Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, May 1969, P. 458-466Copyright @ 1969 American Society for Microbiology
Vol. 98, No. 2Printed In U.S.A.
Macromolecular Synthesis in Saccharomyces cerevisiaein Different Growth Media
C. T. WEHR AND L. W. PARKSDepartment of Microbiology, Oregon State University, Corvallis, Oregon 97331
Received for publication 24 February 1969
Synthesis of ribonucleic acid (RNA), deoxyribonucleic acid (DNA), and proteinwas determined in Saccharomyces cerevisiae during amino acid and pyrimidine star-vation and during shift-up and shift-down conditions. During amino acid starvation,cell mass, cell number, and RNA continued to increase for varying periods. Duringamino acid and pyrimidine starvation, cell mass and RNA showed little increase,whereas total DNA increased 11 to 17%. After a shift from broth medium to a mini-mal defined medium, increase in RNA and protein remained at the preshift rate be-fore assuming a lower rate. DNA increase remained at an intermediate rate duringshift-down, and then dropped to a low rate. During shift-up from minimal to brothmedium, increase in cell number, protein, and DNA showed varying lag periods be-fore increasing to the new rate characteristic of broth medium; each of these quan-tities exhibited a step sometime in the first 2 hr after transfer to rich medium, sug-gesting a partial synchronous division. Immediately after shift-up, RNA synthe-sis assumed a high rate, and then dropped to a rate characteristic of growth in therich medium after about 1 hr.
In the past decade, much work has been doneon the dynamics of bacterial growth and the con-trol of macromolecular synthesis in bacteria. Forcells in balanced growth, the content of variouscellular constituents and their relationships toeach other vary as a function of the growth rate;the cell may show a continuum of physiologicalstates, depending on the growth rate supportedby the medium. Schaechter et al. (12) have shownthat in Salmonella typhimurium, cell mass, ribo-nucleic acid (RNA) and deoxyribonucleic acid(DNA) content, and the number of nucleii percell during balanced growth can be expressed asexponential functions of the growth rate.During a transition between different states of
balanced growth, the close relationship main-tained among the syntheses of various macro-molecular constituents of the cell is temporarilydisrupted. In this period, the rates of synthesis ofthe different classes of macromolecules are dis-sociated until their quantitative relationships arereestablished at a ratio characteristic of balancedgrowth in the new environment. By observing thepattern of synthesis of cell components in a transi-tion between different steady-state conditions,some information can be obtained about themanner in which their synthesis is controlled.During a shift-up from a medium supporting alow growth rate to one supporting a higher rate,
the synthesis of RNA, DNA, protein, and cellmass exhibits a characteristic pattern which is in-dependent of the magnitude of the shift (4). Therate of RNA synthesis is immediately increasedto a high value and subsequently is reestablishedat a rate characteristic of the new medium (3);there is a preferential synthesis of ribosomal pro-tein during this period (13). Increases in DNA,protein, and colony counts exhibit varying lagperiods during which they remain at the preshiftrate. In a shift-down from a medium supportinga high growth rate to one supporting a low rate,net increases in cell mass and RNA immediatelycease, while cell number and DNA continue toincrease at the preshift rate for a period beforedropping to a lower rate (4). The amount of in-crease in DNA during conditions under whichprotein and RNA synthesis is inhibited is con-sistent with the idea that, upon shift-down, allrounds of DNA replication in progress are com-pleted at the preshift rate and that protein synthe-sis is required for the initiation of new rounds.Maal0e and Hanawalt (8) and Hanawalt et al.(1) have shown that during amino acid starvationthere is at least a 40% increase in total DNA; inthis period the fraction of cells synthesizing DNAgradually decreases. Moreover, incubation underconditions which inhibit RNA and protein syn-thesis is accompanied by a gradual increase in the
458
on August 28, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

MACROMOLECULAR SYNTHESIS IN YEAST
fraction of cells immune to thymineless death,suggesting that cells complete replication roundsin progress but require the addition of aminoacids to reinitiate DNA replication. The experi-ments of Lark et al. (6) confirmed the hypothesisthat, in Escherichia coli, DNA synthesis duringamino acid starvation represents continuation ofrounds in progress at the time of amino aciddeprivation to a definite terminus; upon readdi-tion of amino acids, new rounds are initiated at adefinite origin which is recognized after subse-quent periods of amino acid starvation.Saccharomyces cerevisiae provides an op-
portunity to study the relationships among themacromolecular components of the cell undervarying growth conditions in a eukaryotic system.In contrast to bacteria, yeasts possess a definednucleus with as many as 18 genetic linkage groups(9). In addition, the generation time can bevaried by the selection of different growth media.It was the purpose of this investigation to observethe effects of amino acid starvation and shift-upand shift-down conditions on DNA, RNA, andprotein synthesis and cell growth by using a strainof S. cerevisiae requiring two amino acids, apurine, and a pyrimidine.
MATERIALS AND METHODSOrganism and growth conditions. S. cerevisiae
strain 5015-D, a haploid yeast requiring methionine,tryptophan, adenine, and uracil, was used in all ex-periments. Cells were grown aerobically in Wicker-ham's complete medium (15), Wickerham's minimalmedium (15) supplemented with the various re-quirements, or in broth medium (2% glucose, 1%tryptone, 0.5% yeast extract). For measurement ofadenine incorporation, the medium was supplementedwith 5 jg of adenine-8-14C/ml (specific activity 0.0125pAc/,g; New England Nuclear Corp., Boston, Mass.)Cells were grown from a loop inoculum in 100 ml ofmedium in a 250-ml flask for 18 hr on a reciprocalshaker at 30 C, then harvested by centrifugation, andwashed with 0.1 M phosphate buffer (pH 6.6). Forgrowth studies, cells were resuspended in 50 to 300ml of fresh medium in 250-ml or 1-liter flasks andincubated at 30 C on a Dubnoff metabolic shakingincubator. Transfer of cultures to different mediawas accomplished by collecting the cells on Bac-T-Flex membrane filters (Carl Schleicher and SchuellCo., Keene, N.H.), then washing the cells with 1 literof prewarmed medium, and suspending them in newmedium.
Measurement of macromolecular constituents. Fordetermination of total protein, cell samples werewashed with 0.1 M phosphate buffer (pH 6.6), hot5% perchloric acid, and finally with 95% ethylalcohol. Protein was extracted from the residue with1 N NaOH for 10 min at 100 C. Protein in the ex-
tract was measured by the method of Lowry et al. (7).For determination of total DNA, cell samples were
washed with phosphate buffer (pH 6.6) followed by
cold 5% trichloroacetic acid. Lipid was extractedtwice with hot 95% ethyl alcohol, and DNA wasdetermined in the residue by using a modified ver-sion of the Kissane and Robins fluorometric pro-cedure (2).
Total RNA was determined either colorimetricallyor by incorporation of adenine-8-'4C. For colorimetricdeterminations, cell samples were washed twice withcold 0.1 M phosphate buffer (pH 6.6), three times withcold 5% trichloroacetic acid, and twice with 95%ETOH. Nucleic acids were extracted from the residuewith 5% perchloric acid at 70 C for 15 min; RNA inthe extract was determined by the orcinol method.For measurement of the adenine-'4C incorporated,samples were suspended in cold trichloroacetic acidto a final concentration of 5%; cells were collected onmembrane filters and washed with cold 5% trichloro-acetic acid. Filters were placed in scintillation vialsand dried; 5 ml of scintillation fluid [3.0 g of 2,5-diphenyloxazole + 0.1 g of 1,4-bis-2-(5-phenyloxa-zole)-benzene per liter of toluene] was added, andradioactivity was determined by use of a PackardTri-Carb liquid scintillation spectrometer.
Growth of cultures was followed by determinationof total cell number by using a model F Coultercounter; cell clumps were broken up by sonic treat-ment of samples for 30 sec with a Branson cell dis-ruptor. In some experiments, growth was followedby the increase in optical density at 520 nm with aBeckman DU spectrophotometer. All chemicalswere reagent grade obtained from commercial sources.
RESULTSTotal adenine-'4C incorporation as a measure of
total RNA synthesis. Since some difficulty wasencountered in achieving reproducible results byusing the orcinol method to determine totalRNA content in growth studies, it was decidedto use incorporation of adenine-8-14C as a measureof total RNA synthesis. An overnight culture ofstrain 5015-D grown in Wickerham's minimalmedium supplemented with methionine, trypto-phan, uracil, and adenine was transferred tomedium of the same composition but containing5 ,ug of adenine-'4C/ml. After 3 and 6 hr ofaerobic incubation, samples were taken to deter-mine total trichloroacetic acid-precipitable radio-activity and radioactivity released by 12 hr ofincubation in 1 N NaOH. The results showed that2.9% of the trichloroacetic acid-precipitableradioactivity was resistant to 1 N NaOH. Theratio of NaOH-resistant radioactivity to acid-precipitable radioactivity is in good agreementwith the DNA/RNA ratio of cells grown insupplemented Wickerham's minimal medium asdetermined by colorimetric determinations ofRNA and DNA. In a second experiment, anovernight culture of 5015-D grown in Wicker-ham's complete medium was transferred toWickerham's complete medium with adenine-'4Cand incubated at 30 C. After 4 hr of incubation,
VOL. 98, 1969 459
on August 28, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
WEHR AND PARKS
samples were taken to determine total trichloro-acetic acid-precipitable radioactivity and theradioactivity released by incubation in 5% per-chloric acid at 90 C for 20 min. Results showedthat 0.9% of the total cold acid-precipitablecounts were resistant to hot perchloric acid. Theseexperiments suggest that about 98% of the totalcold acid-precipitable radioactivity can be ac-counted for as RNA; in the remaining experi-ments, incorporation of adenine-14C into the coldacid-precipitable fraction was considered to re-flect total RNA synthesis. Measurement of DNAsynthesis by NaOH-resistant incorporation oflabeled adenine gave ambiguous results either byacidification of the NaOH digestion or by neu-tralization followed by adsorption of DNA tomembrane ifiters in the presence of 6X saline-sodium citrate. The values obtained by thefluorimetric procedure of Kissane and Robinswere more consistent, and this method was usedin succeeding experiments.Macromolecular constituents of strain 5015-D at
different growth rates. To determine the RNA,DNA, and protein content of strain 5015-D atdifferent growth rates, overnight cultures ofyeast were grown in Wickerham's minimalmedium supplemented with methionine, trypto-phan, adenine, and uracil, Wickerham's completemedium, or broth medium; they were then washedand resuspended in fresh medium. The cultureswere monitored until well into logarithmicgrowth, and samples were taken for measurementof the various constituents and for total cellcounts (Table 1). The total amount of RNA,DNA, and protein per cell, as measured by theprocedures described above, increased with in-creasing growth rate (except for protein contentof cells grown in the synthetic media). The valuesreported here are higher than those of Ogur et al.(10), perhaps owing to strain differences anddifferent methods of analysis.Amino acid and pyrimidine starvation of strain
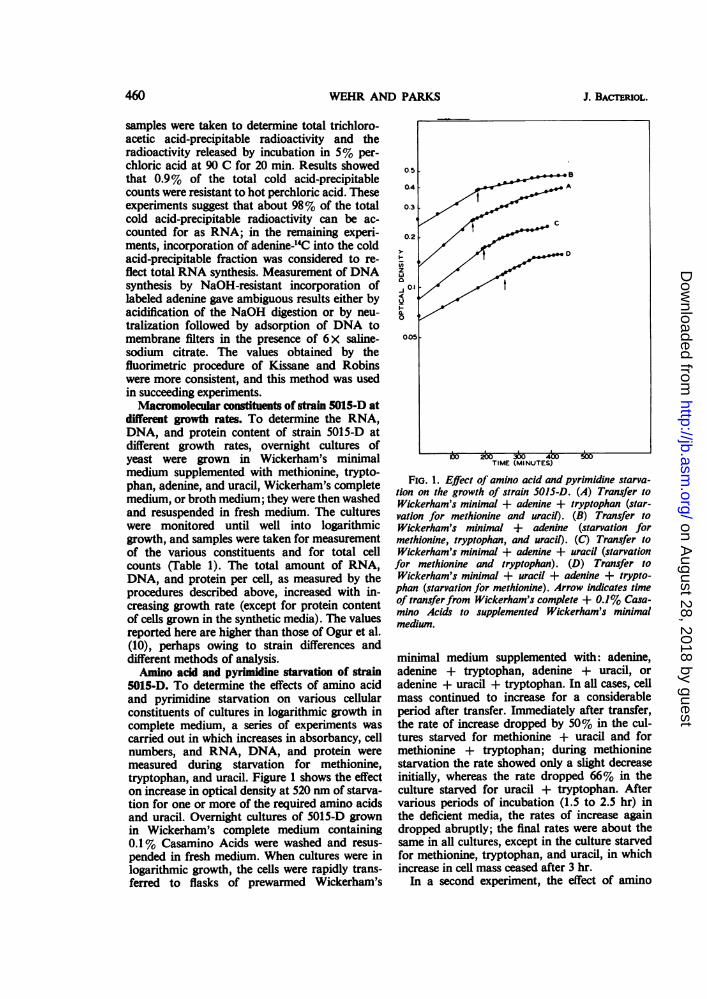
5015-D. To determine the effects of amino acidand pyrimidine starvation on various cellularconstituents of cultures in logarithmic growth incomplete medium, a series of experiments wascarried out in which increases in absorbancy, cellnumbers, and RNA, DNA, and protein weremeasured during starvation for methionine,tryptophan, and uracil. Figure 1 shows the effecton increase in optical density at 520 nm of starva-tion for one or more of the required amino acidsand uracil. Overnight cultures of 5015-D grownin Wickerham's complete medium containing0.1% Casamino Acids were washed and resus-pended in fresh medium. When cultures were inlogarithmic growth, the cells were rapidly trans-ferred to flasks of prewarmed Wickerham's
FIG. 1. Effect of amino acid and pyrimidine starva-tion on the growth of strain 5015-D. (A) Transfer toWickerham's minimal + adenine + tryptophan (star-vation for methionine and uracil). (B) Transfer toWickerham's minimal + adenine (starvation formethionine, tryptophan, and uracil). (C) Transfer toWickerham's minimal + adenine + uracil (starvationfor methionine and tryptophan). (D) Transfer toWickerham's minimal + uracil + adenine + trypto-phan (starvation for methionine). Arrow indicates timeof transferfrom Wickerham's complete + 0.1% Casa-mino Acids to supplemented Wickerham's minimalmedium.
minimal medium supplemented with: adenine,adenine + tryptophan, adenine + uracil, oradenine + uracil + tryptophan. In all cases, cellmass continued to increase for a considerableperiod after transfer. Immediately after transfer,the rate of increase dropped by 50% in the cul-tures starved for methionine + uracil and formethionine + tryptophan; during methioninestarvation the rate showed only a slight decreaseinitially, whereas the rate dropped 66% in theculture starved for uracil + tryptophan. Aftervarious periods of incubation (1.5 to 2.5 hr) inthe deficient media, the rates of increase againdropped abruptly; the final rates were about thesame in all cultures, except in the culture starvedfor methionine, tryptophan, and uracil, in whichincrease in cell mass ceased after 3 hr.
In a second experiment, the effect of amino
460 J. BACTERIOL.
on August 28, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

MACROMOLECULAR SYNTHESIS IN YEAST
acid starvation on increase in total cell numberand total adenine incorporation was investigated.Overnight cultures of 5015-D grown in Wicker-ham's complete medium were washed and re-suspended in fresh medium; when cultures werein logarithmic growth, they were rapidly trans-ferred to Wickerham's minimal medium supple-mented with all requirements except methionine,tryptophan, or methionine and tryptophan (Fig.2). During methionine starvation, the rate of in-crease of total cell number dropped about 20%immediately after transfer to the deficientmedium; after 1.5 hr of incubation the rate againdecreased about 80%. Total adenine incorpora-tion continued at a high rate for about 45 min,then gradually leveled off until, after 2.5 hr ofincubation, adenine incorporation ceased. Totalcounts incorporated during 4 hr of methioninestarvation increased by 140%. During tryptophanstarvation, increase in total cell number droppedto a low rate after 1 hr of starvation. Similarly,adenine incorporation continued at a high ratefor about 30 min and then assumed a low rate forthe balance of the experiment. During 4 hr oftryptophan starvation, total counts incorporatedincreased by about 30%. During starvation forboth methionine and tryptophan, increase in cellnumber assumed a low rate immediately aftertransfer; the rate was again reduced after 2 hr ofstarvation. The rate of adenine incorporation
20000
10000
5000
12
0 0
2 9
0e
'
S4
continued at a high rate initially; after 30 min ofincubation, adenine incorporation ceased.
Figure 3 shows the effect of tryptophan,adenine, and uracil starvation of a culture of5015-D grown on broth medium. An overnightculture of yeast grown in broth medium waswashed and resuspended in fresh medium; whenthe culture was in logarithmic growth, cells wererapidly transferred to Wickerham's minimalmedium with methionine. Increases in opticaldensity and total RNA immediately ceased aftertransfer. However, DNA continued to increaseat approximately the same rate during the first30 min of starvation and showed no further in-crease for the remainder of the incubation period.Total increase in DNA during starvation fortryptophan, adenine, and uracil was about 11 %.The effect of starvation of 501 5-D for
methionine, tryptophan, and uracil on RNA,DNA, and protein synthesis is shown in Fig. 4.An overnight culture of yeast grown in Wicker-ham's complete medium was washed and re-suspended in fresh medium; when the culture wasin logarithmic growth it was rapidly transferredto Wickerham's minimal medium containingadenine. After transfer to the deficient medium,increase in RNA and protein ceased immediately;cell mass increased at a lower rate. For the first10 to 15 min after transfer, DNA increase seemedto continue at about the same rate; it then
100 200 300 400TIME (MINUTES)
150001
10000
n
U sooa~
03J
4
TIME (MINUTES5w
FIG. 2. Effect ofamino acid starvation on cell count and RNA synthesis. (A) Transfer to Wickerham's minimalmedium + adenine + uracil + tryptophan (starvation for methionine). (B) Transfer to Wickerham's minimal +adenine + uracil + methionine (starvationfor tryptophan). (C) Transfer to Wickerham's minimal + adenine +uracil (starvation for methionine and tryptophan). Symbols: (@) cell count, (-) RNA synthesis. Arrow indicatestime oftransferfrom Wickerham's complete to supplemented Wickerham's minimal medium.
B C
VOL. 98, 1969 461
t
on August 28, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

WEHR AND PARKS
0.5
> 04Q
§_ Q4Z 03
0
4
Y02
a.
7
5
2 4
Z 30
2
TIME (MINUTfE%
3'
2 <z
FiG. 3. Synthesis of macromolecular constituentsduring starvation for tryptophan, adenine, and uracil.Symbols: (0) optical density, (A) RNA synthesis, (0)DNA synthesis. Arrow indicates time of transfer frombroth medium to Wickerham's minimal medium +methionine.
dropped to a lower rate for the rest of the in-cubation period. Total increase in DNA duringthe incubation period was about 17%.
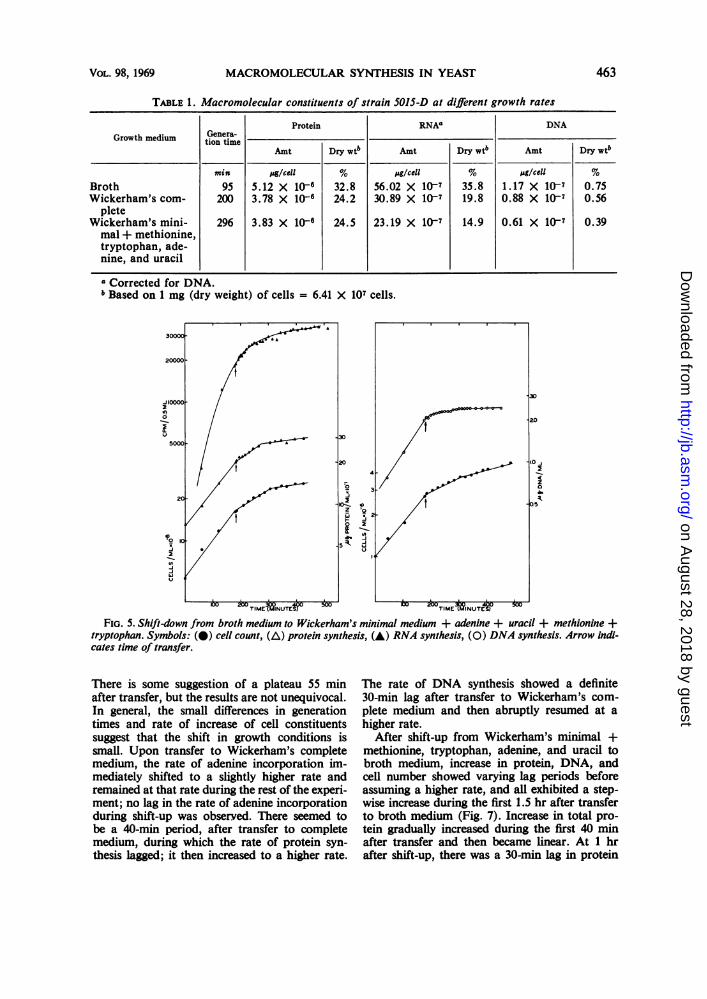
Macromolecular synthesis during shift-downconditions. The amount of DNA, RNA, andprotein per cell varied as a function of the growthrate (Table 1). On the basis of these results, theeffect of macromolecular synthesis of shifting aculture growing logarithmically at one rate to amedium supporting a different growth rate wasinvestigated. In the first case, the effect on RNA,DNA, and protein synthesis was observed duringa shift-down from broth medium to Wickerham'sminimal medium supplemented with methionine,adenine, tryptophan, and uracil. An overnightculture of 5015-D grown in broth medium waswashed and resuspended in fresh medium. Whenthe culture was in logarithmic growth, cells wererapidly transferred to Wickerham's minimal +methionine, tryptophan, adenine, and uracil (Fig.5). Increase in total cell counts showed a con-sistent pattern similar to that observed in theamino acid starvation experiments; the rate ofincrease of cell number dropped 60%, remainedconstant for the first 120 min after shift-down,and then assumed a lower rate. The rate of ade-
TIME (MINUTES)
Fio. 4. Synthesis of macromolecular constituentsduring starvation for methionine, tryptophan, and uracil.Symbols: (0) optical density, (A) protein synthesis,(A) RNA synthesis, (0) DNA synthesis. Arrow indi-cates time of transfer from Wickerham's complete toWickerham's minimal medium + adenine.
nine incorporation remained close to the preshiftrate for the first 30 min after transfer and thengradually decreased over the next 60 min ofincubation in minimal medium. During the re-mainder of the incubation time, adenine in-corporation was roughly linear at a low rate.After transfer to minimal medium, the rate of in-crease in total DNA immediately dropped toabout 14% of the rate in broth medium; after60 min of incubation, increase in DNA exhibiteda brief lag and then continued at a reduced rateduring the rest of the incubation period. Increasein total protein during shift-down conditions re-sembles increase in total cell number; the rate ofincrease dropped immediately upon transfer andwas again reduced after 2 hr of incubation.
Macromolecular synthesis during shift-up condi-tions. In a similar set of experiments, the effect onRNA, DNA, and protein synthesis was investi-gated during transfer of yeast growing at a lowgrowth rate to media supporting a rapid growthrate. Figure 6 shows the results of a shift-up fromWickerham's minimal medium supplementedwith methione, tryptophan, adenine, and uracilto Wickerham's complete medium. Increase incell numbers showed a very brief lag, if any,before being reestablished at a slightly higher rate.
462 J. BACrERIOL.
a -0
30
on August 28, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
MACROMOLECULAR SYNTHESIS IN YEAST
TABLE 1. Macromolecular constituents of strain 501S-D at different growth rates
Protein RNAa DNAGrowth medium Genera-t
tion timeb
Amt Dry wtb Amt Dry wt6 Amt Dry wt6
min pg/cell % pg/cell %g/cellBroth 95 5.12 X 10- 32.8 56.02 X 107 35.8 1.17 X 107 0.75Wickerham's com- 200 3.78 X 10-6 24.2 30.89 X 107 19.8 0.88 X 107 0.56
pleteWickerham's mini- 296 3.83 X 10- 24.5 23.19 X 107 14.9 0.61 X 107 0.39mal + methionine,tryptophan, ade-nine, and uracil
a Corrected for DNA.b Based on 1 mg (dry weight) of cells = 6.41 X 107 cells.
FIG. 5. Shift-down from broth medium to Wickerham's minimal medium + adenine + uracil + methionine +tryptophan. Symbols: (@) cell count, (A) protein synthesis, (A) RNA synthesis, (0) DNA synthesis. Arrow indi-cates time of transfer.
There is some suggestion of a plateau 55 minafter transfer, but the results are not unequivocal.In general, the small differences in generationtimes and rate of increase of cell constituentssuggest that the shift in growth conditions issmall. Upon transfer to Wickerham's completemedium, the rate of adenine incorporation im-mediately shifted to a slightly higher rate andremained at that rate during the rest of the experi-ment; no lag in the rate of adenine incorporationduring shift-up was observed. There seemed tobe a 40-min period, after transfer to completemedium, during which the rate of protein syn-thesis lagged; it then increased to a higher rate.
The rate of DNA synthesis showed a definite30-min lag after transfer to Wickerham's com-plete medium and then abruptly resumed at ahigher rate.
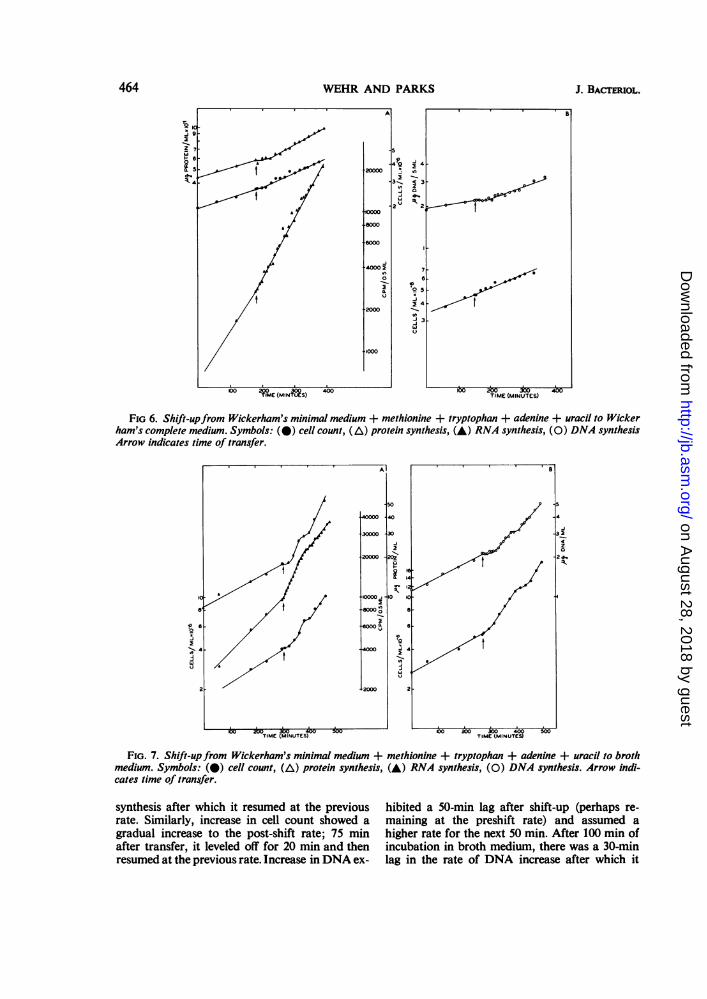
After shift-up from Wickerham's minimal +methionine, tryptophan, adenine, and uracil tobroth medium, increase in protein, DNA, andcell number showed varying lag periods beforeassuming a higher rate, and all exhibited a step-wise increase during the first 1.5 hr after transferto broth medium (Fig. 7). Increase in total pro-tein gradually increased during the first 40 minafter transfer and then became linear. At 1 hrafter shift-up, there was a 30-min lag in protein
463VOL. 98, 1969
on August 28, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
WEHR AND PARKS J. BACTERIOL.
_,__,A-~~~~~ ' -~~j40
10022300 400 100 200400000
4 4 3~~~~~~~~~~~
2 25
21000 -
1.000
00200 300 400 1002000000 40
TIME (MINTUES) TIME (MINUTES)
FIG 6. Shift-upfrom Wickerham's minimal medium + methionine + tryptophan + adenine + uracil to Wickerham's complete medium. Symbols: (@) cell count, (A) protein synthesis, (A) RNA synthesis, (O) DNA synthesisArrow indicates time of transfer.
84J
0 6
*0
-- 4
TIME (MINUTES)00o 300 400 500
TIME (MINUTES)
33
2 a-
FIG. 7. Shift-up from Wickerham's minimal medium + methionine + tryptophan + adenine + uracil to brothmedium. Symbols: (@) cell count, (A) protein synthesis, (A) RNA synthesis, (0) DNA synthesis. Arrow indi-cates time of transfer.
synthesis after which it resumed at the previousrate. Similarly, increase in cell count showed agradual increase to the post-shift rate; 75 minafter transfer, it leveled off for 20 min and thenresumed at the previous rate. Increase in DNA ex-
hibited a 50-min lag after shift-up (perhaps re-maining at the preshift rate) and assumed ahigher rate for the next 50 min. After 100 min ofincubation in broth medium, there was a 30-minlag in the rate of DNA increase after which it
464I ~~~~~B
Al
40000.0.
AW .0 ~~~~~~~~~-
/~~~~~~~~~~~~~~~~O _jI0100I.000
2000
30
10 10/
4J0
3
2
on August 28, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

MACROMOLECULAR SYNTHESIS IN YEAST
assumed the earlier rate. In contrast to the be-havior of other cellular constituents duringshift-up, the rate of RNA synthesis increaseddramatically and remained at a higher rate for thefirst hour after shift-up. After this time the rateabruptly dropped about 50% and remained con-stant for the rest of the experiment. The decreasein the rate of adenine incorporation occurredsimultaneously with the step in cell increase. Thefinal rate of adenine incorporation in brothmedium was only slightly higher than that insupplemented Wickerham's minimal; this wasprobably caused by dilution of the label in thebroth medium.
DISCUSSIONDuring the starvation of S. cerevisiae for amino
acids or amino acids and uracil, cell mass in-creased for a considerable period (as much as a50% increase over 4.5 hr of incubation). Similarly,striking increases in cell count and adenine in-corporation were noted during amino acid anduracil starvation. Yeasts are known to possesslarge intracellular amino acid pools and mayunder some circumstances accumulate exogenousamino acids up to 1000-fold (14); the continuedsynthesis of RNA during amino acid starvationmay represent exhaustion of such intracellularpools. It is of interest that the greatest residualRNA synthesis (140% increase in adenine in-corporated during the first 135 min of incubation)was observed during methionine starvation. It isknown that yeast can accumulate high levels ofmethionine as S-adenosylmethionine, which isthrought to be unavailable to the cell under nor-mal growth conditions (11). The results wouldsuggest that the cell has a relatively largeravailable pool of methionine than of tryptophan.During starvation for methionine, tryptophan,and uracil, however, adenine incorporation im-mediately ceased; the amount of total RNA may,in fact, have dropped slightly (see Fig. 4). Duringstarvation for tryptophan, uracil, and adenine,cell mass and RNA showed little increase; on theother hand, DNA increased at the original ratefor a brief period before leveling off. Total in-crease in DNA was about 11% during starvationfor adenine, uracil, and tryptophan, and about17% during starvation for uracil, methionine,and tryptophan. Williamson (17) showed that, inyeast growing at 25 C with a doubling time ofabout 2 hr, DNA synthesis occupies about thefirst 27% of the cell cycle. If roughly 30% of thecells in a random population were engaged inDNA synthesis at the time of transfer to theamino acid-deficient media, the observed increasein DNA during amino acid and pyrimidinestarvation would be consistent with the idea that
all cells engaged in DNA synthesis will continue,whereas older cells in the population will notinitiate a new cycle of DNA synthesis. It cannotbe determined from the data whether cells activelysynthesizing DNA at the time of amino acidstarvation continue to the completion of a fullcomplement of DNA or complete only a fractionof the genome.
In a shift-down from broth medium to supple-mented Wickerham's minimal medium, adenineincorporation and increase in total protein re-mained at the preshift rate but gradually leveledoff to a rate lower than that characteristic for cellsin balanced growth in minimal medium. Theinitial high rate may, again, represent a largeintracellular amino acid pool so that, within thecell, actual shift-down conditions are not reacheduntil the pools have been depleted by incubationin minimal medium. Upon transfer to minimalmedium, the rate of increase in cell numbershowed an immediate drop to a rate comparableto that of cells in balanced growth in minimalmedium, but after 2 hr of incubation abruptlyunderwent a second drop to a much lower rate.In general, there seems to be a lag in responseamong all three quantities, after which the ratesof increase drop to a low value; the condition ofbalanced growth in minimal medium was not re-established during the course of the experiments.In contrast to the results of the amino acid starva-tion experiments, the rate of DNA synthesisduring shift-down from broth medium to minimalmedium immediately dropped to a low value.When total DNA increased about 10%, therewas a brief lag before DNA synthesis resumed ata lower rate. Here again, the rate characteristicof balanced growth in minimal medium was notattained. However, the increase in total DNAprior to assumption of final low rate was the sameas that observed during tryptophan, adenine, anduracil starvation (Fig. 3).
Although there was little difference betweenthe pre- and post-shift rates of increase in RNA,DNA, protein, and cell number during a shift-upfrom supplemented Wickerham's minimal toWickerham's complete medium, it is of interestthat all quantities except RNA exhibited a slightlag period after shift-up. The rate of RNA syn-thesis immediately assumed a higher rate whichwas maintained for the balance of the experiment.This phenomenon was much more evident in ashift-up from supplemented Wickerham's mini-mal to broth medium. Here RNA synthesisassumed a high rate which was maintained forthe first 70 min after the shift. Kudo and Imahori(5) showed that, upon a shift-up of nitrogen-starved yeast cells to a rich medium, newly syn-thesized RNA appears in the polysome fraction
VOL. 98, 1969 465
on August 28, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

WEHR AND PARKS
and later moves to the 80S fraction, and that thebase composition of this RNA is similar to thatof ribosomal RNA. Thus the increased rate ofRNA synthesis during a shift-up from minimal torich medium may represent a synthesis of riboso-mal RNA. The stepwise increases in total DNAand cell count following shift-up to broth mediumresemble a partially synchronous division, andsuggest that shift-up conditions may bring a por-tion of the cell population into phase with regardto DNA synthesis and cell division. However, astepwise increase in total protein was also ob-served; in a synchronous culture of yeast, proteinincreases continuously throughout the cell cycle(16).
ACKNOWLEDGMENT
During the course of these studies C. T. Wehr has held a NASApredoctoral traineeship.
LITATURE CITED
1. Hanawalt, P. C., 0. Maaloe, D. J. Cumming, and M.Schaechter. 1961. The normal DNA replication cycle. II.J. Mol. Biol. 3:156-165.
2. Kissane, J. M., and E. Robins. 1958. The fluorometric meas-urements of deoxyribonucleic acid in animal tissues withspecial reference to the central nervous system. J. Biol.Chem. 233:184-188.
3. Kjeldgaard, N. 0. 1961. The kinetics of ribonucleic acid andprotein formation in Salmonella typhinurium during thetransition between different states of balanced growth.Biochim. Biophys. Acta 49:64-76.
4. Kjeldgaard, N. O., 0. Maaloe, and M. Schaechter. 1958. Thetransition between different physiological states duringbalanced growth of Salmonella typhhnurlwn. J. Gen.Microbiol. 19:607416.
5. Kudo, Y., and K. Imahori. 1965. The characterization ofnewly synthesized RNA in step up cultures of yeast cells. J.Biochem. 58:364-372.
6. Lark, K. G., T. Repko, and E J. Hoffman. 1963. The effectof amino acid deprivation on subsequent deoxyribonucleicacid replication. Biochim. Biophys. Acta 76:9-24.
7. Lowry, 0. H., N. J. Rosebrough, A. L. Faff, and R. J.Randall. 1951. Protein measurement with the Folin phenolreagent. J. Biol. Chem. 193:265-275.
8. Maal0e, O., and P. C. Hanawalt. 1961. Thymine deficiencyand the normal DNA replication cycle. J. Mol. Biol. 3:144-155.
9. Mortimer, R. K., and D. C. Hawthorne. 1966. Geneticmapping in Saccharomyces. Genetics 53:165-173.
10. Ogur, M., S. Minckler, G. Lindegren, and C. Lindegren. 1962.The nucleic acids in a polyploid series of Sacchrwomyces.Arch. Biochem. Biophys. 40:175-184.
11. Pigg, C. J., W. A. Sorsoli, and L. W. Parks. 1964. Induction ofthe methionine-activating enzyme in Saccharomyces cerevi-siae. J. Bacteriol. 87:920-923.
12. Schaechter, M., 0. Maaloe, and N. 0. Kjeldgaard. 1958. De-pendency on medium and temperature of cell size andchemical composition during balanced growth of Salmonellatyphimurium. J. Gen. Microbiol. 19:5924606.
13. Schleif, R. 1967. Control of production of ribosomal protein.J. Mol. Biol. 27:41-55.
14. Surdin, Y., W. Sly, J. Sire, A. M. Bordes, and H. DeRobichon-Szulmajster. 1965. Proprietes et controle geneti-que du systeme d'accumulation des acides amines chez Sac-charomyces cerevislac. Biochim. Biophys. Acta 167:546-566.
15. Wickerham, L. J. 1946. A critical evaluation of the nitrogenassimilation tests commonly used in the classification ofyeasts. J. Bacteriol. 52:293-301.
16. Williamson, D. H. 1964. Division synchrony in yeasts, p. 372-373. In E. Zeuthen (ed.), Synchrony in cell division andgrowth. John Wiley & Sons, Inc., New York.
17. Williamson, D. H. 1965. The timing of deoxyribonucleic acidsyntheds in the cell cycle of Saccharomyces cerevlslae. J.Cell Biol. 25:517-528.
466 J. BACTERIOL.
on August 28, 2018 by guest
http://jb.asm.org/
Dow
nloaded from





























