Embed Size (px)
Citation preview

7.32/7.81J/8.591J
Systems BiologySystems Biology‘modeling biological networks’‘modeling biological networks’
1
IntroducingIntroducing ...
L t R it tiLectures: Recitations:
TR 1:00 -2:30 PM W 4:00 - 5:00 PMRm. 6-120 Rm. 26-204
(under construction)
Alexander van Oudenaarden Jialing LiRm. 68-371B [email protected]@mit edu Rm 68 [email protected] Rm. 68-383
Bernardo Pando
2
[email protected]. 68-365
Take-home Exams:
Exam 1 due 10/06/09 15 %Exam 1 due 10/06/09 15 %Exam 2 due 10/20/09 15 %Exam 3 due 11/03/09 15 %E 4 d 11/17/09 15 %Exam 4 due 11/17/09 15 %Exam 5 due 12/01/09 15 %Final due 12/10/09 25 %
Final Problem Set will be a take-home PSbroadly covering the course material
3
Text books: noneText books: none
Handouts will be available on-line
Good reference (biology textbook):Molecular biology of the cellAlberts et al.
Matlab will be used intensively during theMatlab will be used intensively during thecourse, make sure you known (or learn) howto use it (necessary for problem sets)
Web-page: web.mit.edu/biophysics/sbio(contains all course materials)
4

Intrinsic challenge of this class:Intrinsic challenge of this class:
mixed audience with wildly different backgrounds
⇒ read up on your biology or math if neededp y gy
⇒ recitations (W 4PM, Rm. 26-204) are intended to close the gaps and prepare for homeworkclose the gaps and prepare for homework
5
Systems Biology ?Systems Biology ?
6
Systems Biology ≈ Network Biology
GOAL: develop a quantitative (mathematical) understanding of thebiological function of genetic and biochemical networks
INPUT
gene A gene B
INPUT
gene C gene D
gene E gene F OUTPUT
- function of gene product A-F can be known in detail but this is notfunction of gene product A F can be known in detail but this is notsufficient to reveal the biological function of the INPUT-OUTPUT relation
- a system approach (looking beyond one gene/protein) is necessary toreveal the biological function of this whole network
7
g- what is the function of the individual interactions (feedbacks and
feedforwards) in the context of the entire network ?
Three levels of complexity
I Systems Microbiology (~14 Lectures)
‘The cell as a well-stirred biochemical reactor’
II Systems Cell Biology (~8 Lectures)II Systems Cell Biology ( 8 Lectures)
‘The cell as a compartmentalized system withconcentration gradients’concentration gradients
III Systems Developmental Biology (~3 Lectures)
‘The cell in a social context communicating withneighboring cells’
8
neighboring cells

I Systems Microbiology (14 Lectures)
‘The cell as a well-stirred biochemical reactor’
L1 IntroductionL2-3 Chemical kinetics, Equilibrium binding, cooperativityL4 Lambda phagep gL5 Cellular memoryL6-7 Genetic switchesL8-9 E. coli chemotaxisL10 Stability analysisL11-12 Genetic oscillatorsL13-14 Stochastic chemical kinetics
9
I Systems Microbiology (14 Lectures)
‘The cell as a well-stirred biochemical reactor’
L1 IntroductionL2-3 Chemical kinetics, Equilibrium binding, cooperativityL4 Lambda phagep gL5 Cellular memoryL6-7 Genetic switchesL8-9 E. coli chemotaxisL10 Stability analysisL11-12 Genetic oscillatorsL13-14 Stochastic chemical kinetics
10
Introduction phage biology
Phage genome:48512 base pairs ~ 12 kB‘phage jpg’ ~ 10 kBphage.jpg ~ 10 kB
Excellent book:
A genetic switch by Mark Ptashne
11
A genetic switch by Mark Ptashne
12
The central dogma defines three major groups of biomolecules (biopolymers):
( 9 /1. DNA (passive library, 6×109 bp, 2 m/cell,75×1012 cells/human, total length150×1012 m/human ~ 1000 rsun-earth)
2. RNA (‘passive’ intermediate)3. Proteins (active work horses)
The fourth (and final) group consists ofso-called ‘small molecules’.
4. Small molecules (sugars, hormones,vitamines, ‘substrates’ etc.)
13
The lysis-lysogeny decision:
As the phage genome is injectedphage genes are transcribed andtranslated by using the host’smachinerymachinery.
Which set of phage proteins areexpressed determines the fate of thephage: lysis or lysogenyphage: lysis or lysogeny
14
The lysis-lysogeny decision is a genetic switch
RNA RNARNAp RNAp
15only ‘space’ for one RNA polymerase (mutual exclusion)
Single repressor dimer bound - three cases:
I Negative control, dimer binding to OR2 inhibitsRNAp binding to right PR promoter.
Positive control, dimer binding to OR2 enhancesRNAp binding to left PRM promoter.
ti ti
cI transcriptinhibition
activation
more repressor monomers
16more repressor dimers
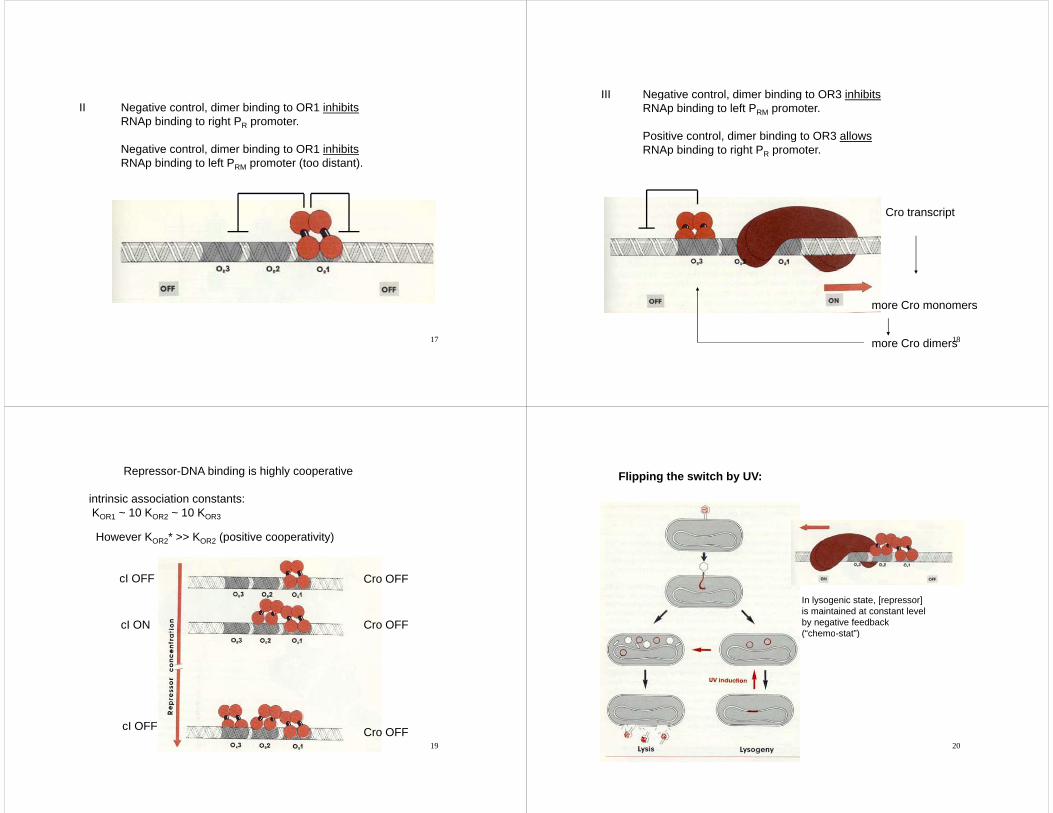
II Negative control, dimer binding to OR1 inhibitsRNAp binding to right PR promoter.
Negative control, dimer binding to OR1 inhibitsRNAp binding to left PRM promoter (too distant).
17
III Negative control dimer binding to OR3 inhibitsIII Negative control, dimer binding to OR3 inhibitsRNAp binding to left PRM promoter.
Positive control dimer binding to OR3 allowsPositive control, dimer binding to OR3 allowsRNAp binding to right PR promoter.
Cro transcript
more Cro monomers
18more Cro dimers
Repressor-DNA binding is highly cooperative
intrinsic association constants:intrinsic association constants:KOR1 ~ 10 KOR2 ~ 10 KOR3
However KOR2* >> KOR2 (positive cooperativity)OR2 OR2
cI OFF Cro OFF
cI ON Cro OFF
O19
cI OFF Cro OFF
Flipping the switch by UV:
In lysogenic state, [repressor]is maintained at constant levelby negative feedback(“chemo stat”)( chemo-stat )
20
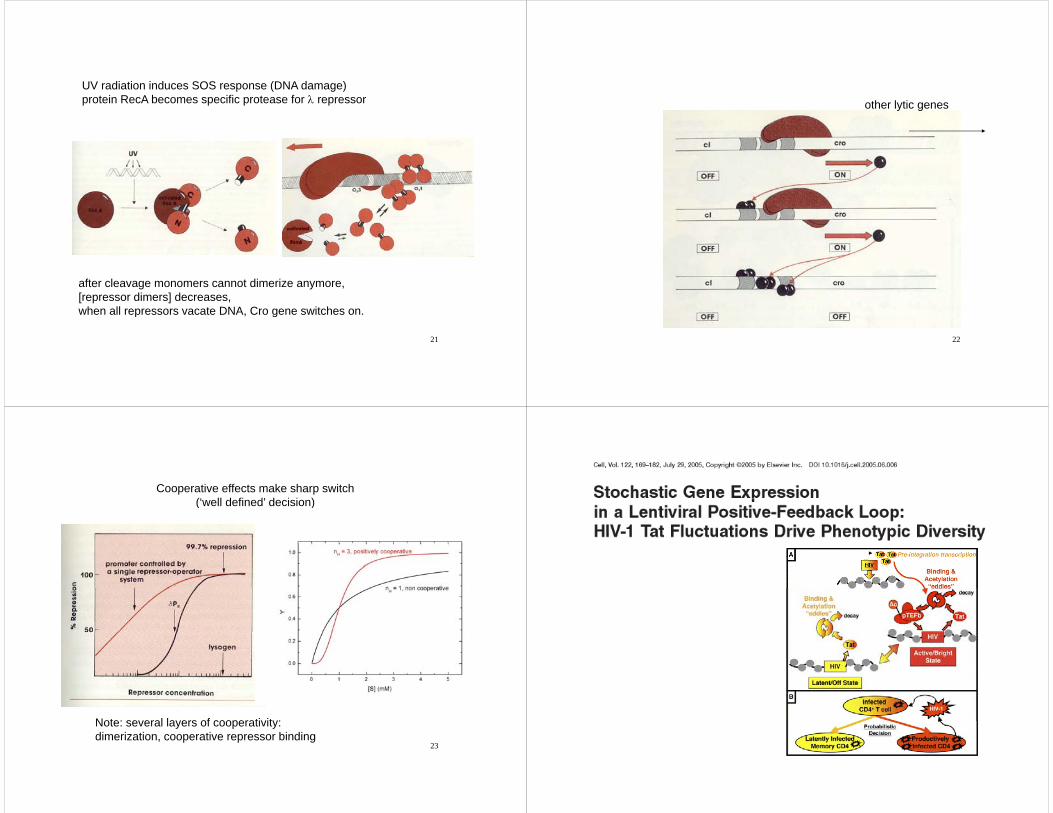
UV radiation induces SOS response (DNA damage)protein RecA becomes specific protease for λ repressor
after cleavage monomers cannot dimerize anymore,[repressor dimers] decreases,when all repressors vacate DNA, Cro gene switches on.
21
p , g
other lytic genes
22
Cooperative effects make sharp switch(‘ ll d fi d’ d i i )(‘well defined’ decision)
Note: several layers of cooperativity:
23
Note: several layers of cooperativity:dimerization, cooperative repressor binding
24
I Systems Microbiology (14 Lectures)
‘The cell as a well-stirred biochemical reactor’
L1 IntroductionL2-3 Chemical kinetics, Equilibrium binding, cooperativityL4 Lambda phagep gL5 Cellular memoryL6-7 Genetic switchesL8-9 E. coli chemotaxisL10 Stability analysisL11-12 Genetic oscillatorsL13-14 Stochastic chemical kinetics
25 26
The Flagellum
1000 H+ / rotation
27> 40 genes involved
Absence of chemical attractant
28
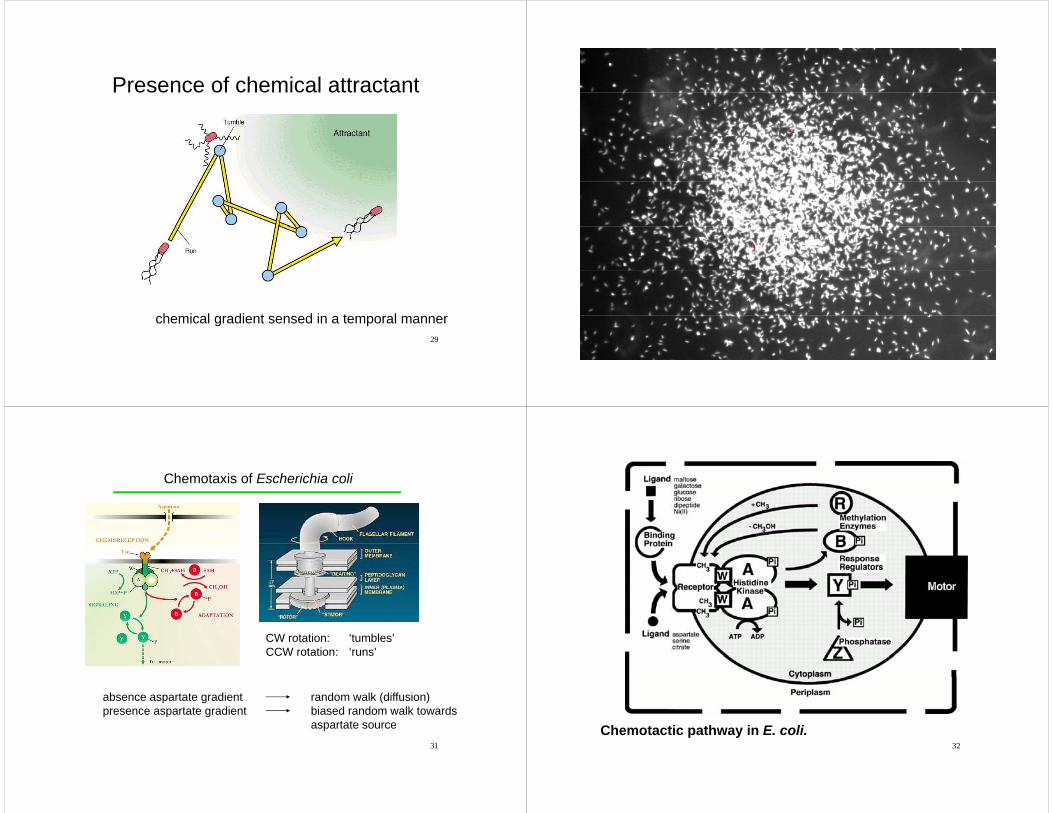
Presence of chemical attractantPresence of chemical attractant
h i l di t d i t l29
chemical gradient sensed in a temporal manner30
Chemotaxis of Escherichia coli
CW rotation: ‘tumbles’CCW rotation: ‘runs’
absence aspartate gradient random walk (diffusion)presence aspartate gradient biased random walk towards
t t
31
aspartate source
32
Chemotactic pathway in E. coli.

Adaptation:
Correlation of receptor methylation with behavioral response. p y p
33
slow
fast
What is the simplest mathematical modelthat is consistent with the biology and
34
that is consistent with the biology andreproduces the experiments ?
I Systems Microbiology (14 Lectures)
‘The cell as a well-stirred biochemical reactor’
L1 IntroductionL2-3 Chemical kinetics, Equilibrium binding, cooperativityL4 Lambda phagep gL5 Cellular memoryL6-7 Genetic switchesL8-9 E. coli chemotaxisL10 Stability analysisL11-12 Genetic oscillatorsL13-14 Stochastic chemical kinetics
35 36
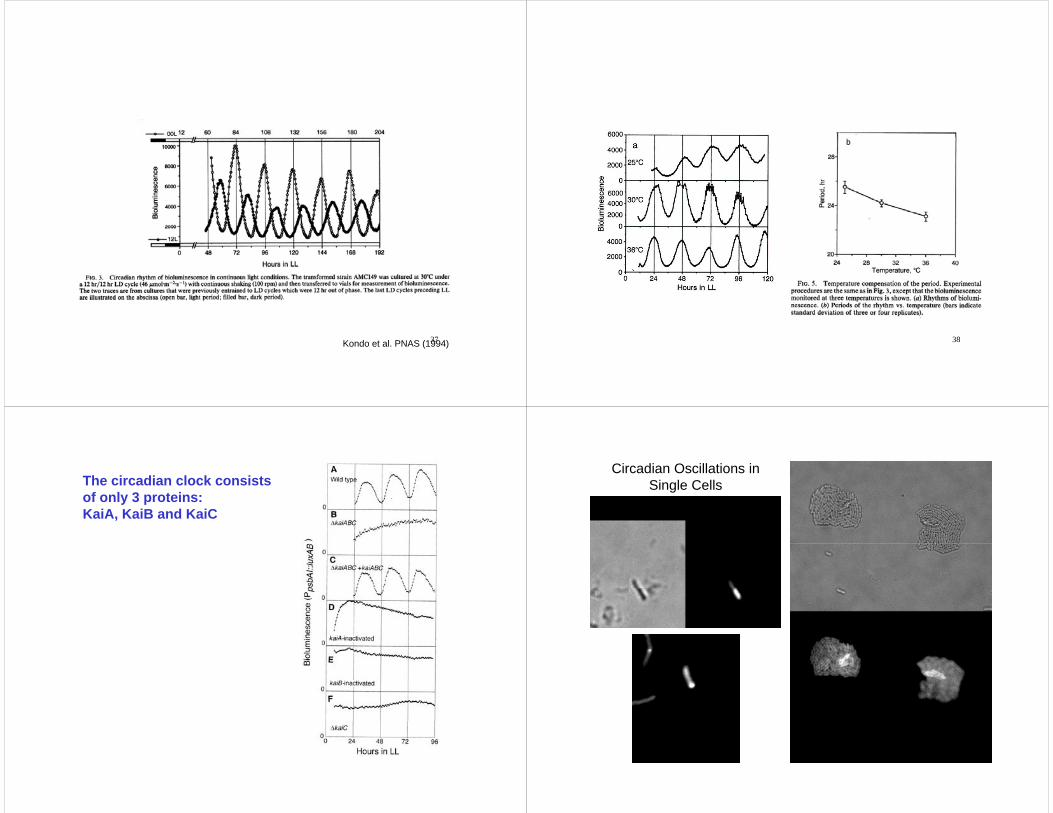
37Kondo et al. PNAS (1994) 38
The circadian clock consists of only 3 proteins:of only 3 proteins:KaiA, KaiB and KaiC
39
Circadian Oscillations inSingle Cells
40
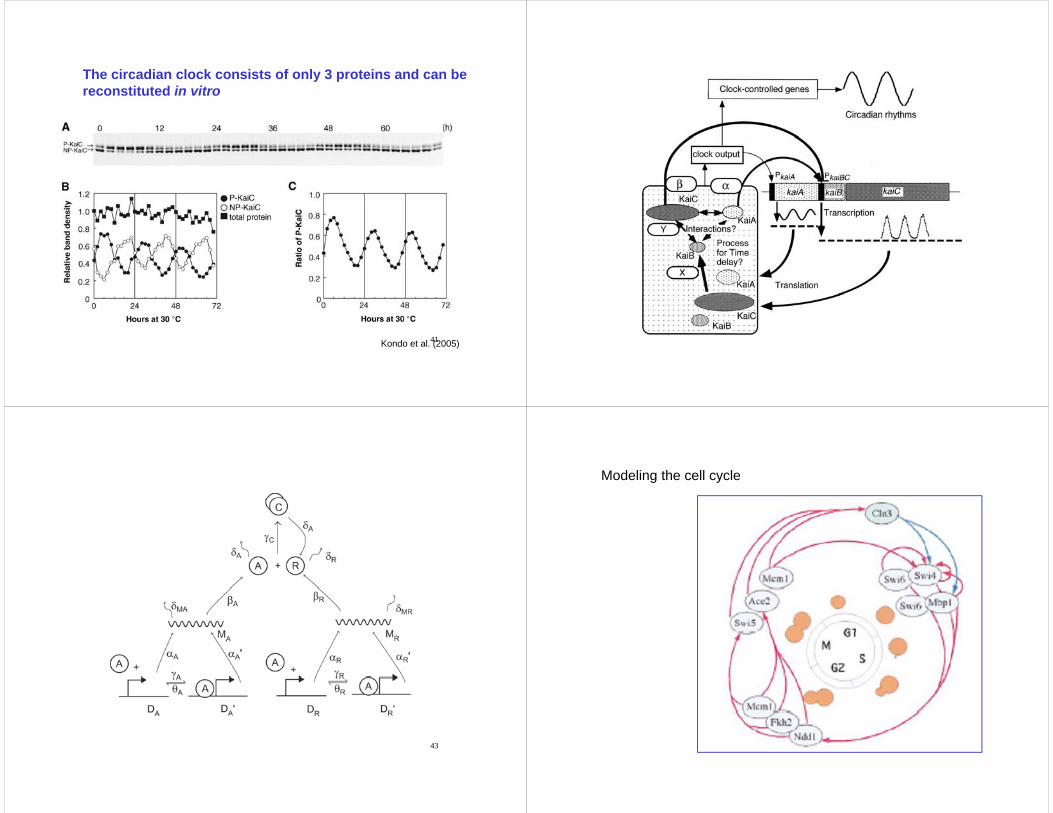
The circadian clock consists of only 3 proteins and can bereconstituted in vitroreconstituted in vitro
41Kondo et al. (2005) 42
43
Modeling the cell cycle
44
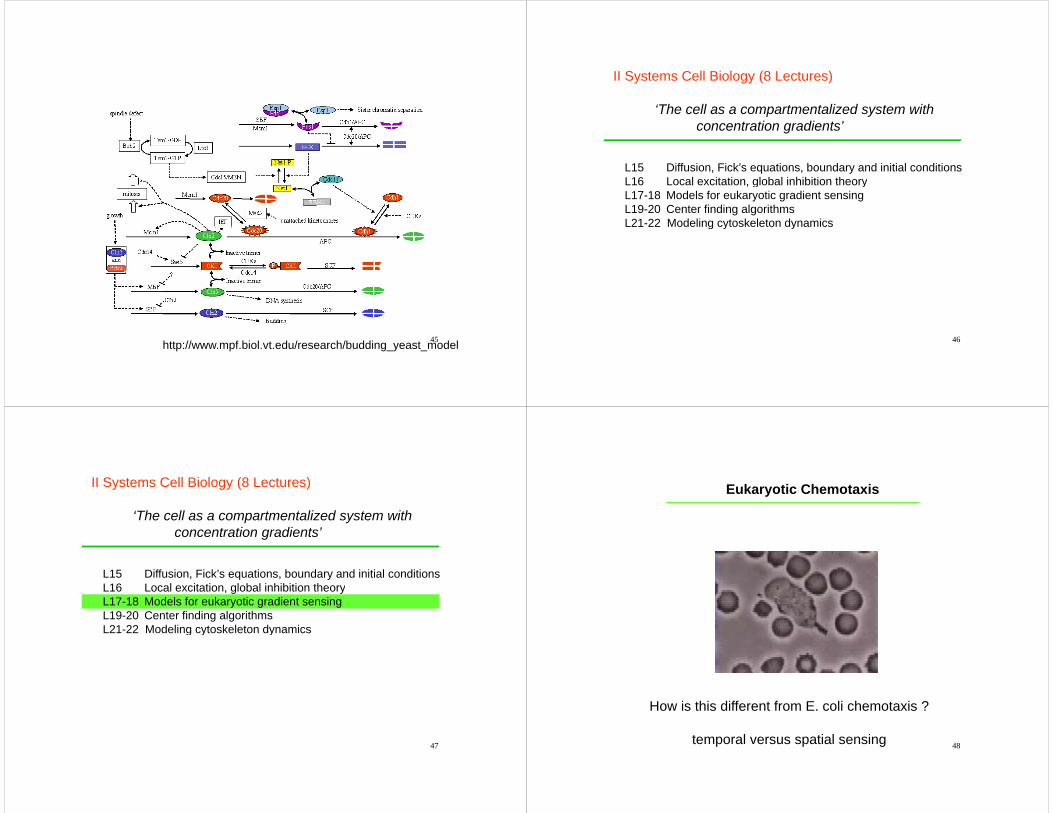
45http://www.mpf.biol.vt.edu/research/budding_yeast_model
II Systems Cell Biology (8 Lectures)
‘The cell as a compartmentalized system withconcentration gradients’
L15 Diffusion, Fick’s equations, boundary and initial conditionsL16 Local excitation global inhibition theoryL16 Local excitation, global inhibition theoryL17-18 Models for eukaryotic gradient sensingL19-20 Center finding algorithmsL21-22 Modeling cytoskeleton dynamicsL21 22 Modeling cytoskeleton dynamics
46
II Systems Cell Biology (8 Lectures)
‘The cell as a compartmentalized system withconcentration gradients’
L15 Diffusion, Fick’s equations, boundary and initial conditionsL16 Local excitation global inhibition theoryL16 Local excitation, global inhibition theoryL17-18 Models for eukaryotic gradient sensingL19-20 Center finding algorithmsL21-22 Modeling cytoskeleton dynamicsL21 22 Modeling cytoskeleton dynamics
47
Eukaryotic Chemotaxis
How is this different from E. coli chemotaxis ?
48temporal versus spatial sensing
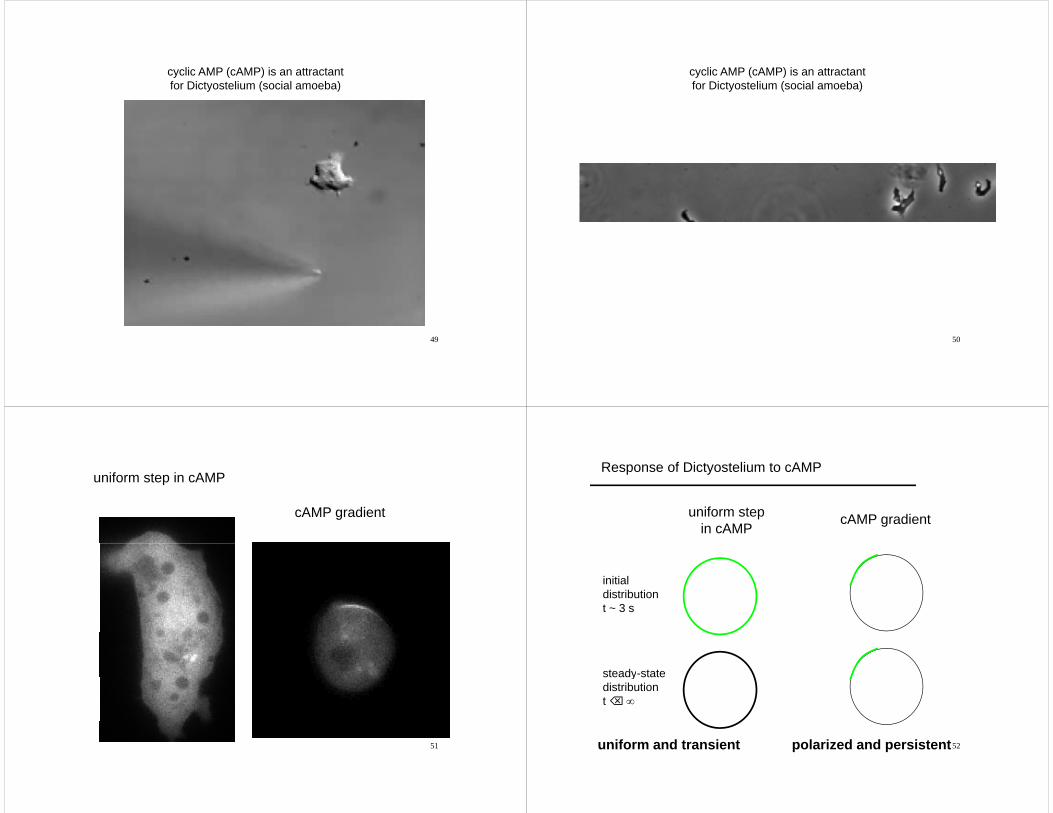
cyclic AMP (cAMP) is an attractantfor Dictyostelium (social amoeba)
49
cyclic AMP (cAMP) is an attractantfor Dictyostelium (social amoeba)
50
uniform step in cAMP
cAMP gradient
51
Response of Dictyostelium to cAMP
uniform stepin cAMP
cAMP gradient
initialdistributiont ~ 3 s
steady-statesteady statedistributiont ⌫ ∞
52uniform and transient polarized and persistent
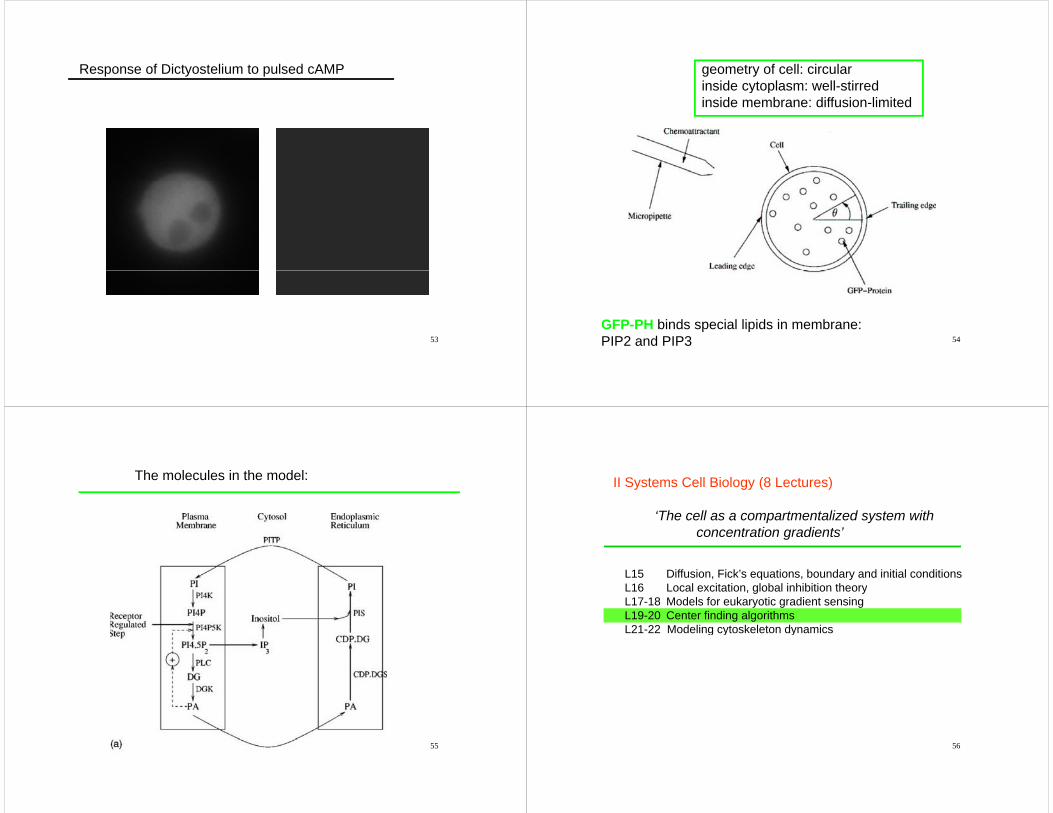
Response of Dictyostelium to pulsed cAMP
53
geometry of cell: circularinside cytoplasm: well-stirredy pinside membrane: diffusion-limited
54GFP-PH binds special lipids in membrane:PIP2 and PIP3
The molecules in the model:
55
II Systems Cell Biology (8 Lectures)
‘The cell as a compartmentalized system withconcentration gradients’
L15 Diffusion, Fick’s equations, boundary and initial conditionsL16 Local excitation global inhibition theoryL16 Local excitation, global inhibition theoryL17-18 Models for eukaryotic gradient sensingL19-20 Center finding algorithmsL21-22 Modeling cytoskeleton dynamicsL21 22 Modeling cytoskeleton dynamics
56
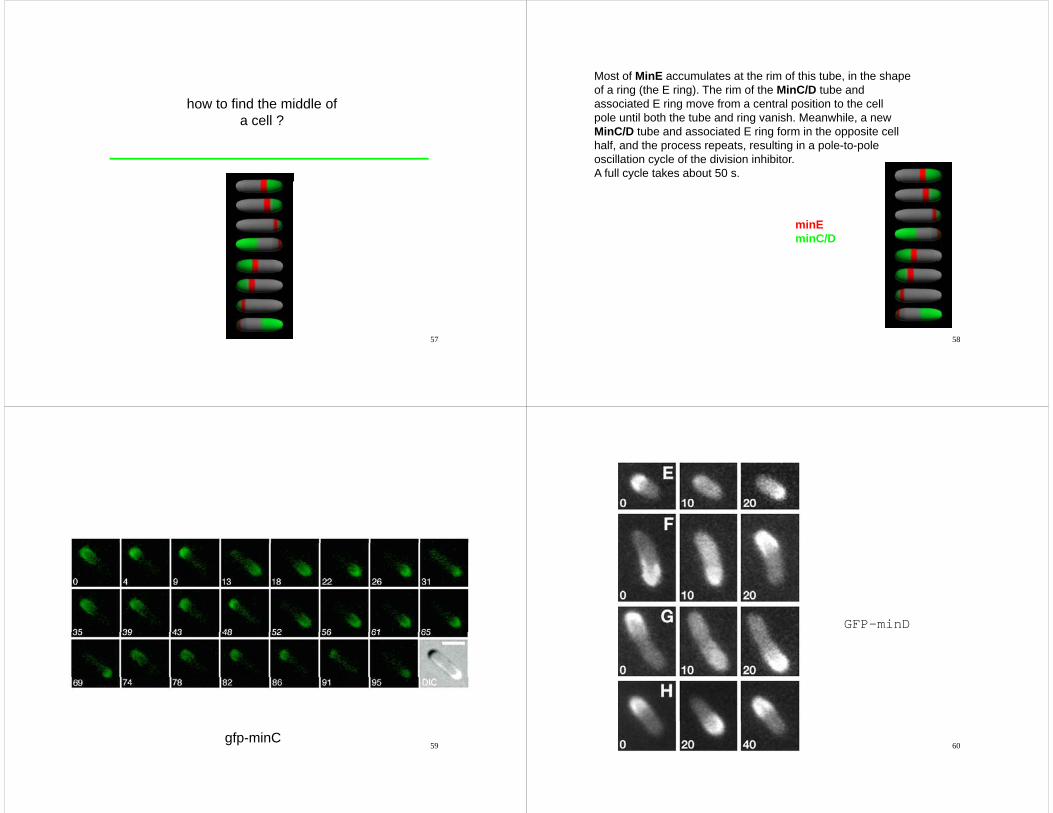
how to find the middle ofa cell ?
57
Most of MinE accumulates at the rim of this tube, in the shapeof a ring (the E ring) The rim of the MinC/D tube andof a ring (the E ring). The rim of the MinC/D tube andassociated E ring move from a central position to the cellpole until both the tube and ring vanish. Meanwhile, a newMinC/D tube and associated E ring form in the opposite cellg pphalf, and the process repeats, resulting in a pole-to-poleoscillation cycle of the division inhibitor.A full cycle takes about 50 s.
minEminEminC/D
58
59gfp-minC
GFP-minD
60
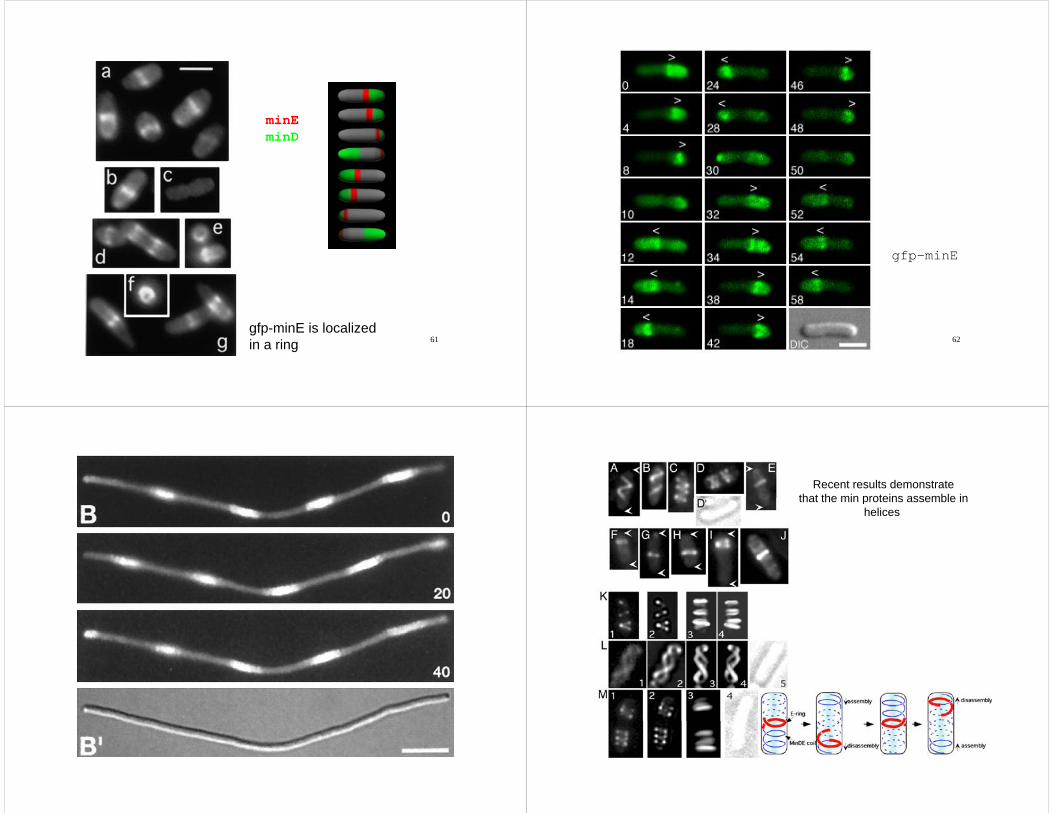
minEminDminD
61gfp-minE is localizedin a ring
gfp-minE
62
63
Recent results demonstratethat the min proteins assemble inthat the min proteins assemble in
helices
64
II Systems Cell Biology (8 Lectures)
‘The cell as a compartmentalized system withconcentration gradients’
L15 Diffusion, Fick’s equations, boundary and initial conditionsL16 Local excitation global inhibition theoryL16 Local excitation, global inhibition theoryL17-18 Models for eukaryotic gradient sensingL19-20 Center finding algorithmsL21-22 Modeling cytoskeleton dynamicsL21 22 Modeling cytoskeleton dynamics
65
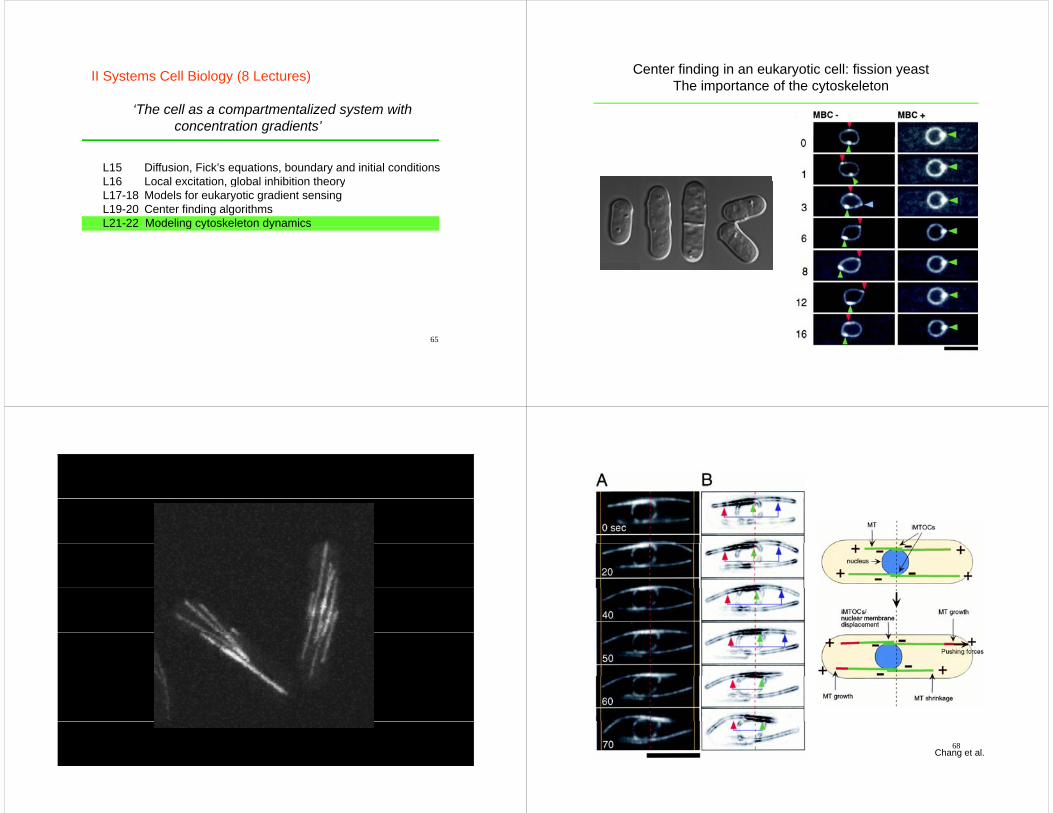
Center finding in an eukaryotic cell: fission yeastThe importance of the cytoskeletonp y
66
67 68Chang et al.
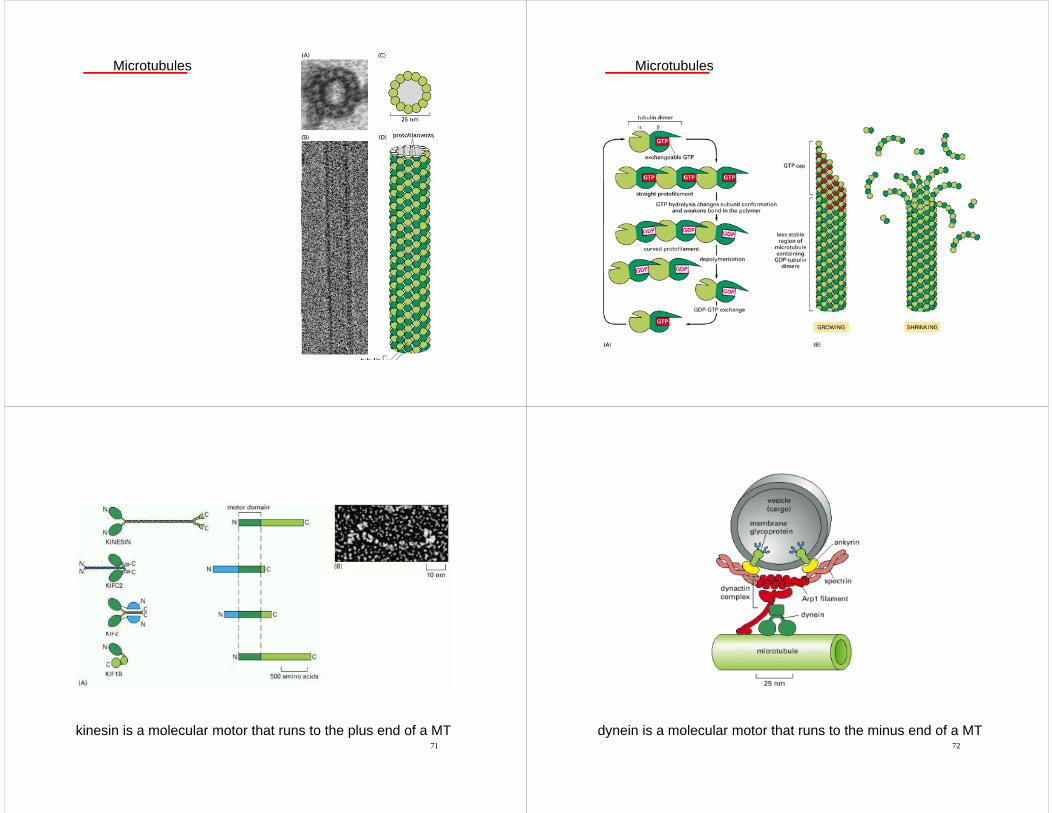
Microtubules
69
Microtubules
70
71
kinesin is a molecular motor that runs to the plus end of a MT72
dynein is a molecular motor that runs to the minus end of a MT

Black tetraBlack tetra
73Movies !
III Systems Developmental Biology (3 Lectures)
‘The cell in a social context communicating withneighboring cells’
L23 Quorum sensingL24 25 Drosophila developmentL24-25 Drosophila development
74
III Systems Developmental Biology (3 Lectures)
‘The cell in a social context communicating withneighboring cells’
L23 Quorum sensingL24 25 Drosophila developmentL24-25 Drosophila development
75 76
77
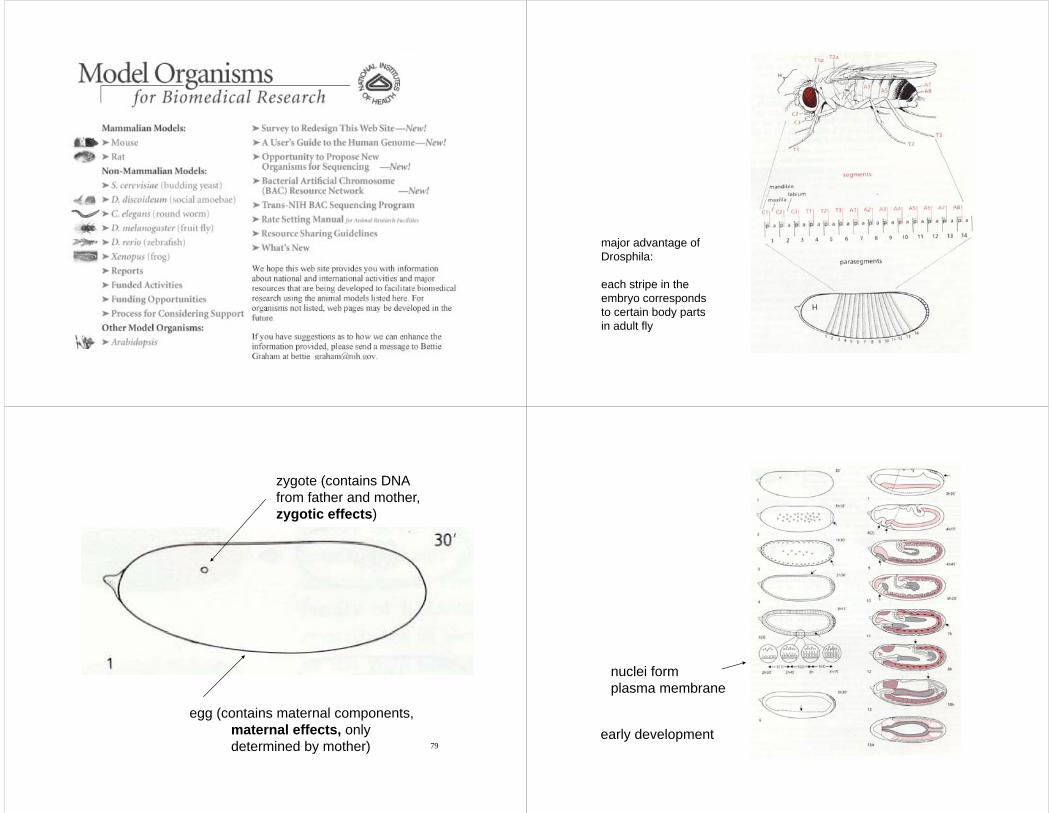
major advantage ofDrosphila:
each stripe in theembryo correspondsto certain body parts
78
to certain body partsin adult fly
zygote (contains DNAfrom father and motherfrom father and mother,zygotic effects)
egg (contains maternal components,
79
maternal effects, onlydetermined by mother)
nuclei formplasma membrane
80early development
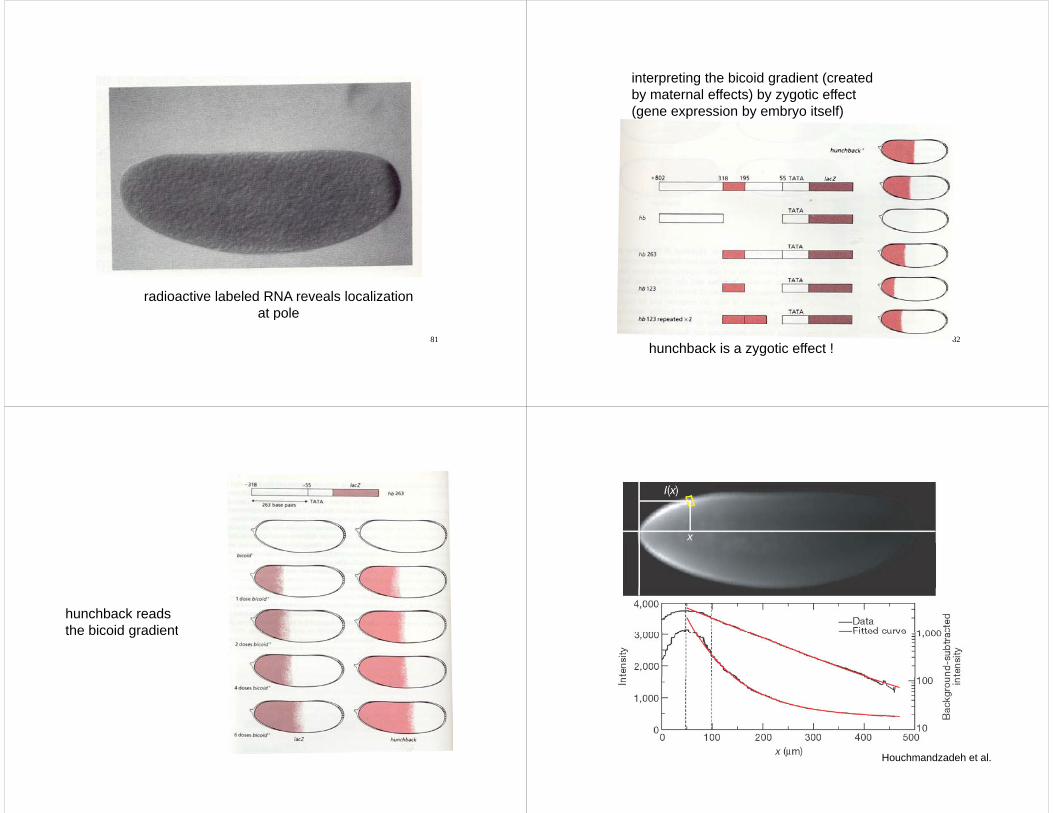
radioactive labeled RNA reveals localizationat pole
81
at pole
interpreting the bicoid gradient (createdby maternal effects) by zygotic effectby maternal effects) by zygotic effect(gene expression by embryo itself)
82hunchback is a zygotic effect !
hunchback readsthe bicoid gradientthe bicoid gradient
83 84Houchmandzadeh et al.
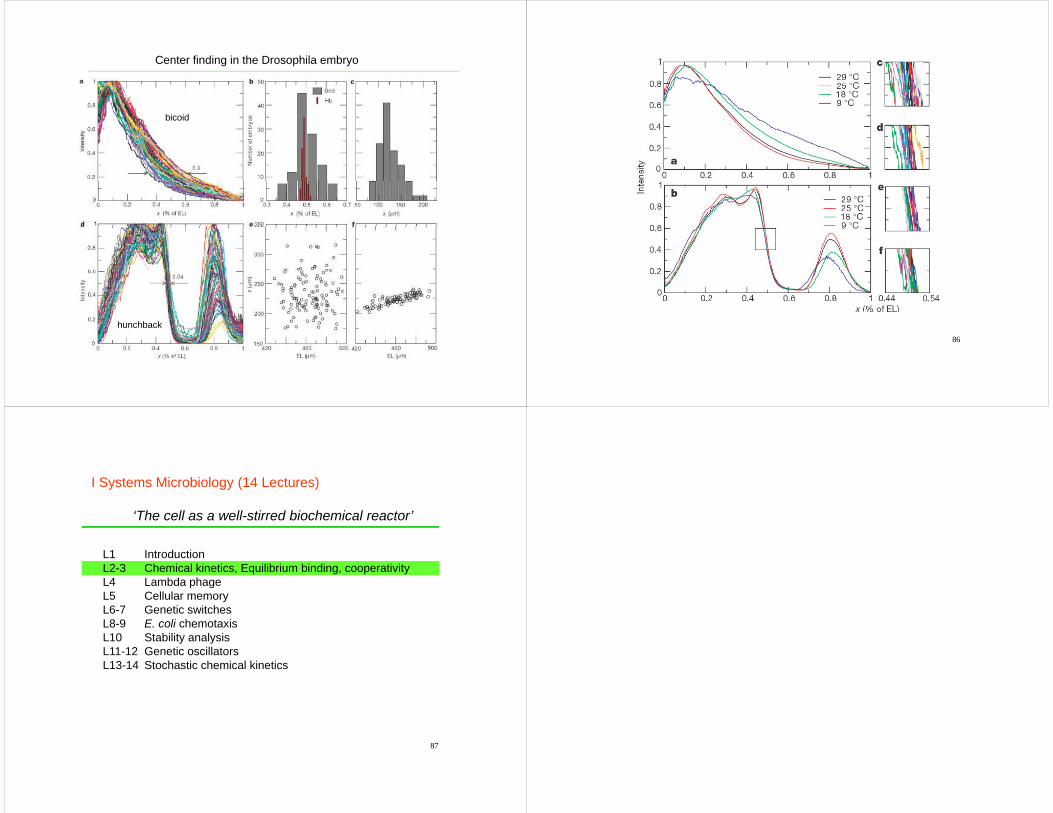
Center finding in the Drosophila embryo
bicoid
85
hunchback86
I Systems Microbiology (14 Lectures)
‘The cell as a well-stirred biochemical reactor’
L1 IntroductionL2-3 Chemical kinetics, Equilibrium binding, cooperativityL4 Lambda phagep gL5 Cellular memoryL6-7 Genetic switchesL8-9 E. coli chemotaxisL10 Stability analysisL11-12 Genetic oscillatorsL13-14 Stochastic chemical kinetics
87





























