Nat Rev Neurosci 14: 626-636
Koboldt D. C., Steinberg K. M., Larson D. E., Wilson R. K. and Mardis E. R. (2013) The next-generation sequencing revolution and its impact on genomics. Cell 155: 27-38
Sandoe J. and Eggan K. (2013) Opportunities and challenges of pluripotent stem cell neurodegenerative disease models. Nat Neurosci 16: 780-789
Trinh J. and Farrer M. (2013) Advances in the genetics of Parkinson disease. Nat Rev Neurol 9: 445-454
Bras J., Guerreiro R. and Hardy J. (2012) Use of next-generation sequencing and other whole-genome strategies to dissect neurological disease. Nat Rev Neurosci 13: 453-464
138. Fan H. C., Chen S. J., Harn H. J. and Lin S. Z. (2013) Parkinson’s disease: from genetics to treatments. Cell Transplant 22: 639-652
139. Fan H. C., Chen S. J., Harn H. J. and Lin S. Z. (2013) Parkinson’s disease: from genetics to treatments. Cell Transplant 22: 639-652
140. Sibley C. R., Seow Y., Curtis H., Weinberg M. S. and Wood M. J. (2012) Silencing of Parkin-son's disease-associated genes with artificial mirtron mimics of miR-1224. Nucleic Acids Res 40: 9863-9875
141. Jowaed A., Schmitt I., Kaut O. and Wullner U. (2010) Methylation regulates alpha-synuclein expression and is decreased in Parkinson's disease patients' brains. J Neurosci 30: 6355-6359
142. Nalls M. A., Pankratz N., Lill C. M., Do C. B., Hernandez D. G., et al. (2014) Large-scale meta-analysis of genome-wide association data identifies six new risk loci for Parkinson's disease. Nat Genet 46: 989-993

21 An Overview of Publications Featuring Illumina® Technology
ReferencesBras J., Guerreiro R., Darwent L., Parkkinen L., Ansorge O., et al. (2014) Genetic analysis implicates APOE, SNCA and suggests lysosomal dysfunction in the etiology of dementia with Lewy bodies. Hum Mol Genet Clinical and neuropathological similarities between dementia with Lewy bodies (DLB), PD, and AD, suggest that these disorders may share the same etiology. To test this hypothesis, this study used Illumina NeuroX custom arrays to test for association of 54 genomic regions, previously implicated in PD or AD in a large cohort of DLB cases and controls. The authors identified APOE as a strong genetic risk factor for DLB, and several other potential risk loci. Overall, the results suggest the etiology of DLB is influenced by some of the same genetic risk factors as AD and PD, but that these loci may act in different manners.
Illumina Technology: Infinium HumanExome BeadChip
Hollerhage M., Goebel J. N., de Andrade A., Hildebrandt T., Dolga A., et al. (2014) Trifluoperazine rescues human dopaminergic cells from wild-type alpha-synuclein-induced toxicity. Neurobiol Aging 35: 1700-1711PD is the most frequent neurodegenerative movement disorder. To study α-synuclein-mediated toxicity in PD progression, the authors developed a new cell-line model in which moderate overexpression of wild-type α-synuclein led to gradual death of human post-mitotic DA neurons. Using Illumina BeadArrays to monitor gene expression, the authors discovered that activating autophagy in human DA mid-brain neurons rescued them from α-synuclein-induced cell death. The phenothiazine neuroleptic trifluoperazine, an activator of macroautophagy, may be a potential therapeutic target.
Illumina Technology: Human Gene Expression BeadArray
Mencacci N. E., Isaias I. U., Reich M. M., Ganos C., Plagnol V., et al. (2014) Parkinson's disease in GTP cyclohydrolase 1 mutation carriers. Brain 137: 2480-2492GTP cyclohydrolase 1 (encoded by the GCH1 gene) is potentially associated with increased risk of PD. The frequency of GCH1 variants was evaluated in WES data of 1,318 cases with PD and 5,935 control subjects. Through their analysis, the authors identified 11 different heterozygous variants in GCH1. These include four previously reported pathogenic variants and seven variants of unknown clinical relevance. The frequency of GCH1 variants was significantly higher than in controls, indicating the clinical relevance of GTP cyclohydrolase 1 deficiency.
Illumina Technology: HiSeq 2000
Nalls M. A., Pankratz N., Lill C. M., Do C. B., Hernandez D. G., et al. (2014) Large-scale meta-analysis of genome-wide association data identifies six new risk loci for Parkinson's disease. Nat Genet 46: 989-993Only a small fraction of genetic heritability has been discovered for PD to date. This study performed a meta-analysis of GWAS for PD in the search for new loci associated with the disease. Using Illumina genotyping arrays, the authors identified 24 loci that were both statistically significant and replicated across experiments. Six of the identified loci had not been previously reported associated with PD, and the authors estimated the cumulative risk of the loci to be substantial (odds ratio = 3.31).
Illumina Technology: ExomeChip, HumanOmniExpress, HumanHap550, Human610-Quad, Human660W-Quad; HumanMethylation27, Human Gene Expression BeadArray
Chung S. J., Armasu S. M., Anderson K. J., Biernacka J. M., Lesnick T. G., et al. (2013) Genetic susceptibility loci, environmental exposures, and Parkinson's disease: a case-control study of gene-environment interactions. Parkinsonism Relat Disord 19: 595-599PD susceptibility is suspected to be a combination of genetic and environmental factors. This study examined the potential interactions between known genetic risk factors and environmental exposures (pesticide application, tobacco smoking, coffee drinking, and alcohol drinking). Using Illumina GoldenGate Genotyping arrays, the authors genotyped 1,098 PD cases and 1,098 controls, and performed an interaction analysis. They found limited evidence for pairwise interactions between the examined genotypes and environmental risk factors; however, larger sample sizes will be needed to confirm any effect on PD susceptibility.
Illumina Technology: BeadArray Reader, DNA Test Panel

22 Neurobiology Research Review
Karlsson M. K., Sharma P., Aasly J., Toft M., Skogar O., et al. (2013) Found in transcription: accurate Parkinson's disease classification in peripheral blood. J Parkinsons Dis 3: 19-29The prevalence of PD increases with age. In an effort to develop a blood-based test for early detection of PD, the authors performed a large-scale gene expression study using Illumina Gene Expression BeadArrays on a cohort of PD patients and controls to develop a prediction model and test its accuracy. Through cross-validation results, they showed that PD can be correctly classified from healthy controls with an accuracy of 88% compared to clinical diagnosis. These results suggest the potential for developing a blood-based gene expression test for PD.
Illumina Technology: Human Gene Expression BeadArray
Ryan S. D., Dolatabadi N., Chan S. F., Zhang X., Akhtar M. W., et al. (2013) Isogenic human iPSC Parkinson's model shows nitrosative stress-induced dysfunction in MEF2-PGC1alpha transcription. Cell 155: 1351-1364An association has been reported between PD and exposure to mitochondrial toxins. In this study, a stem cell model was used to characterize the response to toxins using Illumina BeadArray for gene expression analysis. The authors identified a pathway whereby basal and toxin-induced nitrosative/oxidative stress results in S-nitrosylation of transcription factor MEF2C. They reported the alteration contributing to mitochondrial dysfunction and apoptotic cell death, indicating a mechanism and potential therapeutic target for PD.
Illumina Technology: Human Gene Expression BeadArray
Ganat Y. M., Calder E. L., Kriks S., Nelander J., Tu E. Y., et al. (2012) Identification of embryonic stem cell-derived midbrain dopaminergic neurons for engraftment. J Clin Invest 122: 2928-2939
Rousseaux M. W., Marcogliese P. C., Qu D., Hewitt S. J., Seang S., et al. (2012) Progressive dopaminergic cell loss with unilateral-to-bilateral progression in a genetic model of Parkinson disease. Proc Natl Acad Sci U S A 109: 15918-15923
Sibley C. R., Seow Y., Curtis H., Weinberg M. S. and Wood M. J. (2012) Silencing of Parkinson's disease-associated genes with artificial mirtron mimics of miR-1224. Nucleic Acids Res 40: 9863-9875

23 An Overview of Publications Featuring Illumina® Technology
GENETIC MECHANISMS
Schizophrenia, autism, PD, and AD are complex diseases driven by an intricate
interplay between multiple genetic and environmental factors. The etiology of a
complex disease is the sum of all these factors, including somatic (non-inherited)
mutations, inherited mutations, epigenetic modifications, small RNAs, immunity, and
many others. NGS provides the tools to measure most of these contributing factors.
The future challenge will be to combine these measurements into a coherent view of
these complex diseases.143
Copy-Number Variants
CNVs are one of the most common mutations in psychiatric diseases. Some
notable successes have been achieved with array-based approaches, particularly
with mapping CNVs.144 However, arrays cannot detect balanced translocations
and fluorescence in situ hybridization (FISH) techniques have limited resolution. The
true extent of balanced translocations in both healthy and diseased genomes was
only discovered with the advent of NGS. Paired-end and mate-pair sequencing
are particularly effective in mapping genomic rearrangements.145 Two groups
have estimated that the number of CNVs relevant to ASD range from 130 to 300
target loci.146,147 CNVs also play a critical role in onset of schizophrenia and bipolar
disorder.148-151 A recent study of single cells and neurons in brain tissue found
that most (≥95%) neurons in normal brain tissue are euploid. However, a patient
with hemimegalencephaly (HMG) due to a somatic CNV of chromosome 1q had
unexpected tetrasomy 1q in 20% of neurons. This observation suggests that different
cells in the brain may have different mutations, and that CNVs in a minority of cells
can cause widespread brain dysfunction.152 This increased complexity can only be
resolved with single-cell sequencing approaches (see Biology: Single Cells).
The majority of de novo structural variations are attributed to new transposon
insertions. Exome sequencing studies have shown that an increased level of ASD-
causing single-nucleotide variants (SNVs) correlates positively with paternal age.153
CNVs can also contribute to disease, in combination with other CNVs or other
point mutations at different loci, but identifying these interactions is challenging.154
To account for these multiple impacts, several groups have devised a “two-hit”
hypothesis, analogous to that for cancer.155,156
“Recent advances in technology have allowed the interrogation of very large numbers of markers dispersed throughout the genome in a highly rapid and inexpensive manner.” Bras et al. 2012
143. Wright J. (2014) Genetics: Unravelling complexity. Nature 508: S6-7
144. Buxbaum J. (2013) The Neuroscience of Autism Spectrum Disorders.
145. Quinlan A. R. and Hall I. M. (2012) Characterizing complex structural variation in germline and somatic genomes. Trends Genet 28: 43-53
146. Levy D., Ronemus M., Yamrom B., Lee Y. H., Leotta A., et al. (2011) Rare de novo and transmitted copy-number variation in autistic spectrum disorders. Neuron 70: 886-897
147. Sanders S. J., Ercan-Sencicek A. G., Hus V., Luo R., Murtha M. T., et al. (2011) Multiple recurrent de novo CNVs, including duplications of the 7q11.23 Williams syndrome region, are strongly associated with autism. Neuron 70: 863-885
148. Malhotra D., McCarthy S., Michaelson J. J., Vacic V., Burdick K. E., et al. (2011) High frequen-cies of de novo CNVs in bipolar disorder and schizophrenia. Neuron 72: 951-963
149. Priebe L., Degenhardt F. A., Herms S., Haenisch B., Mattheisen M., et al. (2012) Genome-wide survey implicates the influence of copy number variants (CNVs) in the development of early-onset bipolar disorder. Mol Psychiatry 17: 421-432
150. Zhang D., Cheng L., Qian Y., Alliey-Rodriguez N., Kelsoe J. R., et al. (2009) Singleton deletions throughout the genome increase risk of bipolar disorder. Mol Psychiatry 14: 376-380
151. Stefansson H., Meyer-Lindenberg A., Steinberg S., Magnusdottir B., Morgen K., et al. (2014) CNVs conferring risk of autism or schizophrenia affect cognition in controls. Nature 505: 361-366
152. Cai X., Evrony G. D., Lehmann H. S., Elhosary P. C., Mehta B. K., et al. (2014) Single-Cell, Genome-wide Sequencing Identifies Clonal Somatic Copy-Number Variation in the Human Brain. Cell Rep 8: 1280-1289
153. Abnet C. C., Wang Z., Song X., Hu N., Zhou F. Y., et al. (2012) Genotypic variants at 2q33 and risk of esophageal squamous cell carcinoma in China: a meta-analysis of genome-wide associa-tion studies. Hum Mol Genet 21: 2132-2141
154. Nava C., Keren B., Mignot C., Rastetter A., Chantot-Bastaraud S., et al. (2014) Prospective diagnostic analysis of copy number variants using SNP microarrays in individuals with autism spectrum disorders. Eur J Hum Genet 22: 71-78
155. Nava C., Keren B., Mignot C., Rastetter A., Chantot-Bastaraud S., et al. (2014) Prospective diagnostic analysis of copy number variants using SNP microarrays in individuals with autism spectrum disorders. Eur J Hum Genet 22: 71-78
156. Gau S. S., Liao H. M., Hong C. C., Chien W. H. and Chen C. H. (2012) Identification of two inher-ited copy number variants in a male with autism supports two-hit and compound heterozygosity models of autism. Am J Med Genet B Neuropsy-chiatr Genet 159B: 710-717

24 Neurobiology Research Review
Scandinavian families have been used extensively as subjects for autism and schizophrenia GWAS. This selection is due to the ethnic uniformity of Scandinavian peoples, and the well-developed registry system of newborn blood samples (such as Danish Newborn Screening Biobank) and health records established in those countries.157
ReviewsCai X., Evrony G. D., Lehmann H. S., Elhosary P. C., Mehta B. K., et al. (2014) Single-Cell, Genome-wide Sequencing Identifies Clonal Somatic Copy-Number Variation in the Human Brain. Cell Rep 8: 1280-1289
Gilissen C., Hehir-Kwa J. Y., Thung D. T., van de Vorst M., van Bon B. W., et al. (2014) Genome sequencing identifies major causes of severe intellectual disability. Nature 511: 344-347
Jacquemont S., Coe B. P., Hersch M., Duyzend M. H., Krumm N., et al. (2014) A higher mutational burden in females supports a "female protective model" in neurodevelopmental disorders. Am J Hum Genet 94: 415-425
Rosti R. O., Sadek A. A., Vaux K. K. and Gleeson J. G. (2014) The genetic landscape of autism spectrum disorders. Dev Med Child Neurol 56: 12-18
Malhotra D. and Sebat J. (2012) CNVs: harbingers of a rare variant revolution in psychiatric genetics. Cell 148: 1223-1241
ReferencesFromer M., Pocklington A. J., Kavanagh D. H., Williams H. J., Dwyer S., et al. (2014) De novo mutations in schizophrenia implicate synaptic networks. Nature 506: 179-184Of the known risk alleles for schizophrenia, the only ones definitively shown to confer considerable increments in risk are rare chromosomal CNVs that involve deletion or duplication of thousands of bases of DNA. This study examined the effect of small de novo mutations affecting one or a few nucleotides. By Illumina HiSeq WES of 623 schizophrenia trios, the authors assessed de novo mutation rates and shared genetic etiology for schizophrenia, intellectual disability, and ASDs. They found several insights to suggest a common etiological mechanism.
Illumina Technology: HiSeq (exome sequencing)
157. Szatkiewicz J. P., O'Dushlaine C., Chen G., Chambert K., Moran J. L., et al. (2014) Copy number variation in schizophrenia in Sweden. Mol Psychiatry 19: 762-773

25 An Overview of Publications Featuring Illumina® Technology
Gaugler T., Klei L., Sanders S. J., Bodea C. A., Goldberg A. P., et al. (2014) Most genetic risk for autism resides with common variation. Nat Genet 46: 881-885Although ASDs have been studied widely, the proportion and nature of genetic heritability is uncertain. This study analyzed the largest autism cohort data to date, including 1.6 million Swedish families with at least two children and ~5,700 individuals with strict autism diagnoses. Using Illumina SNP arrays, the authors investigated the contribution of rare versus common genetic variants to the disease. They conclude that the heritability is ~52.4%, with common variation as the biggest contributor. Rare, de novo mutations contribute substantially to individual liability, yet their contribution to variance in liability is modest at 2.6%.
Illumina Technology: OmniExpress
Karayannis T., Au E., Patel J. C., Kruglikov I., Markx S., et al. (2014) Cntnap4 differentially contributes to GABAergic and dopaminergic synaptic transmission. Nature 511: 236-240In an effort to understand the pathological development of neurological disorders, genetic effects have been studied in the context of proteins that are expressed in neural cells. In this study, the authors characterized the effect of CNTNAP4 knockouts on mouse behavior and development, and relate these results to the findings of CNVs in humans across a region including the CNTNAP2 gene. The authors found that CNTNAP4 is localized presynaptically, and its loss leads to a reduction in the output of cortical parvalbumin (PV)-positive GABAergic basket cells. In addition, CNTNAP4-mutant mice showed defects in these neuronal populations and exhibited sensory-motor gating and grooming endophenotypes.
Illumina Technology: HumanHap550, HumanOmni1-Quad
Stefansson H., Meyer-Lindenberg A., Steinberg S., Magnusdottir B., Morgen K., et al. (2014) CNVs conferring risk of autism or schizophrenia affect cognition in controls. Nature 505: 361-366Certain CNVs contribute to the pathogenesis of schizophrenia and autism. In this study, the authors investigated the influence of these CNVs on phenotypes separate from those of the mentioned diseases. In a big population-wide study of nearly a third of the Icelandic population (n = 101,655), the authors used Illumina SNP microarrays to test for associations of CNVs with cognitive deficits, dyslexia, dyscalculia, and brain structure changes. The authors found that the 15q11.2(BP1-BP2) deletion affects brain structure in a pattern consistent with first-episode psychosis in both schizophrenia and dyslexia.
Illumina Technology: HumanHap300, HumanCNV370-Duo, HumanHap650Y, Human1M, HumanOmni2.5, HumanOmniExpress, HumanOmni1S
Chapman J., Rees E., Harold D., Ivanov D., Gerrish A., et al. (2013) A genome-wide study shows a limited contribution of rare copy number variants to Alzheimer's disease risk. Hum Mol Genet 22: 816-824AD is the most common form of dementia, and although genetically complex, it is also highly heritable. This study examined the prevalence of CNVs using Illumina Infinium arrays to determine whether rare CNVs play a role in AD susceptibility. Although the authors examined loci that had been highlighted in previous AD studies, they did not find a significant contribution of CNVs to the development of AD.
Illumina Technology: Human610-Quad
Lionel A. C., Tammimies K., Vaags A. K., Rosenfeld J. A., Ahn J. W., et al. (2014) Disruption of the ASTN2/TRIM32 locus at 9q33.1 is a risk factor in males for autism spectrum disorders, ADHD and other neurodevelopmental phenotypes. Hum Mol Genet 23: 2752-2768
McGrath L. M., Yu D., Marshall C., Davis L. K., Thiruvahindrapuram B., et al. (2014) Copy number variation in obsessive-compulsive disorder and tourette syndrome: a cross-disorder study. J Am Acad Child Adolesc Psychiatry 53: 910-919
Morris D. W., Pearson R. D., Cormican P., Kenny E. M., O'Dushlaine C. T., et al. (2014) An inherited duplication at the gene p21 Protein-Activated Kinase 7 (PAK7) is a risk factor for psychosis. Hum Mol Genet 23: 3316-3326
Nava C., Keren B., Mignot C., Rastetter A., Chantot-Bastaraud S., et al. (2014) Prospective diagnostic analysis of copy number variants using SNP microarrays in individuals with autism spectrum disorders. Eur J Hum Genet 22: 71-78
Ramos-Quiroga J. A., Sanchez-Mora C., Casas M., Garcia-Martinez I., Bosch R., et al. (2014) Genome-wide copy number variation analysis in adult attention-deficit and hyperactivity disorder. J Psychiatr Res 49: 60-67
Rees E., Kirov G., Sanders A., Walters J. T., Chambert K. D., et al. (2014) Evidence that duplications of 22q11.2 protect against schizophrenia. Mol Psychiatry 19: 37-40

26 Neurobiology Research Review
Alternative Splicing
It has been established that up to 94% of multi-exon genes are alternatively spliced,
and that incorrect alternative splicing can lead to at least 15% of disease cases in
humans.158,159 Alternative splicing is a mechanism by which exons of pre-mRNA
can be grouped (spliced) into different arrangements to produce mature mRNA that
codes structurally and functionally distinct protein variants. The advent of high-
throughput genomic analysis tools, such as exon arrays and RNA-Seq, has allowed
the identification of alternative splicing events which were undetectable previously by
conventional microarrays.
Exon arrays can distinguish between different isoforms.160 This technology also has
some intrinsic limitations, such as the ability to detect only known splice variants of
previously sequenced genomes, low signal-to-noise ratio, limited dynamic range,
and cross-hybridization. The full power of this technology can be realized when
combined with whole mRNA sequencing, which allows the identification of the exon
and transcript boundaries at single-base resolution and detection of novel transcripts.
In this approach, the mRNA is first converted into cDNA, which is further ligated to
unique adapters and sequenced in a massively parallel fashion.161
Most genes commonly associated with AD have multiple splice variants, and some
of those variants are pathogenic.162,163,164 In PD, alternative splicing was detected
for PARK2, SNCA, and SRRM2 genes. Although all three splice variants of PARK2
are believed to be non-pathogenic, it was hypothesized that the variable ratio of
these three transcripts may determine disease susceptibility.165 Finally, there is
some evidence that unspliced mRNA corresponding to AD susceptibility genes can
accumulate in brains of AD patients as a result of mutations in U1 small nuclear
ribonucleoprotein (U1 snRNP), a component of the spliceosome complex. Multiple
U1 snRNP subunits form cytoplasmic tangled agglomerations in AD.166
RNA samples from human brains are usually acquired post-mortem, which raises
the problem of poor RNA quality for genetic analysis. However, correlative studies of
RNA and protein expression in brain and in peripheral organs and tissues, such as
blood, may provide the means for early, non-invasive diagnosis of neurodegenerative
diseases, similar to the approach once established for the diagnosis of
prostate cancer.167-170
158. Mills J. D. and Janitz M. (2012) Alternative splicing of mRNA in the molecular pathology of neurodegenerative diseases. Neurobiol Aging 33: 1012 e1011-1024
159. Martin C. L., Duvall J. A., Ilkin Y., Simon J. S., Arreaza M. G., et al. (2007) Cytogenetic and molecular characterization of A2BP1/FOX1 as a candidate gene for autism. Am J Med Genet B Neuropsychiatr Genet 144B: 869-876
160. Mills J. D. and Janitz M. (2012) Alternative splicing of mRNA in the molecular pathology of neurodegenerative diseases. Neurobiol Aging 33: 1012 e1011-1024
161. Mills J. D. and Janitz M. (2012) Alternative splicing of mRNA in the molecular pathology of neurodegenerative diseases. Neurobiol Aging 33: 1012 e1011-1024
162. Mills J. D. and Janitz M. (2012) Alternative splicing of mRNA in the molecular pathology of neurodegenerative diseases. Neurobiol Aging 33: 1012 e1011-1024
163. Raj T., Ryan K. J., Replogle J. M., Chibnik L. B., Rosenkrantz L., et al. (2014) CD33: increased inclusion of exon 2 implicates the Ig V-set domain in Alzheimer's disease suscepti-bility. Hum Mol Genet 23: 2729-2736
164. Bai B., Hales C. M., Chen P. C., Gozal Y., Dammer E. B., et al. (2013) U1 small nuclear ribonucleoprotein complex and RNA splicing alterations in Alzheimer's disease. Proc Natl Acad Sci U S A 110: 16562-16567
165. Mills J. D. and Janitz M. (2012) Alternative splicing of mRNA in the molecular pathology of neurodegenerative diseases. Neurobiol Aging 33: 1012 e1011-1024
166. Bai B., Hales C. M., Chen P. C., Gozal Y., Dammer E. B., et al. (2013) U1 small nuclear ribonucleoprotein complex and RNA splicing alterations in Alzheimer's disease. Proc Natl Acad Sci U S A 110: 16562-16567
167. Carter H. B., Morrell C. H., Pearson J. D., Brant L. J., Plato C. C., et al. (1992) Estimation of prostatic growth using serial prostate-spe-cific antigen measurements in men with and without prostate disease. Cancer Res 52: 3323-3328
168. Carter H. B., Pearson J. D., Metter E. J., Brant L. J., Chan D. W., et al. (1992) Longitudinal evaluation of prostate-specific antigen levels in men with and without prostate disease. JAMA 267: 2215-2220
169. Mills J. D. and Janitz M. (2012) Alternative splicing of mRNA in the molecular pathology of neurodegenerative diseases. Neurobiol Aging 33: 1012 e1011-1024
170. Mutez E., Nkiliza A., Belarbi K., de Broucker A., Vanbesien-Mailliot C., et al. (2014) Involve-ment of the immune system, endocytosis and EIF2 signaling in both genetically determined and sporadic forms of Parkinson's disease. Neurobiol Dis 63: 165-170

27 An Overview of Publications Featuring Illumina® Technology
ReferencesAkula N., Barb J., Jiang X., Wendland J. R., Choi K. H., et al. (2014) RNA-sequencing of the brain transcriptome implicates dysregulation of neuroplasticity, circadian rhythms and GTPase binding in bipolar disorder. Mol Psychiatry. In press.
Fogel B. L., Cho E., Wahnich A., Gao F., Becherel O. J., et al. (2014) Mutation of senataxin alters disease-specific transcriptional networks in patients with ataxia with oculomotor apraxia type 2. Hum Mol Genet 23: 4758-4769
Gomez-Herreros F., Schuurs-Hoeijmakers J. H., McCormack M., Greally M. T., Rulten S., et al. (2014) TDP2 protects transcription from abortive topoisomerase activity and is required for normal neural function. Nat Genet 46: 516-521
Lionel A. C., Tammimies K., Vaags A. K., Rosenfeld J. A., Ahn J. W., et al. (2014) Disruption of the ASTN2/TRIM32 locus at 9q33.1 is a risk factor in males for autism spectrum disorders, ADHD and other neurodevelopmental phenotypes. Hum Mol Genet 23: 2752-2768
Morihara T., Hayashi N., Yokokoji M., Akatsu H., Silverman M. A., et al. (2014) Transcriptome analysis of distinct mouse strains reveals kinesin light chain-1 splicing as an amyloid-beta accumulation modifier. Proc Natl Acad Sci U S A 111: 2638-2643
Norris A. D., Gao S., Norris M. L., Ray D., Ramani A. K., et al. (2014) A pair of RNA-binding proteins controls networks of splicing events contributing to specialization of neural cell types. Mol Cell 54: 946-959
Oldmeadow C., Mossman D., Evans T. J., Holliday E. G., Tooney P. A., et al. (2014) Combined analysis of exon splicing and genome wide polymorphism data predict schizophrenia risk loci. J Psychiatr Res 52: 44-49
Screen M., Jonson P. H., Raheem O., Palmio J., Laaksonen R., et al. (2014) Abnormal splicing of NEDD4 in myotonic dystrophy type 2: possible link to statin adverse reactions. Am J Pathol 184: 2322-2332

28 Neurobiology Research Review
Epigenetic Modifications
The extensive role of epigenetic changes in the onset and progression of psychiatric
diseases has recently become apparent due to the development of microarray-
and NGS-based protocols for detecting genome-wide epigenetic patterns. This
role appeared to be especially strong in neurodegenerative diseases; however,
some recently discovered epigenetic patterns in psychiatric disorders may be
critical for better understanding the causes and patterns of these diseases. One of
the complications of studying the epigenetics in neurological diseases is that the
signature can only be detected post-mortem, and stability of samples and these
modifications is often compromised.171
Genomic imprinting is an example of an epigenetic modification that occurs
throughout life.172 SHANK3, the first gene associated with ASD, has five CpG
islands across the gene that display brain- and cell-type–specific DNA methylation
patterns.173,174,175 Similar specificity was observed for histone acetylation in this
gene.176 These modifications regulate the expression of the SHANK3 gene in an
isoform-specific manner.177 Several groups have independently identified other
multiple differentially methylated regions (DMRs) in post-mortem samples of autistic
patients representing biologically diverse gene regions, such as DNase-hypersensitive
sites and an alternative transcript termination site.178,179,180 These studies provide an
additional level of evidence to unravel the mechanisms of complex diseases,
such as ASD.
171. Bakulski K. M., Rozek L. S., Dolinoy D. C., Paulson H. L. and Hu H. (2012) Alzheimer's disease and environmental exposure to lead: the epidemiologic evidence and potential role of epigenetics. Curr Alzheimer Res 9: 563-573
172. Banerjee S., Riordan M. and Bhat M. A. (2014) Genetic aspects of autism spectrum disorders: insights from animal models. Front Cell Neurosci 8: 58
173. Beri S., Tonna N., Menozzi G., Bonaglia M. C., Sala C., et al. (2007) DNA methylation regulates tissue-specific expression of Shank3. J Neurochem 101: 1380-1391
174. Ching T. T., Maunakea A. K., Jun P., Hong C., Zardo G., et al. (2005) Epigenome analyses using BAC microarrays identify evolutionary conservation of tissue-specific methylation of SHANK3. Nat Genet 37: 645-651
175. Maunakea A. K., Nagarajan R. P., Bilenky M., Ballinger T. J., D'Souza C., et al. (2010) Conserved role of intragenic DNA methylation in regulating alternative promoters. Nature 466: 253-257
176. Kriukiene E., Labrie V., Khare T., Urbanaviciute G., Lapinaite A., et al. (2013) DNA unmeth-ylome profiling by covalent capture of CpG sites. Nat Commun 4: 2190
177. Jiang Y. H. and Ehlers M. D. (2013) Modeling autism by SHANK gene mutations in mice. Neuron 78: 8-27
178. Ladd-Acosta C., Hansen K. D., Briem E., Fallin M. D., Kaufmann W. E., et al. (2013) Common DNA methylation alterations in multiple brain regions in autism. Mol Psychiatry
179. Ladd-Acosta C., Hansen K. D., Briem E., Fallin M. D., Kaufmann W. E., et al. (2013) Common DNA methylation alterations in multiple brain regions in autism. Mol Psychiatry
180. Wong C. C., Meaburn E. L., Ronald A., Price T. S., Jeffries A. R., et al. (2014) Methylomic analysis of monozygotic twins discordant for autism spectrum disorder and related be-havioural traits. Mol Psychiatry 19: 495-503

29 An Overview of Publications Featuring Illumina® Technology
Aging and AD—in particular, LOAD—are also associated with a spectrum
of epigenetic changes, including abnormal DNA methylation and histone
modifications.181,182 These changes can be precipitated by physiological and
environmental conditions, such as stroke,183 hypertension, type II diabetes, obesity,184
exposure to heavy metals,185,186 and head injury.187 Oxidative stress, for example, can
cause an imbalance between methylation and demethylation of DNA in AD brains.188
Changes in histone tail modifications (primarily decreased levels of H3 acetylation in
the temporal lobe, and increased levels of histone deacetylases HDAC2 and HDAC6)
have also been observed in post-mortem brains of AD patients.189,190,191 Targeting
these histone modifications by therapeutic agents is a potential strategy for the
treatment of AD.192 Experiments in mouse have shown that pharmacological inhibition
of DNA methylation in the hippocampus of mice after a learning task impaired
memory consolidation,193 and promotion of histone acetylation had an opposite
effect: it increased learning and memory through increased learning-related gene
expression in aged mice.194,195
181. Chouliaras L., Rutten B. P., Kenis G., Peer-booms O., Visser P. J., et al. (2010) Epigenetic regulation in the pathophysiology of Alzhei-mer's disease. Prog Neurobiol 90: 498-510
182. Wang S. C., Oelze B. and Schumacher A. (2008) Age-specific epigenetic drift in late-on-set Alzheimer's disease. PLoS One 3: e2698
183. Savva G. M., Stephan B. C. and Alzheimer's Society Vascular Dementia Systematic Review G. (2010) Epidemiological studies of the effect of stroke on incident dementia: a systematic review. Stroke 41: e41-46
184. Huang Y. and Mucke L. (2012) Alzheimer mechanisms and therapeutic strategies. Cell 148: 1204-1222
185. Bakulski K. M., Rozek L. S., Dolinoy D. C., Paulson H. L. and Hu H. (2012) Alzheimer's disease and environmental exposure to lead: the epidemiologic evidence and potential role of epigenetics. Curr Alzheimer Res 9: 563-573
186. Basha M. R., Wei W., Bakheet S. A., Benitez N., Siddiqi H. K., et al. (2005) The fetal basis of amyloidogenesis: exposure to lead and latent overexpression of amyloid precursor protein and beta-amyloid in the aging brain. J Neurosci 25: 823-829
187. Vidaki A., Daniel B. and Court D. S. (2013) Forensic DNA methylation profiling--potential opportunities and challenges. Forensic Sci Int Genet 7: 499-507
188. Lu H., Liu X., Deng Y. and Qing H. (2013) DNA methylation, a hand behind neurodegenerative diseases. Front Aging Neurosci 5: 85
189. Zhang K., Schrag M., Crofton A., Trivedi R., Vinters H., et al. (2012) Targeted proteomics for quantification of histone acetylation in Alz-heimer's disease. Proteomics 12: 1261-1268
190. Ding H., Dolan P. J. and Johnson G. V. (2008) Histone deacetylase 6 interacts with the micro-tubule-associated protein tau. J Neurochem 106: 2119-2130
191. Graff J., Rei D., Guan J. S., Wang W. Y., Seo J., et al. (2012) An epigenetic blockade of cognitive functions in the neurodegenerating brain. Nature 483: 222-226
192. Coppede F. (2014) The potential of epigenetic therapies in neurodegenerative diseases. Front Genet 5: 220
193. Kilgore M., Miller C. A., Fass D. M., Hennig K. M., Haggarty S. J., et al. (2010) Inhibitors of class 1 histone deacetylases reverse contextual memory deficits in a mouse model of Alzheimer's disease. Neuropsychopharma-cology 35: 870-880
194. Fischer A., Sananbenesi F., Wang X., Dobbin M. and Tsai L. H. (2007) Recovery of learning and memory is associated with chromatin remodelling. Nature 447: 178-182
195. Peleg S., Sananbenesi F., Zovoilis A., Burkhardt S., Bahari-Javan S., et al. (2010) Altered histone acetylation is associated with age-dependent memory impairment in mice. Science 328: 753-756

30 Neurobiology Research Review
Epigenetic modifications are also critical for the development of PD. Reduced
methylation levels of the SNCA gene that encodes α-synuclein protein were detected
in the substantia nigra, cortex, and putamen regions of brain in patients with sporadic
PD.196,197 The protein α-synuclein binds directly to histone H3 and inhibits
histone acetylation.198
ReviewsCoppede F. (2014) The potential of epigenetic therapies in neurodegenerative diseases. Front Genet 5: 220
ReferencesDe Jager P. L., Srivastava G., Lunnon K., Burgess J., Schalkwyk L. C., et al. (2014) Alzheimer's disease: early alterations in brain DNA methylation at ANK1, BIN1, RHBDF2 and other loci. Nat Neurosci 17: 1156-1163DNA methylation is a genetic mechanism that may affect gene expression and, as such, may be implicated in disease susceptibility. The epigenomic influence on AD onset and progression was examined in this study using Illumina HumanMethylation450 arrays and bisulfite sequencing on Illumina HiSeq 2000. The authors found several replicated, functionally validated associations between altered DNA methylation and the pre-symptomatic accumulation of AD pathology. They hypothesized that the observed DNA methylation changes may be involved in the onset of AD.
Illumina Technology: HumanMethylation450, HiSeq 2000
Hollerhage M., Goebel J. N., de Andrade A., Hildebrandt T., Dolga A., et al. (2014) Trifluoperazine rescues human dopaminergic cells from wild-type alpha-synuclein-induced toxicity. Neurobiol Aging 35: 1700-1711PD is the most frequent neurodegenerative movement disorder. To study α-synuclein-mediated toxicity in PD progression, the authors developed a new cell-line model in which moderate overexpression of wild-type α-synuclein led to gradual death of human post-mitotic dopaminergic neurons. Using Illumina BeadArrays to monitor gene expression, the authors discovered that activating autophagy in human dopaminergic mid-brain neurons rescued them fromα-synuclein-induced cell death. The phenothiazine neuroleptic trifluoperazine, an activator of macroautophagy, may be a potential therapeutic target.
Illumina Technology: Human Gene Expression BeadArray
Ladd-Acosta C., Hansen K. D., Briem E., Fallin M. D., Kaufmann W. E., et al. (2014) Common DNA methylation alterations in multiple brain regions in autism. Mol Psychiatry 19: 862-871Genetic as well as environmental factors are responsible for ASDs. In this study, the authors measured over 485,000 CpG loci in human brain tissue and identified 4 genome-wide significantly different DMRs from 19 autism cases. This study highlights a new selected set of affected genes.
Illumina Technology: HumanMethylation450
Lunnon K., Smith R., Hannon E., De Jager P. L., Srivastava G., et al. (2014) Methylomic profiling implicates cortical deregulation of ANK1 in Alzheimer's disease. Nat Neurosci 17: 1164-1170In this study of AD methylomic variation, the authors used Illumina HumanMethylation450k arrays to characterize the genome-wide DNA methylation state across multiple tissues. Based on the results from 122 donor samples, the authors compared the methylation state in four brain regions and whole blood where available. The authors identified evidence for cortex-specific hypermethylation at CpG sites in the ANK1 gene associated with AD neuropathology.
Illumina Technology: HumanMethylation450k, Human Gene Expression BeadArray
196. Jowaed A., Schmitt I., Kaut O. and Wullner U. (2010) Methylation regulates alpha-synuclein expression and is decreased in Parkinson's disease patients' brains. J Neurosci 30: 6355-6359
197. Matsumoto L., Takuma H., Tamaoka A., Kuri-saki H., Date H., et al. (2010) CpG demeth-ylation enhances alpha-synuclein expression and affects the pathogenesis of Parkinson's disease. PLoS One 5: e15522
198. Coppede F. (2014) The potential of epigenetic therapies in neurodegenerative diseases. Front Genet 5: 220

31 An Overview of Publications Featuring Illumina® Technology
Nalls M. A., Pankratz N., Lill C. M., Do C. B., Hernandez D. G., et al. (2014) Large-scale meta-analysis of genome-wide association data identifies six new risk loci for Parkinson's disease. Nat Genet 46: 989-993Only a small fraction of genetic heritability has been discovered for PD to date. This study performed a meta-analysis of GWAS for PD in the search for new loci associated with the disease. Using Illumina genotyping arrays, the authors identified 24 loci that were both statistically significant and replicated across experiments. Six of the identified loci had not been previously reported associated with PD, and the authors estimated the cumulative risk of the loci to be substantial (odds ratio = 3.31).
Illumina Technology: ExomeChip, HumanOmniExpress, HumanHap550, Human610-Quad, Human660W-Quad; HumanMethylation27, Human Gene Expression BeadArray
Wong C. C., Meaburn E. L., Ronald A., Price T. S., Jeffries A. R., et al. (2014) Methylomic analysis of monozygotic twins discordant for autism spectrum disorder and related behavioural traits. Mol Psychiatry 19: 495-503

32 Neurobiology Research Review
Small RNAs
MicroRNA (miRNA) is enriched in the brain, and neuronal-specific miRNA controls
neuronal differentiation, excitability, and function.199 Other RNAs, such as non-coding
RNA (ncRNA), appear to play a role in neurodevelopment.
A number of miRNAs are associated with AD and PD, and were detected not only
in brains, but also in peripheral tissues of affected individuals.200,201 This observation
suggests that minimally invasive diagnostic tools could be developed for early
prediction of neurodegenerative disorders. miRNAs have also been considered as
therapeutic agents against AD pathogens, such as APP202. miRNA molecules can
be conjugated to—or otherwise associated with—aptamers, monoclonal antibodies,
peptides, or exosomes for delivery in vivo to specific cell types and tissues.203
ReviewsSekiyama K., Takamatsu Y., Waragai M. and Hashimoto M. (2014) Role of genomics in translational research for Parkinson's disease. Biochem Biophys Res Commun 452: 226-235
Maciotta S., Meregalli M. and Torrente Y. (2013) The involvement of microRNAs in neurodegenerative diseases. Front Cell Neurosci 7: 265
Mills J. D. and Janitz M. (2012) Alternative splicing of mRNA in the molecular pathology of neurodegenerative diseases. Neurobiol Aging 33: 1012 e1011-1024
ReferencesRaj T., Ryan K. J., Replogle J. M., Chibnik L. B., Rosenkrantz L., et al. (2014) CD33: increased inclusion of exon 2 implicates the Ig V-set domain in Alzheimer's disease susceptibility. Hum Mol Genet 23: 2729-2736The identification of possible therapeutic targets requires a functional identification follow-up to known genetic disease variants. In this study, a previously identified risk allele for AD was examined in detail. The authors used expression data in population cohorts stratified by their genetic risk variant, identified by Illumina Infinium arrays. They found that the risk allele rs3865444(C) results in a higher surface density of CD33 on monocytes. The risk allele is strongly associated with greater expression of CD33 exon 2, which is likely to be the functional consequence of the risk variant.
Illumina Technology: OmniExpress
Bai B., Hales C. M., Chen P. C., Gozal Y., Dammer E. B., et al. (2013) U1 small nuclear ribonucleoprotein complex and RNA splicing alterations in Alzheimer's disease. Proc Natl Acad Sci U S A 110: 16562-16567Many neurodegenerative diseases are characterized by deposition of insoluble protein aggregates. The universal presence of ß-amyloid and tau proteins in AD has facilitated advancement of the amyloid cascade and tau hypotheses that have dominated AD pathogenesis research and therapeutic development. This study investigated the human brain-insoluble proteome in AD by mass spectrometry and transcriptome sequencing. The authors identified 36 proteins that accumulate in the disease and found similarities with protein aggregates in mild cognitive impairment.
Illumina Technology: HiSeq 2000 (mRNA sequencing)
Li M. M., Jiang T., Sun Z., Zhang Q., Tan C. C., et al. (2014) Genome-wide microRNA expression profiles in hippocampus of rats with chronic temporal lobe epilepsy. Sci Rep 4: 4734
199. Maciotta S., Meregalli M. and Torrente Y. (2013) The involvement of microRNAs in neu-rodegenerative diseases. Front Cell Neurosci 7: 265
200. Maciotta S., Meregalli M. and Torrente Y. (2013) The involvement of microRNAs in neu-rodegenerative diseases. Front Cell Neurosci 7: 265
201. Sekiyama K., Takamatsu Y., Waragai M. and Hashimoto M. (2014) Role of genomics in translational research for Parkinson's disease. Biochem Biophys Res Commun 452: 226-235
202. Maciotta S., Meregalli M. and Torrente Y. (2013) The involvement of microRNAs in neu-rodegenerative diseases. Front Cell Neurosci 7: 265
203. Maciotta S., Meregalli M. and Torrente Y. (2013) The involvement of microRNAs in neu-rodegenerative diseases. Front Cell Neurosci 7: 265

33 An Overview of Publications Featuring Illumina® Technology
Genetic Variants
Genome Wide Association StudiesGenome-wide association studies (GWAS) are conducted to identify common
disease susceptibility alleles by comparing allele frequencies across the genome
in large groups of cases and controls.204,205 This approach has yielded an
unprecedented amount of clinically relevant data, including the majority of mutations
and variants associated with AD and PD.207-211 However, in spite of over 9,000 GWAS
published to date, this approach has uncovered only a fraction of the true heritability.
Additionally, GWAS can miss epigenetic patterns, such as methylation, and it
can mistakenly identify genes in the vicinity of pathogenic213 SNVs as pathogenic.
The sequencing of entire genomes in large cohorts at affordable prices is likely to
generate additional genes, pathways, and biological insights, as well as the potential
to identify causal mutations.214 NGS, both alone or in combination with microarrays,
can address most of these limitations and significantly improve the results from
these studies.215
ReviewsSharma M., Kruger R. and Gasser T. (2014) From genome-wide association studies to next-generation sequencing: lessons from the past and planning for the future. JAMA Neurol 71: 5-6
Keogh M. J. and Chinnery P. F. (2013) Next generation sequencing for neurological diseases: new hope or new hype? Clin Neurol Neurosurg 115: 948-953e>
Koboldt D. C., Steinberg K. M., Larson D. E., Wilson R. K. and Mardis E. R. (2013) The next-generation sequencing revolution and its impact on genomics. Cell 155: 27-38
ReferencesKarayannis T., Au E., Patel J. C., Kruglikov I., Markx S., et al. (2014) Cntnap4 differentially contributes to GABAergic and dopaminergic synaptic transmission. Nature 511: 236-240In an effort to understand the pathological development of neurological disorders, genetic effects have been studied in the context of proteins that are expressed in neural cells. In this study, the authors characterized the effect of CNTNAP4 knockouts on mouse behavior and development, and relate these results to the findings of CNVs in humans across a region including the CNTNAP2 gene. The authors found that Cntnap4 is localized presynaptically, and its loss leads to a reduction in the output of cortical parvalbumin (PV)-positive GABAergic basket cells. In addition, CNTNAP4-mutant mice showed defects in these neuronal populations and exhibited sensory-motor gating and grooming endophenotypes.
Illumina Technology: HumanHap550, HumanOmni1
Nalls M. A., Pankratz N., Lill C. M., Do C. B., Hernandez D. G., et al. (2014) Large-scale meta-analysis of genome-wide association data identifies six new risk loci for Parkinson's disease. Nat Genet 46: 989-993Only a small fraction of genetic heritability has been discovered for PD to date. This study performed a meta-analysis of GWAS for PD in the search for new loci associated with the disease. Using Illumina genotyping arrays, the authors identified 24 loci that were both statistically significant and replicated across experiments. Six of the identified loci had not been previously reported associated with PD, and the authors estimated the cumulative risk of the loci to be substantial (odds ratio = 3.31).
Illumina Technology: ExomeChip, HumanOmniExpress, HumanHap550, Human610-Quad, Human660W-Quad; HumanMethylation27, Human Gene Expression BeadArray
204. Koboldt D. C., Steinberg K. M., Larson D. E., Wilson R. K. and Mardis E. R. (2013) The next-generation sequencing revolution and its impact on genomics. Cell 155: 27-38
205. Keogh M. J. and Chinnery P. F. (2013) Next generation sequencing for neurological diseases: new hope or new hype? Clin Neurol Neurosurg 115: 948-953e>
206. Sharma M., Kruger R. and Gasser T. (2014) From genome-wide association studies to next-generation sequencing: lessons from the past and planning for the future. JAMA Neurol 71: 5-6
207. Harold D., Abraham R., Hollingworth P., Sims R., Gerrish A., et al. (2009) Genome-wide as-sociation study identifies variants at CLU and PICALM associated with Alzheimer's disease. Nat Genet 41: 1088-1093
208. Hollingworth P., Harold D., Sims R., Gerrish A., Lambert J. C., et al. (2011) Common variants at ABCA7, MS4A6A/MS4A4E, EPHA1, CD33 and CD2AP are associated with Alzheimer's disease. Nat Genet 43: 429-435
209. Lambert J. C., Heath S., Even G., Campion D., Sleegers K., et al. (2009) Genome-wide association study identifies variants at CLU and CR1 associated with Alzheimer's disease. Nat Genet 41: 1094-1099
210. Naj A. C., Jun G., Beecham G. W., Wang L. S., Vardarajan B. N., et al. (2011) Common variants at MS4A4/MS4A6E, CD2AP, CD33 and EPHA1 are associated with late-onset Alzheimer's disease. Nat Genet 43: 436-441
211. Bras J., Guerreiro R. and Hardy J. (2012) Use of next-generation sequencing and other whole-genome strategies to dissect neurologi-cal disease. Nat Rev Neurosci 13: 453-464
212. Sharma M., Kruger R. and Gasser T. (2014) From genome-wide association studies to next-generation sequencing: lessons from the past and planning for the future. JAMA Neurol 71: 5-6
213. Sekiyama K., Takamatsu Y., Waragai M. and Hashimoto M. (2014) Role of genomics in translational research for Parkinson's disease. Biochem Biophys Res Commun
214. Visscher P. M., Brown M. A., McCarthy M. I. and Yang J. (2012) Five years of GWAS discovery. Am J Hum Genet 90: 7-24
215. Sharma M., Kruger R. and Gasser T. (2014) From genome-wide association studies to next-generation sequencing: lessons from the past and planning for the future. JAMA Neurol 71: 5-6

34 Neurobiology Research Review
Cruchaga C., Kauwe J. S., Harari O., Jin S. C., Cai Y., et al. (2013) GWAS of cerebrospinal fluid tau levels identifies risk variants for Alzheimer's disease. Neuron 78: 256-268The progress of AD can be monitored by the tau phosphorylated threonine 181 (ptau) in CSF. To identify the genetic mechanism associated with elevated ptau, the authors performed the largest GWAS to date, enrolling 1,269 participants. The participants were genotyped using Illumina OmniExpress arrays and tau/ptau levels measured. The authors identified three genome-wide significant loci for CSF tau and ptau; one of these showed a strong association with AD risk in independent data sets.
Illumina Technology: Human610-Quad, HumanOmniExpress
Next-Generation SequencingNext-generation sequencing for the analysis of eukaryotic DNA genome consists
of two modalities: WGS and WES.216 A combination of WES with custom-designed
microarrays is the method of choice for large sample sizes.217 This combinatory
approach allows the effective resolution of such common genetic analysis
problems as pseudogenes, repeated exons, and a failure to detect rare and/
or novel mutations.218 Additionally, WES per se is not very adequate currently for
addressing CNVs, because sample preparation relies on non-quantitative PCR
amplification.219 However, an approach that combines WES with genotyping resolves
this complication.220
A multi-pronged approach—supplementing WES with WGS, proteomics, and
epigenomics—is anticipated to deliver full understanding of the effects of newly
discovered genetic variability. Rare mutations often have significantly stronger effects
on disease pathology than common mutations, and that effect has been especially
remarkable in the field of complex diseases. The power of NGS was further proved in
the discovery of de novo mutations, which arise in individuals during their life and are
more likely to have functional roles in rare diseases. Although the appearance of such
mutations seems stochastic, the mutation rate and its dependence on parental age222
and other environmental factors are only some of the important results of using
this technology.223
Sequencing is becoming indispensable for diagnosing disease by analyzing
circulating tumor DNA. Not only the sequence of that DNA, but also fluctuations in
cell count, often correlate with the disease pathology and state. The “liquid biopsy
approach” allows the detection of somatic mutations for a specific tumor type in
plasma or liquid Papanicolaou (Pap) smears.224,225 This approach is currently being
developed to detect PD226 and AD227,228 in peripheral blood. The recent discovery
that the whole fetal genome is present in maternal plasma during pregnancy230 has
launched the era of pre-natal non-invasive genetic diagnostics.
“Unlike GWAS which examines common mutations, sequencing facilitates the discovery of rare mutations which often associate with complex phenotypes.”Koboldt et al. 2013
216. Medland S. E., Jahanshad N., Neale B. M. and Thompson P. M. (2014) Whole-genome analyses of whole-brain data: working within an expanded search space. Nat Neurosci 17: 791-800
217. Bras J., Guerreiro R. and Hardy J. (2012) Use of next-generation sequencing and other whole-genome strategies to dissect neurologi-cal disease. Nat Rev Neurosci 13: 453-464
218. Bras J., Guerreiro R. and Hardy J. (2012) Use of next-generation sequencing and other whole-genome strategies to dissect neurologi-cal disease. Nat Rev Neurosci 13: 453-464
219. Bras J., Guerreiro R. and Hardy J. (2012) Use of next-generation sequencing and other whole-genome strategies to dissect neurologi-cal disease. Nat Rev Neurosci 13: 453-464
220. Bras J., Guerreiro R. and Hardy J. (2012) Use of next-generation sequencing and other whole-genome strategies to dissect neurologi-cal disease. Nat Rev Neurosci 13: 453-464
221. Bras J., Guerreiro R. and Hardy J. (2012) Use of next-generation sequencing and other whole-genome strategies to dissect neurologi-cal disease. Nat Rev Neurosci 13: 453-464
222. Kong A., Frigge M. L., Masson G., Besen-bacher S., Sulem P., et al. (2012) Rate of de novo mutations and the importance of father's age to disease risk. Nature 488: 471-475
223. Koboldt D. C., Steinberg K. M., Larson D. E., Wilson R. K. and Mardis E. R. (2013) The next-generation sequencing revolution and its impact on genomics. Cell 155: 27-38
224. Forshew T., Murtaza M., Parkinson C., Gale D., Tsui D. W., et al. (2012) Noninvasive identi-fication and monitoring of cancer mutations by targeted deep sequencing of plasma DNA. Sci Transl Med 4: 136ra168
225. Kinde I., Bettegowda C., Wang Y., Wu J., Agrawal N., et al. (2013) Evaluation of DNA from the Papanicolaou test to detect ovarian and endometrial cancers. Sci Transl Med 5: 167ra164
226. Karlsson M. K., Sharma P., Aasly J., Toft M., Skogar O., et al. (2013) Found in transcription: accurate Parkinson's disease classification in peripheral blood. J Parkinsons Dis 3: 19-29
227. Booij B. B., Lindahl T., Wetterberg P., Skaane N. V., Saebo S., et al. (2011) A gene expres-sion pattern in blood for the early detection of Alzheimer's disease. J Alzheimers Dis 23: 109-119
228. Rye P. D., Booij B. B., Grave G., Lindahl T., Kristiansen L., et al. (2011) A novel blood test for the early detection of Alzheimer's disease. J Alzheimers Dis 23: 121-129
229. Fehlbaum-Beurdeley P., Sol O., Desire L., Touchon J., Dantoine T., et al. (2012) Validation of AclarusDx, a blood-based transcriptomic signature for the diagnosis of Alzheimer's disease. J Alzheimers Dis 32: 169-181
230. Lo Y. M., Chan K. C., Sun H., Chen E. Z., Jiang P., et al. (2010) Maternal plasma DNA sequencing reveals the genome-wide genetic and mutational profile of the fetus. Sci Transl Med 2: 61ra91

35 An Overview of Publications Featuring Illumina® Technology
ReviewsRabbani B., Tekin M. and Mahdieh N. (2014) The promise of whole-exome sequencing in medical genetics. J Hum Genet 59: 5-15
Koboldt D. C., Steinberg K. M., Larson D. E., Wilson R. K. and Mardis E. R. (2013) The next-generation sequencing revolution and its impact on genomics. Cell 155: 27-38
Bras J., Guerreiro R. and Hardy J. (2012) Use of next-generation sequencing and other whole-genome strategies to dissect neurological disease. Nat Rev Neurosci 13: 453-464
Malhotra D. and Sebat J. (2012) CNVs: harbingers of a rare variant revolution in psychiatric genetics. Cell 148: 1223-1241
ReferencesDe Jager P. L., Srivastava G., Lunnon K., Burgess J., Schalkwyk L. C., et al. (2014) Alzheimer's disease: early alterations in brain DNA methylation at ANK1, BIN1, RHBDF2 and other loci. Nat Neurosci 17: 1156-1163DNA methylation is a genetic mechanism that may affect gene expression and, as such, may be implicated in disease susceptibility. The epigenomic influence on AD onset and progression was examined in this study using Illumina HumanMethylation450 arrays and bisulfite sequencing on Illumina HiSeq 2000. The authors found several replicated, functionally validated associations between altered DNA methylation and the pre-symptomatic accumulation of AD pathology. They hypothesized that the observed DNA methylation changes may be involved in the onset of AD.
Illumina Technology: HumanMethylation450, HiSeq 2000
Fromer M., Pocklington A. J., Kavanagh D. H., Williams H. J., Dwyer S., et al. (2014) De novo mutations in schizophrenia implicate synaptic networks. Nature 506: 179-184Of the known risk alleles for schizophrenia, the only ones definitively shown to confer considerable increments in risk are rare chromosomal CNVs that involve deletion or duplication of thousands of bases of DNA. This study examined the effect of small de novo mutations affecting one or a few nucleotides. By Illumina HiSeq WES of 623 schizophrenia trios, the authors assessed de novo mutation rates and shared genetic etiology for schizophrenia, intellectual disability, and ASDs. They found several insights to suggest a common etiological mechanism.
Illumina Technology: HiSeq (exome sequencing)
Lee H., Lin M. C., Kornblum H. I., Papazian D. M. and Nelson S. F. (2014) Exome sequencing identifies de novo gain of function missense mutation in KCND2 in identical twins with autism and seizures that slows potassium channel inactivation. Hum Mol Genet 23: 3481-3489Numerous studies have reported comorbidity of autism and epilepsy, but the relationship between the two disorders is unknown. In this study, identical twins, affected by both autism and severe intractable seizures, were studied using exome sequencing. A novel variant in the KCND2 gene was observed in both twins. The de novo mutation is located in the protein coding the Kv4.2 potassium channel, and the authors expressed the mutant protein in Xenopus oocytes to observe functional effects. Expression analysis showed that the mutation dominantly impairs the closed-state inactivation of the potassium channel, strongly supporting KCND2 as the causal gene for epilepsy in this family.
Illumina Technology: HiSeq 2000, Illumina Paired-End Sequencing Library Prep Protocol
King I. F., Yandava C. N., Mabb A. M., Hsiao J. S., Huang H. S., et al. (2013) Topoisomerases facilitate transcription of long genes linked to autism. Nature 501: 58-62Topoisomerases are expressed throughout the developing and adult brain, and are mutated in some individuals with ASD. However, the mechanism by which topoisomerases impact ASD is unknown. By transcriptome sequencing, in combination with genome-wide mapping of RNA polymerase II density in neurons, the authors found that expression of long genes was reduced after knockdown of topoisomerase in neurons. The authors noted that many high-confidence ASD candidate genes are exceptionally long and were reduced in expression after TOP1 inhibition. This observation suggests that topoisomerases could contribute commonly to ASD.
Illumina Technology: HiSeq 2000, TruSeq RNA, TruSeq for ChIP-Seq

36 Neurobiology Research Review
Willsey A. J., Sanders S. J., Li M., Dong S., Tebbenkamp A. T., et al. (2013) Coexpression networks implicate human midfetal deep cortical projection neurons in the pathogenesis of autism. Cell 155: 997-1007Recent studies employing WES and WGS have identified nine high-confidence ASD genes. This study examined the contribution of these nine genes to the common phenotype by combining Illumina WES and RNA-Seq data into co-expression networks. The authors explain how these networks will guide future ASD research by indicating which genes are most likely to have overlapping molecular, cellular or circuit-level phenotypes.
Illumina Technology: HiSeq 2000, Genome Analyzer
Yu T. W., Chahrour M. H., Coulter M. E., Jiralerspong S., Okamura-Ikeda K., et al. (2013) Using whole-exome sequencing to identify inherited causes of autism. Neuron 77: 259-273Steel syndrome is a developmental structural disorder first described in 1993 in 23 Hispanic children from Puerto Rico. This paper presents the genomic analysis of a family with two affected siblings. The authors used whole-exome sequencing using the Baylor College of Medicine Human Genome Sequencing Center (BCM-HGSC) core design followed by sequencing of both affected siblings, parents, and an affected cousin and her unaffected parents. By filtering the detected genetic variants by segregation, the authors discovered a single homozygous missense variant segregating with the disorder. The variant disrupts the collagen gene COL27A1, which codes for a protein expressed in developing cartilage.
Illumina Technology: HiSeq 2000, HumanOmniExpress

37 An Overview of Publications Featuring Illumina® Technology
MODEL SYSTEMS
Modeling of neurological diseases has been challenging for two primary reasons:
extremely limited access to the primary brain tissue of individuals affected by these
diseases, and polygenicity of these disorders. Traditional knockout models reiterate
only a fraction of the disease phenotype, leaving room for speculation about the
relevance of research results to real human diseases. The introduction of triple-
knockout mice, transgenic rats, and stem cells as in vitro models has significantly
broadened the arsenal of tools available to researchers in the field and moved closer
to the “hypothetical” ideal disease model.231 The development of adequate model
systems is essential for developing accurate diagnostic and therapeutic strategies.
Animal Models
The development of animal models for schizophrenia and ASD has been challenging,
because over 90% of these diseases are polygenic. Therefore, a standard single-
knockout mouse model can only partially mimic the disease phenotype. Introduced
mutations are only tangentially relevant to the disease, and symptoms observed in
animals may represent other diseases in this spectrum.232 Another complication is
associated with the recognition and quantification of the symptoms: animal behavioral
patterns are different from those of a human; therefore, interpretation of behavioral
changes, feelings, and intentions of animals may be highly subjective.
SchizophreniaRodents have been the most commonly used models for studying schizophrenia and
ASD. Until recently, these models were limited to mice, but now some rat knockouts
have also become available.233 Rats may be beneficial over mice, as they are highly
social animals and possess a rich acoustic communication system (including
frequencies in the ultrasonic range) and have a closer resemblance to human neural
processes.234 Additionally, pre-clinical toxicology studies are normally carried out in
rats; therefore, use of these animals as a research model can significantly streamline
the drug development process.235
“Given the low penetrance of schizophrenia-associated alleles, and their ability to contribute to different diseases, inserting one or even several into an animal model may yield a phenotype that is ambiguous with respect to human disease — or no phenotype at all.” Hyman 2014
231. Lyons M. R. and West A. E. (2011) Mecha-nisms of specificity in neuronal activity-regulat-ed gene transcription. Prog Neurobiol 94: 259-295
232. Kvajo M., McKellar H. and Gogos J. A. (2012) Avoiding mouse traps in schizophrenia genet-ics: lessons and promises from current and emerging mouse models. Neuroscience 211: 136-164
233. Wohr M. and Scattoni M. L. (2013) Behavioural methods used in rodent models of autism spectrum disorders: current standards and new developments. Behav Brain Res 251: 5-17
234. Lipska B. K. and Weinberger D. R. (2000) To model a psychiatric disorder in animals: schizophrenia as a reality test. Neuropsycho-pharmacology 23: 223-239
235. Wohr M. and Scattoni M. L. (2013) Behavioural methods used in rodent models of autism spectrum disorders: current standards and new developments. Behav Brain Res 251: 5-17

38 Neurobiology Research Review
Several genetic mouse models of schizophrenia have been developed that use
one of the three traditional methods: conventional gene targeting, conditional gene
targeting, or point mutation by chemical mutagens.236 However, these techniques
allow for generation of phenotypes only scarcely similar to schizophrenia. For that
reason, Cre/loxP-based chromosome engineering technique was used to generate
models reflecting complex genomic rearrangements, such as large deletions,
inversions, and duplications.237
One of the oldest models of schizophrenia is the dominant-negative disrupted in
schizophrenia 1 (DISC1) gene. DISC1 mice are a good model for studying not only
schizophrenia, but also a dual diagnosis of schizophrenia and substance-abuse
disorder.238 The effects of this mutation on brain structure or function remain to
be studied.239
Autism Spectrum DisorderMouse models have allowed for the demonstration of fundamental principles of ASD
diseases. Mouse knockout models, with mutations in a range of genes, are making
a significant contribution to understanding disease onset. These genes include
SHANK3 (Phelan-McDermid syndrome, idiopatic ASD), MeCP2 (Rett syndrome),
Fragile X chromosome (FMR1), PTEN (autism), and others.240,241 SHANK3 is a good
example of the importance of reproducing the exact type and point of mutation of the
gene: some mutations in this gene are also associated with other diseases, including
schizophrenia and intellectual disability.242,243 Microduplications of SHANK3 were
also associated with developmental delay and dysmorphic features in children.244
This phenomenon underpins the need to use genomic analysis tools for accurate
identification of mutations in spectrum disorders, and for the verification of their
accurate reproduction in animal models.
Additional models of ASD include non-human primates, songbirds, zebrafish,
Drosophila, and C. elegans. Non-human primates have been instrumental in studying
the behavioral patterns in this disorder, as the anatomy of their neural circuits
responsible for mediating social behavior is very similar to that of humans.245 Like
humans, they possess mirror neurons—cells responsible for repeating the actions of
others, commonly damaged in autism. For ethical reasons, introduction of genetic
mutations into primates is currently not feasible.
Songbirds have been used as a model due to their well-developed vocal machinery.
As in humans, vocal learning is an important element of language in this species, and
its impairment is commonly associated with ASD.246 Finally, zebrafish, Drosophila,
and C. elegans have been used extensively to study the genetic fundamentals of
psychiatric diseases.247
236. Nomura J. and Takumi T. (2012) Animal models of psychiatric disorders that reflect human copy number variation. Neural Plast 2012: 589524
237. Nomura J. and Takumi T. (2012) Animal models of psychiatric disorders that reflect human copy number variation. Neural Plast 2012: 589524
238. Ng E., McGirr A., Wong A. H. and Roder J. C. (2013) Using rodents to model schizophrenia and substance use comorbidity. Neurosci Biobehav Rev 37: 896-910
239. Kvajo M., McKellar H. and Gogos J. A. (2012) Avoiding mouse traps in schizophrenia genetics: lessons and promises from current and emerging mouse models. Neuroscience 211: 136-164
240. Banerjee S., Riordan M. and Bhat M. A. (2014) Genetic aspects of autism spectrum disorders: insights from animal models. Front Cell Neurosci 8: 58
241. Gadad B. S., Hewitson L., Young K. A. and German D. C. (2013) Neuropathology and animal models of autism: genetic and environ-mental factors. Autism Res Treat 2013: 731935
242. Gauthier J., Champagne N., Lafreniere R. G., Xiong L., Spiegelman D., et al. (2010) De novo mutations in the gene encoding the synaptic scaffolding protein SHANK3 in patients ascer-tained for schizophrenia. Proc Natl Acad Sci U S A 107: 7863-7868
243. Tucker T., Zahir F. R., Griffith M., Delaney A., Chai D., et al. (2014) Single exon-resolution targeted chromosomal microarray analysis of known and candidate intellectual disability genes. Eur J Hum Genet 22: 792-800
244. Okamoto N., Kubota T., Nakamura Y., Murakami R., Nishikubo T., et al. (2007) 22q13 Microduplication in two patients with common clinical manifestations: a recognizable syn-drome? Am J Med Genet A 143A: 2804-2809
245. Watson K. K. and Platt M. L. (2012) Of mice and monkeys: using non-human primate models to bridge mouse- and human-based investigations of autism spectrum disorders. J Neurodev Disord 4: 21
246. Clayton D. F., Balakrishnan C. N. and London S. E. (2009) Integrating genomes, brain and behavior in the study of songbirds. Curr Biol 19: R865-873
247. Banerjee S., Riordan M. and Bhat M. A. (2014) Genetic aspects of autism spectrum disorders: insights from animal models. Front Cell Neurosci 8: 58

39 An Overview of Publications Featuring Illumina® Technology
Animal models used in research of psychiatric and neurodegenerative diseases: mice, rats, non-human primates, Drosophila, songbirds, and zebrafish.
Alzheimer’s DiseaseAnimal models of AD are challenging to develop, as spontaneous amyloidosis is not
common in laboratory animals. Aged non-human primates can develop ß-amyloidosis
and tau fibrillary inclusions; however, these animals do not develop the clinical signs
of human AD.248 Currently used animal models of AD are mostly limited to genetically
engineered mice.249 These models allowed for the successful mimicking of most
human cerebral amyloidosis, including ß-amyloidosis and taupathies.250 In all cases,
overexpression of human amyloidogenic protein was required.251
A mouse model overexpressing ß-amyloid protein has also been established.
Although neurofibrillary tangles are not produced in brains of these mice, tau
pathology is still observed, because ß-amyloid pathology activates kinases,
down-regulates phosphatases, and impairs tau degradation.252 Mice expressing
both mutated APP and tau demonstrate greater neurofibrillary tangle pathology
than mutated tau mice, thus suggesting a role of ß-amyloid accumulation in the
development of tau pathology.253
Apart from the previously mentioned models, knockout models for genes involved
in APP processing—such as presenelin-1, presenelin-2, and b-secretase enzyme
(BACE1) are now available. Unfortunately, none of them accurately mimics all the key
symptoms and molecular signatures of this disease. Specifically, increased neuronal
death has been one of such symptoms, and it seems to be necessary for selecting
and testing drugs against AD. A more advanced model was obtained by crossing
APP-overexpressing mice with transgenic animals expressing mutated presenilin-1
248. Jucker M. (2010) The benefits and limitations of animal models for translational research in neurodegenerative diseases. Nat Med 16: 1210-1214
249. Eisenberg D. and Jucker M. (2012) The amy-loid state of proteins in human diseases. Cell 148: 1188-1203
250. Eisenberg D. and Jucker M. (2012) The amy-loid state of proteins in human diseases. Cell 148: 1188-1203
251. Eisenberg D. and Jucker M. (2012) The amy-loid state of proteins in human diseases. Cell 148: 1188-1203
252. Blurton-Jones M. and Laferla F. M. (2006) Pathways by which Abeta facilitates tau pa-thology. Curr Alzheimer Res 3: 437-448
253. Lewis J., Dickson D. W., Lin W. L., Chisholm L., Corral A., et al. (2001) Enhanced neu-rofibrillary degeneration in transgenic mice expressing mutant tau and APP. Science 293: 1487-1491
254. Oddo S., Caccamo A., Shepherd J. D., Murphy M. P., Golde T. E., et al. (2003) Triple-transgenic model of Alzheimer's disease with plaques and tangles: intracellular Abeta and synaptic dysfunction. Neuron 39: 409-421

40 Neurobiology Research Review
or presenelin-2, with the third transgene (mutated tau) added to the system. These
mice featured accelerated ß-amyloid pathology, formation of neurofibrillary tangles,
neuronal loss and cognitive decline, and tau pathology. 254
Although significant progress has been made in the development of mouse models
of AD, the available models do not take into account genetic and epigenetic variability
or the immunological factors common in LOAD. Recent advances in NGS technology
are anticipated to intensify building of these models, which are essential to develop
disease-modifying drugs for AD.255
Parkinson’s diseaseThe etiopathogenesis of PD is not yet clarified, and existing animal models have many
limitations. Nevertheless, they allow for unraveling some fundamental mechanisms
underlying the molecular and cellular basis of this neurodegenerative disorder. Most
animal models of PD developed to date are toxic, rather than genetic, models. Toxic
(also known as pharmacological) models—in particular, neurotoxin-based models—
were most effective in reproducing dopaminergic neuron death and striatal dopamine
deficit in non-human primates and rodents. The additional advantage of toxic models
is the feasibility of their use in non-human primates, whose motor symptoms and
neuronal structure are very similar to those of a human. The only limitation is the lack
of formation of classic Lewy bodies in primate brains.
Genetic models of PD are very limited. The reason for this limited availability is the
low contribution of the genetic component to this disorder: only 5% to 10% of all
PD cases are inherited. The most common mutations in this form of PD are those
in LRRK2 (which encodes an enzyme that may be involved in the deregulation of
α-synuclein phosphorylation), 256 PINK1 (PTEN-induced putative kinase 1), and
Parkin (which participates in the ubiquitin proteasome system). Transgenic mice with
knockouts in one of these genes exhibit only part of the PD phenotype, such as
motility abnormalities, mitochondrial and nigrostriatal neurotransmission deficits,
and others.257
255. Ribeiro F. M., Camargos E. R., Souza L. C. and Teixeira A. L. (2013) Animal models of neurodegenerative diseases. Rev Bras Psiqui-atr 35 Suppl 2: S82-91
256. Scholz S. W., Mhyre T., Ressom H., Shah S. and Federoff H. J. (2012) Genomics and bioin-formatics of Parkinson's disease. Cold Spring Harb Perspect Med 2: a009449
257. Blandini F. and Armentero M. T. (2012) Animal models of Parkinson's disease. FEBS J 279: 1156-1166

41 An Overview of Publications Featuring Illumina® Technology
None of the single-gene transgenic mouse models featured substantial nigrostriatal
degeneration.258 Multi-gene transgenic mouse models are also available (with
α-synuclein and parkin or DJ-1 knockouts, or simultaneous silencing of PINK-1,
DJ-1, and parkin), but they too have only limited relevance to the symptomatic and
phenotypical spectrum of PD.259
Recently developed rat models with monogenic PD mutations are believed to be
advantageous over mouse models. Rat neuronal circuitry is closer to that of a
human, and they are less prone to anxiety than mice, which is important for the
evaluation of behavioral patterns. Transgenic rats with mutated α-synuclein do not
have major motor deficits, but do exhibit significant olfactory deficits.260 Rats with a
neuron-specific mutation in LRRK2, driven by adenoviral vectors, exhibit progressive
degeneration of nigral dopaminergic neurons.261,262
258. Chesselet M. F. (2008) In vivo alpha-synuclein overexpression in rodents: a useful model of Parkinson's disease? Exp Neurol 209: 22-27
259. Blandini F. and Armentero M. T. (2012) Animal models of Parkinson's disease. FEBS J 279: 1156-1166
260. Lelan F., Boyer C., Thinard R., Remy S., Usal C., et al. (2011) Effects of Human Alpha-Synu-clein A53T-A30P Mutations on SVZ and Local Olfactory Bulb Cell Proliferation in a Transgenic Rat Model of Parkinson Disease. Parkinsons Dis 2011: 987084
261. Zhou H., Huang C., Tong J., Hong W. C., Liu Y. J., et al. (2011) Temporal expression of mutant LRRK2 in adult rats impairs dopamine reuptake. Int J Biol Sci 7: 753-761
262. Dusonchet J., Kochubey O., Stafa K., Young S. M., Jr., Zufferey R., et al. (2011) A rat model of progressive nigral neurodegeneration in-duced by the Parkinson's disease-associated G2019S mutation in LRRK2. J Neurosci 31: 907-912

42 Neurobiology Research Review
ReviewsBanerjee S., Riordan M. and Bhat M. A. (2014) Genetic aspects of autism spectrum disorders: insights from animal models. Front Cell Neurosci 8: 58
Katsnelson A. (2014) Drug development: The modelling challenge. Nature 508: S8-9
Ebert D. H. and Greenberg M. E. (2013) Activity-dependent neuronal signalling and autism spectrum disorder. Nature 493: 327-337
Gadad B. S., Hewitson L., Young K. A. and German D. C. (2013) Neuropathology and animal models of autism: genetic and environmental factors. Autism Res Treat 2013: 731935
Jiang Y. H. and Ehlers M. D. (2013) Modeling autism by SHANK gene mutations in mice. Neuron 78: 8-27
Ng E., McGirr A., Wong A. H. and Roder J. C. (2013) Using rodents to model schizophrenia and substance use comorbidity. Neurosci Biobehav Rev 37: 896-910
Ribeiro F. M., Camargos E. R., Souza L. C. and Teixeira A. L. (2013) Animal models of neurodegenerative diseases. Rev Bras Psiquiatr 35 Suppl 2: S82-91
Wohr M. and Scattoni M. L. (2013) Behavioural methods used in rodent models of autism spectrum disorders: current standards and new developments. Behav Brain Res 251: 5-17
Blandini F. and Armentero M. T. (2012) Animal models of Parkinson's disease. FEBS J 279: 1156-1166
Crabtree D. M. and Zhang J. (2012) Genetically engineered mouse models of Parkinson's disease. Brain Res Bull 88: 13-32
Kvajo M., McKellar H. and Gogos J. A. (2012) Avoiding mouse traps in schizophrenia genetics: lessons and promises from current and emerging mouse models. Neuroscience 211: 136-164
Nomura J. and Takumi T. (2012) Animal models of psychiatric disorders that reflect human copy number variation. Neural Plast 2012: 589524
Pratt J., Winchester C., Dawson N. and Morris B. (2012) Advancing schizophrenia drug discovery: optimizing rodent models to bridge the translational gap. Nat Rev Drug Discov 11: 560-579

43 An Overview of Publications Featuring Illumina® Technology
STEM-CELL MODELS
One of the most promising models for studying neurological diseases is patient-
derived iPSCs. These cells would allow testing of new therapeutic approaches
directly on human neural tissue that contains molecular alterations typical for ASD.263
The use of iPSCs in animal models has already provided a tool to correct abnormal
synaptic morphology and physiology, and to reverse behavioral alterations, even in
symptomatic animals.264
Human iPSC have been used as a model for studying PD.265 Because only a limited
population of dopaminergic neurons located in the midbrain is most prone to
degeneration, the engrafted stem cells must be matched to those affected
by degeneration.266
Another stem-cell model has been used for the treatment of ASD in clinical trials—
mesenchymal stem cells (MSCs).267 Reportedly, they overturn ASD symptoms by
restoring integration into the neural network, facilitating the recovery of synaptic
plasticity, and releasing anti-inflammatory cytokines.268
263. Ghosh A., Michalon A., Lindemann L., Fontou-ra P. and Santarelli L. (2013) Drug discovery for autism spectrum disorder: challenges and opportunities. Nat Rev Drug Discov 12: 777-790
264. Marchetto M. C., Carromeu C., Acab A., Yu D., Yeo G. W., et al. (2010) A model for neural development and treatment of Rett syndrome using human induced pluripotent stem cells. Cell 143: 527-539
265. Sandoe J. and Eggan K. (2013) Opportunities and challenges of pluripotent stem cell neu-rodegenerative disease models. Nat Neurosci 16: 780-789
266. Sandoe J. and Eggan K. (2013) Opportunities and challenges of pluripotent stem cell neu-rodegenerative disease models. Nat Neurosci 16: 780-789
267. Siniscalco D., Bradstreet J. J., Sych N. and Antonucci N. (2014) Mesenchymal stem cells in treating autism: Novel insights. World J Stem Cells 6: 173-178
268. Siniscalco D., Bradstreet J. J., Sych N. and Antonucci N. (2014) Mesenchymal stem cells in treating autism: Novel insights. World J Stem Cells 6: 173-178

44 Neurobiology Research Review
ReviewsGhosh A., Michalon A., Lindemann L., Fontoura P. and Santarelli L. (2013) Drug discovery for autism spectrum disorder: challenges and opportunities. Nat Rev Drug Discov 12: 777-790
Sandoe J. and Eggan K. (2013) Opportunities and challenges of pluripotent stem cell neurodegenerative disease models. Nat Neurosci 16: 780-789
Jung Y. W., Hysolli E., Kim K. Y., Tanaka Y. and Park I. H. (2012) Human induced pluripotent stem cells and neurodegenerative disease: prospects for novel therapies. Curr Opin Neurol 25: 125-130
ReferencesYoon K. J., Nguyen H. N., Ursini G., Zhang F., Kim N. S., et al. (2014) Modeling a genetic risk for schizophrenia in iPSCs and mice reveals neural stem cell deficits associated with adherens junctions and polarity. Cell Stem Cell 15: 79-91Defects in brain development can contribute to the onset of neuropsychiatric disorders. This study set out to identify the functional role of the 15q11.2 deletion on neural development using induced pluripotent stem cell (iPSC)-derived human neural precursor cells (hNPCs) by RNA-Seq and SNP-genotyping arrays. They found that haploinsufficiency of CYFIP1, a gene within 15q11.2, affects radial glial cells, leading to their ectopic localization outside of the ventricular zone.
Illumina Technology: HumanOmni2.5S
Ryan S. D., Dolatabadi N., Chan S. F., Zhang X., Akhtar M. W., et al. (2013) Isogenic human iPSC Parkinson's model shows nitrosative stress-induced dysfunction in MEF2-PGC1alpha transcription. Cell 155: 1351-1364An association has been reported between PD and exposure to mitochondrial toxins. In this study, a stem cell model was used to characterize the response to toxins using Illumina BeadArray for gene expression analysis. The authors identified a pathway whereby basal and toxin-induced nitrosative/oxidative stress results in S-nitrosylation of transcription factor MEF2C. They reported the alteration contributing to mitochondrial dysfunction and apoptotic cell death, indicating a mechanism and potential therapeutic target for PD.
Illumina Technology: Human Gene Expression BeadArray
Zhang Y., Schulz V. P., Reed B. D., Wang Z., Pan X., et al. (2013) Functional genomic screen of human stem cell differentiation reveals pathways involved in neurodevelopment and neurodegeneration. Proc Natl Acad Sci U S A 110: 12361-12366

45 An Overview of Publications Featuring Illumina® Technology
BIOLOGY
Neurological and neurodegenerative diseases occur and develop as a result of
genetic and epigenetic mutations. However, many of these mutations are somatic
(non-inherited) and occur under the influence of biological factors, such as immune
activity, gut microbiome activity, and environmental factors. The contribution of
these factors at a multi-cell and single-cell level has long remained hypothetical and
controversial. However, the advent of high-resolution sequencing techniques now
allows the uncovering of these mechanisms and the significance of their contribution
to disease.
Immunity
The immune system, which was long believed to be independent from the central
nervous system (CNS), is now acknowledged to have an important contribution to
normal CNS function as well as to multiple neurological disorders.269,270 For example,
in PD, elevated levels of inflammatory cytokines are associated with more severe
forms of the disease, such as PD with dementia.271 In AD, an analysis of the two
largest GWAS has shown a significant overlap between disease-associated genes in
pathways associated with AD, cholesterol metabolism, and immune response.272 In
schizophrenia, a number of immune genes have been identified as genetic risk
factors associated with this disease.273,274,275 In autism, ongoing inflammation has
been determined as one of the common components of the disease. Interestingly,
fever in some autistic children was associated with improvement of their social
behavior, underpinning the involvement of inflammation in the symptomatic profile.276
“Both inflammation and oxidative stress tend to increase with age and are associated with a variety of chronic diseases. They have also been linked to neurodegeneration and proposed as factors that might contribute to schizophrenia.” Anthes 2014
269. Lindqvist D., Hall S., Surova Y., Nielsen H. M., Janelidze S., et al. (2013) Cerebrospinal fluid inflammatory markers in Parkinson's disease-associations with depression, fatigue, and cognitive impairment. Brain Behav Immun 33: 183-189Lynch M. A. and Mills K. H. (2012) Immunology meets neuroscience--opportuni-ties for immune intervention in neurodegenera-tive diseases. Brain Behav Immun 26: 1-10
270. Girgis R. R., Kumar S. S. and Brown A. S. (2014) The cytokine model of schizophrenia: emerging therapeutic strategies. Biol Psychia-try 75: 292-299
271. Lindqvist D., Hall S., Surova Y., Nielsen H. M., Janelidze S., et al. (2013) Cerebrospinal fluid inflammatory markers in Parkinson's disease--associations with depression, fatigue, and cognitive impairment. Brain Behav Immun 33: 183-189
272. Jones L., Harold D. and Williams J. (2010) Genetic evidence for the involvement of lipid metabolism in Alzheimer's disease. Biochim Biophys Acta 1801: 754-761
273. Stefansson H., Meyer-Lindenberg A., Steinberg S., Magnusdottir B., Morgen K., et al. (2014) CNVs conferring risk of autism or schizophrenia affect cognition in controls. Nature 505: 361-366
274. Sinkus M. L., Adams C. E., Logel J., Freed-man R. and Leonard S. (2013) Expression of immune genes on chromosome 6p21.3-22.1 in schizophrenia. Brain Behav Immun 32: 51-62
275. Meyer U. (2011) Anti-inflammatory signaling in schizophrenia. Brain Behav Immun 25: 1507-1518
276. Curran L. K., Newschaffer C. J., Lee L. C., Crawford S. O., Johnston M. V., et al. (2007) Behaviors associated with fever in children with autism spectrum disorders. Pediatrics 120: e1386-1392

46 Neurobiology Research Review
The blood-brain barrier prevents penetration of many types of immune cells into the
brain; however, a very limited number of certain immune cell populations, such as
dendritic cells and microglia, reside in the brain and facilitate the removal of dead
neurons.277,278 Neurodegeneration is most often accompanied by the accumulation of
microglia and monocytes around amyloid plaques and dying neurons.279 According
to one hypothesis of PD origin, the death of dopaminergic neurons in PD per se may
be facilitated by neuroinflammation.280 Infection of neurons and neighboring glial cells
with viruses, such as Japanese encephalitis virus (JIV), can increase the vulnerability
of neurons to factors such as aging, oxidative stress, environmental stress, and
genetic predisposition.281 Furthermore, neurons express some molecules normally
attributed to the immune system, thus uncovering an intricate interplay between
neuronal and immune systems.282
Immunotherapy is considered one of the promising approaches to treating
neurological diseases. Examples of such treatment include celecoxib, an inhibitor of
cyclooxygenase-2, which has shown some improvement of schizophrenia symptoms
in four studies283, and anti-ß-amyloid antibody, used in multiple trials up to phase III as
an agent to remove pathogenic ß-amyloid plaques.284,285
ReviewsGirgis R. R., Kumar S. S. and Brown A. S. (2014) The cytokine model of schizophrenia: emerging therapeutic strategies. Biol Psychiatry 75: 292-299
Horvath S. and Mirnics K. (2014) Immune system disturbances in schizophrenia. Biol Psychiatry 75: 316-323
Lynch M. A. and Mills K. H. (2012) Immunology meets neuroscience--opportunities for immune intervention in neurodegenerative diseases. Brain Behav Immun 26: 1-10
Onore C., Careaga M. and Ashwood P. (2012) The role of immune dysfunction in the pathophysiology of autism. Brain Behav Immun 26: 383-392
Meyer U. (2011) Anti-inflammatory signaling in schizophrenia. Brain Behav Immun 25: 1507-1518
ReferencesSchizophrenia Working Group of the Psychiatric Genomics C. (2014) Biological insights from 108 schizophrenia-associated genetic loci. Nature 511: 421-427Schizophrenia is a highly heritable disorder, but the heritability is not found in a single gene effect. In this largest GWAS for schizophrenia to date, the authors used SNP arrays for 36,989 cases and 113,075 controls to determine genetic risk factors for the disorder. The authors found that the significant genetic associations were not randomly spread across the genome, but enriched among genes expressed in brain and genes that have been associated with typical co-morbidity diagnoses, such as ASD and intellectual disability. Interestingly, links were also enriched within genes related to immunity, which fits the existing hypothesis of immune dysregulation in schizophrenia.
Illumina Technology: Human1M, HumanOmni2.5, HumanOmniExpress, HumanHap550, Human610, HumanHap650Y, HumanHap300
277. Lynch M. A. and Mills K. H. (2012) Immunol-ogy meets neuroscience--opportunities for immune intervention in neurodegenerative diseases. Brain Behav Immun 26: 1-10
278. Meyer U. (2011) Anti-inflammatory signaling in schizophrenia. Brain Behav Immun 25: 1507-1518
279. Lynch M. A. and Mills K. H. (2012) Immunol-ogy meets neuroscience--opportunities for immune intervention in neurodegenerative diseases. Brain Behav Immun 26: 1-10
280. Lindqvist D., Hall S., Surova Y., Nielsen H. M., Janelidze S., et al. (2013) Cerebrospinal fluid inflammatory markers in Parkinson's disease--associations with depression, fatigue, and cognitive impairment. Brain Behav Immun 33: 183-189
281. Deleidi M. and Isacson O. (2012) Viral and inflammatory triggers of neurodegenerative diseases. Sci Transl Med 4: 121ps123
282. Boulanger L. M. (2009) Immune proteins in brain development and synaptic plasticity. Neuron 64: 93-109
283. Girgis R. R., Kumar S. S. and Brown A. S. (2014) The cytokine model of schizophrenia: emerging therapeutic strategies. Biol Psychia-try 75: 292-299
284. Lynch M. A. and Mills K. H. (2012) Immunol-ogy meets neuroscience--opportunities for immune intervention in neurodegenerative diseases. Brain Behav Immun 26: 1-10
285. www.clinicaltrials.gov

47 An Overview of Publications Featuring Illumina® Technology
METAGENOMICS
Metagenomics refers to the study of genomic DNA obtained from multiple micro-
organisms that often cannot be cultured in the laboratory. Humans carry 10 times
more bacterial cells than human cells, and 100 times more bacterial genes than the
human genome.286 The new generation of sequencing technology, with its ability to
sequence thousands of organisms in parallel, has proved to be uniquely suited to this
application. Recent technical improvements allow nearly complete genome assembly
from individual microbes directly from environmental samples or clinical specimens,
without the need for cultivation methods.287 This accumulation of sequence
information has greatly expanded the appreciation of the dynamic nature of microbial
populations, as well as their impact on the environment and human health. With this
extraordinary and powerful set of sequencing tools now available, it is no surprise
that metagenomics has become one of the fastest-growing scientific disciplines.
The concept of a gut-brain axis has been developed recently, to highlight the
important influence of the metagenome on mental processes. It has been shown
that the gut microbiome plays an unexpectedly important role in the development
of depression, anxiety, irritable bowel syndrome, and neurological diseases, such
as autism289,290 and schizophrenia.291 The gut-microbial products may impose their
effect on the brain through chromatin plasticity, which causes changes in neuronal
transcription.292 Hsiao et al. proposed the use of microbiome-mediated therapies for
treating neurodevelopmental disorders.293
Interestingly, some gut bacteria (defined as psychobiotics) have a positive effect
on mental health and foster brain activity.294 These living organisms produce such
neuroactive substances as g-aminobutyric acid and serotonin, and are beneficial not
only for healthy individuals, but also for those with psychiatric disorders.295
Reviews Malkki H. (2014) Neurodevelopmental disorders: human gut microbiota alleviate behavioural symptoms in a mouse model of autism spectrum disorder. Nat Rev Neurol 10: 60
Stilling R. M., Dinan T. G. and Cryan J. F. (2014) Microbial genes, brain & behaviour - epigenetic regulation of the gut-brain axis. Genes Brain Behav 13: 69-86
Dinan T. G., Stanton C. and Cryan J. F. (2013) Psychobiotics: a novel class of psychotropic. Biol Psychiatry 74: 720-726
Lamendella R., VerBerkmoes N. and Jansson J. K. (2012) 'Omics' of the mammalian gut--new insights into function. Curr Opin Biotechnol 23: 491-500
Moon C. and Stappenbeck T. S. (2012) Viral interactions with the host and microbiota in the intestine. Curr Opin Immunol 24: 405-410
Nicholson J. K., Holmes E., Kinross J., Burcelin R., Gibson G., et al. (2012) Host-gut microbiota metabolic interactions. Science 336: 1262-1267
Reyes A., Semenkovich N. P., Whiteson K., Rohwer F. and Gordon J. I. (2012) Going viral: next-generation sequencing applied to phage populations in the human gut. Nat Rev Microbiol 10: 607-617
Zarowiecki M. (2012) Metagenomics with guts. Nat Rev Microbiol 10: 674
286. Barwick B. G., Abramovitz M., Kodani M., Moreno C. S., Nam R., et al. (2010) Prostate cancer genes associated with TMPRSS2-ERG gene fusion and prognostic of biochemical recurrence in multiple cohorts. Br J Cancer 102: 570-576
287. Lasken R. S. (2013) Single-cell sequencing in its prime. Nat Biotechnol 31: 211-212
288. Stilling R. M., Dinan T. G. and Cryan J. F. (2014) Microbial genes, brain & behaviour - epigenetic regulation of the gut-brain axis. Genes Brain Behav 13: 69-86
289. Hsiao E. Y., McBride S. W., Hsien S., Sharon G., Hyde E. R., et al. (2013) Microbiota modulate behavioral and physiological abnor-malities associated with neurodevelopmental disorders. Cell 155: 1451-1463
290. Malkki H. (2014) Neurodevelopmental disorders: human gut microbiota alleviate behavioural symptoms in a mouse model of autism spectrum disorder. Nat Rev Neurol 10: 60
291. Girgis R. R., Kumar S. S. and Brown A. S. (2014) The cytokine model of schizophrenia: emerging therapeutic strategies. Biol Psychia-try 75: 292-299
292. Stilling R. M., Dinan T. G. and Cryan J. F. (2014) Microbial genes, brain & behaviour - epigenetic regulation of the gut-brain axis. Genes Brain Behav 13: 69-86
293. Hsiao E. Y., McBride S. W., Hsien S., Sharon G., Hyde E. R., et al. (2013) Microbiota modulate behavioral and physiological abnor-malities associated with neurodevelopmental disorders. Cell 155: 1451-1463
294. Dinan T. G., Stanton C. and Cryan J. F. (2013) Psychobiotics: a novel class of psychotropic. Biol Psychiatry 74: 720-726
295. Dinan T. G., Stanton C. and Cryan J. F. (2013) Psychobiotics: a novel class of psychotropic. Biol Psychiatry 74: 720-726

48 Neurobiology Research Review
ReferencesMarkle J. G., Frank D. N., Mortin-Toth S., Robertson C. E., Feazel L. M., et al. (2013) Sex differences in the gut microbiome drive hormone-dependent regulation of autoimmunity. Science 339: 1084-1088Microbial exposure and sex hormones exert potent effects on autoimmune diseases. This study examined the effect of early-life microbial exposure on sex hormone levels and autoimmune disease in a non-obese diabetic (NOD) mouse model. The microbiome was characterized using 16S rRNA Illumina sequencing. Comparing the effects across both male and female mice, the results indicate that alteration of the gut microbiome in early life potently suppresses autoimmunity in animals at high genetic risk for disease.
Illumina Technology: MiSeq
Maurice C. F., Haiser H. J. and Turnbaugh P. J. (2013) Xenobiotics shape the physiology and gene expression of the active human gut microbiome. Cell 152: 39-50The microorganisms in the human gut microbiome are known to impact digestive health, but their influence on health by the metabolism of xenobiotics, including antibiotics and drugs, is still unclear. In this metagenomics study, xenobiotic-responsive genes were found across multiple bacterial phyla involved in various metabolic and stress-response pathways. The results suggest that xenobiotics may have important implications for the human gut microbiome.
Illumina Technology: HiSeq to sequence the V4 region of the 16S rRNA gene
Schloissnig S., Arumugam M., Sunagawa S., Mitreva M., Tap J., et al. (2013) Genomic variation landscape of the human gut microbiome. Nature 493: 45-50Subjects sampled at varying time intervals exhibited individuality and temporal stability of SNV patterns, despite considerable composition changes of their gut microbiota. This observation indicates that individual-specific strains are not easily replaced, and that an individual might have a unique metagenomic genotype, which may be exploitable for personalized diet or drug intake.
Illumina Technology: Illumina reads obtained from European MetaHIT study296 and U.S. Human Microbiome Project297
296. Qin J., Li R., Raes J., Arumugam M., Burgdorf K. S., et al. (2010) A human gut microbial gene catalogue established by metagenomic sequencing. Nature 464: 59-65
297. Group N. H. W., Peterson J., Garges S., Giovanni M., McInnes P., et al. (2009) The NIH Human Microbiome Project. Genome Res 19: 2317-2323

49 An Overview of Publications Featuring Illumina® Technology
ENVIRONMENTAL FACTORS
Exposure to microbes298,299, as well as various industrial and agricultural chemicals
(especially pesticides), has been implicated as conferring risk for multiple
diseases, including neurodegenerative disorders . Environmental pesticides serve
as mitochondrial toxins and induce nitrosative stress that inhibits activity of the
myocyte-specific enhancer factor 2C (MEF2C). This factor is involved in cardiac
morphogenesis, myogenesis, and vascular development. MEF2C, in turn, inhibits the
expression of peroxisome proliferator-activated receptor-gamma coactivator 1 alpha
(PGC-1 α), and hence suppresses the neuroprotective function of this transcriptional
co-activator.302
Neurodegenerative disorders, including PD, often co-exist with metabolic diseases.
For example, type 2 diabetes may stimulate the development of PD.303 Sekiyama et
al. suggest that genomic studies should contribute to better understanding of the
mechanism of this interaction, and help to develop strategies for new
therapeutic approaches.304
ReviewsSekiyama K., Takamatsu Y., Waragai M. and Hashimoto M. (2014) Role of genomics in translational research for Parkinson's disease. Biochem Biophys Res Commun 452: 226-235
Chung S. J., Armasu S. M., Anderson K. J., Biernacka J. M., Lesnick T. G., et al. (2013) Genetic susceptibility loci, environmental exposures, and Parkinson's disease: a case-control study of gene-environment interactions. Parkinsonism Relat Disord 19: 595-599
Sun Y., Christensen J. and Olsen J. (2013) Childhood epilepsy and maternal antibodies to microbial and tissue antigens during pregnancy. Epilepsy Res 107: 61-74
Bakulski K. M., Rozek L. S., Dolinoy D. C., Paulson H. L. and Hu H. (2012) Alzheimer's disease and environmental exposure to lead: the epidemiologic evidence and potential role of epigenetics. Curr Alzheimer Res 9: 563-573
Brown A. S. and Derkits E. J. (2010) Prenatal infection and schizophrenia: a review of epidemiologic and translational studies. Am J Psychiatry 167: 261-280
298. Brown A. S. and Derkits E. J. (2010) Prenatal infection and schizophrenia: a review of epidemiologic and translational studies. Am J Psychiatry 167: 261-280
299. Torrey E. F. and Yolken R. H. (2003) Toxoplas-ma gondii and schizophrenia. Emerg Infect Dis 9: 1375-1380
300. Jones B. C., Miller D. B., O'Callaghan J. P., Lu L., Unger E. L., et al. (2013) Systems analysis of genetic variation in MPTP neurotoxicity in mice. Neurotoxicology 37: 26-34
301. Basha M. R., Wei W., Bakheet S. A., Benitez N., Siddiqi H. K., et al. (2005) The fetal basis of amyloidogenesis: exposure to lead and latent overexpression of amyloid precursor protein and beta-amyloid in the aging brain. J Neurosci 25: 823-829
302. Ryan S. D., Dolatabadi N., Chan S. F., Zhang X., Akhtar M. W., et al. (2013) Isogenic human iPSC Parkinson's model shows nitrosative stress-induced dysfunction in MEF2-PGC1al-pha transcription. Cell 155: 1351-1364
303. Sekiyama K., Takamatsu Y., Waragai M. and Hashimoto M. (2014) Role of genomics in translational research for Parkinson's disease. Biochem Biophys Res Commun 452: 226-235
304. Sekiyama K., Takamatsu Y., Waragai M. and Hashimoto M. (2014) Role of genomics in translational research for Parkinson's disease. Biochem Biophys Res Commun 452: 226-235

50 Neurobiology Research Review
SINGLE CELLS
Each cell type has a distinct lineage and function, which contributes to the functioning
of the tissue, the organ, and—ultimately—the organism. The lineage and developmental
stage of each cell determine how they respond to each other and to their environment.
While the ultimate goal of an exhaustive understanding of tissues at their cellular level is
still elusive, recent progress in single-cell analysis is offering a glimpse at the future.
A recent study of single cells and neurons in brain tissue found that most (≥ 95%)
neurons in normal brain tissue are euploid. However, a patient with hemimegalencephaly
(HMG) due to a somatic CNV of chromosome 1q had unexpected tetrasomy 1q in
20% of neurons. This observation suggests that CNVs in a minority of cells can cause
widespread brain dysfunction.305 This complexity can only be resolved with single-cell
sequencing approaches.
Recent advances in research have highlighted the mosaic genomes of individual neurons,
exhibiting CNVs even among cells that make up a specific region of the brain.306 Even
though genetic variations in the brain arise during fetal development,307 the functional
relevance of this mosaicism is unclear at present. It will be of interest not only to discover
the significance of mosaicism in the normal brain, but also to study its role in neurological
diseases and psychological disorders.308-311
Research has just begun to shed light mosaicism, where heterogeneity among cells is
notable at the genome level. If mosaicism exists in the genetic code among single cells,312
there are likely also variations in protein expression, epigenetic changes,313 and RNA
isoforms.314 Sequencing single cells provides a larger integrated image of the collected
data, helping to account for the effects of mosaicism on individual cellular phenotypes
within a given region of the brain.315,316,317
The high accuracy and specificity of NGS lends itself well to single-cell and low-level
DNA/RNA sequencing. The growing armamentarium of published techniques includes
the detection of DNA mutations, CNVs, DNA-protein binding, RNA splicing, and the
measurement of RNA expression values.
ReviewsChi K. R. (2014) Singled out for sequencing. Nat Methods 11: 13-17
Macaulay I. C. and Voet T. (2014) Single cell genomics: advances and future perspectives. PLoS Genet 10: e1004126
Weaver W. M., Tseng P., Kunze A., Masaeli M., Chung A. J., et al. (2014) Advances in high-throughput single-cell microtechnologies. Curr Opin Biotechnol 25: 114-123
Poduri A., Evrony G. D., Cai X. and Walsh C. A. (2013) Somatic mutation, genomic variation, and neurological disease. Science 341: 1237758
305. Cai X., Evrony G. D., Lehmann H. S., Elhosary P. C., Mehta B. K., et al. (2014) Single-Cell, Genome-wide Sequencing Identifies Clonal Somatic Copy-Number Variation in the Human Brain. Cell Rep 8: 1280-1289
306. McConnell M. J., Lindberg M. R., Brennand K. J., Piper J. C., Voet T., et al. (2013) Mosaic copy number variation in human neurons. Science 342: 632-637
307. Iourov I. Y., Vorsanova S. G. and Yurov Y. B. (2012) Single cell genomics of the brain: focus on neuronal diversity and neuropsychiatric diseases. Curr Genomics 13: 477-488
308. Iourov I. Y., Vorsanova S. G. and Yurov Y. B. (2012) Single cell genomics of the brain: focus on neuronal diversity and neuropsychiatric diseases. Curr Genomics 13: 477-488
309. Evrony G. D., Cai X., Lee E., Hills L. B., Elhosary P. C., et al. (2012) Single-neuron sequencing analysis of L1 retrotransposition and somatic mutation in the human brain. Cell 151: 483-496
310. Poduri A., Evrony G. D., Cai X. and Walsh C. A. (2013) Somatic mutation, genomic variation, and neurological disease. Science 341: 12377
311. Eberwine J. and Bartfai T. (2011) Single cell transcriptomics of hypothalamic warm sensi-tive neurons that control core body tempera-ture and fever response Signaling asymmetry and an extension of chemical neuroanatomy. Pharmacol Ther 129: 241-259
312. Evrony G. D., Cai X., Lee E., Hills L. B., Elhosary P. C., et al. (2012) Single-neuron sequencing analysis of L1 retrotransposition and somatic mutation in the human brain. Cell 151: 483-496
313. Lister R., Mukamel E. A., Nery J. R., Urich M., Puddifoot C. A., et al. (2013) Global epigenom-ic reconfiguration during mammalian brain development. Science 341: 1237905
314. Macaulay I. C. and Voet T. (2014) Single cell genomics: advances and future perspectives. PLoS Genet 10: e1004126
315. Evrony G. D., Cai X., Lee E., Hills L. B., Elhosary P. C., et al. (2012) Single-neuron sequencing analysis of L1 retrotransposition and somatic mutation in the human brain. Cell 151: 483-496
316. Poduri A., Evrony G. D., Cai X. and Walsh C. A. (2013) Somatic mutation, genomic variation, and neurological disease. Science 341: 1237758
317. Eberwine J. and Bartfai T. (2011) Single cell transcriptomics of hypothalamic warm sensi-tive neurons that control core body tempera-ture and fever response Signaling asymmetry and an extension of chemical neuroanatomy. Pharmacol Ther 129: 241-259

51 An Overview of Publications Featuring Illumina® Technology
ReferencesLovatt D., Ruble B. K., Lee J., Dueck H., Kim T. K., et al. (2014) Transcriptome in vivo analysis (TIVA) of spatially defined single cells in live tissue. Nat Methods 11: 190-196RNA sequencing methods that rely on RNA extracted from cell mixtures do not convey the individual variability in expression among cells of the same tissue. In this paper, the authors present a transcriptome in vivo analysis (TIVA), which is applicable to single-cell studies. In combination with Illumina sequencing technology, the authors capture and analyze the transcriptome variance across single neurons both in culture and in vivo. This method is furthermore non-invasive and may be applied to intact tissue. It will enable detailed studies of cell heterogeneity in complex tissues that have been intractable previously, and it opens up the possibility of use in conjunction with in vivo live functional imaging.
Illumina Technology: 670k BeadChip Array
Young G. T., Gutteridge A., Fox H. D., Wilbrey A. L., Cao L., et al. (2014) Characterizing Human Stem Cell-derived Sensory Neurons at the Single-cell Level Reveals Their Ion Channel Expression and Utility in Pain Research. Mol Ther 22: 1530-1543Research into the biology of pain is commonly performed on animal models due to the lack of sensory neuronal cell lines. In this paper, the authors present human stem-cell derived sensory neurons and use a combination of population and single-cell techniques to perform detailed molecular, electrophysiological, and pharmacological phenotyping. The directed differentiation was monitored over 6 weeks and the gene expression characterized using Illumina BeadArrays. The authors show the derived neurons are both molecularly and functionally comparable to human sensory neurons derived from mature dorsal root ganglia.
Illumina Technology: BeadArrays
Lister R., Mukamel E. A., Nery J. R., Urich M., Puddifoot C. A., et al. (2013) Global epigenomic reconfiguration during mammalian brain development. Science 341: 1237905DNA methylation is implicated in mammalian brain development and plasticity underlying learning and memory. This paper reports the genome-wide composition, patterning, cell specificity and dynamics of DNA methylation at single-base resolution in human and mouse frontal cortex throughout their lifespan. The extensive methylome profiling was performed with ChIP-Seq on Illumina HiSeq, revealing methylation profiles at single-base resolution.
Illumina Technology: TruSeq RNA, TruSeq DNA, HiSeq
McConnell M. J., Lindberg M. R., Brennand K. J., Piper J. C., Voet T., et al. (2013) Mosaic copy number variation in human neurons. Science 342: 632-637The genome of a species varies not only between individuals, but also between mother and daughter cells as a result of errors and uneven distribution of DNA material after cell division. The extent of such genetic variation between somatic cells of the same individual has been hitherto unknown. With the advent of single-cell sequencing, it is now possible to examine variations within cells of the same tissue. In this study, the CNVs in individual neuron cells were studied using Illumina genome-wide sequencing. The authors discovered that CNVs are abundant even between neuronal cells from the same tissue.
Illumina Technology: Genome AnalyzerIIx, MiSeq, Nextera DNA Sample Prep Kit
Pan X., Durrett R. E., Zhu H., Tanaka Y., Li Y., et al. (2013) Two methods for full-length RNA sequencing for low quantities of cells and single cells. Proc Natl Acad Sci U S A 110: 594-599Gene expression profiling by RNA-Seq is a powerful tool for understanding the molecular activity of specific tissues. However, the heterogeneity of gene expression within a tissue requires RNA-Seq technology that can manage single-cell amounts of input RNA. This study presents two methods: Phi29 DNA polymerase–based mRNA transcriptome amplification (PMA), and semi–random-primed PCR-based mRNA transcriptome amplification (SMA). Both techniques are coupled with Illumina sequencing for expression detection from low RNA input amounts. Both protocols produced satisfactory detection/coverage of abundant mRNAs, even from a single cell.
Illumina Technology: HiSeq
Binder V., Bartenhagen C., Okpanyi V., Gombert M., Moehlendick B., et al. (2014) A New Workflow for Whole-Genome Sequencing of Single Human Cells. Hum Mutat 35: 1260-1270
Cai X., Evrony G. D., Lehmann H. S., Elhosary P. C., Mehta B. K., et al. (2014) Single-Cell, Genome-wide Sequencing Identifies Clonal Somatic Copy-Number Variation in the Human Brain. Cell Rep 8: 1280-1289

52 Neurobiology Research Review
Abnet C. C., Wang Z., Song X., Hu N., Zhou F. Y., et al. (2012) Genotypic variants at 2q33 and risk of esophageal squamous cell carcinoma in China: a meta-analysis of genome-wide association studies. Hum Mol Genet 21: 2132-2141
Aguzzi A. and Rajendran L. (2009) The transcellular spread of cytosolic amyloids, prions, and prionoids. Neuron 64: 783-790
Akula N., Barb J., Jiang X., Wendland J. R., Choi K. H., et al. (2014) RNA-sequencing of the brain transcriptome implicates dysregulation of neuroplasticity, circadian rhythms and GTPase binding in bipolar disorder. Mol Psychiatry
Anthes E. (2014) Ageing: Live faster, die younger. Nature 508: S16-17
Baba M., Nakajo S., Tu P. H., Tomita T., Nakaya K., et al. (1998) Aggregation of α-synuclein in Lewy bodies of sporadic Parkinson's disease and dementia with Lewy bodies. Am J Pathol 152: 879-884
Bai B., Hales C. M., Chen P. C., Gozal Y., Dammer E. B., et al. (2013) U1 small nuclear ribonucleoprotein complex and RNA splicing alterations in Alzheimer's disease. Proc Natl Acad Sci U S A 110: 16562-16567
Bakulski K. M., Rozek L. S., Dolinoy D. C., Paulson H. L. and Hu H. (2012) Alzheimer's disease and environmental exposure to lead: the epidemiologic evidence and potential role of epigenetics. Curr Alzheimer Res 9: 563-573
Banerjee S., Riordan M. and Bhat M. A. (2014) Genetic aspects of autism spectrum disorders: insights from animal models. Front Cell Neurosci 8: 58
Barwick B. G., Abramovitz M., Kodani M., Moreno C. S., Nam R., et al. (2010) Prostate cancer genes associated with TMPRSS2-ERG gene fusion and prognostic of biochemical recurrence in multiple cohorts. Br J Cancer 102: 570-576
Basha M. R., Wei W., Bakheet S. A., Benitez N., Siddiqi H. K., et al. (2005) The fetal basis of amyloidogenesis: exposure to lead and latent overexpression of amyloid precursor protein and beta-amyloid in the aging brain. J Neurosci 25: 823-829
Beri S., Tonna N., Menozzi G., Bonaglia M. C., Sala C., et al. (2007) DNA methylation regulates tissue-specific expression of Shank3. J Neurochem 101: 1380-1391
Bertram L., Lill C. M. and Tanzi R. E. (2010) The genetics of Alzheimer disease: back to the future. Neuron 68: 270-281
Bien-Ly N., Andrews-Zwilling Y., Xu Q., Bernardo A., Wang C., et al. (2011) C-terminal-truncated apolipoprotein (apo) E4 inefficiently clears amyloid-beta (Abeta) and acts in concert with Abeta to elicit neuronal and behavioral deficits in mice. Proc Natl Acad Sci U S A 108: 4236-4241
Binder V., Bartenhagen C., Okpanyi V., Gombert M., Moehlendick B., et al. (2014) A New Workflow for Whole-Genome Sequencing of Single Human Cells. Hum Mutat 35: 1260-1270
Blandini F. and Armentero M. T. (2012) Animal models of Parkinson's disease. FEBS J 279: 1156-1166
Blurton-Jones M. and Laferla F. M. (2006) Pathways by which Abeta facilitates tau pathology. Curr Alzheimer Res 3: 437-448
Booij B. B., Lindahl T., Wetterberg P., Skaane N. V., Saebo S., et al. (2011) A gene expression pattern in blood for the early detection of Alzheimer's disease. J Alzheimers Dis 23: 109-119
Borglum A. D., Demontis D., Grove J., Pallesen J., Hollegaard M. V., et al. (2014) Genome-wide study of association and interaction with maternal cytomegalovirus infection suggests new schizophrenia loci. Mol Psychiatry 19: 325-333
Boulanger L. M. (2009) Immune proteins in brain development and synaptic plasticity. Neuron 64: 93-109
Bras J., Guerreiro R. and Hardy J. (2012) Use of next-generation sequencing and other whole-genome strategies to dissect neurological disease. Nat Rev Neurosci 13: 453-464
Bras J., Guerreiro R., Darwent L., Parkkinen L., Ansorge O., et al. (2014) Genetic analysis implicates APOE, SNCA and suggests lysosomal dysfunction in the etiology of dementia with Lewy bodies. Hum Mol Genet. In Press
Brody H. (2014) Schizophrenia. Nature 508: S1
Brookmeyer R., Johnson E., Ziegler-Graham K. and Arrighi H. M. (2007) Forecasting the global burden of Alzheimer's disease. Alzheimers Dement 3: 186-191
Brown A. S. and Derkits E. J. (2010) Prenatal infection and schizophrenia: a review of epidemiologic and translational studies. Am J Psychiatry 167: 261-280
Buxbaum J. (2013) The Neuroscience of Autism Spectrum Disorders.
Cai X., Evrony G. D., Lehmann H. S., Elhosary P. C., Mehta B. K., et al. (2014) Single-Cell, Genome-wide Sequencing Identifies Clonal Somatic Copy-Number Variation in the Human Brain. Cell Rep 8: 1280-1289
Carter H. B., Morrell C. H., Pearson J. D., Brant L. J., Plato C. C., et al. (1992) Estimation of prostatic growth using serial prostate-specific antigen measurements in men with and without prostate disease. Cancer Res 52: 3323-3328
Carter H. B., Pearson J. D., Metter E. J., Brant L. J., Chan D. W., et al. (1992) Longitudinal evaluation of prostate-specific antigen levels in men with and without prostate disease. JAMA 267: 2215-2220
Chapman J., Rees E., Harold D., Ivanov D., Gerrish A., et al. (2013) A genome-wide study shows a limited contribution of rare copy number variants to Alzheimer's disease risk. Hum Mol Genet 22: 816-824
Chen-Plotkin A. S., Hu W. T., Siderowf A., Weintraub D., Goldmann Gross R., et al. (2011) Plasma epidermal growth factor levels predict cognitive decline in Parkinson disease. Ann Neurol 69: 655-663
Chesselet M. F. (2008) In vivo alpha-synuclein overexpression in rodents: a useful model of Parkinson's disease? Exp Neurol 209: 22-27
Chi K. R. (2014) Singled out for sequencing. Nat Methods 11: 13-17
Ching T. T., Maunakea A. K., Jun P., Hong C., Zardo G., et al. (2005) Epigenome analyses using BAC microarrays identify evolutionary conservation of tissue-specific methylation of SHANK3. Nat Genet 37: 645-651
Chouliaras L., Rutten B. P., Kenis G., Peerbooms O., Visser P. J., et al. (2010) Epigenetic regulation in the pathophysiology of Alzheimer's disease. Prog Neurobiol 90: 498-510
Chung S. J., Armasu S. M., Anderson K. J., Biernacka J. M., Lesnick T. G., et al. (2013) Genetic susceptibility loci, environmental exposures, and Parkinson's disease: a case-control study of gene-environment interactions. Parkinsonism Relat Disord 19: 595-599
Clayton D. F. and George J. M. (1998) The synucleins: a family of proteins involved in synaptic function, plasticity, neurodegeneration and disease. Trends Neurosci 21: 249-254
Clayton D. F., Balakrishnan C. N. and London S. E. (2009) Integrating genomes, brain and behavior in the study of songbirds. Curr Biol 19: R865-873
Cookson M. R. (2005) The biochemistry of Parkinson's disease. Annu Rev Biochem 74: 29-52
Cooper A. A., Gitler A. D., Cashikar A., Haynes C. M., Hill K. J., et al. (2006) Alpha-synuclein blocks ER-Golgi traffic and Rab1 rescues neuron loss in Parkinson's models. Science 313: 324-328
BIBLIOGRAPHY

53 An Overview of Publications Featuring Illumina® Technology
Coppede F. (2014) The potential of epigenetic therapies in neurodegenerative diseases. Front Genet 5: 220
Crabtree D. M. and Zhang J. (2012) Genetically engineered mouse models of Parkinson's disease. Brain Res Bull 88: 13-32
Cross-Disorder Group of the Psychiatric Genomics C., Lee S. H., Ripke S., Neale B. M., Faraone S. V., et al. (2013) Genetic relationship between five psychiatric disorders estimated from genome-wide SNPs. Nat Genet 45: 984-994
Cruchaga C., Kauwe J. S., Harari O., Jin S. C., Cai Y., et al. (2013) GWAS of cerebrospinal fluid tau levels identifies risk variants for Alzheimer's disease. Neuron 78: 256-268
Cruchaga C., Karch C. M., Jin S. C., Benitez B. A., Cai Y., et al. (2014) Rare coding variants in the phospholipase D3 gene confer risk for Alzheimer's disease. Nature 505: 550-554
Cukier H. N., Dueker N. D., Slifer S. H., Lee J. M., Whitehead P. L., et al. (2014) Exome sequencing of extended families with autism reveals genes shared across neurodevelopmental and neuropsychiatric disorders. Mol Autism 5: 1
Curran L. K., Newschaffer C. J., Lee L. C., Crawford S. O., Johnston M. V., et al. (2007) Behaviors associated with fever in children with autism spectrum disorders. Pediatrics 120: e1386-1392
De Jager P. L., Srivastava G., Lunnon K., Burgess J., Schalkwyk L. C., et al. (2014) Alzheimer's disease: early alterations in brain DNA methylation at ANK1, BIN1, RHBDF2 and other loci. Nat Neurosci 17: 1156-1163
de Lau L. M. and Breteler M. M. (2006) Epidemiology of Parkinson's disease. Lancet Neurol 5: 525-535
de Leon J. and Diaz F. J. (2005) A meta-analysis of worldwide studies demonstrates an association between schizophrenia and tobacco smoking behaviors. Schizophr Res 76: 135-157
De Strooper B. and Voet T. (2012) Alzheimer's disease: A protective mutation. Nature 488: 38-39
Deas E., Plun-Favreau H. and Wood N. W. (2009) PINK1 function in health and disease. EMBO Mol Med 1: 152-165
Deleidi M. and Isacson O. (2012) Viral and inflammatory triggers of neurodegenerative diseases. Sci Transl Med 4: 121ps123
Dinan T. G., Stanton C. and Cryan J. F. (2013) Psychobiotics: a novel class of psychotropic. Biol Psychiatry 74: 720-726
Ding H., Dolan P. J. and Johnson G. V. (2008) Histone deacetylase 6 interacts with the microtubule-associated protein tau. J Neurochem 106: 2119-2130
n L. (1999) Dual diagnosis of substance abuse in schizophrenia: prevalence and impact on outcomes. Schizophr Res 35 Suppl: S93-100
Dolgin E. (2014) Therapeutics: Negative feedback. Nature 508: S10-11
Durkin M. S., Maenner M. J., Newschaffer C. J., Lee L. C., Cunniff C. M., et al. (2008) Advanced parental age and the risk of autism spectrum disorder. Am J Epidemiol 168: 1268-1276
Dusonchet J., Kochubey O., Stafa K., Young S. M., Jr., Zufferey R., et al. (2011) A rat model of progressive nigral neurodegeneration induced by the Parkinson's disease-associated G2019S mutation in LRRK2. J Neurosci 31: 907-912
Ebert D. H. and Greenberg M. E. (2013) Activity-dependent neuronal signalling and autism spectrum disorder. Nature 493: 327-337
Eberwine J. and Bartfai T. (2011) Single cell transcriptomics of hypothalamic warm sensitive neurons that control core body temperature and fever response Signaling asymmetry and an extension of chemical neuroanatomy. Pharmacol Ther 129: 241-259
Eisenberg D. and Jucker M. (2012) The amyloid state of proteins in human diseases. Cell 148: 1188-1203
Elert E. (2014) Aetiology: Searching for schizophrenia's roots. Nature 508: S2-3
Emre M., Aarsland D., Brown R., Burn D. J., Duyckaerts C., et al. (2007) Clinical diagnostic criteria for dementia associated with Parkinson's disease. Mov Disord 22: 1689-1707; quiz 1837
Evrony G. D., Cai X., Lee E., Hills L. B., Elhosary P. C., et al. (2012) Single-neuron sequencing analysis of L1 retrotransposition and somatic mutation in the human brain. Cell 151: 483-496
Fan H. C., Chen S. J., Harn H. J. and Lin S. Z. (2013) Parkinson's disease: from genetics to treatments. Cell Transplant 22: 639-652
Fehlbaum-Beurdeley P., Sol O., Desire L., Touchon J., Dantoine T., et al. (2012) Validation of AclarusDx, a blood-based transcriptomic signature for the diagnosis of Alzheimer's disease. J Alzheimers Dis 32: 169-181
Fischer A., Sananbenesi F., Wang X., Dobbin M. and Tsai L. H. (2007) Recovery of learning and memory is associated with chromatin remodelling. Nature 447: 178-182
Fogel B. L., Cho E., Wahnich A., Gao F., Becherel O. J., et al. (2014) Mutation of senataxin alters disease-specific transcriptional networks in patients with ataxia with oculomotor apraxia type 2. Hum Mol Genet 23: 4758-4769
Forshew T., Murtaza M., Parkinson C., Gale D., Tsui D. W., et al. (2012) Noninvasive identification and monitoring of cancer mutations by targeted deep sequencing of plasma DNA. Sci Transl Med 4: 136ra168
Fromer M., Pocklington A. J., Kavanagh D. H., Williams H. J., Dwyer S., et al. (2014) De novo mutations in schizophrenia implicate synaptic networks. Nature 506: 179-184
Gadad B. S., Hewitson L., Young K. A. and German D. C. (2013) Neuropathology and animal models of autism: genetic and environmental factors. Autism Res Treat 2013: 731935
Gadow K. D. (2013) Association of schizophrenia spectrum and autism spectrum disorder (ASD) symptoms in children with ASD and clinic controls. Res Dev Disabil 34: 1289-1299
Ganat Y. M., Calder E. L., Kriks S., Nelander J., Tu E. Y., et al. (2012) Identification of embryonic stem cell-derived midbrain dopaminergic neurons for engraftment. J Clin Invest 122: 2928-2939
Gau S. S., Liao H. M., Hong C. C., Chien W. H. and Chen C. H. (2012) Identification of two inherited copy number variants in a male with autism supports two-hit and compound heterozygosity models of autism. Am J Med Genet B Neuropsychiatr Genet 159B: 710-717
Gaugler T., Klei L., Sanders S. J., Bodea C. A., Goldberg A. P., et al. (2014) Most genetic risk for autism resides with common variation. Nat Genet 46: 881-885
Gauthier J., Champagne N., Lafreniere R. G., Xiong L., Spiegelman D., et al. (2010) De novo mutations in the gene encoding the synaptic scaffolding protein SHANK3 in patients ascertained for schizophrenia. Proc Natl Acad Sci U S A 107: 7863-7868
Genin E., Hannequin D., Wallon D., Sleegers K., Hiltunen M., et al. (2011) APOE and Alzheimer disease: a major gene with semi-dominant inheritance. Mol Psychiatry 16: 903-907
Ghosh A., Michalon A., Lindemann L., Fontoura P. and Santarelli L. (2013) Drug discovery for autism spectrum disorder: challenges and opportunities. Nat Rev Drug Discov 12: 777-790
Gilbert J. A., Krajmalnik-Brown R., Porazinska D. L., Weiss S. J. and Knight R. (2013) Toward effective probiotics for autism and other neurodevelopmental disorders. Cell 155: 1446-1448
Gilissen C., Hehir-Kwa J. Y., Thung D. T., van de Vorst M., van Bon B. W., et al. (2014) Genome sequencing identifies major causes of severe intellectual disability. Nature 511: 344-347
Gilman S. R., Iossifov I., Levy D., Ronemus M., Wigler M., et al. (2011) Rare de novo variants associated with autism implicate a large functional network of genes involved in formation and function of synapses. Neuron 70: 898-907
Girgis R. R., Kumar S. S. and Brown A. S. (2014) The cytokine model of schizophrenia: emerging therapeutic strategies. Biol Psychiatry 75: 292-299

54 Neurobiology Research Review
Glessner J. T., Wang K., Cai G., Korvatska O., Kim C. E., et al. (2009) Autism genome-wide copy number variation reveals ubiquitin and neuronal genes. Nature 459: 569-573
Gomez-Herreros F., Schuurs-Hoeijmakers J. H., McCormack M., Greally M. T., Rulten S., et al. (2014) TDP2 protects transcription from abortive topoisomerase activity and is required for normal neural function. Nat Genet 46: 516-521
Graff J., Rei D., Guan J. S., Wang W. Y., Seo J., et al. (2012) An epigenetic blockade of cognitive functions in the neurodegenerating brain. Nature 483: 222-226
Group N. H. W., Peterson J., Garges S., Giovanni M., McInnes P., et al. (2009) The NIH Human Microbiome Project. Genome Res 19: 2317-2323
Guerreiro R. J., Gustafson D. R. and Hardy J. (2012) The genetic architecture of Alzheimer's disease: beyond APP, PSENs and APOE. Neurobiol Aging 33: 437-456
Guffanti G., Torri F., Rasmussen J., Clark A. P., Lakatos A., et al. (2013) Increased CNV-region deletions in mild cognitive impairment (MCI) and Alzheimer's disease (AD) subjects in the ADNI sample. Genomics 102: 112-122
Guilmatre A., Dubourg C., Mosca A. L., Legallic S., Goldenberg A., et al. (2009) Recurrent rearrangements in synaptic and neurodevelopmental genes and shared biologic pathways in schizophrenia, autism, and mental retardation. Arch Gen Psychiatry 66: 947-956
Guo J. L. and Lee V. M. (2014) Cell-to-cell transmission of pathogenic proteins in neurodegenerative diseases. Nat Med 20: 130-138
Hafner M., Landgraf P., Ludwig J., Rice A., Ojo T., et al. (2008) Identification of microRNAs and other small regulatory RNAs using cDNA library sequencing. Methods 44: 3-12
Harold D., Abraham R., Hollingworth P., Sims R., Gerrish A., et al. (2009) Genome-wide association study identifies variants at CLU and PICALM associated with Alzheimer's disease. Nat Genet 41: 1088-1093
Hollerhage M., Goebel J. N., de Andrade A., Hildebrandt T., Dolga A., et al. (2014) Trifluoperazine rescues human dopaminergic cells from wild-type alpha-synuclein-induced toxicity. Neurobiol Aging 35: 1700-1711
Hollingworth P., Harold D., Sims R., Gerrish A., Lambert J. C., et al. (2011) Common variants at ABCA7, MS4A6A/MS4A4E, EPHA1, CD33 and CD2AP are associated with Alzheimer's disease. Nat Genet 43: 429-435
Horvath S. and Mirnics K. (2014) Immune system disturbances in schizophrenia. Biol Psychiatry 75: 316-323
Hsiao E. Y., McBride S. W., Hsien S., Sharon G., Hyde E. R., et al. (2013) Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell 155: 1451-1463
Huang Y. (2010) Abeta-independent roles of apolipoprotein E4 in the pathogenesis of Alzheimer's disease. Trends Mol Med 16: 287-294
Huang Y. and Mucke L. (2012) Alzheimer mechanisms and therapeutic strategies. Cell 148: 1204-1222
Hyman S. E. (2014) Perspective: Revealing molecular secrets. Nature 508: S20
Iafrate A. J., Feuk L., Rivera M. N., Listewnik M. L., Donahoe P. K., et al. (2004) Detection of large-scale variation in the human genome. Nat Genet 36: 949-951
Ionita-Laza I., Xu B., Makarov V., Buxbaum J. D., Roos J. L., et al. (2014) Scan statistic-based analysis of exome sequencing data identifies FAN1 at 15q13.3 as a susceptibility gene for schizophrenia and autism. Proc Natl Acad Sci U S A 111: 343-348
Iourov I. Y., Vorsanova S. G. and Yurov Y. B. (2012) Single cell genomics of the brain: focus on neuronal diversity and neuropsychiatric diseases. Curr Genomics 13: 477-488
Irwin D. J., Lee V. M. and Trojanowski J. Q. (2013) Parkinson's disease dementia: convergence of alpha-synuclein, tau and amyloid-beta pathologies. Nat Rev Neurosci 14: 626-636
Jacquemont S., Coe B. P., Hersch M., Duyzend M. H., Krumm N., et al. (2014) A higher mutational burden in females supports a "female protective model" in neurodevelopmental disorders. Am J Hum Genet 94: 415-425
Jiang Y. H. and Ehlers M. D. (2013) Modeling autism by SHANK gene mutations in mice. Neuron 78: 8-27
Jiang Y. H., Yuen R. K., Jin X., Wang M., Chen N., et al. (2013) Detection of clinically relevant genetic variants in autism spectrum disorder by whole-genome sequencing. Am J Hum Genet 93: 249-263
Jones B. C., Miller D. B., O'Callaghan J. P., Lu L., Unger E. L., et al. (2013) Systems analysis of genetic variation in MPTP neurotoxicity in mice. Neurotoxicology 37: 26-34
Jones L., Harold D. and Williams J. (2010) Genetic evidence for the involvement of lipid metabolism in Alzheimer's disease. Biochim Biophys Acta 1801: 754-761
Jonsson T., Atwal J. K., Steinberg S., Snaedal J., Jonsson P. V., et al. (2012) A mutation in APP protects against Alzheimer's disease and age-related cognitive decline. Nature 488: 96-99
Jowaed A., Schmitt I., Kaut O. and Wullner U. (2010) Methylation regulates alpha-synuclein expression and is decreased in Parkinson's disease patients' brains. J Neurosci 30: 6355-6359
Jucker M. (2010) The benefits and limitations of animal models for translational research in neurodegenerative diseases. Nat Med 16: 1210-1214
Jucker M. and Walker L. C. (2013) Self-propagation of pathogenic protein aggregates in neurodegenerative diseases. Nature 501: 45-51
Jung Y. W., Hysolli E., Kim K. Y., Tanaka Y. and Park I. H. (2012) Human induced pluripotent stem cells and neurodegenerative disease: prospects for novel therapies. Curr Opin Neurol 25: 125-130
Karayannis T., Au E., Patel J. C., Kruglikov I., Markx S., et al. (2014) Cntnap4 differentially contributes to GABAergic and dopaminergic synaptic transmission. Nature 511: 236-240
Karlsson M. K., Sharma P., Aasly J., Toft M., Skogar O., et al. (2013) Found in transcription: accurate Parkinson's disease classification in peripheral blood. J Parkinsons Dis 3: 19-29
Katsnelson A. (2014) Drug development: The modelling challenge. Nature 508: S8-9
Keogh M. J. and Chinnery P. F. (2013) Next generation sequencing for neurological diseases: new hope or new hype? Clin Neurol Neurosurg 115: 948-953
Kilgore M., Miller C. A., Fass D. M., Hennig K. M., Haggarty S. J., et al. (2010) Inhibitors of class 1 histone deacetylases reverse contextual memory deficits in a mouse model of Alzheimer's disease. Neuropsychopharmacology 35: 870-880
Kinde I., Bettegowda C., Wang Y., Wu J., Agrawal N., et al. (2013) Evaluation of DNA from the Papanicolaou test to detect ovarian and endometrial cancers. Sci Transl Med 5: 167ra164
King C. and Murphy G. H. (2014) A Systematic Review of People with Autism Spectrum Disorder and the Criminal Justice System. J Autism Dev Disord
King I. F., Yandava C. N., Mabb A. M., Hsiao J. S., Huang H. S., et al. (2013) Topoisomerases facilitate transcription of long genes linked to autism. Nature 501: 58-62
Kirik D., Rosenblad C., Burger C., Lundberg C., Johansen T. E., et al. (2002) Parkinson-like neurodegeneration induced by targeted overexpression of alpha-synuclein in the nigrostriatal system. J Neurosci 22: 2780-2791
Kirov G., Rees E., Walters J. T., Escott-Price V., Georgieva L., et al. (2014) The penetrance of copy number variations for schizophrenia and developmental delay. Biol Psychiatry 75: 378-385

55 An Overview of Publications Featuring Illumina® Technology
Koboldt D. C., Steinberg K. M., Larson D. E., Wilson R. K. and Mardis E. R. (2013) The next-generation sequencing revolution and its impact on genomics. Cell 155: 27-38
Kong A., Frigge M. L., Masson G., Besenbacher S., Sulem P., et al. (2012) Rate of de novo mutations and the importance of father's age to disease risk. Nature 488: 471-475
Kriukiene E., Labrie V., Khare T., Urbanaviciute G., Lapinaite A., et al. (2013) DNA unmethylome profiling by covalent capture of CpG sites. Nat Commun 4: 2190
Kvajo M., McKellar H. and Gogos J. A. (2012) Avoiding mouse traps in schizophrenia genetics: lessons and promises from current and emerging mouse models. Neuroscience 211: 136-164
Ladd-Acosta C., Hansen K. D., Briem E., Fallin M. D., Kaufmann W. E., et al. (2013) Common DNA methylation alterations in multiple brain regions in autism. Mol Psychiatry
Ladd-Acosta C., Hansen K. D., Briem E., Fallin M. D., Kaufmann W. E., et al. (2014) Common DNA methylation alterations in multiple brain regions in autism. Mol Psychiatry 19: 862-871
Lambert J. C., Heath S., Even G., Campion D., Sleegers K., et al. (2009) Genome-wide association study identifies variants at CLU and CR1 associated with Alzheimer's disease. Nat Genet 41: 1094-1099
Lambert J. C., Ibrahim-Verbaas C. A., Harold D., Naj A. C., Sims R., et al. (2013) Meta-analysis of 74,046 individuals identifies 11 new susceptibility loci for Alzheimer's disease. Nat Genet 45: 1452-1458
Lamendella R., VerBerkmoes N. and Jansson J. K. (2012) 'Omics' of the mammalian gut--new insights into function. Curr Opin Biotechnol 23: 491-500
Lasken R. S. (2013) Single-cell sequencing in its prime. Nat Biotechnol 31: 211-212
Lee H., Lin M. C., Kornblum H. I., Papazian D. M. and Nelson S. F. (2014) Exome sequencing identifies de novo gain of function missense mutation in KCND2 in identical twins with autism and seizures that slows potassium channel inactivation. Hum Mol Genet 23: 3481-3489
Lelan F., Boyer C., Thinard R., Remy S., Usal C., et al. (2011) Effects of Human Alpha-synuclein A53T-A30P Mutations on SVZ and Local Olfactory Bulb Cell Proliferation in a Transgenic Rat Model of Parkinson Disease. Parkinsons Dis 2011: 987084
Levy D., Ronemus M., Yamrom B., Lee Y. H., Leotta A., et al. (2011) Rare de novo and transmitted copy-number variation in autistic spectrum disorders. Neuron 70: 886-897
Lewis J., Dickson D. W., Lin W. L., Chisholm L., Corral A., et al. (2001) Enhanced neurofibrillary degeneration in transgenic mice expressing mutant tau and APP. Science 293: 1487-1491
Li M. M., Jiang T., Sun Z., Zhang Q., Tan C. C., et al. (2014) Genome-wide microRNA expression profiles in hippocampus of rats with chronic temporal lobe epilepsy. Sci Rep 4: 4734
Lindqvist D., Hall S., Surova Y., Nielsen H. M., Janelidze S., et al. (2013) Cerebrospinal fluid inflammatory markers in Parkinson's disease--associations with depression, fatigue, and cognitive impairment. Brain Behav Immun 33: 183-189
Lionel A. C., Tammimies K., Vaags A. K., Rosenfeld J. A., Ahn J. W., et al. (2014) Disruption of the ASTN2/TRIM32 locus at 9q33.1 is a risk factor in males for autism spectrum disorders, ADHD and other neurodevelopmental phenotypes. Hum Mol Genet 23: 2752-2768
Lipska B. K. and Weinberger D. R. (2000) To model a psychiatric disorder in animals: schizophrenia as a reality test. Neuropsychopharmacology 23: 223-239
Lister R., Mukamel E. A., Nery J. R., Urich M., Puddifoot C. A., et al. (2013) Global epigenomic reconfiguration during mammalian brain development. Science 341: 1237905
Lo Y. M., Chan K. C., Sun H., Chen E. Z., Jiang P., et al. (2010) Maternal plasma DNA sequencing reveals the genome-wide genetic and mutational profile of the fetus. Sci Transl Med 2: 61ra91
Losh M., Childress D., Lam K. and Piven J. (2008) Defining key features of the broad autism phenotype: a comparison across parents of multiple- and single-incidence autism families. Am J Med Genet B Neuropsychiatr Genet 147B: 424-433
Lovatt D., Ruble B. K., Lee J., Dueck H., Kim T. K., et al. (2014) Transcriptome in vivo analysis (TIVA) of spatially defined single cells in live tissue. Nat Methods 11: 190-196
Lu H., Liu X., Deng Y. and Qing H. (2013) DNA methylation, a hand behind neurodegenerative diseases. Front Aging Neurosci 5: 85
Lunnon K., Smith R., Hannon E., De Jager P. L., Srivastava G., et al. (2014) Methylomic profiling implicates cortical deregulation of ANK1 in Alzheimer's disease. Nat Neurosci 17: 1164-1170
Lynch M. A. and Mills K. H. (2012) Immunology meets neuroscience--opportunities for immune intervention in neurodegenerative diseases. Brain Behav Immun 26: 1-10
Lyons M. R. and West A. E. (2011) Mechanisms of specificity in neuronal activity-regulated gene transcription. Prog Neurobiol 94: 259-295
Macaulay I. C. and Voet T. (2014) Single cell genomics: advances and future perspectives. PLoS Genet 10: e1004126
Maciotta S., Meregalli M. and Torrente Y. (2013) The involvement of microRNAs in neurodegenerative diseases. Front Cell Neurosci 7: 265
Malhotra D., McCarthy S., Michaelson J. J., Vacic V., Burdick K. E., et al. (2011) High frequencies of de novo CNVs in bipolar disorder and schizophrenia. Neuron 72: 951-963
Malhotra D. and Sebat J. (2012) CNVs: harbingers of a rare variant revolution in psychiatric genetics. Cell 148: 1223-1241
Malkki H. (2014) Neurodevelopmental disorders: human gut microbiota alleviate behavioural symptoms in a mouse model of autism spectrum disorder. Nat Rev Neurol 10: 60
Marchetto M. C., Carromeu C., Acab A., Yu D., Yeo G. W., et al. (2010) A model for neural development and treatment of Rett syndrome using human induced pluripotent stem cells. Cell 143: 527-539
Markle J. G., Frank D. N., Mortin-Toth S., Robertson C. E., Feazel L. M., et al. (2013) Sex differences in the gut microbiome drive hormone-dependent regulation of autoimmunity. Science 339: 1084-1088
Martin C. L., Duvall J. A., Ilkin Y., Simon J. S., Arreaza M. G., et al. (2007) Cytogenetic and molecular characterization of A2BP1/FOX1 as a candidate gene for autism. Am J Med Genet B Neuropsychiatr Genet 144B: 869-876
Matsumoto L., Takuma H., Tamaoka A., Kurisaki H., Date H., et al. (2010) CpG demethylation enhances alpha-synuclein expression and affects the pathogenesis of Parkinson's disease. PLoS One 5: e15522
Maunakea A. K., Nagarajan R. P., Bilenky M., Ballinger T. J., D'Souza C., et al. (2010) Conserved role of intragenic DNA methylation in regulating alternative promoters. Nature 466: 253-257
Maurice C. F., Haiser H. J. and Turnbaugh P. J. (2013) Xenobiotics shape the physiology and gene expression of the active human gut microbiome. Cell 152: 39-50
McCarroll S. A. and Hyman S. E. (2013) Progress in the genetics of polygenic brain disorders: significant new challenges for neurobiology. Neuron 80: 578-587
McCarthy S. E., Gillis J., Kramer M., Lihm J., Yoon S., et al. (2014) De novo mutations in schizophrenia implicate chromatin remodeling and support a genetic overlap with autism and intellectual disability. Mol Psychiatry 19: 652-658
McConnell M. J., Lindberg M. R., Brennand K. J., Piper J. C., Voet T., et al. (2013) Mosaic copy number variation in human neurons. Science 342: 632-637
McGrath L. M., Yu D., Marshall C., Davis L. K., Thiruvahindrapuram B., et al. (2014) Copy number variation in obsessive-compulsive disorder and tourette syndrome: a cross-disorder study. J Am Acad Child Adolesc Psychiatry 53: 910-919

56 Neurobiology Research Review
Medland S. E., Jahanshad N., Neale B. M. and Thompson P. M. (2014) Whole-genome analyses of whole-brain data: working within an expanded search space. Nat Neurosci 17: 791-800
Mencacci N. E., Isaias I. U., Reich M. M., Ganos C., Plagnol V., et al. (2014) Parkinson's disease in GTP cyclohydrolase 1 mutation carriers. Brain 137: 2480-2492
Meyer U. (2011) Anti-inflammatory signaling in schizophrenia. Brain Behav Immun 25: 1507-1518
Miles J. H. (2011) Autism spectrum disorders--a genetics review. Genet Med 13: 278-294
Mills J. D. and Janitz M. (2012) Alternative splicing of mRNA in the molecular pathology of neurodegenerative diseases. Neurobiol Aging 33: 1012 e1011-1024
Mitsui J. and Tsuji S. (2014) Genomic Aspects of Sporadic Neurodegenerative Diseases. Biochem Biophys Res Commun 452: 221-225
Moon C. and Stappenbeck T. S. (2012) Viral interactions with the host and microbiota in the intestine. Curr Opin Immunol 24: 405-410
Morihara T., Hayashi N., Yokokoji M., Akatsu H., Silverman M. A., et al. (2014) Transcriptome analysis of distinct mouse strains reveals kinesin light chain-1 splicing as an amyloid-beta accumulation modifier. Proc Natl Acad Sci U S A 111: 2638-2643
Morris D. W., Pearson R. D., Cormican P., Kenny E. M., O'Dushlaine C. T., et al. (2014) An inherited duplication at the gene p21 Protein-Activated Kinase 7 (PAK7) is a risk factor for psychosis. Hum Mol Genet 23: 3316-3326
Mutez E., Nkiliza A., Belarbi K., de Broucker A., Vanbesien-Mailliot C., et al. (2014) Involvement of the immune system, endocytosis and EIF2 signaling in both genetically determined and sporadic forms of Parkinson's disease. Neurobiol Dis 63: 165-170
Naj A. C., Jun G., Beecham G. W., Wang L. S., Vardarajan B. N., et al. (2011) Common variants at MS4A4/MS4A6E, CD2AP, CD33 and EPHA1 are associated with late-onset Alzheimer's disease. Nat Genet 43: 436-441
Nalls M. A., Pankratz N., Lill C. M., Do C. B., Hernandez D. G., et al. (2014) Large-scale meta-analysis of genome-wide association data identifies six new risk loci for Parkinson's disease. Nat Genet 46: 989-993
Nalls M. A., Pankratz N., Lill C. M., Do C. B., Hernandez D. G., et al. (2014) Large-scale meta-analysis of genome-wide association data identifies six new risk loci for Parkinson's disease. Nat Genet 46: 989-993
Nava C., Keren B., Mignot C., Rastetter A., Chantot-Bastaraud S., et al. (2014) Prospective diagnostic analysis of copy number variants using SNP microarrays in individuals with autism spectrum disorders. Eur J Hum Genet 22: 71-78
Ng E., McGirr A., Wong A. H. and Roder J. C. (2013) Using rodents to model schizophrenia and substance use comorbidity. Neurosci Biobehav Rev 37: 896-910
Nicholson J. K., Holmes E., Kinross J., Burcelin R., Gibson G., et al. (2012) Host-gut microbiota metabolic interactions. Science 336: 1262-1267
Nomura J. and Takumi T. (2012) Animal models of psychiatric disorders that reflect human copy number variation. Neural Plast 2012: 589524
Norris A. D., Gao S., Norris M. L., Ray D., Ramani A. K., et al. (2014) A pair of RNA-binding proteins controls networks of splicing events contributing to specialization of neural cell types. Mol Cell 54: 946-959
Oddo S., Caccamo A., Shepherd J. D., Murphy M. P., Golde T. E., et al. (2003) Triple-transgenic model of Alzheimer's disease with plaques and tangles: intracellular Abeta and synaptic dysfunction. Neuron 39: 409-421
Okamoto N., Kubota T., Nakamura Y., Murakami R., Nishikubo T., et al. (2007) 22q13 Microduplication in two patients with common clinical manifestations: a recognizable syndrome? Am J Med Genet A 143A: 2804-2809
Oldmeadow C., Mossman D., Evans T. J., Holliday E. G., Tooney P. A., et al. (2014) Combined analysis of exon splicing and genome wide polymorphism data predict schizophrenia risk loci. J Psychiatr Res 52: 44-49
Onore C., Careaga M. and Ashwood P. (2012) The role of immune dysfunction in the pathophysiology of autism. Brain Behav Immun 26: 383-392
Pan X., Durrett R. E., Zhu H., Tanaka Y., Li Y., et al. (2013) Two methods for full-length RNA sequencing for low quantities of cells and single cells. Proc Natl Acad Sci U S A 110: 594-599
Peleg S., Sananbenesi F., Zovoilis A., Burkhardt S., Bahari-Javan S., et al. (2010) Altered histone acetylation is associated with age-dependent memory impairment in mice. Science 328: 753-756
Persico A. M. and Napolioni V. (2013) Autism genetics. Behav Brain Res 251: 95-112
Poduri A., Evrony G. D., Cai X. and Walsh C. A. (2013) Somatic mutation, genomic variation, and neurological disease. Science 341: 1237758
Pratt J., Winchester C., Dawson N. and Morris B. (2012) Advancing schizophrenia drug discovery: optimizing rodent models to bridge the translational gap. Nat Rev Drug Discov 11: 560-579
Priebe L., Degenhardt F. A., Herms S., Haenisch B., Mattheisen M., et al. (2012) Genome-wide survey implicates the influence of copy number variants (CNVs) in the development of early-onset bipolar disorder. Mol Psychiatry 17: 421-432
Purcell S. M., Moran J. L., Fromer M., Ruderfer D., Solovieff N., et al. (2014) A polygenic burden of rare disruptive mutations in schizophrenia. Nature 506: 185-190
Qin J., Li R., Raes J., Arumugam M., Burgdorf K. S., et al. (2010) A human gut microbial gene catalogue established by metagenomic sequencing. Nature 464: 59-65
Quinlan A. R. and Hall I. M. (2012) Characterizing complex structural variation in germline and somatic genomes. Trends Genet 28: 43-53
Rabbani B., Tekin M. and Mahdieh N. (2014) The promise of whole-exome sequencing in medical genetics. J Hum Genet 59: 5-15
Raj T., Ryan K. J., Replogle J. M., Chibnik L. B., Rosenkrantz L., et al. (2014) CD33: increased inclusion of exon 2 implicates the Ig V-set domain in Alzheimer's disease susceptibility. Hum Mol Genet 23: 2729-2736
Ramos-Quiroga J. A., Sanchez-Mora C., Casas M., Garcia-Martinez I., Bosch R., et al. (2014) Genome-wide copy number variation analysis in adult attention-deficit and hyperactivity disorder. J Psychiatr Res 49: 60-67
Redon R., Ishikawa S., Fitch K. R., Feuk L., Perry G. H., et al. (2006) Global variation in copy number in the human genome. Nature 444: 444-454
Rees E., Kirov G., Sanders A., Walters J. T., Chambert K. D., et al. (2014) Evidence that duplications of 22q11.2 protect against schizophrenia. Mol Psychiatry 19: 37-40
Reyes A., Semenkovich N. P., Whiteson K., Rohwer F. and Gordon J. I. (2012) Going viral: next-generation sequencing applied to phage populations in the human gut. Nat Rev Microbiol 10: 607-617
Ribeiro F. M., Camargos E. R., Souza L. C. and Teixeira A. L. (2013) Animal models of neurodegenerative diseases. Rev Bras Psiquiatr 35 Suppl 2: S82-91
Ripke S., O'Dushlaine C., Chambert K., Moran J. L., Kahler A. K., et al. (2013) Genome-wide association analysis identifies 13 new risk loci for schizophrenia. Nat Genet 45: 1150-1159
Rosti R. O., Sadek A. A., Vaux K. K. and Gleeson J. G. (2014) The genetic landscape of autism spectrum disorders. Dev Med Child Neurol 56: 12-18
Rousseaux M. W., Marcogliese P. C., Qu D., Hewitt S. J., Seang S., et al. (2012) Progressive dopaminergic cell loss with unilateral-to-bilateral progression in a genetic model of Parkinson disease. Proc Natl Acad Sci U S A 109: 15918-15923
Ryan S. D., Dolatabadi N., Chan S. F., Zhang X., Akhtar M. W., et al. (2013) Isogenic human iPSC Parkinson's model shows nitrosative stress-induced

57 An Overview of Publications Featuring Illumina® Technology
dysfunction in MEF2-PGC1alpha transcription. Cell 155: 1351-1364
Rye P. D., Booij B. B., Grave G., Lindahl T., Kristiansen L., et al. (2011) A novel blood test for the early detection of Alzheimer's disease. J Alzheimers Dis 23: 121-129
Sakai Y., Shaw C. A., Dawson B. C., Dugas D. V., Al-Mohtaseb Z., et al. (2011) Protein interactome reveals converging molecular pathways among autism disorders. Sci Transl Med 3: 86ra49
Sanders S. J., Ercan-Sencicek A. G., Hus V., Luo R., Murtha M. T., et al. (2011) Multiple recurrent de novo CNVs, including duplications of the 7q11.23 Williams syndrome region, are strongly associated with autism. Neuron 70: 863-885
Sandoe J. and Eggan K. (2013) Opportunities and challenges of pluripotent stem cell neurodegenerative disease models. Nat Neurosci 16: 780-789
Savva G. M., Stephan B. C. and Alzheimer's Society Vascular Dementia Systematic Review G. (2010) Epidemiological studies of the effect of stroke on incident dementia: a systematic review. Stroke 41: e41-46
Schaaf C. P. and Zoghbi H. Y. (2011) Solving the autism puzzle a few pieces at a time. Neuron 70: 806-808
Schizophrenia Working Group of the Psychiatric Genomics C. (2014) Biological insights from 108 schizophrenia-associated genetic loci. Nature 511: 421-427
Schloissnig S., Arumugam M., Sunagawa S., Mitreva M., Tap J., et al. (2013) Genomic variation landscape of the human gut microbiome. Nature 493: 45-50
Scholz S. W., Mhyre T., Ressom H., Shah S. and Federoff H. J. (2012) Genomics and bioinformatics of Parkinson's disease. Cold Spring Harb Perspect Med 2: a009449
Schreiber M., Dorschner M. and Tsuang D. (2013) Next-generation sequencing in schizophrenia and other neuropsychiatric disorders. Am J Med Genet B Neuropsychiatr Genet 162B: 671-678
Screen M., Jonson P. H., Raheem O., Palmio J., Laaksonen R., et al. (2014) Abnormal splicing of NEDD4 in myotonic dystrophy type 2: possible link to statin adverse reactions. Am J Pathol 184: 2322-2332
Sekiyama K., Takamatsu Y., Waragai M. and Hashimoto M. (2014) Role of genomics in translational research for Parkinson's disease. Biochem Biophys Res Commun 452: 226-235
Selkoe D. J. (2012) Preventing Alzheimer's disease. Science 337: 1488-1492
Sharma M., Kruger R. and Gasser T. (2014) From genome-wide association studies to next-generation sequencing: lessons from the past and planning for the future. JAMA Neurol 71: 5-6
Shen J., Bronson R. T., Chen D. F., Xia W., Selkoe D. J., et al. (1997) Skeletal and CNS defects in Presenilin-1-deficient mice. Cell 89: 629-639
Sherva R., Tripodis Y., Bennett D. A., Chibnik L. B., Crane P. K., et al. (2014) Genome-wide association study of the rate of cognitive decline in Alzheimer's disease. Alzheimers Dement 10: 45-52
Sibley C. R., Seow Y., Curtis H., Weinberg M. S. and Wood M. J. (2012) Silencing of Parkinson's disease-associated genes with artificial mirtron mimics of miR-1224. Nucleic Acids Res 40: 9863-9875
Siderowf A., Xie S. X., Hurtig H., Weintraub D., Duda J., et al. (2010) CSF amyloid {beta} 1-42 predicts cognitive decline in Parkinson disease. Neurology 75: 1055-1061
Singh S., Kumar A., Agarwal S., Phadke S. R. and Jaiswal Y. (2014) Genetic insight of schizophrenia: past and future perspectives. Gene 535: 97-100
Siniscalco D., Bradstreet J. J., Sych N. and Antonucci N. (2014) Mesenchymal stem cells in treating autism: Novel insights. World J Stem Cells 6: 173-178
Sinkus M. L., Adams C. E., Logel J., Freedman R. and Leonard S. (2013) Expression of immune genes on chromosome 6p21.3-22.1 in schizophrenia. Brain Behav Immun 32: 51-62
Smith K. R. and Matson J. L. (2010) Social skills: differences among adults with intellectual disabilities, co-morbid autism spectrum disorders and epilepsy. Res Dev Disabil 31: 1366-1372
Stefansson H., Meyer-Lindenberg A., Steinberg S., Magnusdottir B., Morgen K., et al. (2014) CNVs conferring risk of autism or schizophrenia affect cognition in controls. Nature 505: 361-366
Stilling R. M., Dinan T. G. and Cryan J. F. (2014) Microbial genes, brain & behaviour - epigenetic regulation of the gut-brain axis. Genes Brain Behav 13: 69-86
Stoll G., Pietilainen O. P., Linder B., Suvisaari J., Brosi C., et al. (2013) Deletion of TOP3beta, a component of FMRP-containing mRNPs, contributes to neurodevelopmental disorders. Nat Neurosci 16: 1228-1237
Sun Y., Christensen J. and Olsen J. (2013) Childhood epilepsy and maternal antibodies to microbial and tissue antigens during pregnancy. Epilepsy Res 107: 61-74
Szatkiewicz J. P., O'Dushlaine C., Chen G., Chambert K., Moran J. L., et al. (2014) Copy number variation in schizophrenia in Sweden. Mol Psychiatry 19: 762-773
Szatmari P. (1999) Heterogeneity and the genetics of autism. J Psychiatry Neurosci 24: 159-165
Todarello G., Feng N., Kolachana B. S., Li C., Vakkalanka R., et al. (2014) Incomplete penetrance of NRXN1 deletions in families with schizophrenia. Schizophr Res 155: 1-7
Torrey E. F. and Yolken R. H. (2003) Toxoplasma gondii and schizophrenia. Emerg Infect Dis 9: 1375-1380
Trinh J. and Farrer M. (2013) Advances in the genetics of Parkinson disease. Nat Rev Neurol 9: 445-454
Tucker T., Zahir F. R., Griffith M., Delaney A., Chai D., et al. (2014) Single exon-resolution targeted chromosomal microarray analysis of known and candidate intellectual disability genes. Eur J Hum Genet 22: 792-800
Vidaki A., Daniel B. and Court D. S. (2013) Forensic DNA methylation profiling--potential opportunities and challenges. Forensic Sci Int Genet 7: 499-507
Visscher P. M., Brown M. A., McCarthy M. I. and Yang J. (2012) Five years of GWAS discovery. Am J Hum Genet 90: 7-24
Wang S. C., Oelze B. and Schumacher A. (2008) Age-specific epigenetic drift in late-onset Alzheimer's disease. PLoS One 3: e2698
Watson K. K. and Platt M. L. (2012) Of mice and monkeys: using non-human primate models to bridge mouse- and human-based investigations of autism spectrum disorders. J Neurodev Disord 4: 21
Weaver W. M., Tseng P., Kunze A., Masaeli M., Chung A. J., et al. (2014) Advances in high-throughput single-cell microtechnologies. Curr Opin Biotechnol 25: 114-123
Willsey A. J., Sanders S. J., Li M., Dong S., Tebbenkamp A. T., et al. (2013) Coexpression networks implicate human midfetal deep cortical projection neurons in the pathogenesis of autism. Cell 155: 997-1007
Wohr M. and Scattoni M. L. (2013) Behavioural methods used in rodent models of autism spectrum disorders: current standards and new developments. Behav Brain Res 251: 5-17
Wong C. C., Meaburn E. L., Ronald A., Price T. S., Jeffries A. R., et al. (2014) Methylomic analysis of monozygotic twins discordant for autism spectrum disorder and related behavioural traits. Mol Psychiatry 19: 495-503
Wright J. (2014) Genetics: Unravelling complexity. Nature 508: S6-7
Yoon K. J., Nguyen H. N., Ursini G., Zhang F., Kim N. S., et al. (2014) Modeling a genetic risk for schizophrenia in iPSCs and mice reveals neural stem cell deficits associated with adherens junctions and polarity. Cell Stem Cell 15: 79-91
Young G. T., Gutteridge A., Fox H. D., Wilbrey A. L., Cao L., et al. (2014) Characterizing Human Stem Cell-derived Sensory Neurons at the Single-cell Level Reveals Their Ion Channel Expression and Utility in Pain Research. Mol Ther 22: 1530-1543

58 Neurobiology Research Review
Yu T. W., Chahrour M. H., Coulter M. E., Jiralerspong S., Okamura-Ikeda K., et al. (2013) Using whole-exome sequencing to identify inherited causes of autism. Neuron 77: 259-273
Zarowiecki M. (2012) Metagenomics with guts. Nat Rev Microbiol 10: 674
Zhang B., Gaiteri C., Bodea L. G., Wang Z., McElwee J., et al. (2013) Integrated systems approach identifies genetic nodes and networks in late-onset Alzheimer's disease. Cell 153: 707-720
Zhang D., Cheng L., Qian Y., Alliey-Rodriguez N., Kelsoe J. R., et al. (2009) Singleton deletions throughout the genome increase risk of bipolar disorder. Mol Psychiatry 14: 376-380
Zhang K., Schrag M., Crofton A., Trivedi R., Vinters H., et al. (2012) Targeted proteomics for quantification of histone acetylation in Alzheimer's disease. Proteomics 12: 1261-1268
Zhang Y., Schulz V. P., Reed B. D., Wang Z., Pan X., et al. (2013) Functional genomic screen of human stem cell differentiation reveals pathways involved in neurodevelopment and neurodegeneration. Proc Natl Acad Sci U S A 110: 12361-12366
Zhou H., Huang C., Tong J., Hong W. C., Liu Y. J., et al. (2011) Temporal expression of mutant LRRK2 in adult rats impairs dopamine reuptake. Int J Biol Sci 7: 753-761
Zhou W., Bercury K., Cummiskey J., Luong N., Lebin J., et al. (2011) Phenylbutyrate up-regulates the DJ-1 protein and protects neurons in cell culture and in animal models of Parkinson disease. J Biol Chem 286: 14941-14951

Illumina • 1.800.809.4566 toll-free (U.S.) • +1.858.202.4566 tel • www.illumina.com
FOR RESEARCH USE ONLY© 2014 Illumina, Inc. All rights reserved. Illumina, IlluminaDx, BaseSpace, BeadArray, BeadXpress, cBot, CSPro, DASL, DesignStudio, Eco, GAIIx, Genetic Energy, Genome Analyzer, GenomeStudio, GoldenGate, HiScan, HiSeq, Infinium, iSelect, MiSeq, Nextera, NuPCR, SeqMonitor, Solexa, TruSeq, TruSight, VeraCode, the pumpkin orange color, and the Genetic Energy streaming bases design are trademarks or registered trademarks of Illumina, Inc. All other brands and names contained herein are the property of their respective owners. Pub. No. 1070-2014-008 Current as of 09 October 2014
This Scientific Publication Review is brought to you by Illumina. Access previous Scientific Publication Reviews in the archive at www.illumina.com/pubreviews





























