Embed Size (px)
Citation preview

INFECTION AND IMMUNITY,0019-9567/01/$04.0010 DOI: 10.1128/IAI.69.4.2596–2603.2001
Apr. 2001, p. 2596–2603 Vol. 69, No. 4
Copyright © 2001, American Society for Microbiology. All Rights Reserved.
Mycobacterium bovis BCG-Induced Granuloma FormationDepends on Gamma Interferon and CD40 Ligand but
Does Not Require CD28LAURA H. HOGAN,* WES MARKOFSKI, ANJA BOCK,† BRITTANY BARGER, JAMES D. MORRISSEY,
AND MATYAS SANDOR
Department of Pathology and Laboratory Medicine, University of Wisconsin Medical School, Madison, Wisconsin
Received 28 September 2000/Returned for modification 6 December 2000/Accepted 4 January 2001
Progressive granuloma formation is a hallmark of chronic mycobacterial infection. Granulomas are local-ized, protective inflammatory reactions initiated by CD41 T cells, which contribute to control of bacterialgrowth and blockade of bacterial dissemination. In order to understand the costimulatory requirements thatallow CD41 T cells to directly or indirectly induce granulomas, we studied granuloma formation after 6 weeksin Mycobacterium bovis BCG-infected CD28- and CD40 ligand (CD40L)-deficient mice and compared it togranuloma formation in infected wild-type inbred mice and infected cytokine-deficient mice. We characterizedgranulomas morphologically in liver sections, analyzed granuloma infiltrating cells by flow cytometry, andmeasured cytokine production by cultured granuloma cells. CD28-deficient mice have no defect at the localinflammatory site, inasmuch as they form protective granulomas and control bacterial growth. However, thereare fewer activated T cells in the spleen compared to infected wild-type animals, and quantitative differencesin the cellular composition of the granuloma are observed by flow cytometry. In CD40L-deficient mice, thegranuloma phenotype is very similar to the phenotype in gamma interferon (IFN-g)-deficient mice. BothIFN-g-deficient and CD40L-deficient mice form granulomas which prevent bacterial dissemination, but con-trol of bacterial growth is significantly impaired. The relative proportion of CD41 T cells in granulomas fromboth CD282/2 and CD40L2/2 mice is significantly decreased compared with wild-type animals. Both modelsdemonstrate that the phenotype and activation stage of systemic T cells do not always correlate with thephenotype and activation stage of the localized granulomatous response.
Granuloma formation around infected macrophage is a de-fining cellular response to mycobacterium infections. Layers ofextracellular matrix enclose a microenvironment of infectedcells and an intense inflammatory infiltrate. Granulomas elim-inate bacteria and also protect surrounding host tissue fromdestructive inflammatory responses. Without granuloma for-mation, mycobacterial infections can become widely dissemi-nated and frequently lethal, as occurs in human AIDS-associ-ated tuberculosis or in the infection of SCID or recombinase-activating gene-deficient mice (22, 27, 33, 34). The involvementof T lymphocytes in initiation, regulation, and resolution ofgranuloma formation has been well documented for both hu-man and murine infections (18, 22).
Murine models of Mycobacterium tuberculosis and Mycobac-terium bovis infection have been used to study the role ofcytokine regulatory networks in T lymphocyte-macrophage in-teractions (11). Mycobacterial infections induce a Th1-typeT-cell response in which gamma interferon (IFN-g) and tumornecrosis factor alpha (TNF-a) play crucial roles in granulomaformation and function (6, 9, 21, 23, 30, 39). In contrast to theextensive characterization of cytokine requirements for protec-tive granuloma formation, the study of essential T-cell co-stimulatory signaling is relatively new. The role of CD28 hasnot been studied, and the role of CD40-CD40 ligand (CD40L)
interactions in protection against mycobacterial infection iscontroversial. One recent study reported that the CD40L-me-diated pathway of T-cell activation is dispensable for resistanceto M. tuberculosis infection (4). This was in contrast to theenhanced susceptibility to infection observed clinically in pa-tients with a defective CD40L gene (2, 45) and to other exper-imental infections with intracellular pathogens, includingLeishmania (3, 20, 43).
In this study, we examined the role of CD28 and the role ofCD40-CD40L for M. bovis bacille Calmette-Guerin (BCG)-induced protective granuloma formation. We took advantageof the availability of both CD28 and CD40L gene knockoutmice to compare their roles in generating the cytokine milieurequired for effective granulomatous immune responses. Ad-ditionally, we focused on characterizing the effector T-lympho-cyte activation phenotype and function in the granuloma in theabsence of specific costimulatory molecules and correlated thatwith control of bacterial dissemination and formation of effec-tive granulomas as assessed by quantitative histopathology.Here we report that there is a striking change in the granulomaphenotype of CD40L knockout mice after intraperitoneal in-fection with BCG. Our data suggest that CD40L costimulatoryfunction, but not CD28 function, is essential for the generationof IFN-g levels adequate for the control of mycobacterial rep-lication at the local inflammatory site. This is consistent with amodel in which CD40L-mediated interactions are essential forthe induction of a strong Th1 immune response to intracellularpathogens. We show that both CD40L-CD40 and CD28-B7interactions influence the cellular composition of granulomas
* Corresponding author. Mailing address: Room 5580 MSC, 1300University Ave., Madison, WI 53706. Phone: (608) 262-2577. Fax:(608) 265-3301. E-mail: [email protected].
† Present address: University of Heidelberg, Heidelberg, Germany.
2596
on January 29, 2019 by guesthttp://iai.asm
.org/D
ownloaded from

and demonstrate that the systemic T-cell response does notclosely correlate with the T-cell response detected in the localinflammatory lesion.
MATERIALS AND METHODS
Animals. In these studies we used C57BL/6, IFN-g gene-deficient, TNF-agene deficient, CD28 gene-deficient, and CD40L gene-deficient strains of micepurchased from Jackson Labs. Animals were housed at animal facilities at boththe University of Wisconsin Medical School and the William S. MiddletonMemorial Veterans Administration Hospital. Both facilities are accredited andmeet Public Health Service policy.
M. bovis BCG infections. BCG (substrain Pasteur, from G. Fennelly) wasgrown in Middlebrook 7H9 (Difco Laboratories, Detroit, Mich.) with 0.05%Tween 80 and 10% oleic acid-dextrose-catalase (OADC) (Difco) supplementand stored in frozen aliquots at 270°C. For infections, ampoules were thawed,and the inoculum was diluted in saline plus 0.05% Tween 80 and briefly exposedto sonic oscillation in order to obtain a single-cell suspension. Mice were infectedintraperitoneally with 107 BOG in 100 ml (12) in order to maximize the produc-tion of liver granulomas. The dose injected is not lethal and induces a diseasethat is cleared with time. Infection was verified by histology of liver tissuesamples. We chose liver granulomas for analysis because they can be isolated inlarger numbers free of surrounding tissue than lung granulomas.
Histology. Small pieces of liver were fixed in 10% formalin prior to beingimbedded in paraffin for thin sectioning (8 to 10 mm). Hematoxylin and eosinstaining and Ziehl-Neelsen staining for acid-fast bacteria were done by theDepartment of Pathology’s Histopathology Service, University of Wisconsin.Quantitative studies were performed by direct microscopic examination using aOlympus reticular eyepiece containing a 10 by 10 grid. Liver granuloma burdenis the number of granulomas per grid in a field under 1003 total magnification.Granuloma size is the number of grid squares covered at 4003 total magnifica-tion. Bacteria per lesion is the number of acid-fast rods visible per granuloma at1,0003 total magnification using an oil immersion lens. Data are presented as themean 6 standard error of the mean for a minimum of 30 counts per mouse liversection. The number of individual mice is indicated in the figure legends.
Isolation of splenocytes and granuloma-infiltrating cells. Isolation of granu-lomas is described in references 29, 36, and 38. Spleens were removed asepticallyfrom 8- to 16-week-old mice, and viable cells were separated by centrifugationthrough Lympholyte M solution (Cedarlane Laboratories, Hornsby, Ontario,Canada) as described (35).
Flow cytometry and antibodies. Splenocytes or granuloma cell suspensionswere incubated for 30 min at 4°C with different labeled antibodies at saturationand then washed and analyzed. Unlabeled anti-Fc receptor antibody 2.4G2 (50mg/ml) was used to block nonspecific binding of Fc receptors. Cell surfacestaining on 10,000 events was measured by fluorescence-activated cell sort-ing (FACS) on a FACS Calibur (Becton Dickinson) and analyzed using theCellquest computer program (Power Macintosh version 3.0; Becton Dickinson).
Monoclonal antibodies used were purified from hybridoma cell lines obtainedfrom the American Type Culture Collection (Rockville, Md.) as indicated. Hy-bridoma cells were cultured in HB-101 serum-free medium, and the antibodieswere precipitated from supernatants by 45% saturated ammonium sulfate. Thepurified antibodies were labeled with biotin, fluorescein isothiocyanate, or Cy5(Amersham Pharmacia Biotech, Inc., Piscataway, N.J.). Antibodies used in-cluded those specific for murine CD4 (GK1.5), CD8 (53-6.7), MAC-1 (MI/70.15), CD44 (Pgp-1) (IM 7.8.1), L-selectin (CD62L) (Mel-14), and NK cells(DX5). Monoclonal antibodies against murine T-cell receptor a/b (a/b TCR)and CD4 were from Sigma Chemical Co. (St. Louis, Mo.) and against murinemajor histocompatibility complex (MHC) class II (aI-Ab) and NK cells werefrom Pharmingen (San Diego, Calif.).
Cytokine measurements. Samples for cytokine analysis were collected from106 liver granuloma cells or spleen cells (live by trypan blue exclusion) seededinto 96-well plates in 0.2 ml of complete medium and stimulated with 10 mg ofa-CD3 antibody per ml. After 72 h, cell culture supernatants were harvested andstored at 270°C until testing. Measurement of secreted IFN-g was made byenzyme-linked immunosorbent assay (ELISA) using standard methods. Briefly,plates were coated with anti-IFN-g capture antibody (Pharmingen). Serial two-fold dilutions of either murine recombinant IFN-g (rIFN-g) (Genzyme, Cam-bridge, Mass.) or test supernatants were added to triplicate wells. Bound cyto-kine was detected with biotinylated anti-mouse IFN-g (Pharmingen) followed byavidin phosphatase (Molecular Probes, Eugene, Oreg.). Wells were developedwith 50 mM 4-methylumbelliferyl phosphate (Molecular Probes), and fluores-cence intensity was measured with an HTS7000 Bioassay reader (Perkin Elmer,
Foster City, Calif.). Units of IFN-g were calculated with reference to the rIFNgstandard. All data are expressed as the amount of secreted cytokine per 106 cells.
Organ load. Bacterial colony formation was determined by plating serial di-lutions of liver homogenates on Middlebrook 7H10 agar plates (Difco) supple-mented with 10% OADC and cycloheximide (10 mg/ml). Colonies were countedafter 3 weeks of incubation at 37°C.
RESULTS
Liver histopathology. To examine the T-cell costimulatoryrequirements for protective granuloma formation in the liver,we infected CD40L and CD28 gene-deficient mice with BCG.At 6 weeks, when a chronic infection had been established, wesacrificed the mice and examined thin liver sections for gran-uloma formation. Figure 1 shows that CD28-deficient mice canform adequate numbers of well-formed lesions similar to thosefound in C57BL/6 mice (compare Fig. 1B and 1A). Staining foracid-fast bacteria illustrates that the extent of infection foundwithin individual granulomas is also very similar to that seen inthe wild-type mice (compare 1L with 1K). This is in starkcontrast to the CD40L-deficient mice, which have substantiallymore bacteria present per lesion (Fig. 1M). These sectionswere compared to the histopathology observed after infectionof IFN-g- and TNF-a-deficient mice (Fig. 1D, E, I, J, N, andO). Both IFN-g and TNF-a are known to be essential forprotective immunity against mycobacterial infection. IFN-g-deficient mice have elevated numbers of bacteria presentwithin lesions, but there are well-formed granulomas and thesurrounding tissue is healthy (Fig. 1D, I, and N). In contrast,TNF-a-deficient mice form very large, poorly organized lesionswhich do not effectively contain the bacterial infection (Fig.1E, J, and O). Bacteria and infected macrophage are observedwhich have no surrounding inflammation. The extensive bac-terial dissemination and organ damage associated with thisresponse was repeatedly lethal before 6 weeks: the TNF-asection illustrated in Fig. 1 is from a 4-week infection. Thetissue pathology that we observed with the CD40L-deficientmice is hence very similar to that of the IFN-g-deficient mice(compare Fig. 1H to I and 1M to N). Both have good granu-loma structures but poor control of bacteria, suggesting thatCD40L2/2 mice may be deficient in IFN-g but provide suffi-cient TNF-a. Histopathology in CD282/2 mice is comparableto that in wild-type mice.
We rigidly quantified our observations by directly countinggranulomas, number of bacteria per granuloma, and averagesize of granulomas. Figure 2 shows that, on average, the gran-ulomas and the number of bacteria per lesion from CD28-deficient mice were the same as from C57BL/6 mice. Granu-lomas in CD40L-deficient mice are significantly larger andmore numerous than in the wild-type mice (P , 0.05, student’st test). Additionally, the number of bacteria per lesion shows athreefold increase in CD40L-deficient mice compared to wild-type and CD28-deficient mice. Granulomas from IFN-g-defi-cient mice are larger and more numerous than wild-typegranulomas and are very comparable to the granulomas ofCD40L-deficient mice. The number of bacteria per lesion inIFN-g-deficient mice is almost 13 times higher than the num-ber of bacteria in wild-type lesions. The data suggest thatIFN-g and CD40L but not CD28 are crucial for effective gran-uloma formation and bacterial killing. Data from TNF-a-defi-cient mice are not presented due both to the significantly
VOL. 69, 2001 T-CELL COSTIMULATION IN BCG INFECTIONS 2597
on January 29, 2019 by guesthttp://iai.asm
.org/D
ownloaded from
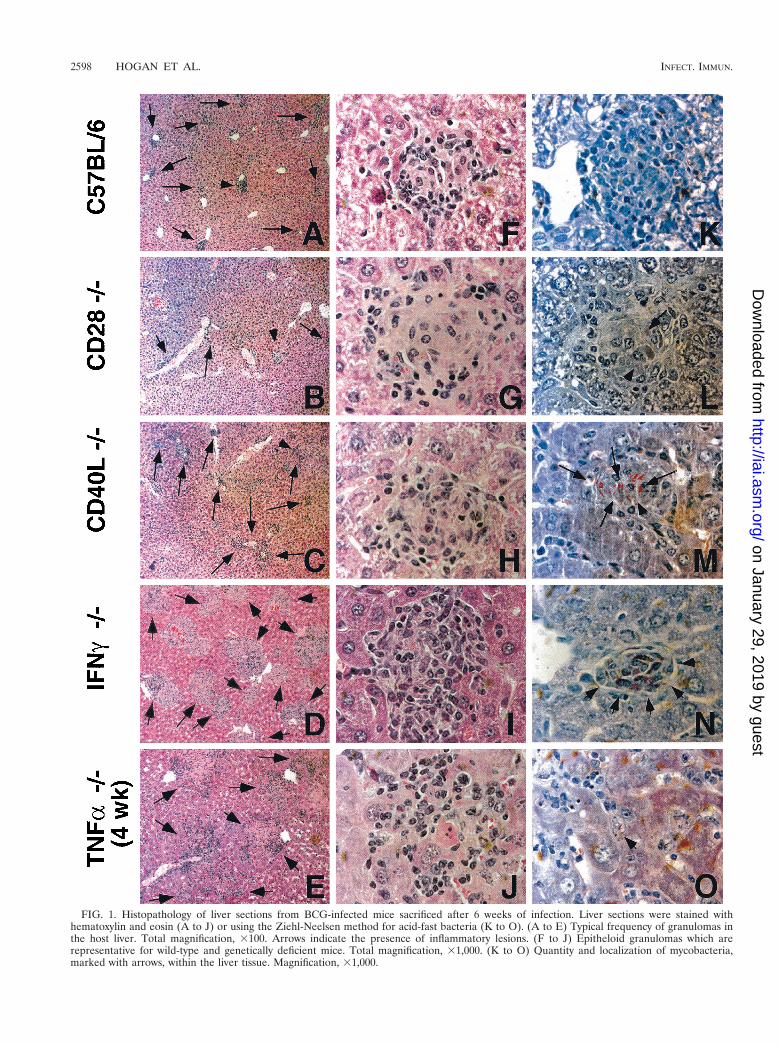
FIG. 1. Histopathology of liver sections from BCG-infected mice sacrificed after 6 weeks of infection. Liver sections were stained withhematoxylin and eosin (A to J) or using the Ziehl-Neelsen method for acid-fast bacteria (K to O). (A to E) Typical frequency of granulomas inthe host liver. Total magnification, 3100. Arrows indicate the presence of inflammatory lesions. (F to J) Epitheloid granulomas which arerepresentative for wild-type and genetically deficient mice. Total magnification, 31,000. (K to O) Quantity and localization of mycobacteria,marked with arrows, within the liver tissue. Magnification, 31,000.
2598 HOGAN ET AL. INFECT. IMMUN.
on January 29, 2019 by guesthttp://iai.asm
.org/D
ownloaded from

smaller sample size and to the fact that the lesions are sodisorganized as to make the comparison questionable. Identi-cal trends were seen in bacterial load measurements made byplating liver homogenates and calculating CFU per liver (Fig.2, last panel).
Comparison of T-cell accumulation and T-cell phenotypebetween local inflammatory site and spleen. Flow cytometrywas used to measure the phenotype and cell surface markerexpression of cell populations found in spleen and in livergranulomas after 6 weeks of BCG infection. Figure 3A showsthat CD40L-deficient mice have a significant reduction in therelative proportion of CD41 T cells present in granuloma-infiltrating cells compared to both C57BL/6 and CD28-defi-cient mice (third row compared to first and second rows).Combined CD44 and CD62L (L-selectin) antibody stainingindicates that activation (CD44 high, CD62L low) of both
CD41 and CD81 T cells is unimpaired. The absolute numberof CD41 T cells on a per-liver basis might be unchanged oreven larger, since Fig. 2 showed that CD40L-deficient micehave more and larger granulomas, but the CD41 T cells areproportionally fewer in the lesions. The lower level of MHCclass II molecules on macrophage populations likely indicatesa lower local concentration of IFN-g in the granulomatouslesion. That correlates well with the abolished bacterial controlshown in Fig. 1 and 2. We conclude that in the absence ofCD40L expression on T cells, insufficient numbers of activatedCD41 T cells capable of secreting IFN-g home to local inflam-matory sites. The phenotype of CD28-deficient granulomacells is almost indistinguishable from that of C57BL/6 granu-loma cells with the exception that a smaller percentage ofCD81 T cells is observed in both CD28-deficient mice andCD40L-deficient mice (first column, 24.8% versus 14.3% and15.2%, respectively).
A very different pattern of activation was observed when wemeasured the same parameters on splenocytes isolated fromthe different mouse strains infected with BCG. Splenocytesfrom infected CD40L-deficient mice have a modest decrease in
FIG. 2. Quantitative analysis of liver granuloma formation 6 weeksafter infection. Data in the top three panels are the means 6 standarderror of the mean of microscopic counts derived from liver sectionsfrom 12 wild-type, 12 CD28-deficient, 5 CD40L-deficient, and 5 IFN-g-deficient mice and represent a minimum of 30 counts per mouse.The last panel shows the mean CFU per liver 6 standard error of themean after growth of serial dilutions of organ homogenates on 7H10,medium for 3 weeks at 37°C. Statistical significance was analyzed bystudent’s t test. p, P , 0.05.
FIG. 3. FACS analysis of cell populations and activation phenotypeof T cells from mice chronically infected with BCG. Infected mice weresacrificed after 6 weeks, and granuloma cells (A) and splenocytes (B)were analyzed for CD41 T-cell, CD81 T-cell, and macrophage cellmarker expression. The first columns of plots represent CD41 andCD81 marker expression on cells from a lymphocyte gate, wherenumbers indicate the percent of gated cells. The middle two columnsshowing CD44 and CD62L expression are gated as indicated on theCD41 and CD81 T-cell populations shown in the left-hand column;numbers indicate the percentage of gated cells in the given quadrant.The right-hand column of histograms represents MHC class II expres-sion on a macrophage gate (Mac-11); numbers indicate the geometricmean of the staining intensity.
VOL. 69, 2001 T-CELL COSTIMULATION IN BCG INFECTIONS 2599
on January 29, 2019 by guesthttp://iai.asm
.org/D
ownloaded from

the fraction of CD41 T cells present, comparable activation ofCD41 and CD81 T cells relative to infected C57BL/6 spleno-cytes, and lowered MHC class II expression on macrophages.In contrast, splenocytes from infected CD28-deficient micehave an indistinguishable CD41-CD81 T-cell ratio but areprofoundly lacking in either CD41 or CD81 T-cell activationand have low MHC class II expression on macrophages relativeto splenocytes from infected C57BL/6 mice. The lack of acti-vated T cells found in splenocytes from CD28-deficient mice isvery different from the infected wild-type mice, in which a largeproportion of T cells are activated. Thus, although CD28-deficient mice are able to form good protective granulomasand control bacteria locally, systemic activation of T cells islargely impaired. These data clearly demonstrate that the studyof T-cell activation in a systemic site is not always predictive ofactivation at local lesions.
Interestingly, there is a lack of granuloma-recruited NK cellsand NK T cells in BCG-infected CD28-deficient mice, as mea-sured by flow cytometry using DX5- and a/b TCR-specificantibodies. Figure 4 shows the relative accumulation of thesecell types in C57BL/6 and CD28-deficient mouse granulomas.NK cell numbers in the spleen of CD28-deficient mice werejust slightly lower than in wild-type and CD40L2/2 mice (datanot shown). This suggests that although NK cells and NK Tcells may play a role in antimycobacterial control in granulo-mas, other mechanisms are able to compensate adequately fortheir absence.
IFN-g measurements. The capacity of BCG-induced granu-loma-infiltrating cells to produce IFN-g was measured directlyby ELISA of cell culture supernatants. As expected, theamount of IFN-g produced by granuloma cells from CD40L-deficient mice after 3 days of in vitro stimulation by a-CD3antibody was below the assay sensitivity and hence significantlylower (P , 0.05, Student’s t test) than levels produced bygranuloma cells from C57BL/6 or CD28-deficient mice (Fig.5A). IFN-g levels produced by CD28-deficient mouse spleno-cytes similarly stimulated with a-CD3 antibody were 5- to10-fold lower than those from CD40L-deficient or C57BL/6mouse stimulated splenocytes, respectively (Fig. 5B). Theoverall level of IFN-g detected in granuloma cell culture su-pernatants was less than in splenocyte culture supernatants.The comparable expression of MHC class II molecules onmacrophages from C57BL/6 infected animals in either com-
partment makes it unlikely that the difference reflects differ-ences in vivo (Fig. 3A and B). We must consider that althoughwe used similar cell numbers in culture, there may be a greaterfraction of T cells present in total splenocytes than in granu-loma cell preparations (;10 to 15%). Additionally, the viabil-ity of granuloma cells in culture may be lower than the viabilityof spleen cells.
DISCUSSION
The essential role of CD41 T cells in granuloma formationin response to intracellular and extracellular mycobacteria isundisputed. BCG infection of class II-deficient mice showedthat without CD41 T cells, infected and dying macrophagesrecruit a damaging neutrophilic inflammatory response that isunable to control bacterial replication (26). T cells, their di-rectly elaborated cytokines, and cytokines from activated mac-rophage are essential both for forming a protective structure tocontain the inflammatory response and for eradicating thesequestered bacteria. Mice deficient in IFN-g or cytokines thatregulate IFN-g levels, including Eta-1 (osteopontin) (1, 31),and interleukin-12 (IL-12) (7, 19), cannot control bacterial
FIG. 4. FACS analysis of NK cell populations from granuloma Tcells in mice chronically infected with BCG. Infected mice were sac-rificed after 6 weeks, and granuloma cells from either C57BL/6 orCD28-deficient mice were analyzed for DX5 and a/b TCR cell markerexpression. Staining analysis is shown from a lymphocyte gate wherenumbers indicate the percentage of gated cells in the given quadrant.
FIG. 5. IFN-g secreted by granuloma cells or spleen cells after invitro culture in the presence of a-CD3 antibody. (A) Granuloma cellsor (B) spleen cells isolated after 6 weeks of BCG infection by theindicated strains were cultured in vitro for 3 days in the presence ofa-CD3 (145-2C11) (5 mg/ml) to activate TCR-mediated signaling.IFN-g secreted into culture supernatants was measured by ELISA.Values shown are the averages of quadruplicate measurements fromone experiment, and error bars represent the standard error of themean. Statistical significance was analyzed by student’s t test. p, P ,0.05. Results shown are representative of three independent experi-ments.
2600 HOGAN ET AL. INFECT. IMMUN.
on January 29, 2019 by guesthttp://iai.asm
.org/D
ownloaded from

growth. In the absence of TNF-a, inflammation is uncontrolledand tissue-damaging abscesses are formed (10, 23, 39).
In this study we examined the costimulatory signal require-ments for activation of CD41 T cells to regulate sufficientproduction of IFN-g and TNF-a to form protective granulo-mas in response to chronic BCG infection. To our knowledge,this is the only study to directly examine the phenotype of cellsthat accumulate in BCG-induced granulomas and correlatethat phenotype to protective histopathology. In the absence ofcostimulatory molecules, our data demonstrated that there canbe substantial differences between splenic responses and localresponses at the granulomatous inflammatory site.
CD40L upregulation after TCR engagement is linked to acascade of events, including production of cytokines and che-mokines, significantly IFN-g and TNF-a in the case of myco-bacteria, and increased expression of accessory molecules onantigen-presenting cells. Recent studies have suggested thatCD28-mediated signals are not always required for resistanceto various infections, but instead are important for the optimalproduction of IL-2 and IFN-g and/or good memory responses(13, 25, 44). Autocrine production of IL-2 after initial CD28costimulation can sustain CD40L expression in the absence ofCD28 or TCR signaling (41). This may represent an importantmechanism for reactivating previously stimulated T cells in theabsence of antigen. Since our work has shown that CD28 andIL-2 are not essential for protective granuloma formation(data not shown), it seems likely that the requirement forCD40L expression in BCG-induced granuloma formation isnot dependent upon CD28 engagement and IL-2 signaling.Our data suggest an alternative model in which CD40L-medi-ated costimulation is required to produce sufficient IFN-g lo-cally. The evidence for this includes the qualitative and quan-titative similarity of histopathology between CD40L-deficientand IFN-g-deficient mice (Fig. 1 and 2). Additionally, IFN-glevels are greatly depressed in granuloma culture supernatantsfrom CD40L-deficient mice (Fig. 5), and MHC class II expres-sion is reduced on granuloma-infiltrating macrophages fromthese mice (Fig. 3), suggesting that the amount of IFN3-g inthe granuloma is not sufficient for normal macrophage activa-tion. A correlation between CD40L expression and IFN-g pro-duction after mycobacterial infection has also been shown byclinical studies of peripheral blood mononuclear cells (PB-MCs) from tuberculosis patients and healthy tuberculin reac-tors (37). It is also clear from our data that sufficient TNF-a isavailable without CD40L or CD28, as the extensive and dam-aging inflammation that results in early death in TNF-a-defi-cient animals is not seen (Fig. 1).
Our flow cytometric data also showed that in the absence ofCD40L, CD41 T-cell recruitment to the local inflammatorysite is substantially reduced (Fig. 3). However, there are acti-vated T cells in the spleens of BCG-infected CD40L-deficientmice. Since activated T cells are capable of homing to localinflammatory sites, it is likely that CD41 T-cell accumulationin the granuloma is regulated not only at the level of T-cellactivation, but also by other factors such as motility, extrava-sation, recruitment or chemotactic sensitivity. The cellularcomposition of granulomas in CD40L2/2 mice indicated thatrecruitment of both CD41 and CD81 T cells is altered.
Although previous immunohistochemical studies haveshown that human tuberculoid granulomatous lesions contain
readily detectable levels of B7-1 and B7-2 on epithelioid cellsand of CD28 on T cells (42), our data indicated that CD28 isdispensable for protective granulomatous responses againstBCG in a murine model. T cells from CD28-deficient micehave reduced expression of IL-2 receptor a(IL-2Ra), reducedT-helper cell function, and reduced proliferation to concanava-lin A but are still able to mount an effective anti-lymphocyticchoriomeningitis virus cytolytic immune response in vivo (40).Although resistance to primary infection by Schistosoma man-soni is impaired in CD28-deficient mice (24), and Th2 protec-tive responses are diminished, egg-induced granulomatous re-actions in the livers of these mice are unaffected. Similarly,B7-1/22/2 mice also form intact granulomas around schisto-some eggs despite limited T-cell proliferative capacity and askew towards Th1-type cytokine production (15).
The results of our study suggested that costimulation medi-ated by either CD40L or other accessory molecules can com-pensate for the lack of CD28 during the local response toBCG. In the microenvironment of the granuloma, local con-centrations of IL-15 and IL-12 may be sufficient to maintainCD40L expression (41) and drive IFN-g production (44) inde-pendent of CD28. Bacteria within granulomas may also altercostimulatory requirements relative to the periphery due tohigher local concentrations of antigen (25). Although CD28-mediated signaling was not required at the local inflammatorysite, the systemic response to BCG infection was much lower,perhaps reflecting either reduced systemic T-cell activation orcomplete recruitment of the few systemically activated T cellsto the local inflammatory site.
The mechanisms responsible for the somewhat lower pro-portions of CD81 T cells and the absence of NK cells and NKT cells found in granuloma cell populations from BCG-in-fected CD282/2 mice are not understood. Since NK cells areactivated by CD28 (5, 17), one explanation is that activation isrequired for NK cell recruitment to the granuloma. Our datasuggest that NK and NK T cells are not absolutely essential forprotective granuloma formation. Additionally, our study makes itclear that the peripheral spleen cell phenotype is not a surepredictor of either granuloma formation or granuloma-infil-trating cell phenotype and underscores the importance of ex-amining the granuloma cells directly.
Our data regarding the susceptibility of CD40L-deficientmice to BCG infection is contrary to the published reports ofCampos-Neto using intravenous infection with M. tuberculosis(4), but consistent with the overall agreement among infectionstudies regarding the role of CD40L (3, 8, 20, 43, 46, 48). Theresults of their study were surprising given that cell lines trans-fected with CD40L genes gain the ability to inhibit the growthof M. avium in human monocyte-derived macrophages in vitro(14). Samten et al. also presented data that CD40L disregula-tion contributes to reduced IFN-g production in PBMCs fromtuberculosis patients (37). It has been shown that BCG re-quires CD40L-CD40 interactions for the IL-12-regulated pro-duction of IFN-g from human macrophages (28). Skin lesionsfrom resistant tuberculoid leprosy patients contain elevatedlevels of CD40 and CD40L mRNA and protein compared tothose from susceptible lepromatous patients (47).
The experimental differences between our work and theprevious study include differences in the bacteria, the dosage,and the route of infection. Our experimental mice were in-
VOL. 69, 2001 T-CELL COSTIMULATION IN BCG INFECTIONS 2601
on January 29, 2019 by guesthttp://iai.asm
.org/D
ownloaded from

fected at high dose intraperitoneally with BCG for the rapidinduction of a chronic immune response and maximum livergranuloma formation. Villegas and coworkers (44) have re-ported differences in the requirement for CD40L betweenacute (CD40L independent) and chronic (CD40L dependent)stages of infection with Toxoplasma gondii. This differencemight be mediated by differences in the initially predominantantigen-presenting cell population encountered via alternativeinfection routes, given that recent reports have demonstratedvariable requirements for CD40-CD40L interactions in differ-ent subpopulations of antigen-presenting cells (32).
Delineating the requirement for CD40L-mediated costimu-lation for granuloma formation has important consequencesfor our approach to managing pathologic granulomatous dis-eases without an infectious etiology. In sarcoidosis and Crohn’sdisease, targeted immunotherapies to downregulate T-cellfunction may be beneficial. This type of approach is alreadybeing taken using anti-CD40L antibodies to treat autoimmunedemyelinating disease models in mice and has shown promis-ing results (16). Our data suggest that granuloma formation inresponse to BCG can compensate for the loss of CD28 func-tion and that CD40L expression plays an antimycobacterialprotective role via the regulation of IFN-g. Additionally, themodels strikingly show that the status of systemic responsesmay not be reflected at local inflammatory sites. Hence, thestudy of immune responses at local lesions, like granulomas,deserves significantly more attention.
ACKNOWLEDGMENTS
This work was supported by an R01 award (AI 48087-01) from theNational Institutes of Health, an American Lung Association award,and an institutional Howard Hughes award to M.S.
We thank Satoshi Kinoshita for his excellent histopathology services,Diane Sewell and Dominic Co for careful reading of the manuscript,and members of our laboratory for many helpful discussions.
REFERENCES
1. Ashkar, S., G. F. Weber, V. Panoutsakopoulou, M. E. Sanchirico, M. Jans-son, S. Zawaideh, S. R. Rittling, D. T. Denhardt, M. J. Glimcher, and H.Cantor. 2000. Eta-1 (osteopontin): an early component of type-1 (cell-me-diated) immunity. Science 287:860–864.
2. Callard, R. E., R. J. Armitage, W. C. Fanslow, and M. K. Spriggs. 1993.CD40 ligand and its role in X-linked hyper-IgM syndrome. Immunol. Today14:559–564.
3. Campbell, K. A., P. J. Ovendale, M. K. Kennedy, W. C. Fanslow, S. G. Reed,and C. R. Maliszewski. 1996. CD40 ligand is required for protective cell-mediated immunity to Leishmania major. Immunity 4:283–289.
4. Campos-Neto, A., P. Ovendale, T. Bement, T. A. Koppi, W. C. Fanslow, M. A.Rossi, and M. R. Alderson. 1998. CD40 ligand is not essential for thedevelopment of cell-mediated immunity and resistance to Mycobacteriumtuberculosis. J. Immunol. 160:2037–2041.
5. Chambers, B. J., M. Salcedo, and H. G. Ljunggren. 1996. Triggering ofnatural killer cells by the costimulatory molecule CD80 (B7–1). Immunity5:311–317.
6. Cooper, A. M., D. K. Dalton, T. A. Stewart, J. P. Griffin, D. G. Russell, andI. M. Orme. 1993. Disseminated tuberculosis in interferon gamma gene-disrupted mice. J. Exp. Med. 178:2243–2247.
7. Cooper, A. M., J. Magram, J. Ferrante, and I. M. Orme. 1997. Interleukin 12(IL-12) is crucial to the development of protective immunity in mice intra-venously infected with mycobacterium tuberculosis. J. Exp. Med. 186:39–45.
8. Cosyns, M., S. Tsirkin, M. Jones, R. Flavell, H. Kikutani, and A. R. Hay-ward. 1998. Requirement of CD40-CD40 ligand interaction for eliminationof Cryptosporidium parvum from mice. Infect. Immun. 66:603–607.
9. Dalton, D. K., S. Pitts-Meek, S. Keshav, I. S. Figari, A. Bradley, and T. A.Stewart. 1993. Multiple defects of immune cell function in mice with dis-rupted interferon-g genes. Science 259:1739–1742.
10. Ehlers, S., J. Benini, S. Kutsch, R. Endres, E. T. Rietschel, and K. Pfeffer.1999. Fatal granuloma necrosis without exacerbated mycobacterial growth intumor necrosis factor receptor p55 gene-deficient mice intravenously in-
fected with Mycobacterium avium. Infect. Immun. 67:3571–3579.11. Flesch, I. E., and S. H. Kaufmann. 1993. Role of cytokines in tuberculosis.
Immunobiology 189:316–339.12. Flynn, J. L., M. M. Goldstein, J. Chan, K. J. Triebold, K. Pfeffer, C. J.
Lowenstein, R. Schreiber, T. W. Mak, and B. R. Bloom. 1995. Tumor ne-crosis factor-a is required in the protective immune response against Myco-bacterium tuberculosis in mice. Immunity 2:561–572.
13. Gause, W. C., S. J. Chen, R. J. Greenwald, M. J. Halvorson, P. Lu, X. D.Zhou, S. C. Morris, K. P. Lee, C. H. June, F. D. Finkelman, J. F. Urban, andR. Abe. 1997. CD28 dependence of T cell differentiation to IL-4 productionvaries with the particular type 2 immune response. J. Immunol. 158:4082–4087.
14. Hayashi, T., S. P. Rao, P. R. Meylan, R. S. Kornbluth, and A. Catanzaro.1999. Role of CD40 ligand in Mycobacterium avium infection. Infect. Immun.67:3558–3565.
15. Hernandez, H. J., A. H. Sharpe, and M. J. Stadecker. 1999. Experimentalmurine schistosomiasis in the absence of B7 costimulatory molecules: rever-sal of elicited T cell cytokine profile and partial inhibition of egg granulomaformation. J. Immunol. 162:2884–2889.
16. Howard, L. M., A. J. Miga, C. L. Vanderlugt, M. C. Dal Canto, J. D. Laman,R. J. Noelle, and S. D. Miller. 1999. Mechanisms of immunotherapeuticintervention by anti-CD40L (CD154) antibody in an animal model of mul-tiple sclerosis. J. Clin. Investig. 103:281–290.
17. Hunter, C. A., L. Ellis-Neyer, K. E. Gabriel, M. K. Kennedy, K. H. Grab-stein, P. S. Linsley, and J. S. Remington. 1997. The role of the CD28/B7interaction in the regulation of NK cell responses during infection withToxoplasma gondii. J. Immunol. 158:2285–2293.
18. Infante-Duarte, C., and T. Kamradt. 1999. Th1/Th2 balance in infection.Springer Semin. Immunopathol. 21:317–338.
19. Jouanguy, E., R. Doffinger, S. Dupuis, A. Pallier, F. Altare, and J. L.Casanova. 1999. IL-12 and IFN-gamma in host defense against mycobacteriaand salmonella in mice and men. Curr. Opin. Immunol. 11:346–351.
20. Kamanaka, M., P. Yu, T. Yasui, K. Yoshida, T. Kawabe, T. Horii, T. Kishi-moto, and H. Kikutani. 1996. Protective role of CD40 in Leishmania majorinfection at two distinct phases of cell-mediated immunity. Immunity 4:275–281.
21. Kamijo, R., J. Le, D. Shapiro, E. A. Havell, S. Huang, M. Aguet, M. Bosland,and J. Vilcek. 1993. Mice that lack the interferon-g receptor have profoundlyaltered responses to infection with bacillus Calmette-Guerin and subsequentchallenge with lipopolysaccharide. J. Exp. Med. 178:1435–1440.
22. Kaufmann, S. H., and C. H. Ladel. 1994. Role of T cell subsets in immunityagainst intracellular bacteria: experimental infections of knock-out mice withListeria monocytogenes and Mycobacterium bovis BCG. Immunobiology191:509–519.
23. Kindler, V., A. P. Sappino, G. E. Grau, P. F. Piguet, and P. Vassalli. 1989.The inducing role of tumor necrosis factor in the development of bactericidalgranulomas during BCG infection. Cell 56:731–740.
24. King, C. L., J. Xianli, C. H. June, R. Abe, and K. P. Lee. 1996. CD28-deficient mice generate an impaired Th2 response to Schistosoma mansoniinfection. Eur. J. Immunol. 26:2448–2455.
25. Kundig, T. M., A. Shahinian, K. Kawai, H. W. Mittrucker, E. Sebzda, M. F.Bachmann, T. W. Mak, and P. S. Ohashi. 1996. Duration of TCR stimulationdetermines costimulatory requirement of T cells. Immunity 5:41–52.
26. Ladel, C. H., S. Daugelat, and S. H. Kaufmann. 1995. Immune response toMycobacterium bovis bacille Calmette-Guerin infection in major histocom-patibility complex class I- and II-deficient knock-out mice: contribution ofCD4 and CD8 T cells to acquired resistance. Eur. J. Immunol. 25:377–384.
27. Ledru, E., S. Ledru, and A. Zoubga. 1999. Granuloma formation and tuber-culosis transmission in HIV-infected patients. Immunol. Today 20:336–337.
28. Mendez-Samperio, P., H. E. Ayala-Verdin, and A. Trejo-Echeverria. 1999.Interleukin-12 regulates the production of Bacille Calmette-Guerin-inducedinterferon-gamma from human cells in a CD40-dependent manner. Scand. J.Immunol. 50:61–67.
29. Metwali, A., D. Elliott, A. M. Blum, J. Li, M. Sandor, R. G. Lynch, N.Noben-Trauth, and J. V. Weinstock. 1996. The granulomatous response inmurine schistomiasis mansoni does not switch to TH1 in IL-4-deficientC57BL/6 mice. J. Immunol. 157:4546–4553.
30. Murray, P. J., R. A. Young, and G. Q. Daley. 1998. Hematopoietic remod-eling in interferon-gamma-deficient mice infected with mycobacteria. Blood91:2914–2924.
31. Nau, G. J., P. Guilfoile, G. L. Chupp, J. S. Berman, S. J. Kim, H. Kornfeld,and R. A. Young. 1997. A chemoattractant cytokine associated with granu-lomas in tuberculosis and silicosis. Proc. Natl. Acad. Sci. USA 94:6414–6419.
32. Ozaki, M. E., B. A. Coren, T. N. Huynh, D. J. Redondo, H. Kikutani, andS. R. Webb. 1999. CD41 T cell responses to CD40-deficient APCs: defectsin proliferation and negative selection apply only with B cells as APCs. J.Immunol. 163:5250–5256.
33. Palmieri, F., A. M. Pellicelli, E. Girardi, A. P. De Felici, P. De Mori, N.Petrosillo, and G. Ippolito. 1999. Negative predictors of survival in HIV-infected patients with culture-confirmed pulmonary tuberculosis. Infection27:331–334.
34. Perlman, D. C., P. El-Helou, and N. Salomon. 1999. Tuberculosis in patients
2602 HOGAN ET AL. INFECT. IMMUN.
on January 29, 2019 by guesthttp://iai.asm
.org/D
ownloaded from

with human immunodeficiency virus infection. Semin. Respir. Infect. 14:344–352.
35. Reiner, S. L., Z. E. Wang, F. Hatam, P. Scott, and R. M. Locksley. 1993. TH1and TH2 cell antigen receptors in experimental leishmaniasis. Science 259:1457–1460.
36. Sacco, R. E., R. J. Jensen, C. O. Thoen, M. Sandor, J. Weinstock, R. G.Lynch, and M. O. Dailey. 1996. Cytokine secretion and adhesion moleculeexpression by granuloma T lymphocytes in mycobacterium avium infection.Am.J. Pathol. 148:1935–1948.
37. Samten, B., E. K. Thomas, J. Gong, and P. F. Barnes. 2000. Depressed CD40ligand expression contributes to reduced gamma interferon production inhuman tuberculosis. Infect. Immun. 68:3002–3006.
38. Sandor, M., A. I. Sperling, G. A. Cook, J. V. Weinstock, R. G. Lynch, andJ. A. Bluestone. 1995. Two waves of gamma delta T cells expressing differentV delta genes are recruited into schistosome-induced liver granulomas.J. Immunol. 155:275–284.
39. Senaldi, G., S. Yin, C. L. Shaklee, P. Piguet, T. W. Mak, and T. R. Ulich.1996. Corynebacterium parvum- and Mycobacterium bovis bacillusCalmette-Guerin-induced granuloma formation is inhibited in TNF receptor1 (TNF-R1) knockout mice and by treatment with soluble TNF-R1. J. Im-munol. 157:5022–5026.
40. Shahinian, A., K. Pfeffer, K. P. Lee, T. M. Kundig, K. Kishihara, A. Wake-ham, K. Kawai, P. S. Ohashi, C. B. Thompson, and T. W. Mak. 1993.Differential T cell costimulatory requirements in CD28-deficient mice. Sci-ence 261:609–612.
41. Skov, S., M. Bonyhadi, N. Odum, and J. A. Ledbetter. 2000. IL-2 and IL-15regulate CD154 expression on activated CD4 T cells. J. Immunol. 164:3500–3505.
42. Soler, P., V. Boussaud, J. Moreau, A. Bergeron, P. Bonnette, A. J. Hance,and A. Tazi. 1999. In situ expression of B7 and CD40 costimulatory mole-cules by normal human lung macrophages and epithelioid cells in tubercu-loid granulomas. Clin. Exp. Immunol. 116:332–339.
43. Soong, L., J. C. Xu, I. S. Grewal, P. Kima, J. Sun, B. J. Longley Jr., N. H.Ruddle, D. McMahonpratt, and R. A. Flavell. 1996. Disruption of CD40-CD40 ligand interactions results in an enhanced susceptibility to Leishmaniaamazonensis. Immunity 4:263–273.
44. Villegas, E. N., M. M. Elloso, G. Reichmann, R. Peach, and C. A. Hunter.1999. Role of CD28 in the generation of effector and memory responsesrequired for resistance to Toxoplasma gondii. J. Immunol. 163:3344–3353.
45. Vogel, L. A., and R. J. Noelle. 1998. CD40 and its crucial role as a memberof the TNFR family. Semin. Immunol. 10:435–442.
46. Wiley, J. A., and A. G. Harmsen. 1995. CD40 ligand is required for resolutionof Pneumocystis carinii pneumonia in mice. J. Immunol. 155:3525–3529.
47. Yamauchi, P. S., J. R. Bleharski, K. Uyemura, J. Kim, P. A. Sieling, A.Miller, H. Brightbill, K. Schlienger, T. H. Rea, and R. L. Modlin. 2000. Arole for CD40-CD40 ligand interactions in the generation of type 1 cytokineresponses in human leprosy. J. Immunol. 165:1506–1512.
48. Zhou, P., and R. A. Seder. 1998. CD40 ligand is not essential for inductionof type 1 cytokine responses or protective immunity after primary or sec-ondary infection with Histoplasma capsulatum. J. Exp. Med. 187:1315–1324.
Editor: J. D. Clements
VOL. 69, 2001 T-CELL COSTIMULATION IN BCG INFECTIONS 2603
on January 29, 2019 by guesthttp://iai.asm
.org/D
ownloaded from





























