Embed Size (px)
Citation preview

ww.sciencedirect.com
j o u rn a l o f s u r g i c a l r e s e a r c h x x x ( 2 0 1 2 ) 1e9
Available online at w
journal homepage: www.JournalofSurgicalResearch.com
Association for Academic Surgery
Inhibition of experimental abdominal aortic aneurysm in a ratmodel by way of tanshinone IIA
Tao Shang, MD,a,1 Zhao Liu, MD, PhD,a,1 Min Zhou, MD, PhD,a
Christopher K. Zarins, MD,b Chengpei Xu, MD, PhD,b and Chang-jian Liu, MDa,*aDepartment of Vascular Surgery, Nanjing Drum Tower Hospital, Affiliated Hospital of Nanjing University Medical School, Nanjing, ChinabDivision of Vascular Surgery, Stanford University, Stanford, CA, USA
a r t i c l e i n f o
Article history:
Received 16 January 2012
Received in revised form
26 April 2012
Accepted 27 April 2012
Available online xxx
Keywords:
Abdominal aortic aneurysm
Tanshinone IIA
Metalloproteinase
Inflammation
Vascular smooth muscle cell
Inducible nitric oxide synthase
* Corresponding author. Department of VasMedical School, No. 321 Zhongshan Road, N
E-mail address: [email protected] (C.1 These authors equally contributed to thi
0022-4804/$ e see front matter ª 2012 Elsevdoi:10.1016/j.jss.2012.04.068
a b s t r a c t
Background: The purpose of the present study was to investigate whether tanshinone
IIA (Tan IIA), one of the major lipophilic components of Salvia miltiorrhiza Bunge, could
inhibit the development of elastase-induced experimental abdominal aortic aneurysms
(AAAs).
Methods: Male Sprague-Dawley rats (n ¼ 12/group) were randomly distributed into three
groups: Tan IIA, control, and sham. The rats from the Tan IIA and control groups under-
went intra-aortic elastase perfusion to induce AAAs, and the rats in the sham group were
perfused with saline. Only the Tan IIA group received Tan IIA (2 mg/rat/d). The maximum
luminal diameter of the abdominal aorta was measured before and 5, 12, 18, and 24 d after
perfusion. The systolic blood pressure was measured twice using the tail cuff technique
before administration and death. Aortic tissue samples were harvested at 24 d and
evaluated using reverse transcriptase-polymerase chain reaction, Western blot, immuno-
histochemistry, and Miller’s elastin-Van Gieson staining.
Results: The rats in the control group had significantly increased aortic sizes compared with
the sham group after 24 days (P < 0.05), and the Tan IIA group had a significant reduction in
aortic size (Tan IIA versus control, P < 0.05) without affecting blood pressure (P > 0.05). The
overexpression of matrix metalloproteinase-2, metalloproteinase-9, monocyte chemo-
tactic protein-1, and inducible nitric oxide synthase and the depletion of elastic fibers and
vascular smooth muscle cells induced by elastase perfusion were significantly decreased
by Tan IIA treatment (P < 0.05).
Conclusions: Tan IIA inhibited the development of elastase-induced experimental AAAs by
suppressing proteolysis, inflammation, and oxidative stress and preserving vascular
smooth muscle cells. It could be a new pharmacologic therapy for AAAs.
ª 2012 Elsevier Inc. All rights reserved.
cular Surgery, Nanjing Drum Tower Hospital, Affiliated Hospital of Nanjing Universityanjing 210008, China. Tel.: þ86 025 8330 4616 11801; fax: þ86 025 8330 8417.-j. Liu).s study.ier Inc. All rights reserved.

j o u r n a l o f s u r g i c a l r e s e a r c h x x x ( 2 0 1 2 ) 1e92
1. Introduction containing 81.6 U of elastase was performed for 10 min using
Abdominal aortic aneurysm (AAA) can cause death from
aneurysm rupture, which has a mortality rate of up to 90% [1].
This disease has been 1 of the 15 leading causes of death in the
United States [2]. Two randomized controlled trials have
shown that early surgery offered no survival advantage for
small AAAs (diameter less than 5.5 cm) in patients diagnosed
with ultrasound surveillance [3,4]. Therefore, surgical repair is
the only recommended treatment for patients with large
AAAs (diameter 5.5 cm or larger). However, from aneurysm
screening programs performed in the United States about
10 years ago, an estimate of 360,000 reported aneurysms were
small AAAs (greater than 90%) [5e7]. Those patients could not
receive effective therapy, except for “watchful waiting” in
clinics. Also, they were more likely to acquire psychological
problems and a reduced quality of life [8]. For the slow
development of small AAAs, it would take a long period before
they reach the threshold size for surgical intervention [9].
Thus, an effective pharmacologic therapy might be a good
option to limit or stop the growth of small AAAs.
Tanshinone IIA (Tan IIA) is one of the major lipophilic
components extracted from the root of Salvia miltiorrhiza
Bunge, which has been used to treat various types of ischemic
disease as a traditional Chinese medicine for many years [10].
Recent studies have shown that Tan IIA might have the
potential to suppress AAA formation and affect AAA devel-
opment [10e15]. Therefore, we studied the effect of Tan IIA on
experimental AAAs and investigated the possible molecular
mechanisms.
2. Methods
2.1. Experimental groups and AAA model
All experiments in our study were conducted in compliance
with the Guide for the Care and Use of Laboratory Animals
approved by the Ethical Committee of Researches of Nanjing
University and were approved by the Committee on the Use
and Care of Animals of Medical School of Nanjing University.
Six-week-old male rats (SLAC Laboratory Animal, Shanghai,
China) weighing 180 to 220 g were used in our study. All rats
(n ¼ 12/group) were randomly distributed into three groups:
Tan IIA, control, and sham. The rats were housed with a 12-h
light/dark cycle and standard diet and water. They were
anesthetized and underwent laparotomy mainly according to
previous methods, with some modifications [16,17]. In brief,
the abdominal aorta was isolated from the level of the left
renal vein to the bifurcation. A tiny incision was made on the
aorta bifurcation. Next, a polyethylene-10 tube (Smiths
Medical International, Ashford, UK) was introduced through
the incision into the abdominal aorta. The aorta was clamped
above the tip level of the polyethylene tube and ligated with
a 4-0 silk suture (Ethicon, Johnson & Johnson, Somerville, NJ)
near the aortic bifurcation, followed by perfusion with
27.2 U/mL type I porcine pancreatic elastase (Sigma-Aldrich,
St. Louis, MO) in the Tan IIA and control groups and isotonic
saline in the sham group. Aortic perfusion with 10 mL saline
a microinfusion pump at 2 atm. After perfusion, the clamp
and ligatures were removed, and the polyethylene tube was
withdrawn. The incision was sutured with an 8-0 poly-
propylene suture (Prolene, Ethicon, Johnson & Johnson).
Aortic segments were harvested for additional study 24 d after
perfusion. The systolic blood pressure of the rats was
measured using the tail cuff technique before administration
and death.
2.2. Drug administration
From the findings of previous reports and experiments, 2 mg
Tan IIA with 0.4 mL saline (Shanghai No.1 Biochemical &
Pharmaceutical, Shanghai, China) was given to each rat daily
by intraperitoneal injection in the Tan IIA group (n ¼ 12) [20].
Tan IIA injection was started 1 d before elastase perfusion
until the rats were killed. The same volume of saline was
given to the sham (n ¼ 12) and control (n ¼ 12) groups for the
same period by intraperitoneal injection.
2.3. Measurement of aortic size using ultrasonography
An ultrasound system (SonoSite, Bothell, WA) containing
a linear transducer (25 MHz) was used to demonstrate the
dilation of the rat aortas. We measured the maximum inner
luminal diameter of aortas. The aortic size was measured
before laparotomy and 5, 12, 18, and 24 d after. Two experi-
enced operators who were unaware of the protocols
performed the quantitative analysis of the ultrasound data.
2.4. Histologic studies
All rats were killed 24 d after the operation. The excised aorta
was fixed in 10% neutral-buffered formalin and processed
using routine paraffin embedding. Aortic tissue cross-sections
(5 mm) were stained with hematoxylin and eosin and Miller’s
elastin-Van Gieson according to standard procedures. The
percentage of the surface area occupied by the elastin-Van
Giesonestained elastic fibers was quantified using the
morphometry system MacScope, version 2.2 (Mitani, Tokyo,
Japan).
2.5. Immunohistochemical staining
Mouse monoclonal antibodies for matrix metalloproteinase
(MMP)-9 (ab58803, Abcam, Cambridge, MA) and MMP-2
(ab3158, Abcam) were used to analyze the local expression of
matrix-degrading proteinases in the aorta walls 24 d after
operation. Also, mouse monoclonal antibody for a-smooth
muscle cell (SMC) actin (ab7817, Abcam) was used to analyze
the depletion of medial SMCs. Immunohistochemical staining
was performed using an immunoperoxidase avidin-biotin
complex system. After blocking the activity of endogenous
peroxidase, the sections were incubated in the primary anti-
bodies (1:100) overnight at 4�C. Next, according to the manu-
facturer’s specifications (Vectastain Elite ABC Kit, Vector
Laboratories, Burlingame, CA), we incubated the sectionswith
biotinylated anti-mouse IgG antibody for 30 min (Vector

Table 1 e Effect of tanshinone IIA (2 mg/rat/d) on systolicblood pressure.
Group (n) Beforeadministration
(mm Hg)
24 d Afterperfusion(mm Hg)
P value
Sham (12) 113 � 3 112 � 3 0.304
Control (12) 113 � 4 115 � 3 0.287
Tan IIA (12) 112 � 4 113 � 4 0.226
P value 0.201 0.275
Tan IIA ¼ tanshinone IIA.
j o u rn a l o f s u r g i c a l r e s e a r c h x x x ( 2 0 1 2 ) 1e9 3
Laboratories) and then with avidin-biotinylated horseradish
peroxidase complex in phosphate-buffered saline for
10 min. Immune complexes were visualized using 0.05%
3,3’-diaminobenzidine (Vector Laboratories). The slides were
counterstained using hematoxylin.
2.6. Quantitative real-time reverse transcriptase-polymerase chain reaction
The expression of MMP-2 and MMP-9 mRNA in the aortic
tissues was determined using quantitative real-time reverse
transcriptase-polymerase chain reaction (PCR). TotalmRNAwas
extracted from the aorta using TRIzol reagent (Invitrogen, Carls-
bad, CA), and cDNA produced by reverse transcription using
oligo-(dT) primer 5.0 and Moloney Murine Leukemia Virus
Reverse Transcriptase (Fermentas, Vilnius, Lithuania). The PCRs
were performed in quadruplicate with SYBR Green PCR Core
Reagents (Toyobo, Osaka, Japan), and the fluorescence signals
were analyzedusing theDA7600 SequenceDetectionSystem (Da
An Gene, Guangzhou, China). The results for each sample were
normalized to the concentration of b-actin mRNA. PCR amplifi-
cation was performed under the following conditions: denatur-
ation for 15 s at 95�C, annealing for 30 s at 60�C, and extension for
30sat72�C.PrimersofMMP-2,MMP-9,andb-actinweredesigned
according to published rat sequences, and the amplified frag-
ments were 135 bp, 228 bp, and 136 bp, respectively. The primer
sequences were as follows: MMP-2 primer (GenBank reference
no. NM031054, 135 bp), sense primer: 50-CTGATAACCTGGATGCAGTCGT-30; antisense primer: 50-CCAGCCAGTCCGATTTGA-30;MMP-9 primer (GenBank reference no. NM031055, 228 bp); sense
primer: 50-TTCAAGGACGGTCGGTATT-30; antisense primer: 50-CTCTGAGCCTAGCCCCAACTTA-30; and b-actin primer (GenBank
referenceno.NM031144, 136bp), senseprimer: 50-GCAGAAGGAGATTACTGCCCT-30; antisense primer: 50-GCTGATCCACATCTGCTGGAA-30.
2.7. Western blot analysis
Aortic tissues were obtained and homogenized 24 d after the
operation, and the total proteins were extracted from the
frozen aorta tissue. The samples (70 mg) were electrophoresed
in sodium dodecyl sulfate-polyacrylamide gel at 80 V, trans-
ferred onto polyvinylidene difluoride membranes at 300 mA,
and incubated for 1 h in Tris-buffered saline, 5% nonfat milk,
and 0.2% Tween-20 at room temperature. The membranes
were then incubated for 24 h at 4�C with different antibodies,
including mouse monoclonal antibodies for MMP-9 (1:2000
dilution; Abcam), MMP-2 (1:2000 dilution; Abcam), and
inducible nitric oxide synthase (iNOS) (1:4000 dilution;
Abcam), rabbit polyclonal antibody for monocyte chemo-
attractant protein-1 (MCP-1) (1:5000 dilution; Abcam).They
were washed in Tris-buffered saline and 0.1% Tween-20 and
then incubated for 2 h at room temperature with sheep anti-
mouse IgG secondary antibody for MMP-2, MMP-9, and iNOS
(1:5000; Amersham Biosciences, Uppsala, Sweden), and goat
anti-rabbit IgG forMCP-1 (1:5000; AmershamBiosciences). The
membranes were visualized using an ECL plus chemilumi-
nescent kit (Amersham Biosciences), according to the manu-
facturer’s instructions, and exposed to X-ray film (Kodak,
Rochester, NY). b-Actin levels were used to standardize
protein loading. To quantify and compare the levels of
proteins, the density of each band was measured using
densitometry (Shimazu, Kyoto, Japan).
2.8. Statistical analysis
All values are presented as the mean � standard error of the
mean. Statistical analysis was performed using SPSS, version
16.0 (SPSS, Chicago, IL). Multiple comparisons were analyzed
using one-way analysis of variance followed by Bonferroni’s
post hoc test. The threshold for significance was P < 0.05.
3. Results
3.1. Blood pressure
At 24 d after intra-aortic perfusion, the systolic blood pressure
in all three groups was not significantly different (Table 1).
This result indicated that Tan IIA (2 mg/rat/d) did not affect
the blood pressure (P > 0.05).
3.2. Development of elastase-induced AAAs
AAA was induced by elastase perfusion. Next, the aortic size
was measured using ultrasonography before perfusion and 5,
12, 18, and 24 d afterward (Fig. 1A1, A2). Compared with the
sham group, we found that the aortic size of the control group
was significantly increased (P < 0.05; Fig. 1A3), which sug-
gested that the AAA model was successfully established.
However, the aortic size of the Tan II A group was obviously
smaller than that of the control group (P < 0.05; Fig. 1A3).
Therefore, Tan IIA inhibited the progression of the AAAs.
3.3. Histologic changes in aortic walls
The aortas of the rats were harvested after 24 d. On hema-
toxylin and eosin staining, it was found that the control group
had a thinner aortic wall and more mural thrombus than
other groups (Fig. 2A1, A2). Elastin-Van Gieson staining
showed observable degeneration of elastic lamellae in the
control group (control versus sham, P < 0.05; Fig. 2B2). In
contrast, no remarkable degeneration of the medial elastic
lamellae was seen in the Tan IIA or sham groups (Tan IIA
versus control, P < 0.05; Fig. 2B2).
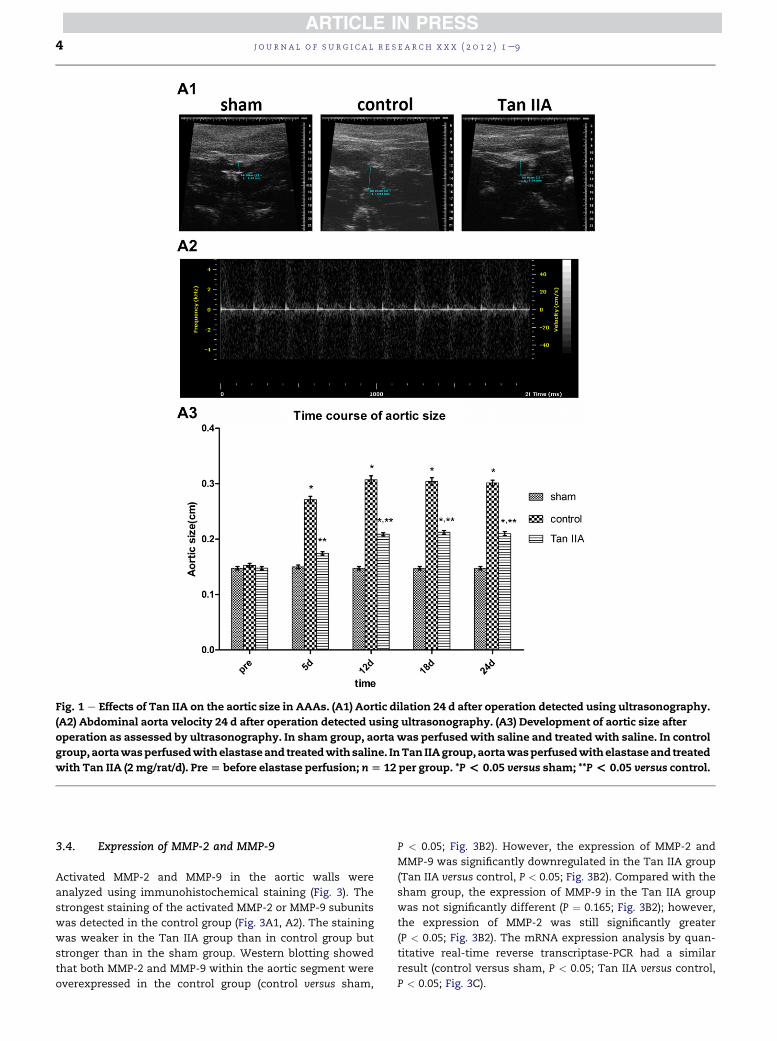
Fig. 1 e Effects of Tan IIA on the aortic size in AAAs. (A1) Aortic dilation 24 d after operation detected using ultrasonography.
(A2) Abdominal aorta velocity 24 d after operation detected using ultrasonography. (A3) Development of aortic size after
operation as assessed by ultrasonography. In sham group, aorta was perfusedwith saline and treatedwith saline. In control
group,aortawasperfusedwithelastaseand treatedwithsaline. InTan IIAgroup,aortawasperfusedwithelastaseand treated
with Tan IIA (2 mg/rat/d). Pre[ before elastase perfusion; n[ 12 per group. *P< 0.05 versus sham; **P< 0.05 versus control.
j o u r n a l o f s u r g i c a l r e s e a r c h x x x ( 2 0 1 2 ) 1e94
3.4. Expression of MMP-2 and MMP-9
Activated MMP-2 and MMP-9 in the aortic walls were
analyzed using immunohistochemical staining (Fig. 3). The
strongest staining of the activated MMP-2 or MMP-9 subunits
was detected in the control group (Fig. 3A1, A2). The staining
was weaker in the Tan IIA group than in control group but
stronger than in the sham group. Western blotting showed
that both MMP-2 and MMP-9 within the aortic segment were
overexpressed in the control group (control versus sham,
P < 0.05; Fig. 3B2). However, the expression of MMP-2 and
MMP-9 was significantly downregulated in the Tan IIA group
(Tan IIA versus control, P < 0.05; Fig. 3B2). Compared with the
sham group, the expression of MMP-9 in the Tan IIA group
was not significantly different (P ¼ 0.165; Fig. 3B2); however,
the expression of MMP-2 was still significantly greater
(P < 0.05; Fig. 3B2). The mRNA expression analysis by quan-
titative real-time reverse transcriptase-PCR had a similar
result (control versus sham, P < 0.05; Tan IIA versus control,
P < 0.05; Fig. 3C).
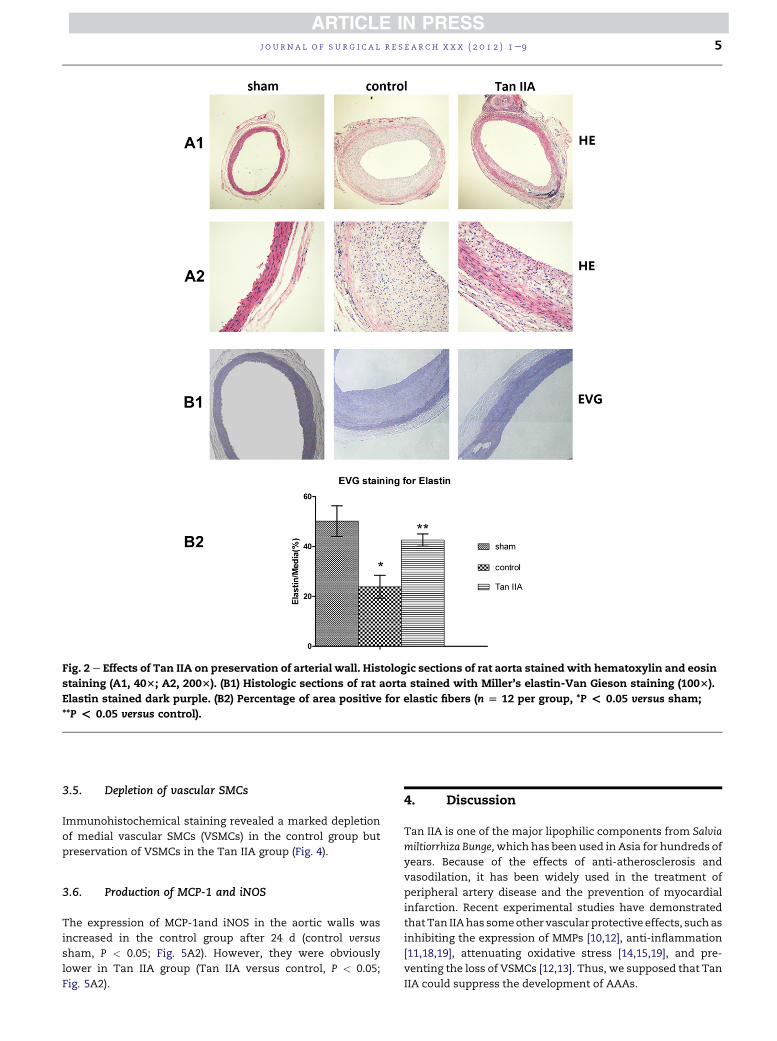
Fig. 2 e Effects of Tan IIA on preservation of arterial wall. Histologic sections of rat aorta stained with hematoxylin and eosin
staining (A1, 403; A2, 2003). (B1) Histologic sections of rat aorta stained with Miller’s elastin-Van Gieson staining (1003).
Elastin stained dark purple. (B2) Percentage of area positive for elastic fibers (n [ 12 per group, *P < 0.05 versus sham;
**P < 0.05 versus control).
j o u rn a l o f s u r g i c a l r e s e a r c h x x x ( 2 0 1 2 ) 1e9 5
3.5. Depletion of vascular SMCs
Immunohistochemical staining revealed a marked depletion
of medial vascular SMCs (VSMCs) in the control group but
preservation of VSMCs in the Tan IIA group (Fig. 4).
3.6. Production of MCP-1 and iNOS
The expression of MCP-1and iNOS in the aortic walls was
increased in the control group after 24 d (control versus
sham, P < 0.05; Fig. 5A2). However, they were obviously
lower in Tan IIA group (Tan IIA versus control, P < 0.05;
Fig. 5A2).
4. Discussion
Tan IIA is one of the major lipophilic components from Salvia
miltiorrhiza Bunge,which has been used in Asia for hundreds of
years. Because of the effects of anti-atherosclerosis and
vasodilation, it has been widely used in the treatment of
peripheral artery disease and the prevention of myocardial
infarction. Recent experimental studies have demonstrated
that Tan IIAhas someother vascular protective effects, suchas
inhibiting the expression of MMPs [10,12], anti-inflammation
[11,18,19], attenuating oxidative stress [14,15,19], and pre-
venting the loss of VSMCs [12,13]. Thus, we supposed that Tan
IIA could suppress the development of AAAs.
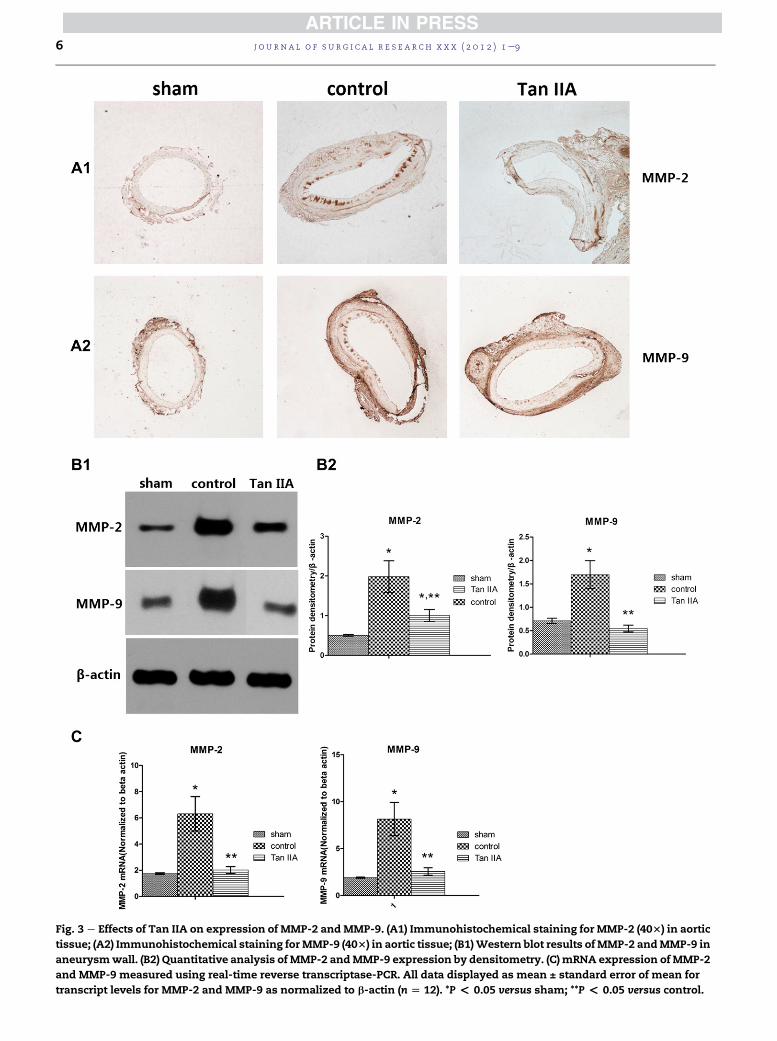
Fig. 3 e Effects of Tan IIA on expression of MMP-2 andMMP-9. (A1) Immunohistochemical staining for MMP-2 (403) in aortic
tissue; (A2) Immunohistochemical staining forMMP-9 (403) in aortic tissue; (B1)Western blot results ofMMP-2 andMMP-9 in
aneurysmwall. (B2) Quantitative analysis of MMP-2 andMMP-9 expression by densitometry. (C) mRNA expression of MMP-2
and MMP-9 measured using real-time reverse transcriptase-PCR. All data displayed as mean ± standard error of mean for
transcript levels for MMP-2 and MMP-9 as normalized to b-actin (n [ 12). *P < 0.05 versus sham; **P < 0.05 versus control.
j o u r n a l o f s u r g i c a l r e s e a r c h x x x ( 2 0 1 2 ) 1e96

Fig. 4 e Effects of Tan IIA on the preservation of VSMCs. Immunohistochemical staining for a-SMC actin in aorta tissues
(4003).
j o u rn a l o f s u r g i c a l r e s e a r c h x x x ( 2 0 1 2 ) 1e9 7
The elastase-induced AAA in our experiments is a stan-
dard small animal aneurysm model. Compared with other
AAA-induced methods, it achieves pathologic characteristics
similar to those of human AAAs [20]. The animal surgery was
performed according to a previously published protocol, with
some modifications [21]. Previous studies have indicated that
AAA formation correlated with the pump pressure and elas-
tase concentration [21]. We increased the perfusion pressure
to 2 atm with an elastase concentration of 8.16 U/mL to
shorten the period of anesthesia and allow the operator to
suture the incision in the bifurcation of the abdominal aorta in
time. These modifications resulted in successful AAAmodels.
AAA is a complex multifactorial disease with an unknown
etiology. It occurs after the development of an inflammatory
response, destruction of elastin and collagen, the loss of
VSMCs, and other histologic changes [21e26]. Among these
characteristics, chronic inflammation is prominent and could
leadmacrophages and lymphocytes to infiltrate into the outer
aortic wall [27]. The recruitedmacrophages could secret MMPs
and pro-inflammatory cytokines, contributing to the devel-
opment of AAAs [28]. Increased expression of MMPs (espe-
cially MMP-2 and MMP-9) could promote elastin degradation,
which is considered to cause the direct enlargement of AAA
[24,29]. In addition, some of the pro-inflammatory cytokines
such asMCP-1 could induce themigration ofmonocytes to the
aneurysm walls [18]. These infiltrated monocytes would
further secret MMPs and pro-inflammatory cytokines and
Fig. 5 e (A1) Western blot results of MCP-1 and iNOS expression
by densitometry (n [ 12 per group, *P < 0.05 versus sham, **P
continue to deteriorate the pathologic process [19,30]. Our
results showed that MMP-2, MMP-9, and MCP-1 were highly
expressed in the aortic walls of AAA rats. In accordance with
previous studies of atherosclerotic and inflammatory animal
models, our studies showed that, along with inhibiting the
overexpression of MMP-2, MMP-9, and MCP-1, Tan IIA could
attenuate the inflammatory response and downregulate the
expression of MMP-2 and MMP-9 in an AAA model.
Apoptosis of VSMCs is another major pathologic feature of
AAA. It impairs the reparation for connective tissues and
reduces the capacity for anti-pressure. The loss of VSMCs is
generally caused by two mechanisms, one associated with
inflammation. Previous research has shown that Fas ligand
and inflammatory cytokines (e.g., interleukin-6) from infil-
trated inflammatory cells play a crucial role in the depletion of
medial VSMCs [21,31]. The other mechanism is related to the
activation of nuclear factor-kB (NF-kB) caused by oxidative
stress [32]. In our study, the loss of VSMCs in the control group
was observed by immunohistochemical staining. Some
investigators have reported that Tan IIAwas able to inhibit the
migration of human aortic SMCs and the proliferation of
VSMCs [12,13]. Our study has demonstrated that Tan IIA
treatment can preserve the medial level of VSMCs in the rat
aortic wall. Therefore, it might inhibit the development of
AAAs by preventing the loss of VSMCs.
Oxidative stress involved in inflammatory response is
associated with aortic aneurysm formation in human tissues
. (A2) Quantitative analysis of MCP-1 and iNOS expression
< 0.05 versus control).

j o u r n a l o f s u r g i c a l r e s e a r c h x x x ( 2 0 1 2 ) 1e98
and animal models [19,33]. Previous studies have suggested
thatnitric oxidesynthesized fromiNOScandamage thearterial
wall by producing reactive oxidative stress. Thus, the expres-
sion of iNOS could promote oxidative vascular injury in AAAs
[34e36]. According toour study, theelastase-inducedAAAshad
overexpressed iNOS, and the Tan IIA-treated rats did not share
this phenomenon. Thus, it can be inferred that Tan IIA might
inhibit the development of AAAs by downregulating the
expression of iNOS and attenuating oxidative stress.
5. Conclusions
Wefound that Tan IIA could significantly inhibit the expansion
of elastase-induced experimental AAAs. The possible mecha-
nisms include downregulating the expression ofMMP-2,MMP-
9, and MCP-1, preventing the loss of VSMCs, suppressing the
infiltration of monocytes, and decreasing oxidative stress by
inhibiting iNOS synthesis. Our findings have shown that it
could be a new potential drug for AAA therapy.
Acknowledgment
The authors thank Dr. Wei Wang and Zhu Cheng-yan for
technical assistance with the duplex ultrasound examination
of the rat aorta. This work was supported by grants from
Nature Science Foundation of Jiangsu Province, China (grant
BK2009035).
r e f e r e n c e s
[1] Pearce WH, Zarins CK, Bacharach JM. Writing Group forAtherosclerotic Peripheral Vascular Disease Symposium II:controversies in abdominal aortic aneurysm repair.Circulation 2008;118:2860.
[2] Hallett JW Jr. Management of abdominal aortic aneurysms.Mayo Clin Proc 2000;75:395.
[3] The UK Small Aneurysm Trial Participants. Mortality resultsfor randomized controlled trial of early elective surgery orultrasonographic surveillance for small abdominal aorticaneurysms. Lancet 1998;352:1649.
[4] Lederle FA, Wilson SE, Johnson GR. Immediate repaircompared with surveillance of small abdominal aorticaneurysms. N Engl J Med 2002;346:1437.
[5] Fleming C, Whitlock EP, Beil TL, et al. Screening forabdominal aortic aneurysm: a best-evidence systematicreview for the U.S. Preventive Services Task Force. AnnIntern Med 2005;142:203.
[6] Lederle FA, Johnson GR, Wilson SE. Prevalence andassociations of abdominal aortic aneurysm detected throughscreening. Aneurysm Detection and Management (ADAM)Veterans Affairs Cooperative Study Group. Ann Intern Med1997;126:441.
[7] Boll AP, Verbeek AL, van de Lisdonk EH, et al. Highprevalence of abdominal aortic aneurysm in a primary carescreening programme. Br J Surg 1998;85:1090.
[8] Lindeman JH, Abdul-Hussien H, van Bockel JH, et al. Clinicaltrial of doxycycline for matrix metalloproteinase-9 inhibitionin patients with an abdominal aneurysm: doxycycline
selectively depletes aortic wall neutrophils and cytotoxic Tcells. Circulation 2009;119:2209.
[9] Rentschler M, Baxter BT. Pharmacological approaches toprevent abdominal aortic aneurysm enlargement andrupture. Ann N Y Acad Sci 2006;1085:39.
[10] Zhi-yuan F, Rong L, Bing-xiang Y, et al. Tanshinone IIAinhibits atherosclerotic plaque formation by down-regulating MMP-2 and MMP-9 expression in rabbits feda high-fat diet. Life Sci 2007;81:1339.
[11] Zhi-yuan F, Rong L, Bing-xiang Y, et al. Tanshinone IIAdownregulates the CD40 expression and decreases MMP-2activity on atherosclerosis induced by high fatty diet inrabbit. J Ethnopharmacol 2008;115:217.
[12] Jin UH, Suh SJ, Chang HW, et al. Tanshinone IIA from Salviamiltiorrhiza bunge inhibits human aortic smooth muscle cellmigration and MMP-9 activity through Akt signalingpathway. J Cell Biochem 2008;104:15.
[13] Hong W, XiuMei G, BoLi Z. Tanshinone: an inhibitor ofproliferation of vascular smooth muscle cells.J Ethnopharmacol 2005;99:93.
[14] Tang F, Wu X, Wang T, et al. Tanshinone IIA attenuatesatherosclerotic calcification in rat model by inhibition ofoxidative stress. Vasc Pharmacol 2007;46:427.
[15] Huang KJ, Wang H, Xie WZ, et al. Investigation of the effect oftanshinone IIA on nitric oxide production in human vascularendothelial cells by fluorescence imaging. Spectrochim ActaA 2007;68:1180.
[16] Anidjar S, Salzmann JL, Gentric D, et al. Elastase-inducedexperimental aneurysms in rats. Circulation 1990;82:973.
[17] Petrinec D, Liao S, Holmes DR, et al. Doxycycline inhibition ofaneurysmal degeneration in an elastase-induced rat modelof abdominal aortic aneurysm: preservation of aortic elastinassociated with suppressed production of 92 kD gelatinase. JVasc Surg 1996;23:336.
[18] Ren ZH, Tong YH, Xu W, et al. Tanshinone IIA attenuatesinflammatory responses of rats with myocardial infarctionby reducing MCP-1 expression. Phytomedicine 2010;17:212.
[19] Jang SI, Jeong SI, Kim KJ, et al. Tanshinone IIA from Salviamiltiorrhiza inhibits inducible nitric oxide synthaseexpression and production of TNF-a, IL-1b and IL-6 inactivated RAW264.7 Cells. Planta Med 2003;69:1057.
[20] Tanaka A, Hasegawa T, Chen Z, et al. A novel rat model ofabdominal aortic aneurysm using a combination ofintraluminal elastase infusion and extraluminal calciumchloride exposure. J Vasc Surg 2009;50:1423.
[21] Thompson RW, Curci JA, Ennis TL, et al. Pathophysiology ofabdominal aortic aneurysms: insights from the elastase-induced model in mice with different genetic backgrounds.Ann NY Acad Sci 2006;1085:59.
[22] Steinmetz EF, Buckley C, Shames ML, et al. Treatment withsimvastatin suppresses the development of experimentalabdominal aortic aneurysms in normal andhypercholesterolemic mice. Ann Surg 2005;241:92.
[23] Shiraya S, Miyake T, Aoki M, et al. Inhibition of developmentof experimental aortic abdominal aneurysm in rat model byatorvastatin through inhibition of macrophage migration.Atherosclerosis 2009;202:34.
[24] Longo GM, Xiong W, Greiner TC, et al. Matrixmetalloproteinases 2 and 9 work in concert to produce aorticaneurysms. J Clin Invest 2002;110:625.
[25] Miller FJ Jr, Sharp WJ, Fang X, et al. Oxidative stress in humanabdominal aortic aneurysms: a potential mediator ofaneurysmal remodeling. Arterioscler Thromb Vasc Biol 2002;22:560.
[26] Teng X, Zhang H, Snead C, et al. Molecular mechanismsof iNOS induction by IL-1 beta and IFN-gamma in rataortic smooth muscle cells. Am J Physiol Cell Physiol 2002;282:144.

j o u rn a l o f s u r g i c a l r e s e a r c h x x x ( 2 0 1 2 ) 1e9 9
[27] Liapis CD, Paraskevas KI. The pivotal role of matrixmetalloproteinases in the development of human abdominalaortic aneurysms. Vasc Med 2003;8:267.
[28] Newman KM, Jean-Claude J, Hong L, et al. Cytokines thatactivate proteolysis are elevated in abdominal aorticaneurysm. Circulation 1994;90:224.
[29] Raffetto JD, Khalil RA. Matrix metalloproteinases and theirinhibitors in vascular remodeling and vascular disease.Biochem Pharmacol 2008;75:346.
[30] Chattopadhyay S, Myers RR, Janes J, et al. Cytokineregulation of MMP-9 in peripheral glia: implications forpathological processes and pain in injured nerve. BrainBehav Immun 2007;21:561.
[31] Watanabe N, Arase H, Kurasawa K, et al. Th1 and Th2subsets equally undergo Fas-dependent and -independentactivation-induced cell death. Eur J Immunol 1997;27:1858.
[32] Li P-F, Dietz R, von Harsdorf R. Reactive oxygen speciesinduce apoptosis of vascular smooth muscle cell. FEBS Lett1997;404:249.
[33] Xiong W, MacTaggart J, Knispel R, et al. Inhibition of reactiveoxygen species attenuates aneurysm formation in a murinemodel. Atherosclerosis 2009;202:128.
[34] Inhibition of inducible nitric oxide synthase limits nitricoxide production and experimental aneurysm expansion.J Vasc Surg 2001;33:579.
[35] Zhang J, Schmidt J, Ryschich E, et al. Inducible nitric oxidesynthase is present in human abdominal aortic aneurysmand promotes oxidative vascular injury. J Vasc Surg 2003;38:360.
[36] Liao M-F, Jing Z-P, Bao J-M, et al. Role of nitric oxide andinducible nitric oxide synthase in human abdominal aorticaneurysms: a preliminary study. Chin Med J 2006;119:312.