Embed Size (px)
Citation preview

Journal of Experimental Botany, Vol. 65, No. 22 pp. 6513–6528, 2014doi:10.1093/jxb/eru369 Advance Access publication 10 September, 2014
v
ReseaRch PaPeR
Mitogen-activated protein kinase 6 regulates NPR1 gene expression and activation during leaf senescence induced by salicylic acid
Jinyu Chai, Jian Liu, Jun Zhou and Da Xing*
MOE Key Laboratory of Laser Life Science & Institute of Laser Life Science, College of Biophotonics, South China Normal University, Guangzhou 510631, China
* To whom correspondence should be addressed. E-mail: [email protected]
Received 28 May 2014; Revised 7 August 2014; Accepted 12 August 2014
Abstract
Plant senescence is a highly regulated process that can be induced by a range of factors. The nonexpressor of patho-genesis-related genes 1 (npr1) mutant is defective in the salicylic acid (SA) signalling pathway, displaying delayed yellowing during developmental senescence. However, the regulating mechanism of NPR1 on exogenous SA-induced senescence in detached Arabidopsis leaves has not yet been clarified. It was shown here that mitogen-activated protein kinase 6 (MPK6) is involved in promoting exogenous SA-induced detached leaf senescence. During the pro-cess of SA-induced senescence, the expression of NPR1 and senescence-related transcription factor WRKY6 was suppressed in mpk6 mutant plants. Further analyses showed that the NPR1 mRNA level is reduced in wrky6 mutants and enhanced in WRKY6 overexpressing lines. Meanwhile, chromatin immunoprecipitation experiments revealed that WRKY6 binds directly to the NPR1 promoter containing W-box motifs. Moreover, inhibition of MPK6 function dimin-ished SA-induced monomerization and nuclear localization of NPR1. In addition, the expression of Trx h5, which catalyses the SA-induced NPR1 activation, was suppressed in the mpk6 mutant, suggesting that MPK6 promotes NPR1 activation, possibly by regulating the expression of Trx h5. Collectively, MPK6-mediated WRKY6 and Trx h5 transcriptional activation co-regulated the expression of the NPR1 gene and the monomerization of NPR1 protein, allowing it to enter the nucleus, thereby promoting SA-induced leaf senescence. These results provide new insight into the mechanism of exogenous SA-induced detached leaf senescence.
Key words: Arabidopsis, NPR1, MPK6, salicylic acid, senescence, Trx h5, WRKY6.
Introduction
Senescence is an intricate process in which a series of regu-lated metabolic events takes place (Abreu and Munné-Bosch, 2008). In plants, senescence can be induced by a wide variety of factors including mineral deficiency, drought, hormones, and infection by pathogens. Several genes have been identi-fied as being senescence-associated (SEN, SAG), and some of these have been isolated and characterized as molecular markers of senescence (Lim et al., 2003). In contrast to the
delaying effect of cytokinins, salicylic acid (SA), methyl jas-monate, ethylene, and brassinosteroid have all been implicated in the onset of senescence (Schippers, 2008). Measurements of endogenous SA during the progression of leaf senescence, the external applications of SA, and the use of transgenic lines deficient in SA have shown that SA participates, either directly or indirectly, in regulating senescence-associated gene expression (Abreu and Munné-Bosch, 2008). In addition to
© The Author 2014. Published by Oxford University Press on behalf of the Society for Experimental Biology. All rights reserved. For permissions, please email: [email protected]
Abbreviations: CaMV, cauliflower mosaic virus; DTT, dithiothreitol; Fv/Fm, maximum variable fluorescence to maximum yield of fluorescence; GFP, green fluorescence protein; GSH, reduced glutathione; GSSG, oxidized glutathione; MAPK, mitogen-activated protein kinase; PMSF, phenylmethanesulfonyl fluoride; PSII, photosystem II; qRT-PCR, quantitative reverse transcription PCR; ROS, reactive oxygen species; SA, salicylic acid; SD, standard deviation; Trx, thioredoxin, WT, wild type.
at South China N
ormal U
nviversity on Novem
ber 28, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

6514 | Chai et al.
playing a role in the senescence process, SA has a central role in plant defence against pathogen attack, and also participates in diverse processes such as thermotolerance acquisition, ger-mination, flowering, and drought stress (Krinke et al., 2007). Arabidopsis plants defective in the SA signalling pathway, such as NahG transgenic plants and pad4 and npr1 mutants, show an altered expression pattern of senescence-enhanced genes, delayed yellowing, and reduced necrosis during devel-opmental senescence (Morris et al., 2000). Moreover, NPR1 plays a role in mediating SA-induced cell death, and this process is regulated by the reductants glutathione and dithi-othreitol (DTT) (Ogawa, 2005).
The mitogen-activated protein kinase (MAPK) cascade is involved in a variety of abiotic and biotic stress responses by sequential phosphorylation (Bartels et al., 2009; Zhou et al., 2009). Recently, the MKK9–MPK6 cascade has been shown to regulate leaf senescence in Arabidopsis (Zhou et al., 2009). It has been shown that Arabidopsis transgenic NahG plants can reduce the protein levels of MPK3, MPK4, and MPK6, which leads to the conclusion that these MAPK proteins are modulated by SA level. Moreover, MPK3 and MPK6 have been shown to lead to SA biosynthesis and the expression of PR1 and PR5 (Bartels et al., 2009). MAPKs have been implicated in upregulation of the thioredoxin (Trx), glutathione S-transferase, and late-embryogenesis-abundant/dehydrin-type genes (Habora et al., 2012). Plant Trxs are involved in regulation of the cellular redox state. Changes in the cellular redox state promote NPR1 oli-gomer disassociation, in turn leading to monomerization of NPR1. Subsequently, the monomeric NPR1 migrates from the cytoplasm to the nucleus to regulate downstream gene expression by interaction with transcription activators such as TGA1 (Cheng et al., 2009). Extensive research has shown that the nuclear targets of three MAPKs (MPK3, MPK6, and MPK4) include many pivotal transcriptional regula-tors such as the WRKY transcription factors, EIN3, and ERF104 (van den Burg and Takken, 2010). There are 74 WRKY transcription factors in Arabidopsis, which act as the second largest family in the senescence transcriptome (Mao et al., 2007). WRKY proteins contain the most conserved amino acid sequence, WRKYGQW, in their N terminus, and zinc-finger motifs in their C terminus, and they can bind to a consensus motif of TTGAC(C/T), called the W-box, which exists in the promoters of certain defence- and senes-cence-related genes (Robatzek and Somssich, 2001, 2002; Zhou et al., 2011). WRKY transcription factors have been shown to be downstream components of MAPK signalling cascades, and nearly all of the WRKY factors studied can recognize W-box motifs present in the promoters of stress-responsive genes (Yu et al., 2001; Ciolkowski et al., 2008). Moreover, expression of members of the WRKY family such as WRKY22/29, WRKY6, WRKY53, and WRKY18 is upregulated during leaf senescence (Gepstein, 2004). WRKY6 is identified as a senescence- and defence-associ-ated transcription factor and, more importantly, it can act both as a positive and a negative regulator to stimulate or repress the transcriptional level of target genes (Robatzek and Somssich, 2002).
In this study, the possible molecular mechanism underly-ing the regulatory effect of exogenous SA on NPR1 during leaf senescence is presented. It was observed that SA-induced activation of MPK6 contributed to higher expression of WRKY6, which in turn enhanced WRKY6 binding to the W-boxes in the NPR1 gene promoter to increase the mRNA level of NPR1. Further analyses demonstrated that the nuclear accumulation of NPR1 was reduced in mpk6 mutant plants. By investigating the MPK6-associated signalling path-ways, we found that MPK6 plays a role in modulating the activity of thioredoxin h (Trx h), which was shown to catalyse SA-induced NPR1 conformational changes and was required in vivo for SA-induced NPR1 monomer release. In general, our research provides evidence that the MPK6-modulated signalling pathway is responsible for the gene expression and activation of NPR1 in the process of SA-induced leaf senescence.
Materials and methods
Plant materials, growth conditions, and chemical treatmentsWild-type (WT) Arabidopsis ecotype Columbia (Col-0) plants, mpk3-1 (SALK_151594), mpk4-2 (SALK_056245), mpk6-3 (SALK_127507), npr1, wrky6-1, and wrky6-2 mutants and the cauli-flower mosaic virus (CaMV) 35S::WRKY6-9 overexpressing line, as well as transgenic 35S::NPR1–GFP in the npr1 mutant (Sun et al., 2012), were used. Arabidopsis mutants and overexpressing lines used in this work were identified using semi-quantitative reverse transcription (RT)-PCR (Supplementary Fig. S1 at JXB online). Following 2 d cold stratification, seeds were grown in an environ-mentally controlled growth room at 22 °C with a 16 h light photo-period (120 μmol quanta m–2 s–1) and 82% relative humidity. The effects of SA on leaf senescence were examined on leaf numbers 4 and 5, detached from plants 3 weeks after sowing.
For hormone treatment, 3-week-old detached leaves were floated in 3 mM 2-(N-morpholino) ethanesulfonic acid (MES) buffer (pH 5.8) with or without 100 µM SA (Sharabi-Schwager et al., 2010; Zhang et al., 2012). Leaf samples were incubated under continu-ous light and harvested at different times after treatment. Before the experiment, leaves were washed three times with distilled water.
The common inhibitor of MAPK cascade, PD98059, was pur-chased from Sigma-Aldrich (Shanghai, China). Before SA treatment, the detached leaves were pre-incubated with PD98059 (dissolved in DMSO) for 60 min, and the final concentration of PD98059 was 150 µM (Li et al., 2012).
Measurement of chlorophyll content and photochemical efficiencyChlorophyll was extracted by boiling the leaves in 95% ethanol at 80 °C. The chlorophyll concentration per unit fresh weight of leaf was calculated as described by Lichtenthaler (Lichtenthaler, 1987). The content was determined spectrophotometrically using the formula Chl (a+b)=5.24 A664.2+22.24 A648.6 and chlorophyll content (%)=Chl (a+b)×V×10–3/m, where Chl (a+b) is the chlo-rophyll concentration in μg ml–1, A is absorption at the indicated wavelength, V is the volume of the 95% ethanol, and m is the mass of the leaves.
The ratio of maximum variable fluorescence to maximum yield of fluorescence (Fv/Fm) that can be affected by the integrity of pho-tosystem II (PSII) and used as an indicator for leaf senescence (Oh et al., 1996) was measured using a PAM fluorometer (Walz GmbH, Effeltrich, Germany) (Ralph et al., 2005).
at South China N
ormal U
nviversity on Novem
ber 28, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

NPR1 promotes SA-induced detached leaf senescence | 6515
Total RNA extraction, RT-PCR and quantitative RT-PCR (qRT-PCR)Total RNA was extracted using TRIZOL reagent (Sigma-Aldrich, Shanghai, China), following the manufacturer’s protocol. The con-centration and purity of RNA were determined by measuring A260. Total RNAs were reverse transcribed into cDNA using a Moloney murine leukemia virus kit (Takara, Dalian, China), and the cDNAs were used as templates in PCRs.
RT-PCR was performed with RT-PCR High-plus- (Toyobo, Osaka, Japan) using gene-specific primers designed to target Actin2, MPK3, MPK4, WRKY6, and NPR1. The gene-specific primers for RT-PCR analysis were used as described previously (Robatzek and Somssich, 2002; Laloi et al., 2004; Li et al., 2012).
For qRT-PCR assay, SYBR Green real-time RT-PCR amplifica-tions were carried out using SYBR Green real-time PCR Master mix (Toyobo, Osaka, Japan). The Actin2 gene was amplified as an endogenous control. Gene-specific primers for qRT-PCR analyses were used as described previously (Chen et al., 2009; Ishikawa et al., 2010; Zhou et al., 2011; Love et al., 2012; Jaradat et al., 2013; Jiang et al., 2014).
Chromatin immunoprecipitation (ChIP)In each experiment, 1.5 g of leaves was treated and analysed as described previously for ChIP (Nelson et al., 2006). An aliquot of 1% formaldehyde was added for cross-linking and the formaldehyde was quenched with 2.5 ml of 2 M glycine under vacuum infiltration and the tissues were then harvested for extraction of chromatin. To shear the chromatin, the samples were sonicated four times for 10 s at 20% power and kept on ice for 1 min during each interval. Before precipitation, 1:10 dilutions of the supernatant were kept aside as the ‘input fraction’ for the cross-linking reversal, DNA purifica-tion, and PCR assay. Appropriate antibodies were added to each diluted sample and incubated overnight at 4 °C with gentle rota-tion. The polyclonal antibody against WRKY6 was rabbit anti-WRKY6 serum (a gift from Dr Weihua Wu, China Agricultural University, China) and anti-acetyl-histone 3 and normal rabbit IgG were used as positive and negative controls. Magnetic beads (Millipore, Billerica, USA) were added to the immune complex and vortexed to mix. This was followed by standing on a magnetic stand for 10 min and the supernatant was then discarded. The magnetic beads were washed for 3–5 min each time with gentle rotation at 4 °C and then centrifuged to discard the supernatants. The com-plexes were then eluted from the magnetic beads and centrifuged at 4 °C. The elution step was repeated once and the elution prod-ucts combined. Next, NaCl was added and it was all incubated at 65 °C for at least 6 h overnight to reverse the cross-linking. RNase was then added and incubated at 37 °C for 15 min. The protein was digested and the DNA purified using a GV-High efficiency DNA fragment purification kit (Genview, Beijing, China). PCR was performed using specific primers to amplify the sequences with W-box regions (from –170 to –2) contained in the NPR1 gene promoter (NPR1-F, 5′-GTGGGACGAGGATGCTTCTT-3′, and NPR1-R, 5′-CAACAGGTTCCGATGAATTGA-3′), the sequences without W-box regions (from –1001 to –821) (NPR1-F, 5′-AACCAAGAGGTGATTATGCAG-3′, and NPR1-R, 5′-GCCTTCAACTTCTCGTTCTG-3′), and the known sequences of PHO1 bound by WRKY6 (Chen et al., 2009) (PHO1-F, 5′-ACAGTTTGCTTTCAAATCAGTTATTG-3′, and PHO1-R, 5′-AAACATCTTCACTTTTCCATCAAA-3′).
Confocal laser-scanning microscopy (CLSM)Green fluorescent protein (GFP) fluorescence in leaves expressing the NPR1 reporter constructs was determined using a Zeiss LSM 510 confocal laser-scanning microscope (LSM510/Con-foCor2; Carl-Zeiss, Jena, Germany) (Pavet et al., 2005). Leaf samples were mounted in distilled water and GFP signals were illuminated with excitation and emission wavelengths of 488 nm and 500–550 nm,
respectively, using a band-pass filter, and chloroplast autofluores-cence (488 nm excitation) was visualized at 650 nm with a long-pass filter. To identify the nucleus, leaves were fixed with a fixation solu-tion and stained with propidium iodide, a fluorescent dye that stains the nucleus (Running et al., 1995).
Protein extraction and nuclear fractionationAmounts of 0.4 g of leaves were ground in liquid nitrogen and the powder resuspended in 1 ml of ice-cold extraction buffer comprising 50 mM Tris/HCl (pH 6.8), 50 mM DTT, 4% (w/v) SDS, 10% (v/v) glycerol, 1% (w/v) polyvinylpolypyrrolidone, and 50 μl phenylmeth-anesulfonyl fluoride (PMSF). The extracts were centrifuged and the protein concentration of the supernatant was determined with a Bio-Rad protein assay.
Nuclear fractionation was performed as described previously (Kinkema et al., 2000). The cytoplasmic samples were standardized using anti-PEPC antibody and the nuclear protein using anti-his-tone H3 antibody.
Western blotting and immunoblot assaySamples of total protein (40 μg) were separated by 8% SDS-PAGE and Western blotting was performed. For detection of NPR1 proteins, blots were performed using antibodies raised against NPR1 (a gift from Dr Xinnian Dong, Duke University, NC, USA) at a 1:1000 dilution. Plant MAPKs are highly homolo-gous to mammalian ERK1/2 MAPKs, and anti-phospho-p44/42 antibody can be used to recognize the phosphorylated active MAPK forms in Arabidopsis (Liu et al., 2010; Yue et al., 2012). Hence, the endogenous kinase activity of MPK6 after SA treat-ment was determined using phospho-P44/42 MAPK antibody. For the immunoblot assay, anti-GFP antibody-bound proteins were precipitated by adding protein A–agarose beads (50 μl ml–1) to the protein extracts.
Assay of Trx h activityTrx h activity was detected using bovine insulin as a substrate in the presence of DTT. The reduction of insulin by thioredoxin led to cleavage of the interchain disulfide bridges and formed a white precipitate from the free B chain of insulin. This phenomenon was used to measure disulfide reduction, and the rate of precipitation was recorded at 650 nm (Cho et al., 2007). Approximately 0.5 g of Arabidopsis leaves was ground to a powder and extracted with 2.5 ml of buffer (50 mM Tris/HCl, pH 7.9, 0.5 mM PMSF, 1 mM EDTA) by stirring for 2 h at 4 °C. The extracts were centrifuged at 25 000g for 20 min at 4 °C and the supernatants removed. Powdered (NH4)2SO4 was added to a final concentration of 30–90% satura-tion and the samples stirred for 16 h at 4 °C. This was centrifuged as above and the supernatants removed to a 10 000 Da molecular weight cut-off dialysis bag in the sucrose solutions environment and then purified by Sephadex G-50 gel at 4 °C. The mixture contained 100 mM Tris/HCl (pH 7.0), 2 mM EDTA, and 0.13 mM bovine insulin with or without 20 μl of Trx h extracted as above. The reac-tion was initiated by adding 0.33 mM DTT. Trx h activity was tested by measuring the turbidity of the solution at 650 nm (Yamazaki et al., 2004).
Results
Loss of MPK6 function delays SA-induced leaf senescence
MPK6 was shown to act as an SA-mediated defence regula-tor during systemic acquired resistance signal transmission and also to play a vital role in regulating programmed cell
at South China N
ormal U
nviversity on Novem
ber 28, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

6516 | Chai et al.
death. Whether the SA-induced senescence phenotype in detached leaves of WT plants alters in mpk6 mutant is unclear (Sharabi-Schwager et al., 2010). To compare the response of WT and the mpk6 mutant to the senescence stimulon, SA treatment was applied for the times indicated, after which various senescence symptoms were looked for.
After 3 d of incubation, the detached leaves showed more serious yellowing in the SA-treated WT plants than in the mpk6-3 mutants (Fig. 1A) and no kinase activity of MPK6 was observed in SA-treated detached leaves in the mpk6-3 mutants (Supplementary Fig. S2 at JXB online). Upon expo-sure to 100 μM SA, the chlorophyll content in the WT leaves decreased by 27% after 2 d and by 75% after 4 d. In contrast, in the mpk6-3 mutants, chlorophyll content was reduced by just 16% after 2 d and by 46% after 4 d (Fig. 1B). At the same time, we examined the Fv/Fm ratio, which reflects the photochemical efficiency of PSII. The decreasing trend in the Fv/Fm ratio showed a symptom similar to chlorophyll con-tent. Upon treatment with SA, the Fv/Fm ratio of WT leaves decreased by 22% after 2 d and by 64% after 4 d, whereas after the same incubation times, the mpk6-3 mutant leaves retained Fv/Fm ratios of 90 and 70%, respectively (Fig. 1C). As a con-trol, the two parameters in both WT and mpk6-3 leaves were examined without incubation with SA. The resulting changes were not obviously different. To check whether mutation of MPK6 affected the expression of senescence-associated genes, the mRNA levels of senescence marker genes (SAG12, SAG18, SAG20, and SIRK) were examined. After treatment with SA, the level of transcripts was higher in the WT than in the mpk6-3 mutant plants (Fig. 1D–G). Inhibition of senes-cence in the mpk6 mutants indicated that in Arabidopsis a deficiency of MPK6 delayed leaf senescence induced by SA.
Effects of SA on the altered expression of WRKY6 in mpk6 mutant leaves
A further study was carried out to investigate the mechanism of the delayed SA-induced leaf senescence by deficiency of MPK6. Four WRKYs (WRKY3, WRKY4, WRKY6, and WRKY72) are regulated by MPK3/MPK6 during pathogen defence responses. Among these WRKYs, WRKY6 has been analysed in more detail, and has been found to be involved in senescence, as well as being induced by wounding and by treatment with SA, jasmonic acid, and ethylene (Robatzek and Somssich, 2001, 2002; van den Burg and Takken, 2010). To determine whether WRKY6 was regulated by the MKK–MPK modules in SA-induced leaf senescence, we examined the expression of WRKY6 in the presence of the common MAPK inhibitor PD98059. As shown in Fig. 2B, detached WT leaves pre-treated with PD98059 displayed a lower expres-sion level of WRKY6 compared with those treated with SA only. As expected, plants pre-treated with PD98059 displayed delayed senescence phenotypes compared with WT plants (Fig. 2A). These results demonstrated that MAPK cascades are involved in SA-induced WRKY6 gene expression. Among a large number of MAPKs, MPK3, MPK4, and MPK6 have been studied in more detail and shown to be involved in Arabidopsis stress responses and programmed cell death.
To validate which specific MAPKs mediate SA-induced gene expression of WRKY6, qRT-PCR was performed on SA-treated WT, mpk3-1, mpk4-2, and mpk6-3 plants. Only mpk6-3 exhibited decreased expression of WRKY6 after exposed to SA compared with the other plants described above (Fig. 2C). This indicated that MPK6 may be the main MAPK involved in SA-triggered WRKY6 expression.
WRKY6 positively regulates SA-induced leaf senescence
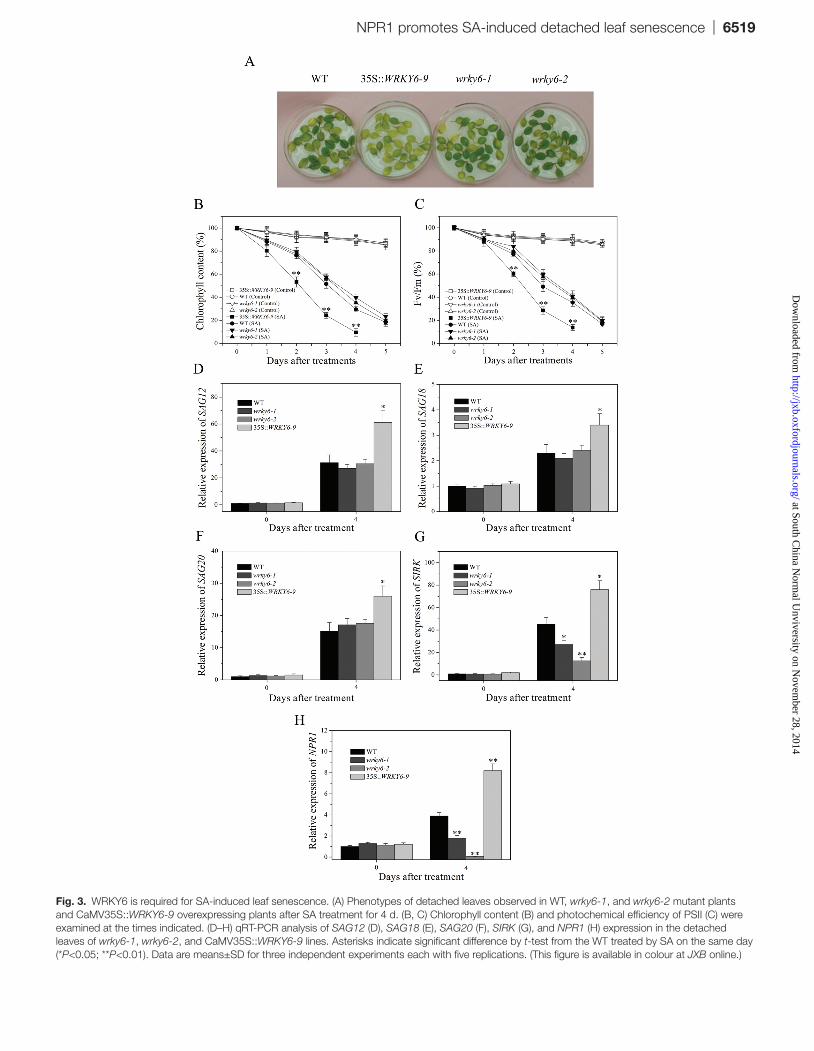
WRKY6 is reportedly upregulated as a transcription fac-tor during age-dependent leaf senescence and also by treat-ment with jasmonic acid, SA, and ethylene. This suggests that it is involved in senescence programmes mediated by both natural and unnatural stresses (Kim et al., 2008). It has been demonstrated that MPK6 regulates the mRNA level of WRKY6 with SA treatment. To confirm that MPK6 functions as an activator of SA-induced leaf senescence by regulating the gene expression of WRKY6, the effect of WRKY6 on SA-induced leaf senescence was examined. We used the CaMV 35S::WRKY6-9 overexpressing lines, wrky6-1 and wrky6-2 mutants to measure the physiological phenotype of detached leaves in response to SA. Under SA treatment, the detached leaves from CaMV 35S::WRKY6-9 overexpressing plants showed more severe senescence phe-notypes compared with WT plants. However, there was lit-tle observed phenotypic difference between WT plants and wrky6 mutants after SA treatment (Fig. 3A). Chlorophyll contents and the Fv/Fm ratio were also is coincident with the phenotypes (Figs 3B, C). A possible reason for this might be that partial functional redundancy exists within multigene families of WRKY. Correspondingly, the tran-script levels of senescence marker genes were higher in the CaMV 35S::WRKY6-9 line than in WT plants after 4 d of incubation with SA (Fig. 3D–G). The relative RNA levels of SIRK were lower in wrky6 mutants than in WT plants, while there was no significant difference in SAG12, SAG18, and SAG20 levels between wrky6 mutants and WT plants under SA treatment (Fig. 3D–G). Taken together, these results led to the conclusion that WRKY6 is involved in SA-induced leaf senescence.
PR1 has previously been shown to be a target of WRKY6, and the expression level of WRKY6 might determine whether transcription of the target gene is stimulated or repressed. The process of PR1 gene expression promoted by WRKY6 mostly involves the function of NPR1 (Robatzek and Somssich, 2002). To verify the function of WRKY6 on NPR1, previously described CaMV 35S::WRKY6-9, wrky6-1, and wrky6-2 plants were used to measure the gene expres-sion of NPR1. In contrast to WT plants, elevated NPR1 mRNA levels were detected in SA-treated detached leaves of WRKY6 overexpressing lines, while wrky6 mutants showed decreased expressions of NPR1 (Fig. 3H). Based on these results, MPK6-mediated WRKY6 expression was shown to act upstream of NPR1 and is likely to be involved in regu-lating its gene expression in the process of SA-induced leaf senescence.
at South China N
ormal U
nviversity on Novem
ber 28, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

NPR1 promotes SA-induced detached leaf senescence | 6517
NPR1 acts as a SA inducible protein involved in leaf senescence
It has been shown that npr1 mutants exhibit decreased expression of the PR1 gene, which is defined as a senescence
and pathogen-defence associated gene, after SA treatment (Robatzek and Somssich, 2002). Therefore, we speculated that NPR1 might participate in exogenous SA-induced leaf senes-cence. To explore the function of NPR1 in leaf senescence,
Fig. 1. Delayed senescence phenotype in detached leaves of mpk6-3 mutant plants upon SA treatment. (A) Phenotypes of detached leaves observed in WT and mpk6-3 mutant plants after SA treatment for 0–5 d. (B, C) Chlorophyll content (B) and photochemical efficiency of PSII (C) were examined in leaves at the times indicated. (D–G) qRT-PCR analysis of the senescence marker genes SAG12 (D), SAG18 (E), SAG20 (F), and SIRK (G) in WT and mpk6-3 leaves. Leaves were treated with 100 µM SA, and total RNA was extracted from control and SA-treated leaves. All measurements were carried out on the detached fourth and fifth rosette leaves harvested 21 d after sowing. Asterisks indicate significant difference by t-test from the WT treated by SA on the same day (*P<0.05; **P<0.01). Data are means±standard deviation (SD) for three independent experiments each with five replications. (This figure is available in colour at JXB online.)
at South China N
ormal U
nviversity on Novem
ber 28, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

6518 | Chai et al.
typical physiological markers of senescence were examined in detached leaves of WT and npr1 mutants.
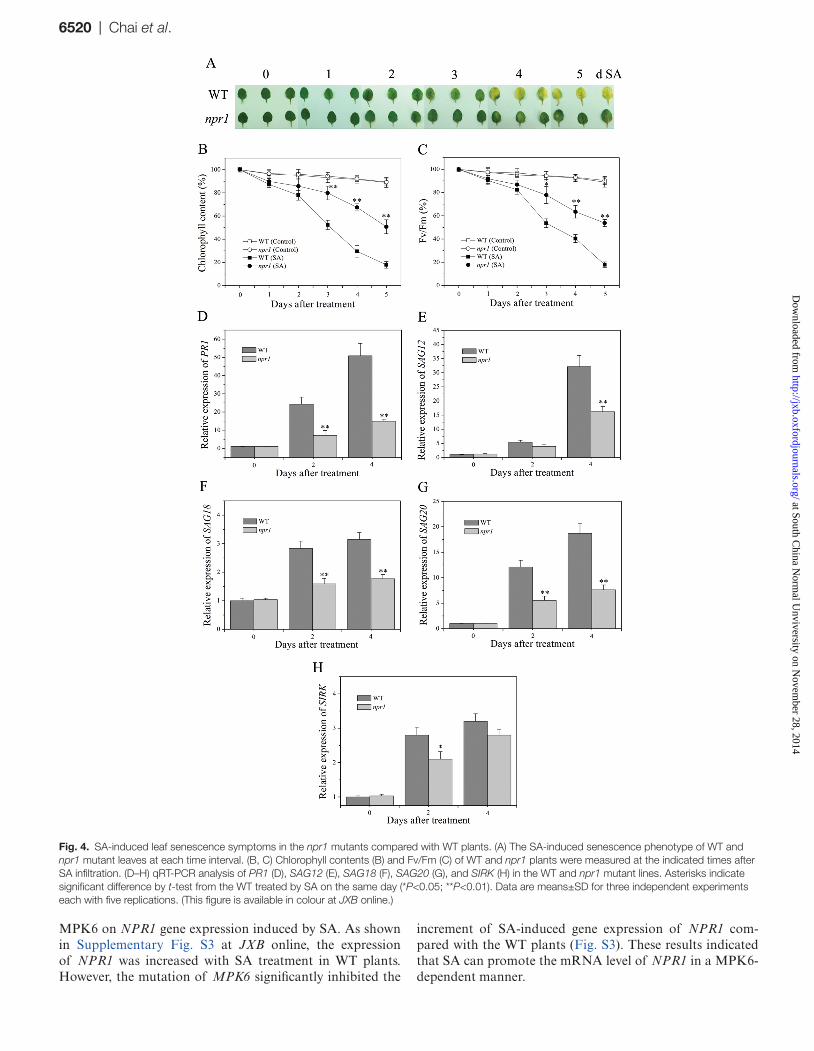
After 3 d of treatment, the expanding leaves of npr1 mutant plants displayed delayed senescence phenotypes compared with WT plants (Fig. 4A). Senescence symptoms were also assayed by measuring typical senescence param-eters as described above. Similarly, the levels of these mark-ers declined more rapidly in WT plants than in npr1 mutants. Upon treatment with SA, the remaining chlorophyll content
in WT leaves was 80% after 2 d and 30% after 4 d, whereas in npr1 leaves it was reduced to 86% after 2 d and to 67% after 4 d (Fig. 4B). After incubation with SA for 2 d, the pho-tochemical efficiency of WT leaves decreased to 82% and to 40% after 4 d, whereas the same values for the npr1 mutant leaves were 87 and 63% after 2 and 4 d, respectively (Fig. 4C). Correspondingly, the transcript levels of PR1 and senescence maker genes were lower in npr1 mutants than in WT plants (Fig. 4D–H). These results suggested that NPR1 plays an important role in SA-mediated leaf senescence.
WRKY6 binds to W-boxes in the promoter of NPR1
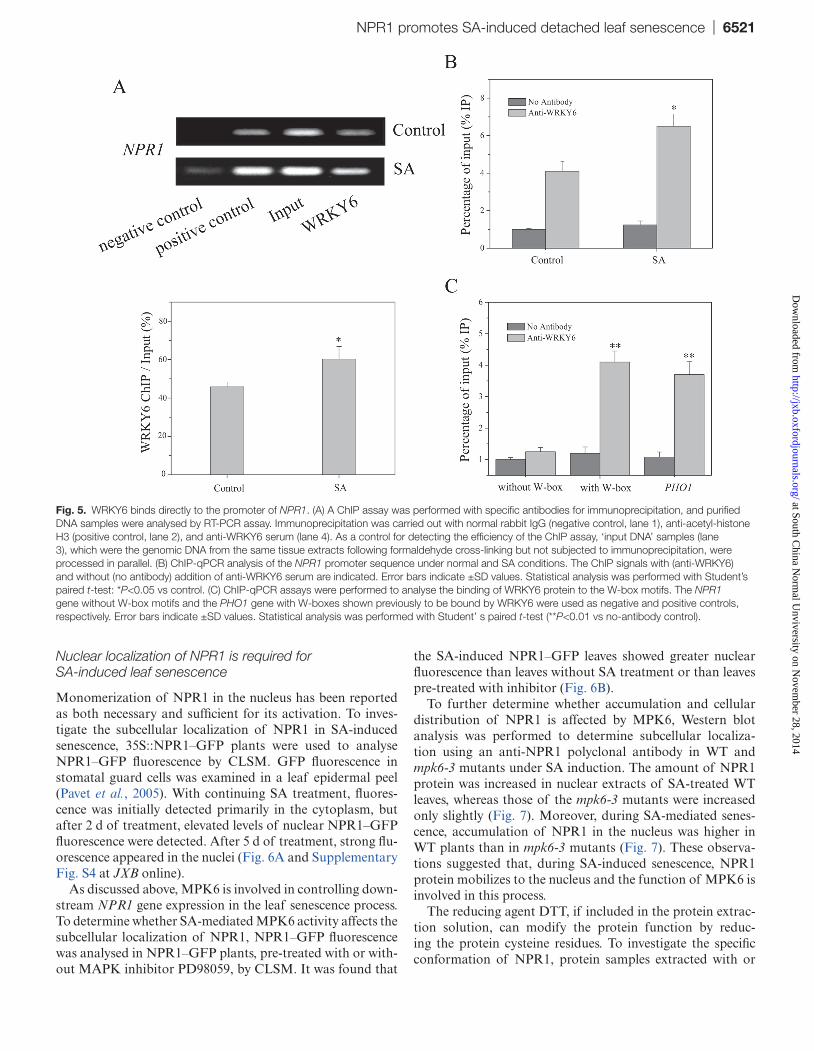
The three W-box motifs involved in the promoter of the NPR1 gene, which are known to be cis-acting elements of WRKYs, play an essential role in regulating NPR1 expression (Yu et al., 2001). We proved that WRKY6 upregulated NPR1 expres-sion in SA-induced leaf senescence. To investigate whether the NPR1 promoter was bound specifically by WRKY6, ChIP was performed to test the DNA-binding activity of WRKY6 to the W-boxes within the NPR1 promoter in vivo.
For the ChIP assay, WT leaves with or without SA treat-ment were first fixed with 1% formaldehyde for cross-linking of protein to the DNA, resulting in DNA–protein complexes. The cross-linked chromatins were sonicated into suitably sized frag-ments to expose the target proteins for subsequent recognition by their corresponding antibodies. After immunoprecipitation of the DNA–protein complexes, primers of the NPR1 gene encompassing the W-boxes within the promoter region were used to analyse the precipitated DNA fragments by RT-PCR. ChIP with either anti-acetyl-histone 3 or anti-WRKY6 anti-body yielded detectable NPR1 promoter amplification prod-ucts (Fig. 5A, lanes 2 and 4, respectively). However, almost no product was detected when ChIP with anti-rabbit IgG (Fig. 5A, lane 1). As a control for ChIP efficiency, the input DNA sam-ples (Fig. 5A, lane 3), the genomic DNA from chromatin preparations before immunoprecipitation, showed the bright-est band. In addition, ChIP-qPCR experiments show that the interaction of the WRKY6 protein and W-boxes within the NPR1 promoter was enhanced under SA conditions (Fig. 5B). As shown in Fig. 5C, WRKY6 strongly interacted with the NPR1 promoter encompassing the W-box motifs and the known WRKY6-binding sequences of the PHO1 promoter, whereas no interaction was observed between WRKY6 and the NPR1 promoter without the W-box motifs. These results proved that WRKY6, as a senescence-associated transcription factor, can bind to W-box motifs within the NPR1 promoter. Moreover, the NPR1 mRNA level was increased in the CaMV 35S::WRKY6-9 line and decreased in the wrky6-1 and wrky6-2 mutants (Fig. 3H). Taken together, upon treatment with SA, WRKY6 positively regulates NPR1 expression by directly binding to the W-boxes upstream of the NPR1 promoter.
The mpk6 mutant shows suppressive transcription of NPR1 under SA treatment
Since the mRNA level of NPR1 was regulated by WRKY6 and MPK6 could influence the expression of WRKY6 with SA treatment, it was interesting to examine the effects of
Fig. 2. MPK6 promotes WRKY6 expression in SA-induced leaf senescence. (A) The SA-induced senescence phenotype was alleviated by pre-treatment with PD98059 in the WT leaves. (B) qRT-PCR analysis of WRKY6 expression with or without PD98059 pre-treated detached WT leaves. Detached leaves were pre-incubated with or without PD98059 (150 μM), and then treated with SA for 4 d. (C) qRT-PCR analysis of WRKY6 expression in WT, mpk3-1, mpk4-2, and mpk6-3 plants. Asterisks indicate significant difference by t-test from the WT treated by SA on the same day (*P<0.05; **P<0.01). Data are means±SD for three independent experiments each with five replications. (This figure is available in colour at JXB online.)
at South China N
ormal U
nviversity on Novem
ber 28, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from
NPR1 promotes SA-induced detached leaf senescence | 6519
Fig. 3. WRKY6 is required for SA-induced leaf senescence. (A) Phenotypes of detached leaves observed in WT, wrky6-1, and wrky6-2 mutant plants and CaMV35S::WRKY6-9 overexpressing plants after SA treatment for 4 d. (B, C) Chlorophyll content (B) and photochemical efficiency of PSII (C) were examined at the times indicated. (D–H) qRT-PCR analysis of SAG12 (D), SAG18 (E), SAG20 (F), SIRK (G), and NPR1 (H) expression in the detached leaves of wrky6-1, wrky6-2, and CaMV35S::WRKY6-9 lines. Asterisks indicate significant difference by t-test from the WT treated by SA on the same day (*P<0.05; **P<0.01). Data are means±SD for three independent experiments each with five replications. (This figure is available in colour at JXB online.)
at South China N
ormal U
nviversity on Novem
ber 28, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from
6520 | Chai et al.
MPK6 on NPR1 gene expression induced by SA. As shown in Supplementary Fig. S3 at JXB online, the expression of NPR1 was increased with SA treatment in WT plants. However, the mutation of MPK6 significantly inhibited the
increment of SA-induced gene expression of NPR1 com-pared with the WT plants (Fig. S3). These results indicated that SA can promote the mRNA level of NPR1 in a MPK6-dependent manner.
Fig. 4. SA-induced leaf senescence symptoms in the npr1 mutants compared with WT plants. (A) The SA-induced senescence phenotype of WT and npr1 mutant leaves at each time interval. (B, C) Chlorophyll contents (B) and Fv/Fm (C) of WT and npr1 plants were measured at the indicated times after SA infiltration. (D–H) qRT-PCR analysis of PR1 (D), SAG12 (E), SAG18 (F), SAG20 (G), and SIRK (H) in the WT and npr1 mutant lines. Asterisks indicate significant difference by t-test from the WT treated by SA on the same day (*P<0.05; **P<0.01). Data are means±SD for three independent experiments each with five replications. (This figure is available in colour at JXB online.)
at South China N
ormal U
nviversity on Novem
ber 28, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from
NPR1 promotes SA-induced detached leaf senescence | 6521
Nuclear localization of NPR1 is required for SA-induced leaf senescence
Monomerization of NPR1 in the nucleus has been reported as both necessary and sufficient for its activation. To inves-tigate the subcellular localization of NPR1 in SA-induced senescence, 35S::NPR1–GFP plants were used to analyse NPR1–GFP fluorescence by CLSM. GFP fluorescence in stomatal guard cells was examined in a leaf epidermal peel (Pavet et al., 2005). With continuing SA treatment, fluores-cence was initially detected primarily in the cytoplasm, but after 2 d of treatment, elevated levels of nuclear NPR1–GFP fluorescence were detected. After 5 d of treatment, strong flu-orescence appeared in the nuclei (Fig. 6A and Supplementary Fig. S4 at JXB online).
As discussed above, MPK6 is involved in controlling down-stream NPR1 gene expression in the leaf senescence process. To determine whether SA-mediated MPK6 activity affects the subcellular localization of NPR1, NPR1–GFP fluorescence was analysed in NPR1–GFP plants, pre-treated with or with-out MAPK inhibitor PD98059, by CLSM. It was found that
the SA-induced NPR1–GFP leaves showed greater nuclear fluorescence than leaves without SA treatment or than leaves pre-treated with inhibitor (Fig. 6B).
To further determine whether accumulation and cellular distribution of NPR1 is affected by MPK6, Western blot analysis was performed to determine subcellular localiza-tion using an anti-NPR1 polyclonal antibody in WT and mpk6-3 mutants under SA induction. The amount of NPR1 protein was increased in nuclear extracts of SA-treated WT leaves, whereas those of the mpk6-3 mutants were increased only slightly (Fig. 7). Moreover, during SA-mediated senes-cence, accumulation of NPR1 in the nucleus was higher in WT plants than in mpk6-3 mutants (Fig. 7). These observa-tions suggested that, during SA-induced senescence, NPR1 protein mobilizes to the nucleus and the function of MPK6 is involved in this process.
The reducing agent DTT, if included in the protein extrac-tion solution, can modify the protein function by reduc-ing the protein cysteine residues. To investigate the specific conformation of NPR1, protein samples extracted with or
Fig. 5. WRKY6 binds directly to the promoter of NPR1. (A) A ChIP assay was performed with specific antibodies for immunoprecipitation, and purified DNA samples were analysed by RT-PCR assay. Immunoprecipitation was carried out with normal rabbit IgG (negative control, lane 1), anti-acetyl-histone H3 (positive control, lane 2), and anti-WRKY6 serum (lane 4). As a control for detecting the efficiency of the ChIP assay, ‘input DNA’ samples (lane 3), which were the genomic DNA from the same tissue extracts following formaldehyde cross-linking but not subjected to immunoprecipitation, were processed in parallel. (B) ChIP-qPCR analysis of the NPR1 promoter sequence under normal and SA conditions. The ChIP signals with (anti-WRKY6) and without (no antibody) addition of anti-WRKY6 serum are indicated. Error bars indicate ±SD values. Statistical analysis was performed with Student’s paired t-test: *P<0.05 vs control. (C) ChIP-qPCR assays were performed to analyse the binding of WRKY6 protein to the W-box motifs. The NPR1 gene without W-box motifs and the PHO1 gene with W-boxes shown previously to be bound by WRKY6 were used as negative and positive controls, respectively. Error bars indicate ±SD values. Statistical analysis was performed with Student’ s paired t-test (**P<0.01 vs no-antibody control).
at South China N
ormal U
nviversity on Novem
ber 28, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

6522 | Chai et al.
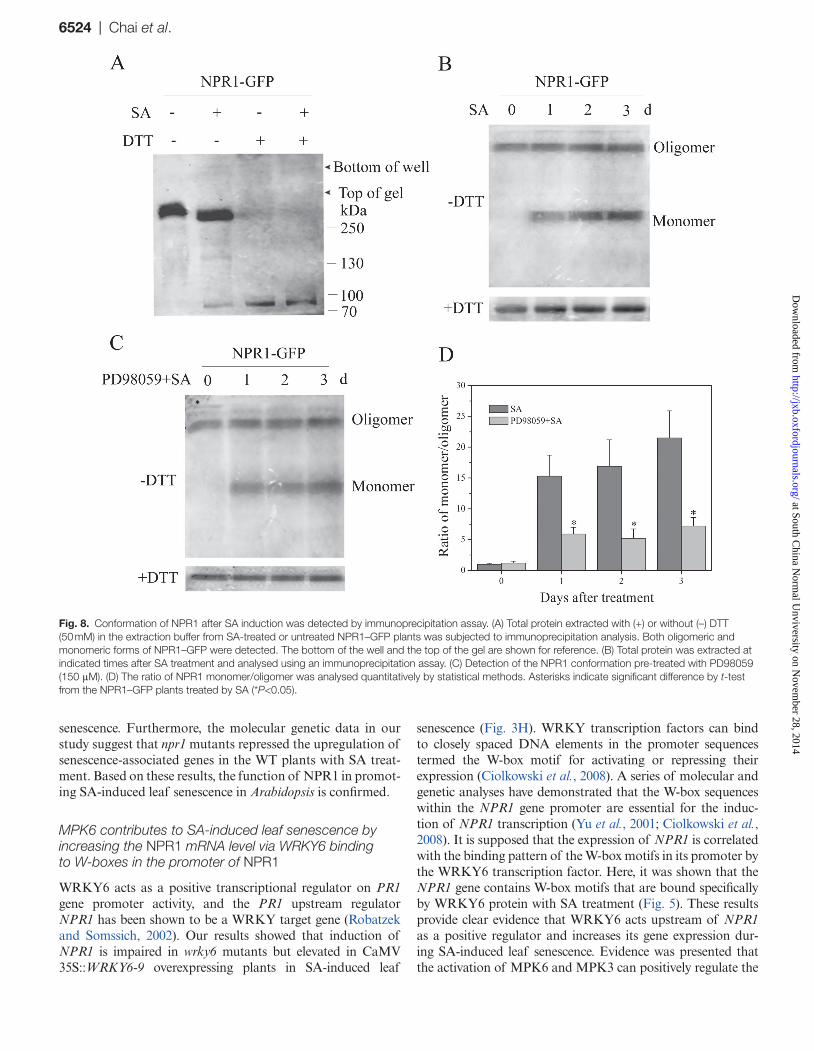
without DTT were subjected to immunoblot analysis. In the absence of DTT, the NPR1–GFP fusion protein was present at a lower molecular weight only in SA-treated samples and not in the untreated ones. Nevertheless, a higher-molecular-weight protein band was detected in both samples. The addi-tion of DTT to both samples reduced all of the NPR1–GFP protein to the monomeric form (Fig. 8A). Following SA treatment, monomeric NPR1–GFP was strongly increased compared with samples pre-treated with PD98059 (Fig. 8B–D). It was concluded that MPK6 is involved in regulating
both the monomerization and the movement of NPR1 dur-ing SA-mediated senescence of Arabidopsis leaves.
Influence of MPK6 on the activation of Trx h upon SA treatment
Cytosolic Trx h3 and Trx h5 are essential to promote mono-merization of NPR1, and the expression of Trx h5 is associ-ated with senescence (Laloi et al., 2004). To test whether the activation of NPR1 by MPK6 involves redox regulation, the
Fig. 6. Effect of MAPK cascade inhibitor (PD98059) on SA-induced nuclear localization of NPR1–GFP. (A) Confocal microscope images of NPR1–GFP fluorescence in representative pairs of guard cells by an epidermal strip bioassay. Detached leaves (21 d) floated in 3 mM MES buffer were used as a control. NPR1–GFP fluorescence in guard cells was observed by CLSM. Representative results are shown. Bars, 10 µm. (B) NPR1–GFP transgenetic plants pre-treated with or without PD98059 (150 μM) were incubated with SA for 4 d. Bars, 10 μm. Arrows indicate the nucleus. For each treatment, fluorescence and bright-field images are shown. (This figure is available in colour at JXB online.)
at South China N
ormal U
nviversity on Novem
ber 28, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

NPR1 promotes SA-induced detached leaf senescence | 6523
expression and activation of Trx h was measured in both WT and mpk6-3 mutant plants.
qRT-PCR analyses revealed that the expression of both Trx h3 and Trx h5 was increased by SA treatment. Induction of Trx h5 expression was suppressed in leaves pre-treated with MAPK inhibitor PD98059, while Trx h3 expression showed no significant difference between the control and inhibitor pre-treated leaves (Fig. 9A). A similar phenom-enon was observed in the mpk6-3 mutant leaves (Fig. 9B).
Furthermore, activity of Trx h was detected. After SA treat-ment, lower levels of Trx h activation were observed in mpk6-3 lines compared with WT plants (Fig. 9C). This indicated positive regulation of Trx h by MPK6. These results indicated that MPK6 is involved in regulating the activation of NPR1, possibly by enhancing the expression of Trx h5.
Discussion
Although much research has focused on the mechanisms of SA signal transduction in Arabidopsis, the specific mecha-nisms by which SA induces detached leaf senescence are not well understood. MPK6 has been reported to be involved in various biotic and abiotic stresses. The SA-responsive gene PR1 has been shown to be induced in senescent leaves and generally upregulated in WRKY6 overexpressing lines, dur-ing which the NPR1 function is probably involved (Robatzek and Somssich, 2002). Our study showed that MPK6 accel-erates SA-mediated leaf senescence not only by stimulating NPR1 gene expression but also by promoting the activation of NPR1.
NPR1 promotes leaf senescence induced by SA
The expression of the SA-responsive PR1 gene is induced during the onset of systemic acquired resistance and is also induced during leaf senescence (Morris et al., 2000; Robatzek and Somssich, 2002). Both the increase in the endogenous levels of SA and the exogenous application of SA coincide with the elevated expression of PR genes and enhanced resistance to pathogens (Dong et al., 2011). PR1 mRNA lev-els are prevented in the npr1 mutants, even after SA treat-ment, and nuclear localization of NPR1 is essential for PR1 gene induction (Dong, 2004). NPR1 protein is a key regu-latory component in pathogen defence reactions. Most of the senescence-enhanced gene expression is also induced in response to pathogen treatment, which reflects extensive over-lap between gene expression during senescence and in patho-gen reactions (Buchanan-Wollaston et al., 2003). Arabidopsis plants defective in SA signalling, such as npr1 and pad4 mutants and NahG transgenic plants, have altered expression of senescence-associated genes and exhibit delayed pheno-types of developmental leaf senescence (Morris et al., 2000; Mukherjee et al., 2010). However, it is still unclear whether NPR1 is involved in the induction of leaf senescence by exogenous SA.
In this study, it was found that, during SA incubation, the detached leaves of the npr1 mutant exhibited a delay in the appearance of senescence-correlative features compared with the WT plants (Fig. 4A–C). The NPR1 gene has previously been shown to influence the manifestation of cell death in two mutants, Arabidopsis accelerated cell death 5 (acd5) and accel-erated cell death 6 (acd6) (Rate et al., 1999; Greenberg et al., 2000). This provides additional support for the proposed role of NPR1 in the activation of senescence-associated processes. It was confirmed here that the mRNA level of NPR1 increases in response to exogenous SA (Supplementary Fig. S3). These findings imply that NPR1 is required for SA-induced leaf
Fig. 7. Cellular distribution of NPR1 in WT and mpk6-3 mutants during the process of SA-induced leaf senescence. (A, B) Western blot analyses of NPR1 in the nuclear-fractionated and cytoplasmic-fractionated protein extracted from leaves of WT and mpk6-3. The blot was probed with antibodies against NPR1. To determine the purity of the fractions, antibody against the cytosol and nuclear markers anti-PEPC and anti-histone H3, respectively, were used in each cytoplasmic and nuclear fraction. (C) Quantitative analyses of the results of NPR1 accumulation in the nucleus by statistical methods. Asterisks indicate significant difference by t-test from the WT treated by SA (*P<0.05).
at South China N
ormal U
nviversity on Novem
ber 28, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from
6524 | Chai et al.
senescence. Furthermore, the molecular genetic data in our study suggest that npr1 mutants repressed the upregulation of senescence-associated genes in the WT plants with SA treat-ment. Based on these results, the function of NPR1 in promot-ing SA-induced leaf senescence in Arabidopsis is confirmed.
MPK6 contributes to SA-induced leaf senescence by increasing the NPR1 mRNA level via WRKY6 binding to W-boxes in the promoter of NPR1
WRKY6 acts as a positive transcriptional regulator on PR1 gene promoter activity, and the PR1 upstream regulator NPR1 has been shown to be a WRKY target gene (Robatzek and Somssich, 2002). Our results showed that induction of NPR1 is impaired in wrky6 mutants but elevated in CaMV 35S::WRKY6-9 overexpressing plants in SA-induced leaf
senescence (Fig. 3H). WRKY transcription factors can bind to closely spaced DNA elements in the promoter sequences termed the W-box motif for activating or repressing their expression (Ciolkowski et al., 2008). A series of molecular and genetic analyses have demonstrated that the W-box sequences within the NPR1 gene promoter are essential for the induc-tion of NPR1 transcription (Yu et al., 2001; Ciolkowski et al., 2008). It is supposed that the expression of NPR1 is correlated with the binding pattern of the W-box motifs in its promoter by the WRKY6 transcription factor. Here, it was shown that the NPR1 gene contains W-box motifs that are bound specifically by WRKY6 protein with SA treatment (Fig. 5). These results provide clear evidence that WRKY6 acts upstream of NPR1 as a positive regulator and increases its gene expression dur-ing SA-induced leaf senescence. Evidence was presented that the activation of MPK6 and MPK3 can positively regulate the
Fig. 8. Conformation of NPR1 after SA induction was detected by immunoprecipitation assay. (A) Total protein extracted with (+) or without (–) DTT (50 mM) in the extraction buffer from SA-treated or untreated NPR1–GFP plants was subjected to immunoprecipitation analysis. Both oligomeric and monomeric forms of NPR1–GFP were detected. The bottom of the well and the top of the gel are shown for reference. (B) Total protein was extracted at indicated times after SA treatment and analysed using an immunoprecipitation assay. (C) Detection of the NPR1 conformation pre-treated with PD98059 (150 μM). (D) The ratio of NPR1 monomer/oligomer was analysed quantitatively by statistical methods. Asterisks indicate significant difference by t-test from the NPR1–GFP plants treated by SA (*P<0.05).
at South China N
ormal U
nviversity on Novem
ber 28, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

NPR1 promotes SA-induced detached leaf senescence | 6525
expression of particular defence genes, such as PR1, PR2, and PR5 (Ichimura et al., 2006). Except for the function in defence responses, MPK6 and its homologues have been shown to be activated in response to other abiotic stresses (Innes, 2001; Yue et al., 2012). Here, we found that exogenous SA promoted MPK6 activation in the process of leaf senescence (Supplementary Fig. S2). This indicates the vital role of MPK6 in SA-induced leaf
senescence. Moreover, detailed characterization of senescence indicates that SA-induced leaf senescence is positively regulated by MPK6 (Fig. 1). The receptor of SA in plants may be mem-bers of the Leu-rich repeat receptor-like protein kinase (LRR-RLK) family, which often contain C-X8-C-X2-C motifs in the putative extracellular domain (Wang and Irving, 2011). Plant LRR-RLK proteins that contain a ligand binding extracellular domain and are localized at the plasma membrane can perceive extracellular ligands, including hormones (Ohtake et al., 2000). The transcripts of numerous RLKs accumulate to a higher level in response to exogenous SA treatment of plants (Ohtake et al., 2000). In plants, LRR-RLKs act upstream of the MAPK cascade, and MAPKs transduce and amplify intracellular and extracellular stimuli into a wide range of intracellular responses (Shan et al., 2007). The possible signalling transduction medi-ated by exogenous SA in leaf senescence might act through SA receptor LRR-RLKs, leading to the activation of MAPK cas-cades. The precise mechanisms require further study.
However, how MPK6 exerts its functions in regulating leaf senescence remains unknown. The TIR-NBS-LRR type R protein SSI4 has been defined as a suppressor of SA insen-sitive. The Arabidopsis ssi4 mutant, which exhibits chlorotic morphology, induces MPK6 activation and then leads to elevated expression of WRKY6 and WRKY29 (Zhou et al., 2004). Among the several WRKYs that are regulated by MPK6 during pathogen defence responses, WRKY6 is found to be involved in plant senescence as well as being induced by treatment with various hormones (Robatzek and Somssich, 2001, 2002; van den Burg and Takken, 2010). It was confirmed here that the mRNA level of WRKY6 increases in conjunc-tion with SA-induced leaf senescence and MPK6 increases the expression of WRKY6 with SA treatment (Fig. 2B, C). It has been shown that WRKY6 is phosphorylated only when it is co-expressed with the MKK5/MPK10 module (Popescu et al., 2009). This indicates that the effect of MPK6 on WRKY6 gene expression may be indirect. This phenomenon is consist-ent with previous study that the increase in W-box binding activity after SIPK/WIPK (SIPK and WIPK are two tobacco MAPKs that are orthologues of Arabidopsis AtMPK6 and AtMPK3) activation is the result of WRKY gene activation instead of phosphorylation modification (Kim and Zhang, 2004). However, the precise mechanisms involved require further study. Having demonstrated that WRKY6 upregu-lated NPR1 expression in SA-induced leaf senescence, we here showed that the mRNA level of NPR1 was significantly suppressed in the mpk6 mutant plants under SA treatment (Supplementary Fig. S3). A signalling cascade is presented in our analyses whereby MPK6 participates in SA-induced Arabidopsis leaf senescence by increasing the transcriptional level of NPR1 via the WRKY6 transcription factor binding to W-box sequences within the promoter of NPR1 gene (Fig. 10).
MPK6-mediated activation of NPR1 via regulation of the expression of Trx h5 modulates SA-induced Arabidopsis leaf senescence
NPR1 is maintained in the cytoplasm as a disulfide-bonded oligomer and is translocated to the nucleus in an active
Fig. 9. Expression and activation of Trx h influenced by MPK6 following SA treatment. (A) qRT-PCR analysis of the Trx h3 and Trx h5 gene expression in PD98059-pre-treated detached leaves of WT plants. Detached leaves were pre-incubated with or without PD98059 (150 μM) and then treated with SA. (B) Expression of Trx h3 and Trx h5 in SA-treated WT and mpk6-3 mutant leaves. Statistical analyses were performed with Student’s paired t-test (*P<0.05 vs control; **P<0.01 vs control; #P<0.01 vs indicated samples). (C) Activity of Trx h was measured according to the procedure described in Materials and methods. Asterisks indicate significant differences (*P<0.05 or **P<0.01) by t-test from the WT treated with SA at the same time point of detection. Data are the means±SD for three independent experiments, each with five replications.
at South China N
ormal U
nviversity on Novem
ber 28, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

6526 | Chai et al.
monomeric form, which can function as a co-activator for downstream gene transcription (Spoel et al., 2009, 2010). Based on analysis of the mRNA levels of NPR1 induced by MPK6, we speculated that the activation of NPR1 may be influenced by MPK6. To understand better the regulation of NPR1, the subcellular localization of NPR1 was ana-lysed by detecting GFP fluorescence in NPR1–GFP plants. As shown in Fig. 6, enhanced nuclear NPR1–GFP fluores-cence was observed in the guard cells upon SA treatment, while pre-treatment with PD98059 decreased the accumula-tion of fluorescence in the nucleus. This is similar to a previ-ous report that strong NPR1–GFP fluorescence is observed in the nuclei of guard cells when seedlings are grown on Murashige and Skoog medium with SA, compared with on Murashige and Skoog medium alone (Kinkema et al., 2000). Previous studies have suggested that the increase in nuclear fluorescence observed in the SA-induced plants was due to the redistribution of NPR1–GFP to the nucleus (Kinkema et al., 2000). Our results based on subcellular fractiona-tion analyses revealed that nuclear accumulation of NPR1 is increased during SA-induced senescence. However, in the mpk6-3 mutants, it remained at a relatively constant level (Fig. 7), suggesting that MPK6 may be involved in the regu-lation of the nuclear localization of NPR1. The polyclonal antibody against NPR1 used in this study can recognize only the monomeric form of the protein; therefore, the conforma-tion of NPR1 was tested as reported previously in transgenic 35S::NPR1–GFP plants (Mou et al., 2003). It was found that increased monomerization of NPR1–GFP plants treated with SA was suppressed in plants pre-treated with PD98059 (Fig. 8). These results indicated that MPK6 is involved in the regulation of NPR1 activation in the process of SA-induced
leaf senescence. However, it is worth noting that the activa-tion of NPR1 was not completely suppressed in the mpk6-3 lines, so other factors may also regulate the NPR1 activity simultaneously. These ideas require further investigation.
The accumulation of endogenous SA, which is probably synthesized in chloroplasts, and application of exogenous SA can both induce changes in the cellular redox state (Dong et al., 2011). It has been shown that Trxs contain two redox-sensitive cysteine residues, which make it possible to dis-rupt disulfide bridges of target proteins and thus maintain the cellular redox equilibrium (Sweat and Wolpert, 2007). Furthermore, Trx h can mediate SA-induced conformational changes of NPR1 from oligomer to monomer, and can regu-late its nuclear localization (Ishikawa et al., 2010). The acti-vation of MAPK pathways can result in the induction of transcription factors and various redox-sensitive genes for animal cell growth, proliferation, and apoptosis (Cakir and Ballinger, 2005). In plants, data on the relationship between MAPK cascades and regulation of cellular redox state is scarce. To determine whether the effect of MPK6 on regulat-ing NPR1 activity is based on the regulation of Trx h, the expressions and activities of Trx h were investigated. The induction of Trx h5 expression was lower in the PD98059 pre-treated WT leaves and mpk6 mutants than in the SA-treated WT plants (Fig. 9A, B). This suggested that MPK6 contrib-utes to the expression of Trx h5, which involves maintaining a cellular redox equilibrium. Interestingly, the expression of the Trx h3 gene, the most close-related Trx to Trx h5, was not changed significantly under the above conditions (Fig. 9A, B). This indicated that the regulation and function of these two closely related genes is divergent, at least during SA-induced leaf senescence. On the other hand, Trx h activation was
Fig. 10. A model for MPK6-mediated gene expression and activation of NPR1 in SA-induced leaf senescence. (This figure is available in colour at JXB online.)
at South China N
ormal U
nviversity on Novem
ber 28, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

NPR1 promotes SA-induced detached leaf senescence | 6527
significantly attenuated in the mpk6-3 mutants compared with in the WT plants (Fig. 9C), indicating that MPK6 acts as a positive regulator of Trx h activity during SA-induced leaf senescence. Hence, during the process of SA-induced leaf senescence, the activation of NPR1 catalysed by Trx h requires the regulation of MPK6 (Fig. 10).
In conclusion, our studies show that, by regulating compo-nents of the SA pathways, MPK6 provides a mechanistic expla-nation as to how SA-mediated promotion of NPR1 is achieved during the course of leaf senescence. This is partly accomplished by the involvement of MPK6 for WRKY6 expression, while WRKY6 specifically recognizes W-box motifs in the NPR1 promoter. Also, by regulating the expression of a modulator of cellular redox status, Trx h5, which in turn leads to the mono-merization and nuclear localization of NPR1 protein. Taken together, these results indicate that the MPK6-modulated signalling pathway is responsible for the gene expression and activation of NPR1 in promoting SA-induced leaf senescence, providing new insights into the signalling cascades that modu-late leaf senescence promoted by exogenous SA.
Supplementary data
Supplementary data are available at JXB online.Supplementary Fig. S1. Identification of npr1, mpk3-
1, mpk4-2, wrky6-1, and wrky6-2 mutants and CaMV 35S::WRKY6-9 overexpressing plants using semi-quantita-tive RT-PCR analysis.
Supplementary Fig. S2. The activity of MPK6 during the process of leaf senescence induced by SA in the WT and mpk6-3 plants.
Supplementary Fig. S3. qRT-PCR analyses of the NPR1 mRNA level in SA-treated detached leaves of WT and mpk6-3 plants.
Supplementary Fig. S4. Nuclear localization of NPR1–GFP.
AcknowledgementsWe thank Dr Imre E. Somssich (the Max-Planck-Institute for Züchtungsforschung, Köln, Germany) for kindly providing seeds of Arabidopsis wrky6-1 and wrky6-2 knockout mutants and the CaMV 35S::WRKY6-9 overexpressing line. We thank Dr Xinnian Dong (Duke University, Durham, USA) for kindly providing seeds of the transgenic 35S::NPR1–GFP in the npr1 mutant of Arabidopsis thaliana. We also thank Professor Xinnian Dong and Professor Weihua Wu (China Agricultural University, Beijing, China) for kindly providing Arabidopsis thaliana NPR1 polyclonal antibody and rabbit anti-WRKY6 serum. This research is sup-ported by the Programs for Changjiang Scholars and Innovative Research Team in University (IRT0829), the Key Program of NSFC-Guangdong Joint Funds of China (U0931005), and the National High Technology Research and Development Program of China (863 Program) (2007AA10Z204).
ReferencesAbreu ME, Munné-Bosch S. 2008. Salicylic acid may be involved in the regulation of drought-induced leaf senescence in perennials: a case study in field-grown Salvia officinalis L. plants. Environmental and Experimental Botany 64, 105–112.
Bartels S, Anderson JC, Besteiro MAG, Carreri A, Hirt H, Buchala A, Métraux JP, Peck SC, Ulm R. 2009. MAP kinase phosphatase1 and
protein tyrosine phosphatase1 are repressors of salicylic acid synthesis and SNC1-mediated responses in Arabidopsis. Plant Cell 21, 2884–2897.
Buchanan-Wollaston V, Earl S, Harrison E, Mathas E, Navabpour S, Page T, Pink D. 2003. The molecular analysis of leaf senescence—a genomics approach. Plant Biotechnology Journal 1, 3–22.
Cakir Y, Ballinger SW. 2005. Reactive species-mediated regulation of cell signaling and the cell cycle: the role of MAPK. Antioxidants and Redox Signaling 7, 726–740.
Chen YF, Li LQ, Xu Q, Kong YH, Wang H, Wu WH. 2009. The WRKY6 transcription factor modulates PHOSPHATE1 expression in response to low Pi stress in Arabidopsis. Plant Cell 21, 3554–3566.
Cheng YT, Germain H, Wiermer M, et al. 2009. Nuclear pore complex component MOS7/Nup88 is required for innate immunity and nuclear accumulation of defense regulators in Arabidopsis. Plant Cell 21, 2503–2516.
Cho MJ, Wong JH, Lemaux PG, Buchanan BB. 2007. Transgenic plants with elevated thioredoxin levels. US Patent 7179964.
Ciolkowski I, Wanke D, Birkenbihl RP, Somssich IE. 2008. Studies on DNA-binding selectivity of WRKY transcription factors lend structural clues into WRKY-domain function. Plant Molecular Biology 68, 81–92.
Dong CJ, Wang XL, Shang QM. 2011. Salicylic acid regulates sugar metabolism that confers tolerance to salinity stress in cucumber seedlings. Scientia Horticulturae 129, 629–636.
Dong X. 2004. NPR1, all things considered. Current Opinion in Plant Biology 7, 547–552.
Gepstein S. 2004. Leaf senescence-not just a ‘wear and tear’ phenomenon. Genome Biology 5, 212–212.
Greenberg JT, Silverman FP, Liang H. 2000. Uncoupling salicylic acid-dependent cell death and defense-related responses from disease resistance in the Arabidopsis mutant acd5. Genetics 156, 341–350.
Habora MEE, Eltayeb AE, Tsujimoto H, Tanaka K. 2012. Identification of osmotic stress-responsive genes from Leymus mollis, a wild relative of wheat (Triticum aestivum L.). Breeding Science 62, 78–86.
Ichimura K, Casais C, Peck SC, Shinozaki K, Shirasu K. 2006. MEKK1 is required for MPK4 activation and regulates tissue-specific and temperature-dependent cell death in Arabidopsis. Journal of Biological Chemistry 281, 36969–36976.
Innes RW. 2001. Mapping out the roles of MAP kinases in plant defense. Trends in Plant Science 6, 392–394.
Ishikawa K, Yoshimura K, Harada K, Fukusaki E, Ogawa T, Tamoi M, Shigeoka S. 2010. AtNUDX6, an ADP-ribose/NADH pyrophosphohydrolase in Arabidopsis, positively regulates NPR1-dependent salicylic acid signaling. Plant Physiology 152, 2000–2012.
Jaradat MR, Feurtado JA, Huang D, Lu Y, Cutler AJ 2013. Multiple roles of the transcription factor AtMYBR1/AtMYB44 in ABA signaling, stress responses, and leaf senescence. BMC Plant Biology 13, 192.
Jiang Y, Liang G, Yang S, Yu D. 2014. Arabidopsis WRKY57 functions as a node of convergence for jasmonic acid- and auxin-mediated signaling in jasmonic acid-induced leaf senescence. Plant Cell 113, 117838.
Kim CY, Bove J, Assmann SM 2008. Overexpression of wound-responsive RNA-binding proteins induces leaf senescence and hypersensitive-like cell death. New Phytologist 180, 57–70.
Kim CY, Zhang S. 2004. Activation of a mitogen-activated protein kinase cascade induces WRKY family of transcription factors and defense genes in tobacco. The Plant Journal 38, 142–151.
Kinkema M, Fan W, Dong X. 2000. Nuclear localization of NPR1 is required for activation of PR gene expression. Plant Cell 12, 2339–2350.
Krinke O, Ruelland E, Valentová O, Vergnolle C, Renou JP, Taconnat L, Flemr M, Burketová L, Zachowski A. 2007. Phosphatidylinositol 4-kinase activation is an early response to salicylic acid in Arabidopsis suspension cells. Plant Physiology 144, 1347–1359.
Laloi C, Mestres-Ortega D, Marco Y, Meyer Y, Reichheld JP 2004. The Arabidopsis cytosolic thioredoxin h5 gene induction by oxidative stress and its W-box-mediated response to pathogen elicitor. Plant Physiology 134, 1006–1016.
Li Z, Yue H, Xing D. 2012. MAP kinase 6-mediated activation of vacuolar processing enzyme modulates heat shock-induced programmed cell death in Arabidopsis. New Phytologist 195, 85–96.
Lichtenthaler HK. 1987. Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Methods in Enzymology 148, 350–382.
at South China N
ormal U
nviversity on Novem
ber 28, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

6528 | Chai et al.
Lim PO, Woo HR, Nam HG. 2003. Molecular genetics of leaf senescence in Arabidopsis. Trends in Plant Science 8, 272–278.
Liu XM, Kim KE, Kim KC, et al. 2010. Cadmium activates Arabidopsis MPK3 and MPK6 via accumulation of reactive oxygen species. Phytochemistry 71, 614–618.
Love AJ, Geri C, Laird J, Carr C, Yun BW, Loake GJ, Tada Y, Sadanandom A, Milner JJ. 2012. Cauliflower mosaic virus protein P6 inhibits signaling responses to salicylic acid and regulates innate immunity. PLoS One 7, e47535.
Mao P, Duan M, Wei C, Li Y. 2007. WRKY62 transcription factor acts downstream of cytosolic NPR1 and negatively regulates jasmonate-responsive gene expression. Plant and Cell Physiology 48, 833–842.
Morris K, Mackerness S AH, Page T, John CF, Murphy AM, Carr JP, Buchanan-Wollaston V. 2000. Salicylic acid has a role in regulating gene expression during leaf senescence. The Plant Journal 23, 677–685.
Mou Z, Fan W, Dong X. 2003. Inducers of plant systemic acquired resistance regulate NPR1 function through redox changes. Cell 113, 935–944.
Mukherjee M, Larrimore KE, Ahmed NJ, Bedick TS, Barghouthi NT, Traw MB, Barth C. 2010. Ascorbic acid deficiency in Arabidopsis induces constitutive priming that is dependent on hydrogen peroxide, salicylic acid, and the NPR1 gene. Molecular Plant–Microbe Interactions 23, 340–351.
Nelson JD, Denisenko O, Bomsztyk K. 2006. Protocol for the fast chromatin immunoprecipitation (ChIP) method. Nature Protocols 1, 179–185.
Ogawa KI. 2005. Glutathione-associated regulation of plant growth and stress responses. Antioxidants and Redox Signaling 7, 973–981.
Oh SA, Lee SY, Chung IK, Lee CH, Nam HG. 1996. A senescence-associated gene of Arabidopsis thaliana is distinctively regulated during natural and artificially induced leaf senescence. Plant Molecular Biology 30, 739–754.
Ohtake Y, Takahashi T, Komeda Y. 2000. Salicylic acid induces the expression of a number of receptor-like kinase genes in Arabidopsis thaliana. Plant and Cell Physiology 41, 1038–1044.
Pavet V, Olmos E, Kiddle G, Mowla S, Kumar S, Antoniw J, Alvarez ME, Foyer CH. 2005. Ascorbic acid deficiency activates cell death and disease resistance responses in Arabidopsis. Plant Physiology 139, 1291–1303.
Popescu SC, Popescu GV, Bachan S, Zhang Z, Gerstein M, Snyder M, Dinesh-Kumar SP. 2009. MAPK target networks in Arabidopsis thaliana revealed using functional protein microarrays. Genes and Development 23, 80–92.
Ralph PJ, Schreiber U, Gademann R, Kühl M, Larkum AW. 2005. Coral photobiology studied with a new imaging pulse amplitude modulated fluorometer. Journal of Phycology 41, 335–342.
Rate DN, Cuenca JV, Bowman GR, Guttman DS, Greenberg JT. 1999. The gain-of-function Arabidopsis acd6 mutant reveals novel regulation and function of the salicylic acid signaling pathway in controlling cell death, defenses, and cell growth. Plant Cell 11, 1695–1708.
Robatzek S, Somssich IE. 2001. A new member of the Arabidopsis WRKY transcription factor family, AtWRKY6, is associated with both senescence- and defence-related processes. Plant Journal 28, 123–133.
Robatzek S, Somssich IE. 2002. Targets of AtWRKY6 regulation during plant senescence and pathogen defense. Genes and Development 16, 1139–1149.
Schippers JHM. 2008. Molecular aspects of ageing and the onset of leaf senescence. PhD thesis, University of Groningen, Groningen, The Netherlands.
Shan L, He P, Sheen J. 2007. Intercepting host MAPK signaling cascades by bacterial type III effectors. Cell Host & Microbe 1, 167–174.
Sharabi-Schwager M, Lers A, Samach A, Guy CL, Porat R. 2010. Overexpression of the CBF2 transcriptional activator in Arabidopsis delays leaf senescence and extends plant longevity. Journal of Experimental Botany 61, 261–273.
Spoel SH, Mou Z, Tada Y, Spivey NW, Genschik P, Dong X. 2009. Proteasome-mediated turnover of the transcription coactivator NPR1 plays dual roles in regulating plant immunity. Cell 137, 860–872.
Spoel SH, Tada Y, Loake GJ. 2010. Post-translational protein modification as a tool for transcription reprogramming. New Phytologist 186, 333–339.
Sun A, Nie S, Xing D. 2012. Nitric oxide-mediated maintenance of redox homeostasis contributes to NPR1-dependent plant innate immunity triggered by lipopolysaccharides. Plant Physiology 160, 1081–1096.
Sweat TA, Wolpert TJ. 2007. Thioredoxin h5 is required for victorin sensitivity mediated by a CC-NBS-LRR gene in Arabidopsis. Plant Cell 19, 673–687.
van den Burg HA, Takken FL. 2010. SUMO- MAPK-, and resistance protein-signaling converge at transcription complexes that regulate plant innate immunity. Plant Signaling and Behavior 5, 1597–1601.
Wang YH, Irving HR. 2011. Developing a model of plant hormone interactions. Plant Signaling and Behavior 6, 494–500.
Running MP, Clark SE, Meyerowitz EM. 1995. Confocal microscopy of the shoot apex. In: Wilson L, Matsudaira PT, Galbraith DW, Bohnert HJ, Bourque DP, eds. Methods in plant cell biology, vol. 49. San Diego, CA: Academic Press, 217–229.
Yamazaki D, Motohashi K, Kasama T, Hara Y, Hisabori T. 2004. Target proteins of the cytosolic thioredoxins in Arabidopsis thaliana. Plant and Cell Physiology 45, 18–27.
Yu D, Chen C, Chen Z. 2001. Evidence for an important role of WRKY DNA binding proteins in the regulation of NPR1 gene expression. Plant Cell 13, 1527–1540.
Yue H, Nie S, Xing D. 2012. Over-expression of Arabidopsis Bax inhibitor-1 delays methyl jasmonate-induced leaf senescence by suppressing the activation of MAP kinase 6. Journal of Experimental Botany 63, 4463–4474.
Zhang B, Jia J, Yang M, Yan C, Han Y. 2012. Overexpression of a LAM domain containing RNA-binding protein LARP1c induces precocious leaf senescence in Arabidopsis. Molecules and Cells 34, 367–374.
Zhou C, Cai Z, Guo Y, Gan S. 2009. An Arabidopsis mitogen-activated protein kinase cascade MKK9-MPK6, plays a role in leaf senescence. Plant Physiology 150, 167–177.
Zhou F, Menke FL, Yoshioka K, Moder W, Shirano Y, Klessig DF. 2004. High humidity suppresses ssi4-mediated cell death and disease resistance upstream of MAP kinase activation, H2O2 production and defense gene expression. The Plant Journal 39, 920–932.
Zhou X, Jiang Y, Yu D. 2011. WRKY22 transcription factor mediates dark-induced leaf senescence in Arabidopsis. Molecules and Cells 31, 303–313.
at South China N
ormal U
nviversity on Novem
ber 28, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from














