Embed Size (px)
Citation preview

NaKR1 regulates long-distance movement ofFLOWERING LOCUS T in ArabidopsisYang Zhu, Lu Liu, Lisha Shen and Hao Yu*
Flowering plants perceive photoperiodic signals in leaves to generate mobile stimuli required for the induction of flowerformation at shoot apices. Although FLOWERING LOCUS T (FT) has been identified as part of the mobile floral stimuli inArabidopsis thaliana, the mechanisms underlying long-distance movement of FT from leaves to shoot apices remain largelyunclear. Here we show that a heavy-metal-associated (HMA) domain-containing protein, SODIUM POTASSIUM ROOTDEFECTIVE 1 (NaKR1), is activated by CONSTANS (CO) under long-day conditions and regulates long-distance movementof FT in Arabidopsis. Loss of function of NaKR1 compromises FT transport to shoot apices through sieve elements, causinglate flowering under long-day conditions. NaKR1 and FT share similar expression patterns and subcellular localization, andinteract with each other in vivo. Grafting experiments demonstrate that NaKR1 promotes flowering through mediating FTtranslocation from leaves to shoot apices. Thus, photoperiodic control of floral induction requires NaKR1-mediated long-distance delivery of florigenic signals.
Flowering plants respond to various environmental and develop-mental signals to determine the timing of the transition fromvegetative to reproductive growth. The relative length of night
and day periods, known as photoperiod, is a key environmentalcue that contributes to plant flowering responses in changingseasons. Classic studies have suggested that perception of photo-period in leaves generates a long-distance mobile floral stimuluscalled florigen that moves to the shoot apex to induce the formationof flowers1,2. The proteins encoded by FLOWERING LOCUS T (FT)in Arabidopsis and its orthologues in other plant species have beensuccessively identified as part of the long-sought florigen3–8.
In Arabidopsis, FT transcription is activated by a nuclear zinc-finger transcriptional regulator, CO, in the vasculature of leaves inresponse to long days9–13. An endoplasmic reticulum (ER) mem-brane protein, FT-INTERACTING PROTEIN 1 (FTIP1), interactswith FT in companion cells of the phloem and mediates FTprotein movement from companion cells to sieve elements14. Thiscontributes to regulation of FT transport through the phloem tothe shoot apical meristem, where FT interacts with a bZIP transcrip-tion factor FD to activate transcription of floral meristem identitygenes to initiate subsequent flower development9,15–17. Althoughthese findings have advanced our understanding of FT functionand transport, it is so far unclear how long-distance movement ofFT in the phloem stream is regulated in response to photoperiod.
In plants, a group of metallochaperone-like proteins contain aheavy metal-associated (HMA) domain characterized by a conservedM/LXCXXC metal-binding motif18–20. They play important roles intransporting metallic ions to specific cellular sites and are dividedinto two groups19. The first group includes heavy metal-associatedisoprenylated plant proteins (HIPPs) that contain an isoprenylationmotif and are only present in vascular plants, whereas the secondgroup includes heavy metal-associated plant proteins (HPPs) thatlack the isoprenylation motif and are found in almost all plantspecies18,19. In Arabidopsis, HIPPs are involved in heavy metalhomeostasis and detoxification as well as transcriptional responsesto stresses18,19,21–23, whereas functional characterization of HPPs hasbeen rather limited. SODIUM POTASSIUM ROOT DEFECTIVE 1(NaKR1) is one of the reported Arabidopsis HPPs, which is
specifically expressed in companion cells of the root phloem20,24–26.The nakr1-1 mutant shows pleiotropic phenotypes relevant todefects in phloem function, such as accumulation of Na+ andstarch in leaves, decreased transport of sucrose, and late floweringunder long-day conditions20.
In this study, we show that NaKR1 expression displays circadianrhythm and is specifically induced by CO in leaves in response tolong days. We provide evidence that NaKR1 interacts in vivo with FT,and promotes flowering by regulating long-distance movement of FTfrom leaves to the shoot apex through the phloem stream. Our resultssuggest that NaKR1 is a hitherto unknown regulator specifically requiredfor photoperiodic control of long-distance delivery of florigenic signals.
ResultsPhotoperiodic control of NaKR1 expression is dependent on CO.It has been shown thatNaKR1 affects flowering time under long-dayconditions20,24–26. To elucidate the function of NaKR1 during thefloral transition, we tested how NaKR1 regulates flowering inresponse to known flowering signals using the reported nakr1-1mutant that contains a 7-bp frameshift deletion in the first exon20
(Supplementary Fig. 1a). nakr1-1 flowered late under long-day butnot under short-day conditions (Fig. 1a,b). We transformednakr1-1 with a genomic construct (gNaKR1) harbouring a 5.5-kbNaKR1 genomic region including the 2.8-kb upstream sequenceand the 2.7-kb coding sequence plus three introns. Most nakr1-1gNaKR1 T1 transformants showed similar flowering time to wild-type plants under long-day conditions (Supplementary Fig. 1b),suggesting that NaKR1 promotes flowering under long-dayconditions. In contrast, there were minimal effects of NaKR1 onaltering the flowering response to vernalization and gibberellin(GA) treatment as well as changes in ambient temperature(Supplementary Fig. 2a–d).
To confirm NaKR1 function in the control of flowering time, wealso generated NaKR1 knockdown transgenic plants by artificialmicroRNA (AmiR) interference27. Most of AmiR-nakr1 indepen-dent lines that expressed an AmiR specifically targeting at thethird exon of the NaKR1 mRNA (Supplementary Fig. 1a) exhibitedlate flowering under long-day conditions (Supplementary Fig. 3a,c).
Department of Biological Sciences and Temasek Life Sciences Laboratory, National University of Singapore, Singapore 117543, Singapore.*e-mail: [email protected]
ARTICLESPUBLISHED: 27 MAY 2016 | ARTICLE NUMBER: 16075 | DOI: 10.1038/NPLANTS.2016.75
NATURE PLANTS | VOL 2 | JUNE 2016 | www.nature.com/natureplants 1
© 2016 Macmillan Publishers Limited. All rights reserved
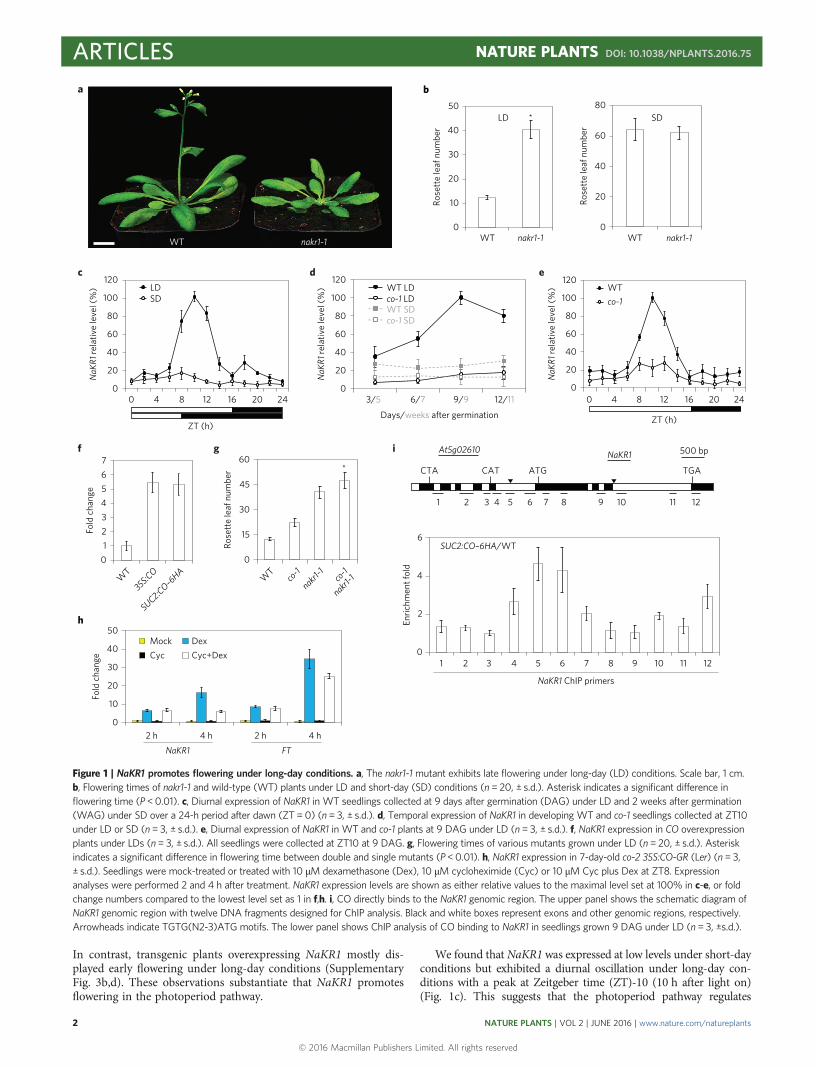
In contrast, transgenic plants overexpressing NaKR1 mostly dis-played early flowering under long-day conditions (SupplementaryFig. 3b,d). These observations substantiate that NaKR1 promotesflowering in the photoperiod pathway.
We found that NaKR1 was expressed at low levels under short-dayconditions but exhibited a diurnal oscillation under long-day con-ditions with a peak at Zeitgeber time (ZT)-10 (10 h after light on)(Fig. 1c). This suggests that the photoperiod pathway regulates
a b50
40LD *
30
20
10Rose
tte
leaf
num
ber
0
80
60
40
20Rose
tte
leaf
num
ber
0
60
30
15
45
Rose
tte
leaf
num
ber
0
80
100
120
60
40
20
NaK
R1 re
lativ
e le
vel (
%)
0
80
100
120
60
40
20
NaK
R1 re
lativ
e le
vel (
%)
0
80
100
120
WT nakr1-1
60
40
20
NaK
R1 re
lativ
e le
vel (
%)
0
0
2 h 4 hNaKR1 FT
WT
35S:CO
SUC2:CO–6HA
WT
co-1 co-1
nakr1-1
nakr1-1
123
*
456
Fold
cha
nge
0
0
2
4
6
10
20
30
40
50Mock DexCyc Cyc+Dex
Fold
cha
nge
7
03/5 6/7 9/9 12/11 4 8 12
ZT (h)ZT (h)
Days/weeks after germination
16 20 240 4 8 12 16 20 24
WT nakr1-1 WT
WTco-1
LDSD
nakr1-1
c d e
f g
h
i
SD
2 h 4 h
1 2 3 4 5 6 7 8 9 10 11 12
1
CTA CAT ATG TGA
2 3 4 5 6 7 8 9 10 11 12
NaKR1 ChIP primers
Enric
hmen
t fol
d
SUC2:CO–6HA/WT
NaKR1At5g02610 500 bp
WT LDco-1 LDWT SDco-1 SD
Figure 1 | NaKR1 promotes flowering under long-day conditions. a, The nakr1-1 mutant exhibits late flowering under long-day (LD) conditions. Scale bar, 1 cm.b, Flowering times of nakr1-1 and wild-type (WT) plants under LD and short-day (SD) conditions (n = 20, ± s.d.). Asterisk indicates a significant difference inflowering time (P <0.01). c, Diurnal expression of NaKR1 in WT seedlings collected at 9 days after germination (DAG) under LD and 2 weeks after germination(WAG) under SD over a 24-h period after dawn (ZT= 0) (n= 3, ± s.d.). d, Temporal expression of NaKR1 in developing WT and co-1 seedlings collected at ZT10under LD or SD (n= 3, ± s.d.). e, Diurnal expression of NaKR1 in WT and co-1 plants at 9 DAG under LD (n = 3, ± s.d.). f, NaKR1 expression in CO overexpressionplants under LDs (n= 3, ± s.d.). All seedlings were collected at ZT10 at 9 DAG. g, Flowering times of various mutants grown under LD (n = 20, ± s.d.). Asteriskindicates a significant difference in flowering time between double and single mutants (P <0.01). h, NaKR1 expression in 7-day-old co-2 35S:CO-GR (Ler) (n = 3,± s.d.). Seedlings were mock-treated or treated with 10 µM dexamethasone (Dex), 10 µM cycloheximide (Cyc) or 10 µM Cyc plus Dex at ZT8. Expressionanalyses were performed 2 and 4 h after treatment. NaKR1 expression levels are shown as either relative values to the maximal level set at 100% in c-e, or foldchange numbers compared to the lowest level set as 1 in f,h. i, CO directly binds to the NaKR1 genomic region. The upper panel shows the schematic diagram ofNaKR1 genomic region with twelve DNA fragments designed for ChIP analysis. Black and white boxes represent exons and other genomic regions, respectively.Arrowheads indicate TGTG(N2-3)ATG motifs. The lower panel shows ChIP analysis of CO binding to NaKR1 in seedlings grown 9 DAG under LD (n = 3, ±s.d.).
ARTICLES NATURE PLANTS DOI: 10.1038/NPLANTS.2016.75
NATURE PLANTS | VOL 2 | JUNE 2016 | www.nature.com/natureplants2
© 2016 Macmillan Publishers Limited. All rights reserved

NaKR1 expression. As NaKR1 was downregulated under long-dayconditions in several photoperiod pathway mutants, particularlyco-1 (Supplementary Fig. 4a), we further examinedNaKR1 expressionin co-1 and wild-type developing seedlings collected at ZT10 underboth long- and short-day conditions. In contrast to constantly lowexpression of NaKR1 in co-1 and wild-type seedlings under short-day conditions, NaKR1 expression under long-day conditionsincreased gradually at the vegetative phase and peaked during thefloral transition occurring between 9 to 13 days after germinationin wild-type seedlings, but was dramatically downregulated in co-1(Fig. 1d). Consistently, there was no circadian rhythm of NaKR1expression in co-1 under long-day conditions (Fig. 1e). In contrast,overexpression of CO upregulated NaKR1 under long-day conditions(Fig. 1f). In addition, induced CO activity upregulated both NaKR1and FT in a steroid-inducible co-2 35S:CO-GR line12 under the singletreatment of dexamethasone or a combined treatment of dexametha-sone and cycloheximide, an inhibitor of translation (Fig. 1h), suggestingthat NaKR1 may be an immediate target of transcriptional pro-motion by CO.
We then performed chromatin immunoprecipitation (ChIP) assayusing a functional SUC2:CO–6HA transgenic plant28 to examinewhether CO physically binds to the NaKR1 locus. We searched theNaKR1 genomic sequence for the TGTG(N2-3)ATGmotif, a potentialbinding site for CO29, and designed 12 primer pairs covering the
NaKR1 locus including two identified motifs to measure DNA enrich-ment (Fig. 1i). ChIP assays revealed that CO–6HAwas associated withthe region containing a TGTG(N2-3)ATG motif (TGTGAGAATG) atthe 5′ region (Fig. 1i). These results suggest that CO directly promotesNaKR1 expression in the photoperiod pathway. As co-1 nakr1-1 doublemutants still flowered much later than either of the single-mutantcounterparts under long-day conditions (Fig. 1g), CO and NaKR1may also affect flowering through other independent targets.
NaKR1 expression was not obviously affected in response to verna-lization and GA treatment and also not by several other flowering timeregulators that mediate flowering signals from various genetic pathways(Supplementary Fig. 4b–d). However, NaKR1 was downregulated invarious autonomous-pathway mutants (Supplementary Fig. 4e), indi-cating that the autonomous pathway also affects NaKR1 expression.
NaKR1 functions in vascular tissues to promote flowering.Quantitative real-time PCR revealed that NaKR1 was expressed inall the tissues examined with the highest expression in rosetteleaves (Fig. 2a). To monitor NaKR1 expression during the floraltransition, we established an NaKR1:β-glucuronidase (GUS)reporter construct (gNaKR1:GUS) in which the NaKR1 genomicfragment used for the gene complementation test (SupplementaryFig. 1b) was fused to the GUS gene. Among 25 independentgNaKR1:GUS lines created, most lines showed similar GUS
80
100
120
60
40
20NaK
R1 re
lativ
e le
vel (
%)
0Rt RL CL St FB OF Sil 6
0
2
4
6
8
10
12WTnakr1-1nakr1-1 SUC2:NaKR1nakr1-1 KNAT1:NaKR1
7 8 9 10 11 12 13 37 38 39 40 41 42 43
Rosette leaf number
Num
ber o
f pla
nts
a
b
e f g h
c d
i
Figure 2 | NaKR1 is expressed and functional in leaf vascular tissues. a, Quantitative real-time PCR analysis of NaKR1 expression in various tissues of WTplants (n=3, ± s.d.). All samples were collected at ZT10. Expression levels are shown as relative values to the maximal level set at 100%. Rt, roots; RL, rosetteleaves; CL, cauline leaves; St, inflorescence stems; FB, flower buds; OF, open flowers; Sil, siliques. b–g, Representative histochemical GUS staining of gNaKR1:GUStransgenic plants displays NaKR1 expression in seedlings at 5 (b), 7 (c), 9 (d) and 11 (e) DAG, a rosette leaf (f), and a primary root (g). Scale bars, 1 mm (b–e),0.5 mm (f) and 10 µm (g). h, An enlarged view of a seedling at 11 DAG showing no NaKR1 expression in the shoot apex. Scale bar, 25 µm. i, Distribution offlowering time in T1 transgenic plants grown under LD carrying the SUC2:NaKR1 or KNAT1:NaKR1 construct in the nakr1-1 background.
NATURE PLANTS DOI: 10.1038/NPLANTS.2016.75 ARTICLES
NATURE PLANTS | VOL 2 | JUNE 2016 | www.nature.com/natureplants 3
© 2016 Macmillan Publishers Limited. All rights reserved

staining patterns. Further investigation of one representative linerevealed specific GUS expression in vascular tissues of cotyledons,rosette leaves and roots in developing seedlings before and duringthe floral transition occurring 9–13 days after germination(Fig. 2b–g). In contrast, no GUS signal was detected in thevegetative shoot apex (Fig. 2h). To investigate the effect of spatialexpression of NaKR1 on flowering time, we transformed nakr1-1
with the construct in which the NaKR1 coding sequence wasdriven by the promoter of either SUCROSE TRANSPORTER 2(SUC2) or KNOTTED-LIKE FROM ARABIDOPSIS THALIANA 1(KNAT1), which is specifically expressed either in phloemcompanion cells30 or shoot apical meristems11, respectively. Mostnakr1-1 KNAT1:NaKR1 T1 transformants exhibited similarflowering time to nakr1-1, whereas most nakr1-1 SUC2:NaKR1 T1
0CC SE XV
P = 5.3 × 10−9
Anti-HA lgG
MC CC SE XV MC
Num
ber o
f gol
d pa
rtic
les
per µ
m2
1
2
3
4
5
6
7
35S:
NaK
R1–G
FP
gNaK
R1–G
FP
gNaK
R1–4
HA
CC SE XV
Anti-HA lgG
MC CC SE XV MC
80
100
60
Freq
uenc
y (%
)
40
20
0
+Gold particles−Gold particles
Bright fieldGFP
ER–RFPBright fieldGFP
CC SE XV MC
Merge
Merge
a
c
d
e f
b
P = 1.2 × 10−8
Rose
tte
leaf
num
ber
0
10
20
30
40
50
WTnakr1-1
nakr1-1 gNaKR1–4HAnakr1-1 gNaKR1–GFP
Figure 3 | NaKR1 is localized in the phloem. a, Flowering times of various transgenic lines grown under LD (n = 20, ± s.d.). b, Confocal analysis of nakr1-1gNaKR1–GFP seedlings at 11 DAG showing NaKR1–GFP localization in the vasculature of leaves. Scale bar, 100 µm. c, Subcellular localization of NaKR1–GFP inN. benthamiana leaf epidermal cells. ER–RFP, RFP fluorescence of an ER marker. Scale bar, 10 µm. d, Analysis of NaKR1–4HA localization by immunogoldelectron microscopy using anti-HA antibody in companion cells (CC), sieve elements (SE) and xylem vessels (XV) in the vasculature in the first rosetteleaves and shoot meristem cells (MC) of nakr1-1 gNaKR1–4HA seedlings at 11 DAG. Arrows indicate gold particles. Scale bars, 1 µm. e, Quantification ofNaKR1–4HA immunogold signals or background signals in sections (as shown in d) probed with anti-HA antibody or mouse IgG control, respectively, invarious cells of nakr1-1 gNaKR1–4HA (n = 60, ± s.d.). The results are considered statistically significant at P < 0.001. f, Frequency histograms of appearance ofNaKR1–4HA immunogold signals or background signals in sections (as shown in d) probed with anti-HA antibody or mouse IgG control, respectively, invarious cells of nakr1-1 gNaKR1–4HA.
ARTICLES NATURE PLANTS DOI: 10.1038/NPLANTS.2016.75
NATURE PLANTS | VOL 2 | JUNE 2016 | www.nature.com/natureplants4
© 2016 Macmillan Publishers Limited. All rights reserved

transformants flowered either earlier than or similar to wild-typeplants (Fig. 2i). These results suggest that NaKR1 expression invascular tissues is required for promoting flowering.
As previous studies have shown that NaKR1 protein is localized inroot companion cells and could be mobile in the root phloem20,24, wefurther explored its protein localization in aerial tissues. To this end,we generated an nakr1-1 gNaKR1–GFP transgenic line, in which thegreen fluorescent protein (GFP) cDNA fused with the NaKR1genomic fragment (Supplementary Fig. 1b) rescued the late-floweringphenotype of nakr1-1 (Fig. 3a). NaKR1–GFP signal notably markedthe vascular tissues of the rosette leaves in nakr1-1 gNaKR1–GFPseedlings (Fig. 3b). We also examined the subcellular localization ofNaKR1 in tobacco leaves using 35S:NaKR1–GFP. NaKR1–GFP wasmostly colocalized with the red fluorescent protein-tagged (RFP)endoplasmic reticulum (ER) marker ER-RFP31 in whole cells exceptthe nuclei (Fig. 3c). To precisely localize NaKR1, we performedimmunoelectronmicroscopy on an nakr1-1 gNaKR1–4HA transgenicline, in which gNaKR1–4HA rescued the late-flowering phenotype ofnakr1-1 (Fig. 3a). Quantitative analysis of labelling density revealedthat NaKR1–4HA signals were specifically detected by anti-HA anti-body (Fig. 3d–f and Supplementary Figs 5,6). Notably, NaKR1–4HAwas localized mostly in the phloem (companion cell-sieve elementcomplexes) in rosette leaves and below shoot apices of nakr1-1gNaKR1–4HA, but barely detectable in xylem vessels and shootmeristem cells during the floral transition.
NaKR1 interacts with FT. To elucidate how NaKR1 affectsflowering time, we performed yeast two-hybrid screening toidentify interacting partners of NaKR1, and found that one of theinteractors encoded FT. Yeast two-hybrid assays confirmed thatthe full-length NaKR1 or its truncated HMA domain interactedwith FT (Fig. 4a). This result, together with the observations onsimilar expression patterns of NaKR1 and FT in vasculartissues13,14 (Fig. 2b–g) and comparable subcellular localization ofNaKR1–GFP and FT–GFP (Fig. 3c and Supplementary Fig. 7),prompted us to carry out a detailed analysis of their interaction.
Glutathione S-transferase (GST) pull-down assays demonstratedthat GST–NaKR1 bound to in vitro-translated full-length myc-FT(Fig. 4b). Bimolecular fluorescence complementation (BiFC) assayfurther revealed colocalization of the enhanced yellow fluorescentprotein (EYFP) fluorescence signal with ER-RFP31 in tobaccoepidermal cells (Fig. 4c), implying an interaction between NaKR1and FT on the ER in living plant cells. More importantly, coimmu-noprecipitation analysis on total extracts from nakr1-1 gNaKR1–4HA SUC2:FT–9myc seedlings collected at 9 days after germinationgrown under long-day conditions confirmed the in vivo interactionof NaKR1–4HA and FT–9myc in Arabidopsis (Fig. 4d). Theseresults demonstrate that NaKR1 interacts with FT in vivo duringthe floral transition. As genetic analysis showed that ft-10 nakr1-1flowered later than ft-10 (Fig. 4e), it is likely to be that NaKR1may also affect other flowering regulators in addition to FT. Theclosest homologue of FT, TWIN SISTER OF FT (TSF)32, could beanother potential target of NaKR1. However, TSF expressionremained unchanged in nakr1-1 and wild-type plants(Supplementary Fig. 8a). Yeast two-hybrid assay also did notreveal an interaction between TSF and NaKR1 (SupplementaryFig. 8b), implying that TSF might not be a direct target of NaKR1.
NaKR1 regulates FT transport. The findings on the interactionbetween NaKR1 and FT, their overlapping expression patterns at thetissue and subcellular levels and the nature of NaKR1 as a phloem-mobile regulator20 led us to hypothesize that NaKR1 may mediateFT transport in vascular tissues. We then examined the effect ofnakr1-1 on the early flowering phenotypes caused by FT expressionin various spatial patterns (Fig. 5a). nakr1-1 did not affect earlyflowering caused by overexpression of FT in whole plants driven by
the 35S promoter or expression of FT in the shoot apical meristemdriven by the KNAT1 promoter. However, flowering time resultingfrom expression of FT, FT–9myc and FT–GFP in phloemcompanion cells driven by the SUC2 promoter was all significantlydelayed in nakr1-1. Thus, NaKR1 may mediate FT effect onflowering prior to FT protein moving to the shoot apical meristem.
319
myc–FT
GST–NaKR1
GST
EYFP Bright field ER–RFP Merge
InputGST
GST–NaKR1250
NaKR1
nEYF
P–FT
cEYF
P–N
aKR1
N
nEYF
P–FT
cEYF
PnE
YFP
cEYF
P–N
aKR1
HMA Empty
Empty
Empty
FT
FT
ADBD
a
c
d e
b1
Input
015
30456075
*Anti-myc IP
FT–9myc
gNaKR1–4HA
nakr1-1
gNaKR1–4HA
nakr1-1
gNaKR1–4HA
SUC2:FT–9
myc
nakr1-1
gNaKR1–4HA
SUC2:FT–9
myc
nakr1-1
Rose
tte
leaf
num
ber
WT
nakr1-1
ft-10
ft-10
nakr1-1
N HMA
NaKR1–4HA
Figure 4 | NaKR1 interacts with FT. a, Yeast two-hybrid assay of theinteraction between NaKR1 and FT. Top panel shows the schematic diagramof the NaKR1 protein with a C-terminal heavy-metal-associated domain(HMA) and an N-terminal region (N). Transformed yeast cells harbouringvarious NaKR1 truncated proteins fused to AD (activation domain) and FTfused to BD (binding domain) were grown on SD/−Leu/−Trp medium(upper panel) and SD/−His/−Leu/−Trp medium supplemented with 4 mM3-amino-1,2,4-triazole (lower panel). b, In vitro GST pull-down assay withNaKR1 and FT. Myc-tagged FT protein generated by in vitro translation wasincubated with immobilized GST or GST–NaKR1. Immunoblot analysis wasperformed using anti-myc antibody. Input, 5% of the in vitro translationproduct. Asterisk indicates a non-specific band. c, BiFC analysis of theinteraction between NaKR1 and FT in N. benthamiana leaf epidermal cells.Scale bars, 10 µm. d, In vivo interaction between NaKR1 and FT shown byco-immunoprecipitation in Arabidopsis. Total protein extracts from nakr1-1gNaKR1–4HA and nakr1-1 gNaKR1–4HA SUC2:FT–9myc plants at 9 DAG wereimmunoprecipitated by anti-myc antibody. The input andcoimmunoprecipitated proteins were detected by either anti-HA or anti-mycantibody. e, Flowering phenotypes of ft-10 nakr1-1 under LD (n = 20, ± s.d.).Asterisk indicates a significant difference in flowering time of ft-10 nakr1-1compared with that of ft-10 or nakr1-1 (P <0.001).
NATURE PLANTS DOI: 10.1038/NPLANTS.2016.75 ARTICLES
NATURE PLANTS | VOL 2 | JUNE 2016 | www.nature.com/natureplants 5
© 2016 Macmillan Publishers Limited. All rights reserved

We found that the abundance of FT–9myc and FT–GFP fusionproteins remained unchanged in wild-type and nakr1-1 backgrounds(Fig. 5b and Supplementary Fig. 9a) in SUC2:FT–9myc and SUC2:FT–GFP, both of which greatly rescued the flowering defect of ft-10(ref. 14), respectively. This indicates that NaKR1 might not be directlyinvolved in regulating FT protein stability. We further performedimmunoelectron microscopy to examine FT–9myc distribution insieve elements during the floral transition. The labelling density ofFT–9myc in sieve elements in the first rosette leaves was comparablein SUC2:FT–9myc and nakr1-1 SUC2:FT–9myc (Fig. 5c,d), whereasthe density significantly decreased in sieve elements below theshoot apical meristem in nakr1-1 SUC2:FT–9myc compared toSUC2:FT–9myc (Fig. 5c,d). These results suggest that NaKR1 affects
FT–9myc transport in the phloem stream to the shoot apical meris-tem rather than its transport from companion cells to sieve elementsin leaves. Although we also observed downregulation of native FTexpression in nakr1-1 (Supplementary Fig. 9b,c), significantlyincreased expression of NaKR1 in phloem companion cells did notobviously affect FT transcription (Supplementary Fig. 9d–f). Thus,downregulation of FT expression in nakr1-1 could be an indirecteffect associated with mis-localization of FT protein in nakr1-1 asobserved in previous studies3,14.
As FTIP1 mediates FT transport from companion cells to sieveelements14, we next examined the relationship between NaKR1and FTIP1. FTIP1 expression was not greatly altered in nakr1-1under long-day conditions (Supplementary Fig. 10a). Yeast
Rose
tte
leaf
num
ber
SE in
leaf
SE in le
af
SE b
elow
SA
M
SE below SAM
SE in le
af
SE below SAM
0
11.2
2.1 2.26.3 6.6
*
**
***
3.1 3.97.9
12.39.4
26.8
40.5
WT35S:FT
nakr1-1 35S:FT
KNAT1:FT
nakr1-1 KNAT1:FT
10
20
30
40
50
SUC2:FT
SUC2:FT–9myc
SUC2:FT–GFP
nakr1-1 SUC2:FT–GFP
nakr1-1
nakr1-1 SUC2:FT–9myc
nakr1-1 SUC2:FT
WT
SUC2:FT–9
myc
nakr1-1
SUC2:FT
–9myc
0
0.0
2.0
1.2
1.6
0.4
0.8
*
1.00 1.02
SUC2:FT–9myc nakr1-1 SUC2:FT–9myc
SUC2:FT–9myc
Num
ber o
fgo
ld p
artic
les
per µ
m2
Freq
uenc
y of
gold
par
ticle
s (%
)
80
100
60
40
20
0
Anti-myc lgG
a b
c d
nakr1-1 SUC2:FT–9myc
Figure 5 | NaKR1 affects FT transport. a, Flowering times of various mutants or transgenic plants (Col background) grown under LD (n = 20, ± s.d.).Mean rosette leaf numbers are indicated at the top of each bar. Asterisks indicate significant difference in flowering time of transgenic plants in the nakr1-1background compared with that in the WT background (*P < 0.05, **P < 0.005, ***P <0.001). b, Western blot analysis using anti-myc antibody shows thecomparable abundance of FT–9myc protein in SUC2:FT–9myc and nakr1-1 SUC2:FT–9myc plants at 11 DAG (upper panel). Coomassie blue staining of the largeRubisco subunit (rbcL) was used as a loading control (lower panel). The numbers below each lane indicate the relative intensity of FT–9myc protein levels indifferent genetic backgrounds, calculated by first normalizing each signal for FT–9myc against the signal for rbcL and then against the value of SUC2:FT–9myc(middle lane). c, Analysis of FT–9myc localization by immunogold electron microscopy using anti-myc antibody in SEs in the first rosette leaves and belowthe shoot apical meristem (SAM) of SUC2:FT–9myc and nakr1-1 SUC2:FT–9myc seedlings at 11 DAG. Arrows indicate gold particles. Scale bars, 1 µm.d, Quantification (upper panel; n = 60, ± s.d.) and frequency histogram of appearance (lower panel) of FT–9myc immunogold signals or background signals inSE sections (as shown in c) probed with anti-myc antibody or mouse IgG control in SUC2:FT–9myc and nakr1-1 SUC2:FT–9myc. Asterisk indicates a significantdifference in gold particle density in SEs below the SAM of nakr1-1 SUC2:FT–9myc compared to that in SUC2:FT–9myc (P < 0.005).
ARTICLES NATURE PLANTS DOI: 10.1038/NPLANTS.2016.75
NATURE PLANTS | VOL 2 | JUNE 2016 | www.nature.com/natureplants6
© 2016 Macmillan Publishers Limited. All rights reserved

Donor plant
Recipient plant
Cot-graftedplant
GFP
WT
ft-10
nakr
1-1
ft-10
nak
r1-1
WT
*
* *
* *
ft-10 nakr1-1Recipient
ft-10 nakr1-1
WT ft-10
nakr1-1 ft-10 nakr1-1
Bright field Merge
0
15
30
45
75
60
Donor:WTSUC2:FT–GFPSUC2:FT–9mycMutant
0
0
WT ft-10nakr1-1
Anti-myc IgG
ft-10 nakr1-1 WT ft-10nakr1-1
ft-10 nakr1-1
20
40
60
80
100
0.2
0.4
Num
ber o
f gol
dpa
rtic
les
per µ
m2
Freq
uenc
y of
gol
dpa
rtic
les
(%)
0.6
0.8
1.0 Donor SUC2:FT–9myc
FT
CC
FT
NaKR1
CO
SE
SE
SE
SECC
Sourcetissues
SAM
1 (phloem in leaf)
2 (phloem below SAM)
1 2
Destinationtissues
Long-distancedelivery
NaKR1 FTIP1
Leaf
SAM
Leaf
CC
CC
PD
PD
a
c
d f
e
b
Rose
tte
leaf
num
ber
Figure 6 | NaKR1 is required for the long-distance movement of FT. a, A schematic illustration of a Cot-grafting procedure. A cotyledon is cut off from adonor (black) and a recipient (white). The donor cotyledon is then grafted to the excised petiole of the recipient. b, Flowering times of various recipientsgrafted with different donor cotyledons (n = 15, ± s.d.). Upright triangles indicate significant differences in WT recipients grafted with SUC2:FT–GFP orSUC2:FT–9myc cotyledons versus WT cotyledons (P < 0.001). Open triangles indicate significant differences in ft-10 recipients grafted with WT, SUC2:FT–GFPor SUC2:FT–9myc cotyledons versus ft-10 cotyledons (P <0.001). Asterisks indicate significant differences in nakr1-1 versus WT recipients grafted withwild-type, SUC2:FT–GFP or SUC2:FT–9myc cotyledons, respectively (P <0.001). Upside-down triangles indicate significant differences in ft-10 nakr1-1 versus ft-10recipients grafted with WT, SUC2:FT–GFP or SUC2:FT–9myc cotyledons, respectively (P <0.001). c, Confocal analysis of FT–GFP protein distribution in variousrecipients at 10 DAG grafted with SUC2:FT–GFP cotyledons. Arrows indicate the petioles in which grafting was performed. Scale bars, 100 µm. d, Analysis ofFT–9myc localization by immunogold electron microscopy in SEs below the shoot apical meristem (SAM) of various recipients at 10 DAG grafted with SUC2:FT–9myc cotyledons. Arrows indicate gold particles. Scale bars, 1 µm. e, Quantification (upper panel; n = 60, ± s.d.) and frequency histogram of appearance(lower panel) of FT–9myc immunogold signals in SE sections (as shown in d) in various recipients. Asterisks indicate significant differences in gold particledensity in SEs below the SAM of nakr1-1 and ft-10 nakr1-1 recipients compared to that of WT and ft-10 recipients, respectively, when probed with anti-mycantibody (P <0.001). f, NaKR1 regulates long-distance movement of FT. Photoperiodic control of CO activates expression of NaKR1 and FT in companion cells(CCs) of the vasculature. FTIP1 mediates the transport of FT protein from CCs to SEs, whereas NaKR1 is required for long-distance movement of FT fromleaves to the SAM through SEs.
NATURE PLANTS DOI: 10.1038/NPLANTS.2016.75 ARTICLES
NATURE PLANTS | VOL 2 | JUNE 2016 | www.nature.com/natureplants 7
© 2016 Macmillan Publishers Limited. All rights reserved

two-hybrid assay revealed no interaction between NaKR1 andFTIP1 (Supplementary Fig. 10b). Furthermore, the late-floweringphenotype of nakr1-1 was enhanced by ftip1-1 (SupplementaryFig. 10c). These observations indicate that NaKR1 and FTIP1may play sequential, but independent roles in regulating FTtransport from leaves to the shoot apex.
NaKR1 is required for long-distance movement of FT. To confirmNaKR1 role in mediating FT long-distance movement during thefloral transition, we applied an established cotyledonmicrografting (Cot-grafting) approach in Arabidopsis33 (Fig. 6a) tomonitor the effect of NaKR1 on FT transport through its nativeroute from leaves to the shoot apex. Grafting of wild-typecotyledons as donors significantly accelerated flowering of ft-10recipient plants compared to the grafting control using ft-10donor cotyledons (Fig. 6b). This is consistent with the previousreport33, indicating that the Cot-grafting surgery successfullyfacilitates the transport of florigen containing FT protein fromcotyledons to the shoot apex. Wild-type recipient plants graftedwith SUC2:FT–GFP or SUC2:FT–9myc cotyledons exhibitedearlier flowering than those with wild-type cotyledons, indicatingthat FT–GFP or FT–9myc are functional and mobile (Fig. 6b).Consistently, ft-10 recipient plants grafted with SUC2:FT–GFP orSUC2:FT–9myc cotyledons flowered earlier than those with graftedft-10 cotyledons (Fig. 6b). Notably, when grafted with wild-type,SUC2:FT–GFP or SUC2:FT–9myc cotyledons, flowering wasconsistently delayed in recipient plants in the nakr1-1 background(nakr1-1 or ft-10 nakr1-1) compared to wild-type or ft-10 recipientplants, respectively (Fig. 6b). These results demonstrate that NaKR1is important for floral induction resulting from long-distancemovement of FT through the graft junction.
We further monitored the movement of FT–GFP and FT–9mycin the grafted plants after graft recovery using confocal and immu-noelectron microscopy, respectively. Confocal microscopy revealedthat FT–GFP signal from SUC2:FT–GFP donor cotyledons wasobservable in the vasculature of the leaf petiole of wild-type andft-10 recipients, but not in the recipients in the nakr1-1 background(nakr1-1 or ft-10 nakr1-1) (Fig. 6c). Furthermore, in all plantsgrafted with SUC2:FT–9myc donor cotyledons, the labellingdensity of FT–9myc in sieve elements below the shoot apical meris-tem of nakr1-1 and ft-10 nakr1-1 recipient plants was significantlylower than that in wild-type and ft-10 recipient plants, respectively(Fig. 6d,e). Taken together, our results suggest that NaKR1 isrequired for long-distance movement of FT from leaves to theshoot apex through the phloem stream.
DiscussionDiscovery of FT-like proteins as part of florigen in various floweringplants has raised some outstanding questions regarding their move-ment from source leaves to sink tissues. Previous studies inArabidopsis have suggested that FT acts as a phloem-mobile signalthat moves from leaves to the shoot apical meristem to induceflowering3–5,34. However, the regulators that directly control long-distance movement of FT in the phloem stream are hithertounknown. Here we show that CO simultaneously activatesexpression of NaKR1 and FT in the leaf vasculature in response tophotoperiod, and that NaKR1 interacts with FT to mediate long-distance movement of FT from leaves to the SAM through thephloem stream (Fig. 6f). Our results suggest that FT transport tothe shoot apical meristem during the floral transition is orchestratedby the timely production of FT and its co-regulatorNaKR1 by CO inleaves in response to seasonal cues.
Several pieces of evidence support that NaKR1 is specificallyrequired for FT transport in the phloem stream. First, NaKR1–4HAis localized mostly in companion cell–sieve element complexes ofthe phloem in leaves and below shoot apices (Fig. 3d–f and
Supplementary Fig. 5). Second, NaKR1 specifically promotesFT–9myc transport in the phloem stream to the shoot apicalmeristem, but does not affect FT–9myc movement from companioncells to sieve elements in leaves (Fig. 5c,d). Third, graftingexperiments demonstrate that NaKR1 is required for floweringpromoted by FT translocation from leaves to shoot apices throughthe graft junction (Fig. 6b–d). In agreement with the specific roleof NaKR1 in mediating FT movement in the phloem stream,nakr1-1 only delays flowering promoted by FT expression in thephloem, but does not affect flowering triggered by FT expressiondirectly in the shoot apical meristem (Fig. 5a).
We envisage that NaKR1 function in mediating FT transport ispossibly relevant to NaKR1 participation in long-distance transportof other molecules in the phloem. A previous study has shown thatnakr1-1 displays several phenotypes pertaining to its defects inphloem function, including compromised long-distance transportof sucrose20. We have found that the sucrose levels are greatlyreduced in shoot apices of nakr1-1 compared with wild-typeplants during the floral transition (Supplementary Fig. 11). This isconsistent with the previous finding showing starch overaccumula-tion in nakr1-1 leaves and a reduced ability of translocating14C-sucrose in nakr1-1 (ref. 20), suggesting that NaKR1 playsan important role in transporting sucrose from leaves to othersink tissues. As the major sugar in both leaf and apical exudates,it was early hypothesized that sucrose is part of the florigenicstimuli that serves as a transmissible flowering signal inresponse to photoperiod35. Recent studies have also revealed thattrehalose-6-phospohate (T6P) derived from sucrose could play acentral role in mediating regulation of flowering time in bothleaves and shoot apical meristems36. Thus, NaKR1 could simul-taneously regulate FT and sucrose transport in the phloem, thusensuring that the supply of carbohydrates is coordinated with theenergy-demanding flowering process occurring in the shootapical meristem.
As NaKR1-like proteins are present in various flowering plants(Supplementary Fig. 12), it will be interesting to investigatewhether the mechanism revealed in this study is applicable to trans-port of FT orthologues in other flowering plants. In addition, as amember in the HPP family, NaKR1 acts in multiple developmentalprocesses relevant to phloem function20. Our yeast two-hybrid assayhas shown that only the HMA domain of NaKR1 interacts with FT(Fig. 3a), indicating that interaction between NaKR1 and FT mightbe closely relevant to HMA function in binding metallic ions in thephloem. Further elucidation of the metal binding capacity of HMAin NaKR1 will not only unravel other unknown mechanisms rel-evant to long-distance transport of flowering stimuli in responseto environmental signals, but also shed light on the molecular func-tion of the members in the HPP family.
MethodsPlant materials and growth conditions. Arabidopsis thaliana ecotypes Columbia(Col) and Landsberg erecta (Ler) were grown under long-day conditions (16-h light/8-h dark) or short day conditions (8-h light/16-h dark) at 23 °C. The mutants nakr1-1,ft-10, ftip1-1, gi-1, soc1-2, flc-3, svp-41, fld-3, flk-1, fve-4 and co-1 are in the Colbackground, whereas fca-1, fpa-1 and fve-1 mutants as well as co-2 35S:CO-GRtransgenic plants are in the Ler background. Transgenic plants of 35S:FT, SUC2:FT,KNAT1:FT, SUC2:FT–GFP and SUC2:FT–9myc have been described previously14.All the other transgenic plants were generated in the Col background by floraldipping with Agrobacterium tumefaciens and selected by Basta on soil37.
Plasmid construction. To construct 35S:NaKR1, SUC2:NaKR1 or KNAT1:NaKR1,the coding region of NaKR1 was amplified and cloned into pGreen–35S, pGreen–SUC2 or pGreen–KNAT1, respectively14,38. For the complementation test, a 5.5-kbNaKR1 genomic fragment ( gNaKR1) was amplified, and the resulting PCR fragmentwas digested and cloned into pHY105 (ref. 38). Based on this construct, thegNaKR1–GUS, gNaKR1–4HA and gNaKR1–GFP plasmids were generated byΩ-PCR39. To construct 35S:NaKR1–GFP, the cDNA encoding GFP was amplifiedand cloned into pGreen-35S (ref. 38) to generate pGreen-35S–GFP. The cDNAencoding NaKR1 was subsequently amplified and cloned into pGreen-35S–GFP.
ARTICLES NATURE PLANTS DOI: 10.1038/NPLANTS.2016.75
NATURE PLANTS | VOL 2 | JUNE 2016 | www.nature.com/natureplants8
© 2016 Macmillan Publishers Limited. All rights reserved

To construct AmiR-NaKR1, design of the artificial microRNA was performed usingthe software on the website (http://wmd3.weigelworld.org/cgi-bin/webapp.cgi?page=Home;project=stdwmd). Based on the NaKR1 sequence, a set of four primerswere generated and used for the PCR amplification according to the publishedprotocol27. The resulting PCR fragment was digested with EcoRI and BamHI, andcloned into pGreen-35S. To construct the vectors for yeast two-hybrid assays, thecoding region of FTIP1 was amplified and cloned into pGBKT7 (Clontech). Primersused for plasmid construction are listed in Supplementary Table 1.
Expression analysis. Total RNAwas isolated using FavorPrep Plant Total RNAminiKit (Favorgen) and used for reverse transcription with M-MLV ReverseTranscriptase (Promega) according to the manufacturer’s instructions. Quantitativereal-time PCR was performed in triplicates on an ABI PRISM 7900HT sequencedetection system (Applied Biosystems) with the Maxima SYBR Green/ROX qPCRMaster Mix (Fermentas). The relative expression levels were determined andnormalized against TUB2 expression as previously described40. Primers used forgene expression analysis are listed in Supplementary Table 1. GUS staining wasperformed as previously reported41.
ChIP assays. Seedlings were collected and then fixed on ice for 30 min in 1%formaldehyde under vacuum. Chromatin was isolated and sonicated to producesmall DNA fragments of 200 to 500 bp. CO–6HA fusion protein wasimmunoprecipitated by anti-HA agarose conjugate (Sigma). Enrichment fold ofeach fragment was calculated as previously reported40 first by normalizing the amountof a target DNA fragment against a genomic fragment of TUB2 as an internal controland then by normalizing the value for SUC2:CO–6HA against that for wild-typeplants. Primer pairs used for ChIP assays are listed in Supplementary Table 1.
BiFC analysis. The full-length coding regions of NaKR1 and FT were cloned intoprimary pSAT1 vectors. The resulting cassettes including fusion proteins and theconstitutive promoters were cloned into pHY105 and transformed intoAgrobacterium tumefaciens. The Agrobacteria were coinfiltrated into Nicotianabenthamiana leaves as previously reported42. The leaves were examined 2 days afterinfiltration under a confocal microscope.
Immunogold transmission electron microscopy. Immunoelectron microscopy wasperformed as previously described14,43. Plants were fixed in paraformaldehyde-glutaraldehyde solution (2 and 2.5%, respectively) overnight at room temperatureand imbedded in LR white resin (EMS). Ultrathin sections were mounted on nickelgrids, then transferred onto blocking drops of TTBS (20 mM Tris, 500 mM NaCl,and 0.05% Tween-20, pH 7.5) supplemented with 1% (w/v) BSA for 30 min andsubsequently incubated with anti-HA or anti-myc antibody at 1:5 (v/v) dilution ofblocking solution for 1 h at room temperature. After washing with TTBS thrice,grids were incubated for 30 min with 15 nm gold-conjugated goat anti-mouseantibody (EMS) at 1:20 (v/v) dilution of blocking solution. The grids weresubsequently washed with TTBS and distilled water. Tissue staining was carried outwith 2% uranyl acetate for 15 min at room temperature, and photos were taken bytransmission electron microscope (Jeol JEM-1230).
To quantitatively analyse immunogold labelling, electron micrographs ofrandomly photographed immunogold-labelled transverse sections were digitized,whereas the number of gold particles and the cell area were calculated by ImageJ. Forcalculating the density of gold particles throughout the projected cell area, weanalysed 60 individual sections from six different plants of each genotype. Theobtained data were presented as the mean number of gold particles per µm2 plusor minus standard deviation and statistically evaluated by a two-tailed pairedStudent’s t test.
Cot-grafting experiments. The Cot-grafting surgery was carried out based on apublished protocol33 using 4-day-old plants grown on MS medium under long-dayconditions under a dissection microscope (Nikon). After the surgery, grafted plantsin Petri dishes recovered for 6 days under long-day conditions. Successfully graftedplants were selected for further investigation. For analysis of flowering time, graftedplants were transferred onto soil under long-day conditions.
Statistics. Statistical analyses were performed using a two-tailed pairedStudent’s t test.
Received 29 October 2015; accepted 26 April 2016;published 27 May 2016
References1. Knott, J. E. Effect of localized photoperiod on spinach. Proc. Am. Soc. Hortic. Sci.
31, 152–154 (1934).2. Chailakhyan, M. K. New facts in support of the hormonal theory of plant
development. Compt. Rend. Acad. Sci. URSS 13, 79–83 (1936).3. Corbesier, L. et al. FT protein movement contributes to long-distance signaling
in floral induction of Arabidopsis. Science 316, 1030–1033 (2007).4. Jaeger, K. E. &Wigge, P. A. FT protein acts as a long-range signal in Arabidopsis.
Curr. Biol. 17, 1050–1054 (2007).
5. Mathieu, J., Warthmann, N., Küttner, F. & Schmid, M. Export of FT proteinfrom phloem companion cells is sufficient for floral induction in Arabidopsis.Curr. Biol. 17, 1055–1060 (2007).
6. Tamaki, S., Matsuo, S., Wong, H. L., Yokoi, S. & Shimamoto, K. Hd3a protein isa mobile flowering signal in rice. Science 316, 1033–1036 (2007).
7. Lin, M.-K. et al. FLOWERING LOCUS T protein may act as the long distanceflorigenic signal in the cucurbits. Plant Cell 19, 1488–1506 (2007).
8. Lifschitz, E. et al. The tomato FT ortholog triggers systemic signals that regulategrowth and flowering and substitute for diverse environmental stimuli.Proc. Natl Acad. Sci. USA 103, 6398–6403 (2006).
9. Wigge, P. A. et al. Integration of spatial and temporal information during floralinduction in Arabidopsis. Science 309, 1056–1059 (2005).
10. Suarez-Lopez, P. et al. CONSTANSmediates between the circadian clock and thecontrol of flowering in Arabidopsis. Nature 410, 1116–1120 (2001).
11. An,H. et al.CONSTANS acts in the phloem to regulate a systemic signal that inducesphotoperiodic flowering of Arabidopsis. Development 131, 3615–3626 (2004).
12. Samach, A. et al. Distinct roles of CONSTANS target genes in reproductivedevelopment of Arabidopsis. Science 288, 1613–1616 (2000).
13. Takada, S. & Goto, K. TERMINAL FLOWER2, an Arabidopsis homolog ofHETEROCHROMATIN PROTEIN1, counteracts the activation ofFLOWERING LOCUS T by CONSTANS in the vascular tissues of leaves toregulate flowering time. Plant Cell 15, 2856–2865 (2003).
14. Liu, L. et al. FTIP1 is an essential regulator required for florigen transport.PLoS Biol. 10, e1001313 (2012).
15. Abe, M. et al. FD, a bZIP protein mediating signals from the floral pathwayintegrator FT at the shoot apex. Science 309, 1052–1056 (2005).
16. Kardailsky, I. et al. Activation tagging of the floral inducer FT. Science 286,1962–1965 (1999).
17. Kobayashi, Y., Kaya, H., Goto, K., Iwabuchi, M. & Araki, T. A pair of relatedgenes with antagonistic roles in mediating flowering signals. Science 286,1960–1962 (1999).
18. Tehseen, M., Cairns, N., Sherson, S. & Cobbett, C. S. Metallochaperone-likegenes in Arabidopsis thaliana. Metallomics 2, 556–564 (2010).
19. de Abreu-Neto, J. B., Turchetto-Zolet, A. C., de Oliveira, L. F. V., BodaneseZanettini, M. H. & Margis-Pinheiro, M. Heavy metal-associated isoprenylatedplant protein (HIPP): characterization of a family of proteins exclusive to plants.FEBS J. 280, 1604–1616 (2013).
20. Tian, H. et al. Arabidopsis NPCC6/NaKR1 is a phloem mobile metal bindingprotein necessary for phloem function and root meristem maintenance.Plant Cell 22, 3963–3979 (2010).
21. Suzuki, N., Yamaguchi, Y., Koizumi, N. & Sano, H. Functional characterizationof a heavy metal binding protein CdI19 from Arabidopsis. Plant J. 32,165–173 (2002).
22. Gao, W., Xiao, S., Li, H.-Y., Tsao, S.-W. & Chye, M.-L. Arabidopsis thaliana acyl-CoA-binding protein ACBP2 interacts with heavy-metal-binding farnesylatedprotein AtFP6. New Phytol. 181, 89–102 (2009).
23. Barth, O., Vogt, S., Uhlemann, R., Zschiesche, W. & Humbeck, K. Stress inducedand nuclear localized HIPP26 from Arabidopsis thaliana interacts via its heavymetal associated domain with the drought stress related zinc finger transcriptionfactor ATHB29. Plant Mol. Biol. 69, 213–226 (2009).
24. Zhang, C., Barthelson, R. A., Lambert, G. M. & Galbraith, D. W. Globalcharacterization of cell-specific gene expression through fluorescence-activatedsorting of nuclei. Plant Physiol. 147, 30–40 (2008).
25. Brady, S. M. et al. A high-resolution root spatiotemporal map reveals doexpression patterns. Science 318, 801–806 (2007).
26. Deeken, R. et al. Identification of Arabidopsis thaliana phloem RNAs providesa search criterion for phloem-based transcripts hidden in complex datasets ofmicroarray experiments. Plant J. 55, 746–759 (2008).
27. Schwab, R., Ossowski, S., Riester, M., Warthmann, N. & Weigel, D. Highlyspecific gene silencing by artificial microRNAs in Arabidopsis. Plant Cell 18,1121–1133 (2006).
28. Hou, X. et al.Nuclear factor Y-mediated H3K27me3 demethylation of the SOC1locus orchestrates flowering responses of Arabidopsis. Nature Commun. 5,4601 (2014).
29. Tiwari, S. B. et al. The flowering time regulator CONSTANS is recruited to theFLOWERING LOCUS T promoter via a unique cis-element. New Phytol. 187,57–66 (2010).
30. Imlau, A., Truernit, E. & Sauer, N. Cell-to-cell and long-distance trafficking ofthe green fluorescent protein in the phloem and symplastic unloading of theprotein into sink tissues. Plant Cell 11, 309–322 (1999).
31. Nelson, B. K., Cai, X. & Nebenführ, A. A multicolored set of in vivo organellemarkers for co-localization studies in Arabidopsis and other plants. Plant J. 51,1126–1136 (2007).
32. Yamaguchi, A., Kobayashi, Y., Goto, K., Abe, M. & Araki, T. TWIN SISTER OFFT (TSF) acts as a floral pathway integrator redundantly with FT. Plant CellPhysiol. 46, 1175–1189 (2005).
33. Yoo, S. J., Hong, S. M., Jung, H. S. & Ahn, J. H. The cotyledons produce sufficientFT protein to induce flowering: evidence from cotyledon micrografting inArabidopsis. Plant Cell Physiol. 54, 119–128 (2013).
NATURE PLANTS DOI: 10.1038/NPLANTS.2016.75 ARTICLES
NATURE PLANTS | VOL 2 | JUNE 2016 | www.nature.com/natureplants 9
© 2016 Macmillan Publishers Limited. All rights reserved

34. Notaguchi, M. et al. Long-distance, graft-transmissible action of ArabidopsisFLOWERING LOCUS T protein to promote flowering. Plant Cell Physiol. 49,1645–1658 (2008).
35. Bernier, G., Havelange, A., Houssa, C., Petitjean, A. & Lejeune, P. Physiologicalsignals that induce flowering. Plant Cell 5, 1147–1155 (1993).
36. Wahl, V. et al. Regulation of flowering by trehalose-6-phosphate signaling inArabidopsis thaliana. Science 339, 704–707 (2013).
37. Clough, S. J. & Bent, A. F. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 16, 735–743 (1998).
38. Liu, C. et al. Specification of Arabidopsis floral meristem identity by repression offlowering time genes. Development 134, 1901–1910 (2007).
39. Chen, L., Wang, F., Wang, X. & Liu, Y.-G. Robust one-tube Ω-PCR strategyaccelerates precise sequence modification of plasmids for functional genomics.Plant Cell Physiol. 54, 634–642 (2013).
40. Li, D. et al. A repressor complex governs the integration of flowering signals inArabidopsis. Dev. Cell 15, 110–120 (2008).
41. Yu, H., Yang, S. H. & Goh, C. J. DOH1, a class 1 knox gene, is required formaintenance of the basic plant architecture and floral transition in orchid.Plant Cell 12, 2143–2159 (2000).
42. Sparkes, I. A., Runions, J., Kearns, A. & Hawes, C. Rapid, transient expression offluorescent fusion proteins in tobacco plants and generation of stablytransformed plants. Nature Protoc. 1, 2019–2025 (2006).
43. Wilson, S. M. & Bacic, A. Preparation of plant cells for transmission electronmicroscopy to optimize immunogold labeling of carbohydrate and proteinepitopes. Nature Protoc. 7, 1716–1727 (2012).
AcknowledgementsWe thank J. M. Ward for providing nakr1-1 seeds. This work was supported by AcademicResearch Funds (MOE2011-T2-2-008) from the Ministry of Education, Singapore, theSingapore National Research Foundation Investigatorship Programme (NRF-NRFI2016-02)and intramural research support from National University of Singapore and Temasek LifeSciences Laboratory.
Author contributionsY.Z. and H.Y. conceived and designed the study. Y.Z., L.L. and L.S. performed theexperiments. Y.Z., L.S. and H.Y. analysed data and wrote the paper.
Additional informationSupplementary information is available online. Reprints and permissions information isavailable online atwww.nature.com/reprints. Correspondence and requests formaterials shouldbe addressed to H.Y.
Competing interestsThe authors declare no competing financial interests.
ARTICLES NATURE PLANTS DOI: 10.1038/NPLANTS.2016.75
NATURE PLANTS | VOL 2 | JUNE 2016 | www.nature.com/natureplants10
© 2016 Macmillan Publishers Limited. All rights reserved

![Floral Induction in Arabidopsis by FLOWERING LOCUS T ... · stored using the transgene pGAS1::FT (GALACTOL SYNTHASE1 [GAS1] promoter fused to FT), which is active only in phloem companion](https://img.dokumen.tips/doc/110x75/5fe346abeba6c44579738d9a/floral-induction-in-arabidopsis-by-flowering-locus-t-stored-using-the-transgene.jpg)









![CENTRORADIALIS Interacts with FLOWERING LOCUS T-Like … · CENTRORADIALIS Interacts withFLOWERING LOCUS T-Like Genes to Control Floret Development and Grain Number1[OPEN] Xiaojing](https://img.dokumen.tips/doc/110x75/604e0934c680ad0da9122568/centroradialis-interacts-with-flowering-locus-t-like-centroradialis-interacts-withflowering.jpg)










![The Chickpea Early Flowering 1 Efl1)LocusThe Chickpea Early Flowering 1 (Efl1)Locus Is an Ortholog of Arabidopsis ELF31[OPEN] Stephen Ridge,a,2 Amit Deokar,a Robyn Lee,b Ketema Daba,a](https://img.dokumen.tips/doc/110x75/610adfb62252a72b375ba4d8/the-chickpea-early-flowering-1-ei1-the-chickpea-early-flowering-1-ei1locus.jpg)






