Embed Size (px)
Citation preview

[CANCER RESEARCH 37, 4352-4360, December 1977]
Morphogenesis of Early 1,2-Dimethylhydrazine-induced Lesions
and Latent Period Reduction of Colon Carcinogenesis inMice by a Variant of Citrobacter freund//1
Stephen W. Barthold and Albert M. Jonas
Section of Comparative Medicine, Yale University School of Medicine, New Haven, Connecticut 06510
SUMMARY
The morphogenesis of 1,2-dimethylhydrazine (DMH)-in-duced lesions in the colon of outbred NIH Swiss mice wasdetermined for up to 5 months of treatment. The effect ofhyperplasia on DMH carcinogenesis was also evaluated byintroducing a transient hyperplastic stimulus to the colonduring the chronic weekly treatment regimen of DMH. Thehyperplastic stimulus was a naturally occurring disease ofmice, transmissible murine colonie hyperplasia, which iscaused by a variant of Citrobacter freundii. In control mice,those not receiving the bacterium, weekly injections of thecarcinogen induced neoplastic changes first detectable attwo months of treatment in all segments of the colon andin both sexes. The changes increased in frequency andseverity with time. Diffuse mucosal hyperplasia and chronicinflammatory and degenerative changes were also associated with DMH after prolonged treatment. The hyperplasticstimulus of C. freundii reduced the latent period for appearance of early DMH tumors, but it had no influence onalready established DMH tumors.
INTRODUCTION
Hyperplasia is believed to precede neoplasia of the colon.Evidence for this observation is based on the higher incidence and earlier age of onset of colonie cancer in patientswith diseases of the bowel with exaggerated proliferativeactivity, such as Crohn's disease, ulcerative colitis, and
familial polyposis (5, 12, 16, 20, 34), and the increasedproliferative activity seen in the flat mucosa between existing tumors of the colon (7, 18). Recently, it has beenshown experimentally that nonspecific mucosal injury tothe large bowel of rats causes mucosal proliferation thatenhances DMH2-induced neoplasia at the site of injury (26).
A unique opportunity exists to explore the interaction ofa hyperplastic stimulus and chemical carcinogenesis in themouse. Mice are susceptible to a natural infectious disease,TMCH, which is caused by a specific variant of Citrobacterfreundii (1). In TMCH there is severe although transientmucosal hyperplasia of the distal colon (2), which is alsothe site of predilection for DMH-induced neoplasia (13, 31,33). Colonie neoplasms induced by DMH or related com-
1 Supported by the National Large Bowel Cancer Project USPHS Research
Grant 5 R26 CA 15405, National Cancer Institute.2 The abbreviations used are: DMH, 1,2-dimethylhydrazine; TMCH, trans
missible murine colonie hyperplasia.Received June 24, 1977; accepted September 7, 1977.
pounds in mice have been described by several authors (6,8-10, 13, 24, 31, 37). A study of the interaction of TMCHand DMH would provide insight into the role of a temporaryalthough strong hyperplastic stimulus on DMH carcinogenesis.
The present experiment was designed to introduce TMCHat different times during a chronic weekly regimen of DMH.Since only a few reports (6, 13, 31, 32) have incompletelydescribed early neoplastic events in the mouse colon, thisstudy also sought to define the early DMH-induced neoplastic and nonneoplastic lesions in the colon of outbred NIHSwiss mice.
MATERIALS AND METHODS
Outbred NIH Swiss mice were obtained from a barrier-maintained production colony in the Division of AnimalCare, Yale University School of Medicine, New Haven,Conn. Mice were 21 to 35 days of age at the onset ofthe experiments and were fed Purina laboratory chowand water ad libitum. They were housed by sex in groupsof 6 in plastic boxes with pine chip bedding.
Experiment 1. Four hundred seventy-one mice were divided into 6 experimental groups (A to E and X), eachconsisting of one-half males and one-half females. Eachexperimental group was further divided into 2 subgroups.One subgroup was treated with weekly injections of 20 mgDMH (1,2-dimethylhydrazine dihydrochloride, Matheson,Coleman & Bell, Norwood, Ohio) per kg s.c. in 0.1 ml 0.001M EDTA, and the other subgroup was treated similarly withEDTA alone. Treatments continued until the animals werekilled for necropsy. Experimental Groups A to E were alsoinoculated with 2 to 3 drops p.o. of a thioglycolate brothculture of C. freundii (1) at different monthly intervalsduring the weekly DMH-EDTA or EDTA treatment regimen.Experimental Group X was not exposed to C. freundii andserved as DMH-EDTA- and EDTA-treated controls. Althoughthey received simultaneous and similar DMH and EDTAtreatments, Groups X and A to E were maintained inseparate rooms to avoid inadvertent exposure to C. freundii. Table 1 illustrates the design of the experiment, thenumber of mice, and the time of C. freundii inoculation.The groups were actually established in staggered monthlyintervals so that Groups A to E were inoculated simultaneously with the same broth culture of C. freundii. In additionto these groups, 20 mice were given an equal volume ofsterile water weekly; 10 of these mice were killed at 1month of treatment, and 10 were killed at 2 months of
4352 CANCER RESEARCH VOL. 37
on June 14, 2019. © 1977 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Morphogenesis and Latency Reduction of DMH Carcinogenesis
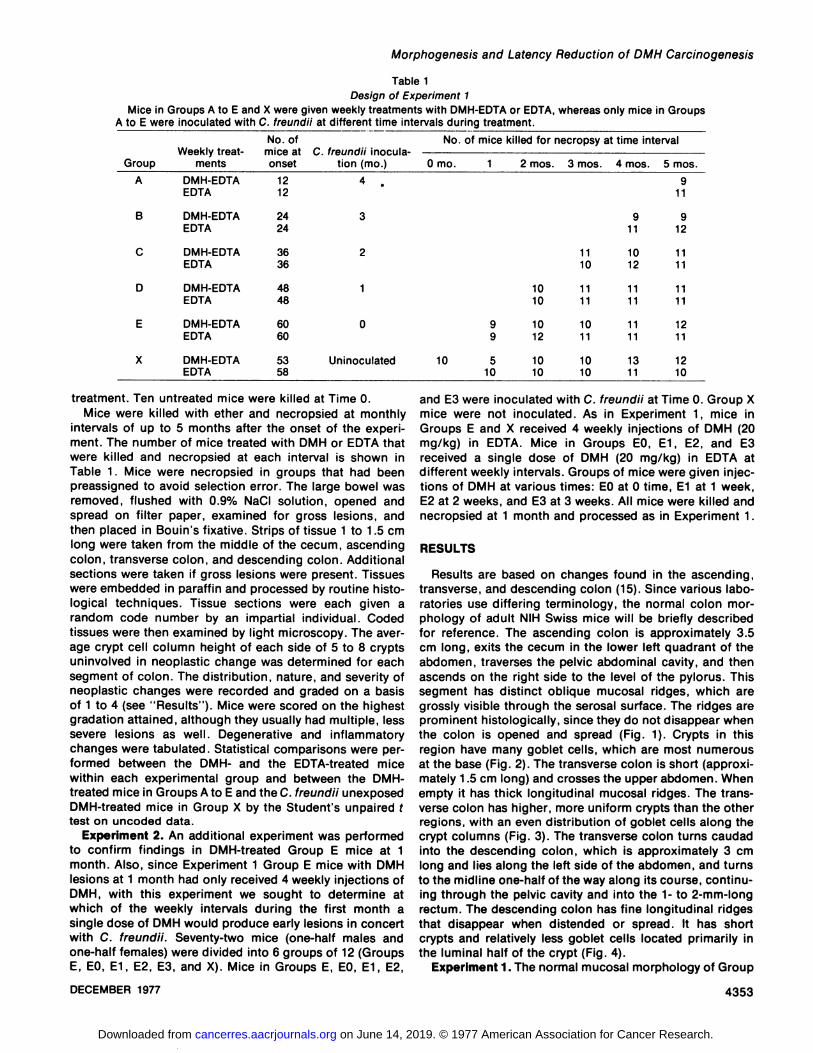
Table 1Design of Experiment 1
Mice in Groups A to E and X were given weekly treatments with DMH-EDTA or EDTA, whereas only mice in Groups
A to E were inoculated with C. freundii at different time intervals during treatment.
GroupABCDEXWeeklytreat
mentsDMH-EDTAEDTADMH-EDTAEDTADMH-EDTAEDTADMH-EDTAEDTADMH-EDTAEDTADMH-EDTAEDTANo.
ofmice atonset121224243636484860605358C.
freundii inoculation (mo.)4
.3210UninoculatedNo.
of mice killed for necropsy at timeinterval0
mo. 1 2mos.10109
1091210
51010103
mos.11101111101110104mos.91110121111111113115mos.9119121111111112111210
treatment. Ten untreated mice were killed at Time 0.Mice were killed with ether and necropsied at monthly
intervals of up to 5 months after the onset of the experiment. The number of mice treated with DMH or EDTA thatwere killed and necropsied at each interval is shown inTable 1. Mice were necropsied in groups that had beenpreassigned to avoid selection error. The large bowel wasremoved, flushed with 0.9% NaCI solution, opened andspread on filter paper, examined for gross lesions, andthen placed in Bouin's fixative. Strips of tissue 1 to 1.5 cm
long were taken from the middle of the cecum, ascendingcolon, transverse colon, and descending colon. Additionalsections were taken if gross lesions were present. Tissueswere embedded in paraffin and processed by routine histo-logical techniques. Tissue sections were each given arandom code number by an impartial individual. Codedtissues were then examined by light microscopy. The average crypt cell column height of each side of 5 to 8 cryptsuninvolved in neoplastic change was determined for eachsegment of colon. The distribution, nature, and severity ofneoplastic changes were recorded and graded on a basisof 1 to 4 (see "Results"). Mice were scored on the highest
gradation attained, although they usually had multiple, lesssevere lesions as well. Degenerative and inflammatorychanges were tabulated. Statistical comparisons were performed between the DMH- and the EDTA-treated micewithin each experimental group and between the DMH-treated mice in Groups A to E and the C. freundii unexposedDMH-treated mice in Group X by the Student's unpaired f
test on uncoded data.Experiment 2. An additional experiment was performed
to confirm findings in DMH-treated Group E mice at 1month. Also, since Experiment 1 Group E mice with DMHlesions at 1 month had only received 4 weekly injections ofDMH, with this experiment we sought to determine atwhich of the weekly intervals during the first month asingle dose of DMH would produce early lesions in concertwith C. freundii. Seventy-two mice (one-half males andone-half females) were divided into 6 groups of 12 (GroupsE, EO, E1, E2, E3, and X). Mice in Groups E, EO, E1, E2,
and E3 were inoculated with C. freundii at Time 0. Group Xmice were not inoculated. As in Experiment 1, mice inGroups E and X received 4 weekly injections of DMH (20mg/kg) in EDTA. Mice in Groups EO, E1, E2, and E3received a single dose of DMH (20 mg/kg) in EDTA atdifferent weekly intervals. Groups of mice were given injections of DMH at various times: EO at 0 time, E1 at 1 week,E2 at 2 weeks, and E3 at 3 weeks. All mice were killed andnecropsied at 1 month and processed as in Experiment 1.
RESULTS
Results are based on changes found in the ascending,transverse, and descending colon (15). Since various laboratories use differing terminology, the normal colon morphology of adult NIH Swiss mice will be briefly describedfor reference. The ascending colon is approximately 3.5cm long, exits the cecum in the lower left quadrant of theabdomen, traverses the pelvic abdominal cavity, and thenascends on the right side to the level of the pylorus. Thissegment has distinct oblique mucosal ridges, which aregrossly visible through the serosal surface. The ridges areprominent histologically, since they do not disappear whenthe colon is opened and spread (Fig. 1). Crypts in thisregion have many goblet cells, which are most numerousat the base (Fig. 2). The transverse colon is short (approximately 1.5 cm long) and crosses the upper abdomen. Whenempty it has thick longitudinal mucosal ridges. The transverse colon has higher, more uniform crypts than the otherregions, with an even distribution of goblet cells along thecrypt columns (Fig. 3). The transverse colon turns caudadinto the descending colon, which is approximately 3 cmlong and lies along the left side of the abdomen, and turnsto the midline one-half of the way along its course, continuing through the pelvic cavity and into the 1- to 2-mm-longrectum. The descending colon has fine longitudinal ridgesthat disappear when distended or spread. It has shortcrypts and relatively less goblet cells located primarily inthe luminal half of the crypt (Fig. 4).
Experiment 1. The normal mucosal morphology of Group
DECEMBER 1977 4353
on June 14, 2019. © 1977 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
S. W. Barthold and A. M. Jonas
X EDTA controls varied with the age of the mice. At theonset of the experiment, when mice were 21 to 35 days ofage and had received no treatment, the average height ofcrypts of the descending and transverse colon was 28cells. Crypt column height later increased in the transversecolon to a level higher than that of the descending colon,as illustrated in Charts 1 and 2. Ascending colon cryptcolumn height did not vary with age or treatment andremained between 25 and 30 cells. No sex differences weredetectable. EDTA produced no noticeable effect on themorphology of the colon compared to that produced bywater treatment for up to 2 months.
Three general categories of changes occurred in thecolon of DMH-treated Group X mice: neoplastic change,hyperplastic change, and degenerative-inflammatorychange. No changes were detectable in the cecum.
Neoplastic changes occurred only in DMH-treated mice.The lesions were graded on a basis of 1 to 4. Grade 1lesions were focal areas of epithelial proliferation in theluminal one-third to one-half of individual crypts. The basesof the affected crypts were normal. Proliferating cellslacked mucin droplets, were mitotically active, and becamecrowded and pseudostratified, but they maintained their
50 r
Xo
40
oo
30
20uo
10
p< .01p <.OI
p <.01
N.S. N.S.
01 2345
MONTHS OF TREATMENT
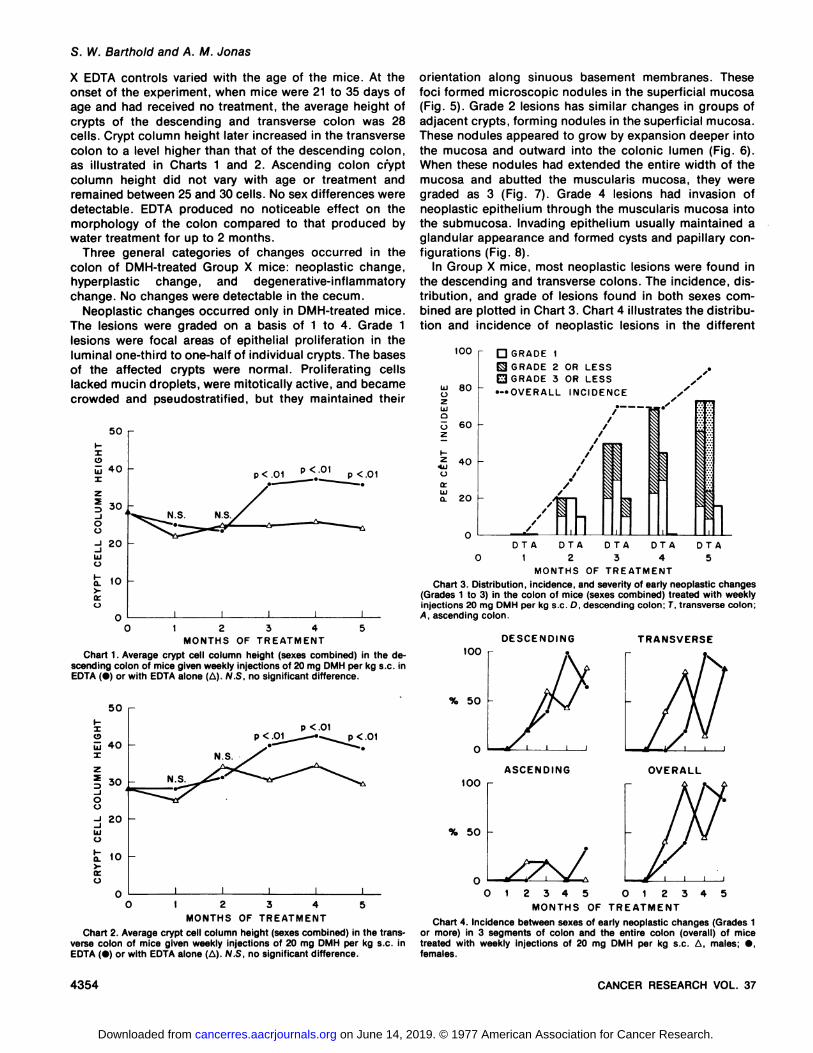
Chart 1. Average crypt cell column height (sexes combined) in the descending colon of mice given weekly injections of 20 mg DMH per kg s.c. inEDTA (•)or with EDTA alone (A). N.S, no significant difference.
50 r
p <.O1p <.01
12345MONTHS OF TREATMENT
Chart 2. Average crypt cell column height (sexes combined) in the transverse colon of mice given weekly injections of 20 mg DMH per kg s.c. inEDTA (•)or with EDTA alone (A). N.S. no significant difference.
orientation along sinuous basement membranes. Thesefoci formed microscopic nodules in the superficial mucosa(Fig. 5). Grade 2 lesions has similar changes in groups ofadjacent crypts, forming nodules in the superficial mucosa.These nodules appeared to grow by expansion deeper intothe mucosa and outward into the colonie lumen (Fig. 6).When these nodules had extended the entire width of themucosa and abutted the muscularis mucosa, they weregraded as 3 (Fig. 7). Grade 4 lesions had invasion ofneoplastic epithelium through the muscularis mucosa intothe submucosa. Invading epithelium usually maintained aglandular appearance and formed cysts and papillary configurations (Fig. 8).
In Group X mice, most neoplastic lesions were found inthe descending and transverse colons. The incidence, distribution, and grade of lesions found in both sexes combined are plotted in Chart 3. Chart 4 illustrates the distribution and incidence of neoplastic lesions in the different
100
BO
D GRADE 1
S GRADE 2 OR LESSE3 GRADE 3 OR LESS—OVERALL INCIDENCE
Illo5
60z¿
40
oo:LUo.
20--/•
~ ~ — _„,X*/////
///
/^^_A^^___DTA0
1^1|118II\\ii1DTA
DTA DTADTA2345
MONTHS OF TREATMENT
Chart 3. Distribution, incidence, and severity of early neoplastic changes(Grades 1 to 3) in the colon of mice (sexes combined) treated with weeklyinjections 20 mg DMH per kg s.c. D, descending colon: T. transverse colon;A. ascending colon.
DESCENDING TRANSVERSEtoo
OVERALL100 r
% 50
012345 012345
MONTHS OF TREATMENT
Chart 4. Incidence between sexes of early neoplastic changes (Grades 1or more) in 3 segments of colon and the entire colon (overall) of micetreated with weekly injections of 20 mg DMH per kg s.c. A, males; •,females.
4354 CANCER RESEARCH VOL. 37
on June 14, 2019. © 1977 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Morphogenesis and Latency Reduction of DMH Carcinogenesis
sexes. Lesions were often multiple, particularly after 3months, and in severely affected mice coalesced to producebroad areas of neoplastic change. No positive correlationof lesions with lymphoid nodules was observed.
Elongation of crypts within the colonie mucosa, whichwere not involved in neoplastic change, was also seen inGroup X DMH-treated mice (Charts 1 and 2). At 3 monthsor more of DMH treatment, mice of both sexes had highercrypt cell column heights in the descending and transversecolon although not in the ascending colon compared tothose in control mice (p < 0.01).
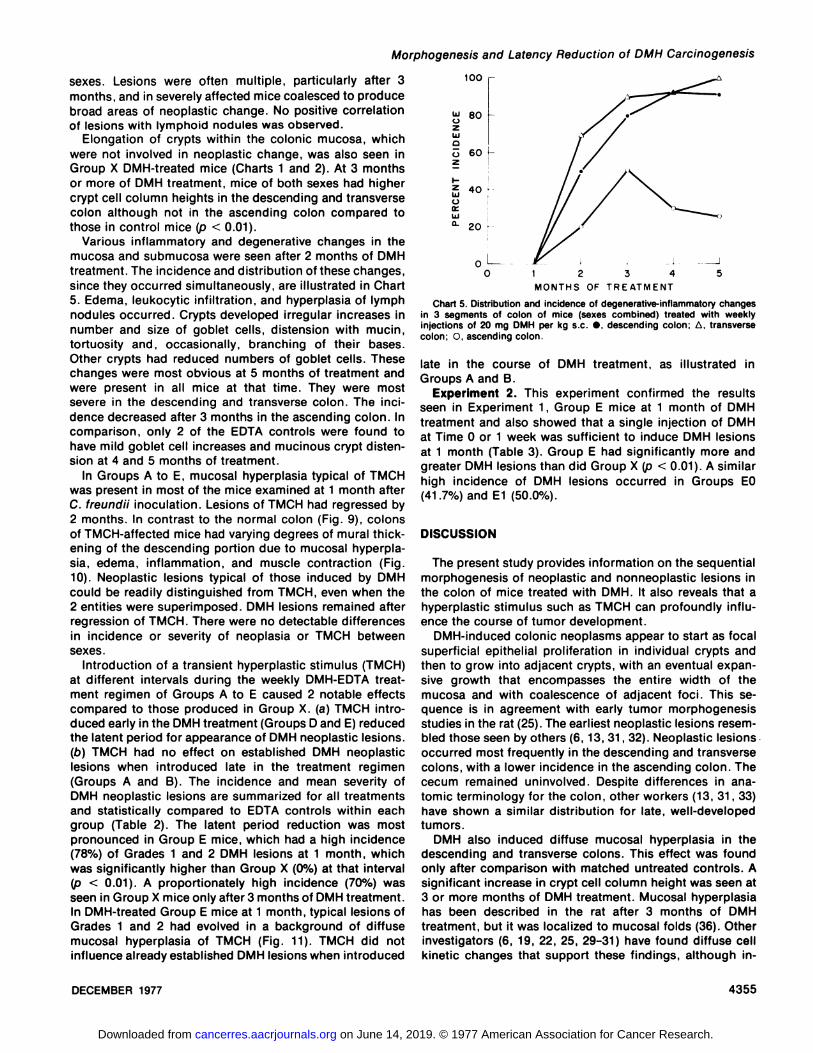
Various inflammatory and degenerative changes in themucosa and submucosa were seen after 2 months of DMHtreatment. The incidence and distribution of these changes,since they occurred simultaneously, are illustrated in Chart5. Edema, leukocytic infiltration, and hyperplasia of lymphnodules occurred. Crypts developed irregular increases innumber and size of goblet cells, distension with mucin,tortuosity and, occasionally, branching of their bases.Other crypts had reduced numbers of goblet cells. Thesechanges were most obvious at 5 months of treatment andwere present in all mice at that time. They were mostsevere in the descending and transverse colon. The incidence decreased after 3 months in the ascending colon. Incomparison, only 2 of the EDTA controls were found tohave mild goblet cell increases and mucinous crypt distension at 4 and 5 months of treatment.
In Groups A to E, mucosal hyperplasia typical of TMCHwas present in most of the mice examined at 1 month afterC. freundii inoculation. Lesions of TMCH had regressed by2 months. In contrast to the normal colon (Fig. 9), colonsof TMCH-affected mice had varying degrees of mural thickening of the descending portion due to mucosal hyperplasia, edema, inflammation, and muscle contraction (Fig.10). Neoplastic lesions typical of those induced by DMHcould be readily distinguished from TMCH, even when the2 entities were superimposed. DMH lesions remained afterregression of TMCH. There were no detectable differencesin incidence or severity of neoplasia or TMCH betweensexes.
Introduction of a transient hyperplastic stimulus (TMCH)at different intervals during the weekly DMH-EDTA treatment regimen of Groups A to E caused 2 notable effectscompared to those produced in Group X. (a) TMCH introduced early in the DMH treatment (Groups D and E) reducedthe latent period for appearance of DMH neoplastic lesions.(b) TMCH had no effect on established DMH neoplasticlesions when introduced late in the treatment regimen(Groups A and B). The incidence and mean severity ofDMH neoplastic lesions are summarized for all treatmentsand statistically compared to EDTA controls within eachgroup (Table 2). The latent period reduction was mostpronounced in Group E mice, which had a high incidence(78%) of Grades 1 and 2 DMH lesions at 1 month, whichwas significantly higher than Group X (0%) at that interval(p < 0.01). A proportionately high incidence (70%) wasseen in Group X mice only after 3 months of DMH treatment.In DMH-treated Group E mice at 1 month, typical lesions ofGrades 1 and 2 had evolved in a background of diffusemucosal hyperplasia of TMCH (Fig. 11). TMCH did notinfluence already established DMH lesions when introduced
100
80
D
O
LUUte.
60
20
"O 1 2 3 4 5
MONTHS OF TREATMENT
Chart 5. Distribution and incidence of degenerative-inflammatory changesin 3 segments of colon of mice (sexes combined) treated with weeklyinjections of 20 mg DMH per kg s c •,descending colon; A. transversecolon; C, ascending colon.
late in the course of DMH treatment, as illustrated inGroups A and B.
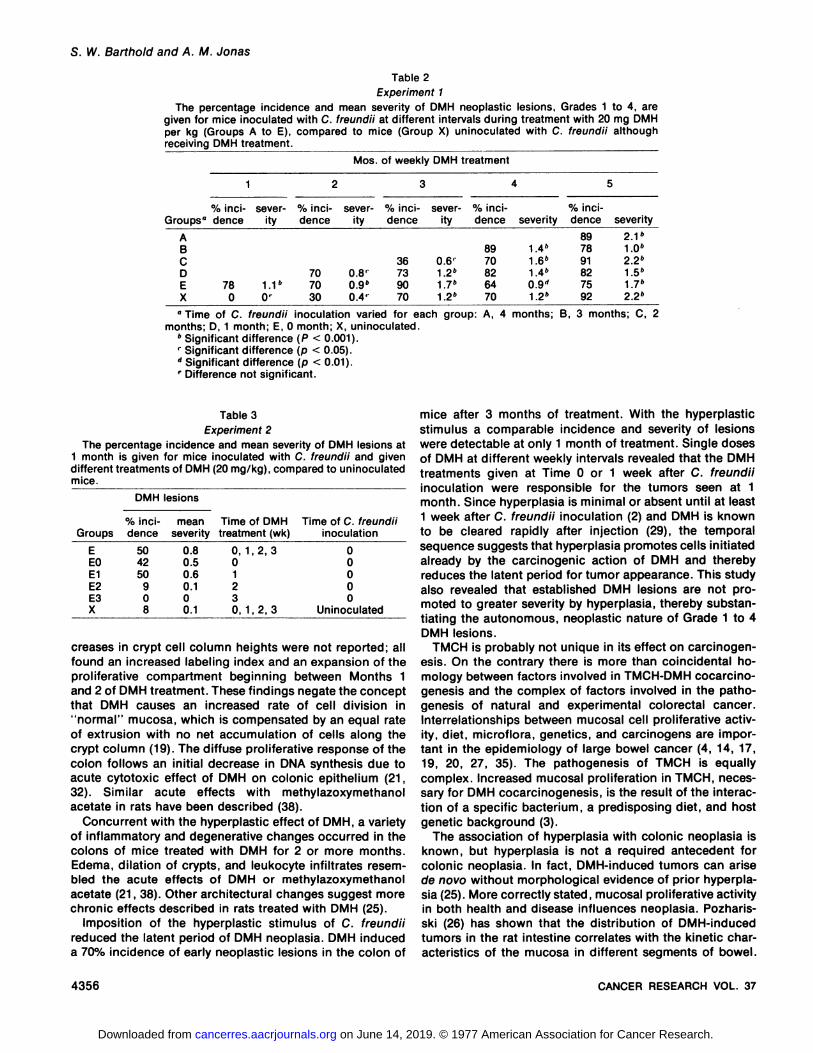
Experiment 2. This experiment confirmed the resultsseen in Experiment 1, Group E mice at 1 month of DMHtreatment and also showed that a single injection of DMHat Time 0 or 1 week was sufficient to induce DMH lesionsat 1 month (Table 3). Group E had significantly more andgreater DMH lesions than did Group X (p < 0.01). A similarhigh incidence of DMH lesions occurred in Groups EO(41.7%) and E1 (50.0%).
DISCUSSION
The present study provides information on the sequentialmorphogenesis of neoplastic and nonneoplastic lesions inthe colon of mice treated with DMH. It also reveals that ahyperplastic stimulus such as TMCH can profoundly influence the course of tumor development.
DMH-induced colonie neoplasms appear to start as focalsuperficial epithelial proliferation in individual crypts andthen to grow into adjacent crypts, with an eventual expansive growth that encompasses the entire width of themucosa and with coalescence of adjacent foci. This sequence is in agreement with early tumor morphogenesisstudies in the rat (25). The earliest neoplastic lesions resembled those seen by others (6, 13, 31, 32). Neoplastic lesionsoccurred most frequently in the descending and transversecolons, with a lower incidence in the ascending colon. Thececum remained uninvolved. Despite differences in anatomic terminology for the colon, other workers (13, 31, 33)have shown a similar distribution for late, well-developedtumors.
DMH also induced diffuse mucosal hyperplasia in thedescending and transverse colons. This effect was foundonly after comparison with matched untreated controls. Asignificant increase in crypt cell column height was seen at3 or more months of DMH treatment. Mucosal hyperplasiahas been described in the rat after 3 months of DMHtreatment, but it was localized to mucosal folds (36). Otherinvestigators (6, 19, 22, 25, 29-31) have found diffuse cellkinetic changes that support these findings, although in-
DECEMBER 1977 4355
on June 14, 2019. © 1977 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
S. W. Barthold and A. M. Jonas
Table 2Experiment 1
The percentage incidence and mean severity of DMH neoplastic lesions, Grades 1 to 4, aregiven for mice inoculated with C. freundii at different intervals during treatment with 20 mg DMHper kg (Groups A to E), compared to mice (Group X) uninoculated with C. freundii althoughreceiving DMH treatment.
Mos. of weekly DMH treatment
Groups"ABCDEX%inci
dence780severity1.1"0'%inci
dence707030severity0.8'0.9"0.4'%inci
dence36739070severity0.6'1.2"1.7*1.2"%inci
dence8970826470severity1.4"1.6"1.4»0.9"1.2"%inci
dence897891827592severity2.1"1.0"2.2"1.5"1.7"2.2»
" Time of C. freundii inoculation varied for each
months; D, 1 month; E, O month; X, uninoculated." Significant difference (P < 0.001).' Significant difference (p < 0.05).'' Significant difference (p < 0.01).' Difference not significant.
group: A, 4 months; B, 3 months; C, 2
Table 3
Experiment 2
The percentage incidence and mean severity of DMH lesions at1 month is given for mice inoculated with C. freundii and givendifferent treatments of DMH (20 mg/kg), compared to uninoculatedmice.
DMH lesions
GroupsE
EOE1E2E3X%
incidence50
4250908mean
severity0.8
0.50.60.100.1Time
of DMHtreatment(wk)0,
1,2,301230, 1,2, 3Time
of C. freundiiinoculation0
0000
Uninoculated
creases in crypt cell column heights were not reported; allfound an increased labeling index and an expansion of theproliferative compartment beginning between Months 1and 2 of DMH treatment. These findings negate the conceptthat DMH causes an increased rate of cell division in"normal" mucosa, which is compensated by an equal rate
of extrusion with no net accumulation of cells along thecrypt column (19). The diffuse proliferative response of thecolon follows an initial decrease in DMA synthesis due toacute cytotoxic effect of DMH on colonie epithelium (21,32). Similar acute effects with methylazoxymethanolacetate in rats have been described (38).
Concurrent with the hyperplastic effect of DMH, a varietyof inflammatory and degenerative changes occurred in thecolons of mice treated with DMH for 2 or more months.Edema, dilation of crypts, and leukocyte infiltrates resembled the acute effects of DMH or methylazoxymethanolacetate (21, 38). Other architectural changes suggest morechronic effects described in rats treated with DMH (25).
Imposition of the hyperplastic stimulus of C. freundiireduced the latent period of DMH neoplasia. DMH induceda 70% incidence of early neoplastic lesions in the colon of
mice after 3 months of treatment. With the hyperplasticstimulus a comparable incidence and severity of lesionswere detectable at only 1 month of treatment. Single dosesof DMH at different weekly intervals revealed that the DMHtreatments given at Time 0 or 1 week after C. freundiiinoculation were responsible for the tumors seen at 1month. Since hyperplasia is minimal or absent until at least1 week after C. freundii inoculation (2) and DMH is knownto be cleared rapidly after injection (29), the temporalsequence suggests that hyperplasia promotes cells initiatedalready by the carcinogenic action of DMH and therebyreduces the latent period for tumor appearance. This studyalso revealed that established DMH lesions are not promoted to greater severity by hyperplasia, thereby substantiating the autonomous, neoplastic nature of Grade 1 to 4DMH lesions.
TMCH is probably not unique in its effect on carcinogen-esis. On the contrary there is more than coincidental ho-mology between factors involved in TMCH-DMH cocarcino-genesis and the complex of factors involved in the patho-genesis of natural and experimental colorectal cancer.Interrelationships between mucosal cell proliferative activity, diet, microflora, genetics, and carcinogens are important in the epidemiology of large bowel cancer (4, 14, 17,19, 20, 27, 35). The pathogenesis of TMCH is equallycomplex. Increased mucosal proliferation in TMCH, necessary for DMH cocarcinogenesis, is the result of the interaction of a specific bacterium, a predisposing diet, and hostgenetic background (3).
The association of hyperplasia with colonie neoplasia isknown, but hyperplasia is not a required antecedent forcolonie neoplasia. In fact, DMH-induced tumors can arisede novo without morphological evidence of prior hyperplasia (25). More correctly stated, mucosal proliferative activityin both health and disease influences neoplasia. Pozharis-ski (26) has shown that the distribution of DMH-inducedtumors in the rat intestine correlates with the kinetic characteristics of the mucosa in different segments of bowel.
4356 CANCER RESEARCH VOL. 37
on June 14, 2019. © 1977 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

Morphogenesis and Latency Reduction of DMH Carcinogenesis
The descending colon, which has the largest stem cellpopulation and shortest life cycle, is the site of greatestpredilection for tumors. When proliferative activity wasincreased by nonspecific injury in a site of low tumorincidence (cecum), tumors developed in high frequency.Similarly, increasing the proliferative activity of the distalcolon mucosa in the present study reduced the latentperiod for tumorigenesis. The high incidence and early ageof onset of colon cancer in humans with hyperplasticmucosal diseases may be explained on this basis. Thecolonie mucosa of mice with TMCH has mitotic activityalong the entire crypt column (including the surface mucosa) similar to ulcerative colitis, familial polyposis, flatmucosa between tumors, and adenomatous and villouspolyps, as well as colonie neoplasms in humans and mice(2, 19).
Two other factors, diet and microflora, are integrallyrelated since one modifies the other. There is strong evidence that bacteria metabolize bile acids, bile salts, andother chemical compounds (including DMH) into carcino-genetic or cocarcinogenetic products that act on the intestinal epithelium (14, 27). These factors also have an important effect on>mucosal proliferative kinetics. The intestinalepithelial replacement rate is modified by gut microflora.The sluggish cell turnover rate of neonates increases withthe establishment of gut microflora, but it remains sluggishif the animal is maintained germ free (23). The lowerincidence of DMH tumors in germ-free versus conventionalrats, which has been attributed to the bacterial deconjuga-tion of carcinogens (28), could also be explained by directmicrofloral effects on mucosal proliferation rates. Diet significantly affects colon crypt cell column heights in miceexposed to C. freundii as well as in normal, unexposedmice (3).
The genetic association to human large bowel cancerhas been reviewed (4). A genetic factor is also involved incolon carcinogenesis with DMH. Certain inbred strains ofmice are susceptible to tumor induction by DMH, whereasothers are resistant (11). Parallel results have been foundwith similar strains of mice in their hyperplastic responseto C. freundii (3). The genetic mechanism is not known, butit is clearly related to mucosal proliferative response.
This study shows that hyperplasia (TMCH) modifies theresponse of colonie epithelim to the chemical carcinogenDMH. TMCH, which depends on microflora, diet, and hostgenetics, provides insight into the same mechanisms involved in cancer of the large bowel in man.
REFERENCES
1. Barthold, S. W.. Coleman, G. L.. Bhatt. P. N.. Osbaldiston, G. W.. andJonas, A. M. The Etiology of Transmissible Murine Colonie Hyperplasia.Lab. Animal Sci., 26: 889-894, 1976.
2. Barthold, S. W., Coleman, G. L., Jacoby, R. 0., Livstone, E., andJonas, A. M. Transmissible Murine Colonie Hyperplasia. Vet. Pathol., inpress.
3. Barthold, S. W., Osbaldiston, G. W. and Jonas, A. M. Dietary, Bacterialand Host Genetic Interactions in the Pathogenesis of TransmissibleMurine Colonie Hyperplasia. Lab. Animal Sci., in press.
4. Burdette, W. J. Heritable Cancer of the Colorectum. In: W. J. Burdette(ed.), Carcinoma of the Colon and Antecedent Epithelium, pp. 78-102.Springfield, III.: Charles C Thomas, Publisher, 1970.
5. Darke, S. G., Parks, A. G.. Grogono, J. L., and Pollock, D. J. Adenocar-cinoma and Crohn's Disease. Brit. J. Surg., 60: 169-175, 1973.
6. Deschner, E. E. Experimentally Induced Cancer of the Colon. Cancer,34: 824-828, 1974.
7. Deschner, E., and Lipkin, M. Study of Human Rectal Epithelial Cells invitro. III. RNA, Protein and DNA Synthesis in Polyps and AdjacentMucosa. J. Nat.Cancer Inst., 44: 175-185, 1970.
8. Double, J. A., Ball, C. R., and Cowen, P. N. Transplantation of Adeno-carcinomas of the Colon in Mice. J. Nati. Cancer Inst. ,54: 271-275,1975.
9. Evans, J. T., Hauschka. T. S., and Mittelman, A. Differential Susceptibility of Four Mouse Strains to Induction of Multiple Large-Bowel Neoplasms by 1,2-Dimethylhydrazine. J. Nati. Cancer Inst., 52. 999-1000,1974.
10. Evans, J. T.. Lutman, G., and Mittelman, A. The Induction of MultipleLarge Bowel Neoplasms in Mice. J. Med.. 3: 212-215, 1972.
11. Evans, J. T., Shows, T. B., Sproul, E. E., Paolini, N. S., Mittelman, A.,and Hauschka, T. S. Genetics of Colon Carcinogenesis in Mice Treatedwith 1,2-Dimethylhydrazine. Cancer Res.,37: 134-136, 1977.
12. Farmer, R. G., Hawk, W. A., and Turnbull, R. B. Carcinoma Associatedwith Mucosal Ulcerative Colitis, and with Transmural Colitis andEnteritis (Crohn's Disease). Cancer, 28: 289-292, 1971.
13. Haase, P., Cowen, D. M., Knowles, J. C., and Cooper, E. H. Evaluationof Dimethylhydrazine Induced Tumors in Mice as a Model System forCplorectal Cancer. Brit. J. Cancer, 28: 530-543, 1973.
14. Hill, M. J. The Role of Colon Anaerobes in the Metabolism of Bile Acidsand Steroids, and Its Relation to Colon Cancer. Cancer, 36: 2387-2400,1975.
15. Hummel, K. P., Richardson, F. L., and Fekete, E. Anatomy. In: E. G.Green (ed.), Biology of the Laboratory Mouse, pp. 247-307. New York:McGraw-Hill Book Company, 1966.
16. Kronman, B. S. Ulcerative Colitis, Autoimmune Epiphenomena. andColonie Cancer. Cancer, 28: 82-88, 1971.
17. Laqueur, G. L. Contributions of Intestinal Macroflora and Microflora toCarcinogenesis. in: W. J. Burdette (ed.), Carcinoma of the Colon andAntecedent Epithelium, pp. 305-313. Springfield. III.: Charles C ThomasPublisher, 1970.
18. Lipkin, M. Proliferation and Differentiation of Gastrointestinal Cells.Physiol. Rev., 53: 891-915, 1973.
19. Lipkin, M. Phase 1 and Phase 2 Proliferative Lesions of Colonie Epithelial Cells in Disease Leading to Colonie Cancer. Cancer, 34: 878-888,1974.
20. Lipkin, M. Biology of Large Bowel Cancer. Present Status and ResearchFrontiers. Cancer, 36. 2319-2324, 1975.
21. Lohrs. U., Wiebecke, B., and Eder, M. Morphologische und autoradiographische Untersuchung der Darmschleim-hautveränderungen nacheinmaliger Injektion von 1,2-Dimethylhydrazin. Z. Ges. Exptl. Med., 151:297-307, 1969.
22. Maskens, A. P. Histogenesis and Growth Pattern of 1,2-Dimethylhydra-zine-induced Rat Colon Adenocarcinoma. Cancer Res.. 36: 1585-1592,1976.
23. Moon, H. W. Intestine. In: N. F. Cheville (éd.).Cell Pathology, pp. 380-407. Ames, Iowa: Iowa State University Press, 1976.
24. Pegg, A. E., and Hawks, A. Increased Transfer Ribonucleic Acid Methyl-ase Activity in Tumors induced in the Mouse Colon by the Administrationof 1.2-Dimethylhydrazine. Biochem. J.. 722: 121-123, 1971.
25. Pozharisski, K. M. Morphology and Morphogenesis of ExperimentalEpithelial Tumors of the Intestine. J. Nat. Cancer Inst., 54. 1115-1135.1975.
26. Pozharisski, K. M. The Significance of Nonspecific Injury for ColonCarcinogenesis in Rats. Cancer Res., 35. 3824-3830, 1975.
27. Reddy, B. S. Role of Bile Metabolites in Colon Carcinogenesis. AnimalModels. Cancer, 36: 2401-2406. 1975.
28. Reddy, B. S.. Weisburger, J. H., Narisawa, T., and Wynder, E. L. ColonCarcinogenesis in Germ-free Rats with 1,2-Dimethylhydrazine and N-Methyl-AT-nitro-N-nitrosoguanidine. Cancer Res., 34: 2368-2372, 1974.
29. Shimizu, E., and Toth, B. Autoradiographic Studies on the Distributionof "C-1,2-Dimethylhydrazine Dihydrochloride and Its Effect on DNASynthesis in Swiss Mice. Gann. 66: 589-601, 1975.
30. Springer, P., Springer, J., and Oehlerte, W. Die Vorstufen das 1,2-Dimethylhydrazininduzierten Dick- und Dünnolarmcarcinomsder Ratte.Z. Krebsforsch., 74: 236-240. 1970.
31. Thurnherr, N.. Deschner. E. E., Stonehill, E. H., and Lipkin, M. Inductionof Adenocarcinomas of the Colon in Mice by Weekly Injections of 1,2-Dimethylhydrazine. Cancer Res., 33: 940-945, 1973.
32. Thurnherr. N., and Reinhart, K. Induktion von Kolonkarzinomen mit 1.2-Dimethylhydrazin-Hydrochlorid bei der Maus. Schweiz. Med. Wochschr.,105: 585-586. 1975.
33. Ward, Ü.M.,Yamamoto. R. S., Benjamin, T.. Brown, C..and Weisburger,J. H. Experimentally Induced Cancer of the Colon in Rats and Mice. J.Am. Vet. Med. Assoc.. 164: 729-732, 1974.
34. Weedon, D. D., Shorter, R. G., Ilstrup, D. M., Huizenga, K. A., andTaylor, W. F. Crohn's Disease and Cancer. New Engl. J. Med., 289:1099-1103, 1973.
35. Weisburger, J. H. Chemical Carcinogens and Their Mode of Action inColonie Neoplasia. Diseases Colon Rectum, 76: 431-437, 1973.
DECEMBER 1977 4357
on June 14, 2019. © 1977 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

S. W. Barthold and A. M. Jonas
36. Wiebecke, B., Krey, U., Lóhrs, U., and Eder, M. Morphological and Med., Õ49:277-278, 1969.Autoradiographical Investigations on Experimental Carcinogenesis and 38. Zedeck, M. S., Grab, D. J., and Sternberg, S. S. Differences in thePolyp Development in the Intestinal Tract of Rats and Mice. Virchows Acute Response of the Various Segments of Rat Intestine to TreatmentArch. Abt. A Pathol. Anat., 360: 179-193, 1973. with the Intestinal Carcinogen, Methylazoxymethanol Acetate. Cancer
37. Wiebecke. B., Lòhrs, U., Gimmy, J., and Eder, M. Erzeugung von Res., 37: 32-36, 1977.Darmtumoren bei Mäusendurch 1,2-Dimethylhydrazin. Z. Ges. Exptl.
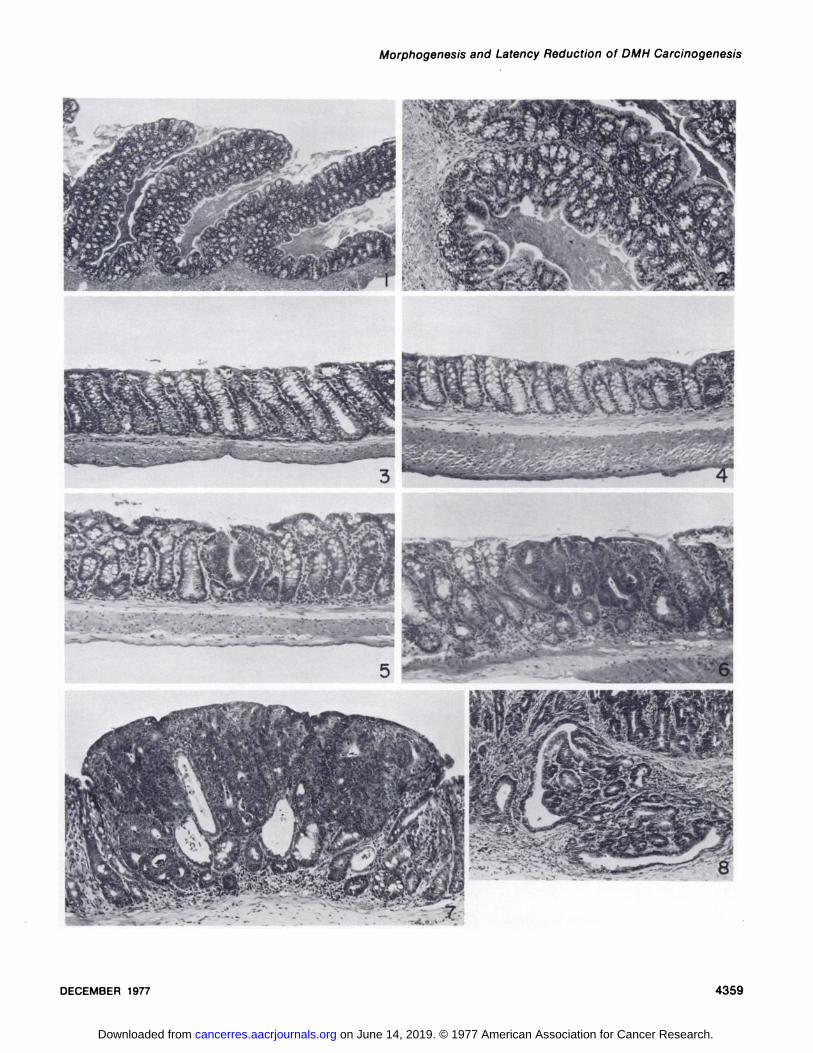
Fig. 1. Ascending colon, normal mouse. Note the prominent mucosal ridges that remain when the colon is opened and spread. Goblet cells areprominent at the base of the crypts. H & E, x 32.
Fig. 2. Ascending colon, normal mouse. Higher magnification of mucosa is shown in Fig. 1. H & E, x 95.Fig. 3. Transverse colon, normal mouse. Crypts are highest in this segment of colon, and goblet cells are evenly distributed along the crypt columns. H &
E. x 95.Fig. 4. Descending colon, normal mouse. Crypts are shorter and goblet cells are fewer than in the transverse colon. Goblet cells are located in the top
half of the crypts. H & E, x 95.Fig. 5. Grade 1 DMH-induced neoplastic lesion, transverse colon. The mucosa contains a nodule of epithelial cell proliferation apparently arising from a
single crypt. Cells lack mucin droplets and are crowded and pseudostratified. H & E, x 95.Fig. 6. Grade 2 DMH-induced neoplastic lesion, descending colon. The nodular growth within the superficial mucosa is composed of several adjacent
abnormal crypts. H & E, x 95.Fig. 7. Grade 3 DMH-induced neoplastic lesion, descending colon. The nodule encompasses the entire width of the mucosa, abuts the muscularis
mucosa, and protrudes into the colonie lumen. H & E, x 95.Fig. 8. Grade 4 DMH-induced neoplastic lesion, descending colon. The base of the neoplasm within the mucosa has infiltrating glandular cords
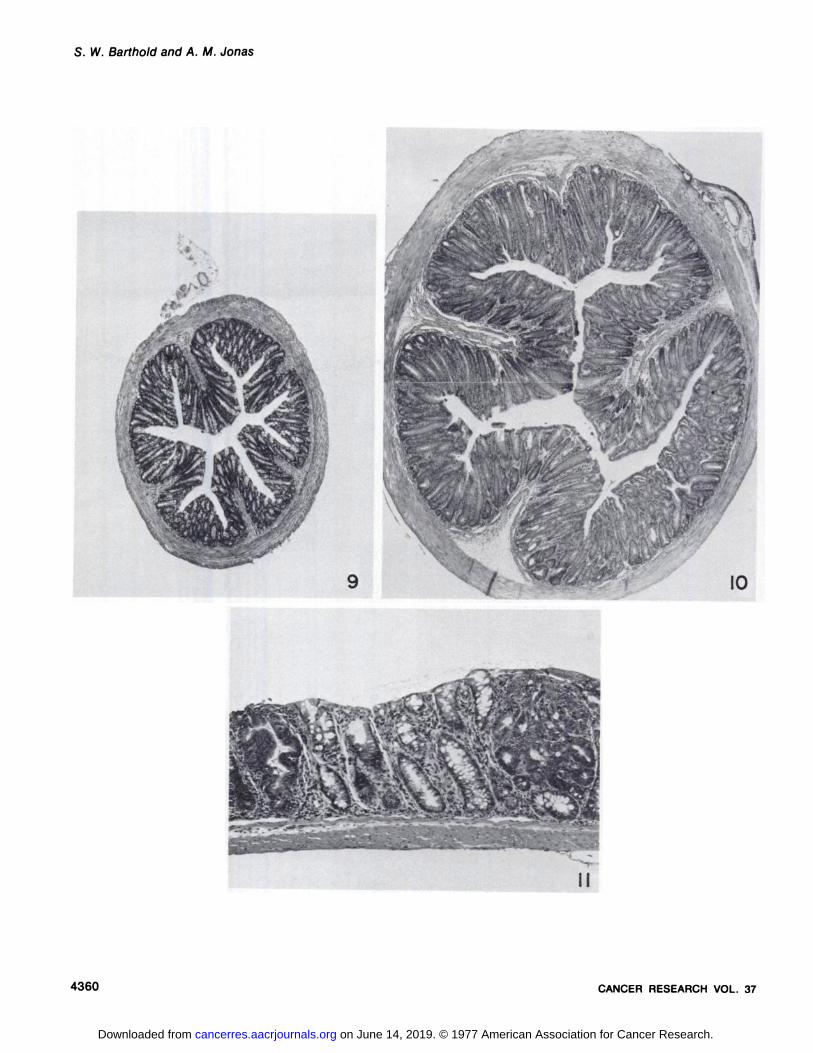
extending downward through the muscularis mucosa into the submucosa. H & E, x 95.Fig. 9. Cross-section of the descending colon from a normal mouse. H & E, x 12.5.Fig. 10. Cross-section of the descending colon from a mouse with well-developed TMCH. The diameter is increased due to marked mucosal epithelial
hyperplasia, inflammation, and muscular contraction. H & E, x 12.5.Fig. 11. Descending colon from a mouse 1 month after C. freundii inoculation and onset of weekly DMH treatments (Group E). The mucosa is mildly and
diffusely hyperplastic with 2 focal DMH neoplastic lesions superimposed. H & E, x 95.
4358 CANCER RESEARCH VOL. 37
on June 14, 2019. © 1977 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Morphogenesis and Latency Reduction of DMH Carcinogenesis
a
DECEMBER 1977 4359
on June 14, 2019. © 1977 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
S. W. Barthold and A. M. Jonas
•*,,
9 IO
4360 CANCER RESEARCH VOL. 37
on June 14, 2019. © 1977 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1977;37:4352-4360. Cancer Res Stephen W. Barthold and Albert M. Jonas
Citrobacter freundiiby a Variant of and Latent Period Reduction of Colon Carcinogenesis in Mice Morphogenesis of Early 1,2-Dimethylhydrazine-induced Lesions
Updated version
http://cancerres.aacrjournals.org/content/37/12/4352
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/37/12/4352To request permission to re-use all or part of this article, use this link
on June 14, 2019. © 1977 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from





























