Embed Size (px)
Citation preview

REGULAR PAPER
Identification and characterization of a carboxysomal c-carbonicanhydrase from the cyanobacterium Nostoc sp. PCC 7120
Charlotte de Araujo • Dewan Arefeen • Yohannes Tadesse • Benedict M. Long •
G. Dean Price • Roger S. Rowlett • Matthew S. Kimber • George S. Espie
Received: 8 May 2014 / Accepted: 19 May 2014 / Published online: 8 June 2014
� Springer Science+Business Media Dordrecht 2014
Abstract Carboxysomes are proteinaceous microcom-
partments that encapsulate carbonic anhydrase (CA) and
ribulose 1,5-bisphosphate carboxylase/oxygenase (Rubi-
sco); carboxysomes, therefore, catalyze reversible HCO3-
dehydration and the subsequent fixation of CO2. The N-
and C-terminal domains of the b-carboxysome scaffold
protein CcmM participate in a network of protein–protein
interactions that are essential for carboxysome biogenesis,
organization, and function. The N-terminal domain of
CcmM in the thermophile Thermosynechococcus elongatus
BP-1 is also a catalytically active, redox regulated c-CA.
To experimentally determine if CcmM from a mesophilic
cyanobacterium is active, we cloned, expressed and puri-
fied recombinant, full-length CcmM from Nostoc sp. PCC
7120 as well as the N-terminal 209 amino acid c-CA-like
domain. Both recombinant proteins displayed ethoxyzola-
mide-sensitive CA activity in mass spectrometric assays, as
did the carboxysome-enriched TP fraction. NstCcmM209
was characterized as a moderately active and efficient c-
CA with a kcat of 2.0 9 104 s-1 and kcat/Km of
4.1 9 106 M-1 s-1 at 25 �C and pH 8, a pH optimum
between 8 and 9.5 and a temperature optimum spanning
25–35 �C. NstCcmM209 also catalyzed the hydrolysis of
the CO2 analog carbonyl sulfide. Circular dichroism and
intrinsic tryptophan fluorescence analysis demonstrated
that NstCcmM209 was progressively and irreversibly
denatured above 50 �C. NstCcmM209 activity was inhib-
ited by the reducing agent tris(hydroxymethyl)phosphine,
an effect that was fully reversed by a molar excess of
diamide, a thiol oxidizing agent, consistent with oxidative
activation being a universal regulatory mechanism of
CcmM orthologs. Immunogold electron microscopy and
Western blot analysis of TP pellets indicated that Rubisco
and CcmM co-localize and are concentrated in Nostoc sp.
PCC 7120 carboxysomes.
Keywords Carboxysome � CcmM � CO2 concentrating
mechanism � Carbonyl sulfide � Gamma-carbonic
anhydrase � Nostoc sp. PCC 7120
Introduction
Cyanobacteria possess a complex, biophysical CO2 con-
centrating mechanism (CCM) which is crucial for photo-
synthesis, growth, and survival. This adaptive mechanism
relies on light-energy-dependent membrane transport sys-
tems to acquire and concentrate HCO3- in the cytosol. The
Electronic supplementary material The online version of thisarticle (doi:10.1007/s11120-014-0018-4) contains supplementarymaterial, which is available to authorized users.
C. de Araujo � D. Arefeen � G. S. Espie
Department of Cell & Systems Biology, University of Toronto,
Mississauga, ON, Canada
Y. Tadesse � G. S. Espie (&)
Department of Biology, University of Toronto, 3359
Mississauga Road, Mississauga, ON L5L 1C6, Canada
e-mail: [email protected]
B. M. Long � G. D. Price
Division of Plant Science, Research School of Biology,
Australian National University, Canberra, ACT, Australia
R. S. Rowlett
Department of Chemistry, Colgate University, Hamilton, NY,
USA
M. S. Kimber
Department of Molecular and Cellular Biology, University of
Guelph, Guelph, ON, Canada
123
Photosynth Res (2014) 121:135–150
DOI 10.1007/s11120-014-0018-4

accumulated HCO3- subsequently enters the carboxysome
where it is dehydrated to CO2 by a specific carbonic
anhydrase (CA) and is then fixed by ribulose 1,5-bisphos-
phate carboxylase/oxygenase (Rubisco) to initiate the
photosynthetic carbon reduction cycle. By facilitating
HCO3-/CO2 equilibration solely within the confines of the
carboxysome, this CA effectively maintains a near-satu-
rating substrate concentration in close proximity to Rubi-
sco (Price et al. 2008). The importance of a carboxysomal
CA to the photoautotrophic lifestyle of cyanobacteria is
highlighted by the fact that the loss of this protein from the
carboxysome results in a conditional lethal phenotype that
can only be rescued by growth at supra-normal CO2
(Fukuzawa et al. 1992; So et al. 2002b; Yu et al. 1992). In
cyanobacteria with form 1A Rubisco (a-cyanobacteria), the
carboxysomal CA is CsoSCA (formerly CsoS3) a highly
divergent member of the b-class of CAs (Sawaya et al.
2006; So et al. 2004). In cyanobacteria with form 1B Ru-
bisco (b-cyanobacteria), like Synechocystis PCC 6803 and
Synechococcus PCC 7942, the carboxysomal CA has been
identified as a 30 kDa protein (CcaA) with closest
homology to the b-class of CAs that is widely distributed in
bacteria, eukaryotic algae, and plants (So et al. 2002b; Yu
et al. 1992; So and Espie 2005).
CcaA is recruited to the carboxysome as part of a larger
protein complex that also contains CcmM and CcmN (Cot
et al. 2008; Long et al. 2007). CcmM is the central com-
ponent of this complex, interacting with both CcaA and
CcmN directly through its N-terminal c-CA-like domain
(Cot et al. 2008; Long et al. 2007; Kinney et al. 2012). In
addition to the N-terminal domain, CcmM also possesses a
C-terminus comprised of a series of 3–5 RbcS-like domains
connected by flexible linkers. These domains interact with
RbcL and help recruit Rubisco to the carboxysome (Cot
et al. 2008; Long et al. 2007). CcmM is also expressed in
one or more ‘‘short forms’’ which arises from an internal
ribosome-binding site and contains only the RbcS-like
repeat domains (Cot et al. 2008; Long et al. 2007, 2010).
This variant interacts with RbcL to create an internal net-
work of protein interactions that organizes multiple Rubi-
sco molecules into an ordered array within the core of the
carboxysome. The CcmM/CcmN/CcaA complex also
appears capable of interacting with CcmK2 in the shell
through either the N-terminal domain of CcmM (Cot et al.
2008), or the extended C-terminus of CcmN (Kinney et al.
2012).The absence of carboxysomes and concomitant high
CO2 requiring phenotype of DccmM mutants attest to the
importance of this multi-domain protein to the structure
and function of b-carboxysomes (Berry et al. 2005;
Hackenberg et al. 2012; Ludwig et al. 2000; Marcus et al.
1992).
As mentioned, the N-terminal domain of CcmM has
detectable sequence homology to c-CAs, including 38 and
29 % identity, respectively, to the Methanosarcina ther-
mophila Cam (MtCam) and CamH enzymes (MtCamH)
(Alber et al. 1999; Alber and Ferry 1994, 1996; Zimmer-
man et al. 2010). However, in most instances, proteins with
similarity to Cam are determined to have no CA activity
(Pena et al. 2010; Zimmerman et al. 2010), and indeed,
previous work had shown that CcmMs from Synechocystis
PCC 6803 (SynCcmM) and Synechococcus PCC 7942
(SycCcmM) do not display CA activity under a variety of
assay conditions (Cot et al. 2008; So and Espie 2005).
However, the N-terminal 209 amino acids of CcmM of
Thermosynechococcus elongatus BP-1 (TeCcmM209) was
recently demonstrated to be catalytically active (Pena et al.
2010); this b-cyanobacterium lacks a ccaA homolog, and
CcmM, in this case, seems to take over the catalytic role of
the carboxysomal CA. The structure of TeCcmM209 is
characterized by the presence of a C-terminal a-helix
which is stabilized by a disulfide bond; this helix, and the
extended loop connecting the first pair of b-strands appears
to act as a redox switch, and was proposed to be the key
feature for predicting which CcmM’s retain CA activity
(Pena et al. 2010). While the presence of known functional
motifs in diverse CcmM homologs predicts that catalytic
activity is widespread (though not universal) among CcmM
homologs, no CA activity has been shown for a CcmM
from any cyanobacterium except T. elongatus. This is
problematic given that the only proven catalytically com-
petent cyanobacterial c-CA is from a thermophilic strain,
while CcmM homologs tested in two mesophilic cyano-
bacteria both proved to be catalytically incompetent. To
establish the enzymatic activity and role in the CCM for
CcmM in mesophilic cyanobacteria, we have examined the
sub-cellular localization, activity profile, and catalytic
properties of CcmM encoded by the diazotrophic cyano-
bacterium Nostoc sp. PCC 7120 (NstCcmM).
Materials and methods
Isolation of Nostoc sp. PCC 7120 carboxysomes
Nostoc sp. PCC 7120 (hereafter Nostoc PCC 7120) was
obtained from the Canadian Phycological Culture Collec-
tion, University of Waterloo, Waterloo, Canada as CPCC
387. The cells were maintained in unbuffered BG11
medium with constant shaking at 120 rpm. The cultures
were continuously illuminated using cool white and gro-
lux fluorescent lamps (25 lmol m-2 s-1, photosynthetic
photon flux density). For carboxysome isolation from 2 to
6 L of illuminated cells were grown to late log phase
(8 days) with 5 % v/v CO2 followed by 3 additional days
with air bubbling to fully induce the CCM. A carboxy-
some-enriched fraction (TP pellet) was obtained from
136 Photosynth Res (2014) 121:135–150
123

crude cell lysates using the Percoll-Mg2? technique as
described previously (Long et al. 2005, 2007).
Protein overexpression and polyclonal antibody
production
The ccmM gene (all0865) encoding a 555 amino acid
protein (NCBI Accession, NP_484908) was PCR amplified
from Nostoc PCC 7120 genomic DNA and cloned into
pET21b or pET28b (Novagen) using standard techniques.
The PCR primers employed for cloning and sequencing are
listed in Table S1. To confirm the presence of the gene of
interest, DNA sequencing was performed at The Centre for
Applied Genomics, The Hospital for Sick Children (Tor-
onto, Canada).
Recombinant E. coli BL21 DE3 cells or Rosetta Blue
DE3 cells were grown at 37 �C in shaker flasks, and
recombinant protein expression was induced with 1 mM
IPTG at 20 �C (16 h) or 37 �C (6 h). The cells were har-
vested by centrifugation, and the pellet was lysed on ice by
five 10 s bursts of ultrasound (50 % maximum setting,
Heat Systems Inc. sonicator W185) separated by 30 s
intervals on ice. Bacterial lysate was clarified by centrifu-
gation and passed through a 0.45 lm filter. The filtrate was
loaded onto an immobilized metal affinity column (IMAC,
Novagen). Weakly bound protein was removed by exten-
sive washing as prescribed in the manufacturer’s protocol.
His-tag-bound protein was eluted with 1 M imidazole,
0.5 M NaCl, 20 mM Tris–HCl, pH 7.9 and fractions con-
taining the purified proteins were pooled and subsequently
dialyzed against 500 mM NaCl, 1 mM EDTA, 20 mM
Tris–HCl, pH 8.0.
Protein inclusion bodies were solubilized in 100 mM
EPPS/NaOH, 20 mM MgSO4, 10 lM ZnSO4, 6.0 M urea,
pH 8.0. After the addition of solid urea, the slurry was
diluted to 5, 4, 3, or 2 M urea and stirred for 2 h at room
temperature. The slurries were centrifuged at 5,0009g for
15 min; the supernatant was collected and immediately
used for enzymatic analysis.
Protein purity was assessed by SDS-PAGE, and the identity
of the CcmM protein was verified by Western blot analysis.
Rabbit polyclonal antibodies were created in house against
two highly conserved regions of the SynCcmM protein. Syn-
thetic peptides corresponding to the N-terminal c-CA-like
domain (APGSSIRADEGTPFWIGGN) and the Rubisco
small subunit-like domain (HAGEYVRIIGIDRVGKRRVL)
were synthesized by Advanced SynTech Corporation (Tor-
onto, Canada) and used as the antigen. The IgG fraction from
immunized serum was separated using a Protein G affinity
column (Sigma). Reactivity of the antibodies (anti-CcmM NT
and anti-CcmM CT) was assessed using bona fide SynCcmM
and Synechocystis sp. PCC 6803 cell lysates. A third CcmM
antibody along with an anti CcmK antibody, raised against
SycCcmM and SycCcmK, respectively (Long et al. 2010), was
used in the analysis of carboxysome-enriched fractions. Anti
Rubisco antibody (anti-RbcL/S) was a gift from Dr. S. Whit-
ney, Australian National University. Protein concentrations
were determined by the Bradford method (Bio-Rad) using
bovine serum albumin as a standard.
Immunogold electron microscopy
Nostoc PCC 7120 cells (1 mL) were centrifuged at
10,000 rpm for 1 min. The pellet was suspended and fixed
with 1 % v/v glutaraldehyde (in 0.1 M Sorensen’s phos-
phate buffer) for 2 h at room temperature. The sample was
washed by centrifugation three times with Sorensen’s
phosphate buffer for 5 min. The cells were then dehydrated
using a graded ethanol series (50, 70, 95 % v/v) at room
temperature followed by 3 9 10 min washes with 100 %
ethanol. Samples were infiltrated with LR white resin in the
following ratios: 1:1 LR white (London Resin Co Ltd,
England) and 100 % ethanol for 30 min, 3:1 LR white
resin and 100 % ethanol for 1 h followed by 100 % LR
white for an hour at 4 �C. An additional overnight incu-
bation with 100 % LR white was performed, preceding a
final treatment for 4 h at 4 �C. Fractions of the pellet were
transferred to 00 gelatin capsules and covered with LR
white resin. Polymerization step was performed overnight
at 58 �C as prescribed previously (Lichtle et al. 1992).
Sections were cut with a Reichert Ultracut E microtome
and collected on 300-nm mesh nickel grids.
Post-embedding immunolabeling
Sample grids were incubated with blocking solution (100
lL of conditioned TBS supplemented with 1 mM CaCl2,
1 mM MgCl2, 0.1 % globulin free BSA, 0.4 % cold water
fish gelatin, pH 7.0) for 2 h. Grids were then transferred to
drops containing 100 lL of the primary antibody, either
anti-RbcL/S or anti-CcmM CT at a 1:100 (v/v) dilution and
incubated for 2 h. The grids were washed four times (2, 2,
4, and 4 min incubations) with TBS and then exposed to
100 lL of the secondary antibody at 1:100 (v/v) dilution
(Aurion Immuno Protein A gold, 15 nm) for an additional
2 h. Grids were re-washed three times for 3 min and
allowed to air dry. Sections were post-stained with satu-
rated uranyl acetate for 3 min and rinsed 10 times for
1 min with distilled water. The samples were viewed using
a Hitachi H7000 transmission electron microscope at an
accelerating voltage of 75 kV.
Carbonic anhydrase activity assays
CA activity was determined by three independent methods.
An aqueous inlet mass spectrometer (model MM 14-80 SC:
Photosynth Res (2014) 121:135–150 137
123

VG Gas Analysis, Middlewich, England) was used to
measure 18O exchange between 13C18O2 and H216O (Miller
et al. 1997; Silverman 1982), by following changes in13C16O16O (m/z = 45), 13C18O16O (m/z = 47), and13C18O18O (m/z = 49) over time in aqueous solution. The18O enrichment of the 13CO2 species (atom %) was cal-
culated as log18O atom % = log [([47] ? 2[49])/(2
([45] ? [47] ? [49]))) 9 100]. Standard assay conditions
employed 100 mM EPPS/NaOH buffer, pH 8, 20 mM
MgSO4, 10 lM ZnSO4, and 0.6 mM K213C18O3 as substrate
at 30 �C. The final protein concentration was 8 lg mL-1.
CA activity was also determined using the electrometric
assay (Wilbur and Anderson 1948). The time required for
the pH of a weakly buffered solution to change from 8.1 to
7.0 following the addition of CO2 was recorded in the
absence or presence of protein. CA activity per mg protein
was expressed as, Activity Units = (to - t)/t, where to and
t are the time required for the pH to change from 8.1 to 7.0
in absence and presence of the protein, respectively.
Steady-state kinetic measurements of CA activity
Initial rates of CO2 hydration were determined by stopped-
flow spectroscopy (Hi-Tech SF-61DX2 spectrophotometer)
at 25 �C using the changing pH-indicator method (Khalifah
1971; Rowlett et al. 1991). Saturated solutions of CO2 were
prepared by bubbling CO2 gas (100 % v/v) into distilled
water in a vessel maintained at 25 �C. Dilutions of the CO2
solution were prepared by mixing with degassed water to
yield the desired CO2 concentration. Mixing was achieved
by coupling two gastight syringes with a luer-lock con-
nector. CO2 concentrations were calculated on the basis of
a 33.8 mM saturated solution at 25 �C (Rowlett et al.
1991). The following buffer/indicator combinations and
absorbance wavelengths were used: at pH 5.5–7.0, MES
(pKa = 6.1)/chlorophenol red (574 nm); pH 7.0–8.1,
MOPS (pKa = 7.2)/p-nitrophenol (400 nm); pH 8.0–9.5
Bicine (pKa = 8.3)/m-creosol purple (578 nm). Buffer
concentration during measurements was 40 mM. The
steady-state kinetic constant kcat, Km, and kcat/Km were
determined by fitting the observed initial rates to the
Michaelis—Menten equation as described previously
(Rowlett et al. 2002).
Alternative substrates
Carbonyl sulfide (COS) is an isoelectronic structural analog
of CO2 and it may, therefore, serve as an alternative sub-
strate for NstCcmM209. A COS stock solution was pre-
pared by sparging ice-cold acidified water with a
compressed gas mixture containing 1.05 % v/v COS in
nitrogen. Based on COS solubility in water, the stock
solution contained 0.631 mM COS (Miller et al. 1989; So
et al. 2002a). The capped solution was stored on ice and
periodically re-sparged during the day. COS can be directly
measured by aqueous inlet mass spectrometry at m/z = 60.
COS stock solution was injected into the mass spectrom-
eter cuvette containing 100 mM EPPS/NaOH pH 7,
20 mM MgSO4 at 30 �C to establish an initial concentra-
tion. The spontaneous hydrolysis of COS was then mea-
sured over time. A protein sample was subsequently
introduced into the reaction cuvette, and the effect of the
protein on the m/z = 60 signal was followed over time.
The ability of NstCcmM209 to hydrolyze p-
nitrophenylacetate was tested at room temperature (about
22 �C). The enzyme was incubated with 1.5 mM p-
nitrophenylacetate in the above buffer, pH 7.2, and the
change in A348 was followed over time.
Effect of temperature on the CA activity
The effect of temperature on NstCcmM209 CA activity
was determined by incubating protein samples at select
temperatures between 5 and 35 �C for 15 min and then
assaying for activity at the same temperature using the 18O
exchange method. CA activity above 35 �C could not be
reliably measured due to excessive permeability of the
membrane inlet. The thermostability of NstCcmM209 was
determined by incubating protein samples for 15 min at
select temperatures between 10 and 75 �C, and then
assaying for CA activity using the 18O exchange method
under standard conditions at 30 �C.
Fluorescence spectroscopy
A Fluoromax 4 (HORIBA Scientific) spectrometer equip-
ped with a Peltier unit for temperature control was used to
obtain intrinsic tryptophan fluorescence emission spectra
from protein samples in 20 mM Tris–HCl, pH 7.5–8.0.
Proteins solutions (1 lM) were excited at 290 nm (Trp
excitation, 1 nm band width), and emission spectra (6 nm
band width) were collected from 300 to 450 nm over a
range of temperatures from 10 to 95 �C. Protein solutions
were incubated for 60 s at each temperature increment
prior to recording the spectrum. Maximum fluorescence
emission was observed at 343 nm in all cases, and this
value was used for subsequent analysis.
Circular dichroism (CD) spectroscopy
The CD spectra (195–300 nm) of NstCcmM209 were
obtained using AVIV 250 CD spectrometer (Aviv Bio-
medical Inc., Lakeview, NJ, USA) in a quartz cuvette with
a 1mm path length. At each temperature from 10 to 80 �C,
the protein solution (10 lM) was scanned three times and
an average spectrum was determined.
138 Photosynth Res (2014) 121:135–150
123

Results
Identification and evaluation of candidate
carboxysomal carbonic anhydrases
The identity of the Nostoc PCC 7120 carboxysomal CA has
not been determined experimentally. Candidate CAs in the
proteome were identified by BlastP searches of the NCBI
database (http://www.ncbi.nlm.nih.gov) using the sequen-
ces of experimentally validated a, b, c, and d CAs as
search query. Proteins directly corresponding to previously
identified carboxysomal CAs, CcaA or CsoSCA, were not
found in the Nostoc PCC 7120 proteome when queried
with Synechocystis PCC 6803 CcaA (NCBI accession
AAC46375) or Halothiobacillus neapolitanus CsoSCA
(EEG96215), respectively. Similarly, no d-CA homolog
was found when the proteome was queried with T.
weissflogii CA1 (AAX08632). Four potential candidates
were identified; the a-CA, EcaA (NP_486969, all2929), the
b-CA, EcaB (NP_486950, all2910), and the c-CAs, CcmM
(NP_484908, all0865) and NP_485067 (alr1024), currently
annotated as a ferripyochelin-binding protein.
EcaA was previously identified as a catalytically inac-
tive a-CA homolog localized to the cell surface (So et al.
1998; Soltes-Rak et al. 1997). Analysis of the amino acid
sequence of EcaA using the TatP 1.0 algorithm (http://
www.cbs.dtu.dk/services/TaTP/) revealed the presence of
an N-terminal twin arginine translocation signal, RRQLL,
consistent with EcaA export to the periplasmic space. The
b-class CA homolog, EcaB, from Synechocystis PCC 6803
has been previously identified as a periplasmic protein in a
large-scale proteomic experiment (Fulda et al. 2000).
However, recombinant EcaB was unable to catalyze 18O
exchange, suggesting that it lacks CA activity (So et al.
1998). TatP analysis of the Nostoc PCC 7120 EcaB
sequence indicated the presence of an N-terminal twin
arginine signal sequence with an RRNFL motif, consistent
with a periplasmic localization.
The two candidate c-CA-like proteins identified in the
Nostoc PCC 7120 proteome are the 555 amino acid CcmM
(NP_484908), and the 202 amino acid polypeptide anno-
tated as ferripyochelin-binding protein (NP_485067).
NstCcmM209 (the first 209 amino acids) has 72, 70, 65,
and 62 % amino acid sequence identity to the same region
of TeCcmM, Synechococcus PCC 7002 CcmM, SycCcmM,
and SynCcmM, respectively, and 40 % identity to MtCam.
This protein has all of the residues currently known to be
required for c-CA activity, and the N- and C-terminal
motifs demonstrated to be necessary in TeCcmM for acti-
vation (Fig. 1). NP_485067, in contrast, is only 30 %
identical to TeCcmM209, and lacks the RbcS-like C-ter-
minal domains, the CcmM activation motifs, and lacks two
residues (E56, N185) demonstrably important for CA
activity. This protein is also only 25 % identical to both
MtCam and MtCamH. While NP_485067 is 40 % identical
to PgiCA, a recently characterized c-CA from the anaer-
obic eubacterium Porphyromonas gingivalis (Del Prete
et al. 2013), it should be noted that PgiCA is inhibited by
almost all anions, including chloride (KI = 0.9 mM), and it
is only in a perchlorate buffer that activity has been dem-
onstrated. The actual physiological efficacy of this enzyme
as a CA within the cell is, therefore, debatable. As
NP_485067 has none of the motifs associated with carb-
oxysome localization or activation, and is only distantly
related to established c-CAs, we do not consider it further.
CA activity in Nostoc is detectable
only in the carboxysomal fraction
The sensitive 18O exchange assay was not able to detect
CA activity on the external cell surface of intact Nostoc
PCC 7120 cells (200 lg Chl mL-1) or in equivalent cel-
lular lysate (Fig. 2a). Following Percoll-Mg2? fraction-
ation of cell lysates, however, a 2.6-fold enhancement in18O exchange was readily detected in the TP pellet
(Fig. 2a). This fraction is enriched in carboxysomes and
carboxysome fragments (Long et al. 2005, 2007). Other
fractions devoid of carboxysomes lacked enhanced 18O
exchange activity, suggesting that CA activity was pre-
dominately localized to the carboxysome fraction. 18O
exchange activity was inhibited by the classic CA inhibitor
ethoxyzolamide (EZ) and by denaturing conditions
(Fig. 2a, c) indicating the presence of a protein catalysis in
the TP pellet.
NstCcmM has CA activity localized within the N-
terminal domain
The full-length ccmM gene product with an N-terminal T7
affinity tag, T7-NstCcmM555, was overexpressed in
E. coli. CA activity was detected in crude cell lysates but
not in cell lysates that lacked the expression plasmid.
However, CA activity was not observed following centri-
fugation of the lysate. Most of T7-NstCcmM555 was
subsequently found in insoluble inclusion bodies. Treat-
ment of the inclusion bodies with urea solubilized T7-
NstCcmM555 and yielded a protein with strong 18O
exchange activity, diagnostic for CA (Fig. 2b). 18O
exchange activity in the urea-treated protein fraction
obtained from control E. coli cells that harbored the
pET21b null construct alone was similar to uncatalyzed
control levels (Fig. 2b). Complete removal of urea from
T7-NstCcmM555 resulted in protein precipitation and loss
of catalytic activity. Similar outcomes were observed for
an N-terminal 6X-His-tagged version of NstCcmM555 (not
shown).
Photosynth Res (2014) 121:135–150 139
123

To circumvent these problems, a 6X-His-tagged version
of CcmM containing the first 209 amino acids
(NstCcmM209) was constructed and purified using IMAC.
NstCcmM209 proved to be highly soluble (at 3–5 mg/mL)
and showed 18O exchange activity (Fig. 2c) comparable to
that observed for T7-NstCcmM555. NstCcmM209 and
TeCcmM209 SS*=
TeCcmM209 1 ------MAVQ---------------SYAA-PP-TPWSRDLAEPEIAPTAYVHSFSNLIGDNstCcmM209 1 ------MAVR---------------STAA-PP-TPWSRSLAEAQIHESAFVHPFSNIIGDMtCam 1 QEITVDEFSN---------------IREN-PV-TPWNPEPSAPVIDPTAYIDPQASVIGEMtCamH 1 -------------------------MKRNFKMHLP-NPHKQHPKVSKRAWISETALIIGNPgiCA 1 ---------MAQRENSDYLTTKMALIQSV-R--------GFTPIIGEDTFLAENATIVGD
TeCcmM209 SS # #* * *# + *
TeCcmM209 38 VRIKDYVHIAPGTSIRADE-GTPFHIGSRT-NIQDGVVIHGLQQ----------GRVIGDNstCcmM209 38 VHIGANVIIAPGTSIRADE-GTPFHIGENT-NIQDGVVIHGLEQ----------GRVVGDMtCam 44 VTIGANVMVSPMASIRSDE-GMPIFVGDRS-NVQDGVVLHALETINEEGEPIEDNIVEVDMtCamH 35 VSIADDVFVGPNAVLRADEPGSSITV-HRGCNVQDNVVVHSLSH--S-------------PgiCA 43 VVMGKGCSVWFNAVLRGDV--NSIRIGDNV-NIQDGSILHTLYQ----------------
TeCcmM209 SS = + +
TeCcmM209 86 DGQEYSVW-IGDNVSITHMALIHGPAYIGDGCFIGFRSTVFN-ARVGAGC-VVMMHVLI-NstCcmM209 86 DNKEYSVW-VGSSASLTHMALIHGPAYVGDNSFIGFRSTVFN-AKVGAGC-IVMMHALI-MtCam 102 -GKEYAVY-IGNNVSLAHQSQVHGPAAVGDDTFIGMQAFVFK-SKVGNNC-VLEPRSAA-MtCamH 79 ---E--VL-IGKNTSLAHSCIVHGPCRIGEDCFIGFGAVVFD-CNIGKDTLVLH-KSIV-PgiCA 84 ----KSTIEIGDNVSVGHNVVIHGA-KICDYALIGMGAVVLDHVVVGEGA-IVAAGSVVL
TeCcmM209 SS * *
TeCcmM209 142 QDVEIPPGKYVPSGMVITTQQQADRLPNVEE--SDIHFAQ-HVVGINEALLSGY-Q--CANstCcmM209 142 KDVEVPPGKYVPSGAIITNQKQADRLPDVQP--QDRDFAH-HVIGINQALRAGY-L--CAMtCam 157 IGVTIPDGRYIPAGMVVTSQAEADKLPEVTDDYAYSHTNE-AVVYVNVHLAEGY-K--ETMtCamH 129 RGVDISSGRMVPDGTVITRQDCADALEDITK--DLTEFKR-SVVKANIDLVEGYIRLREEPgiCA 138 TGTQIEPNSIYA-GAP------ARFIKKVDP--EQSREMNFRIAHNYRMYASWF-KDESS
TeCcmM209 SS
TeCcmM209 196 ENIACIAPIRNELQ 209NstCcmM209 196 ADSKCIAPLRNDQV 209MtCam 213 S------------- 213MtCamH 187 S------------- 187PgiCA 188 E-------IDNP-- 192
Fig. 1 Structure-based multiple sequence alignment of the currently
known, catalytically active c-CAs: TeCcmM209, Thermosynecho-
coccus elongatus BP1 CcmM (NCBI Accession NP_681734);
NstCcmM209, Nostoc PCC 7120 CcmM (NP_484908) this study;
MtCam, Methanosarcina thermophilia Cam (P40881); MtCamH, M.
thermophilia CamH (ACQ57353), and PgiCA, Porphyromonas
gingivalis W83 PgiCA (NP_905402). Sequences were aligned using
T-Coffee—Expresso (http://www.tcoffee.org) and PDB (http://www.
rcsb.org) model structures 3kwcF (TeCcmM209 & NstCcmM209),
1qreA (MtCam), 3ou9A (MtCamH), and 3vnpC (PgiCA). The SS of
TeCcmM209 is shown; b, b-sheet; a, a-helix. Symbols: red inverse,
80 % or greater sequence identity; yellow black, 80 % or greater
sequence similarity; bold and bold underline strain specific sequences
relevant to catalysis; hash mark structurally relevant residues identi-
fied in MtCam; asterisk catalytically relevant residues identified in
MtCam; ?, metal ligands identified in MtCam and TeCcmM209; C C,
Cys194–Cys200 disulfide bond identified in TeCcmM209 as catalyt-
ically relevant; =, source of Trp fluorescence in NstCcmM 209
140 Photosynth Res (2014) 121:135–150
123
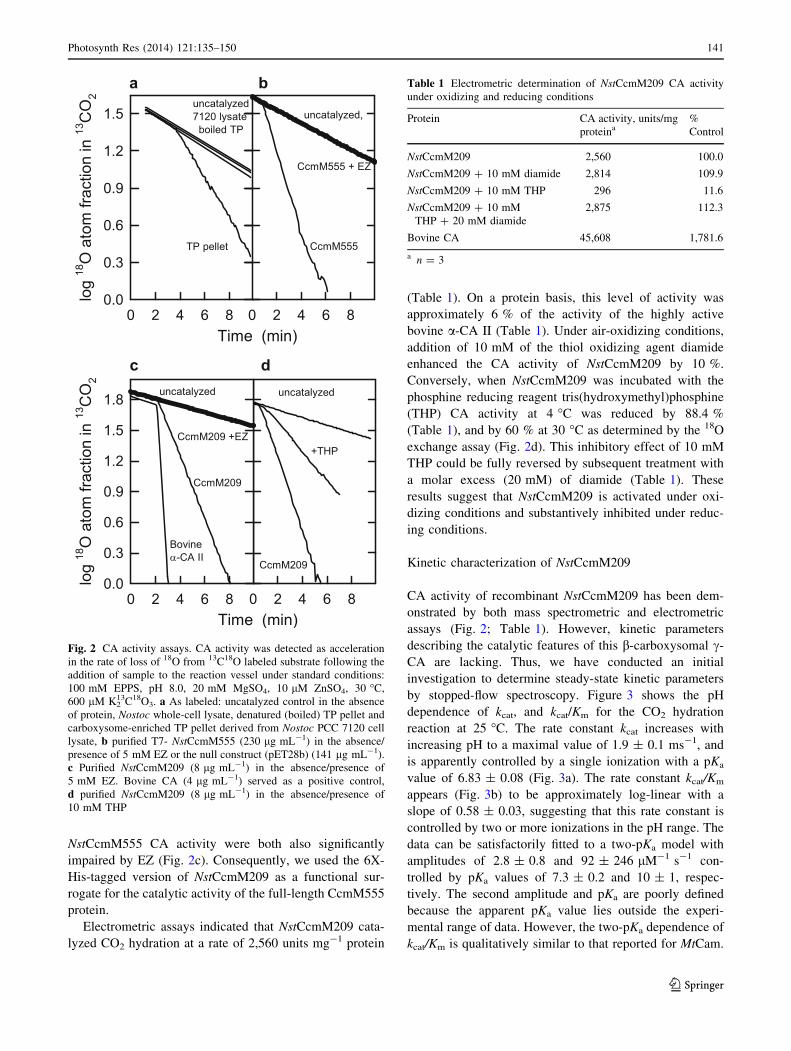
NstCcmM555 CA activity were both also significantly
impaired by EZ (Fig. 2c). Consequently, we used the 6X-
His-tagged version of NstCcmM209 as a functional sur-
rogate for the catalytic activity of the full-length CcmM555
protein.
Electrometric assays indicated that NstCcmM209 cata-
lyzed CO2 hydration at a rate of 2,560 units mg-1 protein
(Table 1). On a protein basis, this level of activity was
approximately 6 % of the activity of the highly active
bovine a-CA II (Table 1). Under air-oxidizing conditions,
addition of 10 mM of the thiol oxidizing agent diamide
enhanced the CA activity of NstCcmM209 by 10 %.
Conversely, when NstCcmM209 was incubated with the
phosphine reducing reagent tris(hydroxymethyl)phosphine
(THP) CA activity at 4 �C was reduced by 88.4 %
(Table 1), and by 60 % at 30 �C as determined by the 18O
exchange assay (Fig. 2d). This inhibitory effect of 10 mM
THP could be fully reversed by subsequent treatment with
a molar excess (20 mM) of diamide (Table 1). These
results suggest that NstCcmM209 is activated under oxi-
dizing conditions and substantively inhibited under reduc-
ing conditions.
Kinetic characterization of NstCcmM209
CA activity of recombinant NstCcmM209 has been dem-
onstrated by both mass spectrometric and electrometric
assays (Fig. 2; Table 1). However, kinetic parameters
describing the catalytic features of this b-carboxysomal c-
CA are lacking. Thus, we have conducted an initial
investigation to determine steady-state kinetic parameters
by stopped-flow spectroscopy. Figure 3 shows the pH
dependence of kcat, and kcat/Km for the CO2 hydration
reaction at 25 �C. The rate constant kcat increases with
increasing pH to a maximal value of 1.9 ± 0.1 ms-1, and
is apparently controlled by a single ionization with a pKa
value of 6.83 ± 0.08 (Fig. 3a). The rate constant kcat/Km
appears (Fig. 3b) to be approximately log-linear with a
slope of 0.58 ± 0.03, suggesting that this rate constant is
controlled by two or more ionizations in the pH range. The
data can be satisfactorily fitted to a two-pKa model with
amplitudes of 2.8 ± 0.8 and 92 ± 246 lM-1 s-1 con-
trolled by pKa values of 7.3 ± 0.2 and 10 ± 1, respec-
tively. The second amplitude and pKa are poorly defined
because the apparent pKa value lies outside the experi-
mental range of data. However, the two-pKa dependence of
kcat/Km is qualitatively similar to that reported for MtCam.
Fig. 2 CA activity assays. CA activity was detected as acceleration
in the rate of loss of 18O from 13C18O labeled substrate following the
addition of sample to the reaction vessel under standard conditions:
100 mM EPPS, pH 8.0, 20 mM MgSO4, 10 lM ZnSO4, 30 �C,
600 lM K213C18O3. a As labeled: uncatalyzed control in the absence
of protein, Nostoc whole-cell lysate, denatured (boiled) TP pellet and
carboxysome-enriched TP pellet derived from Nostoc PCC 7120 cell
lysate, b purified T7- NstCcmM555 (230 lg mL-1) in the absence/
presence of 5 mM EZ or the null construct (pET28b) (141 lg mL-1).
c Purified NstCcmM209 (8 lg mL-1) in the absence/presence of
5 mM EZ. Bovine CA (4 lg mL-1) served as a positive control,
d purified NstCcmM209 (8 lg mL-1) in the absence/presence of
10 mM THP
Table 1 Electrometric determination of NstCcmM209 CA activity
under oxidizing and reducing conditions
Protein CA activity, units/mg
proteina%
Control
NstCcmM209 2,560 100.0
NstCcmM209 ? 10 mM diamide 2,814 109.9
NstCcmM209 ? 10 mM THP 296 11.6
NstCcmM209 ? 10 mM
THP ? 20 mM diamide
2,875 112.3
Bovine CA 45,608 1,781.6
a n = 3
Photosynth Res (2014) 121:135–150 141
123

The maximal kcat value and the low-pH amplitude of kcat/
Km values fall between the values previously reported for
the Zn2?-containing forms of MtCam and MtCamH
(Zimmerman et al. 2010), but are more than an order of
magnitude lower than those reported for PgiCA (Del Prete
et al. 2013) (Table 2).
Alternative substrates for CcmM
In addition to the reversible hydration of CO2, a-CA enzymes
are known to possess other catalytic activities including
phosphatase and esterase activities with 4-nitrophenyl phos-
phate or 4-nitrophenyl acetate as substrate (Pocker and
Sarkanen 1978). However, b- or c-CAs generally do not seem
to catalyze these reactions (Heinhorst et al. 2006; Innocenti
and Supuran 2010; Innocenti et al. 2004). Similarly,
NstCcmM209 did not display esterase activity against
4-nitrophenyl acetate (not shown). However, like a-CAs
(Chengelis and Neal 1979; Miller et al. 1989; Protoschill-
Krebs and Kesselmeier 1992), NstCcmM209 did catalyze the
decomposition of COS in a substrate and protein concentra-
tion-dependent manner while heat denatured enzyme lost this
ability (Fig. 4) indicating that native protein structure was
essential for expression of this activity. The uncatalyzed
hydrolysis of COS proceeds through an acid-labile mono-
thiocarbonate intermediate that slowly decomposes to CO2
and H2S (Miller et al. 1989). Acidification of the reaction
medium following the consumption of COS by NstCcmM209
did not result in any COS reappearance as would be expected
if there had been a build-up of the reaction intermediate. Our
initial data, therefore, indicate that NstCcmM209 catalyzes
the complete hydrolysis of COS.
Fig. 3 pH dependence of the steady-state kinetic parameters kcat (a),
and kcat/Km (b) for the CO2 hydration reaction at 25 �C catalyzed by
NstCcmM209. Data for kcat were fit to the logarithmic form of the
equation k(obs) = k(max)/(1 ? [H?]/Ka), where k(obs) is the observed
value of kcat, k(max) is the maximal value of kcat at high pH, and Ka
relates to the ionization constant controlling the reaction. For ak(max) = 1.93 9 104 s-1 ± 0.15, pKa = 6.83 ± 0.08. Data for kcat/
Km were fit to a log-linear function with slope 0.58 ± 0.03 (solid line)
and to the logarithmic form of the equation kobs ¼ k1
1þ½Hþ�
Ka1
þ k2
1þ½Hþ�
Ka2
,
where kobs is the value of kcat/Km at any given pH, k1
(2.9 9 106 ± 0.8 M-1 s-1) and k2 (92 9 106 ± 246 M-1 s-1) are
the low and high-pH amplitudes of kcat/Km, and Ka1 and Ka2 are the
controlling acid dissociation constants (pKa1 = 7.3 ± 0.2;
pKa2 = 10 ± 1) (dashed line)
Table 2 Comparison of steady-state kinetic parameters for the CO2
hydration reaction
Enzyme kcat (s-1) Km
(mM)
kcat/Km
(M-1 s-1)
pH/
(�C)
NstCcmM209a 2.2 9 104 5.2 4.1 9 106 8.0/25
NstCcmM209a 1.9 9 104 10.2 1.9 9 106 7.5/25
Zn-MtCamb 11.6 9 104 15.6 7.5 9 106 7.5/25
Zn-MtCamHb 0.05 9 104 3.1 0.16 9 106 7.5/25
PgiCAc 41 9 104 7.6 5.4 9 107 7.5/20
CsoSCAd 8.9 9 104 3.2 2.8 9 107 8.0/25
Data was obtained using stopped-flow spectroscopy at the indicated
pH and temperature and are derived from a this study, b Zimmerman
et al. (2010), c Del Prete et al. (2013) and d Heinhorst et al. (2006)
Fig. 4 Time courses of COS consumption mediated by
NstCcmM209. Dissolved COS (5.5 lM) was introduced into the
reaction chamber of an aqueous inlet mass spectrometer, and COS
disappearance was measured over time at m/z = 60 in the absence or
presence (upward arrow) of NstCcmM209 (45–354 lg mL-1). Fol-
lowing COS consumption the reaction medium was acidified to pH 4
with HCl (asterisk) to determine residual [COS]
142 Photosynth Res (2014) 121:135–150
123
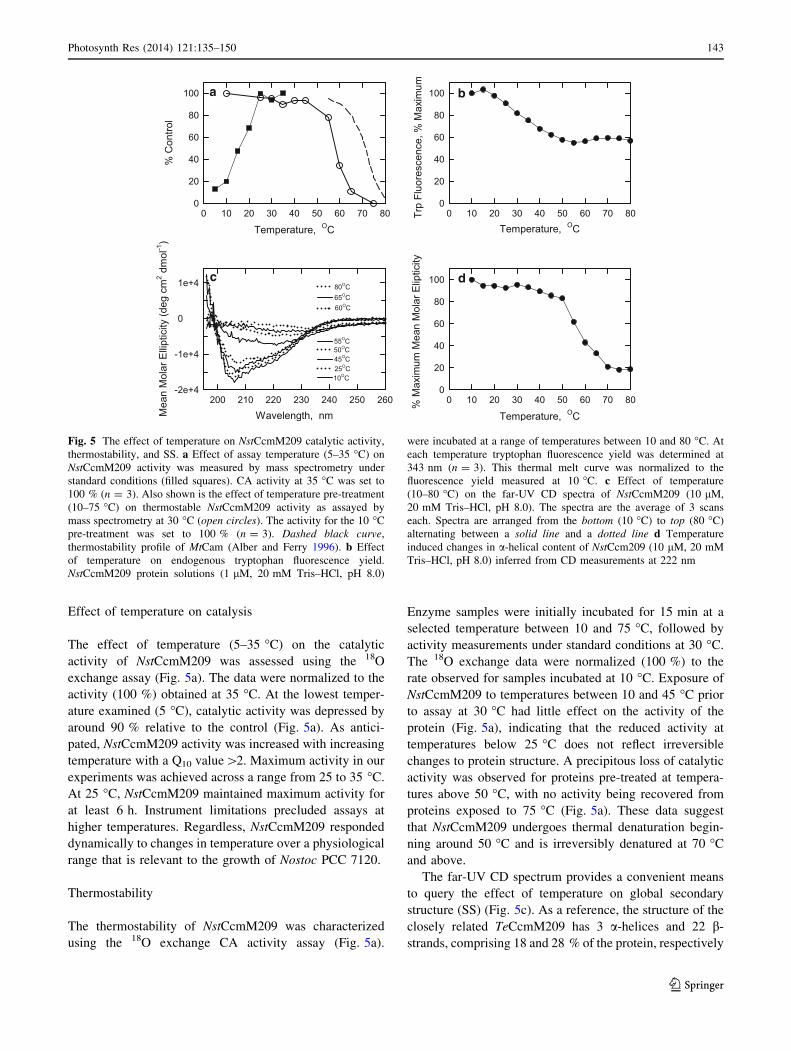
Effect of temperature on catalysis
The effect of temperature (5–35 �C) on the catalytic
activity of NstCcmM209 was assessed using the 18O
exchange assay (Fig. 5a). The data were normalized to the
activity (100 %) obtained at 35 �C. At the lowest temper-
ature examined (5 �C), catalytic activity was depressed by
around 90 % relative to the control (Fig. 5a). As antici-
pated, NstCcmM209 activity was increased with increasing
temperature with a Q10 value[2. Maximum activity in our
experiments was achieved across a range from 25 to 35 �C.
At 25 �C, NstCcmM209 maintained maximum activity for
at least 6 h. Instrument limitations precluded assays at
higher temperatures. Regardless, NstCcmM209 responded
dynamically to changes in temperature over a physiological
range that is relevant to the growth of Nostoc PCC 7120.
Thermostability
The thermostability of NstCcmM209 was characterized
using the 18O exchange CA activity assay (Fig. 5a).
Enzyme samples were initially incubated for 15 min at a
selected temperature between 10 and 75 �C, followed by
activity measurements under standard conditions at 30 �C.
The 18O exchange data were normalized (100 %) to the
rate observed for samples incubated at 10 �C. Exposure of
NstCcmM209 to temperatures between 10 and 45 �C prior
to assay at 30 �C had little effect on the activity of the
protein (Fig. 5a), indicating that the reduced activity at
temperatures below 25 �C does not reflect irreversible
changes to protein structure. A precipitous loss of catalytic
activity was observed for proteins pre-treated at tempera-
tures above 50 �C, with no activity being recovered from
proteins exposed to 75 �C (Fig. 5a). These data suggest
that NstCcmM209 undergoes thermal denaturation begin-
ning around 50 �C and is irreversibly denatured at 70 �C
and above.
The far-UV CD spectrum provides a convenient means
to query the effect of temperature on global secondary
structure (SS) (Fig. 5c). As a reference, the structure of the
closely related TeCcmM209 has 3 a-helices and 22 b-
strands, comprising 18 and 28 % of the protein, respectively
a b
c d
Fig. 5 The effect of temperature on NstCcmM209 catalytic activity,
thermostability, and SS. a Effect of assay temperature (5–35 �C) on
NstCcmM209 activity was measured by mass spectrometry under
standard conditions (filled squares). CA activity at 35 �C was set to
100 % (n = 3). Also shown is the effect of temperature pre-treatment
(10–75 �C) on thermostable NstCcmM209 activity as assayed by
mass spectrometry at 30 �C (open circles). The activity for the 10 �C
pre-treatment was set to 100 % (n = 3). Dashed black curve,
thermostability profile of MtCam (Alber and Ferry 1996). b Effect
of temperature on endogenous tryptophan fluorescence yield.
NstCcmM209 protein solutions (1 lM, 20 mM Tris–HCl, pH 8.0)
were incubated at a range of temperatures between 10 and 80 �C. At
each temperature tryptophan fluorescence yield was determined at
343 nm (n = 3). This thermal melt curve was normalized to the
fluorescence yield measured at 10 �C. c Effect of temperature
(10–80 �C) on the far-UV CD spectra of NstCcmM209 (10 lM,
20 mM Tris–HCl, pH 8.0). The spectra are the average of 3 scans
each. Spectra are arranged from the bottom (10 �C) to top (80 �C)
alternating between a solid line and a dotted line d Temperature
induced changes in a-helical content of NstCcm209 (10 lM, 20 mM
Tris–HCl, pH 8.0) inferred from CD measurements at 222 nm
Photosynth Res (2014) 121:135–150 143
123

(Pena et al. 2010) (Fig. 1). At 10 �C, the CD spectrum of
NstCcmM209 is dominated by a large swing in mean molar
ellipticity from a high positive value at 195 nm to a low
negative value at 206–208 nm. The positive band at 195 nm
and negative bands at 208 and 222 nm are characteristic of
a-helical proteins, while proteins with well-defined b-sheets
typically have a positive band at 195 nm and a minimum at
218 nm (Greenfield 2007; Kelly et al. 2005). The latter
features are not resolved in our spectra; however, the
atypical phi-psi angles seen in left-handed b-helices along
with the large numbers of identically structured turns may
result in spectral features that differ from those seen with
more typical, twisted b-sheets. The CD spectra change little
between 10 and 45 �C, but above 50 �C show a progressive
loss of mean molar ellipticity and a shifting of the minimum
upstream, indicative of protein unfolding (Fig. 5c, d)
(Greenfield 2007). The loss of CD signals at 208 and
222 nm in particular indicates a rapid decline in a-helical
content above 50 �C (Fig. 5d) and provides a mechanistic
explanation for the loss of CA activity in NstCcmM209
exposed to temperatures over 50 �C (Fig. 5a). Near com-
plete denaturation of NstCcmM209 SS was observed at
70 �C (Fig. 5c, d), with the presence of protein aggregates
indicated from an increase in molar ellipticity observed
between 240 and 260 nm.
We also used intrinsic tryptophan fluorescence to follow
structural changes in NstCcmM209 as a function of tem-
perature. Tryptophan fluorescence yield decreases as a
result of increasing solvent quenching and, therefore, pro-
vides information on the extent to which this hydrophobic
residue is buried by native protein structure. NstCcmM209
has two tryptophan residues. Trp13 is located in the b1-b2
loop within a conserved motif (APPTPWS, Fig. 1). This
loop extends to within 1.5 nm of the active site and also
makes contact with the aC helix (Pena et al. 2010). Trp93
is located on b13 of the left-handed b-helix and appears to
be shielded by the b11-b12 loop and, therefore, is protected
from the solvent. Trp13 and/or Trp93 were increasingly
solvent exposed as the temperature increased from 10 to
55 �C (Fig. 5b). There is an abrupt transition in the fluo-
rescence yield curve at 55 �C suggesting a major reorga-
nization of the protein. The fact that tryptophan
fluorescence remained relatively constant from 60 �C
onward suggests that these residues are fully solvent
exposed and disordered.
CcmM and Rubisco are present in Nostoc
carboxysomes
SDS-PAGE and Western blot analysis indicated that purified
NstCcmM209 (MW = 24.9 kDa) and T7-NstCcmM555
(MW = 60.7 kDa) immunologically cross-reacted with an
antibody (anti-CcmM NT) directed against the N-terminal
domain of CcmM (Fig. 6a). A second antibody, anti-CcmM
CT (directed against the C-terminal region of CcmM), was
also tested against Nostoc PCC 7120 cell lysate and E. coli
cell lysate expressing recombinant 6X-His-NstCcmM555
(Fig. 6b). Two protein bands were observed to cross-react in
the Nostoc PCC 7120 lysate with approximate MWs of 59
and 35 kD. The first protein corresponds to the predicted
size of the full-length NstCcmM555 (59.4 kD), while the
second protein corresponds to the C-terminal ‘‘short form’’
of CcmM (35.4 kD) that is translated from an internal
ribosome-binding site within the ccmM transcript (Long
et al. 2010). No other protein bands were observed. The
same two immune-reactive bands were also observed in
E. coli lysate harboring the ccmM expression plasmid, but
not in the null control.
The presence of CcmM in the TP pellet was verified by
SDS-PAGE analysis which revealed 23 major protein
bands (Fig. 7). TP pellets were also analyzed by 2D LC–
MS/MS and a number of carboxysomal proteins were
positively identified as significant matches to proteins from
the Nostoc PCC 7120 proteome database (Table S2). These
included RbcL, RbcS, CcmM555, CcmK1, and CcmK2.
The other proteins identified included hypothetical pro-
teins, transcriptional regulator proteins, and a set of 30S
and 50S ribosomal proteins, similar to the contaminants
found in TP pellets from Synechococcus PCC 7942 (Long
et al. 2007). CcmL, CcmN, and CcmO were not among the
identified proteins.
Western blot analysis of the TP pellet confirmed the
presence of RbcL, CcmM555, and the 35.4 kDa short form
of CcmM (Fig. 7). RbcS was detected as a faint band using
anti-RbcL/S (Fig. 7). A single protein around the mass
expected for CcmK was identified using an antibody to the
shell protein CcmK2 raised against the Synechococcus
PCC 7942 protein. The CcmM short form has been pre-
viously identified in Nst-TP pellets by Western analysis
and its identity verified by Edman sequencing of the
N-terminal 7 amino acids (Long et al. 2007).
The cellular distribution of Rubisco and CcmM was then
analyzed by immunogold electron microscopy using either
anti-RbcL/S or anti-CcmM CT. A total of 100 medial cell
sections were analyzed in each instance. On average, each
cross section of an air-grown Nostoc PCC 7120 cell had 1.6
carboxysomes (range of 0 to 7) distributed through the cell.
Nostoc PCC 7120 carboxysomes are typically larger (400-
600 nm on the long axis) and more irregularly shaped (Fig.
S1) than the smaller, polygonal carboxysomes (100-
200 nm in diameter) found in many unicellular cyanobac-
teria (Espie and Kimber 2011; Long et al. 2007; Rae et al.
2013). When Nostoc PCC 7120 cells were labeled with
anti-RbcL/S, 49 % of the gold particles were found local-
ized within carboxysomes (Table 3; Fig. S1) while the
remaining gold particles were distributed throughout the
144 Photosynth Res (2014) 121:135–150
123

cytoplasm and sporadically over the thylakoid membranes.
Gold particle density was 15.2 times higher in the carb-
oxysome than in the rest of the cell, indicating that Rubisco
is highly concentrated within these structures, in agreement
with the observations of Jager and Bergman (1990). When
Nostoc PCC 7120 cells were labeled with anti-CcmM CT
about one-third of the gold particles were located in visu-
ally identified carboxysomes (Table 3; Fig. S1). Again, the
remainder of the gold particles was distributed throughout
the cell. Similar to Rubisco, CcmM was also found to be
concentrated (ninefold) within the carboxysome relative to
the cytoplasm (Table 3). In general, CcmM seemed to be
distributed throughout the carboxysome lumen and was not
preferentially associated with the shell. This antibody
detects both CcmM555 and the short form and cannot,
therefore, resolve the specific location of either form within
the carboxysome.
Discussion
Carboxysomes are specialized microcompartments that
support the photoautotrophic growth of cyanobacteria under
normal environmental conditions. These structures are
encapsulated by a unique protein shell in which both Rubisco
and its substrate, CO2, are concentrated. Once encapsulated,
Rubisco is a permanent resident of the carboxysome (Chen
et al. 2013) where it mediates CO2 fixation. In contrast,
carboxysomal CO2 must be constantly generated from the
cytosolic pool of HCO3-, which is itself created by the active
transport of inorganic carbon from the environment into the
cell. Crucial to the function of the cyanobacterial CCM is a
carboxysome localized, luminal CA that generates the CO2
required to sustain Rubisco at near maximal rates of fixation.
The behavior and catalytic characteristics of the carbox-
ysomal CA are, therefore, important for understanding both
the CCM and carboxysome function and factors that limit
primary productivity. In contrast to a-cyanobacteria which
have only CsoSCA, b-cyanobacteria have two dis-
tinct carboxysomal CAs (CcaA and CcmM) that have been
identified and characterized to various degrees. Given the
diversity of b-cyanobacteria, however, we cannot yet
exclude the possibility that there are additional, unidentified
carboxysomal CAs within this lineage. Identification of b-
carboxysomal CAs is complicated by our inability to isolate
b-carboxysomes in pure form. Experimental investigations
have also been confined primarily to three unicellular model
organisms, which represent only a small fraction of the
genetic diversity of b-cyanobacteria. To address these lim-
itations, we undertook research to identify and characterize
the carboxysomal CA from the filamentous, diazotrophic,
and mesophilic cyanobacterium Nostoc sp PCC 7120.
Fig. 6 Western blot analysis using polyclonal antibodies directed
against a the N-terminal c-CA domain of CcmM: lane 1 molecular
weight markers; lane 2 purified 6X-His NstCcmM209
(0.89 lg mL-1); lane 3 purified 6X-His NstCcmM555
(0.102 lg mL-1); and b C-terminal domain of CcmM: lane 1
molecular weight markers; lane 2 pET21b vector null construct
(0.110 lg mL-1) or 6X-His NstCcmM209, lane 3 purified 6X-His
NstCcmM555 (140 lg mL-1); lane 4, E. coli cell lysate expressing
6X-His NstCcmM555; lane 5 molecular weight markers; lane 6, cell
lysate (200 lg mL-1) from low inorganic-carbon-acclimated Nostoc
PCC 7120
Photosynth Res (2014) 121:135–150 145
123

CcmM is a b-carboxysomal c-carbonic anhydrase
Full length, recombinant NstCcmM555 catalyzed the
reversible hydration of CO2 and by all criteria is a func-
tional CA. CA activity was readily detected in the carb-
oxysome-enriched TP pellet which was shown by Western
blot and 2D-LC–MS/MS to contain CcmM, along with
Rubisco. Immunogold electron microscopy provided fur-
ther evidence that CcmM is concentrated within the carb-
oxysome and co-localizes with Rubisco. On the other hand,
other candidate CAs (EcaA, EcaB, NP_485067) were
systematically excluded as the carboxysomal CA based on
bioinformatics analysis and the results of previous studies.
Consequently, the collective evidence indicates that CcmM
is the sole functional carboxysomal CA of the mesophilic
cyanobacterium Nostoc PCC 7120 and that CcmM cata-
lytic function is not restricted to thermophilic
cyanobacteria.
Based on gold particle distribution, CcmM was also
found in the cytoplasm in unexpectedly large amounts and
in proximity to thylakoid and plasma membranes (Table 3;
Fig. S1). Western blots of Nostoc PCC 7120 cell lysates
using the same anti-CcmM CT antibody as used in the
immunogold experiments indicated the presence of only
two immune-reactive proteins that corresponded to the
CcmM555 and the 35 kDa short form of CcmM. Thus, a
non-specific antibody reaction complex cannot account for
the observed distribution of CcmM in the cytoplasm of
Nostoc PCC 7120 cells. Two large-scale proteomics
experiments have also noted the presence of CcmM (but
not Rubisco) in isolated outer membrane fractions from
both Nostoc PCC 7120 and Synechocystis PCC 6803
(Huang et al. 2004; Moslavac et al. 2005), while both RbcL
and the CcmM short form were found in peripheral
membrane fractions from Synechocystis PCC 6803 using
TrxA-linked/mass spectrometric identification method
(Mata-Cabana et al. 2007). These findings provide some
evidence for a membrane-associated CcmM in addition to
the carboxysome-localized CcmM. In their transmission
electron microscope study of the architecture and biogen-
esis of carboxysomes from the closely related Anabaena
PCC 7119, Orus et al. (2001) observed a variety of carb-
oxysome shapes and textures ranging from electron-trans-
lucent to electron-intermediate and mature, electron-dense
structures. In our experiments, preservation of immature
carboxysomes may have been compromised due to the
reduced amount of fixative used to preserve antigenicity of
the thin sections employed for the immunogold experi-
ments. Consequently, some of the cytoplasmic CcmM may
be associated with developing carboxysomes whose
structures are poorly resolved under our conditions. In
1 2 3 4250150100
75
50
37
2520
15
10
CcmM555
CcmMshort
CcmK2
RbcL
RbcS
Fig. 7 SDS-PAGE (lane 1) and Western blot analysis (lane 2, 3, 4) of
the carboxysome-enriched TP pellet obtained from Nostoc PCC 7120
cell lysate following Percoll-Mg2? fractionation. Lane 1 Bio-Rad
stain-free acrylamide gel showing protein-banding pattern from
enriched carboxysome preparations (TP pellets). Lanes 2–4, Western
blots of SDS-PAGE separated TP pellet probed with; lane 2, anti-
CcmK2; lane 3 anti-CcmM; lane 4 anti-RbcL/S. Blots for lane 4 are
for different loadings of protein for RbcL and RbcS due to the relative
difficulty in detecting RbcS. In this case the antibodies were raised
against the homologous proteins from Synechococcus sp. PCC 7942.
Molecular weight markers are indicated. CcmK2 appears as an
indistinct band below RbcS with Coomassie staining and is not visible
on Bio-Rad stain-free gels due to the absence of tryptophan residues.
Additional proteins identified from TP pellets are indicated in Table
S2
Table 3 Immuno-localization of Rubisco and CcmM in Nostoc PCC
7120
Parametera Anti-
Rubisco
Anti-C-terminal
CcmM
Carboxysome
Gold particles in cbx 1,274 501
Total cbx area (lm2) 11.3 18.9
Average cbx area (lm2)/section 0.11 0.19
Cbx: gold particles (lm2) 112.6 26.4
Cbx/section 1.42 1.77
Cytosol
Gold particles cytosol 1,674 874
Total cytosol area (lm2) 225.3 290.0
Average cytosol area (lm2)/section 2.3 2.9
Cytosol: gold particles (lm2) 7.4 3.0
Total gold particles 2,948 1,375
Cytosol refers to the area of the section minus carboxysome area and
includes the thylakoid membranes, carboxysome = cbxa A total of 100 medial cell sections were analyzed for gold particle
distribution in each case. The sections contained 142 (anti Rubisco)
and 177 (anti-CcmM) carboxysomes respectively. In control experi-
ments lacking antibody, gold particle density was \0.1/lm2 and was
subtracted from the experimental value
146 Photosynth Res (2014) 121:135–150
123

support of this is the observation that nearly 50 % of the
immunogold labeled Rubisco was also found in the cyto-
plasm. The presence of Rubisco outside the mature carb-
oxysome has been noted frequently (Friedberg et al. 1993)
and for Nostoc PCC 7120 in particular (Jager and Bergman
1990). Recently Chen et al. (2013), using live cell fluo-
rescence microscopy, have shown that the first step in the
biogenesis of Synechococcus sp. PCC 7942 carboxysomes
involves the growth of un-encapsulated Rubisco aggregates
in the cytoplasm, providing direct evidence for a second
cellular reservoir for Rubisco outside the mature carboxy-
some. Depending on the study and the cyanobacterial
species investigated, the cytosolic Rubisco content ranges
from about 5–65 % of the total (Agarwal et al. 2009;
McKay et al. 1993). The co-occurrence of CcmM and
Rubisco in the cytoplasm is consistent with a Rubisco-
centric model for b-carboxysome biogenesis (Chen et al.
2013; Rae et al. 2013; Long et al. 2007, 2010) which
proposes that b-carboxysomes are formed around a Rubi-
sco-CcmM organizing complex that is subsequently
encapsulated by shell proteins. Additional experiments are
required, however, to determine if cytosolic CcmM and
Rubisco actually co-localize and form these complexes in
the cytoplasm of Nostoc PCC 7120 cells.
The presence of CcmM in the cytoplasm could potentially
thwart CCM function by prematurely dissipating the cyto-
solic pool of HCO3- (Price and Badger 1989). Indeed, Nostoc
PCC 7120 has substantial capacity to concentrate HCO3-
with a cytosolic pool as high as 60 mM and a [Ci]in/[Ci]out as
high as 360 (McGinn et al. 1997). However, we did not detect
CA activity in concentrated whole-cell lysates. Only when
the TP pellet was separated from other cytoplasmic compo-
nents did the CA activity of the carboxysome become
apparent (Fig. 2). The activity of NstCcmM209 was signifi-
cantly diminished in the presence of the phosphine reducing
agent THP (Table 1; Fig. 2). Thus, the highly reducing
environment of the cytoplasm, particularly in the light, may
prohibit CcmM catalysis outside the mature carboxysome
(Pena et al. 2010) thereby preventing a short circuit in the
CCM. Pena et al. (2010) have shown that reversible Cys194–
Cys200 disulfide/thiol formation mediates the oxidative
activation/reductive inhibition of TeCcmM 209. These resi-
dues are conserved in NstCcmM and provide a potential
mechanism to explain the redox activation/inhibition of this
enzyme. Chen et al. (2013) have recently shown that the
carboxysome remains a reducing environment as it assem-
bles, and only switches to being oxidizing late in its bio-
genesis following recruitment of the shell proteins. Our initial
data indicate that this could be the case for NstCcmM and
suggest that redox activation of the carboxysomal c-CA may
be a general feature of b-carboxysomes that rely on CcmM
catalysis. A detailed analysis of the redox regulation of
NstCcmM209 is currently in preparation.
Kinetic and biophysical characterization of CcmM
Kinetic analysis of NstCcmM209 was conducted in an air-
oxidizing environment as experimental evidence indicated
that sustained, near-maximum activity was maintained
under these conditions (Table 1). The kinetic parameters
for CO2 hydration, kcat and kcat/Km, increased with
increasing pH, indicating that an unprotonated form of
NstCcmM209 is required for activity, consistent with the
widely accepted metal-hydroxide mechanism of CA
catalysis (Alber et al. 1999; Rowlett et al. 2002). The
overall rate-pH profile for the kinetic parameters was
similar to those previously reported for MtCam though the
absolute values were consistently lower (Alber et al. 1999)
(Fig. 3; Table 2), while the NstCcmM209 kinetic parame-
ters were 10–40-fold higher than those reported for
MtCamH (Zimmerman et al. 2010). Under similar condi-
tions, the kcat and kcat/Km for CO2 hydration of
NstCcmM209 were four-fold and 6.8-fold lower than the
corresponding values for CsoSCA from the a-carboxysome
(Heinhorst et al. 2006). Possibly, this similarity in catalytic
values reflects the common function both enzymes play in
the CCM, though the differing size of the carboxysomes
(a-carboxysomes are considerably smaller), organization
(CsoSCA associates tightly with the shell) and Rubisco:
CA stoichiometry complicates direct comparisons. Like
NstCcmM209, CsoSCA is inhibited by thiol reducing
agents (So et al. 2004) though the underlying mechanism is
unknown; it is, therefore, possible that oxidative activation
of carboxysomal CAs is a general phenomenon.
The rate constant kcat is typically controlled by the pKa
of the proton shuttle residue when proton transfer is rate-
limiting; this is typical of most CAs (Rowlett et al. 2002).
For NstCcmM209, the pKa controlling kcat is 6.83 ± 0.08,
which is consistent with Glu78 performing this role. Glu78
in NstCcmM209 is analogous to Glu84 in MtCam (Fig. 1),
which has been shown through mutagenesis to be the
proton shuttle residue in that enzyme (Tripp and Ferry
2000).
The pH-rate profile of kcat/Km for NstCcmM209 is
apparently log-linear, and is not consistent with a single
controlling pKa. The data suggest that two or more pKa
values control kcat/Km in NstCcmM209. In MtCam, kcat/Km
is controlled by two-pKa values with two distinct ampli-
tudes for kcat/Km at low and high pH. We have fit the data
of Fig. 3 in the same fashion, although the high-pH
amplitude and pKa values are not well determined. The
low-pH pKa (7.3 ± 0.2) should correspond to the pKa value
of the metal-bound water molecule, and this value is
qualitatively similar to that of MtCam (Tripp and Ferry
2000).
In addition to the reversible hydration of CO2, our initial
investigation shows that NstCcmM209 also catalyzes the
Photosynth Res (2014) 121:135–150 147
123

hydrolysis of COS, the most abundant sulfur-containing
compound in the atmosphere and a greenhouse gas. This is
the first example of this activity in a c-CA. The carbox-
ysomal b-CA, CcaA, from Synechocystis PCC 6803 (So
et al. 2002a) and a-CAs from diverse organisms including
rat, cow, and the green alga Chlamydomonas also catalyze
COS hydrolysis (Chengelis and Neal 1979; Miller et al.
1989; Protoschill-Krebs and Kesselmeier 1992) suggesting
that this reaction may be a general biochemical feature of
all CAs arising from a common reaction mechanism, as is
the case for the CO2 hydration reaction. COS is also an
alternative substrate, but not an activator, of Rubisco
yielding 3-phosphoglycerate and 1-thio-3-phosphoglycer-
ate as products of the thiocarboxylation of RuBP (Lorimer
and Pierce 1989). The presence of both CA and Rubisco in
carboxysomes suggests that this bacterial organelle may,
therefore, play a role in the biological degradation of COS.
The kinetic effectiveness of carboxysomes in this process
is yet to be demonstrated, but given the sheer number and
global distribution of cyanobacteria, carboxysomes could
be an important environmental sink for COS.
NstCcmM209 displayed a broad pH optimum (8–9.5)
for CO2 hydration activity. Evaluation of enzyme activity
at different temperatures by the 18O exchange assay also
revealed a broad temperature optimum (25–35 �C), which
spans the optimal temperature range for the growth of
Nostoc PCC 7120. Enzyme activity and protein SS were
reversibly stable to thermal treatment between 5 and 45 �C,
a range broader than likely experienced during the normal
physiological growth of Nostoc PCC 7120. A precipitous
loss of activity occurred following temperature treatments
above 50 �C. CD spectra and intrinsic fluorescence emis-
sion spectra revealed a significant loss in a-helical content
and exposure of buried Trp residues that tracked the loss in
activity. These results demonstrate that irreversible thermal
denaturation of NstCcmM209 occurred above 50 �C that
accounts directly for the loss in enzyme activity and may,
therefore, be one factor that limits the growth of Nostoc
PCC 7120 to mesophilic temperatures. By comparison, the
thermal denaturation activity curve for MtCam is shifted
about 10 �C higher (Alber and Ferry 1996), (Fig. 5) con-
sistent with the moderate thermophilic nature of M. ther-
mophila (optimum growth temperature of 55 �C). A similar
shift in the thermal denaturation activity curve of TeCcmM
might, therefore, be anticipated as this enzyme also
evolved and functions in a moderately thermophilic envi-
ronment, unlike NstCcmM. However, we have not found
substantive structural or compositional differences between
the two enzymes to account for any difference in thermo-
tolerance. Further experimentation and analysis are
required to resolve this issue.
CcmM is a characteristic protein of all b-carboxysomes
with an established role as a structural protein and as a
binding partner for CcaA, the conventional carboxysomal
CA in the model organisms Synechocystis sp. PCC 6803
and Synechococcus sp. PCC 7942. Our demonstration that
NstCcmM and TeCcmM (Pena et al. 2010) are both func-
tional c-CAs indicate that the use of CcmM as the carb-
oxysomal CA is likely widespread in cyanobacterial strains
that lack a ccaA gene. Many strains, however, encode what
appear to be a functional CcaA and a functional CcmM
(Pena et al. 2010). The advantage of expressing two dis-
tinct CA enzymes within the same b-carboxysome is
unknown and is currently under investigation.
Acknowledgments This study was supported by grants from the
Natural Sciences and Engineering Research Council of Canada
(NSERC) to G. S. E. and M. S. K. and an NSERC CGSD to C. A.; by
a grant from the Australian Research Council to B. M. L. and G.
D. P., and by a grant from the U.S.A. National Science Foundation,
MCB-1157332, to R. S. R.
References
Agarwal R, Ortleb S, Sainis JK, Melzer M (2009) Immunoelectron
microscopy for locating Calvin cycle enzymes in the thylakoids
of Synechocystis 6803. Mol Plant 2:32–42
Alber BE, Ferry JG (1994) A carbonic anhydrase from the archaeon
Methanosarcina thermophila. Proc Natl Acad Sci USA
91:6909–6913
Alber BE, Ferry JG (1996) Characterization of heterologously
produced carbonic anhydrase from Methanosarcina thermophila.
J Bacteriol 178:3270–3274
Alber BE, Colangelo CM, Dong J, Stalhandske CMV, Baird TT, Tu
C, Fierke CA, Silverman DN, Scott RA, Ferry JG (1999) Kinetic
and spectroscopic characterization of the gamma-carbonic
anhydrase from the methanoarchaeon Methanosarcina thermo-
phila. Biochem 38:13119–13128
Berry S, Fischer JH, Kruip J, Hauser M, Wildner GF (2005)
Monitoring cytosolic pH of carboxysome-deficient cells of
Synechocystis sp PCC 6803 using fluorescence analysis. Plant
Biol 7:342–347
Chen AH, Robinson-Mosher A, Savage DF, Silver PA, Polka JK
(2013) The bacterial carbon-fixing organelle is formed by shell
envelopment of preassembled cargo. PLoS One 8(9):e76127.
doi:10.1371/journal.pone.0076127
Chengelis CP, Neal RA (1979) Hepatic carbonyl sulfide metabolism.
Biochem Biophys Res Commun 90:993–999
Cot SSW, So AKC, Espie GS (2008) A multiprotein bicarbonate
dehydration complex essential to carboxysome function in
cyanobacteria. J Bacteriol 190:936–945
Del Prete S, Vullo D, De Luca V, Carginale V, Scozzafava A,
Supuran CT, Capasso C (2013) A highly catalytically active c-
carbonic anhydrase from the pathogenic anaerobe Porphyro-
monas gingivalis and its inhibition profile with anions and small
molecules. Bioorg Med Chem Lett 23:4067–4071
Espie GS, Kimber MS (2011) Carboxysomes: cyanobacterial Rubi-
sCO comes in small packages. Photosyn Res 109:7–20
Friedberg D, Jager KM, Kessel M, Silman NJ, Bergman B (1993)
Rubisco but not Rubisco activase is clustered in the carboxy-
somes of the cyanobacterium Synechococcus sp. PCC 7942:
mud-induced carboxysomeless mutants. Mol Microbiol
9:1193–1201
148 Photosynth Res (2014) 121:135–150
123

Fukuzawa H, Suzuki E, Komukai Y, Miyachi S (1992) A gene
homologous to chloroplast carbonic anhydrase (icfA) is essential
to photosynthetic carbon dioxide fixation by Synechococcus
PCC7942. Proc Natl Acad Sci USA 89:4437–4441
Fulda S, Huang F, Nilsson F, Hagemann M, Norling B (2000)
Proteomics of Synechocystis sp. strain PCC 6803: identification
of periplasmic proteins in cells grown at low and high salt
concentrations. Eur J Biochem 267:5900–5907
Greenfield NJ (2007) Using circular dichroism spectra to estimate
protein secondary structure. Nat Protocols 1:2876–2890
Hackenberg C, Huege J, Engelhardt A, Wittink F, Laue M, Matthijs
HCP, Kopka J, Bauwe H, Hagemann M (2012) Low-carbon
acclimation in carboxysome-less and photorespiratory mutants
of the cyanobacterium Synechocystis sp. strain PCC 6803.
Microbiology 158:398–413
Heinhorst S, Williams EB, Cai F, Murin CD, Shively JM, Cannon GC
(2006) Characterization of the carboxysomal carbonic anhydrase
CsoSCA from Halothiobacillus neapolitanus. J Bacteriol
188:8087–8094
Huang F, Hedman E, Funk C, Kieselbach T, Schroder WP, Norling B
(2004) Isolation of outer membrane of Synechocystis sp PCC
6803 and its proteomic characterization. Mol Cell Proteomics
3:586–595
Innocenti A, Supuran CT (2010) Paraoxon, 4-nitrophenyl phosphate
and acetate are substrates of a- but not of b-, c- and f-carbonic
anhydrases. Bioorg Med Chem Lett 20:6208–6212
Innocenti A, Zimmerman S, Ferry JG, Scozzafava A, Supuran CT
(2004) Carbonic anhydrase inhibitors. Inhibition of the b-class
enzyme from the methanoarchaeon Methanobacterium thermo-
autotrophicum (Cab) with anions. Bioorg Med Chem Lett
14:4563–4567
Jager KM, Bergman B (1990) Localization of a multifunctional chaperonin
(GroELprotein) in nitrogen-fixing Anabaena PCC 7120: presence in
vegetative cells and heterocysts. Planta 183:120–125
Kelly SM, Jess TJ, Price NC (2005) How to study proteins by circular
dichroism. Biochim Biophys Acta 1751:119–139
Khalifah RG (1971) Carbon dioxide hydration activity of carbonic
anhydrase. J Biol Chem 246:2561–2573
Kinney JN, Salmeen A, Cai F, Kerfeld CA (2012) Elucidating
essential role of conserved carboxysomal protein CcmN reveals
common feature of bacterial microcompartment assembly. J Biol
Chem 287:17729–17736
Lichtle C, McKay RML, Gibbs SP (1992) Immunogold localization
of Photosystem-I and Photosystem-II light-harvesting complexes
in cryptomonad thylakoids. Biol Cell 74:187–194
Long BM, Price GD, Badger MR (2005) Proteomic assessment of an
established technique for carboxysome enrichment from Syn-
echococcus PCC7942. Can J Bot 83:746–757
Long BM, Badger MR, Whitney SM, Price GD (2007) Analysis of
carboxysomes from Synechococcus PCC7942 reveals multiple
rubisco complexes with carboxysomal proteins CcmM and
CcaA. J Biol Chem 282:29323–29335
Long BM, Tucker L, Badger MR, Dean Price G (2010) Functional
cyanobacterial b-carboxysomes have an absolute requirement for
both long and short forms of the CcmM protein. Plant Physiol
153:285–293
Lorimer GH, Pierce J (1989) Carbonyl Sulfide -An alternate substrate
for but not an activator of ribulose-1,5-bisphosphate carboxylase.
J Biol Chem 264:2764–2772
Ludwig M, Sultemeyer D, Price GD (2000) Isolation of ccmKLMN
genes from the marine cyanobacterium, Synechococcus sp.
PCC7002 (Cyanophyceae), and evidence that CcmM is essential
for carboxysome assembly. J Phycol 36:1109–1118
Marcus Y, Berry JA, Pierce J (1992) Photosynthesis and photores-
piration in a mutant of the cyanobacterium Synechocystis PCC
6803 lacking carboxysomes. Planta 187:511–516
Mata-Cabana A, Florencio FJ, Lindahl M (2007) Membrane proteins
from the cyanobacterium Synechocystis sp PCC 6803 interacting
with thioredoxin. Proteomics 7:3953–3963
McGinn PJ, Canvin DT, Coleman JR (1997) Influx and efflux of
inorganic carbon during steady-state photosynthesis of air-grown
Anabaena variabilis. Can J Bot 75:1913–1926
McKay RML, Gibbs SP, Espie GS (1993) Effect of dissolved
inorganic carbon on the expression of carboxysomes, localiza-
tion of Rubisco and the mode of inorganic carbon transport in
cells of the cyanobacterium Synechococcus UTEX 625. Arch
Microbiol 159:21–29
Miller AG, Espie GS, Canvin DT (1989) Use of carbon oxysulphide,
a structural analog of CO2, to study active CO2 transport in the
cyanobacterium Synechococcus UTEX 625. Plant Physiol
90:1221–1231
Miller AG, Salon C, Espie GS, Canvin DT (1997) Measurement of
the amount and isotopic composition of the CO2 released from
the cyanobacterium Synechococcus UTEX 625 after rapid
quenching of the active CO2 transport system. Can J Bot
75:981–997
Moslavac S, Bredemeier R, Mirus O, Granvogl B, Eichacker LA,
Schleiff E (2005) Proteomic analysis of the outer membrane of
Anabaena sp strain PCC 7120. J Proteome Res 4:1330–1338
Orus MI, Rodriguez-Buey ML, Marco E, Fernandez-Valiente E
(2001) Changes in carboxysome structure and grouping and in
photosynthetic affinity for inorganic carbon in Anabaena strain
PCC 7119 (cyanophyta) in response to modification of CO2 and
Na? supply. Plant Cell Physiol 42:46–53
Pena KL, Castel SE, De Araujo C, Espie GS, Kimber MS (2010)
Structural basis of the oxidative activation of the carboxysomal
c-carbonic anhydrase, CcmM. Proc Natl Acad Sci USA
107:2455–2460
Pocker Y, Sarkanen S (1978) Carbonic anhydrase: structure catalytic
versatility, and inhibition. Adv Enzymol Relat Areas Mol Biol
47:149–274
Price GD, Badger MR (1989) Isolation and characterization of high
CO2-requiring-mutants of the cyanobacterium Synechococcus
PCC 7942–2 phenotypes that accumulate inorganic carbon but
are apparently unable to generate CO2 within the carboxysome.
Plant Physiol 91:514–525
Price GD, Badger MR, Woodger FJ, Long BM (2008) Advances in
understanding the cyanobacterial CO2-concentrating- mecha-
nism (CCM): functional components, Ci transporters, diversity,
genetic regulation and prospects for engineering into plants.
J Expt Bot 59:1441–1461
Protoschill-Krebs G, Kesselmeier J (1992) Enzymatic pathways for
the comsumption of carbonyl sulphide (COS) by higher plants.
Bot Acta 105:206–212
Rae BD, Long BM, Badger MR, Price GD (2013) Functions,
compositions, and evolution of the two types of carboxysomes:
polyhedral microcompartments that facilitate CO2 fixation in
cyanobacteria and some proteobacteria. Microbiol Mol Biol Rev
77:357–379
Rowlett RS, Gargiulo NJ, Santoli FA, Jackson JM, Corbett AH (1991)
Activation and inhibition of bovine carbonic anhydrase-III by
dianions. J Biol Chem 266:933–941
Rowlett RS, Tu C, McKay MM, Preiss JR, Loomis RJ, Hicks KA,
Marchione RJ, Strong JA, Donovan GS, Chamberlin JE (2002)
Kinetic characterization of wild-type and proton transfer-
impaired variants of b-carbonic anhydrase from Arabidopsis
thaliana. Arch Biochem Biophys 404:197–209
Sawaya MR, Cannon GC, Heinhorst S, Tanaka S, Williams EB,
Yeates TO, Kerfeld CA (2006) The structure of b-carbonic
anhydrase from the carboxysomal shell reveals a distinct
subclass with one active site for the price of two. J Biol Chem
281:7546–7555
Photosynth Res (2014) 121:135–150 149
123

Silverman DN (1982) Carbonic anhydrase: oxygen-18 exchange
catalyzed by an enzyme with rate-contributing proton-transfer
steps. Methods Enzymol 87:732–753
So AKC, Espie GS (2005) Cyanobacterial carbonic anhydrases. Can J
Bot 83:721–734
So AKC, Van Spall HGC, Coleman JR, Espie GS (1998) Catalytic
exchange of 18O from 13C18O-labelled CO2 by wild-type cells
and ecaA, ecaB, and ccaA mutants of the cyanobacteria
Synechococcus PCC7942 and Synechocystis PCC6803. Can J
Bot 76:1153–1160
So AKC, Cot SSW, Espie GS (2002a) Characterization of the
C-terminal extension of carboxysomal carbonic anhydrase from
Synechocystis sp. PCC6803. Funct Plant Biol 29:183–194
So AKC, John-McKay M, Espie GS (2002b) Characterization of a
mutant lacking carboxysomal carbonic anhydrase from the
cyanobacterium Synechocystis PCC6803. Planta 214:456–467
So AKC, Espie GS, Williams EB, Shively JM, Heinhorst S, Cannon
GC (2004) A novel evolutionary lineage of carbonic anhydrase
(e Class) is a component of the carboxysome shell. J Bacteriol
186:623–630
Soltes-Rak E, Mulligan ME, Coleman JR (1997) Identification and
characterization of a gene encoding a vertebrate a- type carbonic
anhydrase in cyanobacteria. J Bacteriol 179:769–774
Tripp BC, Ferry JG (2000) A structure-function study of a proton
transport pathway in the gamma-class carbonic anhydrase from
Methanosarcina thermophila. Biochemistry 39:9232–9240
Wilbur KM, Anderson NG (1948) Electrometric and colorimetric
determination of carbonic anhydrase. J Biol Chem 176:147–154
Yu JW, Price GD, Song L, Badger MR (1992) Isolation of a putative
carboxysomal carbonic anhydrase gene from the cyanobacterium
Synechococcus PCC7942. Plant Physiol 100:794–800
Zimmerman SA, Tomb JF, Ferry JG (2010) Characterization of
CamH from Methanosarcina thermophila, founding member of a
subclass of the c class of carbonic anhydrases. J Bacteriol
192:1353–1360
150 Photosynth Res (2014) 121:135–150
123





























