Embed Size (px)
Citation preview

Kloning av möjlig promotorsekvens uppströms kloritdismutas i Ideonella dechloratans.
Cloning of a possible promoter sequence upstream of chlorite dismutase in
Ideonella dechloratans.
Lisa Ljungberg
Faculty of Technology and Science
Biovetenkapligt program, Biokemi
Kandidatuppsats i Kemi 15hp
Maria Rova
Jörgen Samuelsson
26 mars, 2014
1

Lisa Ljungberg
2
2
Sammanfattning
Perklorat och klorater finns naturligt i atmosfären, varifrån det sedimenterar ner i grundvatten
och jord. Utsläpp av klorat och perklorat ifrån jordbruket och pappersbruk leder också till
ökade halter i vår miljö. Dessa föroreningarna kan renas med bakterier vilka kan reducera
perklorat och klorat till syrgas och kloridjoner. Denna reduktion fungerar dock bäst i en
anaerob, syrefri miljö. Med en utökad kunskap om generna, vilka är ansvariga för regleringen
och uttrycket av reducerande enzymer kan man modifiera dessa att fungera lika bra i en
aerob, syrerik miljö. Ett av dessa enzymer är kloritdismutas. Uppströms genen för
kloritdismutas i bakterien Ideonella dechloratans finns en möjlig promotorsekvens. Syftet
med detta arbete var att korta ner denna DNA-sekvens för att komma närmare inpå möjliga
reglerande sekvenser och inbindningssäte för RNA-polymeras. Därefter kontrollera om
sekvensen var en aktiv promotor. Den möjliga promotor region kortades ner och
amplifierades med PCR. Sedan ligerades den in i plasmiden pRU1103, som bär på en reporter
gen. Konstruktionen klonades i Escherichia coli och visade sig kunna bilda produkten av
reportergenen. Detta tyder på att sekvensen innehöll en fungerande promotor.

Lisa Ljungberg
3
3
Abstract
Perchlorate and chlorate are naturally occurring in the atmosphere, from here it sediments
into groundwater and soil. The pollution is increased by discharges of perchlorate and
chlorate from agriculture and paper mills. Bacteria capable of reducing perchlorate and
chlorate to chloride and oxygen can be used to get rid of these contaminants. However an
anaerobic environment needs to be sustained in order for this reaction to be used. For this
reduction to work in an aerobic environment as well, a greater knowledge of the reducing
enzymes, regulating factors and their corresponding genes is needed. One of these enzymes is
called chlorite dismutase. The upstream region of this gene in Ideonella dechloratans bacteria
is likely to contain a promoter sequence. The purpose of this work was to shorten this DNA
sequence to get closer to the potential regulatory sequences and binding-site for RNA
polymerase. Subsequently control if the sequence was active as an promoter. The potential
promoter region was shortened and amplified by PCR. It was then ligated into the plasmid
pRU1103which is carrying a reporter gene. Cloning of the construction into Escherichia coli
resulted in a production of the reporter gene product. This indicates that the sequence
contained a working promoter.

Lisa Ljungberg
4
4
Innehållsförteckning
SAMMANFATTNING ........................................................................................................................... 2
ABSTRACT ............................................................................................................................................ 3
FÖRKORTNINGAR .............................................................................................................................. 4
1. INLEDNING....................................................................................................................................... 6
2. MATERIAL OCH METODER ........................................................................................................ 10
2. 1 PRIMERDESIGN ............................................................................................................................................ 10 2. 2 PLASMIDPREPARATION .................................................................................................................................. 11 2. 3 RESTRIKTIONSKLYVNING ................................................................................................................................ 11 2. 4 RENING AV KLUVEN PLASMID .......................................................................................................................... 11 2. 5 PCR .......................................................................................................................................................... 12 2. 6 LIGERING .................................................................................................................................................... 13 2. 7 ELEKTROKOMPETENTA CELLER ......................................................................................................................... 13 2. 8 TRANSFORMATION OCH ELEKTROPORERING ........................................................................................................ 13
3. RESULTAT OCH DISKUSSION .................................................................................................... 14
3.1 PRIMERDESIGN ............................................................................................................................................. 14 3. 2 PLASMIDPREPARATION .................................................................................................................................. 15 3. 3 RESTRIKTIONSKLYVNING ................................................................................................................................ 15 3. 4 RENING AV KLUVEN PLASMID .......................................................................................................................... 18 3. 5 PCR OCH TRANSFORMATION .......................................................................................................................... 19
SAMMANFATTANDE SLUTSATS ................................................................................................... 24
REFERENSER ..................................................................................................................................... 25
APPENDIX I ........................................................................................................................................ 27
APPENDIX II ....................................................................................................................................... 28
APPENDIX III ...................................................................................................................................... 29

Lisa Ljungberg
5
5
Förkortningar
PCR - Polymerase chain reaction
MCS - Multiple cloning site
X-gal - 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside
Ö. N – Över natt
PRB - Perklorat reducerande bakterier
CRB - Klorat reducerande bakterier
FNR - Fumarat och nitrat reduktas
bp - baspar
cfu - colony-forming unit

Lisa Ljungberg
6
6
1. Inledning
Perklorat och klorat är kemiskt stabila i vattenmiljöer och extremt lösliga. Perkloraters stora
spridning på jorden kan tyda på en naturlig källa, detta har likaså hittats i grundvatten och
ökenmark [1]. Möjligen genereras perklorat och klorat i atmosfären, sedan lagras det i jord
och grundvatten. Det faktum att många bakterier har kapaciteten att metabolisera klorater och
leva i en kloratrik miljö kan förklaras av att en naturlig källa finns [2]. Klorat och perklorat är
starka oxidationsmedel och kan därför ha en mycket negativ inverkan på biologiska system.
Perklorat och klorat är farligt för både djur och växter, exempelvis kan perklorat påverka
upptaget av jod i sköldkörteln (tyreoidea), jod är essentiellt för att bilda sköldkörtelhormoner.
Perklorat binder till natrium-jod symportern och inhiberar på så vis upptaget av jod till
sköldkörteln. Detta kan leda till sköldkörtelhormonbrist, hypotyreos [3]. Då
sköldkörtelhormoner styr metabolismen kan en felfunktion leda till defekter i tidig utveckling
hos barn och foster [4]. Växter som utsatts för klorater fick en förändring i tillväxten,
speciellt hos brunalger. Antalet alger minskade kraftigt och i några miljöer slutade algerna att
växa [2,5,6]. Klorat verkar dock vara mer toxiskt för brunalgerna jämfört med många andra
marina växter som saknar enzymet nitratreduktas. Detta kan bero på kloratets strukturella
likhet med nitrat och att det nitratreducerande enzymet nitratreduktas reducerar klorat till dess
toxiska form klorit i brunalgerna[2,7]. Trots att det finns belägg för en naturlig källa kan
klorater i vatten och jord bäst förklaras genom mänsklig påverkan. Klorater har producerats
industriellt under många år, då dessa har använts inom jordbruket som ett ogräs- eller
avlövningsmedel. Klorat bildas också som en oönskad biprodukt vid vattendesinfektion och
ifrån cellulosablekning av pappersmassa. Trots detta finns för närvarande inga regleringar av
klorathalt i drickvatten [8]. Kloraters höga löslighet och mobilitet leder till att de traditionella
teknikerna för behandling av avloppsvatten inte är effektiva nog. Biologisk rening har ansetts
vara den bästa lösningen då det betraktas vara effektivt och ekonomiskt gynnsamt [1].
Denna biologiska rening tar hjälp av mikrobiell nedbrytning av perklorat eller klorat genom
bakteriers respiration. Reduktion av klorat (ClO3-) och perklorat (ClO4
-) till kloritjoner (ClO2
-
) är en tvåstegsreaktion där perklorat först reduceras till klorat vilket i sin tur reduceras till
klorit. Klorit sönderdelas därefter till kloridjoner och syrgas (O2). Syret kan sedan användas
som respiratorisk elektronacceptor [9].

Lisa Ljungberg
7
7
→
→
→
Nedbrytningen av perklorat och klorat sker med tre olika enzymer, perkloratreduktas,
kloratreduktas och kloritdismutas. Den ena gruppen bakterier är kapabla att reducera
perklorat eller klorat i elektrontransportkedjan för cellandningen. Dessa benämns
perkloratreducerande bakterier (PRB). Den andra gruppen kallas kloratreducerande bakterier
(CRB) och dessa bakterier har enzymet kloratreduktas. Detta gör att de kan reducera klorat
men inte perklorat [10]. Gemensamt för de båda är kloritdismutas, vilket gör detta till ett
protein av intresse. För att en klorat- och perkloratreducerande reaktionsväg ska utnyttjas
krävs idag en anaerob miljö. Med en ökad kunskap om hur perklorat- och kloratreduktionen
regleras kan möjliga regulatorer manipuleras. På så vis kan bakterierna eventuellt utnyttja
denna reaktionsväg även under aeroba förhållanden. Detta skulle leda till minskade kostnader
och en ökad effektivitet [9].
Ideonella dechloratans är en gramnegativ och stavformad bakterie. Den kan inte bryta ner
perklorat men den är däremot en CRB och har förmågan att använda klorat som
elektronacceptor i andningskedjan vid frånvaro av syre. Bakterien är fakultativt anaerob. Om
syre finns tillgängligt används detta som elektronacceptor men vid syrebrist kan en alternativ
källa utnyttjas som klorat eller nitrat. Dock förlorar I. dechloratans förmågan att använda
nitrat i elektrontransportkedjan efter ett par subkultiveringar anaerobt på klorat [11]. Det har
isolerats omkring 50 arter som tillsammans med Ideonella dechloratans kan reducera
perklorat eller klorat. Reduktionen sker i periplasman, utrymmet mellan yttermembran och
cellmembran. Det är också här de två kloratnedbrytande enzymerna finns. Kloratreduktas
består av tre subenheter vilka kodas av generna clr A, clr B, clr C. Till subenheterna finns
även av ett specifikt chaperon som kodas av genen clr D. Kloritdismutas kodas enbart av
genen cld. Uppströms om clr A genen i Ideonella dechloratans ligger en möjlig promotor
sekvens (Figur 1). Promotor sekvenser är specifika sekvenser i DNA-molekylen till vilka
RNA-polymeras binder in. På så sätt leds transkriptionen av DNA till intilliggande
gensekvenser. I Escherichia coli binder RNA-polymeraset till en region ungefär 70 baspar
innan transkriptionsstart till 30 baspar efter. De baspar i DNA molekylen som korresponderar
till de första basparen i RNA molekylen får positiva nummer, medans de som kommer innan
RNA start sätet får negativa nummer. Promotorn sträcker sig alltså mellan -70 och +30.
Genom analys och jämförelse av promotor regioner i vanliga bakteriestammar har man sett

Lisa Ljungberg
8
8
likheter i två korta sekvenser i positionerna -10 och -35. Konsensus sekvensen vid -10 är (5’)
TATAAT (3’) och vid -35 (5’) TTGACA (3’).
Transkriptionen av en gen regleras noggrant i cellen för att endast bilda proteiner i nödvändig
mängd. Dessa regleringar sker främst vid RNA-polymeras inbindning och transkriptions
initiering, för att spara energi. Proteiner kan binda till regioner både nära och långt ifrån
promotorn och på så sätt påverka genuttrycket. Ett repressor protein hindrar RNA-
polymerasets rörelse igenom eller inbindning till promotorn genom blockering. Repressorn
binder till specifika platser i DNA molekylen, så kallad operator. Operatorn sitter generellt
nära promotorn. Andra transkriptionsfaktorer kan öka affiniteten eller specificiteten för RNA-
polymeraset. En aktivator är ett protein som ökar affiniteten till RNA-polymeraset och därför
också ökar transkriptionen av en eller flera gener [12]. Det är visat att transkriptionen av cld
ökar under anaeroba förhållanden [13], detta kan vara en effekt av transkriptions reglerande
sekvenser.
Escherichia coli har flera sensormekanismer som ger svar på syretillgångar och närvaro av
olika elektronacceptor. Dessa regleringar samordnas av en grupp globala regulatorer vilka
inkluderar FNR-proteinet (fumarat och nitrat reduktas) och två-komponent arcAB systemet.
ArcA, en transkiptionell regulator och arcB, ett sensorkinas. ArcA fosforyleras av arcB vid
anaeroba förhållanden. Det aktiverade arcA reglerar uttrycket av ett flertal operon,
genkluster, involverade i respiratoriska och fermentativa metabolismer [14,15]. I sekvensen
uppströms om cld finns en möjlig arcA bindande sekvens (Figur 1) och även en FNR-
liknande sekvens (figur 1). FNR-proteinet hos E.coli är en syrekänslig regulator och aktivator
som krävs för att kunna byta ifrån aerob till anaerob metabolism. FNR proteinet binder till
den återkommande palindrom liknande sekvensen TTGA(T/C)NNNN(A/G)TCAA vilken
ofta är centrerad vid -40 [15]. FNR kontrollerar alltså transkriptionen genom att binda till
promotorer under anaeroba förhållanden. FNR homologer har identifierats i många andra
bakterier, där dess roll också verkar vara reglering av genuttryck som svar på syre
förhållanden [16,17].

Lisa Ljungberg
9
9
5'GCGAGGTCGGCGTCAGTGTCGGGCGGTTGTCCCCGAGGGTCATGTTTGTTGAGAAGTCAATTTCC
ACTGGAACGGCATAGAGCCAAAAATATAGTGTTAAGTTTCGGAGAAACTCTTGACTTAAATCAAA
CACATAAAAAAGGAGAAATGCGAAGATTCGTGCACTTTCCGCTTGCTCGGCGACC AAGCCGCCCA
GC 3'
Experimentell Transkriptionsstart Möjlig FNR-Box Möjliga -10 och -35 sekvenser Möjlig arcA bindande sekvens
I Ideonella dechloratans har även en ArsR-gen hittats i genklustret för kloratmetabolism.
Möjligen kan denna gen vara involverad i den ökade transkriptionen av cld eller clrA under
anaeroba förhållanden [13]. Sekvensen i Ideonella dechloratans (understruket i figur 1) har
likheter med DNA-inbindningssätet för ArsR-regulatorn Rv2034, i Mycobacterium
tuberculosis. Rv2034 visades här reglera uttrycket av dosR genen, vilken ger hypoxi, istället
för att agera som metallsensor, vilket ArsR främst är förknippat med [18,19]. Dessa
sekvenser kan ha en koppling till kloratmetabolismen i anaeroba förhållanden hos Ideonella
dechloratans.
Ovanstående sekvens, Figur 1, har tidigare klonats i Escherichia coli. Under dessa försök
klonades sekvensen i plasmiden pQlacZ [20]. PQlacZ är en så kallad broad-host-range
plasmid vilket betyder att den kan användas i många olika bakterier, den begränsas främst av
sin stora storlek, 17.1 kbp. PQlacZ är en reporterplasmid och bär på reportergenen lacZ, Med
hjälp av denna reportergen har man visat att sekvensen uppströms cld innehåller en promotor.
LacZ uttrycker β-galactosidase, transkriptionen av denna gen startar om en uppströms
promotorsekvens sätts in.
Genom att korta ner sekvensen med promotorn kan man få en tydligare bild av denna och
dess innehåll. Plasmiden pRU1103 (figur 2) är en betydligt mindre plasmid, 7990 bp lång,
med 17 klyvningssäten för olika restriktionsenzym. Den ingår i en serie promotor probe
vektorer, avsedda för att undersöka närvaron av promotor regioner i gramnegativa bakterier
[22]. En gen för lacZ finns även i pRU1103, tillsammans med en gen för gentamicin
resistens. Antibiotika resistensen kan utnyttjas vid detektion av transformanter. pRU1103 har
en komplett lacZ gen som endast saknar dess promotor och reglerande sekvenser. Om en
Figur 1 Sekvens uppströms cld i Ideonella dechloratans. Understruket i figuren ses möjlig ArsR bindande sekvens

Lisa Ljungberg
10
10
promotor sekvens sätts in framför lacZ genen kommer de bakterier som tagit upp plasmiden
med insert att uttrycka β-galactosidase, precis som hos pQlacZ. Tillför man sedan x-gal, en
laktos analog, hydrolyseras denna av β-galaktosidas och bildar färglös galaktos och blåfärgad
4-chloro-3-brom-indigo [23]. Den blåa färgen ses som blåa kolonier på agar platta.
Figur 2 Restriktionssäten, ORF's och gener hos PRU1103. Bilden kommer Ifrån Addgene, www.addgene.org/14467/
Syftet med detta examensarbete var att skala ner sekvensen uppströms om cld och samtidigt
behålla de sekvenser identifierade som möjliga promotor och reglerande sekvenser. Därefter
undersöka om den nedkortade sekvensen fortfarande fungerade som promotor i Escherichia
coli [20].
2. Material och metoder
2. 1 Primerdesign
Design av primerpar gjordes med Primer-BLAST ifrån NCBI. De resulterande fem primer
paren analyserades med avseende på dess benägenhet att bilda hairpin struktur, self-dimer
och hetero-dimer. Analysen gjordes med OlgioAnalyser 3.1 ifrån Integrated DNA

Lisa Ljungberg
11
11
Technologies®. Till uppströms primern adderades ett restriktionssäte för restriktionsenzymet
PstI och i nedströms primers adderades ett säte för HindIII.
2. 2 Plasmidpreparation
Renodling gjordes av E.coli XL1-Blue innehållande plasmiden pRU1103. Kolonierna
odlades på LB-agar platta (Appendix I) under 16h, 37°C och med en tillsats av 10µg/ml
gentamicin. Mer än tolv stycken kolonier plockades till varsin Ö. N kultur och utav dessa
gjordes plasmid preparationer med E.Z.N.A®, Plasmid Mini Kit I, från Omega Bio-Tek
enligt Plasmid mini Kit I spin Protocol med en elueringsvolym på 40-100µl. Plasmid
koncentrationen och renheten mättes med Infinite® 200 PRO NanoQuant vid 280nm samt
260nm. Kvoten av A260/A280 visar förhållandet mellan DNA och proteiner och ger ett mått på
provets renhet, vid ett värde över 1,8 anses provet vara rent.
2. 3 Restriktionsklyvning
För att kunna ligera in rätt sekvens behövde plasmiden, pRU1103, först klyvas med rätt
restriktionsenzymer, vilka matchar de säten som amplifierats i PCR'en. 5,3µg plasmid
(pRU1103) klyvs med restriktionsenzymer HindIII (8u/µg) ifrån Thermo Scientific och PstI
(8u/µg) ifrån New England Biolabs. Klyvningen utfördes med Buffer R (1x), enligt
rekommendationer ifrån Thermo Scientific DoubleDigest tool. Klyvningen, med en total
volym på 100µl, inkuberades under 60 minuter i 37 °C vattenbad. De båda
restriktionsenzyms sätena sitter i multipelt kloninssite (MCS) vilket resulterade i att ett 85 bp
långt fragment klyvdes bort, se figur 2. En inkubering i 80 °C under 20 minuter gjordes
avslutningsvis för att inaktivera HindIII och PstI innan gelrening. Vid kontroller av okluven
och kluven plasmid användes DNA stegen lambda Eco1301 (styI) 16, ifrån Fermentas, se
Appendix III.
2. 4 Rening av kluven plasmid
Resultatet ifrån restriktionsklyvningen fördes över till en 0,8% agaros gel med 1µl 6x loading
dye per brunn i gelen. Alla gelelektroforeser utfördes med 1x TEA-Buffer (Appendix I).
Provet kördes på gelen under 75V i 90 minuter. Till gelen sattes även kontrollklyvningar av
enbart HindIII respektive PstI för att kontrollera att dessa var aktiva och gav en korrekt
klyvning av plasmiden, pRU1103. Bandet med DNA ifrån gelen renades därefter med
QIAEX II Gel Extraction Kit, enligt medföljande protokoll för rening ur agarose gel.

Lisa Ljungberg
12
12
2. 5 PCR
PCR (tabell 2, 3 och 4) av genomiskt DNA användes för att amplifiera sekvensen av intresse,
figur 1. Alla PCR reaktioner utfördes med KAPA2G Fast HotStart ReadyMix, enligt
medföljande anvisningar för maximal yield reaction. Ungefär 30 ng genomiskt Ideonella
dechloratans DNA användes till varje reaktion. Alla kontroller av PCR produkter gjordes på
3-4% agarose gel med DNA stegen O'Gene ruler™, low range DNA ladder, Ready-to-use
(0,1µg/µl) ifrån Fermentas (Se AppendixIII).
Tabell 1 Temperaturer och tider för de olika stegen och cyklerna i PCR 1 (touchdown).
Steg Temperatur Tid Cykler
Initial denaturation 95 °C 3 min 1
Denaturation 95 °C 15 sek
Annealing 1* 65-55 °C 15 sek 35
Annealing 2 55 °C 15 sek
Final extension 72 °C 1 min 1
* Annealingtemperaturen sänktes, -1 °C/Cykel, under 10
cykler.
En annealingtemperatur på 55 °C användes till de sista 25 cyklerna.
Tabell 2 Temperaturer och tider för de olika stegen och cyklerna i PCR 2.
Steg Temperatur Tid Cykler
Initial denaturation 95 °C 3 min 1
Denaturation 95 °C 15 sek
Annealing 63 °C 15 sek 35
Final extension 72 °C 1 min 1
Tabell 3 Tempereturer och tider för de olika cyklerna i PCR 3.
Steg Temperatur Tid Cykler
Initial denaturation 95 °C 3 min 1
Denaturation 95 °C 15 sek
Annealing 56 °C 15 sek 35
Final extension 72 °C 1 min 1
PCR produkten renades ifrån salter och Taq-polymeras med GeneJet PCR Purification Kit
(Thermo Scientific). Den framrenade PCR produkten kontrollklyvdes med

Lisa Ljungberg
13
13
restriktionsenzymerna HindIII (Thermo Scientific) och PstI (New england Biolabs) enligt
återförsäljares rekommendationer och tidigare klyvning av pRU1103.
2. 6 Ligering
Genom en ligering kunde den amplifierade PCR produkten föras in i pRU1103. Ligeringen
gjordes med T4 Ligase och 10x T4 ligase buffer ifrån Thermo Scientific. Återförsäljarens
protokoll följdes för sticky-end ligation. 1µg plasmid ligerades med 28ng insert-DNA för 2:1
förhållande och med 14ng insert-DNA för 1:1 förhållande. Ligeringen inaktiveras i 65 °C
under 10 minuter. En volym på 60µl ligerat DNA erhölls och denna fälldes ut med etanol (se
Appendix II). Resulterande DNA löstes i TE-Buffer.
2. 7 Elektrokompetenta celler
XL1-Blue, E. coli, gjordes elektrokompetenta för att kunna ta upp plasmid genom
elektroporering. Alla tillsatser och centrifugeringar under framställningen av
elektrokompetenta celler gjordes på is eller i 4 °C. Först odlades en kultur av XL1-blue under
natt (250 ml LB-medium, se Appendix I). Cellerna odlades kommande dag till ett OD på
0,62. Efter att cellerna fått kylas på is under ca 15 minuter centrifugerades de i 15 minuter,
4500 rpm i rotor SLA-3000 (Sorvall™). Pelleten som bildades resuspenderades i ungefär
100ml iskallt vatten. Provet centrifugerades som innan och resuspenderades med ytterligare
40ml iskallt vatten. Efter ännu en centrifugering, som ovan, resuspenderades cellerna i 5ml
10% glycerol två gånger. Sista gången resuspenderades de i 500µl 10% glycerol. Cellerna
överfördes i 40µl portioner till eppendorfrör.
2. 8 Transformation och elektroporering
Transformation med XL1-Blue Competent Cells ifrån STRATAGENE utfördes, enligt
medföljande protokoll med pRU1103 innehållande insert ifrån PCR 3 (tabell 3).
Elektroporeringen gjordes med Gene Pulser II®
ifrån Bio-Rad. Fältstyrkan sattes till 2,5
kV/cm, resistensen till 200 Ω och kapacitansen till 25 μF. I kyvetten (Bio-rad) var avståndet
mellan elektroderna 0,2 cm. 40µl kompetenta celler pulsades en gång tillsammans ~40 ng
plasmid. En tidskonstans på 5,01 erhölls. Direkt efter pulsen var skickad tillsattes 1ml SOC-
medium (Appendix I). Cellerna inkuberades därefter i 5ml SOC-medium, 37°C under 1h. En
positiv kontroll av pUC18 gjordes under samma kriterier. Även otransformerade celler
odlades upp i 5 ml SOC-medium för att beräkna mängden celler vilka överlevt
elektroporeringen. Efter inkubering spreds de elektroporerade, transformerade, cellerna ut på

Lisa Ljungberg
14
14
LB-agar plattor innehållande gentamicin (10µg/ml) för pRU1103 och ampicilin (100µg/ml)
för pUC18. De otransformerade, elektrokompetenta cellerna spreds ut på LB-agar plattor utan
antibiotika. På plattorna med XL-1 Blue och pRU1103 sattes även X-gal (20mg/ml) för att
kontrollera vilka kloner som tagit upp plasmid innehållande insert genom blå/vit screening.
3. Resultat och diskussion
3.1 Primerdesign
De primer par som användes vid PCR såg ut enligt följande:
Upstream: 5'GCC TGC AGC AAG CGG AAA GTG CAC GAA T 3'
Den maximala frigjorda energin, ΔGmax värdet, för hairpin och själv-dimer bildning var
följande:
Hairpin ΔGmax: -1.93 kcal/mol
Self-dimer ΔGmax: -10.24 kcal/mol
Downstream: 5' GGC AAG CTT CTG GAA CGG CAT AGA GCC AA 3'
Hairpin ΔGmax: -3.02 kcal/mol
Self-dimer ΔGmax: -10.23 kcal/mol
ΔGmax värdet för hetero-dimer bildning mellan de båda primrarna var: -6.68 kcal/mol
5'GCGAGGTCGGCGTCAGTGTCGGGCGGTTGTCCCCGAGGGTCATGTTTGTTGAGAAGTCAATTTCCACTGGAA
CGGCATAGAGCCAAAAATATAGTGTTAAGTTTCGGAGAAACTCTTGACTTAAATCAAACACATAAAAAAGGA
GAAATGCGAAGATTCGTGCACTTTCCGCTTGCTCGGCGACC AAGCCGCCCA GC 3'
Experimentell Transkriptionsstart Möjlig FNR-Box Möjliga -10 och -35 sekvenser Möjlig arcA bindande sekvens
Figur 3 Sekvens uppströms cld med primer sekvenser markerade i rött. Understruket ses möjlig ArsR bindande sekvens.
Rödmarkerat ses inbindningssäten för uppströms och nedströms primer.
Rödmarkerat i figur 3 är sekvenserna vid vilka primer paren kommer binda in. Den nya
sekvensen ses i figuren nedan (figur 4). Detta är den avskalade sekvensen vilken klonades i
plasmiden pRU1103 i E. coli:

Lisa Ljungberg
15
15
(5’)CTGGAACGGCATAGAGCCAAAAATATAGTGTTAAGTTTCGGAGAAACTCTTGACTTAAATCAA
ACACATAAAAAAGGAGAAATGCGAAGATTCGTGCACTTTCCGCTTG (3’)
Experimentell Transkriptionsstart Möjlig FNR-Box Möjliga -10 och -35 sekvenser Möjlig arcA bindande sekvens
Figur 4 Den förkortade sekvensen som klonas i pRU1103. Understruket i figur ses möjligt ArsR bindande sekvens.
3. 2 Plasmidpreparation
Plasmiden pRU1103 preparerades fram i olika fraktioner, dessa poolades samman till en
slutvolym på 390 µl. Koncentrationen mättes till 63,35ng/µl, alltså en totalmängd på ~24µg
DNA. Provets renhet mättes till 2,04 (A260/280) vilket anses som rent (se tabell 3).
Tabell 4 Absorbansmätning vid 260 och 280nm för koncentrations- och renhetsbestämning av pRU1103.
Medelvärde av pRU1103 absorbans
260nm (A) 280nm (A) Konc (ng/µl) Ratio Volym (µl)
0,06335 0,0308 63,35 2,04 390
3. 3 Restriktionsklyvning
För att kontrollera att restriktionsenzymerna PstI och HindIII var aktiva och resulterade i en
klyvning av plasmiden gjordes kontrollklyvningar i samma buffer som användes till
dubbelklyvningar (5).

Lisa Ljungberg
16
16
Figur 5 Ifrån vänster till höger: 1: Stege, Lambda Eco1301 . 3: Oklyvd plasmid, pRU1103 5: pRu1103 klyvd med HindIII. 7:
pRU1103 klyvd med PstI. Resten av brunnarna är blanka
I brunn nummer 3, i figur 5, ses två band för den okluvna plasmiden. Ett band i toppen av
stegen, vid 19000 bp och ett strax under vid ungefär 7700 bp. Två band uppkommer eftersom
plasmiden finns i cirkulärt ( relaxerat) och supercoilat tillstånd. Plasmid i cirkulärt tillstånd
rör sig långsammare på gelen. I brunn nummer 7, i samma figur, ses ett band vid ungefär
8000 bp för pRU1103 kluven med PstI. Plasmiden ser ut att ha kluvits fullständigt av PstI
eftersom endast ett band ses i gelen vid önskad storlek, 7990 bp, med klyvd plasmid i linjär
form. Om två band uppkommit även här hade plasmiden fortfarande varit okluven. I brunn
nummer 3, i figur 5, syns inget band. Detta beror troligen på att en för liten mängd DNA
användes till klyvningen med HindIII. Hind III klyvningen kontrollerades därför ytterligare
en gång (figur 6). Den oklyvda plasmiden i cirkulärt tillstånd kunde förväntas röra sig enligt
en dubbel så stor storlek. Det är därför troligt att detta är vad som ses på gelen i brunn 3
(figur 5).
Lisa Ljungberg
17
17
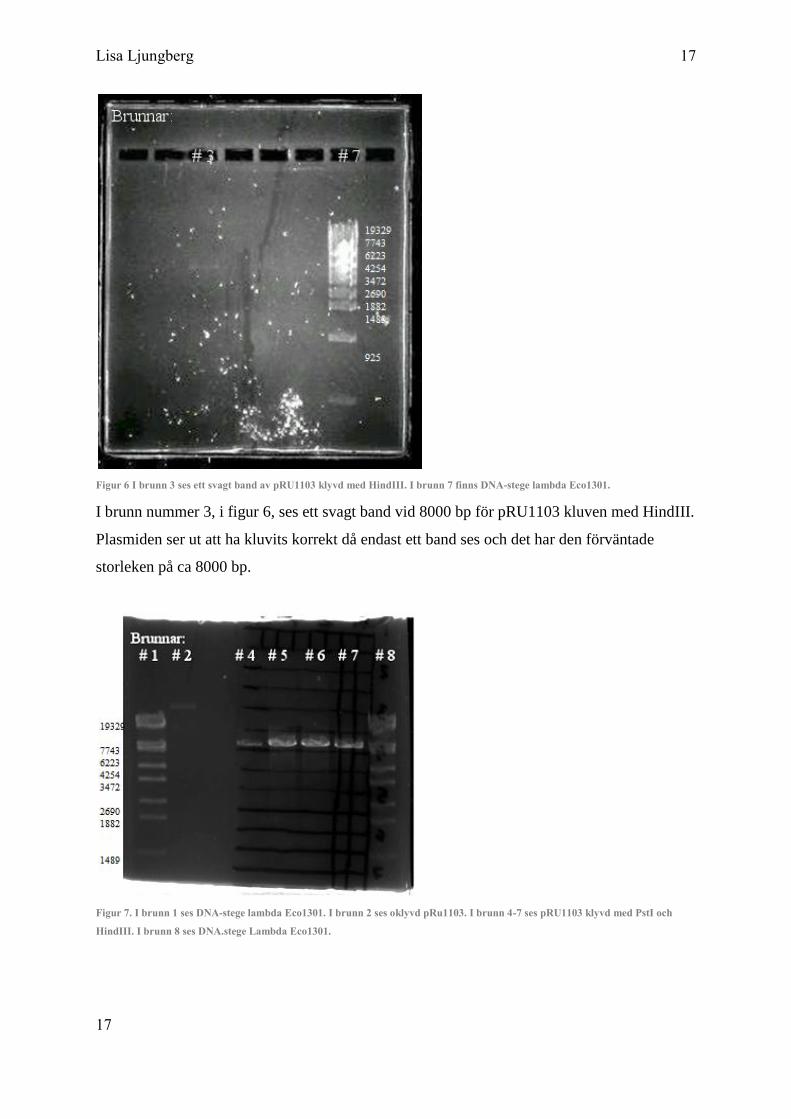
Figur 6 I brunn 3 ses ett svagt band av pRU1103 klyvd med HindIII. I brunn 7 finns DNA-stege lambda Eco1301.
I brunn nummer 3, i figur 6, ses ett svagt band vid 8000 bp för pRU1103 kluven med HindIII.
Plasmiden ser ut att ha kluvits korrekt då endast ett band ses och det har den förväntade
storleken på ca 8000 bp.
Figur 7. I brunn 1 ses DNA-stege lambda Eco1301. I brunn 2 ses oklyvd pRu1103. I brunn 4-7 ses pRU1103 klyvd med PstI och
HindIII. I brunn 8 ses DNA.stege Lambda Eco1301.

Lisa Ljungberg
18
18
I figur 7 ses i brunn nummer 4 till 7 pRU1103 kluven med både HindIII och PstI. Ett band ses
i respektive brunn vid lite mindre än 8000bp. Dubbelklyvningen ser korrekt ut eftersom ett
band ses i brunnarna 4, 5, 6 och 7 och dem har den förväntade storleken på ca 7990 bp.
I Brunn 2 ses ett band större än 19329 bp med okluven pRU1103. Bandet med okluven
pRU1103 ligger över den förväntade storleken, på 7990 bp, eftersom den troligtvis befinner
sig i relaxerad form och därför rör sig långsammare på gelen. Ifrån denna gel renades den
dubbelklyvda plasmiden.
3. 4 Rening av kluven plasmid
Tabell 5 Absorbansmätning vid 260 och 280nm för koncentrations- och renhetsbestämning av pRU1103 efter gelrening.
Medelvärde av renad pRU1103
260nm (A) 280nm (A) Konc (ng/µl) Ratio Volym (µl)
0,0195 0,0104 19,4 1,87 90
Efter gelreningen poolades produkterna samman till en totalvolym på 90µl som
koncentrationsbestäms till 19ng/µl (tabell 4). Detta ger en sammanlagd DNA mängd på 1,7µg
DNA. Ett utbyte på 30 % erhölls ifrån gelreningen. Renheten mättes till 1,87 (tabell 4), vilket
antyder att provet bör vara rent ifrån andra ämnen som proteiner och acetat. Reningen av
gelen fick optimeras ett par gånger då utbytet blev mycket lågt vid de första försöken. Den
största skillnaden mellan körningarna var i lufttorkningssteget. Pelleten fick torka under 15
minuter och resuspenderingen gjordes direkt då den vita färgen började träda fram. Pelleten
blev lätt över- eller under-torkad. Detta resulterade antingen i att etanol fanns kvar och störde
eller att pelleten blev för svår att resuspendera. Utbytet jämfördes med tidigare
dokumenterade gelreningar ifrån samma kit där det maximala utbytet också visade sig vara
30 %.

Lisa Ljungberg
19
19
3. 5 PCR och transformation
Figur 8 PCR 2, annealingtemperatur = 63° C. I brunn 2: "Prov 1". Brunn 5: "Prov 2". Brunn 6: Endast templat DNA (I.
dechloratans). Brunn 7: Negativ kontroll, utan templat. Brunn 8: DNA-stege O’Gene ruler, low range DNA ladder.
Under de första körningarna av PCR används en annealing-(hybridisering) temperatur på 63°
C (figur 8). I figur 8, brunnarna 2, 4 och 6 ses inga band. Ytterligare en reaktion gjordes,
denna gång med en annealingtemperatur på 56° C (figur 9). I brunn 3, figur 19, ses ett band
strax under 150bp för PCR produkten. I samma figur, brunn 4, ses ett band strax under 150
bp för den negativa kontrollen. I brunn 5 ses inget band för den positiva kontrollen. PCR
produkten, i brunn 3, ligger i samma storlek som den väntade produkten på 127 bp. I brunnen
med den negativa kontrollen, brunn 4, antogs det vara en kontaminering av genomiskt I.
dechloratans DNA. Detta eftersom ett band med ungefär önskad produktstorlek, 126 bp,
uppkom även här.
Figur 9 PCR 3, annealingtemperatur = 56° C. I brunn 1 och 8 ses DNA-stege, O’gene ruler, low range DNA- ladder. I bunn 3:
"Prov". Brunn 4: Negativ kontroll utan templat. Brunn 5: Endast templat DNA (I. dechloratans).

Lisa Ljungberg
20
20
Figur 10 Kolonier spädda med 100µl vatten ifrån koloni PCR. Brunn 1: Stege, O’Gene ruler low range DNA-ladder. Brunn 2-11
kolonier 1-10 ifrån transformation av pRU1103 med insert ifrån PCR 3. Brunn 15: Negativ kontroll.
I figur 10 ses PCR av kolonier ifrån transformation av XL1-blue superkompetenta celler med
pRU1103 innehållandes produkt ifrån PCR 3 (figur 9). I brunn nummer 5, 6, 8 och 11 ses
tydliga band vid lite under 150 bp. I Brunn 9, samma figur, ses ett svagt band vid lite under
150 bp. I brunn nummer 15 ses också ett band vid lite under 150 bp. I brunnarna 5, 6, 8 och
11 ser det önskade insertet ut att finnas då banden ligger vid önskad storlek, ungefär 126 bp. I
den negativa kontrollen, brunn 15, ses också ett tydligt band vid ungefär 126 bp. Under tiden
av koloni PCR’en kontrollerades allt material använt vid PCR för kontamineringar. Olika
koncentrationer detta materialet kördes ut på agarose geler, inga band uppkom och någon
kontaminering hittades inte. Produkten ifrån PCR reaktionerna bör vara resultatet av en
primer-dimer. För att komma runt detta problem användes en touchdown PCR, där
annealingtemperaturen fick gå ifrån 65-55° C (figur 11). Denna varierande
annealingtemperatur ger de båda primrarna en chans att binda in vid sin optimala
hybridiseringstemperatur. Temperaturintervallet valdes efter rekommendationer ifrån tidigare
användare av metoden. Detta intervall var också baserat på det faktum att ingen produkt
bildades vid en annealingtemperatur på 63 °C. Mycket produkt bildades däremot vid 56 °C,
den optimala temperaturen ansågs därför finnas någonstans emellan de båda
annealingtemperaturerna.

Lisa Ljungberg
21
21
Figur 11 PCR 1, Touchdown. I brunn 1: stege, O’gene ruler low range DNA-ladder. Brunn 2: prov ifrån PCR 1. Brunn 3: Negativ
kontroll.
I figur 11 ses resultatet ifrån PCR 1, av touchdown modell, av genomiskt I. dechloratans
DNA. I brunn 2, ett band vid ungefär 126 bp. I brunn, negativa kontroll, 3 ses inget band.
Produkten i brunn 2 ser ut att vara den rätta eftersom den negativa kontrollen nu är blank och
bandet ligger vid storleken för den önskade produkten, 126 bp.
Figur 12 LB- platta med XL1-blue transformerad med pRU1103 innehållandes insert ifrån PCR 1 (touchdown).
Materialen ifrån touchdown PCR'en (figur 11) ligerades in i pRU1103. Sedan
transformerades E. coli, XL1-Blue, genom elektroporering och kolonier odlades upp på x-gal.
En av Ö. N kulturerna ses i figur 12, med många blåfärgade, men små, kolonier. Blåfärgade
kolonier uppkom i alla Ö. N kulturer. Ifrån en spädning av transformanter med en

Lisa Ljungberg
22
22
spädningsfaktor på 1:70 000 växte 60 kolonier på plattan. Detta gav 1,67*108
transformanter/µg DNA och 4,20*106 cfu/ml. De elektrokompetenta cellerna, vilka ej hade
elektroporerats, späddes med en spädningsfaktor på 1:1 000 000 och gav 11 kolonier,
1,10*107 cfu/ ml. Vid processen då cellerna gjordes elektrokompetenta överlevde endast 2%
(cellerna innan de gjorde elektrokompetenta hade 6,00*108 cfu/ml). Efter elektroporeringen
och uppodling på antibiotika hade 38 % av cellerna överlevt.
Figur 13 pRU1103 med långt insert (promotor) odlad på X-gal
Figur 14 - pRU1103 utan insert odlad på X-gal
I figurerna ovan (figur 13 och 14) ses pRU1103 med det långa insertet (figur 1) och pRU1103
utan insert odlade på LB-agar plattor med X-gal. I figur 13 ses blåfärgade kolonier. I figur 14
är kolonierna färglösa. De blåa kolonierna har producerat β-galactosidase och kan därför
bryta ner X-gal till den blåfärgade produkten. Det långa insertet innehåller alltså en promotor
sekvens. Det nedkortade sekvensen uppströms cld, i figur 12, innehåller troligtvis också en
fungerande promotor eftersom kolonierna blivit blåfärgade även här.

Lisa Ljungberg
23
23
Figur 15 PCR av kolonier ifrån transformation av XL1-blue med pRU1103 (+ insert ifrån PCR 1 (touchdown)) I brunn 1 och 9:
Stege, O'Generuler Low rangen DNA -ladder. Brunn 2, 4, 6, 10 och 12: Kolonier 1-5 spädda i 100 µl H2O. Brunn 3, 5, 7 och 11:
Kolonier 1-4 spädda i 1000µl H2O. Brunn 8: Negativ kontroll, utan templat DNA.
I figur 15 ses resultatet ifrån PCR på kolonier ifrån transformationen av XL1-Blue med
pRU1103 innehållandes det korta insertet ifrån PCR 1 (touchdown). I brunnarna 2, 4, 6, 10
och 12 ses kolonierna 1-5, spädda i 100 µl vatten, Här finns ett band vid ungefär 126 bp. I
brunnarna 3, 5, 7 och 11 ses kolonier 1-4 spädda i 1000µl vatten. Här finns ett band vid 126
bp i alla brunnar utom nummer 10. I brunn nummer 8 finns den negativa kontroller, här ses
inget band. I alla kolonier ser insertet ut att finnas då banden ligger vid storleken för
produkten, 126 bp. Detta visar att transformationen har fungerat och att pRU1103 har tagit
upp det önskade fragmentet. Att det finns en band i brunn 11 men inte i brunn 10 kan bero på
att cellerna ifrån koloni 4 spädda i 100µl inte lyserats på grund av att LB funnits kvar och
inhiberat. De kloner som odlats upp på plattor är väldigt små, tillväxten ser ut av avstanna
efter att de börjat växa på LB-plattan. Samma resultat sågs i alla kloner ifrån
transformationen. En möjlighet är att en stor produktion av β-galaktosidas verkar toxiskt för
cellen, eller att det finns inhiberande regioner vilka tagits bort i och med förkortningen.

Lisa Ljungberg
24
24
Sammanfattande slutsats
Att kolonierna ifrån elektroporeringen blivit blåfärgade tyder på att en fungerande promotor
finns. Plasmiden pRU1103 har alltså tagit upp ett insert innehållandes en promotorsekvens.
Till denna promotorsekvens i plasmiden kan RNA-polymeras binda in och börja transkription
av den efterföljande lacZ genen vilket till att cellerna producerar β- Galaktosidas och färgar
kolonierna blå i närvaro av x-gal. De designade primerparen har därmed fungerat och
amplifierat det önskade området.
Genom att PCR-reaktionen gjord på de transformerade klonerna ger en produkt av förväntad
längd är det troligt att önskad sekvens har tagits upp. Ett nästa steg vore att jämföra promotor
aktiviteten i det korta insertet med det tidigare klonade, långa, insertet. Promotor aktiviteten
kan mätas och jämföras med avseende på enzymaktivitet i en β- Galaktosidas-assay.
Ytterligare ett steg vore att klona den nya, kortare, promotor sekvensek i Ideonella
dechloratans. Om framtida studier på klonerna ska vara möjligt behövs dock kolonier som
växer bättre. Ett försök vore att odla dem på SOC-medium istället för LB-medium, då de får
tillgång till mer mineraler och glukos.

Lisa Ljungberg
25
25
Referenser
1. Ahn S. C, Hubbard B, Cha D, Kim B. J : Simultaneous removal of perchlorate and
energetic compounds in munitions wastewater by zero-valent iron and perchlorate-respiring
bacteria. Journal of Environmental Science and Health, Part A 2014;49:575-583.
2. Schwarz AO, Urrutia H, Vidal JM, Perez N: Chlorate reduction capacity and
characterisation of chlorate reducing bacteria communities in sediments of the rio cruces
wetland in southern chile. WATER RESEARCH 2012;46:3283-3292.
3. Wolff J: Perchlorate and the thyroid gland. Pharmacol Rev 1998;50:89-105.
4. Health implications of perchlorate ingestion. The National Academies Press, 2005.
5. Li H, Zhang X, Lin C, Wu Q: Toxic effects of chlorate on three plant species inoculated
with arbuscular mycorrhizal fungi. Ecotoxicology and Environmental Safety 2008;71:700-
705.
6. Palma AT, Henríquez LA, Alvarez X, Fariña JM, Schwarz A, Lu Q: Do subtoxic levels of
chlorate influence the desiccation tolerance of egeria densa? Environmental Toxicology and
Chemistry 2013;32:417-422.
7. DJ: Chlorate toxicity in aspergillus nidulans. Studies of mutants altered in nitrate
assimilation. Molecular & General Genetics: MGG 1976;146:147-159.
8. WSH: Chlorite and chlorate in drinking water. Background document for development of
who guidelines for drinking-water quality. World Health Organization, Water Sanitation and
Health, 2005.
9. Nilsson T, Rova M, Smedja Bäcklund A: Microbial metabolism of oxochlorates: A
bioenergetic perspective. Biochimica et Biophysica Acta (BBA) - Bioenergetics
2013;1827:189-197.
10. Mehboob F, Wolterink AFM, Vermeulen AJ, Jiang B, Stams AJM, Kengen SWM,
Hagedoorn PL: Purification and characterization of a chlorite dismutase from pseudomonas
chloritidismutans. FEMS Microbiology Letters 2009;293:115-121.
11. Malmqvist Å, Welander T, Moore E, Ternström A, Molin G, Stenström I-M: Ideonella
dechloratans gen.Nov., sp.Nov., a new bacterium capable of growing anaerobically with
chlorate as an electron acceptor. Systematic and Applied Microbiology 1994;17:58-64.
12. Nelson DL, Cox MM, Lehninger AL: Lehninger principles of biochemistry / david l.
Nelson, michael m. Cox. New York : W. H. Freeman ; Basingstoke : Palgrave [distributor],
cop. 2008
5th ed., 2008.
13. Hellberg Lindqvist M, Johansson N, Nilsson T, Rova M: Expression of chlorite dismutase
and chlorate reductase in the prescence of oxygen and/or chlorate as the terminal electron

Lisa Ljungberg
26
26
acceptor in ideonella dechloratans. Applied and Environmental Microbiology 2012;78:4380-
4385.
14. Bell A, Busby S: Location and orientation of an activating region in the escherichia coli
transcription factor, fnr. Molecular Microbiology 1994;11:383-390.
15. Lynch AS, Lin ECC: Transcriptional control mediated by the arca two-component
response regulator protein of escherichia coli: Characterization of dna binding at target
promoters. Journal of Bacteriology 1996;178:6238-6249.
16. Joshi M, Dikshit KL: Oxygen dependent regulation of vitreoscilla globin gene: Evidence
for positive regulation by fnr. Biochemical And Biophysical Research Communications
1994;202:535-542.
17. Vasilieva SV, Strel'tsova DA, Moshkovskaya EY, Vanin AF, Mikoyan VD, Sanina NA,
Aldoshin SM: Reversible no-catalyzed destruction of the fe-s cluster of the fnr[4fe-4s]
transcription factor: A way to regulate the aidb gene activity in escherichia coli cells cultured
under anaerobic conditions. Doklady Biochemistry & Biophysics 2010;435:283-286.
18. Gao Ch, Yang M, He ZG: Characterization of a novel arsr-like regulator encoded by
rv2034 in mycobacterium tuberculosis. PLoS ONE 2012;7.
19. Park H-D, Guinn KM, Harrell MI, Liao R, Voskuil MI, Tompa M, Schoolnik GK,
Sherman DR: Rv3133c/dosr is a transcription factor that mediates the hypoxic response
ofmycobacterium tuberculosis. Molecular Microbiology 2003;48:833.
20. Ölund D: Konstruktion av reporterplasmider innehållande möjliga promotorregioner för
kloratreduktas respektive kloritdismutas från Ideonella dechloratans, 2012, pp 26.
21. Miller JH: Experiments in molecular genetics. Cold Spring Harbor Laboratory, 1972.
22. Karunakaran R, Mauchline TH, Hosie AH, Poole PS: A family of promoter probe vectors
incorporating autofluorescent and chromogenic reporter proteins for studying gene
expression in gram-negative bacteria. Microbiology 2005;151:3249-3256.
23. Horwitz JP, Chua J, Curby RJ, Tomson AJ, Da Rooge MA, Fisher BE, Mauricio J,
Klundt I: Substrates for cytochemical demonstration of enzyme activity. I. Some substituted
3-indolyl-β-d-glycopyranosides1a. Journal of Medicinal Chemistry 1964;7:574-575.

Lisa Ljungberg
27
27
Appendix I
Soc-Medium:
0,5% Jästextrakt
2% Tryptone
10mM NaCl
2.5mM KCl
10mM MgCl2
10mM MgSO4
20 mM Glukos
1 x TEA Buffer:
1mM EDTA
20mM Ättiksyra
40mM Trizma Base
LB-Medium, 1 liter
10g NaCl
5g Jästextrakt
10g Tryptone
fylls till 1 liter med H2O

Lisa Ljungberg
28
28
Appendix II
Etanol fällning:
Uppskatta provets volym
Tillsätt 1/10 volymer natriumacetat, pH 5,2 till en slutkoncentration på 0,3M
(Förutsatt att DNA't finns i enbart TE-buffer. Om inte beräkna med avseende på
saltkoncentrationen)
Blanda väl
Tillsätt 2 till 2.5 volymer iskall 99,9% Etanol
Blanda väl
Inkubera på is > 20 minuter
Centrifugeras på14,5 x g, 10-15 minuter
Häll av supernatant
Tillsätt 1ml 70% Etanol, centrifugeras snabbt och ta sedan bort supernatant
Lufttorka pellet
Resuspendera i önskad mängd TE-buffer

Lisa Ljungberg
29
29
Appendix III
Figur 16 DNA stege, O'gene Ruler. Bild ifrån ThermoScientific.
Figur 17 DNA stege Lambda Eco1301. Bilden är en skärmdump ifrån tillverkarens (Fermentas) meföljande protokoll.





























