Embed Size (px)
Citation preview

K
CFa
b
a
ARR2A
KACIPNKE
1
to
psccntdif
m1T
CT
1d
Pharmacological Research 65 (2012) 347– 357
Contents lists available at SciVerse ScienceDirect
Pharmacological Research
jo ur n al hom epage: www.elsev ier .com/ locate /yphrs
nock-down of endothelial connexins impairs angiogenesis
hristiane Gärtnera, Barbara Ziegelhöffera, Martin Kostelkaa, Holger Stepanb,riedrich-Wilhelm Mohra, Stefan Dheina,∗
Department of Cardiac Surgery, Heart Centre Leipzig, University of Leipzig, Struempellstr. 39, 04289 Leipzig, GermanyDepartment of Obstetrics, University Hospital of Leipzig, Liebigstr. 20a, 04103 Leipzig, Germany
r t i c l e i n f o
rticle history:eceived 3 June 2011eceived in revised form2 November 2011ccepted 22 November 2011
eywords:ngiogenesisonnexin
ntercellular communication
a b s t r a c t
Connexins (Cx) are suggested to play important roles in growth and differentiation. Aim of our study wasto investigate the role of endothelial Cx in the angiogenic process.
Several parameters of angiogenesis were assessed in 18 h Matrigel in vitro angiogenesis assays withhuman umbilical vein endothelial cells (HUVEC). Prior to culture on Matrigel cells were treated withnicotine or the gap junction inhibitor palmitoleic acid (PA), or siRNA-knock-down of either Cx37, Cx40 orCx43 was performed. Changes in Cx expression and their effects on gap-junctional communication wereinvestigated using immunofluorescence microscopy, Western blot and Lucifer Yellow dye transfer.
Knock-down of each Cx-isoform significantly reduced the amount of specific Cx protein in HUVEC. Cx-knock-down as well as treatment with PA impaired intercellular communication via gap junctions and
almitoleic acidicotinenock-downndothelium
diminished significantly the number of capillary branches. Knock-down of Cx43 and Cx40 or treatmentwith PA reduced complexity pattern in the angiogenesis assay.
Nicotine significantly reduced expression of Cx43 and Cx37 as well as average length of capillarybranches, number of branches and pattern in the Matrigel assay. We can conclude that connexins areinvolved in angiogenesis, in particular in branch formation. This can partly explain the changes in angio-genesis seen under nicotine treatment.
. Introduction
Intercellular communication via gap junction channels is knowno be an important modulatory factor for growth and differentiationf cells.
Gap junctions, as main mediator of cell–cell communication, arelasma membrane channels that allow the passages of ions andmall molecules up to a molecular weight of 1000 Da between adja-ent cells. Gap junctions are built by two hemi-channels, calledonnexon. Each connexon consists of six transmembrane span-ing proteins named connexin that dock to their counterparts onhe adjacent cell membrane. There are 21 isoforms of connexins
escribed, that are tissue-dependently expressed. In human umbil-cal vein endothelial cells (HUVECs) Cx37, Cx40 and Cx43 wereound [1].
Abbreviations: PA, palmitoleic acid; ECGS, endothelial cell growth supple-ent; NFS, Nanoparticle Formation Solution; NCX1, sodium–calcium exchanger
; CHAPS, 3-[(3-cholamidopropyl)dimethylammonio]-1-propanesulfonate; TBST,ris–buffered saline with Tween-20; scr-si, scrambled-siRNA.∗ Corresponding author at: University of Leipzig, Heart Centre Leipzig, Clinic forardiac Surgery, Struempellstr. 39, 04289, Leipzig, Germany.el.: +49 0 341 8651651; fax: +49 0 341 8651452.
E-mail address: [email protected] (S. Dhein).
043-6618/$ – see front matter © 2011 Elsevier Ltd. All rights reserved.oi:10.1016/j.phrs.2011.11.012
© 2011 Elsevier Ltd. All rights reserved.
Several groups investigated the importance of connexins for thecardiovascular system. Thus, Cx43 was reported to prolong G1 andS-phases of the cell cycle [2], whereas Cx37 was shown to slow allcell cycle phases [3]. In Cx40-deficient mice cardiac malformationscould be seen [4]. Combined endothelial Cx40- and Cx37-knockoutmice developed severe abnormalities of the vascular system, hem-orrhages and abnormally large and distended vessels in certainareas [5]. Additionally, mutations in the gene for Cx43 (GJA1) werefound to cause a hypoplastic left heart syndrome [6]. Moreover,blood pressure can be influenced by the expression of connexinsin different ways: knockout of Cx43 in mice caused hypotension,whereas Cx40-knockout in mice led to hypertension [7]. More-over, it is generally known, that gap junctions are involved in theregulation of growth and differentiation. On account of this wefocused our study first on the following question: do the variousendothelial connexins exert different roles in the angiogenic pro-cess? Nicotine was reported to down-regulate Cx43 and Cx37 incultivated HUVECs under 1 �M nicotine-treatment and concomi-tantly to impair cell–cell communication [8,9]. Thus, we wantedto test a possible relevance of our knock-down-results by incubat-
ing HUVECs for 5 days with nicotine to induce a pharmacologicaldown-regulation of connexins and to evaluate the Cx expressionin both 2D petri dishes- and a 3D- Matrigel culture as well as thecapillary formation in the angiogenesis assay in vitro.
348 C. Gärtner et al. / Pharmacological R
Table 1Sequences of the target-siRNA used for knock-down of the different connexins.
Cx37 SenseAntisense
5′-CAGACAGAGUCAGCAUGGA [dT] [dT]5′-UCCAUGCUGACUCUGUCUG [dT] [dT]
Cx40 SenseAntisense
5′-GAGAGGUUUGUCUGCAACA [dT] [dT]5′-UGUUGCAGACAAACCUCUC [dT] [dT]
2
atb
2
u1aB3mssG(5wsgm1GcM
ts
2
RTt
wNr
wfG3NmtMHpowm
Cx43 SenseAntisense
5′-GAAAGUGCACCAGGUGUUA [dT] [dT]5′-UAACACCUGGUGCACUUUC [dT] [dT]
. Materials and methods
The umbilical cords were obtained from Obstetrics Clinics aftern informed written consent was given. The investigation confirmso the principles of the Declaration of Helsinki and was approvedy the Ethical Committee of the University of Leipzig.
.1. Cell culture (2D culture)
HUVECs were prepared as previously described [6]. Briefly, freshmbilical cords were rinsed with phosphate buffered saline (PBS:.5 mM KH2PO4, 0.5 mM Na2HPO4, 120 mM NaCl, 2.7 mM KCl)nd the HUVECs were isolated using collagenase IV (Biochrom,erlin, Germany) dissolved in PBS. After 17 min incubation at7 ◦C the collagenase activity was stopped by addition of 10 mledium I. Medium I consisted of M199 with Earle’s balanced salt
olution with l-glutamine and Hepes (Lonza, Verviers, Belgium)upplemented with 10% fetal calf serum, Biochrom (FCS, Berlin,ermany) and 100 000 U l−1 penicillin/100 mg l−1 streptomycin
Sigma, Steinheim, Germany). Cell suspension was centrifuged min at 1200 rpm and pellet was resolved in medium I. Cellsere disseminated onto gelatine-coated 6-well-plates at a den-
ity of 300 000 cells in 2 ml medium per well. To minimizeenetic varieties cells were pooled before plating. After 2–3 hedium I was replaced by medium II (medium I and supplemental
6.7 �g l−1 endothelial cell growth supplement (c.c. pro, Oberdorla,ermany)). Cells were grown at 37 ◦C in a humified atmosphereontaining 5% CO2 for 5 ± 0.5 days until they were nearly confluent.edium II was changed every 48 h.Treatment of cells with nicotine started on the first day of cul-
ivation, whereas the protein knock-down via RNA-interferencetarted after cultivated HUVECs were nearly confluent.
.2. siRNA experiments
Nearly confluent cells were incubated with small interferingNA (siRNA) directed against either Cx37, Cx40 or Cx43 for 24 h.hereafter, cells were either (a) lysed for protein isolation or (b)rypsinized and transferred to Matrigel for angiogenesis-assay.
For optimization of siRNA-transfer HUVECs in 2D-cultureere treated with siRNA for Cx37, Cx40 or Cx43 using-TER/siRNA nanoparticles (Sigma, Steinheim, Germany). The
espective sequences are shown in Table 1.For siRNA Transfection Nanoparticle Formation Solution (NFS)
as prepared by mixing 5 �M siRNA in siRNA dilution bufferrom the N-TER/siRNA nanoparticles system (Sigma, Steinheim,ermany) with N-TER peptide dissolved in DEPC-water. After0 min incubation at room temperature appropriate volumes ofFS were added to 1.5 ml serum-free medium M199 (supple-ented with penicillin/streptomycin solution as described before)
o achieve the final siRNA-concentration of 10 or 20 nM per well.edium with NFS was spread on gelatine-coated plates. Thereafter,UVECs in serum-free medium were seeded on the pre-treated
lates at two-fold of the desired final cell density. After 3 hf incubation at 37 ◦C and 5% CO2 Medium 199 supplementedith four-fold FCS and four-fold endothelial cell growth supple-ent (ECGS) was added, so that the normal concentration of FCSesearch 65 (2012) 347– 357
and ECGS was reached. After 24 h NFS-containing medium wasremoved, cells were washed with PBS (containing Ca2+ and Mg2+)and medium II was added for further cultivation. Cells were har-vested directly, 12 h or 24 h after the end of siRNA exposure andWestern blot experiments were performed. The most effectiveknock-down protocols were used for the subsequent angiogene-sis assay, i.e. for Cx37 20 nM, for Cx40 20 nM and for Cx43 10 nM.Cells were used directly after 24 h incubation with NFS containingthe specific-siRNA for angiogenesis experiments. In control exper-iments commercially available scrambled siRNA (Mission® siRNAUniversal negative control 1, Sigma, Steinheim, Germany) in a con-centration of 20 nM or NFS only were used.
2.3. Nicotine treatment
In order to simulate a subchronic exposition to nicotine, forthese experiments HUVECs were immediately after isolation incu-bated with (−)-nicotine (Sigma, Steinheim, Germany) at finalconcentration of 1 �M nicotine in medium II. Nearly confluent cellswere after 5 ± 0.5 days trypsinized and angiogenesis-assay was per-formed still in the presence of nicotine in medium. Control cellswere treated with the vehicle ethanol at the final concentration of0.1 vol%.
2.4. Treatment with palmitoleic acid
We used the gap junction blocker palmitoleic acid to inhibitthe gap junctional communication of the HUVECs. Therefore wetrypsinized nearly confluent HUVECs and during the in vitro angio-genesis assay we treated the cells for 18 h with 10 �M palmitoleicacid that is a concentration above EC50 [10].
2.5. Angiogenesis assay (3D culture)
Testing for HUVECs tube formation was performed in a chamber(A = 0.8 cm2) containing 100 �l Matrigel (in vitro angiogenesis assayECM625, Millipore, USA) according to the manufacturer’s protocol.Briefly, 100 �l Matrigel-solution were added to one chamber of theLab-Tek II Chamber Slide system (Lab-Tek, Naperville, USA) andallowed to solidify and polymerize at 37 ◦C and 5% CO2 for 1 h.
Nearly confluent HUVECs were stained 30 min with 5 �l 1% FITC-lectin (from Ulex europaeus UEA I, Sigma, Steinheim, Germany)dissolved in medium I, than trypsinized, centrifuged at 1200 rpmfor 10 min and resuspended in medium II. Finally, 200 �l mediumII containing 200 000 cells were layered on top of the Matrigel. Fornicotine experiments either nicotine (final concentration 1 �M) orvehicle (ethanol: final concentration 1 mM) was added to mediumII.
After 18 h incubation at 37 ◦C and 5% CO2 cells were stainedwith Hoechst 33258 (bisBenzimide, 0.1 mg/ml in dH2O, molecularprobes, Eugene, USA) for 10 min and mounted with DakoCytoma-tion Fluorescent Mounting medium (Dako, Hamburg, Germany).Tube formation was inspected using a Zeiss Axiolab fluorescencemicroscope (Zeiss, Jena, Germany) and a commercial image analysissystem (Zeiss Vision).
We evaluated 25 pictures per object plate (magnification 200×)and the number of cells, number of branches, average length ofthe capillary branches, number of branches to the length of capil-laries ratio and the pattern were assessed using Zeiss Axio VisionSoftware 4.7. For pattern assessment pictures were assigned pointsaccording to the following scale: 0 = single cells; 1 = cells start to
grow and orientate; 2 = cells form tubes, but there are no branches;3 = cells form tubes and there are branches; 4 = nearly complexnetwork; 5 = complex network. Additional phase contrast pictureswere taken (magnification 100×).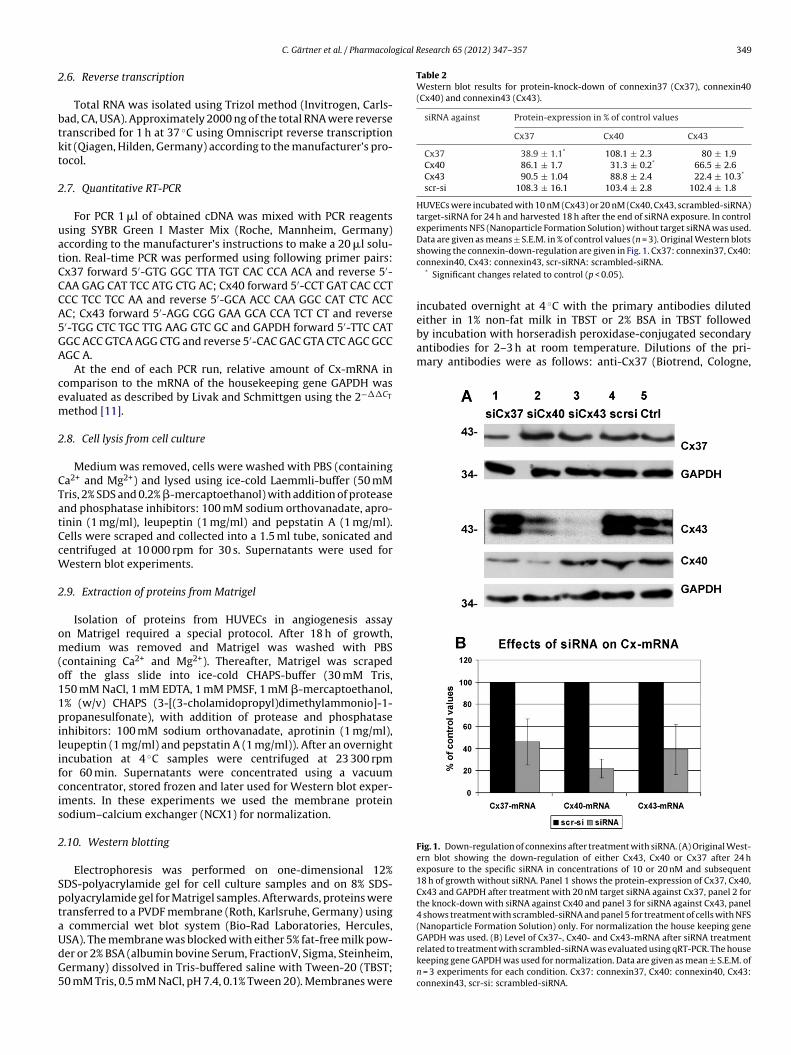
gical Research 65 (2012) 347– 357 349
2
btkt
2
uatCCCA5GA
cem
2
CTatCcW
2
om(o11pilifcis
2
SptaUdG5
Table 2Western blot results for protein-knock-down of connexin37 (Cx37), connexin40(Cx40) and connexin43 (Cx43).
siRNA against Protein-expression in % of control values
Cx37 Cx40 Cx43
Cx37 38.9 ± 1.1* 108.1 ± 2.3 80 ± 1.9Cx40 86.1 ± 1.7 31.3 ± 0.2* 66.5 ± 2.6Cx43 90.5 ± 1.04 88.8 ± 2.4 22.4 ± 10.3*
scr-si 108.3 ± 16.1 103.4 ± 2.8 102.4 ± 1.8
HUVECs were incubated with 10 nM (Cx43) or 20 nM (Cx40, Cx43, scrambled-siRNA)target-siRNA for 24 h and harvested 18 h after the end of siRNA exposure. In controlexperiments NFS (Nanoparticle Formation Solution) without target siRNA was used.Data are given as means ± S.E.M. in % of control values (n = 3). Original Western blots
either in 1% non-fat milk in TBST or 2% BSA in TBST followedby incubation with horseradish peroxidase-conjugated secondaryantibodies for 2–3 h at room temperature. Dilutions of the pri-mary antibodies were as follows: anti-Cx37 (Biotrend, Cologne,
Fig. 1. Down-regulation of connexins after treatment with siRNA. (A) Original West-ern blot showing the down-regulation of either Cx43, Cx40 or Cx37 after 24 hexposure to the specific siRNA in concentrations of 10 or 20 nM and subsequent18 h of growth without siRNA. Panel 1 shows the protein-expression of Cx37, Cx40,Cx43 and GAPDH after treatment with 20 nM target siRNA against Cx37, panel 2 forthe knock-down with siRNA against Cx40 and panel 3 for siRNA against Cx43, panel4 shows treatment with scrambled-siRNA and panel 5 for treatment of cells with NFS(Nanoparticle Formation Solution) only. For normalization the house keeping gene
C. Gärtner et al. / Pharmacolo
.6. Reverse transcription
Total RNA was isolated using Trizol method (Invitrogen, Carls-ad, CA, USA). Approximately 2000 ng of the total RNA were reverseranscribed for 1 h at 37 ◦C using Omniscript reverse transcriptionit (Qiagen, Hilden, Germany) according to the manufacturer’s pro-ocol.
.7. Quantitative RT-PCR
For PCR 1 �l of obtained cDNA was mixed with PCR reagentssing SYBR Green I Master Mix (Roche, Mannheim, Germany)ccording to the manufacturer’s instructions to make a 20 �l solu-ion. Real-time PCR was performed using following primer pairs:x37 forward 5′-GTG GGC TTA TGT CAC CCA ACA and reverse 5′-AA GAG CAT TCC ATG CTG AC; Cx40 forward 5′-CCT GAT CAC CCTCC TCC TCC AA and reverse 5′-GCA ACC CAA GGC CAT CTC ACCC; Cx43 forward 5′-AGG CGG GAA GCA CCA TCT CT and reverse′-TGG CTC TGC TTG AAG GTC GC and GAPDH forward 5′-TTC CATGC ACC GTCA AGG CTG and reverse 5′-CAC GAC GTA CTC AGC GCCGC A.
At the end of each PCR run, relative amount of Cx-mRNA inomparison to the mRNA of the housekeeping gene GAPDH wasvaluated as described by Livak and Schmittgen using the 2−��CT
ethod [11].
.8. Cell lysis from cell culture
Medium was removed, cells were washed with PBS (containinga2+ and Mg2+) and lysed using ice-cold Laemmli-buffer (50 mMris, 2% SDS and 0.2% �-mercaptoethanol) with addition of proteasend phosphatase inhibitors: 100 mM sodium orthovanadate, apro-inin (1 mg/ml), leupeptin (1 mg/ml) and pepstatin A (1 mg/ml).ells were scraped and collected into a 1.5 ml tube, sonicated andentrifuged at 10 000 rpm for 30 s. Supernatants were used for
estern blot experiments.
.9. Extraction of proteins from Matrigel
Isolation of proteins from HUVECs in angiogenesis assayn Matrigel required a special protocol. After 18 h of growth,edium was removed and Matrigel was washed with PBS
containing Ca2+ and Mg2+). Thereafter, Matrigel was scrapedff the glass slide into ice-cold CHAPS-buffer (30 mM Tris,50 mM NaCl, 1 mM EDTA, 1 mM PMSF, 1 mM �-mercaptoethanol,% (w/v) CHAPS (3-[(3-cholamidopropyl)dimethylammonio]-1-ropanesulfonate), with addition of protease and phosphatase
nhibitors: 100 mM sodium orthovanadate, aprotinin (1 mg/ml),eupeptin (1 mg/ml) and pepstatin A (1 mg/ml)). After an overnightncubation at 4 ◦C samples were centrifuged at 23 300 rpmor 60 min. Supernatants were concentrated using a vacuumoncentrator, stored frozen and later used for Western blot exper-ments. In these experiments we used the membrane proteinodium–calcium exchanger (NCX1) for normalization.
.10. Western blotting
Electrophoresis was performed on one-dimensional 12%DS-polyacrylamide gel for cell culture samples and on 8% SDS-olyacrylamide gel for Matrigel samples. Afterwards, proteins wereransferred to a PVDF membrane (Roth, Karlsruhe, Germany) using
commercial wet blot system (Bio-Rad Laboratories, Hercules,
SA). The membrane was blocked with either 5% fat-free milk pow-er or 2% BSA (albumin bovine Serum, FractionV, Sigma, Steinheim,ermany) dissolved in Tris-buffered saline with Tween-20 (TBST;0 mM Tris, 0.5 mM NaCl, pH 7.4, 0.1% Tween 20). Membranes wereshowing the connexin-down-regulation are given in Fig. 1. Cx37: connexin37, Cx40:connexin40, Cx43: connexin43, scr-siRNA: scrambled-siRNA.
* Significant changes related to control (p < 0.05).
incubated overnight at 4 ◦C with the primary antibodies diluted
GAPDH was used. (B) Level of Cx37-, Cx40- and Cx43-mRNA after siRNA treatmentrelated to treatment with scrambled-siRNA was evaluated using qRT-PCR. The housekeeping gene GAPDH was used for normalization. Data are given as mean ± S.E.M. ofn = 3 experiments for each condition. Cx37: connexin37, Cx40: connexin40, Cx43:connexin43, scr-si: scrambled-siRNA.

350 C. Gärtner et al. / Pharmacological Research 65 (2012) 347– 357
Fig. 2. Dye coupling in HUVECs after treatment with siRNA against Cx43, Cx40 or Cx37. Cells were treated with siRNA against Cx43, Cx40, Cx37 or scrambled-siRNA for 24 h.After subsequent 18 h growth without siRNA the dye transfer was measured using Lucifer Yellow. Number of communicating cells was assessed 5 min after dye injection.s umbeg s betwC
G1(UDslSeifi
RcGs
iRNA-treatment against Cx37 (B), Cx40 (C) or Cx43 (D) significantly diminished niven as mean ± S.E.M. of n = 8 experiments for each condition. Significant differencex37: connexin37, Cx40: connexin40, Cx43: connexin43, scr-si: scrambled-siRNA.
ermany) 1:500, anti-Cx40 (Santa Cruz, Heidelberg, Germany):1000, anti-Cx43 (Sigma, Steinheim, Germany) 1:2000, anti-NCX1Abcam, Cambridge, UK) 1:1000, caveolin-1 (Abcam, Cambridge,K) 1:1000 and GAPDH (HyTest Ltd., Turku, Finland) 1:10 000.ilution of secondary antibodies was 1:5000. For detection of the
pecific chemiluminescent signal either Uptilight HRP Blot Chemi-uminescent Substrate (Interchim, Montlucon Cedex, France), oruperSignal West Pico Chemiluminescent Substrate (Thermo Sci-ntific, Rockford, USA) were used according to manufacturer’snstructions and membranes were exposed to Kodak XOmat ARlm.
The results were evaluated using Quantity One Software (Bio-
ad Laboratories, Hercules, USA) and expressed as factor ofhange from control values and normalized to the respectiveAPDH-signal for cell culture samples or NCX1-signal for Matrigel-amples.r of communicating cells related to treatment with scrambled-siRNA (A). Data areeen siRNA- and scrambled-siRNA-treatment are indicated by an asterisk (p < 0.05).
2.11. Dye coupling
For dye transfer experiments cells were grown on a cover-slip with the beginning of treatment with siRNA. Then they weretransferred to a 1 ml organ bath superfused with Tyrode’s solu-tion (135 mM NaCl, 4 mM KCl, 2 mM CaCl2, 1 mM MgCl2, 0.33 mMNaH2PO4, 10 mM HEPES and 10 mM glucose, pH 7.4) at 37 ◦C. Cellswere patched using glass pipettes of 5–6 M� filled with “intracel-lular” solution (140 mM KCl, 4 mM MgCl2, 0.06 mM CaCl2, 5 mMEGTA, 3.1 mM Na2ATP, 5 mM Na2creatinephosphate, 10 mM HEPES,pH 7.1, and 0.1% Lucifer Yellow (Sigma, Steinheim, Germany)). Afterbreak-in and establishing the whole cell configuration cells were
injected with Lucifer Yellow and the dye transfer in the adjacentcells was measured 5 min after dye injection. The number of com-municating cells was measured using computer-assisted imageanalysis.
C. Gärtner et al. / Pharmacological Research 65 (2012) 347– 357 351
Fig. 3. Effect of connexin-knock-down on capillary like network. (A)–(F) In vitro angiogenesis assays. HUVECs were treated with scrambled-siRNA (A) or with 20 nM siRNAa wereg st pictc
2
wwwbfp(Gww1hAtDwb
gainst Cx37 (B) or Cx40 (C) or 10 nM siRNA against Cx43 (D). Additionally HUVECsrown in the presence of 1 �M nicotine for 5 days (F). Representative phase contraonnexin43, scr-siRNA: scrambled-siRNA.
.12. Immunohistochemistry
After 18 h growth of cells into the Matrigel, Matrigel wasashed with PBS for three times, fixed with formalin for 10 min,ashed three times 5 min with PBS and permeabilized for 30 minith Triton-X 100 (1% in PBS). After that Matrigel was incu-
ated with 0.1% bovine serum albumin (BSA, dissolved in PBS)or 20 min. Then Matrigels were incubated with the followingrimary antibodies at a dilution of 1:100 overnight: anti-Cx37Biotrend, Cologne, Germany), anti-Cx40 (Santa Cruz, Heidelberg,ermany) and anti-Cx43 (Sigma, Steinheim, Germany). Matrigelas washed three times with 0.1% BSA-solution and incubatedith the following secondary antibodies in a dilution of 1:100 for
h: anti-rabbit-IgG conjugated with AlexaFluor488 (Sigma, Stein-eim, Germany) for Cx37, Cx43 and anti-goat-IgG conjugated withlexaFluor488 (Sigma, Steinheim, Germany) for Cx40. Matrigel was
hen washed with 0.1% BSA-solution and nuclei were stained withAPI (Roche Diagnostics, Mannheim, Germany) for 1 min. Matrigelas then embedded in Fluorescent Mounting medium (Dako, Ham-
urg, Germany). Connexin-distribution was analyzed using a Zeiss
either treated with 10 �M palmitoleic acid (E) during the 18 h of Matrigel-assay orures were taken. Scale bars = 200 �m. Cx37: connexin37, Cx40: connexin40, Cx43:
Axiolab fluorescence microscope (Zeiss, Jena, Germany) and acommercial image analysis system (Zeiss Axio Vision, Zeiss, Jena,Germany).
2.13. Statistics
All data are given as mean ± S.E.M. Statistical analysis was per-formed with Systat 11.0 (Systat Software GmbH, Erkrath, Germany)using analysis of variance followed Mann–Whitney rank sum test.For dye transfer experiments Kruskal–Wallis test was used. Differ-ences were considered statistically significant if p was <0.05.
3. Results
3.1. siRNA interference
Efficiency of siRNA interference in reducing the amount of targetproteins Cx37, Cx40 and Cx43 in the cells was assessed by West-ern blot (Table 2 and Fig. 1A) and reduced amount of Cx-mRNAwas assessed using qRT-PCR (Fig. 1B). Our experiments revealed

352 C. Gärtner et al. / Pharmacological Research 65 (2012) 347– 357
Fig. 4. Effect of connexin-knock-down on angiogenesis. (A)–(D) In vitro angiogenesis assay. HUVECs were treated with 20 nM Cx37-target-siRNA (B), 20 nM Cx40-target-siRNA(C), 10 nM Cx43-target-siRNA (D) or with transfection reagent only (A) for 24 h. Cells were stained with lectin (FITC-conjugated, green fluorescence) and nuclei are stainedwith Hoechst 33258 (blue fluorescence). Cx37-target-siRNA treated cells build capillaries with a small number of branches. Knock-down of the single connexins results ininhibition of network formation as seen by small number of branches. Greatest effects were seen by knock-down of Cx43 > Cx40 > Cx37. Knock-down of Cx43 and Cx40 alsoc illarieu nts foi is figu
t2tIososnpbmcao
hange overall pattern and the ratio of number of branches/average length of capnder target siRNA-treatment. All data are given as mean ± S.E.M. of n = 6 experime
ndicated by an asterisk (p < 0.05). (For interpretation of the references to color in th
hat the biggest effects of RNA interference were observed after4 h exposure of the cells. For Cx43, Cx40 and Cx37 the most effec-ive concentrations of siRNA varied between 10 nM and 20 nM.n subsequent experiments we used a siRNA transfection timef 24 h with 20 nM siRNA for Cx37 and Cx40 and with 10 nMiRNA for Cx43 knock-down, followed by a 3D growth phasef 18 h in the angiogenesis assay. Knock-down with scramblediRNA did affect neither the Cx expression (Fig. 1A and Table 2)or the investigated angiogenic parameters significantly as com-ared to treatment with NFS only. Average length of capillaryranches and pattern were 100.8 ± 7.5% and 102.7 ± 11.7% vs. treat-
ent with NFS only; whereas number of branches, number ofells and branches per length ratio were 94.8 ± 5.8%, 95.4 ± 12.5%nd 94.7 ± 7.1% respectively related to treatment with NFSnly.
s. Scale bar = 50 �m. (E) Quantitative and semi quantitative evaluation of changesr each condition. Significant differences between siRNA- and control-treatment are
re legend, the reader is referred to the web version of the article.)
3.2. Dye coupling after siRNA treatment of HUVECs
To test functional impairment of gap junction communicationafter 24 h treatment of cells with siRNA against Cx43, Cx40 or Cx37we investigated the coupling of adjacent cells by dye transfer withLucifer Yellow. We could see a significant decrease of intercellularcommunication after knock-down of Cx43, Cx40 or Cx37 as com-pared to cells treated with scrambled-siRNA (p < 0.05, Fig. 2). Thehighest effect was seen for knock-down of Cx43 (Fig. 2D), where thenumber of communicating cells declined from 7.1 ± 1.3 for treat-ment with scrambled-siRNA to 2.3 ± 0.6.
3.3. Effect of Cx37-knock-down on angiogenesis
Western blot showed that after 24 h treatment with 20 nM tar-get siRNA Cx37 was reduced down to 38.9 ± 1.1% related to control.

C. Gärtner et al. / Pharmacological Research 65 (2012) 347– 357 353
Fig. 5. Effect of palmitoleic acid and nicotine on angiogenesis. (A)–(C) In vitro angiogenesis assay. Matrigel-assay was performed in presence of the gap junction blockerpalmitoleic acid (B) or HUVECs were grown in the presence of 1 �M nicotine (C) or vehicle (A) for 5 days. Cells are stained with lectin (FITC-conjugated, green fluorescence)and nuclei are stained with Hoechst 33258 (blue fluorescence). Treatment with palmitoleic acid (B) resulted in attenuated angiogenesis seen by a lower number of branches,a worse overall pattern and a reduced branches/average length of capillaries ratio as related to control treatment (A; Ctrl). Nicotine treated cells form a less complex networkw lei peo ven as( der is
Taa(
tnecais
3
Ci6ets
B
ith fewer branches and shorter capillaries, but in contrast a higher number of nucf changes under palmitoleic acid (n = 3) and nicotine treatment (n = 11). Data are gip < 0.05). (For interpretation of the references to color in this figure legend, the rea
he other connexins, i.e. Cx40 and Cx43, were not significantlyltered (Table 2, Fig. 1). Also qRT-PCR experiments showed thatmount of Cx37-mRNA was reduced to 46 ± 20.8% of control valuesFig. 1B).
As shown in Figs. 3B and 4B, reduced abundance of Cx37 pro-ein after Cx37-siRNA treatment resulted in significantly decreasedumber of branches (75.4 ± 13.9% of control values, p < 0.05). How-ver, average length of the capillary branches and the number ofells were not significantly affected. In consequence, the patternnd the ratio of the number of branches to the average length of cap-llary branches was also slightly diminished, but without reachingtatistical significance (Fig. 4E).
.4. Influence of Cx40-knock-down on angiogenesis
Western blot showed that siRNA-treatment maximally reducedx40 down to 31.3 ± 0.2% of control value. The other connexins,
.e. Cx37 and Cx43, were diminished (Cx37 86.1 ± 1.7% and Cx436.5 ± 2.6% vs. control treatment; Table 2 and Fig. 1A). qRT-PCRxperiments showed that also amount of Cx40-mRNA was reduced
o 21.9 ± 8.4% after siRNA knock-down compared to treatment withcrambled-siRNA (Fig. 1B).In the Matrigel-assay an impairment of angiogenesis was seen.oth, number of branches as well as the ratio of number of
r picture (C). Scale bar = 50 �m. (D) Quantitative and semi- quantitative evaluation mean ± S.E.M. Significant changes to control treatment are indicated by an asteriskreferred to the web version of the article.)
branches to their average length were significantly diminishedas shown in Figs. 3C and 4C. While average length of the capil-lary branches and number of cells were not affected, the patternwas also significantly reduced (80 ± 5.4% of control values, p < 0.05;Fig. 4E).
3.5. Effect of Cx43-knock-down on angiogenesis
Treatment with siRNA resulted in a maximum Cx43-protein-knock-down to 22.4 ± 10.3% of control values. The other connexins,i.e. Cx37 and Cx40, remained nearly unaltered (Cx37 90.5 ± 1.04%and Cx40 88.8 ± 2.4% vs. control; Fig. 1A and Table 2). In qRT-PCR experiments we could see a reduction of Cx43-mRNA to39.4 ± 22.7% after treatment with siRNA against Cx43-mRNA(Fig. 1B).
The effects of Cx43-protein-knock-down on angiogenesis wereeven more pronounced than the effects of Cx37- or Cx40-protein-knock-down (Figs. 3D and 4D). Number of branches was decreasedto 41 ± 5.7% of control values (p < 0.05; Fig. 4E). Both, the pattern
and ratio of branches to the average length of capillary brancheswere also significantly reduced as compared to controls (p < 0.05).However, there were no significant differences in the average cap-illary branches length or number of cells.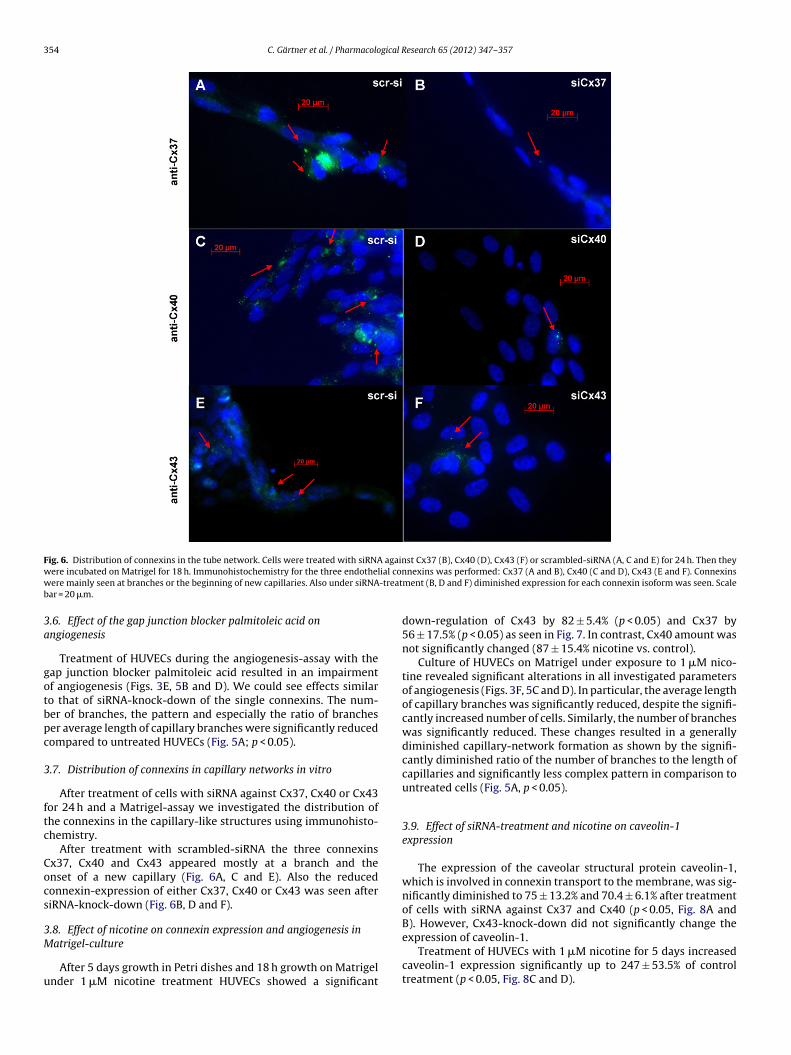
354 C. Gärtner et al. / Pharmacological Research 65 (2012) 347– 357
Fig. 6. Distribution of connexins in the tube network. Cells were treated with siRNA against Cx37 (B), Cx40 (D), Cx43 (F) or scrambled-siRNA (A, C and E) for 24 h. Then theyw ial conw -treatb
3a
gotbpc
3
ftc
Cocs
3M
u
ere incubated on Matrigel for 18 h. Immunohistochemistry for the three endothelere mainly seen at branches or the beginning of new capillaries. Also under siRNA
ar = 20 �m.
.6. Effect of the gap junction blocker palmitoleic acid onngiogenesis
Treatment of HUVECs during the angiogenesis-assay with theap junction blocker palmitoleic acid resulted in an impairmentf angiogenesis (Figs. 3E, 5B and D). We could see effects similaro that of siRNA-knock-down of the single connexins. The num-er of branches, the pattern and especially the ratio of brancheser average length of capillary branches were significantly reducedompared to untreated HUVECs (Fig. 5A; p < 0.05).
.7. Distribution of connexins in capillary networks in vitro
After treatment of cells with siRNA against Cx37, Cx40 or Cx43or 24 h and a Matrigel-assay we investigated the distribution ofhe connexins in the capillary-like structures using immunohisto-hemistry.
After treatment with scrambled-siRNA the three connexinsx37, Cx40 and Cx43 appeared mostly at a branch and thenset of a new capillary (Fig. 6A, C and E). Also the reducedonnexin-expression of either Cx37, Cx40 or Cx43 was seen afteriRNA-knock-down (Fig. 6B, D and F).
.8. Effect of nicotine on connexin expression and angiogenesis in
atrigel-cultureAfter 5 days growth in Petri dishes and 18 h growth on Matrigelnder 1 �M nicotine treatment HUVECs showed a significant
nexins was performed: Cx37 (A and B), Cx40 (C and D), Cx43 (E and F). Connexinsment (B, D and F) diminished expression for each connexin isoform was seen. Scale
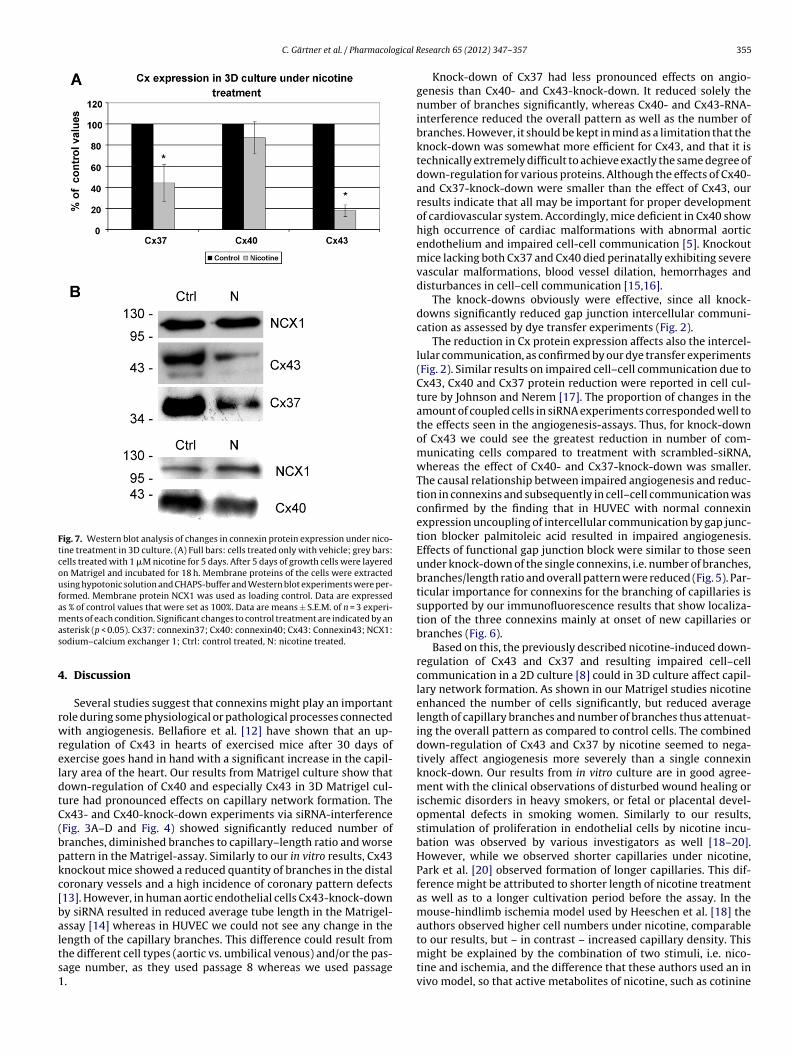
down-regulation of Cx43 by 82 ± 5.4% (p < 0.05) and Cx37 by56 ± 17.5% (p < 0.05) as seen in Fig. 7. In contrast, Cx40 amount wasnot significantly changed (87 ± 15.4% nicotine vs. control).
Culture of HUVECs on Matrigel under exposure to 1 �M nico-tine revealed significant alterations in all investigated parametersof angiogenesis (Figs. 3F, 5C and D). In particular, the average lengthof capillary branches was significantly reduced, despite the signifi-cantly increased number of cells. Similarly, the number of brancheswas significantly reduced. These changes resulted in a generallydiminished capillary-network formation as shown by the signifi-cantly diminished ratio of the number of branches to the length ofcapillaries and significantly less complex pattern in comparison tountreated cells (Fig. 5A, p < 0.05).
3.9. Effect of siRNA-treatment and nicotine on caveolin-1expression
The expression of the caveolar structural protein caveolin-1,which is involved in connexin transport to the membrane, was sig-nificantly diminished to 75 ± 13.2% and 70.4 ± 6.1% after treatmentof cells with siRNA against Cx37 and Cx40 (p < 0.05, Fig. 8A andB). However, Cx43-knock-down did not significantly change the
expression of caveolin-1.Treatment of HUVECs with 1 �M nicotine for 5 days increasedcaveolin-1 expression significantly up to 247 ± 53.5% of controltreatment (p < 0.05, Fig. 8C and D).
C. Gärtner et al. / Pharmacological R
Fig. 7. Western blot analysis of changes in connexin protein expression under nico-tine treatment in 3D culture. (A) Full bars: cells treated only with vehicle; grey bars:cells treated with 1 �M nicotine for 5 days. After 5 days of growth cells were layeredon Matrigel and incubated for 18 h. Membrane proteins of the cells were extractedusing hypotonic solution and CHAPS-buffer and Western blot experiments were per-formed. Membrane protein NCX1 was used as loading control. Data are expressedas % of control values that were set as 100%. Data are means ± S.E.M. of n = 3 experi-mas
4
rwreldtC(bpkc[balts1
ents of each condition. Significant changes to control treatment are indicated by ansterisk (p < 0.05). Cx37: connexin37; Cx40: connexin40; Cx43: Connexin43; NCX1:odium–calcium exchanger 1; Ctrl: control treated, N: nicotine treated.
. Discussion
Several studies suggest that connexins might play an importantole during some physiological or pathological processes connectedith angiogenesis. Bellafiore et al. [12] have shown that an up-
egulation of Cx43 in hearts of exercised mice after 30 days ofxercise goes hand in hand with a significant increase in the capil-ary area of the heart. Our results from Matrigel culture show thatown-regulation of Cx40 and especially Cx43 in 3D Matrigel cul-ure had pronounced effects on capillary network formation. Thex43- and Cx40-knock-down experiments via siRNA-interferenceFig. 3A–D and Fig. 4) showed significantly reduced number ofranches, diminished branches to capillary–length ratio and worseattern in the Matrigel-assay. Similarly to our in vitro results, Cx43nockout mice showed a reduced quantity of branches in the distaloronary vessels and a high incidence of coronary pattern defects13]. However, in human aortic endothelial cells Cx43-knock-downy siRNA resulted in reduced average tube length in the Matrigel-ssay [14] whereas in HUVEC we could not see any change in the
ength of the capillary branches. This difference could result fromhe different cell types (aortic vs. umbilical venous) and/or the pas-age number, as they used passage 8 whereas we used passage.esearch 65 (2012) 347– 357 355
Knock-down of Cx37 had less pronounced effects on angio-genesis than Cx40- and Cx43-knock-down. It reduced solely thenumber of branches significantly, whereas Cx40- and Cx43-RNA-interference reduced the overall pattern as well as the number ofbranches. However, it should be kept in mind as a limitation that theknock-down was somewhat more efficient for Cx43, and that it istechnically extremely difficult to achieve exactly the same degree ofdown-regulation for various proteins. Although the effects of Cx40-and Cx37-knock-down were smaller than the effect of Cx43, ourresults indicate that all may be important for proper developmentof cardiovascular system. Accordingly, mice deficient in Cx40 showhigh occurrence of cardiac malformations with abnormal aorticendothelium and impaired cell-cell communication [5]. Knockoutmice lacking both Cx37 and Cx40 died perinatally exhibiting severevascular malformations, blood vessel dilation, hemorrhages anddisturbances in cell–cell communication [15,16].
The knock-downs obviously were effective, since all knock-downs significantly reduced gap junction intercellular communi-cation as assessed by dye transfer experiments (Fig. 2).
The reduction in Cx protein expression affects also the intercel-lular communication, as confirmed by our dye transfer experiments(Fig. 2). Similar results on impaired cell–cell communication due toCx43, Cx40 and Cx37 protein reduction were reported in cell cul-ture by Johnson and Nerem [17]. The proportion of changes in theamount of coupled cells in siRNA experiments corresponded well tothe effects seen in the angiogenesis-assays. Thus, for knock-downof Cx43 we could see the greatest reduction in number of com-municating cells compared to treatment with scrambled-siRNA,whereas the effect of Cx40- and Cx37-knock-down was smaller.The causal relationship between impaired angiogenesis and reduc-tion in connexins and subsequently in cell–cell communication wasconfirmed by the finding that in HUVEC with normal connexinexpression uncoupling of intercellular communication by gap junc-tion blocker palmitoleic acid resulted in impaired angiogenesis.Effects of functional gap junction block were similar to those seenunder knock-down of the single connexins, i.e. number of branches,branches/length ratio and overall pattern were reduced (Fig. 5). Par-ticular importance for connexins for the branching of capillaries issupported by our immunofluorescence results that show localiza-tion of the three connexins mainly at onset of new capillaries orbranches (Fig. 6).
Based on this, the previously described nicotine-induced down-regulation of Cx43 and Cx37 and resulting impaired cell–cellcommunication in a 2D culture [8] could in 3D culture affect capil-lary network formation. As shown in our Matrigel studies nicotineenhanced the number of cells significantly, but reduced averagelength of capillary branches and number of branches thus attenuat-ing the overall pattern as compared to control cells. The combineddown-regulation of Cx43 and Cx37 by nicotine seemed to nega-tively affect angiogenesis more severely than a single connexinknock-down. Our results from in vitro culture are in good agree-ment with the clinical observations of disturbed wound healing orischemic disorders in heavy smokers, or fetal or placental devel-opmental defects in smoking women. Similarly to our results,stimulation of proliferation in endothelial cells by nicotine incu-bation was observed by various investigators as well [18–20].However, while we observed shorter capillaries under nicotine,Park et al. [20] observed formation of longer capillaries. This dif-ference might be attributed to shorter length of nicotine treatmentas well as to a longer cultivation period before the assay. In themouse-hindlimb ischemia model used by Heeschen et al. [18] theauthors observed higher cell numbers under nicotine, comparable
to our results, but – in contrast – increased capillary density. Thismight be explained by the combination of two stimuli, i.e. nico-tine and ischemia, and the difference that these authors used an invivo model, so that active metabolites of nicotine, such as cotinine
356 C. Gärtner et al. / Pharmacological Research 65 (2012) 347– 357
Fig. 8. Western blot analysis of changes in caveolin-1 (Cav-1) protein expression under treatment with siRNA or nicotine. (A) and (B) HUVECs were treated with target-siRNAfor Cx37, Cx40, Cx43 or with scrambled-siRNA for 24 h. After 18 h additional growth without siRNA proteins were extracted using RIPA-buffer and Western blots wereperformed. Knock-down of Cx37 (B, panel 1) and Cx40 (B, panel 2) significantly reduced amount of Cav-1 related to scrambled-siRNA (B, panel4), whereas knock-down ofCx43 did not alter Cav-1 expression (B, panel 3). (C) and (D) Cells grown for 5 days in presence of 1 �M nicotine showed significantly enhanced Cav-1 expression. Data aree expeC Ctrl: v
(roa
iwsaifiaaaaiiTeapa
tiotA
xpressed as % of control values that were set as 100%. Data are means ± S.E.M. of fivex37: connexin37; Cx40: connexin40; Cx43: connexin43; scr-si: scrambled-siRNA,
which can act with higher affinity on certain, so-called cotinineeceptors [21]), might have contributed as well. In further supportf our findings, enhanced proliferation after Cx43 knock-down haslso been observed in cardiomyocytes [22].
To further investigate the mechanisms that may contribute tompaired angiogenesis seen under nicotine- and siRNA treatment
e analyzed the caveolar structural protein caveolin-1. It has beenhown to co-localize and interact with Cx43 in the Golgi apparatusnd in the plasma membrane in lipid rafts [23] and with Cx37, Cx40n endothelial cells [24]. Also cav-1 is shown to be involved in traf-cking of Cx43 to the plasma membrane [25]. Koval et al. [26] found
decrease of cav-1 Cx40- and Cx43-knockout-mice. We could find down-regulation of cav-1 after transfection of cells with siRNAgainst either Cx40 or Cx37, while down-regulation of both Cx43nd Cx37 after nicotine treatment leads to a significant increasen cav-1 expression. These results confirm the thesis, that cav-1s involved in trafficking of connexins to the plasma membrane.he increased expression under nicotine treatment could be due tonhanced degradation of Cx43 and Cx37 under nicotine treatment,s suggested by Tsai et al. [9], so that the increase in cav-1 could beart of a counter regulatory process. In contrast, down-regulationfter siRNA treatment might indicate reduced demand for cav-1.
In summary, in this study we – to the best of our knowledge – forhe first time show the effects of knock-down of the single connex-
ns on in vitro angiogenesis. Our data suggest that down-regulationf Cx and impaired intercellular communication are relevant fac-ors influencing in vitro angiogenesis due to nicotine treatment.mong the three connexins types in endothelial cells Cx43 seems toriments of each condition, significant changes are indicated by an asterisk (p < 0.05).ehicle treated, N: nicotine treated.
play the major role in angiogenesis. We could see that knock-downof Cx43 had a more pronounced effect on the number of branches,the formation of the capillary-like network and most of all on therelation of branches to the capillary length than reduction in Cx37or Cx40.
Thus, we conclude from our data, that the three endothelialconnexins Cx37, Cx40 and Cx43 all participate in the angiogenicprocesses, regulating primarily the branching of the capillaries, andthat the effects of nicotine on angiogenesis at least in parts involveconnexin-down-regulation.
Disclosure
There is no conflict of interest to declare.
Acknowledgment
This paper was generously supported by grants to ChristianeGärtner given by ProCordis (Leipzig, Germany) and Konrad Ade-nauer Stiftung (Sankt Augustin, Germany).
References
[1] Van Rijen H, van Kempen MJ, Analbers LJ, Rook MB, van Ginneken AC, Gros D,
et al. Gap junctions in human umbilical cord endothelial cells contain multipleconnexins. Am J Physiol 1997;272:C117–30.[2] Zhang YW, Nakayama K, Nakayama K, Morita I. A novel route for connexin 43to inhibit cell proliferation: negative regulation of S-phase kinase-associatedprotein (Skp 2). Cancer Res 2003;63:1623–30.

gical R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[regulation of gap-junctions in endothelial cells. In: Proceedings of the interna-
C. Gärtner et al. / Pharmacolo
[3] Burt JM, Nelson TK, Simon AM, Fang JS. Connexin 37 profoundly slowscell cycle progression in rat insulinoma cells. Am J Physiol Cell Physiol2008;295:C1103–12.
[4] Gu H, Smith FC, Taffet SM, Delmar M. High incidence of cardiac malformationsin connexin40-deficient mice. Circ Res 2003;93:201–6.
[5] Simon AM, McWhorter AR. Vascular abnormalities in mice lacking theendothelial gap junction proteins connexin37 and connexin40. Dev Biol2002;251:206–20.
[6] Krüger O, Maxeiner S, Kim JS, van Rijen HV, de Bakker JM, Eckardt D, et al.Cardiac morphogenetic defects and conduction abnormalities in mice homozy-gously deficient for connexin40 and heterozygously deficient for connexin45.J Mol Cell Cardiol 2006;41:787–97.
[7] De Wit C, Roos F, Bolz SS, Pohl U. Lack of vascular connexin 40 is associ-ated with hypertension and irregular arteriolar vasomotion. Physiol Genomics2003;13:169–77.
[8] Haussig S, Schubert A, Mohr FW, Dhein S. Sub-chronic nicotine expo-sure induces intercellular communication failure and differential down-regulation of connexins in cultured human endothelial cells. Atherosclerosis2008;196:210–8.
[9] Tsai CH, Yeh IH, Tian TY, Lee YN, Lu CS, Ko YS. Down-regulating effect of nico-tine on connexin43 gap junctions in human umbilical vein endothelial cells isattenuated by statins. Eur J Cell Biol 2004;82:589–95.
10] Dhein S, Krüsemann K, Schaefer T. Effects of the gap junction uncoupler palmi-toleic acid on the activation and repolarization wavefronts in isolated rabbithearts. Br J Pharmacol 1999;128:1375–84.
11] Livak KJ, Schmittgen TD. Analysis of relative gene expression data usingreal-time quantitative PCR and the 2(−Delta Delta C(T)) method. Methods2001;25:402–8.
12] Bellafiore M, Sivverini G, Palumbo D, Macaluso F, Bianco A, Palma A, et al.Increased Cx43 and angiogenesis in exercised mouse hearts. Int J Sports Med2007;28:749–55.
13] Walker DL, Vacha SJ, Kirby ML, Lo CW. Connexin43 deficiency causes dysregu-lation of coronary vasculogenesis. Dev Biol 2005;284:479–98.
14] Wang HH, Kung CI, Tseng YY, Lin YC, Chen CH, Tsai CH, et al. Activation ofendothelial cells to pathological status by down-regulation of connexin43.Cardiovasc Res 2008;79:509–18.
[
esearch 65 (2012) 347– 357 357
15] Simon AM, McWhorter AR. Role of connexin37 and connexin40 in vasculardevelopment. Cell Commun Adhes 2003;10:379–85.
16] Simon AM, McWhorter AR. Decreased intercellular dye-transfer and down-regulation on non-ablated connexins in aortic endothelium deficient inconnexin37 and sonnexin40. J Cell Sci 2003;116:2223–36.
17] Johnson TL, Nerem RM. Endothelial connexin 37, connexin 40, and connexin43 respond uniquely to substrate and shear stress. Endothelium 2007;14:215–26.
18] Heeschen C, Jang JJ, Weis M, Pathak A, Kaji S, Hu RS, et al. Nicotine stimu-lates angiogenesis and promotes tumor growth and atherosclerosis. Nat Med2001;7:833–9.
19] Mousa S, Mousa SA. Cellular and molecular mechanisms of nicotine’s pro-angiogenesis activity and its potential impact on cancer. J Cell Biochem2006;97:1370–8.
20] Park YJ, Lee T, Ha J, Jung IM, Chung JK, Kim SJ. Effect of nicotine on humanumbilical vein endothelial cells (HUVECs) migration and angiogenesis. VasculPharmacol 2008;49:32–6.
21] Riah O, Dousset JC, Bofill-Cardona E, Corrirère P. Isolation and microsequencingof a novel cotinine receptor. Cell Mol Neurobiol 2000;20:653–64.
22] Matsuyama D, Kawahara K. Proliferation of neonatal cardiomyocytes by con-nexin43 knockdown via synergistic inactivation of p38 MAPK and increasedexpression of FGF1. Basic Res Cardiol 2009;104:631–42.
23] Langlois S, Cowan KN, Shao Q, Cowan BJ, Laird DW. Caveolin-1 and -2 interactwith connexin43 and regulate gap junctional intercellular communication inkeratinocytes. Mol Biol Cell 2008;19(3):912–28.
24] Saliez J, Bouzin C, Rath G, Ghisdal P, Desjardins F, Rezzani R, et al. Role of caveolarcompartmentation in endothelium-derived hyperpolarizing factor-mediatedrelaxation: Ca2+ signals and gap junction function are regulated by caveolin inendothelial cells. Circulation 2008;117:1065–74.
25] Behets Wydemans G, Rath G, Bouzin C, Dessy C. Caveolin-1 participates to the
tional gap junction conference. 2011. p. 167.26] Koval M, Billaud M, Straub AC, Johnstone SR, Zarbock A, Duling BR, et al.
Spontaneous lung dysfunction and fibrosis in mice lacking connexin 40 andendothelial cell connexin 43. Am J Pathol 2011;178:2536–46.





























