Embed Size (px)
Citation preview

MICROVASCULAR PATTERNS OF BLUBBER IN SHALLOW AND DEEP DIVING
ODONTOCETES
Sara J. McClelland
A Thesis Submitted to the
University of North Carolina Wilmington in Partial Fulfillment
of the Requirements for the Degree of
Master of Science
Department of Biology and Marine Biology
University of North Carolina Wilmington
2010
Approved by
Advisory committee
Richard M. Dillaman D. Ann Pabst
Heather N. Koopman
Chair
Accepted by
Dean, Graduate School

TABLE OF CONTENTS
ABSTRACT...................................................................................................................................iii
ACKNOWLEDGMENTS...............................................................................................................v
DEDICATION...............................................................................................................................vii
LIST OF TABLES.......................................................................................................................viii
LIST OF FIGURES........................................................................................................................ix
INTRODUCTION...........................................................................................................................1
METHODS......................................................................................................................................6
Tissue Collection.................................................................................................................6
Histological Analysis...........................................................................................................7
Lipid Analysis....................................................................................................................12
Fatty Acid Stratification.....................................................................................................13
Statistics.............................................................................................................................13
RESULTS......................................................................................................................................14
Histological Analysis.........................................................................................................14
Lipid Analysis....................................................................................................................15
DISCUSSION................................................................................................................................16
Vascular Distribution and Morphology.............................................................................17
Relationship of Vascular Patterns to Biochemical Characteristics of Blubber..................18
Potential Influences on Blubber Vasculature Structure.....................................................22
Conclusions and Future Directions....................................................................................27
LITERATURE CITED..................................................................................................................29

ABSTRACT
Blubber is the hypertrophied subdermal adipose layer surrounding the bodies of marine
mammals and is an important and dynamic living tissue. Typically, adipose tissue is perfused by
networks of capillaries. Surprisingly, there is little information on the vascularization of blubber.
The goal of this study was to describe and characterize, for the first time, the microvasculature
(capillaries, microarterioles and microvenules) of the blubber of marine mammals, using toothed
whales (Odontoceti) as models. Marine mammals experience vasoconstriction during a dive as
part of the dive response, resulting in decreased blood flow to peripheral tissues (including
blubber). I predicted that deeper divers would have reduced vasculature in their blubber, since
they theoretically would not perfuse their blubber vasculature while on a dive. I used
histochemical techniques and digital image analysis to estimate the density and distribution of
the microvasculature in the blubber of two species of odontocetes, the shallow-diving bottlenose
dolphin (Tursiops truncatus; n=6), and deeper-diving pygmy sperm whale (Kogia breviceps;
n=6). Tursiops blubber showed significant differences in the density of the microvasculature
among the superficial (3.26±0.39%), middle (8.40±0.65%) and deep (9.31±0.72%) layers. Kogia
blubber differed, with its microvascular density more uniformly distributed across the blubber
(superficial 3.26±0.53%, middle 4.38±0.55%, deep 4.50±0.54%). Overall, Tursiops had
significantly (P<0.001) higher microvascular densities than Kogia, with this difference largely
due to the higher values in the inner and middle layers. Two beaked whales (Mesoplodon
densirostris, n=1; Ziphius cavirostris, n=1) examined had microvascular densities much lower
than either Tursiops or Kogia , with a relatively uniform distribution (all layers with values
between 1.5 and 2.8%). To place the data in a broader mammalian context, I also examined
microvascular density in the subcutaneous adipose tissue of the domestic pig (Sus scrofa, n=4).
The pig subdermal adipose exhibited no stratification and comparatively low vessel densities

(superficial 1.75±0.22%, middle 1.87±0.22%, deep 1.88±0.46%). Vascular data were then
compared with biochemical features of the blubber (lipid content and composition) to identify
any links between vessel distribution and lipids. Overall, the species that had the lowest lipid
content (Tursiops ~46% overall) had the highest microvascular densities. The species with the
highest lipid content (beaked whales ~80% overall) had the lowest microvascular densities, and
Kogia was intermediate in both categories. Lipid class also appeared to correlate with the
microvascular densities in blubber; Kogia and the beaked whales had blubber composed mainly
of wax esters and had the lowest microvascular densities. Pigs had low microvascular densities
but had no wax esters; the relationship between lipid class and vascularity is likely not direct.
The deepest divers had the lowest microvascular densities and the shallow-divers had the most
microvascular densities of the odontocetes analyzed to date. While dive depth and microvascular
densities appear to be correlated, the conserved nature of the microvascularity in the blubber of
beaked whales suggests that the microvascular densities in blubber is not a direct consequence of
dive depth. It is likely that other factor(s), such as energetics or thermoregulation, may be
influencing the vessel densities in the blubber of marine mammals.

ACKNOWLEDGEMENTS
I could not have finished this thesis without the help and support of many people, all of
whom deserve to be acknowledged and to receive a very special thank you. First and foremost, I
want to thank my advisor, Dr. Heather Koopman. Her guidance and support have carried me
through the difficulties that come with graduate school and completing a thesis. She has gone
above and beyond the duties of a mentor in helping me think through ideas, work on writing,
become better at public speaking, and just overall teaching me about science. More important
than all of that, she has helped me to learn how to think, which is a skill I am still trying to
master. Her advice and friendship are worth more than gold to me and without her help I would
never have been able to do this.
I would also like to express my deepest gratitude to Mark Gay. He spent countless hours
teaching me exciting, new, and useful laboratory techniques, helped me track down mounds of
information, took the time to think through numerous problems with me and was always there to
talk about my project with eagerness and intelligence. Without his assistance, I never would
have found a method to complete this work, nor would I have had the patience to sustain myself
through frustrating days in the lab when 'science just would not work'.
I want to acknowledge and thank Dr. Ann Pabst for all of her time and attention. She
always treated me as one of her own, and was always available to sit down and think about what
all of my data actually meant. She is an expert at asking the right questions and her input has
been invaluable in the process of completing this project.
Many more thanks go to Dr. Richard Dillaman for helping me to learn about histological
processes beyond the scope of my work, and in doing so, helping me to understand my own
work that much better. Without his time and help, this work would have suffered severely.
I would also like to acknowledge Dr. Andrew Westgate, who should have been a

committee member on this project; he has devoted the time and intellectual input above and
beyond what would be expected of a committee member. I am grateful for all of the time he has
devoted to this project, his patience, and his thoughtful comments on my work. I'd also like to
thank both him and Heather for genuinely caring about me and spending countless hours
encouraging me, without which I would still be floundering.
I would like to acknowledge the other students in my lab, both past and present, as well
as everyone who is a member of the VABLAB. It is rare to find one lab that cares so much
about each other and gets along so well, but to find two and have both accept me is something I
have been blessed with. I'd like to thank Zach Swaim, Hillary Lane, Caitlin McKinstry, Sandy
Camilleri, Bill McLellan, Dr. Sentiel "Butch" Rommel, Brian Balmer, Marina Piscitelli, Laura
Bagge, Caitlin Kielhorn, Ryan McAlarney, and Peter Nielsen for always providing a wonderful
audience for presentations and for being an important sounding board for ideas. I'd also like to
thank them for the friendship they've provided, which has enriched my life and kept me sane
while working on my project.
For sample collection, I'd like to thank the UNCW Marine Mammal Stranding Network,
the Cape Cod stranding Network, the Marine Mammal Center, and Ron and Shannon at Wells
Pork Products in Burgaw, NC. For funding I'd like to thank ONR for funding this project and
Prescott Grants for funding tissue collection. For additional help with methods and advice for
working with blubber, I'd like to thank D.J. Struntz and Eric Montie.
Lastly I would like to point out that the most important acknowledgement now and
forever, as well as all thanks, should go directly to God. He blessed me with a good mind and
has surrounded me by even better minds. I fail daily in my attempts to be a Christian, but He
continues to shower me with blessings, the most recent of which is this thesis.

DEDICATION
This thesis is dedicated to my family. My parents, Thomas William Rager and Carolyn
Jeanine Rager, have always stood beside me and given me their utmost support in every
adventure I have chosen to undertake no matter the emotional or financial cost to them. My
brother, Joshua Clarence Rager, has filled my life with friendship, laughter, and inspiration.
From the very first sentence my brother taught me to write, my family has worked to instill in me
a love of education and a belief that I could accomplish anything if I worked hard enough.
Finally, this thesis is also dedicated to my husband, Jeffrey Bronson McClelland. He is the love
of my life. He has always been there to hold on to me during my darkest moments and to pick
me up and dust me off afterwards. His support and his help in all things have enabled me to
write this thesis and to be where and who I am today. I will forever be grateful that we get to
share our lives together and I look forward to each new day we get to spend together.

LIST OF TABLES
Table Page
1. Specimen information for individual animals used in this study.......................................40
2. Microvascular characteristics of blubber/adipose tissue....................................................41
3. Biochemical characteristics of blubber/adipose tissue......................................................42
4. Average stratification Index for fatty acids in the blubber/adipose tissue.........................43
5. Select fatty acids and fatty alcohols found in blubber/adipose tissue................................44
6. Mean percent area consisting of microvascularity in a diversity of adipose
tissues of terrestrial mammals............................................................................................46

LIST OF FIGURES
Figure Page
1. Sampling site of blubber/adipose tissue.............................................................................47
2. Determining total number of terminal branches/vessel.....................................................48
3. Images of microvasculature in blubber/adipose tissue......................................................49
4. Percent area of microvessels analyzed by depth (Tursiops truncatus)..............................52
5. Percent area of microvessels analyzed by depth (Kogia breviceps)..................................53
6. Percent area of microvessels analyzed by depth (Sus scrofa)............................................54
7. Comparison of Tursiops truncatus, Kogia breviceps, beaked whales,
and Sus scrofa percent area of microvessels analyzed by depth........................................55
8. Microvasculature diameter of microvessels in blubber/adipose tissue..............................56
9. Microvascular branching of microvessels in blubber/adipose tissue.................................57
10. Percent lipid (wet weight) of blubber/adipose tissue.........................................................58
11. Percent of wax ester in blubber/adipose tissue..................................................................59
12. Fatty Acid Stratification Index...........................................................................................60

INTRODUCTION
Adipose tissue is a specialized form of loose connective tissue that is composed mainly of
adipocytes (Ross and Pawlina, 2006). Adipocytes are specialized lipid storage cells. The center
of these cells is filled with lipids; the cell’s nucleus and cytoplasm are pushed to one side
forming a signet ring shape (Ross and Pawlina, 2006). Small aggregations of adipocytes as well
as loose lipid droplets can commonly be found in numerous types of tissue throughout the body,
but the majority of fat in the vertebrate body is located in adipose tissue depots (Pond, 1998).
Adipose tissue can experience large changes in size, with most of the expansion or shrinkage of
adipose depots being due to changes in adipocyte dimensions (Young, 1976; Miller et al., 1983;
Groscolas, 1990; Ramsay et al., 1992; Koopman et al., 2002). Rather than new cell formation or
loss of existing cells (apoptosis), adipocytes may undergo tenfold changes in volume depending
on the amount of resources available and the energetic demands, such as migration or lactation,
of the animal (Young, 1976; Miller et al., 1983; Ryg et al., 1988; Groscolas, 1990; Pond, 1998;
Koopman et al., 2002). With increased food intake, lipid can be stored in adipose tissue to be
used during times of inadequate energy intake.
Marine mammals (cetaceans, pinnipeds, and sirenians) possess a highly modified type of
adipose tissue in the form of blubber. Blubber, a specialized hypodermal layer, forms the bulk of
lipid storage in these animals and is distributed almost solely subcutaneously around the body
(Bryden, 1968; Pond, 1978). Blubber is more rigid and organized than the adipose tissue of
terrestrial mammals (Parry, 1949; Ling, 1974; Ackman et al., 1975; Lockyer et al., 1984; Pond,
1998; Pabst et al., 1999; Ross and Pawlina, 2006). The structural difference between blubber
and adipose tissue of terrestrial mammals is due to the networks of collagen and elastin
surrounding the adipocytes (Parry, 1949; Ling, 1974; Ackman et al., 1975; Lockyer et al., 1984;
Pond, 1998; Pabst et al., 1999; Hamilton et al., 2004; Struntz et al., 2004; Ross and Pawlina,

2006; Montie et al., 2008b). In addition, the bodies of marine mammals contain more fat than
terrestrial mammals. Healthy terrestrial mammals in the wild average between 4-8% dissectible
adipose tissue (Pond, 1998), and most healthy marine mammals average around 30% dissectible
adipose tissue (Pabst et al., 1999; McLellan et al., 2002). It is not unusual to see even higher
values: harbour porpoise calves 37% fat; right whales 43% (Lockyer, 1991; McClellan et al.,
2002).
The increased fat stores in marine mammals serve multiple functions that enable marine
mammals to live in water. Marine mammals face higher thermoregulatory demands than those
living on land. Water conducts heat away from the body 25 times faster than air at the same
temperature (Parry, 1949; Scholander et al., 1950; Schmidt-Nielsen, 1997). Blubber provides
insulation that reduces heat loss, making it possible for marine mammals to expend less energy
to maintain their body temperature (Worthy and Edwards, 1990; Dunkin et al., 2005).
Movement in water is also associated with increased drag forces on the body, and blubber plays
a role in decreasing the amount of those forces experienced by marine mammals. Water is three
times denser than air and sixty times more viscous, leading to increased drag on the body (Webb,
1984; Fish, 1993; Vogel, 2003). Blubber sculpts the body, creating a more streamlined shape
and thereby reducing drag and conserving energy (Ryg et al., 1988; Pabst, 1990; Koopman et al.,
2002; Pabst et al., 1999; Hamilton et al., 2004). Blubber is also often less dense than water,
aiding the body by decreasing the overall density of the body and therefore increasing its
buoyancy (Taylor, 1994; Kipps et al., 2002).
There are two different types of blubber: metabolic and structural (Ackman et al., 1965;
Ackman et al., 1971; Koopman et al., 2002; Struntz et al., 2004; Montie et al., 2008b). Metabolic
blubber is used for energy storage (Young, 1976; Lockyer, 1986; Ryg et al., 1988; Aguilar and

Borrell, 1990; Pond, 1998; Koopman et al., 2002; Montie et al., 2008b). In certain locations of
the body, such as the thoracic region, blubber is stratified in terms of its structure and its
biochemical composition. Histological analysis of adipocytes and structural fibers suggests that
the inner layers of blubber from the thoracic region of a number of cetacean species are
metabolically active, but that the outer layer of blubber from this region is structural (Ackman et
al., 1965; Ackman et al., 1971; Koopman et al., 2002; Struntz et al., 2004; Montie et al., 2008b).
In the metabolic blubber layer of odontocetes in good body condition, the adipocytes are larger
than those of the metabolically inert stores (Koopman et al., 2002; Struntz et al., 2004; Montie et
al., 2008b). During starvation, the metabolically active adipose tissue shrinks significantly, but
the metabolically inert (“structural”) blubber shows no change from that of a healthy cetacean
(Koopman et al., 2002; Struntz et al., 2004).
Blubber is also stratified biochemically. Lipid content varies throughout the depth of
blubber, with the highest lipid content generally found in the innermost and middle layers
(Ackman et al., 1975; West et al., 1979; Kakela et al., 1993; Kakela and Hyvarinen, 1996;
Koopman, 2007). Data from bottlenose dolphins suggests that, during lipid mobilization, lipids
are only mobilized from the middle and deep layers, with fat from the deep layer being
mobilized first (Struntz et al., 2004; Montie et al., 2008b). The blubber that is metabolically
“active” contains more fatty acids that are from a dietary origin, while the possibly inert
structural layer of blubber has a higher degree of fatty acids from an endogenous origin; such a
pattern of fatty acid distribution has been observed across a wide range of marine mammal
species including seals, walruses and other cetaceans (Ackman et al., 1965; Ackman et al., 1971;
Ackman et al., 1975; West et al., 1979; Lockyer et al., 1984; Aguilar and Borrel, 1990; Lockyer
et al., 1991; Kakela et al., 1993; Kakela and Hyvarinen, 1996; Koopman et al., 1996; Koopman

et al., 2002; Cooper, 2004; Iverson et al., 2004; Koopman, 2007).
All of the above properties and functions of blubber point to its being an important and
dynamic living tissue. Typically, adipose tissue is perfused by networks of capillaries,
microarterioles and microvenules (Gersh and Still, 1945; Herd et al., 1968; Hausman and
Wright, 1996; Crandall et al., 1997) so that the vascular system can transport molecules to and
away from the tissue with both nutrient and gas exchange occurring at capillaries (Campbell et
al., 1987). In terrestrial animals the adipose tissue is highly vascularized (Gersh and Still, 1945;
Herd et al., 1968; Hausman and Wright, 1996; Crandall et al., 1997) and can experience high
rates of blood flow (Rossell et al., 1974). Surprisingly there is little information on the
vascularization of blubber. One of the few studies that tried to analyze capillaries in blubber
(Parry 1949) was unable to do so and instead only briefly described large vessels travelling
through the blubber to the epidermis; this description is based upon a single harbor porpoise.
The virtual absence of data on vascular density, morphology and anatomy in blubber is
remarkable given the importance of this tissue and the fact that other aspects of blubber have
been very well studied (e.g. Bryden, 1968; Pond, 1978; Ryg et al., 1988; Worthy and Edwards,
1990; Kipps et al., 2002; Koopman et al., 2002; Montie et al., 2008). Many factors may influence
the microvasculature of blubber, such as thermoregulation, habitat, metabolic demands,
reproductive strategies, or phylogeny. Another unique aspect of marine mammal life to consider
is diving and the mammalian dive response. During a dive, marine mammals undergo
bradycardia, apnea, and ischemia, in which vasoconstriction is believed to control blood flow to
the peripheral tissues (Irving et al., 1941). If the blubber experiences peripheral
vasoconstriction, the blood supply to the blubber could be significantly reduced, if not stopped.
However, our ability to evaluate blubber’s behavior during diving, or the metabolic activity of

blubber, is hampered by a lack of foundational information on the relationship between this
tissue and the circulatory system.
Therefore the first step in considering the relationship between blubber and blood is to
characterize the vascular patterns in blubber. The overall goal of this study was to provide the
first detailed descriptions of the microvasculature (capillaries, microarterioles and microvenules)
in blubber, using the bottlenose dolphin (Tursiops truncatus) as a model. Tursiops is often used
as a model cetacean as much is known about the species (e.g. Scholander and Schevill, 1955;
Ridgeway et al., 1969; Noren et al, 1999; Meagher et al., 2002; Rommel et al., 2002; Wells and
Scott, 2002; Houser et al., 2004; Gannon et al., 2005; Harper et al., 2008). The first objective of
this study was to determine microvascular characteristics (density, vessel size, branching
patterns) using histological analysis. Vascular characteristics were then compared with other
lipid content, lipid class and fatty acid composition to determine whether the distribution of
blood vessels was linked to the biochemical features of blubber. Since the blubber of Tursiops
has been determined to be stratified by histological examination of adipocytes and structural
fibers (Struntz et al., 2004; Montie et al., 2008b) as well as biochemically (Samuel and Worthy,
2004; Koopman, 2006), I measured the microvasculature density in three layers of blubber: the
superficial layer (nearest to the epidermis), the middle layer, and the deep layer (nearest the
muscle), with the working hypothesis that there would also be stratification of the
microvasculature.
Although there are many potential factors controlling or affecting patterns of blood vessel
distribution in blubber, one influence that unites all marine mammals is the act of diving. The
second objective of this study was to compare the blubber microvasculature of Tursiops, a short-
duration (approximately 20-40 seconds), shallow diving (approximately 1-10 meters) cetacean

(Wursig, 1978; Irvine et al., 1981; Shane, 1990; Bassos, 1993), with that of the blubber of the
pygmy sperm whale, Kogia breviceps. Kogia makes dives to approximately 400 meters for
likely around 18 minutes which is a moderate dive duration (Scott et al., 2001; Beatson, 2007)
allowing a possible view of how dive depth may affect microvasculature density. The third
objective of this study was to put the cetaceans into a larger mammalian context by comparing
microvasculature of blubber with that of the subcutaneous adipose depots of a terrestrial
mammal, the domestic pig (Sus scrofa).
I hypothesized that the deeper divers would have reduced microvascular densities in their
blubber, given that these tissues would likely be experiencing chronic reduced perfusion during a
dive. Furthermore, at depth, diving mammals experience high pressure and consequently
elevated nitrogen partial pressures in their tissues (Serway and Faughn, 1992). Nitrogen is a
lipophilic molecule that is five times more soluble in lipid than blood and as such will move into
fat (Ikels, 1964; Gerth, 1985). Therefore, I reasoned that having a reduced amount of
microvasculature could be a means to allow deep divers to limit the amount of nitrogen that
could potentially enter the blubber. However, it may be possible that adipose tissue vascularity
is conserved across species and, with our choice of three diverse species we may elucidate this
relationship.
Data from this project will provide new and much needed data concerning the anatomy of
the microvasculature in blubber. It will also enhance our understanding of diving physiology
and provide insight into the specialized adaptations of being a marine mammal.
MATERIALS AND METHODS
Tissue Collection
Measurements were made on the thoracic blubber of six Tursiops truncatus, six Kogia

breviceps, two beaked whales (Blainville’s beaked whale, Mesoplodon densirostris and Cuvier’s
beaked whale, Ziphius cavirostris), and the subcutaneous layer of back fat from four pigs, Sus
Scrofa (see Table 1 and Figure 1). All odontocetes sampled had either stranded or had been
killed incidentally in commercial fishing operations, and only blubber obtained from animals
with a Smithsonian stranding code of 1 (live stranded and died or euthanized) or 2 (fresh dead)
were used in this study (Geraci and Lounsbury, 1993). All samples were from adult specimens
that were classified as having a normal body condition on the basis of an examination during
necropsy (Cox et al., 1998). All pig samples were obtained from a porcine butcher (Wells Pork
Products, Burgaw, NC) from animals that had been sacrificed for human consumption. After
collection all samples were wrapped in plastic and frozen at -20°C prior to further analysis.
Freezing effects on blubber are negligible (Pond and Mattacks, 1985). However, to
eliminate any possible freezing artifacts in blubber or pig fat samples, pieces selected for either
histological or lipid analysis were taken from the centre of the sample (i.e. not the edges touching
the sample storage bag) of the tissue. The epidermis was removed and the blubber section was
then cut into equal thirds to allow analysis of the samples based on blubber depth: superficial
(nearest the epidermis), middle, and deep (nearest the subdermal connective tissue sheath and
muscle) (Samuel and Worthy, 2004; Struntz et al., 2004; Montie et al., 2008b). Four samples
were removed from each layer, two for histological analysis of vessels: one in the longitudinal
plane and one in the transverse plane, one for histological analysis of adipocytes, and one for
lipid biochemistry.
Histological Analysis of Vessels
As this was the first study to quantify the microvascular densities of blubber, I first had to

develop the methodology. While numerous techniques have worked to distinguish vessels from
surrounding tissue, not all mammalian tissues nor those from different species respond the same
to various staining and histological techniques requiring us to attempt numerous techniques to
find a suitable method for detecting blood vessels in blubber. I attempted to visualize the vessels
in blubber using various lectins to stain for specific carbohydrates found on the cell surfaces of
endothelial cells (Alroy et al., 1986). The lectins that were used to try to identify the vessels in
blubber were GSL-I, RCA, and WGA, all of which have been shown to stain vessels in cats,
cows, dogs, goats, horses, mice, pigs, rats, sheep and humans (Alroy et al., 1986; Hausman and
Wright, 1996 ). Discerning the vessels in blubber was not possible with the lectins because the
lectins bound to adipocyte cell membrane components in addition to the vasculature. When
Tursiops samples of muscle and spleen were used, staining occured the same as it had for
blubber, making visulization of the vasculature difficult. Each lectin worked as previously
described on mouse tissues; however this method did not work with any dolphin tissues.
Antibodies known to bind with endothelial cells were also used to try to visualize the vasculaure
of blubber; trials were conducted with CD-31, MECA-32, and vWF on Tursiops blubber and
kidney samples; these are antibodies that bind endothelial cells in humans (CD-31, MECA-32,
vWF), mice (CD-31, MECA-32, vWF), rats (CD-31, vWF), guinea pigs (CD-31, vWF), rabbit
(CD-31, vWF), monkeys (CD-31), pigs (CD-31), and chickens (CD-31) (Developmental Studies
Hybridoma Bank, Iowa City, Iowa; Vector Laboratories, Inc., Burlingame, CA; Abcam, Inc.,
Cambridge, MA: Hallmann et al., 1985; Wong et al., 2000; Zanetta et al., 2000; Norrby and
Ridell, 2003; Young and Black, 2004). The antibodies all demonstrated non-specific binding in
blubber and when secondary binding was blocked with BSA, no binding occurred. The same
results were obtained with Tursiops kidney tissue with the exception of MECA-32, which did

have light staining of Tursiops kidney tissue and may be a possibility for viewing endothelial
cells in this tissue. When all of these techniques failed to reliably stain for blood vessels, I
turned to standard histochemical techniques. Techniques such as H&E, H&E with alcian blue,
PAF, and PAS were all tried (Groat, 1949; Parry, 1949; Gomori, 1950; Hausman and Thomas,
1983; Hausman and Richardson, 1983; Presnell and Schreibman, 1997), and none provided
sufficient contrast with the surrounding tissue to enable accurate blood vessel
counts/measurements. Finally, samples were incubated with nitro blue tetrazolium chloride plus
5-bromo-4-chloro-3-indolyl phosphate (NBT/BCIP). NBT/BCIP stains for alkaline phosphatase,
which is located in the microarterioles, microvenules, and capillaries (Foley et al., 1954;
McComb and Bowers, 1979; Hansen-Smith et al., 1992) and has been used to view blood vessels
in people, rats, rabbits, pigs and other mammals (Foley et al., 1954; Wachstein and Meisel, 1959;
Cros et al., 1980; Hausman and Richardson, 1983; Werner et al., 1987). NBT/BCIP proved to be
a viable option for fresh, frozen tissue samples. According to Gomori, samples fixed with
alcohol or acetone and paraffin embedded at 56-60°C still have alkaline phosphatase activity
(Gomori, 1939; Presnell and Schreibman, 1997). However, when attempted with acetone fixed,
paraffin embedded tissues ,no staining was visible requiring us to use fresh, frozen tissue
sections.
Frozen, unfixed tissue samples were placed in a Leica Cryocut 1800 (Leica Microsystems
Inc., Bannockburn, IL), covered with Optimal Cutting Temperature Compound (OCT) (Ted
Pella, Inc., Redding, CA), and allowed to freeze to approximately -27°C (temperature
determined through preliminary trials with blubber samples). The samples were sectioned at
30μm thick and placed on a Superfrost Gold Plus slide (Fisher Scientific, Pittsburgh, PA). The
slides were then stored in the freezer at -20°C in a slide box enclosed in a freezer bag until

staining could be completed (not longer than 12 hours).
Samples were rinsed in Sorensen’s phosphate-buffered saline (PBS) for 15 minutes
(Presnell and Schreibman, 1997), and then incubated in a nitro blue tetrazolium chloride plus 5-
bromo-4-chloro-3-indolyl phosphate (NBT/BCIP) solution made out of NBT/BCIP ready-to-use
tablets dissolved in 10ml of distilled water (Roche Diagnostics, Mannheim, Germany). Different
parameters, such as temperature, pH, and time can all affect the binding of NBT/BCIP to the
alkaline phosphatase enzymes found in the microvasculature (McComb et al., 1979). Roche
Diagnostics advises to incubate samples at room temperature and a pH of 9.5 (Roche
Diagnostics, Mannheim, Germany). Preliminary incubations of 20-30 minutes, which should
provide good staining intensity (Vector Laboratories, Inc., Burlingame, CA), did not provide
optimal staining for all species. This was obvious due to the faint coloration of the samples for
all of the species in the study except Tursiops. According to Vector Laboratories, Inc.,
lengthening incubation times provides significantly more sensitivity of NBT/BCIP (Vector
Laboratories, Inc., Burlingame, CA). Incubation times were therefore varied and optimal
incubation times were determined by a series of trials of increasing incubation times. Specific
times for each species were chosen based on qualitative analysis of stained sections. Incubation
times varied: 20 - 25 minutes for Tursiops, approximately two hours for Kogia and 36 hours for
beaked whales and pigs. This could be due to a number of reasons. There are different families
of alkaline phosphatases and these may vary among the test species. (McComb et al., 1979). The
species in the study may possibly have different concentrations of those enzymes in their
microvasculature (McComb et al., 1979). Furthermore, NBT/BCIP is "little soluble in water and
lipid" (Vector Laboratories, Inc., Burlingame, CA), and the incubation times needed for the
species analyzed in this study correlated with the percent lipid of the tissue samples (the tissue

with the higher percent lipid had the longest incubation times; see Table 3; Figure 10). The
samples were rinsed in PBS for seven minutes and placed under coverslips with trisglycerol
(Presnell and Schreibman, 1997).
Stained sections were viewed with an Olympus BX60 (Olympus America Inc., Center
Valley, PA) and digital pictures were taken of each section using a Diagnostic Instruments SPOT
RT digital camera (Diagnostics Instruments, Sterling Heights, MI). All images were analyzed
using Image ProPlus software (Media Cybernetics, Inc., Bethesda, MD). All vessel image
analyses were carried out on sections cut from two orthogonal body planes (longitudinal and
transverse) and then averaged together to eliminate any possible orientation bias between the two
planes.
To calculate percent vascularity and describe vascular branching, one image was captured
from five different, noncontiguous tissue sections for each layer of each sample using a 4x
objective. Percent vascularity of each layer was determined by counting each vessel that touched
the intersecting point of the horizontal and vertical lines of a proportional, orthogonal grid
consisting of eleven horizontal and 15 vertical grid lines, which was overlaid on each image in
Image ProPlus (Howard and Reed, 1998). This grid size allowed for a minimum count of 100
vessels when the counts from each of the 5 tissue sections were pooled.
Vascular branching was determined by counting the total number of terminal end points
for the first five vessels that touched a grid intersection for each of the five noncontiguous tissue
section images (see Figure 2). This resulted in the number of branches per vessel being counted
for 25 vessels, which were averaged to determine the number of branches per vessel for each
layer.
Images were captured with a 10x objective for all vessel diameter measurements and

diameters were measured for each layer by using the linear measurement tool in Image ProPlus.
Vessels were found under a microscope and then followed (in the x, y, and z planes) until the
vessel was cut. If the terminal point of the vessel had been cut in a cross section then the vessel
was used to determine vessel diameter. The first ten vessels that were found cut in cross section
were measured for each layer and the average of those measurements was used to determine
vessel diameter for that layer.
Lipid Analysis
Lipid was extracted from blubber subsamples weighing approximately 0.5 g using a
modified Folch method (Folch et al., 1957; Koopman et al., 1996) to yield percent lipid (wet
weight). Lipid classes [triacylglycerols (TAG), wax esters (WE), cholesterol and phospholipids]
were identified and quantified using a Mark VI Iatroscan TLC/FID (Mitsubishi Kagaku Iatron,
Inc, Tokyo, Japan). Samples were developed in a solvent system of 94/6/1 hexane/ethyl
acetate/formic acid. Lipid class peaks were integrated and quantified using Peak Simple 329
Iatroscan software (SRI Instruments, Torrance, CA). Identification of peaks was confirmed using
known standards (NuChek Prep, Elysian, MN) of WE, TAG, free alcohols, phospholipids,
cholesterol and diacylglycerols (see Koopman, 2007). Fatty acids and alcohols of lipid class
components were analyzed using temperature-programmed capillary gas-liquid chromatography
(GC) of butyl esters on a Varian 3800 GC (Varian, Inc., Palo Alto, CA). Components were
separated and analyzed with a flame ionization detector (ID) in a fused silica column (30 x
0.25mm ID) (Zebron FFAP; Phenomenex). Helium was used as the carrier gas and the gas line
was equipped with an oxygen/water scrubber. The following temperature program was used:
65°C for 2 min, hold at 165°C for 0.40 min after ramping at 20°C/min, hold at 215°C for 6.6 min

after ramping at 2°C/min, and hold at 250°C for 5 min after ramping at 5°C/min. Identities of
individual components were confirmed using prepared standards (NuCheck Prep, Elysian, MN)
and GCMS (Trace GC Ultra coupled to a Polar Q mass spectrometer, Thermo Electron
Corporation with X calibur software) (Suzanne Budge, Dalhousie University). Fatty acids and
fatty alcohols were described using the nomenclature of A:Bn-X, where A is the number of
carbon atoms, B is the number of double bonds, and X is the carbon position of the double bond
closest to the terminal methyl group. These results were integrated with Galaxie (version Varian
1.8.501.1) GC software.
Fatty Acid Stratification
To determine fatty acid (FA) stratification in the blubber a stratification index (SI) was
computed (Koopman, 2007). To calculate the SI, the absolute values of the differences between
deep and superficial layers in a subset of 16 select FA were added together. The 16 FA used
were iso5:0, 14:0; 14:1n-5; 16:0; 16:1n-7; 18:0; 18:1n-11; 18:n-9; 18:1n-7; 18:2n-6; 20:1n-11;
20:1n-9; 20:5n-3; 22:1n-11; 22:5n-3; and 22:6n-3 (Koopman, 2007). These FA were chosen to
compare the SI of this study to those calculated by Koopman (2007).
Statistics
To determine whether there were species differences, or differences associated with depth
of blubber layer in the variables measured, univariate analysis of variance (general linear model)
were conducted on all variables. Repeated measures tests were used to account for the fact that
each individual animal was sampled multiple times (three layers) (SPSS, 1997; Tabachnick and
Fidele, 2007). The within subject factors examined were percent vascularity, number of vascular

branches, vessel diameter, percent lipid, lipid class composition, and stratification index. If there
was significant interaction between depth and species effects, then repeated measures tests were
run separately on each layer to determine a species effect, and on each species to determine a
layer effect.
For each ANOVA, the data were tested for sphericity using Mauchly's test of sphericity.
If sphericity could not be assumed then the Greenhouse-Geisser correction factor, a conservative
value for low n-numbers, was applied to the F-statistic when testing for significance (SPSS,
1997; Tabachnick and Fidele, 2007). For each test, groups were evaluated for equality of
variance using Levene’s test. When within-subject effects were significant, post-hoc tests were
carried out to determine where differences existed using either a Tukey post-hoc (equal
variances) test or a Tamhane T-2 test (unequal variances).
All stastistical tests were carried out using SPSS (SPSS Inc., Chicago, IL). All means are
presented with SEM.
RESULTS
Histological Analysis of Vessels
Blubber microvasculature in Tursiops truncatus was stratified throughout the depth of the
blubber (p<0.001). On average, the superficial layer contained approximately 7% fewer
microvessels than the middle and deep layers (p<0.001), which were not significantly different
from each other (Figure 4; Table 2; p=0.073). This pattern was not found in either Kogia
breviceps or in Sus scrofa (pigs). Kogia showed an insignificant (p=0.160) trend of increasing
microvaculature density from superficial to deep blubber (Figure 5; Table 2). There was no
difference (p=0.898) in the density of the microvasculature of pig subcutaneous back fat based
on depth (Figure 6; Table 2). Two beaked whales that had been analyzed (M. densirostris, n=1;
Z. cavirostris, n=1) had similar microvasculature densities to each other and to the pigs, with no

obvious difference observed throughout the depth in either species (note that statistics were not
performed on beaked whales due to only single individuals being examined) (Table 2).
When comparing across species, Tursiops had greater microvasculature densities in their
middle and deep blubber layers compared to those layers in Kogia (p=0.005), pigs (p<0.001), or
the two individual beaked whales analyzed (Figure 7). Kogia had greater blubber
microvasculature density than either the pigs (p=0.017) or the beaked whales (Figure 7). The
blubber microvasculature densities of the two beaked whales mirrored the microvasculature
density found in pig fat (Figure 5).
There were no significant species (p=0.157) or depth (p=0.176) related differences in the
diameter of the microvessels (Table 2; Figure 8). The average range of vessel diameter size was
generally between 9 – 11 micrometers.
In Tursiops, there were significantly more branches per microvessel in the deep (5.55
branches/microvessel) and middle (5.76 branches/microvessel) layers than in the superficial
(2.75 branches/microvessel) layer (Figure 9; Table 2; p=0.006); this was the only species to
exhibit stratification in branching and the only species to differ from the others in these layers
(p=0.015). All layers of the remaining species contained an average of 2.92 branches/vessel
(Figure 9; Table 2) with no significant differences among Kogia or the pigs (p=0.761), and the
beaked whales followed a similar trend.
Lipid Analysis
In Tursiops, lipid content was stratified throughout the depth of the blubber (p=0.039).
The middle layer, consisting on average of 59.9% wet weight lipid, had more lipid than did the
superficial layer, 39.4% (p=0.001), and the deep layer, 41.9% (Figure 10; Table 3; p=0.001).

Kogia blubber was also stratified in the amount of lipid per layer, with the superficial layer
(consisting of 33.7% lipid), having fewer lipids than the middle (75.1%; p=0.010) and deep
layers (69.2%; Figure 10; Table 3; p=0.030). There was no difference (p=0.683) in the amount
of lipid in pig fat based on depth (Figure 10; Table 3). There was also no obvious difference in
the amount of lipid throughout the depth of blubber in either of the beaked whales (Figure 10;
Table 3).
Tursiops blubber and pig fat contained only triacylglycerols (Table 3). The blubber of
beaked whales contained around 98% wax esters and less than 1% triacylglycerols (Figure 11;
Table 3). Kogia blubber was, on average, 75% wax esters and 22% triacylglycerols (Figure 11;
Table 3). There was no stratification of lipid type in any species (Figure 11; Table 3; all p>0.05).
Overall the FA Stratification Index (SI) showed considerable stratification in fatty acids
in Tursiops (p<0.0001), Kogia (p=0.001), and the beaked whales (see Table 4; Figures 12); there
was no difference in FA stratification between Tursiops and Kogia (p=0.147). There was less
stratification of fatty acids in pig adipose tissue than for Tursiops or Kogia (p= 0.009; see Table
4; figure 12). In all odontocetes examined, dietary fatty acids occurred in the highest
concentrations in the deep layers of the blubber, and decreased towards the superficial layers.
Fatty acids with an endogenous origin decreased from the superficial to the deep layers of
blubber.
DISCUSSION
This study provides the first quantitative description of microvessels in the blubber of any
marine mammal. Given the importance of this tissue for these animals, this is surprising.
Studies on the adipose tissue in rats, dogs, fetal pigs and humans have all shown that each
adipocyte is in contact with at least one capillary (Gersh and Still, 1945; Herd et al., 1968;

DiGiralamo et al., 1971; Rossell and Belfrage, 1979; Hausman and Richardson, 1983; Hausman
and Thomas, 1985; Anstrom et al., 2004; Hausman and Richardson, 2004). However, because
blubber is a highly specialized form of adipose tissue it cannot be assumed that blubber
vasculature is the same as that found in typical mammalian adipose tissue.
Vascular Distribution and Morphology
The microvasculature densities of pig adipose tissue found in this study were nearly
identical to those of other terrestrial mammals, mice and humans (Table 6; Lijnen et al., 2006;
Hemmeryckx et al., 2008; Lijnen et al., 2009; Pasarica et al., 2009). It therefore seems likely
that the microvasculature density of adipose tissue is conserved in terrestrial mammals.
Furthermore, this same relatively low density was found in the superficial layer across three
families of odontocetes (Delphinidae, Kogiidae, Ziphidae) suggesting some level of conservation
in the vascularity of fat that is located closest to the epidermis. The same vascular characteristics
(density, branching) observed in the superficial layer of fat were maintained throughout the depth
the blubber of both beaked whales, making this tissue fairly uniform in these species. However,
some deviations from this pattern were observed in Tursiops, and to a lesser degree in Kogia.
Tursiops showed the greatest divergence from the other species. The middle and deep
layers of blubber in Tursiops had significantly higher microvascular densities, and the
microvessels in these layers also had significantly more microvascular branching, than did all of
the other species. Thus, it is the middle and deep layers of Tursiops blubber that showed the
greatest departure from the other mammals in this study and in the literature.
In Kogia there was a slight but statistically insignificant increase in vascular density
towards the inner layers of blubber. One of the Kogia analyzed (Kb03) exhibited a dramatically

different pattern of vessel distribution than the other conspecifics (Figure 5). Excluding this
individual from the statistical analysis made the trend of greater vascular density in the inner and
deeper layers, compared to the superficial layer, significant (p=0.031). Thus, it is likely that
Kogia blubber does exhibit an increase in vascular density in the innermost blubber layers,
although not as pronounced as that observed in Tursiops.
Parry (1949) was unable to identify any capillaries in the blubber of a harbor porpoise
(Phocoena phocoena). Other studies that have examined the portion of the microvascular system
that lies close to, but not in, the blubber of marine mammals have concentrated on the vessels in
the appendages that are believed to contribute significantly to the animals’ thermoregulatory
abilities (Fawcett, 1942; Tomilin, 1957; Palmer and Weddell, 1964; Elsner, 1969; Elsner et al.,
1974; Bryden and Molyneux, 1978). However, the microvessels located within the blubber itself
have been largely overlooked.
The diameter of the microvessels was the same for all species in this study. The average
diameter was between 9 and 12 micrometers with no clear patterns throughout the depth in any
species. This result is not surprising, because the size of capillaries is likely constrained based
on the size of a red blood cell, which in humans falls between 6-9 m (Persons, 1929). This
result confirms that capillary size is conserved across species, at least in mammals (Hausman and
Richardson, 1983; Anstrom et al., 2004; Ross and Pawlina, 2006).
Relationship of Vascular Patterns to Biochemical Characteristics of Blubber
I hypothesized that I would see correlations between the blubber microvasculature and
blubber biochemical characteristics because the vascular system transports molecules, such as
fatty acids (Campbell et al., 1987), to and away from the blubber. Much more is known about

the biochemical characteristics of blubber, not only for the species in this study but for other
marine mammals in general, than on the vascular properties of blubber. In general, the
biochemical data obtained here agree with previous studies on the physiological aspects of
blubber of odontocetes (Struntz et al., 2004; Samuel and Worthy, 2004; Montie et al., 2008b;
Koopman, 2007).
In this study, the blubber of Tursiops was biochemically stratified among the superficial,
middle and deep layers in terms of total lipid content (see also Samuel and Worthy, 2004;
Montie et al., 2008b; Koopman, 2007), as well as in microvascularity. While the middle and
deep layers of blubber had higher microvascular densities than the superficial, the percent lipid
did not match this pattern. The middle had significantly more lipid than the superficial layer and
deep layers of blubber. Other studies have shown that the superficial layer of blubber in
Tursiops is structural, while the middle and deep layers are metabolic (Struntz et al., 2004;
Montie et al., 2008b). These studies have also suggested that the middle layer forms a more
stable lipid store, and that it is the adipocytes in the deep layer that are accessed first with
adipocytes in that layer showing more variety in cell size/volume (i.e. lots of small and large
cells) than in the middle layer (mostly large cells) and the superficial layer (mostly small cells)
(Struntz et al., 2004; Montie et al., 2008b). Thus in Tursiops the microvascular densities are
highest in the blubber layers where lipid thought to be primarily mobilized, and lowest in the
superficial layer of blubber where little or no lipid mobilization is believed to occur.
In agreement with past work, this study also showed that Kogia breviceps blubber was
stratified in terms of lipid content, with the superficial layer having fewer lipids than the middle
and deep layers (Koopman, 2007). In Kogia the microvascularity exhibited a slight increase
through the depth of the blubber. As mentioned above, higher microvascular densities in

Tursiops were found in the layers of blubber that are thought to be more metabolically active.
While much less is known about the blubber of Kogia, the slightly higher vascular densities in
the middle and deep layers may be due to lipid being mobilized more frequently in these layers
than the in the superficial layer. However the microvascular density differences are much closer
among all three layers in Kogia than in Tursiops, possibly suggesting that there are less
differences among the layers in how lipid is mobilized.
The blubber of M. densirostris and Z. cavirostris has not been previously analyzed for
differences in lipid content between outer and inner blubber layers. However, Koopman (2007)
analyzed the blubber of two other species of beaked whales, M. bidens and M. europaeus, and
found no stratification in lipid content between those layers. This study showed fairly uniform
lipid content among all layers in the blubber of M. densirostris and Z. cavirostris. It also showed
a uniform amount of microvascularity throughout the depth of the blubber in these two species.
There was, overall, an inverse relationship between lipid content and microvascular
densities of each species. The species with the highest lipid content (the beaked whales and
pigs) had the lowest microvascular densities. The species with the lowest lipid content
(Tursiops) had the highest microvascular densities; Kogia was intermediate in both categories.
This relationship was not due to the vasculature limiting the space that could be occupied by
lipid, as the small difference (~7.5%) in vascular densities cannot account for the larger
differences seen in lipid content (~49%).
Previous studies have shown that in odontocetes, lipid class composition of blubber is
strongly linked to phylogeny (e.g. Litchfield et al, 1975; Koopman, 2007). In agreement with
previous studies, the blubber of Kogia and both beaked whales was dominated by wax esters
(WE). The blubber of Tursiops and the adipose of pigs only contained triacylglycerols (TAG).

A prevalence of WE seemed to be associated with lower vascular densities. The blubber of
beaked whales and Kogia (both WE-dominated) had low vascular densities, but pigs, which
contained only TAG, had vascular densities similar to beaked whales and, in fact, lower than
Kogia. This comparison suggests that the correlation between vascularity and WE is more likely
a coincidence or an indirect relationship and that lipid class likely does not influence
microvascularity.
The final biochemical parameters I measured were fatty acid composition and fatty acid
stratification. The stratification indices I observed were slightly higher than those previously
reported by Koopman (2007). However, this result was not surprising because Koopman (2007)
separated the blubber into only two layers (thus providing a less extreme comparison), while this
study looked at the more dramatic differences between the deep and superficial blubber layers.
The blubber of all of the cetaceans was stratified to varying degrees, but the pig adipose showed
no stratification. The pig adipose tissue also contained many fewer fatty acids than any of the
cetaceans, which is not all that surprising given that the dietary intake of these animals was
constrained as they were not wild or free-ranging, and the fact that marine systems tend to
contain a diverse array of a larger number of fatty acids (Budge et al. 2006). There was no
obvious correlation between fatty acid stratification and microvascularity (data not shown). This
result was very surprising because initially I had thought that microvascular stratification may be
one cause of the fatty acid stratification. While it might be logical to assume that some aspect of
the vasculature was linked to controlling fatty acid stratification, it may be that the two are
unrelated and that there is another mechanism, or mechanisms (e.g. differential enzyme activity
or expression), controlling the fatty stratification found in blubber. At the current time our lack
of knowledge concerning the biochemical mechanisms of fatty acid deposition in adipose tissue,

as well as the limited number of animals and species analyzed in this study, precludes our ability
to determine the exact nature, if any, of a relationship between vascularity and fatty acid
stratification.
Potential Influences on Blubber Vasculature Structure
We have a generally limited understanding of the biology, behavior and physiology of the
majority of odontocete species, particularly those with pelagic or deep-diving lifestyles. Without
adequate ancillary data it is difficult to make any strong inferences or conclusions about which
factors (environmental, biological, phylogenetic) might have the greatest influences on
microvascular characteristics of blubber. However, I do wish to explore some possible causes
and correlaries of the data, with the caveat that these are merely potential relationships that we
cannot fully evaluate at this time.
I had originally assumed that blubber’s microvascular patterns would be associated with
the diving characteristics (depth, length) of a given species. Given the constraints associated
with the dive response, I hypothesized that the deeper divers would have reduced microvascular
densities. At depth, diving mammals experience high pressure and consequently elevated
nitrogen partial pressures in their tissues (Serway and Faughn, 1992). Nitrogen is a lipophilic
molecule that is five times more soluble in lipid than blood and as such will move into fat (Ikels,
1964; Gerth, 1985). Therefore, I reasoned that having a reduced amount of microvasculature
could be a means to allow deep divers to limit the amount of nitrogen that could potentially enter
the blubber. Tursiops, the shallowest diver in the study, had the greatest amount of
microvasculature, and also the highest degree of microvascular stratification. Kogia, an
intermediate diver, had a lower amount of microvasculature. The deepest divers in the study, the

beaked whales, had the least amount of microvasculature. Even though only a limited number
of species have been analyzed to date, it would seem that microvascular densities seem to
correlate with dive depth, at least in odontocetes. However, after analyzing the microvascular
densities of adipose tissue in a terrestrial mammal, a pig, and comparing these with literature
values from other terrestrial mammals, this idea loses support (Table 6; Lijnen et al., 2006;
Hemmeryckx et al., 2008; Lijnen et al., 2009; Pasarica et al., 2009). The terrestrial mammals
had the same amount of microvasculature as the deepest divers (the beaked whales), suggesting
that lower microvascular densities may be the “ancestral” mammalian condition and remain
conserved in beaked whales. The "reduced vasculature" seen in the deep divers may be the more
conserved state, while the blubber microvasculaure of Tursiops seems to show the greatest
alteration from the typical mammalian pattern. These data may well indicate that rather than
thinking about Tursiops as a “model” ooontocete, we should really regard Tursiops as a species
showing significant departures from members of other families of odontocetes and other
mammals, at least in terms of its blubber vasculature. A logical extension is then to consider
possible reasons for the observed increased vasculature density and branching in Tursiops.
The coastal Tursiops utilized in this study, live in a coastal environment where they
experience many seasonal changes; water temperatures fall from around 30˚C in summer to
around 11˚C in winter and availability of prey also changes, making food supply less consistent
(Irvine et al. 1981; Wells et al., 1987; Thayer et al., 2003; Barros and Odell, 1990; Cockcroft and
Ross, 1990; Barros and Wells, 1998). Thus these animals must cope with variation in both
energy availability and thermoregulatory challenges imposed by seasonal fluctuations. This
higher degree of environmental variation is reflected in the seasonal mobilization of energy from
Tursiops truncatus blubber (Samuel and Worthy, 2004; Meagher et al., 2008). This study

showed that the microvascular density is amplified in the blubber of Tursiops, possibly
enhancing their ability to adapt to both the changing energy availability and temperature of their
coastal habitat. In summer, fish are abundant, Tursiops are able to consume more and increase
their blubber stores in preparation for winter (Barros and Odell, 1990; Cockcroft and Ross 1990;
Barros and Wells, 1998). In winter, a thick insulating layer is important in preventing heat loss,
the additional lipid stores can provide energy when food is unpredictable (Barros and Odell,
1990; Cockcroft and Ross 1990; Barros and Wells, 1998; Samuel and Worthy, 2004; Dunkin et
al., 2005; Meagher et al., 2008). By summer, lipid has been mobilized from the middle and deep
blubber layers decreasing the thickness of the blubber by 39% (Samuel and Worthy, 2004;
Meagher et al., 2008).
In contrast, Kogia live and forage in the deep waters of a pelagic environment (Caldwell
and Caldwell, 1989; Scott et al., 2001; Beatson, 2007). In this environment seasonal effects are
minimized, as the temperatures faced are more consistent at depth. In these more stable waters,
Kogia feed on pelagic cephalopods, which migrate vertically in the water column on a daily basis
and are available year round (Raun et al., 1970; Roper and Young, 1975; Martins et al., 1985).
In addition, there is no published evidence of seasonal changes in the blubber of Kogia, and
some researchers believe that the blubber of Kogia does not change seasonally (personal
correspondance with W.A. McLellan). This study found that Kogia had a lower microvascular
density that consisted of vessels with fewer branches than seen in Tursiops. Perhaps a more
energetically and thermally stable environment did not place the same selective pressure on
Kogia to have enhanced “access” to their blubber energy stores.
Beaked whales also live in the more energetically and thermally stable pelagic
environment and also feed on pelagic cephalopods (Heyning, 1989; Mead, 1989). As with

Kogia, there is no evidence in the literature or from whaling records that beaked whales use their
blubber lipids for energy. Although this study was limited to only two beaked whales, and those
of two different species, their blubber microvascular densities and branching patterns were not
significantly different than those found in terrestrial mammals (Lijnen et al., 2006; Hemmeryckx
et al., 2008; Lijnen et al., 2009; Pasarica et al., 2009), again lending support to the idea that they
have retained the typical mammalian characteristics of adipose vasculature in their blubber in the
absence of selection pressures for needing to mobilize blubber lipids as part of their life history
strategies.
Additional support for the idea that Kogia and the beaked whales may not make
significant use of their blubber for energy can be found in the lipid classes of the species in this
study. The blubber of Kogia and the beaked whales consisted predominantly of waxes. These
animals all had significantly less microvasculature than Tursiops, whose blubber consists solely
of TAG. While the specific reasons for the presence of waxes in the blubber of the deepest-
diving odontocetes have not yet been established, we do know that mammals are typically
incapable of digesting WE (Savory, 1971; Place, 1992; Pond, 1998). Thus, it is possible that
these animals may not be metabolizing the WE once they have been stored in the blubber,
supporting the hypothesis that Kogia and beaked whales are not mobilizing lipid from their
blubber to the same extent as Tursiops. The blubber of Kogia actually consisted of more TAG
than that of the beaked whales (see Table 3). If the idea that blubber mobilization occurs
primarily for TAG is true, then conceivably, Kogia may be mobilizing more lipid from their
blubber than the beaked whales. Such a concept is supported by the vascular data, which suggest
that, at least in the middle and deep layers of blubber, more deposition and mobilization of lipids
may be occurring in Kogia.

Despite the species differences described above, it is quite remarkable that all the
odontocetes in this study had the same microvascular densities, and number of vascular
branches, in the superficial layer of blubber. The conservative nature of vessels in the superficial
layer suggests that there may be constraints on vasculature in the outermost blubber layer, and I
hypothesize that this may be linked to thermoregulation. As a consequence of living in a coastal
habitat in temperate waters, Tursiops experiences significant fluctuations in water temperature.
During the winter, the water in the coastal habitat of Tursiops falls to about 11˚C (Irvine et al.
1981; Wells et al., 1987; Thayer et al., 2003). Kogia and the beaked whales live in a more
consistent environment, with greater exposure to colder water as a function of their deep diving
behavior (Caldwell and Caldwell, 1989; Scott et al., 2001; Beatson, 2007). A thick, lipid rich
layer of insulation is highly important for all species (Worthy and Edwards, 1990; Struntz et al.,
2004; Dunkin et al., 2005; Meagher et al., 2008). It could be that the thermoregulatory need to
prevent heat loss acts as physiological constraint for the amount of microvessels in the
superficial layer of blubber. The thermoregulatory consequences of increased vasculature in the
superficial blubber would be two-fold. First, blood travelling through vessels nearer the surface
of the body would lead to heat loss to the environment (Scholander and Schevill, 1955; Elsner et
al., 1974; Meagher et al., 2008). Second, if increasing the number of capillaries in the superficial
layer of blubber would cause the lipid in that layer to be mobilized, then an important layer of
insulation would be reduced allowing even more heat loss to the environment.
Another possible physiological constraint preventing increased microvascular densities
and metabolic use of the superficial layers of blubber might be the maintenance of a streamlined
body shape. By having a layer of blubber that is not metabolized surround the body, odontocetes
may maintain their streamlined form, which reduces the drag forces acting upon their bodies and

thus decreasing the energy needed when swimming (Ryg et al., 1988; Pabst, 1990; Koopman et
al., 2002; Pabst et al., 1999; Hamilton et al., 2004).
Conclusions and Future Directions
Here I have provided the first detailed, quantitative description of the microvasculature in
the blubber of marine mammals. Although the distribution and characteristics of blubber
microvasculature in this study did correlate with dive depth when only the marine mammals
were considered, the strong similarities across vasculature patterns in terrestrial mammals and all
odontocetes except Tursiops make it more likely that the microvascular densities are not directly
related or the consequence of dive depth. Rather, it seems plausible that the microvascular
densities observed here instead reflect varying degrees of lipid mobilization, an idea that begs
further investigation. Our limited understanding of the species analyzed and the limited number
of species used in this study prevents us from fully evaluating the relationships between the
microvasculature of blubber and the biology and life history of marine mammals.
This study has provided a good foundation for future work on vasculature in marine
mammals. In addition, it has raised many questions. To test the hypotheses formed in this study,
and to elucidate the relationships between blubber vasculature and food supply, lipid
mobilization, phylogeny, and other environmental conditions. The first goal of future studies
should be to analyze more species with consideration of the influence of phylogeny, life history
strategies, habitat, behavior, thermoregulation, and energy demands. In addition, analysis of a
series of life history stages within a species would allow us to evaluate how blubber vasculature
develops. Finally, placing these data within the broader mammalian context, including not only
the closely related baleen whales, but also the unrelated pinnipeds and sirenians, would provide

significant insight into evolutionary patterns and shared constraints across these convergent
species.

LITERATURE CITED
Ackman, R.G., Eaton, C.A., and C.A. Litchfield. 1971. Composition of wax esters, triglycerides
and diacyl glyceryl ethers in the jaw and blubber fats of the Amazon River dolphin (Inia
geoffrensis). Lipids 6: 69-77.
Ackman, R.G., Eaton, C.A., and P.M. Jangaard. 1965. Lipids of the fin whale (Balaenoptera
physalus) from north Atlantic waters. Canadian Journal of Biochemistry 43: 1513-1520.
Ackman, R.G., Hingley, J.H., Eaton, C.A., Sipos, J.C., and E.D. Mitchell. 1975. Blubber fat
deposition in mysticeti whales. Canadian Journal of Zoology 53: 1332-1339.
Aguilar, A. and A. Borrell. 1990. Patterns of lipid content and stratification in the blubber of fin
whales (Balaenoptera physalus). Journal of Mammology 71: 544-554.
Alroy, J. Goyal, V., and E. Skutelsky. 1986. Lectin histochemistry of mammalian endothelium.
Histochemistry 86: 603-607.
Andersen, P. and J. Henriksson. 1977. Capillary supply of the quadriceps femoris muscle of
man: adaptive response to exercise. Journal of Physiology 270: 677-690.
Anstrom, J.W., Thore, C.R., Moody, D.M., Challa, V.R., Block, S.M., and W.R. Brown. 2004.
Histological analysis of vascular patterns and connections in the ganglionic eminence of
premature neonates. Nueroembryology 5(3): 4-12.
Barbieri, M. 2005. Physiological and behavioral mechanisms of thermoregulation in bottlenose
dolphins (Tursiops truncatus) in Sarasota, Florida. MS thesis, Biology and Marine
Biology ,University of North Carolina Wilmington, pp 69.
Barros, N.B. and D. K. Odell. 1990. Food habits of bottlenose dolphins in the southeastern
United States. In: S. Leatherwood and R. Reeves (eds.) The bottlenose dolphin.
Academic Press, London, U.K.
Barros, N.B., and R.S. Wells. 1998. Prey and feeding patterns of resident bottlenose dolphins
(Tursiops truncatus) in Sarasota Bay, Florida. Journal of Mammalogy 79:1045-1059.
Bassos, M.K. 1993. A behavioral assessment of the reintroduction of two bottlenose dolphins.
M.Sc. Thesis, University of California, Santa Cruz, CA. 84 pp.
Beatson, E. 2007. The diet of pygmy sperm whales, Kogia breviceps, stranded in New
Zealand: implications for conservation. Reviews in Fish Biology and
Fisheries 17: 295-303.
Bryden, M.M. 1968. Growth and function of the subcutaneous fat of the elephant seal.
Nature 220(5167): 597-599.

Bryden, M. M. and G. S. Molyneux. 1978. Arteriovenous anastomoses in the skin of seals II.
Anatomical Record 191: 253-260.
Budge, S.M., Iverson, S.J., and H.N. Koopman. 2006. Studying trophic ecology in marine
ecosystems using fatty acids: A primer on analysis and interpretation. Marine Mammal
Science 22(4): 759-801.
Caldwell, D.K. and M.C. Caldwell. 1989. Pygmy sperm whale, Kogia breviceps (de Blainville,
1838); dwarf sperm whale, Kogia simus (Owen, 1866). In: S.H. Ridgway and R. Harrison
(eds.) Handbook of Marine Mammals, Vol. 4 River Dolphins and the larger toothed
whales. Academic Press, San Diego, CA.
Campbell, N.A., Reece, J.B., and L.G. Mitchell. 1987. Biology fifth edition. Addison Wellsley
Longman, Inc. Melo Park, CA. 1175 pp.
Cockcroft, V.G., and G.J.B. Ross. 1990. Food and feeding of the Indian Ocean bottlenose
dolphin off Southern Natal, South Africa. In: S. Leatherwood and R.R. Reeves (eds.) The
bottlenose dolphin. Academic Press, San Diego, CA.
Cooper, M.H. 2004. Fatty acid metabolism in marine carnivores: implications for quantitative
estimation of predator diets. PhD thesis. Dalhousie University, Halifax, Canada.
Cox, T.M., Read, A.J., Barco, S., Evans, J., Gannon, D.P., Koopman, H.N., McLellan, W.A.,
Murray, K., Nicolas, J., Pabst, D.A., Potter, C.W., Swingle, W.M., Thayer, V.G., Touhey,
K.M., and A.J. Westgate. 1998. Documenting the bycatch of harbor porpoises, Phocoena
phocoena, in coastal gill net fisheries from stranded carcases. Fishery Bulletin
96: 727-734.
Crandall, D.L., Hausman, G.J., and J.G. Kral. 1997. A Review of the microcirculation of adipose
tissue: anatomic, metabolic, and angiogenic perspectives. Microcirculation 4(2): 211-232.
Cros, D., Pearson, C., and M.A. Verity. 1980. Polymyositis-Dermatomyositis diagnostic and
prognostic significance of muscle alkaline phosphatase. American Journal of Pathology
101: 159-176.
Di Giralamo, M., Skinner, N.S.J., Hanley, H.G., and R.G. Sachs. 1971. Relationship of adipose
tissue blood flow to fat cell size and number. American Journal of
Physiology 220(4): 932-937.
Dunkin, R.C., McLellan, W.A., Blum, J.E., and D.A. Pabst. 2005. The ontogenetic changes in
the thermal properties of blubber from Atlantic bottlenose dolphin Tursiops truncatus.
The Journal of Experimental Biology 208: 1469-1480.
Elsner, R. W. 1969. Cardiovascular adjustments to diving. In: H.T. Andersen (ed.) The Biology
of Marine Mammals. Academic Press, New York, NY.

Elsner, R.W., Pirie, J., Kenney, D.D., and S. Schemmer. 1974. Functional circulatory anatomy
of cetacean appendages. In: R. J. Harrison (ed.) Functional Anatomy of Marine
Mammals Vol. 2. Academic Press, New York, NY. 366 pp.
Fawcett, D. W. 1942. A comparative study of the blood vascular bundles in the Florida manatee
(Trichechus latirostris) and in certain cetaceans and edentates. Journal of
Morphology 71:105-133.
Fish, F.E. 1993. Influence of hydrodynamic design and propulsive mode on mammalian
swimming energetics. Australian Journal of Zoology 42:79-101.
Folch, J., Lees, M., and G.H. Sloan-Stanley. 1957. A simple method for the isolation and
purification of total lipids from animal tissues. Journal of Biological
Chemistry 226: 497-509.
Foley, R.C., Reece, R.P., and J.H. Leathem. 1954. Histochemical observations of the bovine
uterus, placenta, and corpus luteum during early pregnancy. Journal of Animal
Science 13: 131-137.
Gannon, D.P., Barros, N.B., Nowacek, D.P., Read, A.J., Waples, D.M., and R.S. Wells. 2005.
Prey detection by bottlenose dolphins, Tursiops truncatus: an experimental test of the
passive listening hypothesis. Animal Behaviour 69(3): 709-720.
Geraci, J. R., and Lounsbury, V. J. 1993. Marine mammals ashore: a field guide for strandings.
Galveston, TX: Texas A&M Sea Grant Publication TAMU-SG-93-601.
Gersh, I.G., and M.A. Still. 1945. Blood vessels in fat tissue relation to problems of gas
exchange. Journal of Experimental Medicine 81: 219-232.
Gerth,W.A. Applicability of Henry's Law to hydrogen, helium, and nitrogen solubility in water
and olive oil at 37°C and pressures up to 300 atmospheres. Archives of Biochemistry
and Biophysics 241: 187-199.
Gomori, G. 1939. Microtechnical demonstration of phosphatase in tissue sections. Proceedings
of the Society for Experimental Biology and Medicine 42: 23-26.
Gomori, G. 1950. A new stain for elastic tissue. American Journal of Clinical Pathology
20: 665-666.
Groat, R.A. 1949. Initial and persisting staining power of solutions of iron hematoxylin lake.
Stain Technology 24: 157-163.
Groscolas, R. 1990. Metabolic adaptations to fasting in emperor and king penguins. In: L.S.
Davis and J.T. Darby (eds.) Penguin Biology. Academic Press, San Diego, CA.

Hallmann, R., Mayer, D.N., Berg, E.L., Broermann, R., and E.L. Butcher. 1995. Novel mouse
endothelial cell surface marker is suppressed during differentiation of the blood brain
barrier. Developmental Dynamics 202: 325-332.
Hamilton, J.L., Dillaman, R.M., McLellan, W.A., and D.A. Pabst. 2004. Structural fiber
reinforcement of keel blubber in harbor porpoise (Phocoena phocoena). Journal of
Morphology 261: 105-117.
Hansen-Smith, F.M., Blackwell, L. H., and G. R. Joswiak. 1992. Expression of muscle capillary
alkaline phosphatase is affected by hypoxia. Journal of Applied Physiology 73(2): 776-
780.
Harper, C.J., McLellan, W.A., Rommel, S.A., Gay, D.M., Dillaman, R.M., and D.A. Pabst. 2008.
Morphology of the melon and its tendinous connections to the facial muscles in
bottlenose dolphins (Tursiops truncatus). Journal of Morphology 269(7): 820-839.
Hausman, G.J. and G.B. Thomas. 1984. The development of the inner layer of back fat in fetal
and young pigs. Journal of Animal Science 58: 1550-1560.
Hausman, G.J., and G.B. Thomas. 1985. Differentiation of blood vessels in the adipose tissue of
lean and obese fetal pigs studied by differential enzyme histochemistry. Acta
Anatomica 123: 137-144
Hausman, G.J. and J.T. Wright. 1996. Cytochemical studies of adipose tissue associated blood
vessels in untreated and thyroxine treated hypophysectomized pig fetuses. Journal of
Animal Science 74: 354-362.
Hausman, G.J., and R.L. Richardson. 1983. Cellular and vascular development in immature rat
adipose tissue. Journal of Lipid Research 24: 522-532.
Hausman, G.J., and R.L. Richardson. 2004. Adipose tissue angiogenesis. Journal of Animal
Science 82: 925-934.
Hemmeryclex, B., Bree, R.V., Hoef, B.V., Vercruysse, L., Lijnen, H.R., and J. Verhaeghe. 2008.
Adverse adipose phenotype and hyperinsulinemia in gravid mice deficient in placental
growth factor. Endocrinology 149(5): 2176-2183.
Herd, J.A., Goodman, H.M., and S.A. Grose. 1968. Blood flow rates through adipose tissue of
unanesthetized rats. American Journal of Physiology 214(2): 263-268.
Heyning, J.E. 1989. Cuvier's beaked whale Ziphius cavirostris G. Cuvier. 1823. In: S.H.
Ridgway and R. Harrison (eds) Handbook of Marine Mammals, Vol 4: River Dolphins
and Larger Toothed Whales. Academic Press, San Diego, CA.

Houser, D.S., Finneran, J., Carder, D., Van Bonn, W., Smith, C., Hoh, C., Mattery, R., and S.
Ridgway. 2004. Structural and functional imaging of bottlenose dolphin (Tursiops
truncatus) cranial anatomy. Journal of Experimental Biology 207: 3657-3665.
Howard, C.V. and M.G. Reed. 1998. Unbiased stereology: three dimensional measurement in
microscopy. Springer-Verlag, New York, NY.
Ikels, K.G. 1964. Determination of the solubility of nitrogen in water and extracted human fat.
USAF School of Aerospace Medicine, Technical Document Report SAM-TDR-64-1.
Ingjer, F. 1979. Effects of endurance training on muscle fiber ATP-ase activity, capillary
supply and mitochondrial content in man. Journal of Physiology 294: 419-432.
Irvine, A.B., Scott, M.D., Wells, R.S., and J.H. Kaufman. 1981. Movements and activities of the
Atlantic bottlenose dolphin, Tursiops truncatus, near Sarasota, Florida. Fishery
Bulletin (U.S.) 79:671-688.
Irving, L, Scholander, P.F., and S.W. Grinnell. 1941. The respiration of the porpoise, Tursiops
truncatus. Journal of Cellular and Comparative Physiology 17:145-168.
Iverson, S.J., Field, C., Bowen, W.D., and W. Blanchard. 2004. Quantitative fatty acids
signature analysis: a new method of estimating predator diets. Ecological
Monographs 74: 211-235.
Käkelä, R. and H. Hyvärinen. 1996. Site-specific fatty acid composition in adipose tissues of
several northern aquatic and terrestrial mammals. Comparative Biochemistry and
Physiology B 115: 501-514.
Käkelä, R., Hyvärinen, H., and P. Vainiotalo. 1993. Fatty acid composition in liver and blubber
of the Saimaa ringed seal (Phoca hispida saimensis) compared with that of the ringed
seal (Phoca hispida botnica) and grey seal (Halichoerus grypus) from the Baltic.
Comparative Biochemistry and Physiology B 105: 553-565.
Kipps, E.K., McLellan, W.A., Rommel, S.A., and D.A. Pabst. 2002. Skin density and its
influence on buoyancy in the manatee (Trichechus manatus latirostris), harbor porpoise
(Phocoena phocoena), and bottlenose dolphin (Tursiops truncatus). Marine Mammal
Science 18(3): 765-778.
Koopman, H.N. 2007. Phylogenetic, ecological, and ontogenetic factors influencing the
biochemical structure of the blubber of odontocetes. Marine Biology 151: 277-291.
Koopman, H.N., Iverson, S.J., and D.E. Gaskin. 1996. Stratification and age-related differences
in blubber fatty acids of the male harbour porpoise (Phocoena phocoena). Journal of
Comparative Physiology B 165: 628-639.

Koopman, H.N., Pabst, D.A., McClellan, W.A., Dillaman, R.M., and A.J. Read. 2002. Changes
in blubber distribution and morphology associated with starvation in the harbor porpoise
(Phocoena phocoena): evidence for regional differences in blubber structure and
function. Physiological and Biochemical Zoology 75(5): 498-512.
Kouidi, E., Albani, M., Natsis, K., Megalopoulos, A., Gigis, P., Guiba-Tziampiri, O.,
Tourkantonis, A., and A. Deligiannis. 1998. The effects of exercise training on muscle
atrophy in haemodialysis patients. Nephrology Dialysis Transplantation 13: 685-699.
Lesser, G., and S. Deutsch. 1967. Measurement of adipose tissue blood flow and perfusion in
man by uptake of 86
Kr. Journal of Applied Physiology 23:621-630.
Lijnen, H.R., Christiaens, V., Scroyen, I., Voros, G., Tjwa, M. Carmeliet, P., and D. Collen.
2006. Impaired adipose tissue development in mice with inactivation of placental growth
factor function. Diabetes 55: 2698-2704.
Lijnen, H.R., Van Hoef, B., Rodrigues, J.A., and J.A. Paramo. 2009. Stromelysin-Z (MMP-10)
deficiency does not affect adipose tissue formation in a mouse model of nutritionally
induced obesity. Biochemical and Biophysical Research Communications 389: 378-381.
Ling, J.K. 1974. The integument of marine mammals. In: R.J. Harrison (ed.) Functional
Anatomy of Marine Mammals, Vol. 3. Academic Press, London, U.K.
Litchfield, C., Greenberg, A.J., Caldwell, D.K., Caldwell, M.C., Sipos J.C., and R.G.
Ackman. 1975. Comparative lipid patterns in acoustical and nonacoustical fatty tissues
of dolphins, porpoises and toothed whales. Journal of Comparative Biochemistry and
Physiology B 50: 591-597.
Lockyer, C.H. 1986. Body fat condition in northeast Atlantic fin whales, Balaenoptera physalus,
and its relationship with reproduction and food resource. Canadian Journal of Fisheries
and Aquatic Sciences 43:142–147.
Lockyer, C.H. 1991. Body composition of the sperm whale Physeter catadon, with special
reference to the possible functions of fat depots. Rit Fiskideildar 12: 1-24.
Lockyer, C.H., McConnell, L.C., and T.D. Waters. 1984. The biochemical composition of fin
whale blubber. Canadian Journal of Zoology 62: 2553-2562.
Martins, H.R., Clarke, M.R., Reiner, F., and R.S. Santos. 1985. A pygmy sperm whale, Kogia
breviceps (Blainville, 1838) (Cetacea: Odontoceti) stranded on Faial Island, Azores,
with notes on cephalopod beaks in stomach. Ciencias Biológicas 6: 63-69.
McComb, R.B., Bowers, G.J.N., and S. Posen. 1979. Alkaline phosphatase. Plenum Press, New
York, NY.

McLellan, W.A., Koopman, H.N., Rommel, S.A., Read, A.J., Potter, C.W., Nicolas, J.R.,
Westgate, A.J., and D.A. Pabst. 2002. Ontogenetic allometry and body composition of
harbour porpoises (Phocoena phocoena) from the western north Atlantic. Journal of
Zoology London 257: 457-472.
Mead, J.G. 1989. Beaked whales of the genus Mesoplodon. In: S.H. Ridgway and R. Harrison
(eds) Handbook of Marine Mammals, Vol 4: River Dolphins and Larger Toothed Whales.
Academic Press, San Diego, CA.
Meagher, E.M., McLellan, W.A., Westgate, A.J., Wells, R.S., Blum, J.E., and D.A. Pabst. 2008.
Seasonal patterns of heat loss in wild bottlenose dolphins (Tursiops truncatus). Journal of
Comparative Physiology B 178: 529-543.
Meagher, E.M., McLellan, W.A., Westgate, A.J., Wells, R.S., Frierson, D.J., and D.A.
Pabst. 2002. The relationship between heat flow and vasculature in the dorsal fin of wild
bottlenose dolphins Tursiops truncatus. The Journal of Experimental
Biology 205: 3475-3486.
Miller, W.H. Jr., Faust, I.M., Goldberger, A.C., and J. Hirsch. 1983. Effects of severe long-term
food deprivation and refeeding on adipose tissue cells in the rat. American Journal of
Physiology Endocrinology and Metabolism 245(1): E74-E80.
Montie, E.W., Fair, P.A., Bossart, G.D., Mitchum, G.B., Houde, M., Muir, D.C.G., Letcher, R.J.,
McFee, W.E., Starczak, V.R., Stegeman, J.J., and M.E. Hahn. 2008. Cytochrome
PY501A1 expression polychlorinated biphenyls and hydroxylated metabolites and
adipocyte size of bottlenose dolphins for the southeast United States. Aquatic
Toxicology 86: 397-412.
Montie, E.W., Garvin, S.R., Fair, P.A., Bossart, G.D., Mitchum, G.B., McFee, W.E., Speakman,
T., Starczak, V.R., and M.E. Hahn. 2008. Blubber morphology in wild bottlenose
dolphins (Tursiops truncatus) from the southeastern United States: influence of
geographic location, age class, and reproductive state. Journal of
Morphology 269(4): 496-511.
Noren, D.P., Williams, T.M., Berry, P., and E. Butler. 1999. Thermoregulation during swimming
and diving in bottlenose dolphins, Tursiops truncatus. The Journal of Comparative
Physiology B 169: 93-99.
Norrby, R., and B. Ridell. 2003. Tumour-type-specific capillary endothelial cell stainability in
malignant B-cell lymphomas using antibodies against CD-31, CD-34, and Factor VIII.
Acta Pathologica, Microbiologica et Immunologica 111: 483-489.
Pabst, D.A. 1990. Axial muscles and connective tissues of the bottlenose dolphin. In: S.
Leatherwood and R.R. Reeves (eds.) The Bottlenose Dolphin. Academic Press,
San Diego, CA.

Pabst, D.A., Rommel, S.A., and W.A. McLellan. 1999. The functional morphology of marine
mammals. In: J.E. Reynolds and S.A. Rommel (eds.) Biology of Marine Mammals.
Smithsonian Institution Press, Washington.
Palmer, E. and G. Weddell. 1964. The relationship between structure, innervation, and function
of the skin of the bottlenose dolphin (Tursiops truncatus). Proceedings of the Zoological
Society of London 143(4): 553-568.
Parry, D.A. 1949. The structure of whale blubber and a discussion of its thermal properties.
Quarterly Journal Microscopical Science 90(1): 13-25.
Pasarica, M., Sereda, O.R., Redman, L.M., Albarado, D.C., Hymel, D.T., Roan, L.E., Rood, J.C.,
Burk, D.H., and S.R. Smith. 2009. Reduced adipose tissue oxygenation in human obesity
evidence for rarefaction, macrophage chemotaxis, and inflammation without an
angiogenic response. Diabetes 58: 718-725.
Persons, E.L. 1929. The relative diameter of immature (reticulocytes) and adult human red blood
cells in health and anemia, especially in pernicious anemia. The Journal of Clinical
Investigation 7(4): 615-629.
Place, A.R. 1992. Comparative aspects of lipid digestion and absorption: physiological correlates
of wax ester digestion. American Journal of Physiology 263:R464–R471.
Pond, C.M. 1978. Morphological aspects and the ecological and mechanical consequences of fat
deposition in wild vertebrates. Annual Review of Ecology and Systematics 9: 519-570.
Pond, C.M. 1998. The Fats of Life. Cambridge University Press, Cambridge, U.K. 337 pp.
Pond, C.M., and C.A. Mattacks. 1985. Body mass and natural diet as determinants of the number
and volume of adipocytes in eutherian mammals. Journal of Morphology 185: 183-193.
Presnell, J.K. and M.P. Schreibman. 1997. Humason’s animal tissue techniques. The Johns
Hopkins University Press, Baltimore, MD.
Ramsay, M.A., Mattacks, C.A., and C.M. Pond. 1992. Seasonal and sex differences in the
structure and chemical composition of adipose tissue in wild polar bears (Ursus
maritimus). Journal of Zoology, London 228: 533-544.
Raun, G.G., Hoese, H.D., and F. Moseley. 1970. Pygmy sperm whales, genus Kogia, on the
Texas coast. Texas Journal of Science 21(3): 269-274.
Ridgway, S.H., Scronce, B.L., and J. Kanwisher. 1969. Respiration and deep diving in the
bottlenose porpoise. Science 166: 1651-1654.

Rommel, S., Pabst, D.A., and W. McLellan. 2002. Skull morphology of marine mammals. In:
W.F. Perrin, B. Wursig, and H. Thewissen (eds.) Encyclopedia of Marine Mammals,
Academic Press, San Diego, CA.
Roper, C.F.E. and R.E. Young. 1975. Vertical Distribution of Pelagic Cephalopods.
Smithsonian Contributions to Zoology 209. Smithsonian Institution Press,
Washington, DC.
Ross, M.H., and W. Pawlina. 2006. Histology: A text and atlas, fifth edition. Lippincott Williams
and Wilkins. Baltimore, Md.
Rossell, S. and E. Belfrage. 1979. Blood Circulation in Adipose Tissue. Physiological
Reviews 59(4): 1078-1104.
Rossell, S., Intaglietta, M., and G.M. Chisolm. 1974. Adrenergic influence on isovolumetric
capillary pressure in canine adipose tissue. American Journal of
Physiology 227: 692-696.
Ryg, M., Smith, T.G., and N.A. Oritsland. 1988. Thermal significance of the topographical
distribution of blubber in ringed seals (Phoca hispida). Canadian Journal of Fisheries and
Aquatic Sciences 45: 985-992.
Samuel, A.M. and G.A.J. Worthy. 2004. Variability in fatty acid composition of bottlenose
dolphin (Tursiops truncatus) blubber as a function of body site, season and reproductive
state. Canadian Journal of Zoology 82: 1933-1942.
Savory, P. 1971. The action of pure pig pancreatic lipase upon esters of long-chain fatty acids
and short-chain primary alcohols. Biochimica et Biophysica Acta 248: 149-155.
Schmidt-Nielsen, K. 1997. Animal Physiology: adaptation and environment. fifth edition.
Cambridge University Press, Cambridge, U.K.
Scholander, P.F. and W.E. Schevill. 1955. Counter-current vascular heat exchange in the fins
of whales. Journal of Applied Physiology 8: 279-282.
Scholander, P.F., Walters, V., Hock, R., and L. Irving. 1950. Body insulation of some arctic and
tropical mammals and birds. Biological Bulletin 99: 259-269.
Scott, M.D., Hohn, A.A., Westgate, A.J., Nicolas, J.R., Whitaker, R.R., and W.B.
Campbell. 2001. A note on the release and tracking of a rehabilitated pygmy sperm whale
(Kogia breviceps). Journal of Cetacean Research and Management 3(1): 87-94.
Serway, R.A. and J.S. Faughn. 1992. College Physics third edition. Sounders College Publishing,
Fort Worth, TX.

Shane, S.H., Wells, R.S., and B. Wursig. 1990. Ecology, behavior, and social organization of
the bottlenose dolphin: A review. Marine Mammal Science 2(1): 34-63.
SPSS, Inc. 1997. SPSS Advanced Statistics 7.5. Chicago, IL.
Struntz, D.J., McLellan, W.A., Dillaman, R.M., Blum, J.E., Kucklick, J.R., and D.A. Pabst.
2004. Blubber development in bottlenose dolphins (Tursiops truncatus). Journal of
Morphology 259: 7-20.
Tabachnick, B.G. and L.S. Fidell. 2007. Using multivariate statistics fifth edition. Pearson
Education, Inc., Boston, MA.
Taylor, M.A. 1994. Stone, bone or blubber? Buoyancy control strategies in aquatic tetrapods. In:
L. Maddock, Q. Bone, and J.M.V. Rayner (eds.) Mechanics and Physiology of Animal
Swimming. Cambridge University Press, Cambridge, U.K.
Thayer, V.G., Read, A. J., Friedlaender, A.S., Colby, D.R., Hohn, A.A., McLellan, W.A., Pabst,
D.A., Dearolf, J.I., Bowles, N.I., Russell, J.R., and K.A. Rittmaster. 2003. Reproductive
seasonality of western Atlantic bottlenose dolphins off North Carolina, USA. Marine
Mammal Science 19 (4): 617-629.
Tomilin, A. G. 1957 (1967 translation). Thermoregulation. In: Cetacea, Vol. IX of Mammals of
the U.S.S.R. and Adjacent Countries. Izdatel'stvo Akademi Nauk SSR, Moskova.
Vincent, M.A., Clerk, L.H., Lindner, J.R., Price, W.J., Jahn, L.A., Leong-Poi, H., and E.J.
Barrett. 2006. Mixed meal and light exercise each recruit muscle capillaries in healthy
humans. American Journal of Physiology Endocrinology and
Metabolism 290: E1191-E1197.
Vogel, S. 2003. Comparative Biomechanics. Princeton University Press, Princeton, NJ. 580 pp.
Wachstein, M. and E. Meisel. 1959. The histochemical demonstration of secretory capillaries in
the pancreas with the aid of substrate specific phosphatases. The Journal of Biophysical
and Biochemical Cytology 6: 119-120.
Webb, P.W. 1984. Body form, locomotion and foraging in aquatic vertebrates. American
Zoologist 24: 107-120.
Wells, R.S. and M.D. Scott. 2002. Bottlenose dolphins. In: Perrin W.F., B. Würsig, & J.G.M.
Thewissen (eds.) Encyclopedia of Marine Mammals. Academic Press, San Diego, CA.
Wells, R. S., M. D. Scott and A. B. Irvine. 1987. The social structure of free-ranging bottlenose
dolphins. In: H. H. Genoways (ed.) Current Mammalogy. Plenum Press, New York, NY.

Werner, J.A., Schunke, M., and B. Tillmann. 1987. Histochemical visualization of lymphatic
capillaries in the rat: A comparison of methods demonstrated at the posterior pharyngeal
surface. Archives of histology, Japan 50(5):505-514.
West, G.C., Burns, J.J., and M. ModaVeri. 1979. Fatty acid composition of Pacific walrus skin
and blubber fats. Canadian Journal of Zoology 57:1249–1255.
Wong, C.W.Y., Wiedle, G., Ballestrem, C. Wehrle-Haller, B., Etteldorf, S., Bruckner, M.,
Engelhardt, B., Gisler, R.H., and B.A. Imhof. 2000. PECAM-1/CD-31 Trans-homophilic
binding at the intercellulaer junctions is independent of its cytoplasmic domain: evidence
for heterophilic interaction with Integrin 3mcis. Molecular Biology of the Cell
11: 3109-3123.
Worthy, G.A.J. and E.F. Edwards. 1990. Morphometric and biochemical factors affecting heat
loss in a small temperate cetacean (Phocoena phocoena) and a small tropical cetacean
(Stenella attenuata). Physiological Zoology 63(2): 432-442.
Würsig, B. 1978. Cetaceans. Science 244: 1550-1557.
Young, H.E. and A.C.Jr. Black. 2004. Adult stem cells. The Anatomical Record Part A
276A:75-102.
Young, R.A. 1976. Fat, energy, and mammalian survival. American Zoologist 16: 699-710.
Zanetta, L., Marcus, S.G., Vasile, J., Dobryansky, M., Cohen, H., Eng, K., Shamamian, P., and
P. Mignatti. 2000. Expression of von Willebrand factor, an endothelial cell marker, is
up-regulated by angiogenesis factors: a potential method for objective assessment of
tumor angiogenesis. International Journal of Cancer 85: 281-288.

Table 1. Specimen information for individual animals used in this study. Species abbreviations
are as follows: Tt, Tursiops truncatus; Kb, Kogia breviceps; Md, Mesoplodon
densirostris; Zc, Ziphius cavirostris; Ss, Sus scrofa.
Species Species ID Age Sex Stranding Location Stranding Date
Tt01 C163 Adult F San Luis Obispo, CA 9-22-2002
Tt02 WAM631 Adult F North Topsail Beach, NC 3-3-2007
Tt03 WAM633 Adult F Sneads Ferry, NC 7-7-2007
Tt04 BRF061 Adult M Nags Head, NC 4-5-2006
Tt05 WAM574 Adult F Shallote, NC 11-11-2001
Tt06 WAM573 Adult F Masonboro Inlet, NC 11-11-2001
Kb01 C71 Adult M Monterey County, CA 5-26-1997
Kb02 C67 Adult M San Mateo, CA 12-17-1996
Kb03 BRF092 Adult F Outer Banks, NC 9-1-2006
Kb04 ASF028 Adult F Masonboro Island, NC 9-13-1998
Kb05 KMS427 Adult F Outer Banks, NC 9-2-2006
Kb06 KMS429 Adult M Outer Banks, NC 9-2-2006
Ss01 Pig01 Adult Na Burgaw, NC 4-2009
Ss02 Pig02 Adult Na Burgaw, NC 4-2009
Ss03 Pig03 Adult Na Burgaw, NC 4-2009
Ss04 Pig05 Adult Na Burgaw, NC 4-2009
Md01 WAM593 Adult M Kure Beach, NC 1-28-2004
Zc02 WAM536 Adult F Long Beach, NC 5-13-1998

Table 2. Microvascular characteristics of blubber in the superficial (S), middle (M), and
deep (D) blubber layers within Tt, Tursiops truncatus; Kb, Kogia breviceps;
Md, Mesoplodon densirostris; Zc, Ziphius cavirostris; and subcutaneous fat of
Ss, Sus scrofa. All values represent means ± SEM.
Percent microvascularity
Tta
Kbb
Ssc
Md Zc
S 3.260±0.391* 3.257±0.526 1.758±0.225 2.770 1.610
M 8.398±0.647 4.377±0.551 1.873±0.223 2.320 1.530
D 9.314±0.723 4.500±0.543 1.883±0.458 1.790 1.990
Microvascular diameter
Tta
Kba
Ssa
Md Zc
S 10.910±0.300 10.655±0.300 10.020±0.280 9.238 10.191
M 9.557±0.171 10.924±0.171 9.825± 0.156 9.091 9.604
D 10.143±0.522 10.534±0.522 9.783± 0.477 9.018 9.824
Microvascular branching
Tta Kb
b Ss
b Md Zc
S 2.753±0.112* 2.917±0.133 2.810±0.059 2.920 2.580
M 5.763±0.639 2.927±0.301 2.635±0.043 2.800 2.540
D 5.547±0.543 2.777±0.148 2.900±0.160 2.440 2.460
* Denotes significant difference among blubber layers/depths. a,b,c
Denotes significant difference among species.
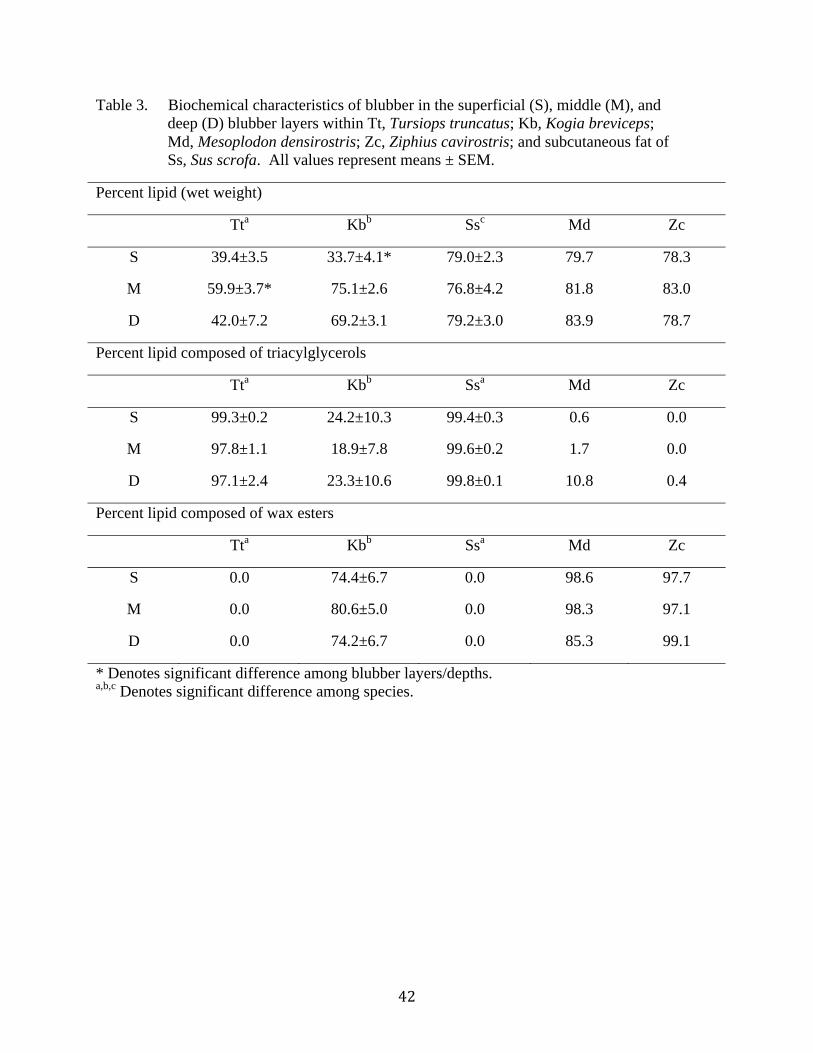
Table 3. Biochemical characteristics of blubber in the superficial (S), middle (M), and
deep (D) blubber layers within Tt, Tursiops truncatus; Kb, Kogia breviceps;
Md, Mesoplodon densirostris; Zc, Ziphius cavirostris; and subcutaneous fat of
Ss, Sus scrofa. All values represent means ± SEM.
Percent lipid (wet weight)
Tta Kb
b Ss
c Md Zc
S 39.4±3.5 33.7±4.1* 79.0±2.3 79.7 78.3
M 59.9±3.7* 75.1±2.6 76.8±4.2 81.8 83.0
D 42.0±7.2 69.2±3.1 79.2±3.0 83.9 78.7
Percent lipid composed of triacylglycerols
Tta Kb
b Ss
a Md Zc
S 99.3±0.2 24.2±10.3 99.4±0.3 0.6 0.0
M 97.8±1.1 18.9±7.8 99.6±0.2 1.7 0.0
D 97.1±2.4 23.3±10.6 99.8±0.1 10.8 0.4
Percent lipid composed of wax esters
Tta Kb
b Ss
a Md Zc
S 0.0 74.4±6.7 0.0 98.6 97.7
M 0.0 80.6±5.0 0.0 98.3 97.1
D 0.0 74.2±6.7 0.0 85.3 99.1
* Denotes significant difference among blubber layers/depths. a,b,c
Denotes significant difference among species.

Table 4. Average Stratification Index (SI) for fatty acids in the blubber/adipose tissue within
Tt, Tursiops truncatus; Kb, Kogia breviceps; Md, Mesoplodon densirostris;
Zc, Ziphius cavirostris; Ss, Sus scrofa. All values represent means ± SEM.
Tta Kb
a Ss
b Md Zc
SI 29.568±2.838 21.455±3.572 5.455±0.727 40.072 23.097
a,b,c Denotes significant difference among species.

Table 5. Select fatty acids and fatty alcohols found in the superficial (S), middle (M), and deep (D) layers of blubber/adipose tissue
in Tt, Tursiops truncatus; Kb, Kogia breviceps; Md, Mesoplodon densirostris; Zc, Ziphius cavirostris; Ss, Sus scrofa.
All values represent means ± SEM.
Tt Kb
Fatty acid S M D S M D
iso 5 1.868±0.663 1.193±0.531 0.618±0.254 0.000 0.000 0.000
14:0 4.645±0.612 5.085±0.506 5.613±0.801 5.045±0.228 5.317±0.220 5.621±0.623
14:1n-5 2.595±0.350 1.277±0.327 0.808±0.289 1.186±0.082 0.894±0.065 0.642±0.097
16:0 6.934±0.941 10.752±0.319 13.147±0.401 11.147±1.186 11.027±0.871 11.191±0.842
16:1n-7 23.508±2.580 18.835±2.048 15.967±1.896 15.861±0.457 12.758±0.797 9.713±1.157
18:0 1.542±0.191 2.355±0.490 2.947±0.269 1.427±0.129 1.399±0.317 1.884±0.228
18:1n-11 0.965±0.263 1.135±0.307 0.917±0.271 0.525±0.382 0.472±0.298 0.343±0.193
18:1n-9 21.137±1.344 19.877±1.239 19.508±1.017 38.368±1.645 37.660±1.908 37.239±2.520
18:1n-7 3.444±0.196 3.582±0.203 3.592±0.213 1.834±0.222 1.755±0.236 1.553±0.275
18:2n-6 1.192±0.046 1.152±0.070 1.157±0.085 0.659±0.083 0.786±0.097 0.612±0.148
20:1n-11 0.670±0.233 0.767±0.289 0.848±0.295 1.568±0.728 2.112±0.991 2.452±0.939
20:1n-9 1.478±0.634 1.713±0.709 1.977±0.710 4.578±0.735 6.470±1.300 8.171±1.567
20:5n-3 3.700±1.006 4.042±0.806 3.625±0.759 0.385±0.120 0.637±0.225 0.674±0.249
22:1n-11 0.618±0.346 0.640±0.448 1.144±0.553 1.990±1.281 3.245±1.941 4.146±2.466
22:5n-3 2.649±0.235 3.405±0.166 3.515±0.215 0.153±0.055 0.246±0.072 0.313±0.064
22:6n-3 6.861±1.641 8.938±1.865 9.810±1.911 0.437±0.115 0.932±0.487 1.209±0.606
Abundant fatty alcohol
alc 16:0 0.000 0.000 0.000 7.587±1.047 9.882±0.999 10.362±1.449
alc 18:1n-9 0.000 0.000 0.000 14.872±2.528 14.622±1.921 13.177±2.065

Table 5 cont.
Ss
Fatty acid S M D
iso 5 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000
14:0 1.290±0.052 1.258±0.048 1.238±0.055 4.051 3.959 3.429 4.378 5.560 6.085
14:1n-5 0.015±0.010 0.003±0.003 0.010±0.010 0.712 0.312 0.177 0.564 0.347 0.262
16:0 20.010±0.713 20.723±0.787 20.950±0.896 4.102 4.337 6.373 7.780 8.859 6.748
16:1n-7 2.338±0.345 2.140±0.303 2.030±0.323 11.712 5.881 4.489 12.226 8.322 5.772
18:0 8.670±0.478 9.928±0.625 10.533±0.723 0.763 1.725 2.237 2.890 3.623 4.621
18:1n-11 0.000 0.000 0.000 5.407 4.107 2.635 0.821 0.468 0.192
18:1n-9 40.168±1.999 39.790±2.011 38.870±2.044 32.949 30.623 28.084 44.237 47.313 45.405
18:1n-7 3.133±0.378 2.948±0.328 2.835±0.303 3.271 3.023 2.723 3.676 3.589 3.121
18:2n-6 19.2125±2.678 18.295±2.501 18.595±2.733 0.814 0.904 0.913 0.393 0.520 0.610
20:1n-11 0.000 0.000 0.000 9.746 11.549 10.686 0.718 0.971 2.755
20:1n-9 0.928±0.045 0.945±0.026 0.940±0.023 9.780 14.523 17.604 3.061 5.010 9.468
20:5n-3 0.000 0.000 0.000 0.153 0.230 0.795 0.000 0.000 0.000
22:1n-11 0.000 0.000 0.000 0.000 9.364 8.611 0.000 0.607 0.453
22:5n-3 0.060±0.014 0.063±0.014 0.063±0.014 0.068 0.181 0.471 0.000 0.503 0.122
22:6n-3 0.000 0.000 0.000 0.186 0.444 1.428 0.000 0.121 0.279
Abundant fatty alcohol
alc 16:0 0.000 0.000 0.000 9.660 10.060 8.330 9.480 11.980 11.060
alc 18:1n-9 0.000 0.000 0.000 13.250 18.220 21.700 22.110 18.670 20.970

Table 6. Mean percent area consisting of microvascularity in a diversity of adipose tissues of
terrestrial mammals.
Species Adipose depot Percent
microvascularity
Source
mouse subcutaneous 0.918% Lijnen et al., 2006
mouse gonadal 0.999% Lijnen et al., 2006
mouse inguinal 2.46% Hemmeryckx et al., 2008
mouse gonadal 2.04% Hemmeryckx et al., 2008
mouse subcutaneous 1.67% Lijnen et al., 2009
human abdominal subcutaneous 1.95-3.48% Pasarica et al., 2009

Figure 1. (a) Sampling site of blubber for all analyses, except those of the pig, Sus scrofa. Image reprinted with permission from
Dr. Sentiel Rommel. (b) Sampling site for adipose tissue of the pig, Sus Scrofa. Image printed with permission from
Jeffrey McClelland. (c) The skin was removed from blubber and adipose tissue samples. These samples were then cut
into thirds and the middle of each third was used for the corresponding depth (superficial, middle, deep). Fat was cut this
way to obtain the different blubber depths for al analyses in this study.

Figure 2. (a) Microvessels in the blubber of Tursiops truncatus (10x). (b) Microvessels in the blubber of Kogia breviceps
blubber (10x). (c) Microvessel in blubber with two terminal branches (d) Microvessel in blubber with three terminal
branches. (e) Microvessel in blubber with five terminal branches. (c,d,e,) Arrows denote terminal branches.
(a) (b)
(d) (c) (e)

Figure 3. Microvasculture in the superficial (S), middle (M), and deep (D) layers of blubber (40x): (a) Tursiops truncatus
(b) Kogia breviceps (c) Sus scrofa (d) Mesoplodon densirostris (e) Ziphius cavirostris

(a) (b) (c)
Blubber
Depth
S
M
D

(d) (e)
Blubber
Depth
S
M
D

Figure 4. Percent area of microvessels analyzed by blubber depth (Tursiops truncatus)
0
2
4
6
8
10
12
Superficial Middle Deep
Mic
rov
ascu
latu
re (
% a
rea)
Depth
Tt01
Tt02
Tt03
Tt04
Tt05
Tt06

Figure 5. Percent area consisting of microvessels analyzed by blubber depth (Kogia breviceps)
0
2
4
6
8
10
12
Superficial Middle Deep
Mic
rov
ascu
latu
re (
% a
rea)
Depth
Kb01
Kb02
Kb03
Kb04
Kb05
Kb06
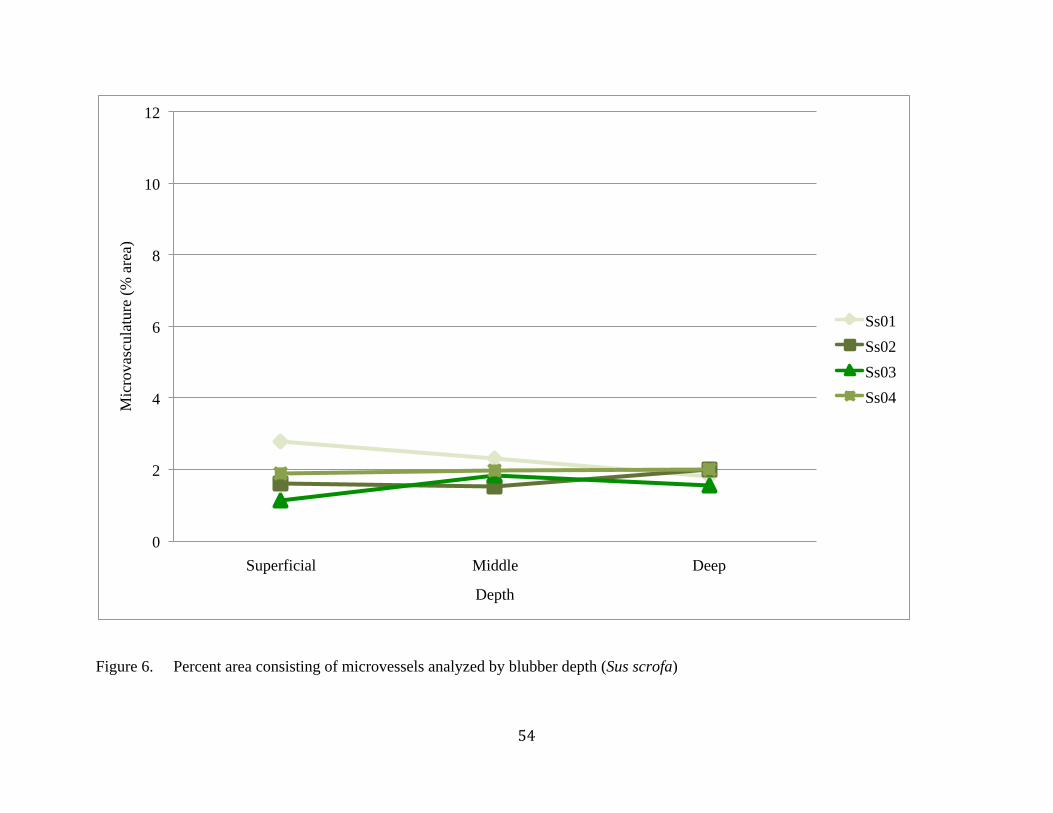
Figure 6. Percent area consisting of microvessels analyzed by blubber depth (Sus scrofa)
0
2
4
6
8
10
12
Superficial Middle Deep
Mic
rov
ascu
latu
re (
% a
rea)
Depth
Ss01
Ss02
Ss03
Ss04

Figure 7. Comparison of Tursiops truncatus, Kogia breviceps, beaked whales (Mesoplodon densirostris and Ziphius
cavirostris), and Sus scrofa percent area of microvessels analyzed by blubber depth (species mean +/- SEM)
0
2
4
6
8
10
12
Superficial Middle Deep
Mic
rov
ascu
latu
re (
% a
rea)
Depth
Tt
Kb
Ss
Md
Zc

Figure 8. Microvasculature diameter of Tursiops truncatus, Kogia breviceps, beaked whales (Mesoplodon densirostris and
Ziphius cavirostris), and Sus scrofa (species mean +/- SEM)
8
8.5
9
9.5
10
10.5
11
11.5
Superficial Middle Deep
Ves
sel
dia
met
er (
m)
Depth
Tt
Kb
Ss
Md
Zc

Figure 9. Microvascular branching of Tursiops truncatus, Kogia breviceps, beaked whales (Mesoplodon densirostris and
Ziphius cavirostris), and Sus scrofa (species mean +/- SEM)
0
1
2
3
4
5
6
7
Superficial Middle Deep
Ter
min
al b
ran
ches
per
ves
sel
Depth
Tt
Kb
Ss
Md
Zc

Figure 10. Percent lipid (wet weight) of Tursiops truncatus, Kogia breviceps, beaked whales (Mesoplodon densirostris and Ziphius
cavirostris), and Sus scrofa (species mean +/- SEM)
0
10
20
30
40
50
60
70
80
90
100
Superficial Middle Deep
Per
cen
t li
pid
(w
et w
eig
ht)
Depth
Tt
Kb
Ss
Md
Zc

Figure 11. Percent of wax esters in the adipose of Tursiops truncatus, Kogia breviceps, beaked whales (Mesoplodon
densirostris and Ziphius cavirostris), and Sus scrofa (species mean +/- SEM)
0
10
20
30
40
50
60
70
80
90
100
Superficial Middle Deep
Wax
est
ers
(% o
f li
pid
)
Depth
Tt
Kb
Ss
Md
Zc

Figure 12. Fatty acid Stratification Index of Tursiops truncatus (Tt), Kogia breviceps (Kb), Sus scrofa (Ss),
Mesoplodon densirostris (Md), and Ziphius cavirostris (Zc) (avg +/- SEM)
0
5
10
15
20
25
30
35
40
45
Tt Kb Ss Md Zc
Str
atif
icat
ion
In
dex
Species





























