Embed Size (px)
Citation preview

La explotación de especies pequeñas en reg-istros arqueológicos de sociedades cazado-ras recolectoras está siendo analizada con
gran interés en diversas regiones del cono sur deAmérica. En la cordillera chilena el pequeño roe-dor Lagidium viscacia (la vizcacha andina) fuecazado acentuadamente y le fueron aplicadasacciones de carnicería complejas durante la tran-sición Pleistoceno-Holoceno (Labarca 2005). Enla región Pampeana de Argentina un roedor semi-acuático de mayor talla, el coypo (Myocastor coy-pus) fue cazado y procesado intensamente en sitiosde la Depresión del Río Salado (Figura 1) dondeocupó un rol central en la economía durante elHoloceno Tardío final (Escosteguy 2007; González1997). En las zonas áridas del Norte de Argentinael roedor Dolichotis patagonum fue consumido
durante el Holoceno Tardío (del Papa et al. 2010).También se está dando a conocer la explotación demicroroedores como los cuises Galea tixiensis yCavia aperea (Caviidae) que fueron cazados en lassierras de Tandilia en tiempos tardíos y aprovecha-dos exhaustivamente, incluso su pequeña piel(Quintana 2005). Otros micro-roedores silvestrescomo Ctenomys sp., Microcavia sp. y cricétidosfueron explotados en diversas regiones deArgentina (Fernández et al. 2009; Medina 2008;Santiago 2004). Además de los roedores, en sitiosarqueológicos de distintas áreas de la región Pam-peana está siendo explorado el uso de otros verte-brados pequeños como armadillos, reptiles, pecescontinentales y aves lacustres (González 2005;Martínez et al. 2005; Quintana y Mazzanti 2001;Stoessel 2007).
LAS VIZCACHAS PAMPEANAS (LAGOSTOMUS MAXIMUS, RODENTIA)EN LA SUBSISTENCIA INDÍGENA DEL HOLOCENO TARDÍO DE LAS
SIERRAS DE TANDILIA ORIENTAL (ARGENTINA)
Carlos A. Quintana y Diana L. Mazzanti
Se describen las evidencias de procesamiento carnicero halladas en los esqueletos del roedor Lagostomus maximus (viz-cacha pampeana) provenientes de sitios arqueológicos del Holoceno Tardío Final (1000 a 600 a.P.) de las sierras de TandiliaOriental (provincia de Buenos Aires, Argentina). La diversidad ósea registrada, la cantidad de ejemplares, las huellas decorte, las de raspado y las fracturas intencionales indican que las vizcachas fueron ingresadas enteras a los sitios y que seles aplicaron procesos de desollado, desarticulación, descarne y fracturas. La comparación con la explotación de la viz-cacha durante milenios anteriores en las mismas sierras muestra que sólo hacia el Holoceno Tardío Final esta especieadquirió mayor importancia numérica y le fueron aplicados procesos carniceros más elaborados. Este fenómeno está vin-culado al desarrollo de una subsistencia de intensificación y diversificación en la cual la caza menor fue preponderante enTandilia.
Butchering evidence found in skeletons of the plains vizcacha (Lagostomus maximus) from Late Holocene (1000 to 600 b.P.)archaeological sites in the Western Edge of Tandilia Range (Buenos Aires Province, Argentina) are described. Skeletal diver-sity, specimen abundance, and cut marks, especially scraping marks and intentional fractures, indicate that entire plains viz-cacha carcasses were introduced to the sites where they were processed through skinning, disarticulation, filleting, andfracturing for marrow. Comparison with earlier vizcacha exploitation in the same area suggests that vizcacha increased innumerical importance and that the butchery process was elaborated only toward the end of the late Holocene. This phenom-enon is linked to the development of diversification and intensification of areal economies in the Pampean Region where smallgame was preponderant in the Tandilia Range.
Carlos A. Quintana y Diana L. Mazzanti � Laboratorio de Arqueología, Universidad Nacional de Mar del Plata. Bélgica2293, 7600 ([email protected]) Mar del Plata, provincia de Buenos Aires, Argentina
Latin American AntiquityP22(2), 2011, pp. 253-270Copyright ©2011 by the Society for American Archaeology
253

254 AmerIcAN ANTIquITy [Vol. 22, No. 2, 2011
Estos antecedentes muestran un panoramaamplio y más importante de la estructura del con-junto de especies incorporado a la subsistencia delo que se pensaba unos pocos años atrás cuando losanálisis zooarqueológicos centralizaban su interésen la explotación de ungulados. Las especiespequeñas como la vizcacha y los cuises en las sier-ras de Tandilia, el coypo en el Río Salado o la viz-cacha andina en la Cordillera fueron recursos que
cumplieron roles críticos por lo que deben ser con-siderados junto a los mamíferos grandes para cono-cer de modo integral la dinámica de la subsistencia.La talla y la anatomía de las presas de menor masaposibilitan la aplicación de técnicas de proce-samiento y de despiece diversas que, a su vez,pueden ser radicalmente diferentes a las conocidasen ungulados. En este sentido el modo y la inten-sidad del procesamiento es un factor que informa
Figura 1. Ubicación geográfica de los sitios analizados. (1) Sistema serrano de Tandilia; (2) Área Interserrana; (3)Sistema serrano de Ventania; (4) Depresión del Río Salado; (5) Cueva El Abra; (6) Cueva Tixi; (7) Lobería I; (8)Localidad Arqueológica Amalia.

sobre la función de este tipo de recursos en laestrategia de explotación (Quintana 2005). Porejemplo en áreas distantes de la región Pampeanael aprovechamiento de animales menores a 15 kgde masa presenta estilos diversos de procesamientocarnicero y de intensidad de captura en momentosdistintos del Holoceno, lo cual es consecuente concada tipo de estrategia de subsistencia (Quintana2009).
En este contexto, el análisis detallado de lasacciones de carnicería aplicadas sobre las carcasasde presas pequeñas específicas se presenta comorelevante para comprender los cambios o la esta-bilidad en las estrategias de subsistencia de ocu-paciones de cazadores recolectores de la regiónPampeana. Así, en este artículo se analiza unaespecie, la vizcacha pampeana (Lagostomus max-imus), en tanto que presenta un registro geográficoextendido y temporalmente amplio de la cual análi-sis preliminares mostraron que su explotación fuevariable a través de la secuencia estratigráfica.
Métodos
Los restos óseos analizados fueron recuperadosdurante campañas de excavaciones y extraídos pormedio de decapados de potencias variables, eltamizado fue en seco o bajo agua con malla de 2mm de poro. Los fechamientos radiocarbónicosfueron realizados en la Universidad de Arizonausando un acelerador de espectroscopia de masas(AMS) sobre muestras de carbón vegetal prove-niente de estructuras de combustión (Mazzanti1997, 1999; Mazzanti et al. 2009). La identificacióntaxonómica de los huesos de Lagostomus maximusse efectuó sobre la base de caracteres diagnósticosy por la comparación con esqueletos de ejemplaresactuales. Los materiales están depositados en lacolección del Laboratorio de Arqueología de laUniversidad Nacional de Mar del Plata.
Para determinar el uso antrópico de los huesosde vizcacha se buscaron evidencias en las super-ficies corticales que validen objetivamente la exis-tencia de una interacción entre esos restos y lassociedades que habitaron las cuevas. Se descar-taron los indicios que pudieron ser consecuenciade procesos secundarios, naturales o posdeposita-cionales (como quemado, distribución espacial,abundancia, etc.) (Stahl 1996). Los mismos crite-rios fueron aplicados para incorporar antecedentes
de otros autores. Para conocer el procesamientoefectuado sobre los animales se evaluaron las huel-las de filos líticos impresas sobre la superficie delos huesos como consecuencia del corte de partesblandas. La identificación de esas trazas como pro-ducto de la acción humana se basó sobre los cri-terios discutidos por Bunn (1981, 1983), PérezRipoll (1992), Potts y Shipman (1981) y Shipmany Rose (1983) entre otros. La búsqueda e identi-ficación de huellas de filos líticos se efectuó entodos los huesos recolectados empleando una lupatriocular de hasta 45 aumentos. Cada rasgo hal-lado fue descrito y registrado en una base de datosy su imagen capturada digitalmente mediante unacámara de video Motic 2000 incorporada a la lupa.Las fracturas antrópicas se identificaron siguiendoel criterio de Johnson (1985), Lyman (1994) y Villay Mahieu (1991) y se registraron en la citada basede datos.
En este artículo el término despiece y proce-samiento carnicero (butchering) se refiere al actocompleto de reducir una carcasa para su consumoo extracción de materias primas (cuero, huesos,grasa, etc.) (Abe et al. 2002), el cual se componede diversas acciones de carnicería (desollado,descarne, desarticulación, etc.). En consecuenciase analizaron dos categorías para reconocer elprocesamiento de una carcasa a partir de las huel-las de corte:
1. la identificación de una o más acciones de car-nicería
2. la integración de esas acciones para determi-nar la secuencia y la regularidad del despiece.
La interpretación de la función de cada corte serealizó vinculando su ubicación con la anatomía dela vizcacha con el fin de conocer el efecto sobre lamusculatura o la piel. En los casos disponibles setuvo en cuenta la recurrencia de huellas en áreassimilares y su distribución en sectores comple-mentarios (articulación distal-proximal,mandíbula-maxilar, etc.).
Para evaluar la preservación diferencial de cadaparte del esqueleto se calculó la abundancia rela-tiva, definida como la proporción entre el númerode elementos registrado de cada huesorespecto del número de elementos esperadoado en función del número mínimo de individuosNISPi x 100/NISPie x MNI, siendo:
NISPi: número del elemento i en la muestra
quintana y mazzanti] LAS VIZcAcHAS PAmPeANAS eN LA SuBSISTeNcIA INDÍGeNA DeL HOLOceNO TArDÍO 255

NISPie: número del elemento i en elesqueleto
MNI: número mínimo de individuos de la muestraEste índice tiene el objetivo de conocer la preser-
vación de cada parte esquelética en función delnúmero mínimo de individuos, lo cual permite lacomparación entre sitios independientemente deltamaño de la muestra (ver una descripción másdetallada en Andrews 1990).
La vizcacha pampeana:Lagostomus maximus
La vizcacha pampeana (Lagostomus maximus) esun roedor caviomorfo que pertenece a la familiaChinchillidae, la cual actualmente se compone detres géneros llamados comúnmente vizcachas ychinchillas: Lagostomus, Lagidium y Chinchilla.El registro fósil de Lagostomus se inicia durante elPleistoceno temprano (Vucetich y Verzi 1995),aunque durante el Plioceno se presenta el géneroLagostomopsis, el cual está poco estudiado y posi-blemente sea un sinónimo de Lagostomus en vir-tud de que las semejanzas entre ambos taxones sontan estrechas como las que hay entre especies deLagidium. Los chinchíllidos son exclusivos deAmérica del Sur, particularmente de la franja and-ina que ocupa desde el extremo sur de Argentina yChile hasta el sur de Ecuador (Spotorno et al. 2004;Werner et al. 2006). Lagostomus maximus es elúnico chinchíllido que habita fuera del área and-ina, desde las planicies y sierras bajas de la regiónPampeana hasta el sur de Bolivia y el oeste deParaguay (Jackson et al. 1996).
La vizcacha pampeana se caracteriza por sushábitos subterráneos y gregarios (Branch 1993). Esun roedor con un fuerte dimorfismo sexual (Llanosy Crespo 1952) manifestado principalmente en eltamaño del cuerpo, las hembras pueden alcanzarlos 5 kg de masa y los machos hasta 9 kg (Jacksonet al. 1996). Se trata de la especie más grande dela familia y con un esqueleto robusto por sus adapta-ciones a la excavación. Sus poblaciones están com-puestas por colonias subterráneas (vizcacheras) quegeneran montículos evidentes en el paisaje forma-dos por la tierra extraída de las galerías. El ciclo devida de esta especie es algo complejo, en tanto queel comportamiento social vinculado a la reproduc-ción condiciona la interacción entre grupos etariosy géneros, y al tiempo de actividad dentro o fuera
de las madrigueras en distintas épocas del año(Branch 1993) lo que impacta sobre su disponibil-idad para la caza.
El registro de la vizcacha en Tandilia Oriental
En los sitios Cueva Tixi, Cueva El Abra, LoberíaI y Amalia Sitio 4 (Figura 1) se identificaron sieteocupaciones con explotación de vizcacha dis-tribuidas entre el Pleistoceno Tardío y tiemposposconquista (Mazzanti 2007; Quintana 2009;Quintana y Mazzanti 2001). Los sitios con ocu-paciones de sociedades cazadoras recolectoras soncuevas ubicadas en lugares estratégicos de las pen-dientes serranas, y con un registro numeroso defauna tanto de especies grandes (ungulados) comopequeñas (roedores, carnívoros, armadillos, mar-supiales, peces, aves y reptiles) (Mazzanti 1997,2009; Quintana et al. 2003). Durante el PleistocenoTardío este roedor fue aprovechado en Cueva ElAbra (Componente Inferior; 9800 a.P.) y en CuevaTixi (Nivel Arqueológico 1; 10,400 a.P.). Se reg-istraron uno (NISP= 12) y dos (NISP= 32) indi-viduos en cada sitio con escasas evidencias deexplotación, lo cual impide proponer una secuen-cia de procesamiento (Quintana y Mazzanti 2001).Durante el Holoceno Medio se cuenta con el reg-istro de Cueva Tixi (Nivel Arqueológico 2; 4800a.P.), el cual presenta un individuo de vizcacha y12 huesos, de los que sólo un omóplato tiene huel-las de corte (Valverde 2001). Mientras que en labase del Holoceno tardío de Cueva Tixi (NivelArqueológico 3; 3200 a.P.) la vizcacha está repre-sentada por cuatro restos atribuibles a un indi-viduo.
De todas las ocupaciones indígenas el registrode la vizcacha es más acentuado durante el Holo-ceno Tardío final preconquista (1000 a 600 a.P.) locual se manifiesta en la mayor cantidad de ejem-plares y en la calidad de las evidencias deexplotación en los tres sitios analizados más abajo.
A partir del contacto hispano indígena las evi-dencias son escasas, en el Sitio 4 de la LocalidadArqueológica Amalia (siglo dieciocho) (Mazzanti2007) se identificaron 19 huesos correspondientesa dos ejemplares con evidencias de exposición alfuego. Todos estos sitios arqueológicos se localizanen sectores serranos con valles amplios y cimasmesetiformes con abundantes sedimentos donde
256 LATIN AmerIcAN ANTIquITy [Vol. 22, No. 2, 2011

actualmente se distribuyen numerosas poblacionesde vizcacha.
La explotación de la vizcacha durante elHoloceno Tardío Final
La vizcacha pampeana fue registrada en tres ocu-paciones de cazadores recolectores correspondi-ente al Holoceno Tardío Final (1000 a 600 a.P.) delos sitios arqueológicos Cueva El Abra, Cueva Tixiy Lobería I. En los tres sitios los restos sonnumerosos, tienen buena calidad de preservacióny claras evidencias de su manipulación antrópica.Las alteraciones naturales son escasas, correspon-den a fracturas posdeposicionales y, en el caso deCueva Tixi, a marcas de raíces. No se registraronfracturas o mordeduras de carnívoros ni morde-duras de humanos. Las marcas de dientes de roe-dores afectaron a menos del uno por ciento de loshuesos de vizcacha sin alterar de modo significa-tivo la anatomía o la superficie cortical.
Cueva El Abra, Componente Superior (958 ± 32 a.P. AA-33419)
Los restos de vizcacha en este sitio son los másnumerosos de la región, estando representadas casitodas las partes del esqueleto. La epífisis proximalde la ulna es la parte mejor preservada propor-cionalmente, seguida por el omóplato, el calcáneoy las ramas mandibulares (Tabla 2).
Hay un buen registro de piezas anatómicas conhuellas de corte (6.83 por ciento). A diferencia delos otros sitios contemporáneos, algunos huesosdel autopodio posterior (metatarsianos y falanges)tienen cortes principalmente en la superficie ante-rior (Tabla 3, Figura 2 y 3). El fémur y el húmeroson las piezas más procesadas (20 por ciento), pre-sentando huellas en casi toda la superficie anterior.Los rasgos se disponen mayormente en posicióntransversal al eje del hueso y son cortos. Algunashuellas de corte se agrupan en conjuntos subpara-lelos que parecen micro-raspados, mientras quedos fémures y tres ulnas tienen cortes transversales
quintana y mazzanti] LAS VIZcAcHAS PAmPeANAS eN LA SuBSISTeNcIA INDÍGeNA DeL HOLOceNO TArDÍO 257
Tabla 1. Diversidad de recursos incorporados a la subsistencia durante el Holoceno Tardío Final de Tandilia Oriental.
Cueva El Abra Lobería I Cueva Tixi
NISP MNI% NISP MNI% NISP MNI%
ArtiodáctilosLama guanicoe (guanaco) 18 1.44 75 11.36 66 2.01Ozotoceros bezoarticus (venado) 113 1.44 178 4.54 100 1.34
RoedoresCaviinae 2814 - 222 - 1850 -Galea tixiensis (cuis) 199 45.3 42 15.9 245 57.1Cavia aperea (cuis) 89 25.1 29 13.63 79 17.4Lagostomus maximus (vizcacha) 468 8.63 382 29.54 180 4.02Myocastor coypus (coypo) 17 2.16 - - 4 1.34
DasipódidosZaedyus pichiy (pichi) 108 .72 32 2.27 228 .67Chaetophractus villosus (peludo) 2230 3.60 769 4.54 1972 2.68Dasypus hybridus (mulita) 783 1.44 1208 6.81 413 .67
CarnívorosPuma concolor (puma) - - - - 1 .67
ReptilesTupinambis cf. merianae (lagarto overo) 279 4.31 45 4.54 150 4.69
PecesRhamdia sapo (bagre) 3 .72 - - 8 .67
AvesRhea americana (ñandú) 1 .72 6 2.27 1 .67Rhea americanaP(huevosS) 252.9 g 2.16 16 g 2.27 269.1 g 2.01Aves indeterminadas 10 2.16 4 2.27 10 4.02

que están asociados a la fractura intencional delhueso (Figura 2 y 5e). En este sitio se preservaronmetatarsianos con huellas de corte (11.5 por ciento)dispuestas transversalmente. Las ramas pélvicas,los maxilares, los radios, las costillas y las tibiascon evidencias de corte son escasas (tres porciento). Las ramas mandibulares presentan huellasde corte en la superficie externa, y sólo un maxilartiene huellas de raspado (Figura 2).
Las ulnas y radios se presentan fracturados haciala mitad de la diáfisis, tienen planos de fracturatransversales o ligeramente oblicuos sin eviden-cias de impacto (Figura 4) y son similares a los real-izados por flexión (Pérez Ripoll 2005–2006). Las
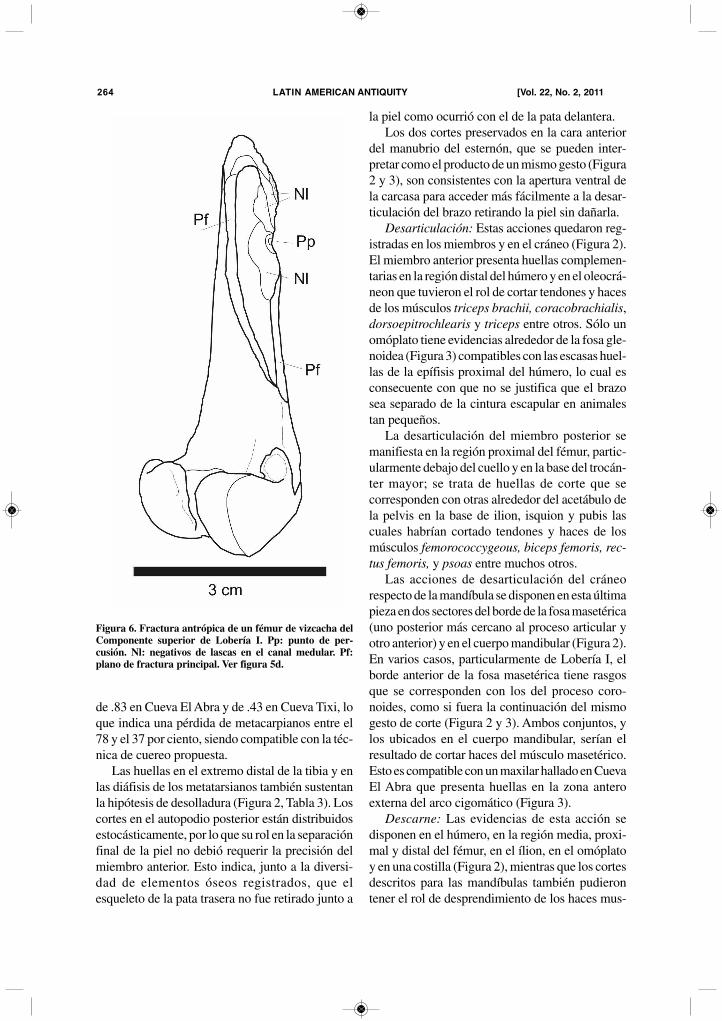
fracturas antrópicas del miembro posterior se dispo-nen en ocho tibias (tres epífisis distales con la diá-fisis, una diáfisis, una epífisis distal y tres epífisisproximales) y en ocho fémures (dos epífisis prox-imales y cuatro distales y dos diáfisis con la epífi-sis distal) (Figura 5). Las fracturas se caracterizanpor presentar planos de fractura lisos, ángulos obtu-sos y negativos de desprendimientos de lascas inter-nos o externos. Tres epífisis distales de húmeropresentan fracturas que parecen de origenantrópico. Una preforma de instrumento óseo fuerealizada sobre un fragmento de tibia (Mazzanti yValverde 2001).
Lobería I, Componente Superior (676 ± 41 AA-81060; 883 ± 41 AA-81059)
En este sitio la vizcacha está representada por casitodo el esqueleto (Tabla 2, Figura 2) con NISP yMNI altos (Tabla 1). Las ramas mandibulares sonlas piezas mejor preservadas proporcionalmenteseguidas por las ulnas enteras, clavículas, radios ypelvis (Tabla 2).
Se hallaron marcas de corte en casi todas laspartes del esqueleto excepto vértebras, costillas yautopodios, correspondiendo al 4.22 por ciento deltotal de los huesos recuperados de esta especie(Figura 2 y 3). Las ramas mandibulares fueron loshuesos más numerosos con registro de rasgos deprocesamiento (23.8 por ciento). Las trazas másconspicuas se disponen de modo oblicuo sobre lafosa masetérica, el proceso coronoides y en elcuerpo mandibular (Figura 2 y 3). Un esternón tienehuellas de corte profundas en su superficie ventral.Un húmero se caracteriza por presentar numerosastrazas oblicuas ubicadas a lo largo de casi toda laparte posterior de la diáfisis, similar al modo en quese presentan algunas marcas realizadas por roe-dores (Quintana 2007). Pero la morfología permiteasignarlas a trazas de filos líticos, posiblemente deraspado. Se trata de conjuntos de huellas aparente-mente realizadas con un filo delgado que generórasgos en forma de V, con estrías y terminacionesramificadas. Las ulnas y radios se caracterizan porhuellas cortas en la mitad de la diáfisis, mientrasque los fémures concentran evidencias de cortes enel sector proximal de la superficie anterior (Figura2 y 3).
Las fracturas del zeugopodio anterior son sim-ilares a las descritas en los mismos huesos de CuevaEl Abra (Figura 4). Se registraron fracturas inten-
258 LATIN AmerIcAN ANTIquITy [Vol. 22, No. 2, 2011
Tabla 2. Abundancia relativa de vizcacha pampeana(Lagostomus maximus) de los sitios del Holoceno Tardío
Final de Tandilia Oriental.
Cueva CuevaEl Abra Lobería I TixiMNI 12 MNI 13 MNI 6
Maxilar 24.99 26 8Rama mandibular 45.82 73 16Húmero entero 16.66 15 50Húmero proximal 37.49 - -Húmero diáfisis 4.16 7.69 -Húmero distal 41.66 19.23 16Ulna entero 12.49 57.69 25Ulna proximal 83.32 11.53 58Ulna diáfisis - - -Ulna distal - - -Radio entero - 7.69 16.6Radio proximal 29.16 42.3 75Radio diáfisis - - -Radio distal - - -Omóplato 62.49 26.92 16Fémur entero 8.33 19.23 33Fémur proximal 16.66 19.23 58Fémur diáfisis - - 8Fémur distal 24.99 19.23 41Tibia entero - - -Tibia proximal 33.32 19.23 25Tibia diáfisis 4.16 3.84 -Tibia distal 20.83 11.53 -Pelvis 33.32 46.15 25Calcáneo 62.49 23.07 41Astrágalo 41.66 30.76 58Costilla 14.1 10.35 21Vértebras 31.48 11.39 23Metatarsianos 58 47 44Metacarpianos 36 10 14Falanges 20.62 10.19 5Clavícula 4.16 50 25Esternón - 11.53 -Rótula - 3.8 -

quintana y mazzanti] LAS VIZcAcHAS PAmPeANAS eN LA SuBSISTeNcIA INDÍGeNA DeL HOLOceNO TArDÍO 259
Elemento
Húmero
Fémur
Ramamandibular
Ulna
Radio
Pelvis
Tibia
Esternón
Maxilar
Falange
Metapodio
Costilla
Rótula
Omóplato
Lobería 1
Anteriores: extremo distalde la diáfisis, transversales.Laterales a la tuberosidaddeltoidea, oblicuas.Posteriores: en casi toda lalongitud de la diáfisis.
Latero anterior, por debajodel cuello. Transversales.
Externas, base y borde de lafosa masetérica, procesocoronoides. Oblicuas.
Diáfisis, en posiciónexterna y medio proximal.Transversales.
Diáfisis, en posición anterior, medio proximal.Transversales.
Alrededor del cuello, sobreel ilion. Transversales yoblicuas.
Diáfisis, anterior. Transversal
Ventral, longitudinal.
-
-
-
-
-
-
Cueva El Abra
Transversales: latero anterior en epífisis dis-tal y diáfisis, anterior en proceso deltoideo,posterior en epífisis proximal.Longitudinales: latero anterior en epífisisproximal.
Principalmente transversales sobre la epífisis proximal y la diáfisis, en posiciónanterior y lateral. Posteriores en la epífisisdistal.
Externas en el borde de la fosa masetérica,en el proceso coronoides, en la rama ascen-dente, y en el diastema.
Todas externas del lado opuesto al radio,longitudinales sobre la base del oleocrá-neon, transversales en la parte más proximalde la diáfisis.
En la base de la cabeza, transversal, anterior.
Isquion: Transversales en posición superior.
Transversales en la epífisis distal, lateroanteriores.
-
En el proceso yugal del maxilar, exterior.
Todas en posición dorsal, transversales.
Todas en posición dorsal, transversales.
En la mitad del cuerpo, transversales.
-
-
Cueva Tixi
Proceso deltoideo: oblicuas ylargas.
Diáfisis: transversales.
Epífisis distal: transversales.
Diáfisis: en toda la longitud deambas áreas anterior y poste-rior.
Externas. Cuerpo mandibular:largas, oblicuas y profundas.Rama ascendente y borde dela fosa masetérica: cortas.
Oleocráneon y porción proxi-mal de la diáfisis, latero externas. Transversales y algunas oblicuas.
En la porción media de la diáfisis, oblicuas.
-
-
-
-
-
-
-
Cara anterior, transversal.
Superficie anterior: en elborde anterior y base de lafosa supraespinosa. Superficieposterior: borde axilar y bordeanterior.
Tabla 3. Ubicación de las huellas de corte en huesos de vizcacha pampeana (Lagostomus maximus) de los sitios del Holoceno Tardío Final de Tandilia Oriental.

cionales en seis tibias (una epífisis proximal con ladiáfisis, cuatro diáfisis y una epífisis distal) y entres fémures (una epífisis proximal y una distal yuna epífisis distal con la diáfisis) (Figura 5). En unatibia y en un fémur se identificó el punto de impactocon negativos de desprendimientos óseos internosen forma radial (Figura 5b y d).
Cueva Tixi, Nivel Arqueológico 4 (715 ± 45 a.P. AA-15809)
En este Nivel Arqueológico la vizcacha presentamenos restos y cantidad de individuos que en lasdos ocupaciones del Holoceno Tardío Finaldescritas anteriormente, aunque su diversidad óseaes importante (Tabla 2). La epífisis proximal deradio es la pieza mejor representada proporcional-
mente, seguida por las epífisis proximales de fémury ulna y por el astrágalo (Tabla 2). Como en losotros sitios, no se registraron tibias enteras ni diá-fisis o epífisis distales de ulnas.
La muestra ofrece una buena información desu procesamiento ya que el 7.78 por ciento de loshuesos tienen evidencias de trozamiento. Casitodos los huesos largos presentan huellas de corte,con patrones claros (Figura 2 y 3, Tabla 3), siendoel fémur y la ulna los más numerosos con huel-las de corte (33.33 por ciento). La rama mandibu-lar muestra una distribución de trazas en losmismos sectores que los otros sitios analizados,particularmente un corte profundo en el cuerpomandibular similar a una rama mandibular deLobería I (Figura 3f y j). En Cueva Tixi el reg-
260 LATIN AmerIcAN ANTIquITy [Vol. 22, No. 2, 2011
Figura 2. Distribución de las huellas de corte en el esqueleto de vizcacha pampeana (Lagostomus maximus) de los sitiosdel Holoceno tardío final de Tandilia Oriental. Cada silueta representa la sumatoria de todos los cortes hallados en elhueso respectivo. (a) radio; (b) ulna; (c-d) húmero; (e-f) fémur; (g) tibia; (h) autopodio posterior; (i) rama pélvica, (j)cráneo, (k) rama mandibular; (l) costilla; (m) esternón; (n-o) omóplato; (p) rótula.
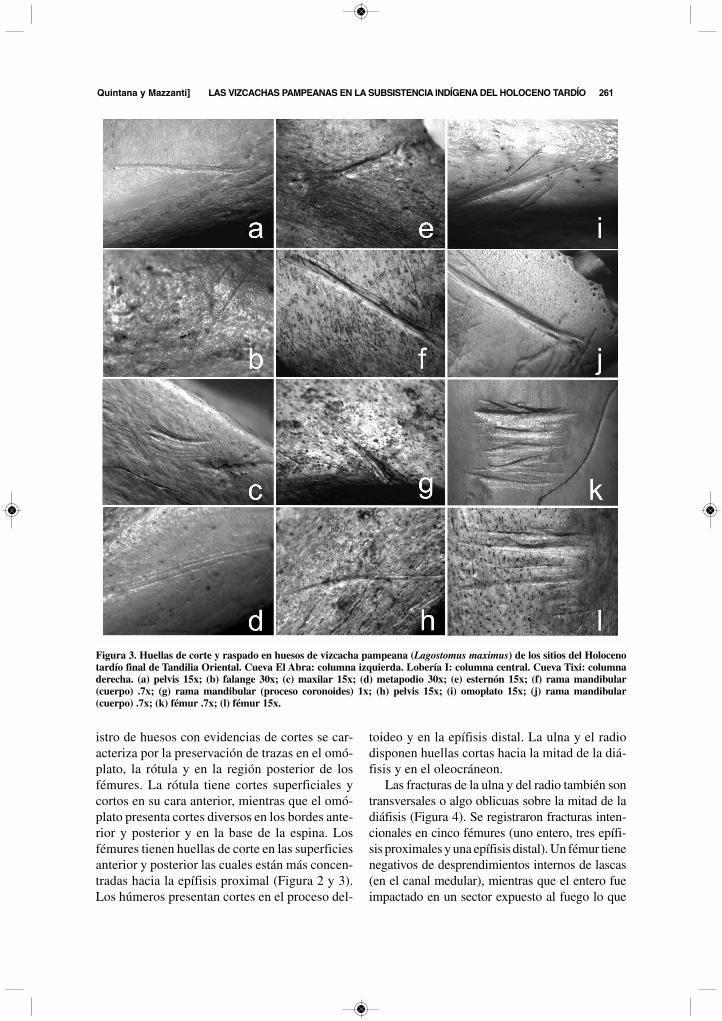
istro de huesos con evidencias de cortes se car-acteriza por la preservación de trazas en el omó-plato, la rótula y en la región posterior de losfémures. La rótula tiene cortes superficiales ycortos en su cara anterior, mientras que el omó-plato presenta cortes diversos en los bordes ante-rior y posterior y en la base de la espina. Losfémures tienen huellas de corte en las superficiesanterior y posterior las cuales están más concen-tradas hacia la epífisis proximal (Figura 2 y 3).Los húmeros presentan cortes en el proceso del-
toideo y en la epífisis distal. La ulna y el radiodisponen huellas cortas hacia la mitad de la diá-fisis y en el oleocráneon.
Las fracturas de la ulna y del radio también sontransversales o algo oblicuas sobre la mitad de ladiáfisis (Figura 4). Se registraron fracturas inten-cionales en cinco fémures (uno entero, tres epífi-sis proximales y una epífisis distal). Un fémur tienenegativos de desprendimientos internos de lascas(en el canal medular), mientras que el entero fueimpactado en un sector expuesto al fuego lo que
quintana y mazzanti] LAS VIZcAcHAS PAmPeANAS eN LA SuBSISTeNcIA INDÍGeNA DeL HOLOceNO TArDÍO 261
Figura 3. Huellas de corte y raspado en huesos de vizcacha pampeana (Lagostomus maximus) de los sitios del Holocenotardío final de Tandilia Oriental. Cueva El Abra: columna izquierda. Lobería I: columna central. Cueva Tixi: columnaderecha. (a) pelvis 15x; (b) falange 30x; (c) maxilar 15x; (d) metapodio 30x; (e) esternón 15x; (f) rama mandibular(cuerpo) .7x; (g) rama mandibular (proceso coronoides) 1x; (h) pelvis 15x; (i) omoplato 15x; (j) rama mandibular(cuerpo) .7x; (k) fémur .7x; (l) fémur 15x.

generó una fractura helicoidal que lo partió en doscon separación de lascas (Figura 5f).
Patrón de procesamiento de la vizcacha
Las huellas producidas por filos de instrumentoslíticos sobre huesos de vizcachas en estos tres sitiosmuestran cierta diversidad de morfologías. En gen-eral son de pequeña longitud y la mayoría sólo sonvisibles con lupa. En todos los casos se identificaronestrías internas. La diversidad de huellas identifi-cadas son:
1. superficiales y largas (corte), se presentan ais-ladas o en grupos de pocos rasgos subparalelos(Figura 3a b d h);
2. profundas y largas (corte) (en los cuerposmandibulares), se trata de varios cortes que sesuperponen y generan terminaciones ramificadas(Figura 3f j);
3. profundas, cortas y superpuestas (corte), pro-ducto de varios cortes sobre un mismo eje que for-man un gran conjunto con forma de V, también conterminaciones ramificadas (Figura 3e g);
4. profundas y superficiales, cortas y paralelas(corte), forman grupos similares a raspados perofueron generadas por varios gestos de corte conaplicación de fuerza variable, con terminacionesramificadas (Figura 3k l);
5. superficiales cortas y paralelas (raspado), pro-ducidas por un único gesto con el filo oblicuo, sinterminaciones ramificadas (Figura 3c).
Sobre la base de esta evidencia se plantea unpatrón de procesamiento para los tres sitios quecomprende: desollado, desarticulación, descarne yfractura del miembro anterior y posterior.Desollado: Los rasgos dispuestos en la región
media de la ulna y del radio se vinculan con laacción final de desprendimiento de la piel de losmiembros anteriores. La mayoría de los radios (87por ciento) y de las ulnas (86.2 por ciento) estánquebrados medialmente (Figura 4). Estas huellasy fracturas corresponden al gesto de separar la pieljunto al esqueleto de la mano y a la parte distal delzeugopodio (ulna y radio). Este comportamientoes frecuente para el procesamiento de animales
262 LATIN AmerIcAN ANTIquITy [Vol. 22, No. 2, 2011
Figura 4. Fracturas mediales del zeugopodio de vizcacha pampeana (Lagostomus maximus) de los sitios del Holocenotardío final de Tandilia Oriental. Fila superior: ulnas. Fila inferior: radios. (a, b, d, e) Cueva El Abra; (c, f-i) Lobería I;(j-l) Cueva Tixi.
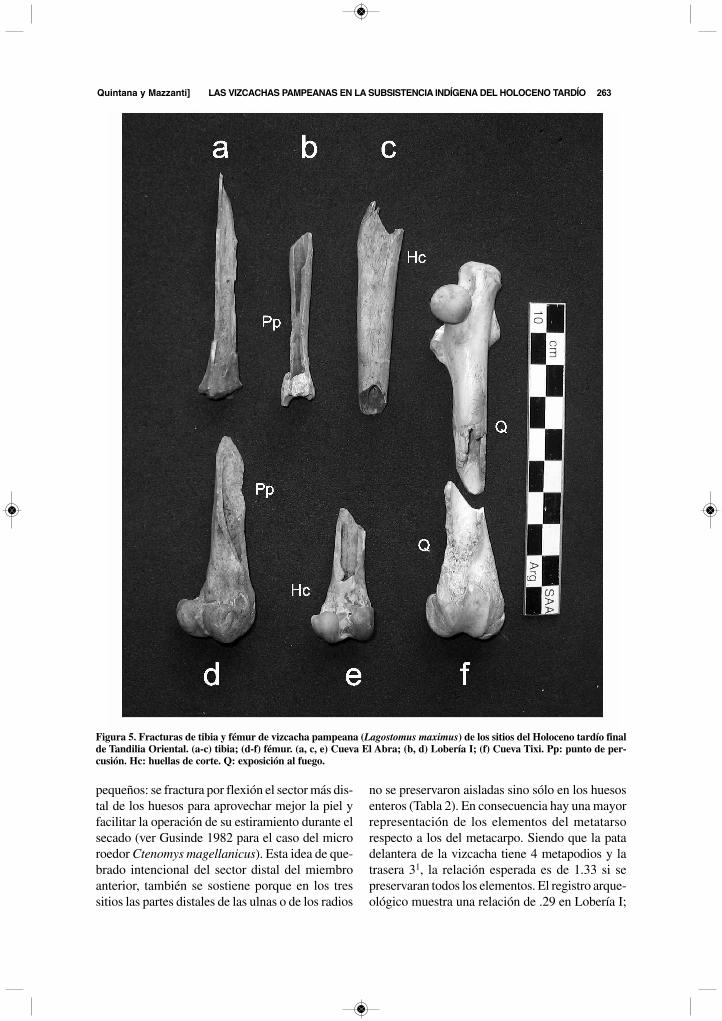
pequeños: se fractura por flexión el sector más dis-tal de los huesos para aprovechar mejor la piel yfacilitar la operación de su estiramiento durante elsecado (ver Gusinde 1982 para el caso del microroedor Ctenomys magellanicus). Esta idea de que-brado intencional del sector distal del miembroanterior, también se sostiene porque en los tressitios las partes distales de las ulnas o de los radios
no se preservaron aisladas sino sólo en los huesosenteros (Tabla 2). En consecuencia hay una mayorrepresentación de los elementos del metatarsorespecto a los del metacarpo. Siendo que la patadelantera de la vizcacha tiene 4 metapodios y latrasera 31, la relación esperada es de 1.33 si sepreservaran todos los elementos. El registro arque-ológico muestra una relación de .29 en Lobería I;
quintana y mazzanti] LAS VIZcAcHAS PAmPeANAS eN LA SuBSISTeNcIA INDÍGeNA DeL HOLOceNO TArDÍO 263
Figura 5. Fracturas de tibia y fémur de vizcacha pampeana (Lagostomus maximus) de los sitios del Holoceno tardío finalde Tandilia Oriental. (a-c) tibia; (d-f) fémur. (a, c, e) Cueva El Abra; (b, d) Lobería I; (f) Cueva Tixi. Pp: punto de per-cusión. Hc: huellas de corte. Q: exposición al fuego.
de .83 en Cueva El Abra y de .43 en Cueva Tixi, loque indica una pérdida de metacarpianos entre el78 y el 37 por ciento, siendo compatible con la téc-nica de cuereo propuesta.
Las huellas en el extremo distal de la tibia y enlas diáfisis de los metatarsianos también sustentanla hipótesis de desolladura (Figura 2, Tabla 3). Loscortes en el autopodio posterior están distribuidosestocásticamente, por lo que su rol en la separaciónfinal de la piel no debió requerir la precisión delmiembro anterior. Esto indica, junto a la diversi-dad de elementos óseos registrados, que elesqueleto de la pata trasera no fue retirado junto a
la piel como ocurrió con el de la pata delantera.Los dos cortes preservados en la cara anterior
del manubrio del esternón, que se pueden inter-pretar como el producto de un mismo gesto (Figura2 y 3), son consistentes con la apertura ventral dela carcasa para acceder más fácilmente a la desar-ticulación del brazo retirando la piel sin dañarla.Desarticulación: Estas acciones quedaron reg-
istradas en los miembros y en el cráneo (Figura 2).El miembro anterior presenta huellas complemen-tarias en la región distal del húmero y en el oleocrá-neon que tuvieron el rol de cortar tendones y hacesde los músculos triceps brachii, coracobrachialis,dorsoepitrochlearis y triceps entre otros. Sólo unomóplato tiene evidencias alrededor de la fosa gle-noidea (Figura 3) compatibles con las escasas huel-las de la epífisis proximal del húmero, lo cual esconsecuente con que no se justifica que el brazosea separado de la cintura escapular en animalestan pequeños.
La desarticulación del miembro posterior semanifiesta en la región proximal del fémur, partic-ularmente debajo del cuello y en la base del trocán-ter mayor; se trata de huellas de corte que secorresponden con otras alrededor del acetábulo dela pelvis en la base de ilion, isquion y pubis lascuales habrían cortado tendones y haces de losmúsculos femorococcygeous, biceps femoris, rec-tus femoris, y psoas entre muchos otros.
Las acciones de desarticulación del cráneorespecto de la mandíbula se disponen en esta últimapieza en dos sectores del borde de la fosa masetérica(uno posterior más cercano al proceso articular yotro anterior) y en el cuerpo mandibular (Figura 2).En varios casos, particularmente de Lobería I, elborde anterior de la fosa masetérica tiene rasgosque se corresponden con los del proceso coro-noides, como si fuera la continuación del mismogesto de corte (Figura 2 y 3). Ambos conjuntos, ylos ubicados en el cuerpo mandibular, serían elresultado de cortar haces del músculo masetérico.Esto es compatible con un maxilar hallado en CuevaEl Abra que presenta huellas en la zona anteroexterna del arco cigomático (Figura 3).Descarne: Las evidencias de esta acción se
disponen en el húmero, en la región media, proxi-mal y distal del fémur, en el ílion, en el omóplatoy en una costilla (Figura 2), mientras que los cortesdescritos para las mandíbulas también pudierontener el rol de desprendimiento de los haces mus-
264 LATIN AmerIcAN ANTIquITy [Vol. 22, No. 2, 2011
Figura 6. Fractura antrópica de un fémur de vizcacha delComponente superior de Lobería I. Pp: punto de per-cusión. Nl: negativos de lascas en el canal medular. Pf:plano de fractura principal. Ver figura 5d.

culares. El fémur tiene huellas de corte en la epífi-sis distal tanto anterior como posteriormente, vin-culables con la desarticulación, sin embargo secarece de rasgos compatibles en las epífisis prox-imales de las tibias lo que sugiere que esas huellastuvieron la función de retirar músculos. La ausen-cia de cortes en los laterales de la epífisis distal delfémur y proximal de la tibia, que serían el productode cortar tendones, sustenta la función de descarnede las huellas citadas.Fractura: El fémur y la tibia son las piezas con
mayor masa de médula y las que sufrieron fracturashelicoidales (Figura 4 y 5), ninguna tibia se recu-peró entera (Tabla 2). Las fracturas presentan ori-entación oblicua o casi paralela al eje longitudinaldel hueso (Figura 5). En algunos ejemplares estánasociadas a huellas de corte, pero ninguna que hayacumplido el rol de aserrado. La fractura que gen-eró cilindros cortando el hueso en los extremos espreponderante seguida de las dispuestas al mediode la diáfisis. En algunos casos se pudo identificarel lugar del impacto y negativos de lascas. Los pun-tos de impacto se presentan a modo de una muescaen el borde de fractura que antecededesprendimientos de lascas óseas en la superficiede la cavidad medular (Figura 5 y 6). Otros nega-tivos de desprendimientos de lascas son máspequeños y dispuestos en los bordes externos defractura. Las fracturas del miembro anterior (Figura4) tuvieron dos características: las ya citadas que-braduras (posiblemente por flexión) transversalesy oblicuas de los elementos del zeugopodio vin-culadas al desollado, y fracturas aparentemente porimpacto de tres húmeros de Cueva El Abra queposiblemente sean atribuibles a la acción humanapero se requiere de casos más claros para confir-mar este procedimiento. En ningún sitio se identi-ficaron fracturas atribuibles a mordeduras humanas,no se hallaron estructuras tales como muescas oca-sionadas por presión, muescas contrapuestas o mar-cas transversales de sección chata como lasdescritas para conejos (Pérez Ripoll 2005_2006;Sanchis y Fernández 2008).
Estos sitios del Holoceno más tardío presentanalgunas particularidades en la distribución de lasevidencias del procesamiento de la vizcacha(Figura 2, Tabla 3). Lobería I tiene un registro másintenso del despiece de la pelvis y es el único conhuellas en el esternón. Sólo en las vizcachas deCueva El Abra hay rasgos en el autopodio poste-
rior, en un maxilar y en una costilla, mientras queCueva Tixi se caracteriza por las huellas preser-vadas en el omóplato, en la rótula y en la regiónposterior de los fémures. De todos modos estasdiferencias no indican procesamientos distintospara cada uno de los sitios, sino que refuerzan lasactividades propuestas a través de las recurrencias,por lo que se las interpreta como parte de la diver-sidad propia de la preservación y de la habilidad ytécnica del operador que realizó el despiece.
Discusión
La vizcacha tiene un registro fósil amplio en sitiosarqueológicos de diversas áreas de la región Pam-peana desde el Pleistoceno Tardío (Martínez yGutiérrez 2004), sin embargo sólo se describieronevidencias de su consumo en el área Interserrana.Particularmente en Paso Otero 3 (5000 a.P.) el análi-sis de la representación esquelética demuestra queesta especie formó parte de la subsistencia indígena(Martínez et al. 2001) y en Laguna Tres Reyes Sitio1 (1800 a 2500 a.P.) se citaron restos de vizcachacon huellas de corte (Salemme y Madrid 2007).
En las sierras de Tandilia Oriental la secuenciaarqueológica contiene evidencias que indican quela vizcacha pampeana fue incorporada a la subsis-tencia de modos diversos desde el Pleistocenotardío hasta tiempos de contacto hispano indígena.Los grupos cazadores recolectores de la parte finaldel Holoceno Tardío preconquista (1000 a 600 a.P.)fueron los que aprovecharon a este roedor de modosistemático e integral. Durante este lapso la preser-vación de las partes esqueléticas revela que los ani-males fueron ingresados enteros a los sitios,mientras que el análisis de las superficies corticalespermitieron conocer una secuencia de proce-samiento que comprende desollado, desarticu-lación, descarne y fractura de los miembros. Laevidencia descrita indica que la secuencia se com-pondría de tres fases principales: 1- el desollado yfractura del miembro anterior; 2- la desarticulacióny el descarne; y 3- la fractura del miembro poste-rior.
La ubicación anatómica de las huellas de corteque indican el descarne en algunos casos se puedeninterpretar también como de desarticulación. Eneste contexto, el descarne se habría aplicado duranteel consumo, en lugar de una acción previa de reti-rar la carne para el secado o para algún proceso más
quintana y mazzanti] LAS VIZcAcHAS PAmPeANAS eN LA SuBSISTeNcIA INDÍGeNA DeL HOLOceNO TArDÍO 265

complejo. La talla pequeña de estas presas permiteque las acciones de reducción de la carcasa seanmenos restringidas que en los animales grandes. Deeste modo, el trozamiento presenta alternativas queno están necesariamente condicionadas por lasarticulaciones ni por el uso de artefactos cortantes(Quintana 2005). En consecuencia, en la colecciónanalizada no es posible establecer el orden de ocur-rencia entre la desarticulación y el descarne, porello se plantea la posibilidad de que la aplicaciónde filos líticos para la desarticulación y el descarnehayan sido alternados durante el despiece y el con-sumo.
La fractura intencional de los huesos largos delmiembro posterior y anterior de la vizcacha es unhallazgo novedoso en el procesamiento de especiespequeñas de la región Pampeana. Entre las especiespequeñas explotadas durante este período (Tabla2) la vizcacha y el lagarto son las únicas con hue-sos fracturados, sin embargo esta acción sólo fuesistemática en la vizcacha. La fractura de huesosde especies pequeñas no es frecuente, aunque sehan registrado otros casos en diversas partes delmundo de animales de masas similares o menoresa las vizcachas como las liebres africanas (Yellen1991), los lagomorfos de Estados Unidos y Europa(Hockett 1994; Hockett y Ferreira 2000; PérezRipoll 2005_2006), las maras del Chaco argentino(del Papa et al. 2010) y las más pequeñas vizcachasandinas (Labarca 2005). La fragilidad de los hue-sos de las presas pequeñas también permite unamayor diversidad de acciones para su fracturacomo flexión, percusión y mordido (Martínez2009; Pérez Ripoll 2005_2006; Sanchis y Fer-nández 2008) de las cuales a las vizcachas sólo leaplicaron las dos primeras, posiblemente por sumayor robustez respecto de huesos de especiescomo conejos o monos. Además, estos tipos defracturas fueron realizadas sobre huesos distintosdel esqueleto con la intencionalidad de cumplirobjetivos específicos: separar el extremo del miem-bro anterior junto a la piel o acceder a la méduladel miembro posterior.
Luego de diez milenios en los que predominóla caza mayor centralizada sobre ungulados (gua-naco y cérvidos), en la región Pampeana se regis-tra un cambio de los modos de vida durante elHoloceno Tardío Final. Este cambio de lassociedades cazadoras-recolectoras consistió en elestablecimiento de estrategias de intensificación
del uso del espacio que involucró una gran diver-sificación en la explotación de las presas pequeñasy el desarrollo de nuevas tecnologías (arco y flecha,alfarería, hilado, etc.). Estos procesos fueron reg-istrados en las sierras de Tandilia, en la Depresióndel Río Salado, en el Área Norte de la provincia deBuenos Aires y, hacia el sur, en la cuenca inferiordel Río Colorado (norte de Patagonia) (González2005; Martínez y Gutiérrez 2004; Martínez et al2005; Quintana y Mazzanti 2001; Quintana 2009)(Figura 1).
En las sierras de Tandilia la caza de presaspequeñas se comporta como un emergente sensi-ble a este nuevo modo de vida (Quintana et al.2002). La relevancia de la caza menor se expresaen la incorporación de nuevas especies a la sub-sistencia que anteriormente estaban disponibles enel paisaje pero que no fueron explotadas (lagar-tos, peces, aves pequeñas, cuises) y en que fueronobtenidas en mayor número las que ya eran usadaspara consumo (vizcacha, nutria, armadillos,huevos de ñandú) (Quintana 2009; Quintana yMazzanti 2001; Quintana et al. 2002). El aumentode la cantidad de ejemplares y de la diversidad depresas pequeñas durante el Holoceno Tardío Finalestá acompañado por el desarrollo de proce-samientos más exhaustivos de sus carcasas, siendopropio de los procesos de intensificación y dediversificación en Tandilia Oriental (Quintana2009). Durante este período el lagarto, los cuisesy la vizcacha son las especies que tienen eviden-cias más claras de procedimientos complejos queinvolucraron acciones de desollado, desmem-bramiento y descarne. Esto las diferencia deltratamiento menos esmerado de otras presaspequeñas como armadillos, aves, peces y coyposregistrados en los mismos sitios (Quintana y Maz-zanti 2001; Quintana 2009). En este espectro dela caza menor las nuevas evidencias indican quela vizcacha es la especie que presenta un proce-samiento sistemático entre los tres sitios y apli-cado de modo exhaustivo. Además de la cantidadde individuos registrados, este aspecto es desta-cado ya que el despiece de la vizcacha estuvo ori-entado hacia la explotación máxima de susrecursos: se aprovechó la carne, la médula y lapiel, mientras que los huesos fueron usados en latecnología y fracturados sistemáticamente paraexponer la médula o para aprovechar la piel.Durante los milenios previos del registro de
266 LATIN AmerIcAN ANTIquITy [Vol. 22, No. 2, 2011

Tandilia Oriental ninguna presa pequeña presentaevidencias de procesamientos tan completos comolas vizcachas del Holoceno Tardío Final (Quin-tana y Mazzanti 2001). A diferencia de otrasespecies pequeñas que fueron preponderantes enla subsistencia como los coypos de los sitios delRío Salado, las vizcachas de este período tardíodesempeñaron un rol destacado en el contexto dela caza menor, siendo esta categoría, compuestapor numerosas especies, la que caracteriza laeconomía de los últimos cazadores recolectoresserranos (Quintana 2009).
La diversidad geomorfológica de estas sierraspresenta micro ambientes que generan discon-tinuidades en la distribución de algunas poblacionesanimales (Goin 2001; Quintana 2009). A pesar deello la vizcacha es una especie que habita en sec-tores distintos del paisaje como lomas, llanuras, cer-canías de peñones rocosos y mesetas. Además, lasmadrigueras subterráneas no son estáticas, sino queocurren migraciones, fusiones o generación decolonias nuevas (Branch et al. 1993). Esta dinámicay distribución espacial facilitó su explotación sis-temática en los tres sitios analizados.
En las ocupaciones tardías de Cueva El Abra yde Cueva Tixi el espectro de presas pequeñas esmás amplio que en Lobería I porque fueron incor-poradas especies vinculadas a cuerpos de aguacomo los coypos, los peces y las aves acuáticas(Tabla 1) (Quintana 2009). En Lobería I la viz-cacha fue capturada con mayor interés en términosrelativos del MNI, lo que estaría indicando un reem-plazo del aprovechamiento de las especies lacus-tres, las cuales eran de obtención difícil en estesector del paisaje por estar alejado de cuerpos deagua importantes. La disponibilidad de este roedortambién está influenciada por su comportamientoestacional ya que durante el invierno aumenta eltiempo de actividad fuera de las madrigueras lo cualestá vinculado al ciclo reproductivo (Branch 1993)facilitando las oportunidades de captura. Si LoberíaI fue ocupado principalmente durante los mesesfríos es concordante con el alto número mínimo deindividuos de vizcacha registrados en este sitio ycon la escasez de lagartos y de huevos de ñandú,comparado con los otros dos sitios (Tabla 1), ya queestos reptiles hibernan entre los meses de abril aseptiembre (Duarte y Cabrera 2000) lo cual coin-cide con el período en que no hay huevos de ñandúdisponibles.
Conclusiones
La vizcacha pampeana (Lagostomus maximus) esun roedor que fue incorporado a la subsistenciaindígena durante 11 milenios en las sierras deTandilia Oriental. A partir de los 1000 a.P. (Holo-ceno Tardío Final) las sociedades cazadorasrecolectoras realizaron un mayor uso de esterecurso, lo que comprendió la captura de más can-tidad de ejemplares y la aplicación de acciones dedespiece orientadas al aprovechamiento integraldel cuerpo (piel, huesos, médula y carne). Durantetiempos posconquista la vizcacha pasó nuevamentea ser un recurso ocasional.
Para la vizcacha pampeana se identificaron lassiguientes acciones de procesamiento durante elHoloceno Tardío Final de Tandilia Oriental:
—desollado con apertura ventral de la piel,siendo seccionada en los autopodios posteriores yen la cabeza para desprenderla del esqueleto. Lapiel mantuvo el esqueleto de los autopodios ante-riores y de parte de la ulna y del radio (por frac-tura), pero no el esqueleto de los miembrosposteriores.
—desarticulación del cráneo y de la mandíbula,del húmero respecto de la ulna y del miembro pos-terior respecto de la cintura pélvica;
—descarne de los músculos de la cinturapélvica, del tórax, de la cabeza y de ambos miem-bros;
—fractura del miembro posterior (y posible-mente del húmero) para acceder a la médula oexponerla durante el hervido y para generar mate-ria prima para la tecnología ósea.
La secuencia de esas acciones fueron: 1, desol-lado y fractura el miembro anterior; 2, desarticu-lación y descarne; y finalmente 3, fractura delmiembro posterior.
La distribución amplia de la vizcacha, sudinámica poblacional y su agregación enmadrigueras subterráneas favoreció su capturadurante todo el año. Su mayor disponibilidad esta-cional coincidente con la disminución de otrosrecursos y su distribución homogénea en el paisajepermitió que la vizcacha fuese aprovechada comoun recurso de carácter amortiguador durante laescasez o dificultad de obtención de otras especiespequeñas. Estas propiedades aumentan su pre-dictibilidad en el paisaje y su posibilidad de encuen-tro, lo cual es consistente con el registro
quintana y mazzanti] LAS VIZcAcHAS PAmPeANAS eN LA SuBSISTeNcIA INDÍGeNA DeL HOLOceNO TArDÍO 267

arqueológico que muestra el uso extendido de estaespecie en momentos de intensificación de la sub-sistencia.
Agradecimientos. A Luis del Papa, Matías Medina yFernando Fernández por permitirme acceder a informacióninédita. A los evaluadores anónimos por sus valiosas sug-erencias. Estas investigaciones son parte de proyectos finan-ciados por la Secretaría de Investigación de la UniversidadNacional de Mar del Plata y por el PICTO 2004 proyecto552 de la Agencia Nacional de Promoción Científica yTecnológica.
Referencias Citadas
Abe, Yoshiko, Curtis Marean, Peter Nilssen, Zelalem Assefa, yElizabeth Stone
2002 The Analysis of Cutmarks on Archaeofauna: A Reviewand Critique of Quantification Procedures and a NewImage-Analysis GIS Approach. American Antiquity67:643_664.
Andrews, Peter1990 Owls, Caves, and Fossils. University of Chicago Press.
Chicago.Branch, Lyn
1993 Seasonal Patterns of Activity and Body Mass in thePlains Vizcacha, Lagostomus maximus (family Chin-chillidae). Canadian Journal of Zoology 71:1041–1045.
Branch, Lyn, Diego Villareal, y Gene Fowler1993 Recruitment, Dispersal, and Group Fusion in a Declin-
ing Population of the Plains Vizcacha (Lagostomus max-imus: chinchillidae). Journal of Mammalogy 74(1):9–20.
Bunn, Henry1981 Archaeological Evidence for Meat-Eating by Plio-
Pleistocene Hominids from Koobi Fora and OlduvaiGorge. Nature 291:574–577.
1983 Comparative Analysis of Modern Bone Assemblagesfrom a San Hunter-Gatherer Camp in the Kalahari Desert,Botswana, and from a Spotted Hyaena Den Near Nairobi,Kenya. En Animals and Archaeology 1. Hunters and theirPrey, editado por Juliet Clutton-Brock y Caroline Grig-son, pp 143–148. BAR International Series 163. BritishArchaeological Reports, Oxford.
del Papa, Luis, Luciano De Santis, y José Togo2010 Consumo de roedores en el sitio Villa La Punta, agro-
alfarero temprano de la región Chaco-Santiagueña. Inter-secciones en Antropología, en prensa.
Duarte, Christian, y Mario Cabrera 2000 Testing Skeletochronology in Black Tegu Lizards
(Tupinambis merianae) of Known Ages. HerpetologicalReview 31(4):224–226.
Escosteguy, Paula2007 Estudios sobre restos de Myocastor coypus en sitios
de la cuenca inferior del río Salado. En Arqueología en lasPampas, tomo II, editado por Cristina Bayón, AlejandraPupio, Isabel González, Nora Flegenheimer y MagdalenaFrère, pp. 471–487. Sociedad Argentina de Antropología,Buenos Aires.
Fernández, Fernando, Germán Moreira, Gustavo Neme, yLuciano De Santis
2009 Microvertebrados exhumados del sitio arqueológico“Cueva Arroyo Colorado” (Mendoza, Argentina): aspec-tos tafonómicos y significación paleoambiental. Archae-ofauna, en prensa.
Goin, Francisco2001 Marsupiales (Didelphidae: Marmosinae y Didelphi-
nae). En: Cueva Tixi: cazadores y recolectores de las sier-ras de Tandilia oriental. I. Geología, Paleontología yZooarqueología. Editado por Diana Mazzanti y CarlosQuintana, pp. 75–114. Publicación Especial 1. ARBO-UNMDP, Mar del Plata.
González, Isabel1997 Potsherds, “coypo” Teeth, and Fish Bones: Hunter-
Gatherer-Fishers in the Río Salado (Pampa Region,Argentina). editado por J. Rabassa y M. Salemme. Qua-ternary of South America and Antarctic Peninsula10:255–278.
2005 Arqueología de alfareros, cazadores y pescadorespampeanos. Sociedad Argentina de Antropología. Colec-ción Tesis Doctorales. Buenos Aires.
Gusinde, Martín1982 Los Indios de Tierra del Fuego. Segunda Parte. Cen-
tro Editor de Etnología Americana, Buenos Aires.Hockett, Bryan
1994 A Descriptive Reanalysis of the Leporid Bones fromHogup Cave, Utah. Journal of California and Great BasinAnthropology 16:106–117.
Hockett, Bryan, y Nuno Ferreyra 2000 The Rabbits of Picareiro Cave: Small Mammal Hunt-
ing During the Late Upper Palaeolithic in the PortugueseEstremadura. Journal of Archaeological Science27:715–723.
Jackson, John, Lyn Branch, y Diego Villareal1996 Lagostomus maximus. Mammalian Species 543:1–6.
Johnson, Eileen 1985 Current Developments in Bone Technology. EnAdvances in Archaeological Method and Theory, editadopor Michael B. Schiffer, pp. 157–235. Academic Press,Orlando.
Labarca, Rafael 2005 Las vizcachas de Piuquenes: evidencia de explotación
de roedores durante la transición Pleistoceno–Holocenoen la cordillera andina de Chile central. Werken 6:63–80.
Llanos, Augusto, y Jorge Crespo1952 Ecología de la vizcacha (Lagostomus maximus max-imus) en el nordeste de la provincia de Entre Ríos. Revistade Investigaciones Agrícolas 6:289–378.
Lyman, R. Lee1994 Vertebrate Taphonomy. Cambridge University Press.
Cambridge.Martínez, Gustavo
2009 Human Chewing Bone Surface Modification and Pro-cessing of Small and Medium Prey Amongst the Nukak(Foragers of Colombian Amazon). Journal of Taphonomy7(1):1–20.
Martínez, Gustavo, y María Gutiérrez 2004 Tendencias en la explotación humana de la fauna
durante el Pleistoceno final y Holoceno en la Región Pam-peana (Argentina). En Zooarchaeology of South America,editado por Guillermo Mengoni Goñalons, pp 81–98. BARInternacional Series 1298. British Archaeological Reports,Oxford.
Martínez, Gustavo, Pablo Messineo, María Piñeyro, CristianKaufmann, y María Barros
2001 Análisis Preliminar de la Estructura Faunística delSitio Paso Otero 3 (Área Interserrana, Región Pampeana,Argentina). Arqueología Uruguaya hacia el fin del Mile-nio 1:505–520.
Martínez, Gustavo, Francisco Zangrando, y Luciana Stoessel 2005 Sitio El Tigre (pdo. de Patagones, pcia. de Buenos
Aires, Argentina): evidencias sobre la explotación de peces
268 LATIN AmerIcAN ANTIquITy [Vol. 22, No. 2, 2011

en el curso inferior del Río Colorado e implicaciones paralos sistemas de subsistencia. Magallania 33(2):99–114.
Mazzanti, Diana1997 Excavaciones arqueológicas en el sitio Cueva Tixi,
Buenos Aires, Argentina. Latin American Antiquity8:55–62.
1999 Secuencia arqueológica del sitio Cueva El Abra,Tandilia Oriental. Provincia de Buenos Aires. Actas del XIIICongreso Nacional de Arqueología Argentina, p. 365. Cór-doba.
2007 Arqueología de las relaciones interétnicas poscon-quista en las sierras de Tandilia. Tesis doctoral inédita.Facultad de Filosofía y Letras, Universidad de BuenosAires.
Mazzanti, Diana y Federico Valverde 2001 Artefactos sobre hueso, asta y valva. En Cueva Tixi:cazadores y recolectores de las sierras de Tandilia orien-tal. I. Geología, Paleontología y Zooarqueología, editadopor Diana Mazzanti y Carlos Quintana, pp. 157–180. Pub-licación Especial 1. ARBO-UNMDP, Mar del Plata.
Mazzanti, D., M. Colobig, A. Zucol, G. Martínez, J. PortoLópez, M. Brea, E. Passeggi, J. Soria, C. Quintana y V.Puente
2009 Investigaciones arqueológicas en el Sitio 1 de la local-idad Lobería I. Actas V Congreso de Arqueología de laRegión Pampeana Argentina, en prensa
Medina, Matías2008 Diversificación económica y uso del espacio en el
tardío prehispánico del Norte del valle de Punilla, Pampade Olaen y Llanura Noroccidental (Córdoba, Argentina).Tesis de Doctorado inédita. Facultad de Filosofía y Letras,Universidad de Buenos Aires, Buenos Aires.
Pérez Ripoll, Manuel 1992 Marcas de carnicería, fracturas intencionadas ymordeduras de carnívoros en huesos prehistóricos delMediterráneo Español. Colección Patrimonio, Instituto deCultura José Gil Alberti, Alicante.
2005–2006 Caracterización de las fracturas antrópicas ysus tipologías en huesos de conejo procedentes de los nive-les gravetienses de la Cova de les Cendres (Alicante).Munibe 57(1): 239–254.
Potts, Richard, y Pat Shipman1981 Cutmarks Made by Stone Tools on Bones from Oldu-
vai Gorge, Tanzania. Nature 291:577–580.Quintana, Carlos
2005 Despiece de micro roedores en el Holoceno Tardío delas sierras de Tandilia, Argentina. Archaeofauna217:227–241.
2007 Marcas de dientes de roedores en huesos de sitiosarqueológicos de las sierras de Tandilia, Argentina. Archae-ofauna 16:185–191.
2009 La subsistencia de los últimos cazadores-recolectoresserranos. En cazadores y recolectores de las sierras deTandilia oriental, editado por Diana Mazzanti. PublicaciónEspecial 3. ARBO-UNMDP. Mar del Plata, en prensa.
Quintana, Carlos, y Diana Mazzanti 2001 Selección y aprovechamiento de recursos faunísticos.
En Cueva Tixi: cazadores y recolectores de las sierras deTandilia oriental. I. Geología, Paleontología y Zooarque-ología, editado por Diana Mazzanti y Carlos Quintana, pp.181–209. Publicación Especial 1. ARBO-UNMDP, Mardel Plata.
Quintana, Carlos, Federico Valverde y Diana Mazzanti 2002 Roedores y lagartos como emergentes de la diversi-
ficación de la subsistencia durante el Holoceno de las sier-ras de Tandilia, Argentina. Latin American Antiquity
13:455–473.Quintana, Carlos, Federico Valverde y Adriana Albino
2003 Registro de fauna del sitio Cueva El Abra, TandiliaOriental, Provincia de Buenos Aires. Actas XIII CongresoNacional de Arqueología Argentina 3:317–324. Cordoba.
Salemme, Mónica y Patricia Madrid2007 The Archaeofaunas from Laguna Tres Reyes 1 Site:
Taxonomic Richness and Abundance during the Beginningof the Late Holocene in the South-East Pampean Region(Argentina). En Taphonomy and Zooarchaeology inArgentina, editado por María Gutiérrez, Gustavo Barrien-tos, Guillermo Mengoni Goñalons, Laura Miotti y MónicaSalemme, pp. 121–143. BAR International Series 1601,British Archaeological Reports, Oxford.
Sanchis Sierra, Alfred, y Josep Fernández Peris 2008 Procesado y consumo antrópico de conejo en la Cova
del Bolomor (Tavernes de la Valldigna, Valencia): El NivelXVIIc (ca 350 ka). Complutum 19(1):25–46.
Santiago, Fernando2004 Los roedores en el “menú” de los habitantes de Cerro
Aguará (provincia de Santa Fe): su análisis arqueofaunís-tico. Intersecciones en Antropología 5:3–18.
Shipman, Pat, y Jennie Rose1983 Evidence of Butchery and Hominid Activities at Tor-
ralba and Ambrona: An Evaluation Using MicroscopicTechniques. Journal of Archaeological Science10:465–474.
Spotorno, Angel E., Carlos A. Zuleta, y J. Pablo Valladares 2004 Chinchilla laniger. Mammalian Species 758:1–9.
Stahl, Peter1996 The Recovery and Interpretation of Microvertebrate
Bone Assemblages from Archaeological Contexts. Jour-nal of Archaeological Method and Theory 3:31–75.
Stoessel, Luciana2007 Análisis arqueofaunísticos de los sitios Loma Ruiz 1
y El Tigre (partidos de Villarino y Patagones, provincia deBuenos Aires): Aportes para el conocimiento de la sub-sistencia en el valle inferior del río Colorado durante elHoloceno tardío. Intersecciones en Antropología8:235–251.
Valverde, Federico2001 Huellas y marcas en restos óseos. En Cueva Tixi:cazadores y recolectores de las sierras de Tandilia orien-tal. I. Geología, Paleontología y Zooarqueología, editadopor Diana Mazzanti y Carlos Quintana, pp. 137–156. Pub-licación Especial 1. ARBO-UNMDP, Mar del Plata.
Villa, Paola, y Erik Mahieu 1991 Breakage Patterns of Human Long Bones. Journal ofHuman Evolution 21(1):27–48.
Vucetich, Guimar, y Diego Verzi 1995 Los roedores caviomorfos. En Evolución biológica yclimática de la región Pampeana durante los últimos cincomillones de años. Un ensayo de correlación con el Mediter-ráneo occidental, editado por María Alberdi, GabrielloLeone y Eduardo Tonni, pp. 211–226. Monografía 12.Museo de Ciencias Naturales de Madrid Madrid.
Werner, Florian, Karim Ledesma, y Rodrigo Hidalgo2006 Mountain Vizcacha (Lagidium cf. peruanum) in
Ecuador - First Record of Chinchillidae from the North-ern Andes, Mendoza. Mastozoología Neotropical13(2):271–274.
Yellen, John1991 Small Mammals: Post-Discard Patterning of !Kung
San Faunal Remains. Journal of Anthropological Archae-ology 10:152–192.
quintana y mazzanti] LAS VIZcAcHAS PAmPeANAS eN LA SuBSISTeNcIA INDÍGeNA DeL HOLOceNO TArDÍO 269

Nota
1. En sentido estricto las vizcachas tienen 5 metatarsianosy tres dedos, pero los metatarsianos correspondientes alprimer y quinto dígito se encuentran extremadamente reduci-dos adquiriendo la forma de láminas muy delgadas. Estasláminas no suelen ser recuperadas en los sitios, ya sea por sudestrucción posdepositacional o por su dificultad dereconocimiento ya que pueden ser confundidos con astillasdiminutas. Así, la cuantificación de metatarsianos se realizasobre los tres bien desarrollados y claramente identificables yrecuperados arqueológicamente que son los correspondientesal dígito 2 al 4.
270 LATIN AmerIcAN ANTIquITy [Vol. 22, No. 2, 2011





























