Embed Size (px)
Citation preview

ELSEVIER
Interphase Cytogenetics in Chronic Lymphocytic Leukemia
Jose A. Garcia-Marco, Cathy M. Price, and Daniel Catovsky
ABSTRACT: The incidence of trisomy 12 and 13q12-qi4 abnormalities in patients with chronic lym- phocytic leukemia (CLL) was determined by conventional cytogenetics and interphase fluorescence in situ hybridization (FISH). In the analysis of 580 consecutive patients, trisomy 12 was detected by con- ventional cytogenetics in 39 cases (9%) and 117 cases (20%) by FISH. Trisomy 12 was shown to be asso- ciated with advanced clinical stage, atypical morphology, and higher proliferative activity. Combined immunophenotyping and FISH showed that trisomy 12 was present only in a proportion of the clonal B-cells. These data suggest that trisomy 12 is a secondary event associated with features of disease pro- gression. Sequential FISH showed clonal progression of the trisomic clone over time. Three hundred patients also were investigated for 13q deletions using FISH analysis of the RB1 locus (13q14). Monoal- lelic RB1 deletion was seen in I04 (34%) of cases. One case had a homozygous deletion in 90% of the cells. Dual-color FISH detected the presence of trisomy 12 and RBI in 17 (5%) cases. DNA probes for 13q12.3 (BRCA2) and 13q14 (RB1 and DBM locus) were used in 35 cases. Twenty-eight (80%) cases showed deletion of a 1Mb 13q12.3 encompassing the BRCA2 locus, whereas 22/35 (63%) were deleted at 13q14. Our data suggest that abnormalities of l3q are more frequent than trisomy 12 in CLL and pro- vide evidence for the presence of a new candidate gene at 13q12.3 that may be involved in the patho- genesis of CLL. © Elsevier Science Inc., 1997
INTRODUCTION
Chronic lymphocytic leukemia (CLL) is the most common leukemia in humans. The disease is characterized by: a clonal proliferation of mature-appearing B Lymphocytes; the common phenotype CD5/CD19+; definite bone mar- row involvement; and a presentation of lymphocytosis, lymphadenopathy or splenomegaly.
Leukemic B lymphocytes usually show a relatively slow growth rate, and CLL often follows a benign course with a long-standing stable disease. However, in some patients there is a higher production of tumor cells leading to a rapidly progressive course and short survival. The cellular mechanisms underlying this heterogeneous clinical course are not known [1].
The low spontaneous mitotic rate of the leukemic cells in CLL has made chromosome analysis difficult. Cytoge-
From the Academic Department of Hematology and Cytoge- netics, The Royal Marsden Hospital and The Leukemia Research Fund Centre, Chester Beatty Laboratories, The Institute of Cancer Research, London, UK.
Address reprint requests to: Jose A. Garcia-Marco, Academic Department of Haematology and Cytogenetics, The Royal Mars- den Hospital, Fulham Road, London SW3 6JJ, UK.
Received August 26, 1996; accepted September 4, 1996.
netic preparations commonly yield metaphases of poor quality that are unsuitable for karyotype analysis. The ad- dition of mitogens may increase metaphase number but of- ten results in the selective stimulation of normal B or T lymphocytes rather than the leukemic cells. Overall, of cases successfully karyotyped, 50% have a clonal abnormal- ity. The most frequent cytogenetic abnormalities reported to date are trisomy 12 and structural abnormalities of chromosome 13 [2]. The significance of these two types of abnormality in the pathogenesis of CLL has not been es- tablished, although several cytogenetic studies indicate that patients with trisomy 12 showed a poorer outcome than cases those with 13q abnormalities [3].
The introduction of fluorescence in situ hybridization (FISH) has had a major impact on the capabilities of cyto- genetic analysis in CLL. The analysis of nonmitotic nuclei (interphase FISH) has been of particular value because it does not rely on an actively dividing cell population and it can be used on fresh patient material without the artefacts of in vitro cell culture. Consequently, an accurate determi- nation of the frequency of clonal chromosome abnormali- ties in a large number of cells is possible [4].
The aim of our investigation is to evaluate the signifi- cance of trisomy 12 and 13q14 deletions in CLL using in- terphase and metaphase FISH and to correlate our laboratory findings with clinico-biologic characteristics.
Cancer Genet Cytogenet 94:52-58 (1997) © Elsevier Science Inc., 1997 655 Avenue of the Americas, New York, NY 10010
0165-4608/97/$17.00 PII S0165-4608(96)00304-4

Interphase Cytogenetics in CLL 53
INTERPHASE CYTOGENETICS FOR THE DETECTION OF TRISOMY 12
Conventional cytogenetics has identified trisomy 12 as the most frequent CLL abnormality, found in about 10% to 15% of all patients and 15% to 28% of cases with evalu- able metaphases. It was generally thought that this inci- dence might be underestimated because of the low mitotic rate, the use of mitogen added to cell cultures, or other technical factors that might influence the yield of analyz- able metaphases. Several studies using interphase FISH support this hypothesis and report a variable incidence of trisomy 12 ranging from 11% to 35% [5-13]. The reasons for this variability might be attributed to selection criteria, the size of the sample, or less likely to different cytoge- netic characteristics in the studied population.
We have performed interphase FISH in peripheral blood samples from a group of 580 consecutive CLL pa- tients submitted for diagnosis or treatment. In 350 cases karyotype studies also were performed. Overall, the inci- dence of trisomy 12 by FISH was 20% (117/580) in con- trast to 9% (32/350) detected by conventional cytogenetics. All cases trisomic for chromosome 12 by cytogenetics were confirmed by FISH. An additional group of 37 cases displaying a normal karyotype or other clonal abnormali- ties also exhibited trisomy 12 in 6% to 59% of the cells. The cut-off level for definition of trisomy 12 was 2% based on published criteria (mean % of 3 signals _+ 3 SD on nor- real controls). Of 117 cases with trisomy 12, more than 90% had a population of trisomic cells higher than 20%. The mean proportion of trisomic cells was 48% ranging from 7% to 85%. These findings clearly demonstrate that FISH is a much more sensitive technique than standard cytogenetics for the detection of numerical abnormalities in CLL. However, based on the analysis of previous cyto- genetic reports, this incidence is to some extent lower than would be expected, as by cytogenetics one-third of the cases with clonal abnormalities showed trisomy 12, whereas by FISH it is found in about 20% of all the cases.
MONITORING OF TRISOMY 12 CLONE
As in other hematologic malignancies there is increasing evidence of clonal evolution in CLL [14-16]. Clonal karyo- typic abnormalities are found both at diagnosis and during the course of the disease. Patients with complex aberra- tions have shorter survival than cases with a normal kary- otype [3]. The presence of trisomy 12 as a result of clonal evolution has been described in few cases, perhaps because of the resistance of the trisomic clone to mitogens and sub- sequent failure to divide or other artefactual factors. From the group of 350 cases with both cytogenetic and FISH studies, we found 7 cases with an abnormal karyotype without tri- somy 12 and additional FISH showed a substantial popu- lation of trisomy 12 cells. All these cases were associated with advanced disease. Moreover, 20 cases with a normal karyotype also had trisomy 12 by FISH in more than 20% of the cells analyzed. Therefore, it appears that by sequen- tial cytogenetics alone trisomy 12 is not detected in some cases and perhaps the number of cases undergoing clonal
evolution is more common than reported and CLL is not a cytogenetically stable disease, as previously thought [17].
We also performed sequential FISH over a 4-year pe- riod in 30 CLL cases. Fourteen cases had trisomy 12, 13 at diagnosis and 1 case in which trisomy 12 was detected during the course of the disease. Fifteen of 16 patients with a normal karyotype at diagnosis remain disomic for chromosome 12. One of 16 cases acquired an extra copy of chromosome 12 in 45% of cells during disease progres- sion. All the cases with trisomy 12 showed progression of the trisomic clone in sequential analysis. Eight cases reached a partial remission after treatment with different combinatory regimes and exhibited a mean percentage of trisomic cells lower than 5%. Four patients died of disease progression showing 68%, 71%, 76%, and 84% of the neoplastic population with 3 copies of chromosome 12. Two patients with resistant disease displayed a minor ini- tial response and a gradual increase of trisomy 12 cells. Although it has been reported that the trisomic clone does not expand when studied over time [7, 18], our findings suggest that there is a clonal expansion of trisomy 12 cells in CLL as the disease progresses. Nine of these patients had a CLL/prolymphocytes (PL) morphology (CLL with more that 10% prolymphocytes) as opposed to 5 cases who showed a typical CLL morphology. Thus, there is a clear association between morphologic transformation and clonal expansion of trisomy 12 suggesting a correlation of this clonal abnormality and disease progression.
Our findings are in agreement with those of Cuneo et al. [19], who reported sequential FISH in 2 CLL cases and showed the persistence of the trisomic clone following chemotherapy in cases that failed to respond. In our series there was a dramatic reduction of trisomy 12 cells in those cases who achieved a partial remission.
Trisomy 12 has been reported to be associated with transformation from typical B-CLL to high-grade lympho- proliferative disease (Richter's syndrome) in a few cases [20]. We have detected one case acquiring trisomy 12 dur- ing transformation to Richter's syndrome. Initially, this patient had a complex karyotype lacking trisomy 12. FISH analysis on bone marrow cells showed 75% large B-CLL cells with trisomy 12 as opposed to 45% detected in the peripheral blood. These data support the concept that tri- somy 12 might play a direct role in the transformation of CLL to a more aggressive disease or diffuse high-grade lymphoma. According to Nowell's model of tumor pro- gression it is likely that trisomy 12 cells arose as a result of acquired genetic instability (mutation, gene activation, or nondisjunction mechanisms) in the expanding neoplastic cells during evolution of the tumor as part of a multistep process resulting in a proliferative advantage to trisomy 12 cells [21].
CLINICAL CORRELATIONS OF TRISOMY 12
Several reports using conventional cytogenetics of FISH have reported trisomy 12 to be associated with a poorer survival than cases with normal karyotype or other single clonal abnormalities [3, 10]. However, the association of trisomy 12 with other clinicobiologic characteristics, such

54 J.A. Garcia-Marco et al.
as antigen expression pattern, clinical stage, cell prolifera- tion, lymphocyte doubling time, and survival, are not in agreement in all the series published. We have examined the association of trisomy 12 with clinical characteristics in a subset of 370 patients. Of them, 76 cases had trisomy 12. As in other studies, atypical B-cell morphology was strongly correlated with the presence of trisomy 12 (52 cases versus 24 cases with typical CLL morphology) [18, 22, 23]. The majority of the trisomy 12 cases with atypical morphology had an increased number of prolymphocytes (CLL/PL) (p < .0001) [24]. There also was a positive corre- lation between the number of prolymphocytes and the proportion of trisomy 12 cells.
We also examined the relationship between clinical stage and trisomy 12 in this group of patients. Trisomy 12 was associated with more advanced clinical stage than cases with diploid chromosome 12 (45 cases stage B and C versus 31 cases with stage A) (p = .0005). Furthermore, patients with stage C exhibited a higher percentage of tri- somic cells (47%) than patients with stage A (38%).
The proliferative activity of CLL cells also was ana- lyzed by measuring the expression pattern of the prolifer- ation-associated nuclear antigen recognized by Ki-67 and correlated with chromosomal abnormalities in 370 pa- tients. Although in general CLL cells showed a low pro- liferative activity, the proportion of Ki-67-positive cells was significantly higher in patients with trisomy 12 (4.23 + 4.37) than in cases displaying a normal karyotype (1.68 + 1.89), other clonal abnormalities (1.37 + 1.40), or no meta- phases (1.18 _+ 1.57)(p < .0001) [25]. In summary, we have observed that trisomy 12 in CLL patients is associated with atypical morphology, higher proliferation rate, and a more advanced clinical stage than in cases without tri- somy 12. Therefore, these findings suggest that trisomy 12 is a secondary event in CLL and also provide evidence of the association of trisomy 12 with features related to dis- ease progression.
IS TRISOMY 12 A PRIMARY OR SECONDARY EVENT IN CLL?
The molecular mechanisms leading to trisomy 12 in CLL remain unclear. Karyotypes with trisomy have long been reported in hematologic malignancies and occasionally in association with disease progression [26]. It has been sug- gested that trisomy contributes to the neoplastic phenotype through a gene dosage effect [2, 27]. In CLL, it is postulated that trisomy 12 arises from the duplication of one chromo- some 12 with the retention of the other copy [28, 29]. How- ever, none of the genes located in chromosome 12 (RAS, ETV6, KIP1, MDM2) have been clearly found to be involved in CLL, although the mechanism of an extra dose of a defec- tive gene conferring a growth advantage during the evolution and expansion of the leukemia clone, such as the ALL1 gene in acute leukemia [30], seems to be the most likely molecular event for the occurrence of trisomy 12 in CLL.
Although trisomy 12 also is seen in a substantial num- ber of patients with stage A (31 of 76 in our series), the fact that its presence is mainly associated with features of dis- ease progression suggests the hypothesis that it is a sec-
ondary rather than a primary event in leukemogenesis in CLL. Previous studies on lineage restriction in CLL based on mitogen-stimulated cell cultures with the morphology- antibody-chromosomes (MAC) method have shown that all metaphases with trisomy 12 were of B-cell origin and all normal metaphases had a T-cell phenotype [31]. How- ever, this method relies on the presence of metaphase spreads, giving a more limited information of the entire clonal population.
We performed simultaneous inmunophenotyping and FISH on interphase cells in a subset of 20 CLL patients to evaluate the lineage restriction of trisomy 12 in a large population of cells without the in vitro stimulus of mito- gen-induced cultures, which in fact might select or induce preferential proliferation of the neoplastic clone, leading to a relative increase of trisomy 12 cells [32, 33]. Because these techniques allow the simultaneous assessment of both cell type and genotype in a single cell, we were able to demon- strate in each patient that trisomy 12 is absent in T lym- phocytes and nonclonal B cells and to confirm its presence in a variable proportion of clonal B cells (range: 30% to 76%) and that the remaining clonal population is disomic for chromosome 12 [34]. The absence of trisomy 12 in T cells and the mosaicism seen in the clonal B-cell popula- tion suggest that this abnormality is a secondary event during the leukemic transformation of CLL and develops in an already established neoplastic B-cell population.
13q CHROMOSOMAL ABNORMALITIES
Cytogenetic studies have shown that structural abnormali- ties of the long arm of chromosome 13 are the most com- mon structural aberrations in CLL (variable incidence of 15% to 30% depending on the published series). In some studies, 13q abnormalities are as frequent as trisomy 12 or even higher. Technical factors or different patient popula- tion may account for this variability. In a review of the lit- erature, we analyzed the breakpoint patterns of 13q in 138 CLL cases published to date. Sixty-eight cases had bal- anced chromosomal translocation affecting the 13q12- 13q32 region, and in 73% of them the 13q14 band, where the retinoblastoma susceptibility gene RB1 is located, was involved. The remaining 70 cases displayed unbalanced chromosome 13q deletions with breakpoints at 13q12 in nearly 60% of the cases and at 13q14 in 31% [14, 35-47]. The extent of the deletion varies, but nearly always includes the 13q14 band.
It has been suggested that the RB1 gene located at 13q14.2 [48, 49] plays a role in leukemogenesis in CLL based on allelic loss analysis determined by Southern blot and FISH [39, 50-52]. The authors demonstrated monoal- lelic loss of RB1 in 30% and 31% of the cases, respec- tively. However, inactivation of the RB1 has not been reported in CLL [50, 53]. Because of the consistent break- point associated with the 13q14 band, it has been thought that genes located at or near RB1 are presumably impor- tant in leukemogenesis. Somatic cell hybrids containing 13q14 translocations were made and showed that loss of heterozygosity at 13q14 is frequently found [41]. Several groups have demonstrated that markers lying more distally

Interphase Cytogenetics in CLL 55
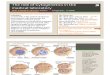
than RB1 often were hemi- or homozygously deleted and frequently encompassed the D13S25 locus, where a putat ive tumor suppressor locus, DBM (deleted in B-cell malignan- cies), has been assigned (Figure 1) [41, 43, 50, 51, 54-56].
There is some discrepancy regarding the incidence of the association of t r isomy 12 and 13q chromosomal abnor- mali t ies in CLL. Therefore, we have performed dual-color interphase FISH with a pericentromeric probe for chromo- some 12 and a unique sequence DNA probe for RB1. This probe was chosen because 13q14 seems to be the region more frequently rearranged or deleted. We s tudied a group of 300 CLL patients to evaluate the inc idence of RB1 dele- t ion and the percentage of cases carrying both t r isomy 12 and allelic RB1 loss. Tr isomy 12 was detected in 61 cases (20%) and monoal le l ic RB1 loss in 104 cases (34%). One case had homozygous delet ion of RB1 in more than 90% of the cells. All cases had allelic loss in more than 10 of the cells (mean: 49%, range: 12%-82%). The cut-off level to define RB1 delet ion was established when more than 3% of the cells had one hybridizat ion signal (based on previously described criteria).
Tr isomy 12 and 13q chromosomal abnormali t ies have been rarely found associated in CLL. Indeed, this might
suggest that they represent two different molecular events occurring in CLL and that its pathogenic role in the dis- ease might follow dis t inct pathways. In our series we have detected 17 cases (5%) carrying both t r isomy 12 and monoal le l ic RB1 deletion. Interphase FISH in these 17 cases enabled us to analyze the clonal dis t r ibut ion of both abnormali t ies in the neoplast ic populat ion, to determine the actual size of any of the clones or subclonal populat ions carrying any of these two aberrations, and also to resolve more accurately which clone was the first cytogenetic event for each patient. This was accompl i shed by scoring an average of 500 cells in each case. In 3 cases t r isomy 12 and monoal le l ic RB1 dele t ion were found as separate clones and three different clones were present in each pa- tient: one clone disomic for both chromosome 12 and RB1, one clone with t r isomy 12 and d ip lo id RB1, and another clone with monoal le l ic RB1 delet ion and d ip lo id chromo- some 12 (Figure 1). In the 3 cases t r isomy 12 cells repre- sented the larger clone. This suggests that in these cases RB1 delet ion is the secondary and acquired cytogenetic event in a smaller popula t ion of cells. The other 14 cases exhibi ted a variable propor t ion of cells carrying both ab- normalit ies. In 6 cases t r isomy 12 cells were the larger
Figure 1 Dual-color FISH in CLL cells from a patient with trisomy 12 (green color) and RB1 deletion (red color) as part of the same clone. The main clone had trisomy 12 and RB1 deletion. There also were subclones with tri- somy 12 and RB1 disomy and monoalleic RB1 loss and chromosome 12 disomy.

56 J.A. Garcia-Marco et al.
Hemi / Homozygous :80 % Hemizygous : 20 - 34%
Hemi / Homozygous : 13 % / 44 %
BRCA2 RB1
D13S319
Hemi / Homozygous :25% / 60% D13S25
Hemizygous : 44% D13S294
Figure 2 Incidence of allelic loss at 13q12.3 and 13q14 in CLL (see text for references).
11.1
12.1 12.2 12.3 13
14.1 14.2 14.3
21.1
21.2
21.3
22
31
32
33
34
clone with a smaller subclone exhibiting RB1 deletion alone or associated with trisomy 12. Eight cases showed RB1 as the larger clone. The fact that none of these 2 ab- normalities have a consistent pattern of presentation sug- gests that either trisomy 12 or RB1 deletion are secondary events in B-CLL leukemogenesis. Mould et al., using a cosmid probe (D13S25) that lies telomeric to RB1, found similar results in 8 patients with trisomy 12 and 13q rear- rangements [46]. In contrast, Arif et al., in a series of 42 Japanese CLL patients, found trisomy 12 and RB1 deletions in different group of patients [44].
Structural abnormalities of chromosome 13 have been associated with early clinical stage, typical morphology, and as good a prognosis as cases with normal karyotypes [3, 23, 57]. Clinical correlations of RB1 deletion in our se- ries showed that, although there are more cases in stage A disease, these differences were not statistically significant when compared with stages B and C. Indeed, 42 of 104 pa- tients with RB1 deletion had stage B or C, and 26 showed an atypical CLL morphology. The latter is of interest, as none of the studies comparing 13q abnormalities with CLL morphology have found any case with atypical morphol- ogy [18, 47, 58].
Several studies have focused on the analysis of the loss of heterozygosity (LOH) at the DBM locus in CLL, which lies approximately 1.6 cM distal to RB1. Several microsat- ellite markers located in that area have been found to be more frequently deleted (both hemi- and homozygously) than RB1 [41, 43, 50, 51, 54, 55]. These findings strongly suggest that this area might be the target for the presence of a tumor-suppressor gene involved in the pathogenesis of CLL. However, cytogenetic studies show that a high proportion of the 13q chromosomal abnormalities have a breakpoint at 13q12, where the recently cloned breast can- cer susceptibility gene BRCA2 is located [59, 60].
Based on the pattern of different breakpoints at chromo- some 13q we have performed interphase in situ hybridiza- tion in 35 CLL patients using a set of probes to characterize allelic loss at 13q12.3 (BRCA2) and 13q14 (RB1 and DBM locus). Dual hybridization with BRCA2 and 13q14 probes was performed in all the cases. Monoallelic deletion of RB1 was detected in 34% (12/35) of the cases. One patient had homozygous RB1 deletion in more than 90% of the cells. A cosmid D13S25 for the DBM locus was found de- leted in 63% (22/35) of the cases. Seven cases exhibited homozygous deletion in a proportion of cells. We also used 7 PAC genomic DNA probes, including a PAC clone and a plasmid probe for BRCA2, to analyze allelic loss at 13q12.3 locus. A minimal deletion consensus region of 1 Megabase encompassing BRCA2 was seen in 80% (28/35) of the patients. In 20 cases, hemi- and homozygous BRCA2 deletion was seen in a proportion of cells, and one case had homozygous BRCA deletion [61]. Our findings suggest that allelic deletion at 13q12.3 encompassing BRCA is the most frequent genetic event at chromosome 13q in CLL (Figure 2) and that BRCA or another underlying gene might play a role in the pathogenesis of CLL.
This work was supported by The Leukemia Research Fund of Great Britain (J.A.G.-M.) and the Key Kendall Leukemia Fund (C.M.P.).
REFERENCES
1. Rozman C, Montserrat E (1995): Chronic lymphocytic leuke- mia. N Engl J Med. 333:1052-1057.
2. ]uliusson G, Gahrton G (1990): Chromosome aberrations in B-cell chronic lymphocytic leukemia. Pathogenetic and clin- ical implications. Cancer Genet Cytogenet 45:143-160.
3. Julinsson G, Gahrton G, Oscier D, Fitchett M, Ross F, Brito-

Interphase Cytogenetics in CLL 5 7
Babapulle V, Catovsky D, Knuutila S, Elonen E, Lechleitner M, Tanzer J, Schoenwald M, Castoldi GL, Cuneo A, Nowell P, Peterson L, Kay N (1991): Cytogenetic findings and survival in B-cell chronic lymphocytic leukemia. Second IWCCLL compilation of data on 662 patients. Leuk Lymphoma 21-25.
4. Price CM (1993): Fluorescence in situ hybridization. Blood Rev 7:127-134.
5. Perez Losada A, Wessman M, Tiainen M, Hopman AHN, Wil- lard HF, Sol~ F, Caballfn MR, Woessner S, Knuuti]a S (1991): Trisomy 12 in chronic lymphocytic leukemia: An interphase cytogenetic study. Blood 78:775-779.
6. Anastasi J, Le Beau MM, Vardiman JW, Fernald AA, Larson RA, Rowley JD (1992): Detection of trisomy 12 in chronic lymphocytic leukemia by fluorescence in situ hybridization to interphase cells: A simple and sensitive method. Blood 79:1796-1801.
7. Raghoebier S, Kibbelaar R, Kleiverda IK, Kluin-Nelemans JC, van Krieken JHJM, Kok F, Kluin PhM (1992): Mosaicism of trisomy 12 in chronic lymphocytic leukemia detected by nonradioactive in situ hybridization. Leukemia 6:1220-1226.
8. D6hner H, Pohl S, Bulgay-M6rschel Stilgenbauer S, Bentz M, Lichter P (1993): Trisomy 12 in chronic lymphoid leuke- mias--A metaphase and interphase cytogenetic analysis. Leukemia 7:516-520.
9. Que TH, Garcia-Marco I, Ellis J, Matutes E, Brito-Babapulle V, Boyle S, Catovsky D (1993): Trisomy 12 in chronic lympho- cytic leukemia detected by fluorescence in situ hybridization: Analysis by stage, immunophenotype, and morphology. Blood 82:571-575.
10. Escudier SM, Pereira-Lehay JM, Drach JW, Weier HU, Good- acre AM, Cork MA, Trujillo JM, Keating MJ, Andreeff M (1993): Fluorescent in situ hybridization and cytogenetic studies of trisomy 12 in chronic lymphocytic leukemia. Blood 81:2702-2707.
11. Bienz N, Cardy DLN, Leyland MJ, Hulten MA (1993): Trisomy 12 in B-cell chronic lymphocytic leukemia: An evaluation of 33 patients by direct fluorescence in situ hybridization (FISH). Br J Haematol 85:819-822.
12. Witzig TE, Borell TJ, Herath JF, Tefferi A, Li C-Y, Jenkins RB (1994): Detection of trisomy 12 by FISH in untreated B-chronic lymphocytic leukemia: Correlation with stage and CD20 anti- gen expression intensity. Leuk Lymphoma 14:447-451.
13. Tabernero MD, San Miguel IF, Garcia JL, Garcia-Isidoro M, Wiegant J, Ciudad J, Gonzalez M, Rios A, Raap A, Orfao A (1995): Clinical, Biological, and immunophenotypical char- acteristics of B-cell chronic lymphocytic leukemia with tri- somy 12 by fluorescence in situ hybridization. Cytometry 22:217-222.
14. Oscier DG, Fitchett M, Herbert T, Lambert R (1991): Karyo- typic evolution in B-cell chronic lymphocytic leukemia. Genes Chromosom Cancer 3:16-20.
15. Cuneo A, Wlodarska I, Sayed Aly M, Piva N, Carli MG, Fagioli F, Tallarico A, Pazzi I, Ferrari L, Cassiman JJ, Van den Berghe H, Castoldi GL (1992): Non-radioactive in situ hybridization for the detection and monitoring of trisomy 12 in B-cell chronic lymphocytic leukemia, Br J Haematol 81:192-196.
16. Peterson L, Blackstadt M, Kay NE (1993): Clonal evolution in chronic lymphocytic leukemia. In: Chronic Lymphocytic Leukemia: Scientific Advances and Clinical Developments. BD Cheson, ed. Marcel Dekker, Inc., New York, pp. 181-196.
17. Iuliusson G, Gahrton G (1993): Cytogenetics in CLL and related disorders. In: Chronic Lymphocytic Leukemia and Related Disorders, Bailli~re's Clinical Hematology, C Rozman, ed. WB Sannders, London, pp. 821-848.
18. Criel A, Wlodarska I, Meeus P, Stul M, Louwagie A, Van Hoof A, Hidajat M, Mecucci C, Van Den Berghe H (1994): Tri- somy 12 is uncommon in typical chronic lymphocytic leuke- mias. Br J Haematol 87:523-528.
19. Cuneo A, Bigoni R, Balboni M, Gretel Carli M, Piva N, Fagioli F, Latorraca A, Wlodarska I, Van den Berghe H, Castoldi GL (1994): Trisomy 12 in chronic lymphocytic leukemia: A cyto- genetic and interphase cytogenetic study. Lank Lymphoma 15:167-172.
20. Brynes RK, McCarty A, Sun NCJ, Koo ChH (1995): Trisomy 12 in Richter's transformation of chronic lymphocytic leuke- mia. Am J Clin Pathol 104:199-203.
21. Nowell PC (1986): Mechanisms of tumor progression. Cancer Res 46:2203-2207.
22. Knauf WU, Knuutila S, Zeigmeister B, Thiel E (1995): Tri- somy 12 in B-cell chronic lymphocytic leukemia: Correlation with advanced disease, atypical morphology, high levels of sCD25, and with refractoriness to treatment. Leuk Lym- phoma 19:289-294.
23. Matutes E, Oscier DG, Garcia-Marco J, Ellis J, Copplestone A, Gillingham R, Hamblin T, Lens D, Swansbury GJ, Catovsky D (1996): Trisomy 12 defines a group of CLL with atypical mor- phology, correlation between cytogenetic, clinical and labo- ratory features in 544 patients. Br J Haematol 92:382-388.
24. Bennett JM, Catovsky D, Daniel MT, Flandrin G, Galton DAG, Gralnick HR, Sultan C (1989): Proposals for the classification of chronic (mature) B and T lymphoid leukemias. J Clin Pathol 42:567-584.
25. Garcia-Marco JA, Price C, Ellis J, Morey M, Matutes E, Lens D, Colman S, Catovsky D (1996): Correlation of trisomy 12 with proliferating cells by combined immunocytochemistry and fluorescence in sitn hybridization in chronic lympho- cytic leukemia. Leukemia 10:1705-1711.
26. Heim S, Mitelman F [1987): Cancer Cytogenetics. Alan R Liss Inc., New York.
27. Mitelman F (1981): Tumor etiology and chromosome pattern: Evidence from human and experimental neoplasms. In: Genes, Chromosomes and Neoplasia. Stubblefield E, ed. Raven Press, New York, pp. 335-350.
28. Crossen PE, Horn HL (1987): Origin of trisomy 12 in B-cell chronic lymphocytic leukemia. Cancer Genet Cytogenet 28:185-186.
29. Mecuci C, Delannoy A, Van den Berghe H (1988): The origin of trisomy 12 in chronic lymphocytic leukemia. Cancer Genet Cytogenet 36:203-204.
30. Schichman SA, Canaani E, Croce C (1995): Self-fusion of the ALL1 gene: A new genetic mechanism for acute leukemia. JAMA 273:571-576.
31. Knuutila S, Elonen E, Teerenhovi L, Rosi L, Leskinen R, Bloomfield C, De la Chapelle A (1986): Trisomy 12 in B cells of patients with B-cell chronic lymphocytic leukemia. N Engl J Med 314:865-869.
32. Risin S, Hopwood VL, Pathak S (1992): Trisomy 12 in Epstein-Barr virus-transformed lymphoblastoid cell lines of normal individuals and patients with nonhaematological malignancies. Cancer Genet Cytogenet 60:164-167.
33. Garcia-Marco J, Price C, Ellis J, Morilla R, Lens D, Matutes E, Oscier D, Catovsky D (1994): "In vitro" proliferation in chronic lymphocytic leukemia. An interphase cytogenetic analysis of clonal growth after mitogen stimulation. Blood 84 (Suppl) 1:452a.
34. Garcia-Marco J, Matutes E, Morilla R, Ellis ], Oscier D, Fantes J, Catovsky D, Price CM (1994): Trisomy 12 in B-cell chronic lymphocytic leukemia: Assessment of lineage restriction by simultaneous analysis of immunophenotype and genotype in interphase cells by fluorescence in situ hybridization. Br J Haematol 87:44-50.
35. Pittman S, Catovsky D (1984): Prognostic significance of chromosome abnormalities in chronic lymphocytic leuke- mia. Br J Haematol 58:649-660.
36. Fitchett M, Griffiths MJ, Oscier DG, Johnson S, Seabright M

58 J.A. Garcia-Marco et al.
(1987): Chromosome abnormalities involving band 13q14 in hematologic malignancies. Cancer Genet Cytogenet 24:143-150.
37. Ross FM, Stockdill G (1987): Clonal chromosome abnormali- ties in chromic lymphocytic leukemia patients revealed by TPA stimulation of whole blood cultures. Cancer Genet Cyto- genet 25:109-121.
38. Bird ML, Ueshima Y, Rowley JD, Haren JM, Vardiman JW (1989): Chromosome abnormalities in B cell chronic lympho- cytic leukemia and their clinical correlations. Leukemia 3:182-191.
39. Liu Y, Grand6r D, S6derhfill S, Juliusson G, Gahrton G, Ein- horn S (1992): Retinoblastoma gene deletions in B-cell chronic lymphocytic leukemia. Genes Chromosom Cancer 4:250-256.
40. Peterson LC, Lindquist LL, Church S, Kay NE, (1992): Fre- quent clonal abnormalities of chromosome band 13q14 in B-cell chronic lymphocytic leukemia: Multiple clones, subclones, and nonclonal alterations in 82 Midwestern patients. Genes Chromosom Cancer 4:273-280.
41. Brown AG, Ross FM, Dunne EM, Steel CM, Weir-Thompson EM (1993): Evidence for a new tumour suppressor locus (DBM) in human B-cell neoplasia telomeric to the retinoblas- toma gene. Nature Genet 3:67-72.
42. Stilgenbauer S, D6hner H, Bulgay-M6rschel M, Weitz S, Bentz M, Lichter P (1993): High frequency of monoallelic ret- inoblastoma gene deletion in B-cell chronic lymphoid leuke- mia shown by interphase cytogenetics. Blood 81:2118-2124.
43. Chapman RM, Corcoran MM, Gardiner A, Hawthorn LA, Cowell JK, Oscier DG (1994): Frequent homozygous deletions of the D13S25 locus in chromosome region 13q14 defines the location of a gene critical in leukaemogenesis in chronic B-cell lymphocytic leukemia. Oncogene 9:1289-1293.
44. Arif M, Tanaka K, Asou H, Ohno R, Kamada N (1995): Inde- pendent clones of trisomy 12 and retinoblastoma gene dele- tion in Japanese B cell chronic lymphocytic leukemia, detected by fluorescence in situ hybridization. Leukemia 9:1822-1827.
45. Fegan C, Robinson H, Thompson P, Whittaker JA, White D (1995): Karyotypic evolution in CLL: identification of a new sub-group of patients with deletions of 11q and advanced or progressive disease. Leukemia 9:2003-2008.
46. Mould S, Gardiner A, Corcoran M, Oscier DG (1996): Tri- somy 12 and structural abnormalities of 13q14 occurring in the same clone in chronic lymphocytic leukemia. Br I Hae- matol 92:389-392.
47. Finn WG, Thangavelu M, Yelavarthi KK, Goolsby ChL, Tall- man MS, Traynor A, Peterson LA (1996): Karyotype correlates with peripheral blood morphology and immunophenotype in chronic lymphocytic leukemia. Am J Clin Pathol 105:458--467.
48. Dryja TP, Rapaport J, Joyce JM, Petersen RA [1986): Molecular detection of deletions involving band q14 of chromosome 13 in retinoblastomas. Proc Natl Acad Sci U S A 83:7391-7394.
49. Mitchell DC, Cowell J (1989): Predisposition to retinoblas- toma due to a translocation within the 4.7R locus. Oncogene 4:253-257.
50. Liu Y, Szekely L, Grand6r D, S6derhgll S, Juliusson G, Gahr- ton G, Linder S, Einhorn S (1993): Chronic lymphocytic leu- kemia cells with allelic deletions at 13q14 commonly have
one intact RB1 gene: Evidence for a role of an adjacent locus. Proc Natl Acad Sci U S A 90:8697-8701.
51. Stilgenbauer S, Leupolt E, Ohl S, Weitz G, Schr6der M, Fis- cher K, Bentz M, Lichter P, D6hner H (1995): Heterogeneity of deletions involving RB-1 and the D13S25 locus in B-cell chronic lymphocytic leukemia revealed by fluorescence in situ hybridization. Cancer Res 55:3475-3477.
52. D6hner H, Pilz T, Fischer K, Cabot G, Diem D, Fink T, Stil- genbauer S, Bentz M, Lichter P (1994): Molecular cytogenetic analysis of RB-1 deletions in chronic B-cell leukemias. Leuk Lymphoma 16:97-103.
53. Kay NE, Suen R, Ranheim E, Peterson AL (1993): Confirma- tion of Rb gene defects in B-CLL clones and evidence for variable predominance of the Rb defective cells within the CLL clone. Br J Haematol 84:257-264.
54. Devilder MC, Francois S, Bosic C, Moreau A, Mellerin MP, Le Paslier D, Baraille R, Moisan JP (1995): Deletion cartogra- phy around the D13S25 locus in B cell chronic lymphocytic leukemia and accurate mapping of the involved tumor sup- pressor gene. Cancer Res 55:1355-1357.
55. Liu Y, Hermanson M, Grand~r D, Merup M, Wu X, Heyman M, Rasool O, Juliusson G, Gahrton G, Detlofsson R, Niki- forova N, Buys C, S6derh~ll S, Yankovsky N, Zabarovsky E, Einhorn S (1995): 13q Deletions in Lymphoid malignancies. B]ood 86:1911-1915.
56. Jabbar SAB, Ganeshaguru K, Wickremasinghe RG, Hoffbrand AV, Foroni L (1995): Deletion of chromosome 13 (bandq14) but not trisomy 12 is a clonal event in B-chronic lymphocytic leukemia (CLL). Br J Haematol 90:476-478.
57. Oscier DG (1994): Cytogenetic and molecular abnormalities in chronic lymphocytic leukemia. Blood Rev 8:88-97.
58. Hernandez JM, Mecucci C, Criel A, Means P, Michaux L, Van Hoof A, Verhoef G, Louwagie A, Scheiff J-M, Michaux J-L, Boogaerts M, Van den Berghe H (1995); Cytogenetic analysis of B cell chronic lymphoid leukemias classified according to morphologic and immunophenotypic (FAB) criteria. Leuke- mia 9:2140-2146.
59. Wooster R, Neuhausen S L, Mangion J, Quirk Y, Ford D, Col- lins N, Nguyen K, Seal Sh, Tran T, Averill D, Fieds P, Mar- shall G, Narod S, Lenoir G.M, Lynch H, Feunteun J, Devilee P, Cornelisse CJ, Menko FH, Daly PA, Ormiston W, McManus R, Pye C, Lewis CM, Cannon-Albright LA, Peto J, Ponder BAJ, Skolnick MH, Easton DF, Goldgar DE, Stratton MR (1994): Localization of a breast cancer susceptibility gene, BRCA2, to chromosome 13q12-13. Science 265:2088-2090.
60. Wooster R, Bignell G, Lancaster J, Swift S, Seal SH, Mangion J, Collins N, Gragory S, Gumbs C, Micklem G, Barfoot R, Hamoudi R, Patel S, Rice C, Biggs P, Hashim Y, Smith A, Con~ nor F, Arason A, Gudmundsson J, Ficenec D, Kelsell D, Ford D, Tonin P, Bishop DT, Spurr NK, Ponder BAJ, Eeles R, Peto J, Devilee P, Cornelisse C, Lynch H, Narod S, Lenoir G, Egilsson V, Barkadottir RB, Easton DF, Bentley DR, Futreal PA, Ash- worth A, Stratton MR (1995): Identification of the breast can- cer susceptibility gene BRCA2. Nature 378:789-792.
61. Garcia-Marco JA, Caldas C, Price CM, Wiedemann LM, Ash- worth A, Catovsky D (1996): Frequent somatic deletion of the 13q12.3 locus encompassing BRCA2 in chronic lymphocytic leukemia. Blood 88:1568-1575.