Embed Size (px)
Citation preview

13
1 はじめに 近年,医学においてコンピューターを用いた診断や予測が活発に行われている。このような診断や予測の結果を深く正しく理解することは,臨床医学にとって非常に重要である。しかしながら,これらの背後には基礎医学,物理学,生物学,数学,情報科学といった多くの分野が密接に絡み合い,これらの分野を横断した統合的理解が必要となる。この統合的理解を促す一つの方法として,各分野の実習課題に積極的に分野横断型のテーマを組み込むことが考えられる。そこで本年,物理学教室では一つの試みとして 1年次基礎物理実験(講義名「自然科学実験Ⅰ」)において,Java言語を用いた神経活動電位の数値シミュレーションに関する課題(課題名「Javaによる活動電位のシミュレーション」)を導入した。 この課題では,神経活動電位(生理学・細胞生物学)の背後にある等価電気回路(物理学)を微分方程式(数学)で表現し,コンピューターを用いて数値的にシミュレーションを行う(情報科学)ことを通して,神経活動電位と数値計算及びプログラミングの基礎を俯瞰的に学習することを目的とする。本報告では,まず本課題が実施された自然科学実験Ⅰと本課題に関連する講義について概説し,続いて導入した課題の原理と内容を説明する。そして実習後に実施した学生アンケートの集計結果に基づき,数値計算と物理現象,生理現象に関する学習効果を検討し,課題の改善案について提起する。
2 基礎物理学実験と本課題に関連する講義 本学では 1年次後期に基礎物理学実験(自然科学実験Ⅰ)を半期 2時限で行っている。全13回から構成され,最初の実習では実験ガイダンス及び測定データの扱い方に関する説明を行う。第 2週以降は 4人 1グループに分かれ,各グループ 1課題を 1週ごとのローテーションによって,順次表 1の 16の実習課題のうち 12課題を進める。本課題「Javaによる活動電位のシミュレーション」は,前年まで行っていた「コンピューターによるオシロスコープ・
Javaによる活動電位のシミュレーション―日本大学医学部 1年次「自然科学実験Ⅰ」における物理・生理学・情報科学の融合課題の導入例報告―
坪 泰 宏理化学研究所,日本大学医学部一般教育学系物理学分野深 作 貴美代,田 代 健 治,糸 井 充 穂
日本大学医学部一般教育学系物理学分野
教育実践

14 坪 泰 宏,深 作 貴美代,田 代 健 治,糸 井 充 穂
シミュレーション」に代わり導入された。
表 1 日本大学医学部 1年次基礎物理実験「自然科学実験Ⅰ」の 16課題(2011年度)
1.ジュール熱による熱の仕事当量の測定 9.固体ダイオードの特性試験と整流回路2.気柱の共鳴 10.電気天秤による微小質量の測定3.オームの法則の検証と抵抗率の測定 11.軟 X線による透視と写真処理4.顕微鏡による屈折率の測定 12.テスターとデジボルの比較実験5.水の粘性係数の測定 13.レーザー光による回折と干渉実験6.サーミスタの特性試験 14.等電位線の観察7.薄いレンズの焦点距離 15.Javaによる活動電位のシミュレーション8.超音波 16.重力加速度の測定
この基礎物理学実験に先立ち,1年次前期に物理学では基礎物理学,医系物理学Ⅰ,医系物理学Ⅱの講義を行っている。本学では約半数の学生が受験時に物理・化学を選択しており,残りの半数が生物・化学を選択している。生物・化学選択受験生の多くは高校物理を未習であり,物理・化学選択受験生は高校生物を未習であるため,主に高校物理未習者を対象としたクラスと既習者を対象としたクラスに分けている。以後,便宜的に高校物理未習者,既習者と呼ぶ。未習者対象の「基礎物理学」及び「医系物理学Ⅰ」(それぞれ 1時限)においては力学,電磁気学,光学,波動,流体を,主に既習者対象の「医系物理学Ⅱ」( 1時限)においては電磁気学,波動,流体を講義している。またこの実験に引き続き,2年次前期において「医系物理学Ⅲ」( 1時限)において原子核,放射線,熱力学,シミュレーションと微分方程式の講義を行っている。つまり,基礎物理学実験のほとんどの課題は講義の後に実験を行い, 2つの課題「ジュール熱による熱の仕事当量の測定」と「軟 X線による透視と写真処理」に関しては実験の後に講義が行われる。 本年度の課題「Javaによる活動電位のシミュレーション」に関係する物理学の講義は,医系物理学Ⅰの第 2講「コンデンサーと生体の静電気現象」,及び医系物理学Ⅱの第 12講「実用回路での電圧-電流の関係」によって行った。ここでは後述するように神経細胞膜の並列等価回路モデルと,準静的近似の元でのホジキンハクスレー(Hodgkin-Huxley)方程式の解釈を説明した。上述の物理学以外の分野では,1,2年次に限っても,「数学Ⅰ」(微積分),「基礎生物学」(神経細胞),「細胞生物学」(細胞膜の電気特性),「情報科学」(概論),「医用工学」(神経活動の検出),「神経科学」と多岐に渡り,本課題と関連した講義が行われている。学生にこれらの知識の関連性を再認識させ,有機的につなぐことで俯瞰的に医学関連知識を体得させることを目指して,本課題を導入した。
3 課題原理3- 1 神経細胞膜とコンデンサー[1]
細胞は脂質二重層からなる薄い細胞膜で覆われている。脂質は極性イオンや水分子を通さ

15Javaによる活動電位のシミュレーション
ない絶縁体であるため,この膜によって細胞の内液と外液が電気的に隔てられている。この細胞膜にはイオンポンプ(ion pump)と呼ばれるタンパク質が存在し,ATP(アデノシン三リン酸;adenosine triphosphate)などのエネルギーを利用して特定のイオンを膜の内外へ相互に輸送している。例えば,ナトリウム-カリウムポンプは 3個のナトリウムイオンを細胞内から細胞外へ送り,2個のカリウムイオンを細胞外から細胞内へ送る。このようなイオンポンプなどの寄与により,細胞内外の液間にはイオン組成に差異が生じている。 一方,細胞膜にはイオンポンプの他に,イオンチャネル(ion channel)と呼ばれるタンパク質が存在する。イオンチャネルは特定のイオンのみを選択的に通す「穴」であって,エネルギーを使わずに特定のイオンを往来させる。またイオンチャネルは膜内外の電位差や化学物質の有無に依存して開閉する[2]。いま,細胞内外の液がそれぞれイオン組成は異なるがお互いに電気的に中性だと仮定しよう。ここで,例えばカリウムのみが往来できる(あるカリウムチャネルのみ開いている)状態を考えると,カリウム濃度が高い細胞内から低い細胞外へ平均的なカリウムイオンの移動が生じる(全イオンが高濃度側から低濃度側へ移動する訳ではなく,平均流入量と流出量の差が濃度差で決まることに注意)。その結果,膜の内外で電荷に偏りが生じ,電位差が生じるとともに,絶縁体である膜はコンデンサー(キャパシター:capacitor)として振る舞う。膜内外の電位勾配と濃度勾配がバランスしたところで平均的な移動は止まる。一般に膜外の電位を基準(0[V])とした膜内の電位を膜電位という。上述の例の状態は神経細胞が刺激を受けていない状態(静止状態)におおよそ対応しており,この状態の膜電位は負になり約-70[mV]となる(静止膜電位)。ちなみに,この静止膜電位は細胞内外の液組成やイオンチャネルの分布に依存して決まり,例えば大脳皮質の神経細胞では細胞により静止膜電位が異なる[3]。 神経細胞が刺激をうけ,(厳密には正しくないが)膜電位がある閾値を超えると,興奮性膜では急激に膜電位が変化して細胞内が約+40[mV]に達する。その後数ミリ秒のうちに再び負の電位へ戻る。これを活動電位という。この活動電位は神経細胞の軸索突起を伝搬し,
図 1 神経細胞の模式図と 3つの大脳皮質錐体細胞(パッチクランプ法 biocytin/ABC/Ni-DAB染色)。軸索突起は極めて細く,目立つ突起はほとんどが樹状突起である。

16 坪 泰 宏,深 作 貴美代,田 代 健 治,糸 井 充 穂
神経終末と呼ばれる末端に到達する。活動電位が神経終末に到達すると,細胞内カルシウムイオン濃度の上昇を引き起こし,グルタミン酸(glutamate)や GABA(γアミノ酪酸;γ-aminobutyric acid),アセチルコリン(acetylcholine)などの神経伝達物質がシナプス(synapse)において細胞外へ放出される(化学シナプス)。この伝達物質を次の神経細胞が受容体により受け取ると,その受容体に応じたイオン電流が次の神経細胞で生じる。このように,活動電位の発生は神経細胞の情報伝達にとって重要であると考えられている。
3- 2 単純な神経細胞膜の等価回路モデル[4]
静止膜電位,活動電位など細胞膜の電位変化を統一的に理解する為に,細胞膜で生じている電気現象を電気回路に置き換えて考える。この方法を等価回路モデルという。図 2に細胞膜とその等価回路を示す。等価回路では,膜の脂質二重層とそれを挟む細胞内外の電解質を電気容量 Cmのコンデンサーと見なし,膜を通過するイオンの流れは内部抵抗が Rm=1/gL(gL
はリークコンダクタンス(conductance)で抵抗の逆数)である起電力 ELの電池による電流と見なす。コンデンサーに蓄えられる電荷 Qは,コンデンサーにかかる電圧(すなわち膜電位 Vm(t))と電気容量 Cmの積で表される。コンデンサーの電流は電荷の時間微分 CmdVm(t)/dt
となり,膜のコンダクタンス gLが時間変化しない細胞膜に電流 I(t)を内向きに流した場合には,図 2の回路に電流保存則(キルヒホッフ(Kirchhoff)の法則)を適用することで,回路に流れる電流に関して
( )( ) ( ( ) )m
m L m LdV t
I t C g V t Edt
= + − ⑴
が成立することがわかる。この微分方程式は,時間に関する膜電位の方程式であり,変形すると単純な等価回路モデルは後述の定数係数非斉次線形一階微分方程式で表される。
( )m L L Lm
m m
dV g g E I tV
dt C C+
= − + ⑵
図 2 細胞膜と等価回路

17Javaによる活動電位のシミュレーション
3- 3 定数係数非斉次線形一階微分方程式 微分方程式とは,式⑵のような導関数を用いて書かれた方程式である。x2-1=0のような代数方程式は,解が x=±1という「値」として求められる。一方,微分方程式は値(定数)が解になることはあるものの,一般的には式(関数)の形で解が求められる。例えば式⑵の場合,この式の関係を満たす時間の関数 Vm(t)=f(t)を求めることが解くべき問題となる。 以下のように未知関数 xの一次導関数 dx/dtと xの項のみからなり,それぞれが定数係数である常微分方程式
dxax
dt= ⑶
を定数係数線形一階微分方程式と呼ぶ。未知関数 xを位置,微分変数 tを時刻だと考えれば,この微分方程式は,例えばa<0ならば,全ての位置で速度はx=0へ向かう方向に向いており,x=0から遠いほど速さが大きくなる。結果としてこの微分方程式は,時間とともに x=0に向かう緩和(減衰)状態を表している。この微分方程式は解として,未知関数 xが微分変数 t
の式の形で明示的に解けて,その解(解析解)は,
x=A・eat ⑷
で与えられる。ここで式⑷を式⑶に代入すればわかるように,定数 Aは任意の値をとりうる(一般解)。この定数 Aは境界条件が与えられることで決定される(例えば「t=0のときに x=1」が境界条件ならば A=1)。物理における微分方程式は,常に境界条件とセットで考える。ちなみに,ほとんどの微分方程式は解析的に解けないが,時間に関する一階微分方程式は必ず発散するか,ある値に収束する( 8章参照)。 さて式⑵を再び眺めると,以下のように式⑶に xを含まない項が加わった形の微分方程式であることがわかる。
( )dx
ax Q tdt
= +
この形の微分方程式を定数係数非斉次線形一階微分方程式と呼ぶ。この場合の解析解は,
( )0( )
tat asx e Q s e ds B−= +∫で与えられる。よって,式⑵の一般解は,
/ / /
0
1( ) ( 1) ( )L m L m L m
tg t C g t C g s Cm L
m
V t e E e I s e ds BC
− ⎛ ⎞= − + +⎜ ⎟⎝ ⎠∫ ⑸
となる。例えば I(t)=I0の場合,初期条件として Vm(t=50)=ELとすると,式⑸は
mtm L
L
IV e E
g0 ( 50) /(1 )τ− −= − + ⑹
となる。ここで,τm=Cm/gL=RmCmは膜時定数と呼ばれ,電流変化に対する膜電位の応答時間を意味する。神経細胞の膜時定数は典型的に数ミリ秒から数十ミリ秒である[5]。

18 坪 泰 宏,深 作 貴美代,田 代 健 治,糸 井 充 穂
3- 4 ホジキンハクスレー(Hodgkin⊖Huxley)方程式[4]
次にホジキン(Hodgkin)とハクスレー(Hux-
ley)が電位固定法を駆使して得た知見をもとに構築したヤリイカの巨大神経軸索の膜電位を記述する数理モデルを示す[6]。このホジキンハクスレー方程式は,先に述べた単純な等価回路モデルに,ナトリウムチャネルとカリウムチャネルに相当する「内部抵抗(コンダクタンス)+電池」を並列に接続したモデルである(図 3)。さらに,このモデルで重要な点は,先ほどの等価回路モデルで一定だったコンダクタンス gLが,ナトリウムコンダクタンスとカリウムコンダクタンスでは膜電位に依存して変化する点である。ホジキンとハクスレーは ① 膜を流れる電流はコンデンサーを流れる電流とイオン電流に分離される(電流保存則) ② イオン電流はナトリウム電流,カリウム電流,リーク電流の 3つに分離される ③ 各イオン電流はオーム(Ohm)の法則 Iion=gion(Vm-Eion)に従って流れる ④ 膜の電気容量 Cm及び各イオンの平衡電位(ENa, EK, EL)は変化しないの条件のもと膜電位(Vm),チャンネルの活性化因子(m, n),および不活性化因子(h)を変数とする 4元連立非線形一階微分方程式をたて,活動電位を再構成する事に成功した。ホジキンとハクスレーはこの一連の仕事によりノーベル生理学賞を受賞したが,驚きなのは当時まだイオンチャネルというタンパク質が発見されていなかった点である。ちなみに非線形な一階微分方程式とは,未知関数 xの一次導関数 dx/dtと xの複雑な(一次式ではない)関数からなる微分方程式である。線形微分方程式では解の関数は比較的単純になるが,連立非線形微分方程式では一般に解は解析的に表せない。ホジキンハクスレー方程式は以下の連立微分方程式である。
( ) ( ) ( ) ( )mm L m L Na m Na K m KdV
I t C g V E g V E g V Edt
= + − + − + − ⑺
⑻Na Na K K
mm m m m m
m
h h h m hm
mn n n
m
g m hG g n G
dm Vm m V
dt V
dhh h V
dt V
dn Vn n
dt V
3 4,
0.1( 50)(1 ) , , 4 exp( ( 75) / 18)
exp( ( 50) / 10) 1
1(1 ) , 0.07exp( ( 75) / 20),
exp( ( 45) / 10) 1
0.01( 65)(1 ) ,
exp( ( 65) / 10)
= =
− += − − = = − +
− + −
= − − = − + =− + +
− += − − =
− + −
α β α β
α β α β
α β α n mV, 0.125exp( ( 75) / 80)1
= − +β
図 3 ホジキンハクスレーモデルの等価回路 Hodgkin and Huxley J Physiol 117 : 500(1952)より引用

19Javaによる活動電位のシミュレーション
式⑺は等価並列回路を表している。この式は,リーク平衡電位 EL,ナトリウム平衡電位ENa,カリウム平衡電位 EKのコンダクタンスの比による重心の電位に向かって,膜電位が緩和することを意味している( 8章参照)。つまり,これら 3つのコンダクタンスの大きい項の平衡電位に近い電位に膜電位は向かう。式⑻はナトリウムコンダクタンス gNaが最大コンダクタンス GNaと活性化変数の 3乗 m3と不活性化変数 hの積で表されること,またカリウムチャネルのコンダクタンスが最大コンダクタンス GKと活性化変数の 4乗 n4の積で表されることを表している。活性化変数,不活性化変数は 0から 1の値をとり,いわば「チャネルの開き具合」を決定する。その下の式では活性(不活性)化変数の時間変化が記述されている。これらは膜電位に依存して変化することから,上述の緩和先の「重心の電位」は現在の膜電位によって決定される。緩和先に向かうと膜電位が変わり,また緩和先が変わり,そこに向かうと膜電位が変わり,ということを繰り返すことで,結果として活動電位のような複雑な電位変化が観察される。ちなみに,αは開きやすさ,βは閉じやすさを意味し,1/(α+β)
は活性(不活性)化変数の時定数,α/(α+β)は活性(不活性)化変数の緩和先となる。なお原論文では膜電位として細胞内静止膜電位に対する細胞外電位 Vをとり,各イオンの平衡電位 Vionも静止電位 Vrestを基準としているので,上記の式では Vrest=-75[mV]として,前節の等価回路の表式にあわせるように変数変換を行った(電流の方向も逆である)。α,βの関数は 6.3℃の条件のものである。一般に温度が上昇すると活性(不活性)化変数の時定数が速くなる(Q10という値で 10℃上昇ごとの時定数変化を記述する)。ホジキンとハクスレーが用いた定数は,式⑺⑻の定数に変換すると,ENa=40[mV],EK=87[mV],EL=-64.387
[mV],GNa=120[mS/cm2],GK=36[mS/cm2],gL=0.3[mS/cm2],Cm=1.0[μF/cm2]である。以下では簡単のため単位は省略する。
3- 5 微分方程式の数値解法(オイラー法)[7]
本課題ではコンピューターを使って微分方程式(式⑴)及び(式⑺⑻)を数値的に解く。式⑴は解析解が求まるので,一般には解析解を用いることが多い。一方式⑺⑻は連立非線形微分方程式であり,特殊な初期条件をとらない限り解析解を求めることはできない。解析解が求まらない場合は微分方程式を数値的に解く必要がある。数値的に解く方法には様々あるが,ここでは最も基本的なオイラー(Euler)法を用いる。オイラー法は基本的な方法であるが,昨今の計算機能力を考えると本格的な数値計算をしない限りはオイラー法で十分であると思われる。またノイズを含むダイナミクスや金融工学のモデルとして用いられる乗法型確率微分方程式[8]などを解く際には,オイラー法を用いるのが比較的安全である。 オイラー法のように,ある物理量の時間発展に関する微分方程式を数値的に解く方法の殆どは,時間を離散ステップに分割する事から始める(図 4)。まず,微分方程式
( , )dx
f x tdt
=
を考える。ここでは物理量 xを球の位置としよう。この微分方程式では時刻 tにおける球の

20 坪 泰 宏,深 作 貴美代,田 代 健 治,糸 井 充 穂
速度は,f(x, t)で与えられる。もし,時刻 t=tnから次の時間ステップの時刻 t=tn-Δtまで球がこの速度で動くとすると,次の時間ステップにおける球の位置は,
x(tn+Δt)=x(tn)+f(x(tn), tn)Δt ⑼となり,前の時間ステップにおける位置 x(tn)と,前の時間ステップにおける速度 f(x(tn), tn)
から求められる。f(x(tn), tn)は x(tn)の関数であるため,結局次の時間ステップにおける位置x(tn+Δt)は前の時間ステップにおける位置 x(tn)から逐次的に求まることになる。繰り返しになるが注目すべき点は,「f(x, t)の x, tに前のステップで計算した xを代入する」という操作を繰り返すだけで,各ステップでの xが逐次的に求まるということである。この計算をもちろん人間が行ってもよいが,非常に多数のステップを計算しなければならない。コンピュータープログラムには一般に同じ計算の繰り返しを指示する命令が存在する。これを援用して上述のオイラー法を効率的に解くことができる。 図 4の示すように,Δtを小さくとるほど真の解に近くなる。しかしながら,計算ステップ数は求めたい時間の長さを Δtで割った回数となるので,Δtを小さくとるほど計算時間がかかる。この問題を解決する様々な方法が提案されている。
3- 6 Javaプログラム説明 本課題では,あらかじめ用意された 2つの Javaアプレットプログラム(Jikken1_RC.java,
Jikken2_HHequation.java)[9]を統合開発環境 Eclipse[10]上で改変,実行する。【Jikken1_RC.java】 単純な神経細胞膜電位の等価回路を記述する微分方程式(式⑴)を境界条件 Vm(t=50)=0
のもとで解いた解析解(式⑹)に Cm=1.0,gL=0.3,I0=10.0を代入したグラフ(図 5上段;提示の図では,3つの時間区間でそれぞれ電流値及び境界条件を場合分けして計算,描画している),及び微分方程式(式⑴)に t=50から t=550までの間に I0=10.0の矩形電流(図5下段)を入力した場合のオイラー法による数値解のグラフ(図 5中段;パラメータは解析解と同じ)を出力するプログラムである。なお,オイラー法の時間ステップは Δt=0.025
とした。プログラムのほとんどはグラフの出力に関する命令である(ソースコードはWeb
図 4 オイラー法
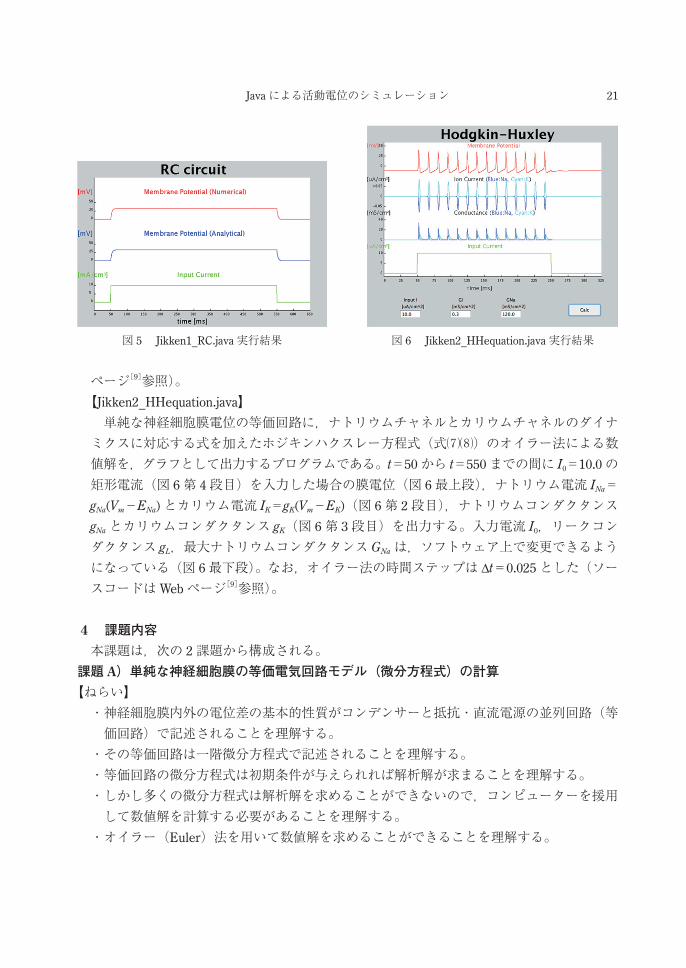
21Javaによる活動電位のシミュレーション
ページ[9]参照)。【Jikken2_HHequation.java】 単純な神経細胞膜電位の等価回路に,ナトリウムチャネルとカリウムチャネルのダイナミクスに対応する式を加えたホジキンハクスレー方程式(式⑺⑻)のオイラー法による数値解を,グラフとして出力するプログラムである。t=50から t=550までの間に I0=10.0の矩形電流(図 6第 4段目)を入力した場合の膜電位(図 6最上段),ナトリウム電流 INa=gNa(Vm-ENa)とカリウム電流 IK=gK(Vm-EK)(図 6第 2段目),ナトリウムコンダクタンスgNaとカリウムコンダクタンス gK(図 6第 3段目)を出力する。入力電流 I0,リークコンダクタンス gL,最大ナトリウムコンダクタンス GNaは,ソフトウェア上で変更できるようになっている(図 6最下段)。なお,オイラー法の時間ステップは Δt=0.025とした(ソースコードはWebページ[9]参照)。
4 課題内容 本課題は,次の 2課題から構成される。課題 A)単純な神経細胞膜の等価電気回路モデル(微分方程式)の計算【ねらい】・神経細胞膜内外の電位差の基本的性質がコンデンサーと抵抗・直流電源の並列回路(等価回路)で記述されることを理解する。
・その等価回路は一階微分方程式で記述されることを理解する。・等価回路の微分方程式は初期条件が与えられれば解析解が求まることを理解する。・しかし多くの微分方程式は解析解を求めることができないので,コンピューターを援用して数値解を計算する必要があることを理解する。
・オイラー(Euler)法を用いて数値解を求めることができることを理解する。
図 5 Jikken1_RC.java実行結果 図 6 Jikken2_HHequation.java実行結果

22 坪 泰 宏,深 作 貴美代,田 代 健 治,糸 井 充 穂
【課題手順】1)単純な神経細胞膜の等価電気回路を記述する微分方程式(式⑴)の解析解を初期条件
Vm(t=50)=0のもとで求める(式⑹)。2)用意されたコンピューター上で統合開発環境 Eclipseを起動し,用意された Javaアプレット(プログラム名:Jikken1_RC.java)を実行する(図 5)。
これらのグラフをワークシートに写し,膜電位 Vmの時間変化を観察する。また,解析解と数値解を比較する。
3)用意された Javaアプレットのプログラムのうち,解析解の計算にかかわる部分を探し出す。反復部分の for文を while文で書き直し,プログラムの実行結果が再現されることを確認する。
課題 B)ホジキンハクスレー方程式の数値計算による神経活動電位の観察【ねらい】・作成済 Javaアプレットによってホジキンハクスレー方程式の数値解を求め,入力電流の増加が神経活動電位にどのような影響を与えるのかを理解する。
・同様に,最大ナトリウムコンダクタンスに対応するパラメーターを0に設定することで,ナトリウムチャネルを閉じてしまう毒物テトロドトキシン(tetrodotoxin : TTX)の効果を予測する。
【課題手順】1)課題 Aに引き続き,統合開発環境 Eclipse上で用意された Javaアプレット(プログラム名:Jikken2_HHequation.java)を実行する(図 6)。
2)入力電流(InputI)を変化させ,初めて活動電位が出る電流強度(Rheobase)を小数第 2位まで調べる。
3)電流を 0.1刻みで変化させ,電流に対する活動電位の本数を記録する。4)Rheobaseの 2倍の電流を入れた時の電位の変化をワークシートに写し,観察する。5)このとき最大ナトリウムコンダクタンスGNaを0にした場合の,電位の時間変化をワークシートに写し,活動電位が発生しなくなることを観察する。
5 演習実践とアンケート結果分析 本課題は最初の 20分程度関連知識を教員が説明し,以降学生同士相談することを許可して課題を行わせた。本章では 2011年 9月 20日から 11月 29日までに行われた 18回の実験を対象に得られたアンケートと学生実験の観察を基に,学生に対する本課題の効果について考察する。アンケートの有効回答数は 61名(男子 41名,女子 20名)であった。アンケートは記名式でレポート提出時に添付してもらった。アンケートの項目は表 2の通りで,単数回答法 5項目,自由回答法 1項目の全部 6項目行った。なお,選択肢の点数はアンケート用紙には記載しなかった。以下では,項目によって該当者数が非常に少数であることや,アン

23Javaによる活動電位のシミュレーション
ケート実施状況の不均一性などを踏まえ,アンケート結果の傾向を述べるにとどめ,統計検定などは行わない。
表 2 学生アンケートの内容(選択肢の点数はアンケート用紙には不記載)
① 今までにプログラムを作った事がありますか?(ある + 1 ない - 1)② JAVAのプログラムを行い,プログラムに興味をもちましたか?(もった + 1 どちらでもない 0 もたない - 1)③ プログラムの構造について理解できましたか?(とても理解できた + 2 理解できた + 1 理解できなかった - 1 全く理解できない - 2)④ 神経の活動電位について,理解が深まりましたか?(とても深まった + 2 深まった + 1 かわらない 0)⑤ 興奮のメカニズムについての物理的解釈を理解できましたか?(とても理解できた + 2 理解できた + 1 理解できなかった - 1 全く理解できない - 2)⑥ この実験に対する感想をおかきください
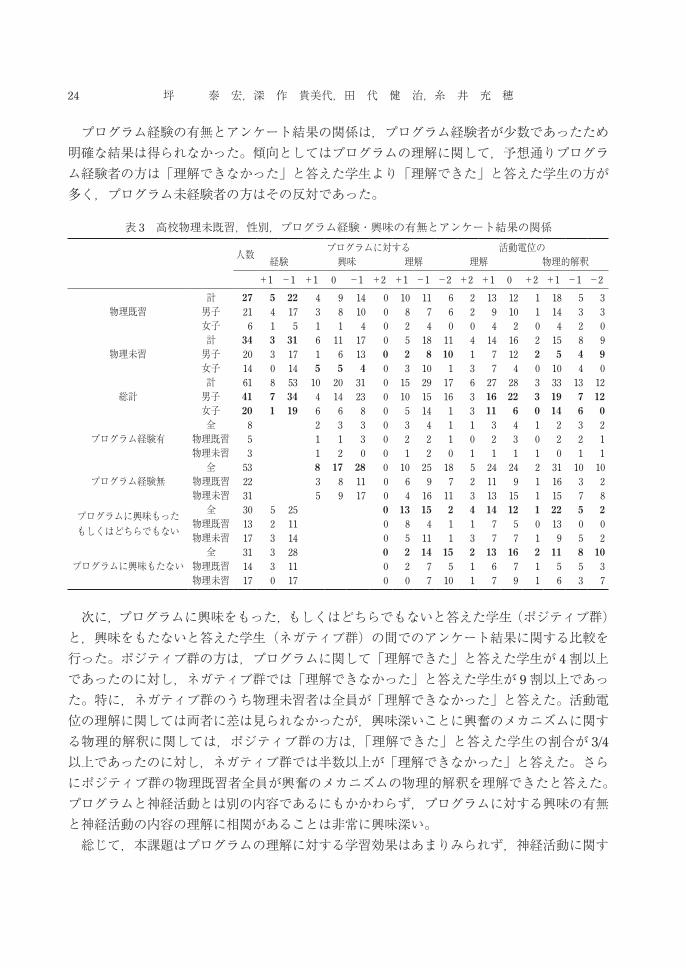
5- 1 性別・高校物理未 /既習とアンケート結果の関係 まず,プログラム経験者の割合は男子と高校物理既習(以後物理既習と表示)女子では約2割であった(表 3)。高校物理未習(以後物理未習と表示)女子にはプログラム経験者はいなかった。また物理未 /既習者の男女構成比を反映して,物理未習者のプログラム経験者の割合が少なかった。 一方で,男子と物理既習女子は「プログラムに興味を持った」,もしくは「どちらでもない」と答えた学生(プログラム興味群)の割合が,「興味を持たなかった」と答えた学生(プログラム無興味群)の割合と同数程度以下であったのに対し,物理未習女子ではプログラム興味群の割合が無興味群の割合の 2倍以上であった。この結果は,本課題はプログラムを初めて眼にする学生にとっては比較的興味を持たせるものであったことを示唆する。ところが興味深いことに,実際にプログラム経験の有無とプログラムに対する興味を調べてみると,プログラム興味群の割合は若干プログラムの経験有の学生の方が高かった。これは,プログラム未経験者のうち物理未習の男子の約 3 /4が「興味持たなかった」と答えたことが影響していると考えられる。 プログラムの理解に関して,多くの学生が理解できなかったと答えた(約 3 /4)。特に,物理未習男子では9割程度の学生が理解できなかったと答えた。活動電位の理解に関しては,女子は 2 /3以上の学生が「理解が深まった」と答えたのに対し,男子は半数以下の学生が「わからなかった」と答えた。同様の傾向は興奮の物理的解釈に関しても見られた。特に,女子と物理既習の男子では「理解が深まった」と答えた学生の割合が「深まらなかった」と答えた学生の 2倍以上いたのに対し,物理未習の男子では逆に「理解が深まった」と答えた学生の割合は「深まらなかった」と答えた学生のほぼ半数であった。
24 坪 泰 宏,深 作 貴美代,田 代 健 治,糸 井 充 穂
プログラム経験の有無とアンケート結果の関係は,プログラム経験者が少数であったため明確な結果は得られなかった。傾向としてはプログラムの理解に関して,予想通りプログラム経験者の方は「理解できなかった」と答えた学生より「理解できた」と答えた学生の方が多く,プログラム未経験者の方はその反対であった。
表 3 高校物理未既習,性別,プログラム経験・興味の有無とアンケート結果の関係
人数 プログラムに対する 活動電位の経験 興味 理解 理解 物理的解釈
+1 -1 +1 0 -1 +2 +1 -1 -2 +2 +1 0 +2 +1 -1 -2
物理既習計 27 5 22 4 9 14 0 10 11 6 2 13 12 1 18 5 3男子 21 4 17 3 8 10 0 8 7 6 2 9 10 1 14 3 3女子 6 1 5 1 1 4 0 2 4 0 0 4 2 0 4 2 0
物理未習計 34 3 31 6 11 17 0 5 18 11 4 14 16 2 15 8 9男子 20 3 17 1 6 13 0 2 8 10 1 7 12 2 5 4 9女子 14 0 14 5 5 4 0 3 10 1 3 7 4 0 10 4 0
総計計 61 8 53 10 20 31 0 15 29 17 6 27 28 3 33 13 12男子 41 7 34 4 14 23 0 10 15 16 3 16 22 3 19 7 12女子 20 1 19 6 6 8 0 5 14 1 3 11 6 0 14 6 0
プログラム経験有全 8 2 3 3 0 3 4 1 1 3 4 1 2 3 2
物理既習 5 1 1 3 0 2 2 1 0 2 3 0 2 2 1物理未習 3 1 2 0 0 1 2 0 1 1 1 1 0 1 1
プログラム経験無全 53 8 17 28 0 10 25 18 5 24 24 2 31 10 10
物理既習 22 3 8 11 0 6 9 7 2 11 9 1 16 3 2物理未習 31 5 9 17 0 4 16 11 3 13 15 1 15 7 8
プログラムに興味もったもしくはどちらでもない
全 30 5 25 0 13 15 2 4 14 12 1 22 5 2物理既習 13 2 11 0 8 4 1 1 7 5 0 13 0 0物理未習 17 3 14 0 5 11 1 3 7 7 1 9 5 2
プログラムに興味もたない全 31 3 28 0 2 14 15 2 13 16 2 11 8 10
物理既習 14 3 11 0 2 7 5 1 6 7 1 5 5 3物理未習 17 0 17 0 0 7 10 1 7 9 1 6 3 7
次に,プログラムに興味をもった,もしくはどちらでもないと答えた学生(ポジティブ群)と,興味をもたないと答えた学生(ネガティブ群)の間でのアンケート結果に関する比較を行った。ポジティブ群の方は,プログラムに関して「理解できた」と答えた学生が 4割以上であったのに対し,ネガティブ群では「理解できなかった」と答えた学生が 9割以上であった。特に,ネガティブ群のうち物理未習者は全員が「理解できなかった」と答えた。活動電位の理解に関しては両者に差は見られなかったが,興味深いことに興奮のメカニズムに関する物理的解釈に関しては,ポジティブ群の方は,「理解できた」と答えた学生の割合が 3/4
以上であったのに対し,ネガティブ群では半数以上が「理解できなかった」と答えた。さらにポジティブ群の物理既習者全員が興奮のメカニズムの物理的解釈を理解できたと答えた。プログラムと神経活動とは別の内容であるにもかかわらず,プログラムに対する興味の有無と神経活動の内容の理解に相関があることは非常に興味深い。 総じて,本課題はプログラムの理解に対する学習効果はあまりみられず,神経活動に関す

25Javaによる活動電位のシミュレーション
る学習効果はある程度見受けられた。また,物理未習の男子に対して比較的学習効果が低かった。
5- 2 電磁気学の試験成績とアンケート結果の関係 続いて,前期講義で行った電磁気学の定期試験の成績とアンケート結果の関係を調べた(表 4)。物理未習者は前期に電磁気学を 6週受講し,既習者は 4週受講した後,それぞれ定期試験を受験している。これらの試験における点数を 5段階に換算し,5群に分類した。成績ランク 5が高得点者群であり,成績ランク 1が低得点者群である。ほとんどの項目で成績とアンケート結果の間に関係は見られなかったが,プログラムの理解と興奮の物理的解釈が理解できなかったと答えた学生のうち物理既習者は,低得点者群の学生が多かった。これは物理既習者対象の定期試験では神経活動電位の等価回路に関する問題を出題しており,その際の理解度が現在に至ってもそのまま反映されていると考えられる(物理未習者対象の定期試験では神経活動電位は出題しなかった)。
表 4 定期試験電磁気学範囲の成績ランクとアンケート結果の関係
成績ランク 5 4 3 2 1 平均 成績ランク 5 4 3 2 1 平均
プログラムに興味(+/0)
計 6 6 3 8 7 2.9活動電位の理解
(+)
計 6 8 4 8 7 2.9物理既習 3 3 1 3 3 3.0 物理未習 2 6 2 4 4 2.9物理未習 3 3 2 5 4 2.8 物理既習 4 2 2 4 3 3.0
プログラムに興味(-)
計 5 6 12 4 4 3.1活動電位の理解
(-)
計 5 4 11 4 4 3.1物理既習 2 2 6 2 2 3.0 物理未習 4 1 6 3 2 3.1物理未習 3 4 6 2 2 3.2 物理既習 1 3 5 1 2 3.0
成績ランク 5 4 3 2 1 平均 成績ランク 5 4 3 2 1 平均
プログラムの理解(+)
計 5 2 2 2 4 3.1 活動電位の物理的解釈(+)
計 8 8 7 7 6 3.1物理既習 4 1 1 2 2 3.3 物理既習 5 4 4 3 3 3.3物理未習 1 1 1 0 2 2.8 物理未習 3 4 3 4 3 3.0
プログラムの理解(- 1)
計 4 8 6 7 4 3.0 活動電位の物理的解釈(-)
計 3 4 8 5 5 2.8物理既習 1 4 3 2 1 3.2 物理既習 0 1 3 2 2 2.4物理未習 3 4 3 5 3 2.9 物理未習 3 3 5 3 3 3.0
プログラムの理解(- 2)
計 2 2 7 3 3 2.8物理既習 0 0 3 1 2 2.2物理未習 2 2 4 2 1 3.2
5- 3 課題実施時期とアンケート結果の関係 さらに,課題の実施時期とアンケート結果との関係を調べた。これは課題導入初期と中期,後期で比較することで,教員側の説明などの原因がアンケート結果に影響している可能性を調べる目的で行った。結果として,実施時期とアンケート結果の間には強い関係は見受けられなかった(データ非表示)。

26 坪 泰 宏,深 作 貴美代,田 代 健 治,糸 井 充 穂
5- 4 自由回答の意見から 最後に,自由回答法から寄せられた意見として,「難しかった」,「説明してもらわないと課題が進められない」,「予習のための資料を欲しい」,などがあった。これらに関しては次章で改善策として考察する。一方で,「生物と物理のつながりがわかってよかった」,「様々な分野が集約されていて面白かった」,というような課題目的が達成されているコメントも見受けられた。また演習時間内に,ある学生が「電流を増加させると活動電位はどうして突然本数が増えるのですか」,「活動電位が発生の有無を,閾値の場合分けなしの 1組の微分方程式によってどうして表現できるのですか」という質問を投げかけてきた。これに関しては8章の「参考」で考察してみたい。
6 今後の改善点 本課題「JAVAによる活動電位のシミュレーション」は,これまで個別の学科として学習してきた医学関連知識の関連性を再認識させることを目的として導入した。学生の実習態度やアンケート結果などをふまえると,課題の詳細にはまだ多くの改善の余地が残されているが,本課題の導入自体には一定の効果が見受けられた。そこで以下では,この課題を生かし,さらに有効なものにするためのいくつかの改善点を挙げる。 まず,教員が実際に実施した際に感じたこと,また学生からのコメントを基にすると,本年度実施形態の問題点は大きく 2点に分類される。 ① 課題に要求される知識の多さとそれに伴う消化不足 ② 課題プロセスの複雑さにより課題意図の不明瞭化つまり,課題内容が絞り切れていなかったために学生がそれぞれのプロセスの意図を消化できなかったことが問題であった。これを踏まえ,次年度以降は以下のような改善策を講じたい。1)学生がしっかりと予習を行えるために実習テキストを改訂する この課題に必要となる生物学,生理学,プログラミング,数値計算,電磁気学に関する知識が各教科のどの講で学習した内容と密接に関係しているかを実習テキストに明示する。さらに,必要な前提知識を確認できるような予習問題をあらかじめ用意しておく。2)課題 Aと課題 Bの順番を入れ替える 最初にシミュレーターを用いた課題 Bを行い,コンピューターシミュレーションで可能なことを概観した上で,その背景知識として課題 Aを行わせる。3)課題 Aで統合開発環境 Eclipseを用いない エディターによるソースコード作成,コンパイラーによるコンパイル,Webブラウザーによる表示,というプロセスを個別に行い,どの段階で何が行われているのかを明確にする。4)課題 Aではさらに単純な微分方程式を扱う 本年度は単純な神経細胞膜の等価回路として,抵抗・コンデンサー並列回路(RC回路)の矩形電流入力に対する応答を解析解と数値シミュレーションで比較させた。このシミュ

27Javaによる活動電位のシミュレーション
レーションには矩形電流入力が時間変化する(入力 ONと OFF)ために,解析解に場合分けが生じてしまい非常に煩雑になってしまった。さらに数値シミュレーション時の時間ステップをあらかじめ十分短い時間幅に固定したために,数値解と解析解の差異や誤差に関して十分な考察をさせることができなかった。次年度は入力 ONのみの緩和過程を取扱い,数値解の時間ステップ幅依存性を調べさせることで,数値解と解析解の差異を明確に意識させる。5)課題 Bのホジキンハクスレー方程式の変更パラメーターを増やす 課題 Bのシミュレーターによるシミュレーションは比較的学生に理解しやすかったようである。本年度は入力電流による活動電位の本数の変化と,ナトリウムコンダクタンスを 0
にすることによるテトロドトキシンの効果の予測を行わせた。次年度はさらに,温度パラメーターやカリウム平衡電位などもソフトウェア上で変更できるようにし,温度と活動電位ダイナミクスの関係,高カリウム血症と電位変化の関係などについても考察させたい。
7 まとめ 本稿では,日本大学医学部 1年次における基礎物理学実験課題「JAVAによる活動電位のシミュレーション」の導入の説明とその報告を行った。医学部生のように非常に多くの知識を必要とされる学生にとって,各知識を有機的につなげ現象をイメージできることは重要である。近年では,廉価なノート型コンピューターと無料のソフトウェアの組み合わせであっても十分な速度で数値シミュレーションや可視化を行うことができ,例えば本課題のような神経活動電位を計算し「眼で見る」ことができるようになった。スーパーコンピューターを用いた非常に緻密な心臓シミュレーションなども行われているが,本質的には同様の計算が行われているわけである。このように今回の実習を通して,学生が医学の進歩に貢献しているコンピューターの利用に関して,詳細はわからないまでも,どのようなことが行われているのかを理解してくれれば幸いである。 最後に,医学教育に携わられている臨床,基礎医学,一般教養の諸先生方から,本稿での試みに関してご助言,苦言などを賜ることができましたら,非常にうれしく思います。本課題を実施するにあたって,内容や実習テキストにアドバイスをいただきました応用システム神経科学分野の山下晶子先生,生理学分野の和田義之先生,生物学分野の大竹伸一先生,阿部健之先生,数学分野の宇田川誠一先生に深く感謝いたします。また,理化学研究所脳回路機能理論研究チームの竹川高志博士には,コンピュータープログラムのソースコードや教授法に関するアドバイスをいただきましたことを感謝いたします。
8 参考 神経細胞の活動電位生成メカニズムに関して,その数理的な側面を中心として補足解説する。
8- 1 一階微分方程式の可視化 以下の一階微分方程式を考える。

28 坪 泰 宏,深 作 貴美代,田 代 健 治,糸 井 充 穂
3( )dx
f x x xdt
= = − + ⑽
いま,球が x軸上の位置 xにあり,球の速度dx/dtは球の位置 xに依存して微分方程式⑽によって与えられるとする。この球は,図 7のように x<-1または 0<x<1では dx/dt>0なので右に動き,-1<x<0または x>1では dx/dt<0なので左に動くことがわかる。よって,ある時刻で x<0にある球は x=-1へ向かい,x>0にある球は x=1へ向かう。x=±1は dx/
dt=0なのでここに到達した球は静止する(安定固定点)。x=0も dx/dt=0であるので厳密に x=0にある球は静止するが,少しでもずれると x=0から離れる方向に動く(不安定固定点)ので実際にはほとんど観測されない。ちょうど安定固定点が谷底,不安定固定点が山の頂上のようなイメージである。 ここで,いくつかの重要なことがわかる。速度が 0 (f(x)=0)となる点には集まる点(安定固定点)と離れる点(不安定固定点)がある。そして,一変数の一階微分方程式はどの点にあってもしばらくすると安定固定点に向かうか,もしくは無限遠(x=±∞)へ飛んでいく。最終的にどこへ向かうか(ベイスン)は不安定固定点(か不連続点)で区切られる,ということである。つまり,一変数の一階微分方程式の挙動をイメージしたければ,f(x)=0である点(と f(x)が不連続で符号が変わる点)を求め,その点が安定固定点であるかどうか(符号の変化)を調べればよい。 さて,話を式⑵に戻す。式⑵は以下の形に変形できる。
( )m Lm L
m L
dV g I tV E
dt C g
⎛ ⎞⎛ ⎞= − − +⎜ ⎟⎜ ⎟
⎝ ⎠⎝ ⎠ ⑾
式⑾は,式⑽でいう f(Vm)が傾き負の直線であり,Vm=EL+I(t)/gLで Vm軸と交わる。従って,任意のVmからスタートしても常にEL+I(t)/gLへ緩和することが解析解を求めなくてもイメージできる(当然 I(t)があまりに速く変化し,変化の時定数が Vmが緩和する時定数 Cm/gLよりも小さい場合には Vmは EL+I(t)/gLを追従できない)。実際,解析解である式⑹はそのようになっている。
図 7 速度 f(x)のグラフと球の位置の時間経過

29Javaによる活動電位のシミュレーション
8- 2 ホジキンハクスレー方程式の近似的可視化 次にホジキンハクスレー方程式(式⑺)を考える。
( ) ( ) ( ) ( )mm L m L Na m Na K m KdV
I t C g V E g V E g V Edt
= + − + − + −
ここで gNa,gKは膜電位に依存して時間変化する。この式は,( )m l Na K l l Na Na K K
mm l Na K
dV g g g g E g E g E I tV
dt C g g g⎛ ⎞+ + + + +
= − −⎜ ⎟+ +⎝ ⎠と書き換えられるので,前節の結果から,膜電位は各チャネルの平衡電位の(コンダクタンスの比での)重心に向かうことがイメージできる。膜電位が平衡膜電位(-75[mV])付近の場合,gNaはほとんど 0であり,膜電位は ELと EKの重心付近に向かう,すなわち平衡膜電位付近に滞在する。一方,シナプス入力などの電流入力 I(t)により重心が脱分極側にずらされ,膜電位が閾値電位(-40[mV])付近に達すると,gNaが急激に大きくなり,重心はほとんど ENaとなる。すなわち活動電位が生じ,膜電位が+30[mV]程度まで上昇する。この電位に達すると,gNaはまた小さくなり,gKが大きくなる。その結果,重心は平衡膜電位付近となり,膜電位は急激に減少する。 電流 I(t)についてであるが,通常の電気生理実験では電流固定法により,電流を生体用アンプで制御し,応答膜電位を記録する(図 8)。ところが生体内の神経細胞では,ギャップジャンクション(gap junction)と呼ばれる細胞同士の電気的な結合は存在するものの,一般的には化学シナプスや代謝型イオンチャネルといった化学的信号によって電流が生じる。主なメカニズムはイオンチャネルの開閉であり,化学物質に依存して変化するコンダクタンスとイオン濃度差による起電力である。例えば,興奮性神経伝達物質のグルタミン酸の最も一般的な受容体である AMPA型受容体では,IAMPA=gAMPA(Vm-ECat)のコンダクタンス gAMPA
がグルタミン酸の濃度によって変化する。余談であるが,この化学シナプスが作る電流を上述の生体用アンプで入力する際には,入力する対象の膜電位 Vmが必要になることがわかる。膜電位を計測するために膜電位が必要なこのような系はフィードバック系と呼ばれ,実際には微小時刻前の膜電位を使って電流信号を生成,入力し,次時刻の膜電位を記録することを行う。このため,高速で電気信号をフィードバックするシステムが必要となる(ダイナミッ
図 8 神経細胞の電気応答を記録するシステムと神経細胞の近赤外微分干渉顕微鏡像

30 坪 泰 宏,深 作 貴美代,田 代 健 治,糸 井 充 穂
ククランプ)。
8- 3 分岐と場合分け 神経活動電位の発生と膜電位の閾値について考察する。活動電位を記述する最も単純なモデルのひとつに LIF(Leaky-Integrate and Fire)モデルがある。このモデルは式⑴に閾値を設定し,膜電位が閾値を超えたら活動電位を生成し,静止膜電位にリセットする,というモデルである。LIFモデルでは場合分けによって活動電位を表現する。ところが,ホジキンハクスレー方程式には閾値も場合分けもない,連立非線形常微分方程式である。なぜ,ホジキンハクスレー方程式から活動電位が生じるかの説明は他の文献に譲るが[4][11],簡単な微分方程式でも場合分けなしで突然解の挙動が変わりうることを説明する。 以下の一階微分方程式を考える。
dxf x x x a a
dt3( ) 3 ( 0)= = − + + ≥ ⒁
図 7と同様の考察を行うと,1つの微分方程式⒁から aの値によりグラフの形が変わるので,「場合分け」の必要が生じることがわかる(図 9)。a<2では,dx/dt=f(x)の安定固定点は 2
つあるが,a>2では安定固定点の数が 1つになる。そこで,a<2のときは最初に x<-1に球があれば小さい方の安定固定点に向かうが,a>2のときは最初に x<-1に球があっても突然離れた安定固定点に向かう。ここで,aを入力電流,xを膜電位と考えれば,活動電位のように電位の上昇と下降は見られないが,閾値が明示的に書かれていない微分方程式でも,「閾値」は存在し(今の場合 a=2),その値を超えると解の挙動が定性的に変化することがわかる。これを分岐と呼ぶ。いまの例は接線分岐と呼ばれるものである。ちなみに,ホジキンハクスレー方程式で入力電流を増加させていくと,ホップ(Hopf)分岐と呼ばれる分岐が
図 9 分岐と速度 f(x)のグラフ

31Javaによる活動電位のシミュレーション
生じ,周期的に活動電位が発生する。この周期的な活動電位は,(Vm, m, h, n)の 4次元空間内では輪のような軌道(リミットサイクル;limit cycle)となっている。安定固定点と同じように,近くのものはこのリミットサイクルに緩和する。ちなみに入力電流を増やしたときに,活動電位の本数が突然増える前の小さな電流の時の活動電位はリミットサイクルではなく,安定固定点からずれた軌道が戻る過渡的な現象を観測したものである。従ってしばらくすると活動電位は止まってしまう。一般に Hopf分岐により説明される活動電位は,入力電流の増加に伴い不連続に本数が増加する( 2型興奮性)。一方,大脳皮質の神経細胞に多いサドルノード(saddle-node)分岐によって説明される活動電位は,連続的に本数が増加する( 1型興奮性)。これらは同期傾向が異なることが報告されているが,実際の脳回路の情報処理においてどのような意味があるのかはよくわかっていない。
文献[ 1] JG Nicholls, AR Martin, BG Wallace, PA Fuchs. From Neuron to Brain : A Cellular and Molecular Ap-
proach to the Function of the Nervous System. 4th. Ed.Sinauer Associates Inc. (2001),本間研一,大森治紀,大橋俊夫編集.標準生理学第 7版.医学書院(2011)
[ 2] EC Conley, WJ Brammar. Ion Channel Fatcsbook. Vol 1~4. Elsevier. (1999), B Hille. Ion Channels of Ex-
citable Membranes. 3rd. Ed. Sinauer Associates Inc. (2001)
[ 3] BW Connors, MJ Gutnick. Intrinsic firing pattern of diverse neocortical neurons. Trends Neurosci 13 : 99-104 (1990), E Dégenètais, AM Thierry, J Glowinski, Y Gioanni. Electrophysiological Properties of Pyra-
midal Neurons in the Rat Prefrontal Cortex : An In Vivo Intracellular Recording Study. Cereb Cortex 12 :
1-16 (2002), A Mason, A Larkman. Correlations between morphology and electrophysiology of pyrami-
dal neurons in slices of rat visual cortex. II. Electrophysiology. J Neurosci. 10 : 1415-1428 (1990)
[ 4] P Dayan, LF Abbott. Theoretical Neuroscience : Computational and Mathematical Modeling of Neural
Systems. MIT Press. (2001), C Koch. Biophysics of Computation : Information Processing in Single Neu-
rons. Oxford Univ Press. (2004),宮川博義,井上雅司.ニューロンの生物物理.丸善(2003)[ 5] A Destexhe, M Rudolph, D Paré. The high-conductance state of neocortical neurons in vivo. Nat Rev
Neurosci. 4 : 739-751 (2003)
[ 6] AL Hodgkin, AF Huxley. J Physiol (London) 117 : 500-544 (1952)
[ 7] WH Press et al.著,丹慶勝市ら訳.ニューメリカルレシピ・イン・シー日本語版―C言語による数値計算のレシピ.技術評論社(1993)
[ 8] BK Oksendal. Stochastic Differential Equation. Springer. (2003), C Gardner. Stochastic Methods : A
Handbook for the Natural and Social Sciences. Springer (2010)
[ 9] http://nct.brain.riken.jp/~tsubo/educ/JikkenHH.html
[10] http://www.eclipse.org/,http://mergedoc.sourceforge.jp/
[11] EM Izhikevich. Dynamical Systems in Neuroscience : The Geometry of Excitability and Bursting. MIT
Press (2006)





























