Embed Size (px)
Citation preview

i
OGWUEGBU, MERCY CHISARA
PG/M.Sc./14/76559
GROWTH PERFORMANCE, CARCASS TRAIT, ORGAN CHARACTERISTICS
AND LIPID PROFILE OF BROILERS FED DIETS CONTAINING
DIFFERENT FAT SOURCES
DEPARTMENT OF ANIMAL SCIENCE
FACULTY OF AGRICULTURE
Godwin Valentine
Digitally Signed by: Content manager’s Name
DN : CN = Webmaster’s name
O= University of Nigeria, Nsukka
OU = Innovation Centre

ii
GROWTH PERFORMANCE, CARCASS TRAIT, ORGAN CHARACTERISTICS
AND LIPID PROFILE OF BROILERS FED DIETS CONTAINING
DIFFERENT FAT SOURCES
BY
OGWUEGBU, MERCY CHISARA
PG/M.Sc./14/76559
DEPARTMENT OF ANIMAL SCIENCE,
FACULTY OF AGRICULTURE,
UNIVERSITY OF NIGERIA, NSUKKA
MARCH, 2016

i
GROWTH PERFORMANCE, CARCASS TRAIT, ORGAN CHARACTERISTICS
AND LIPID PROFILE OF BROILERS FED DIETS CONTAINING
DIFFERENT FAT SOURCES
A PROJECT SUBMITTED TO THE DEPARTMENT OF ANIMAL SCIENCE,
UNIVERSITY OF NIGERIA, NSUKKA, IN PARTIAL FULFILLMENT OF THE
REQUIREMENTS FOR AWARD OF DEGREE OF MASTER OF SCIENCE (M.Sc.) IN
ANIMAL SCIENCE, (ANIMAL NUTRITION AND BIOCHEMISTRY) OF THE
UNIVERSITY OF NIGERIA, NSUKKA.
BY
OGWUEGBU, MERCY CHISARA
PG/M.Sc./14/76559
SUPERVISORS: DR. A.O. ANI
MARCH, 2016.

ii
CERTIFICATION
I, OGWUEGBU, MERCY CHISARA, a postgraduate student in the Department of Animal
Science, with registration number, PG/M.Sc/14/76559, have satisfactorily completed the
requirements for research work for the award of the degree of Masters of Science in the
Department of Animal Science (Nutrition and Biochemistry). This work is original and has
not been submitted in part or full for any other diploma or degree of this or any other
University.
_________________ ______________
Ogwuegbu, Mercy C.
Student Date
__________________ ________________
Dr. A.O. Ani Date
Supervisor
________________ ________________
Prof. A.G. Ezekwe Date
H.O.D
__________________ ________________
External examiner Date

iii
DEDICATION
This work is dedicated to God the Father, God the Son and God the Holy Spirit.

iv
ACKNOWLEDGEMENT
My profound gratitude and indebtedness are first and foremost to His Majesty, God Almighty
for His sufficient grace and incomparable mercy towards me even before the world began. I
am also indebted to a number of distinguished personalities who helped me in measures to
make this research work a success. I owe a lot of gratitude to my supervisor, Dr. A.O. Ani for
his kindness, support, assistance, encouragement, patience and unique approach to
instructions and teachings. My sincere prayer is that your aim of bringing out the best in me
will not be thwarted. God bless you daddy.
I wish to appreciate my Head of Department, Prof. A.G Ezekwe for providing a sound
atmosphere for learning. Remain blessed Sir. I wish also to appreciate these dynamic and
erudite Professors- D.N. Onah of Veterinary Medicine, C.C Nwosu, G.C. Okeke and S.O.C.
Ugwu of the Department of Animal Science, University of Nigeria, Nsukka for their humane
approach to teaching and fatherly advice. Also recognized are: Dr. A.E. Onyimonyi, Dr.
(Mrs.) H.N. Folhngu, Mr. O.C. Osita and all members and staff of the Department of Animal
Science, UNN for their concern and input in one way or the other which have enabled me
achieve this feat, May God bless them.
To my parents, Mr. and Mrs. I.O Ogwuegbu, and my lovely siblings: Onyekachukwu and
Makuochukwu who always believed in me and my ability to attain greater height in life, I
want to say a big thank you for your support and encouragement, I love you all.
I will not forget my friends and colleagues: Micheal, Akumjika, Ogumba, Prisca,
Omumabuike, Jecinta, Egom, Marvin, Odo, Kenneth, Ilekuba, Chineye, etc. for all your
constructive criticisms, suggestions and ideas. Space will fail me to mention all. Thank you
enough.
My special thanks go also to all the members of Graduate Students’ Fellowship (GSF) UNN
for your prayers and encouragement. You are indeed a wonderful family. Remain blessed.
Meet you all at the top.

v
TABLE OF CONTENTS
Title page i
Certification ii
Dedication iii
Acknowledgement iv
Table of Contents v
List of Tables vii
Abstract viii
CHAPTER ONE: INTRODUCTION
1.1 Background of the study 1
1.2 Objectives of the study 2
1.3 Justification 2
CHAPTER TWO: LITERATURE REVIEW
2.1 Poultry Nutrition 3
2.2 Broilers 3
2.3 Nutrient Requirements for broilers 5
2.3.1 Energy 5
2.3.2 Protein and amino acids 7
2.3.3 Water 10
2.3.4 Minerals 10
2.3.5 Vitamins 11
2.4 Lipids in poultry nutrition 11
2.4.1 Lipid digestion 13
2.42 Lipid absorption 13
2.5 Dietary fats in poultry nutrition 15
2.5.1 Importance of dietary fats to poultry 16
2.5.2 Advantages of dietary fat supplementation 16
2.5.3 Fats Digestibility in young chicks 19
2.5.4 Efficient utilization of dietary fats 20
2.6 Sources of dietary fats in poultry nutrition 21
2.7 Effects of dietary fats on poultry performance 30

vi
2.8 Lipid profile 32
CHAPTER THREE: MATERIALS AND METHODS
3.1 Location and duration of study 37
3.2 Experimental materials 37
3.3 Experimental diets 37
3.4 Experimental animals and managements 37
3.5 Data collection and measurements 40
3.6 Carcass and Organ evaluation 40
3.7 Blood collection and analysis 40
3.8 Experimental Design 41
3.9 Proximate Analysis 42
3.10 Statistical analysis 42
CHAPTER FOUR: RESULTS AND DISCUSSION
4.1 Results 43
4.1.1 Proximate composition of experimental diets 43
4.1.2 Growth performance of broiler birds fed diets containing different fat sources 44
4.1.3 Carcass and organ characteristics of broiler birds fed diets containing different fat sources 47
4.1.4 Relative organ weight of broiler birds fed diets containing different fat sources 49
4.1.5 Lipid profile of broiler birds fed diets containing different fat sources 50
4.2 Discussion 52
4.2.1 Growth performance of broiler birds fed diets containing different fat sources 52
4.2.2 Carcass and organ characteristics of broiler birds fed diets containing different fat sources 53
4.2.3 Relative organ weights of broiler birds fed diets containing different fat sources 55
4.2.4 Lipid profile of broiler birds fed diets containing different fat sources 55
CHAPTER FIVE: SUMMARY AND DISCUSSION
5.1 Summary 57
5.2 Conclusion 58
5.3 Recommendation 58
References 59
Appendices 73

vii
LIST OF TABLES
Table page
1: Amino acid requirement at different ages of broiler birds 9
2: Percentage composition of broiler starter diets 38
3: Percentage composition of broiler birds finisher diets 39
4: Proximate composition of experimental diets 43
5: Growth performance of broiler birds fed diets containing different fat sources 44
6: Carcass and organ characteristics of broiler birds fed diets containing different fat
sources 47
7: Relative organ weights of broiler birds fed diet containing different fat sources 49
8: Lipid profile of broiler birds fed diets containing different fat sources 50

viii
ABSTRACT
This study was conducted to determine the effect of four different fat sources on growth performance, carcass trait, and organ characteristics and lipid profile of broiler birds reared to 8weeks of age. One hundred and twenty day-old broiler chicks were randomly assigned to five treatments, each with three replicates of 8 chicks in a completely randomized design experiment. The dietary treatments consisted of 1% added fat from four different sources as follows: T1, diet containing no oil; T2 diet containing 1% palm oil; T3, diet containing 1% soybean oil; T4, diet containing 1% palm kernel oil and T5, diet containing 1% groundnut oil. Dietary fat type affected significantly (p<0.05) body weight gain as well as feed efficiency in birds fed the palm kernel oil diet compared with those fed the other diets. The live weight, carcass weight and dressing % of birds on T4 was significantly different (p<0.05) from all other treatments. Most of the internal organ weights of birds on T4 and T1 were significantly (p<0.05) higher than in other treatments. The abdominal fat weight of birds on T4 was significantly (p<0.05) higher than those in the other treatment. Dietary treatment had significant effect on all the parameters evaluated in the lipid profile. Birds on T4 had significantly (p<0.05) higher level of total cholesterol while birds on T3 had lower level of triglyceride. Birds on T2 and T3 had similar (p>0.05) values of Low Density Lipoprotein cholesterol while the least value was observed in the control group. Birds on T2 had the highest High Density Lipoprotein Cholesterol though this was similar to the values observed in T1 and T4. Based on the results obtained in the present study it was concluded that addition of vegetable fat from palm kernel oil and groundnut oil in broiler diets enhanced the growth performance, carcass and organ weights of birds. Birds that consumed the diets containing soybean oil had the least very low density lipoprotein, while those that consumed diet containing palm kernel oil had similar High Density Lipoprotein Cholesterol value with birds on control.

1
CHAPTER ONE
INTRODUCTION
1.1 Background of the Study
The broiler industry has been developed worldwide to the most intensive and efficient model
in comparison to other animal production sectors. The rapid growth, efficient utilization of
feed, tender meat production, ability to thrives and produce under varied environmental
conditions have been achieved through advanced genetic improvement of the modern broiler
chicken. The energy intake of the fast growing broiler chickens fed ad libitum is estimated to
reach two or three times greater than their maintenance needs; and so carcass fat deposition
increases, causing broiler metabolic and skeletal disorders, beside its undesirable effect on
human health. Current commercial hybrids of broiler with high performance require high
energy diets, which would enable the maximum exploitation of their genetic potential.
Increasing demands for energy in growing chickens can be satisfied by the addition of fat to
the feeding mixtures. An increasing supplementation of diets with animal or plant fats or oils
for intensive poultry production has been observed (Sadeghi and Tabeidian, 2005).
The inclusion of fats and oil in animal feed is a common practice in modern poultry
production to increase the energy content of diet. Besides supplying energy, it improves the
absorption of fat-soluble vitamins (Baiao and Lara, 2005), provides essential fatty acids in
large quantity, diminishes the dustiness, increases the palatability of the rations and improves
the energy efficiency (Moav, 1995; Palmquist, 2002). Furthermore, dietary fat reduces
passage rate of the digesta through the gastrointestinal trait, allowing for better nutrient
absorption and utilization (Peebles et al., 2000; Baião and Lara, 2005; Latshaw, 2008).
Digestibility of dietary fats is affected by the fatty acid profiles and several studies have
shown better utilization of unsaturated fats leading to a higher metabolizing energy than
saturated fats (Celebi and Utlu, 2004). Jeffri et al. (2010) reported that increasing fat in
broiler diet will increase the amount of feed intake and feed efficiency.

2
To date, a number of different fat sources are available for poultry from both animal and
vegetable sources and from the rendering industry (Sanz et al., 2000). According to Rose
(2001), animal fat or vegetable oils may be used as a source of energy up to a maximum of
6% in the diet. Above this level it becomes difficult to maintain pellet quality, or to move
mechanically the sticky feed when it is not pelleted.
1.2 Objectives of the Study
The broad objective of the study is to evaluate the effect of dietary fat sources on growth
performance, carcass trait, organ characteristics and lipid profile of broilers.
Specific Objectives:
1. To determine the growth performance of broiler birds fed diets containing different
fat sources.
2. To determine the carcass and organ characteristics of broiler birds fed diets
containing different fat sources.
3. To evaluate the lipid profile of broiler birds fed diets containing different fat sources.
1.3 Justification of the Study
Broilers have unique potential as possible solution to Nigeria meat supply situation because
of some attributes like high growth rate, high efficient carcass yield, short production cycle
and good quality meat. Despite all these attributes, the productivity of poultry in Nigeria has
been low due to the problem of poor utilization of feed and high cost of feed. Current
commercial hybrids with high performance require high energy diets, which enable the
maximum exploitation of their genetic potential. Because of the increasing demand of energy
in growing chicks and the high cost of energy feed ingredient, the addition of fat or oil in low
energy feed can help increase the energy content of the feed and as well reduce the cost.
Besides fat metabolism and deposition in poultry can be affected by different dietary fat
intake and this can lead to poor nutrient absorption and utilization (Sanz et al., 1999; Peebles
et al., 2000; Balao and Lara, 2005).

3
CHAPTER TWO
LITERATURE REVIEW
2.1 Poultry Nutrition
The nutrition of animal involves various activities that result in the conversion of feed into
animal tissues or animal products, for example egg, milk, meat etc (Richard and Church,
1998). Pond et al. (1995) defined nutrition as a series of processes by which an organism
takes and assimilates food for promoting growth and replacing worn-out tissue. Also Obioha
(1992) defined nutrition as a process which a living organism takes in is used primarily for
two major purposes; maintenance and productive function.
Olomu (1995) emphasized that poultry nutrition is more critical than other farm animal
(except pig) because poultry birds are usually reared in confinement. Poultry are more active
and more sensitive to environmental influences than other farm animals. According to
Obioha (1992), Poultry have high growth rate, intensive metabolic rates and rapid developed
reproductive organ. He further reported that during the first ten weeks of post embryonic
growth, the weight of broilers increases for about thirty to forty times and that such rate of
growth has not been encountered in other farm animals.
The ultimate aim of poultry nutrition is to increase the productive efficiency of the poultry.
The economic importance of poultry feeding has become apparent, when it was realized that
feed is the highest single cost factor, about two third of the total production cost. Many cases
of poultry business failures can be traced to poor or improper feeding of the birds. Therefore,
it is pertinent that the right nutrients in adequate quantities and proportions must be supplied
to the birds. According to Richard and Church (1998), quality of ingredients as well as
quality of nutrients present must be evaluated.
2.2. Broilers
Broilers, also known as Cornish Cross, are a type of chicken raised specifically for meat
production. Produced by fast-growing breeds with low mortality, broilers can be reared
successfully in standard housing conditions on readily available, custom-formulated broiler
feed rations. Consumers expect the meat from broilers to be tender and of high quality. The
whole broiler production process is designed for this requirement but the same inputs are at

4
odds with those required for egg production by broiler breeders. The three main steps and
stages in the whole broiler production process are
• Rearing and managing broiler breeders (i.e. the birds that produce eggs for hatching
into broiler chicks)
• Fattening of broiler chicks
• Marketing and processing of finished broiler birds.
The broiler producer clearly requires birds that will achieve a high body weight, with good
carcass quality, over the shortest possible period of time using the minimum amount of
regular feed. In addition the producer also wants birds that posses the correct body
conformation, which will feather rapidly and have a minimal mortality rate.
Selection and breeding for fast growth rates in broilers form the most important processes in
the world poultry industry. Male broilers achieve rapid gain from the start, and at 6 weeks of
age can weigh at 2kg (live weight). Female birds will tend to grow at a slower rate but this
has definite marketing advantages because overall consumer demand is for broiler carcasses
of various weights. It is not the amount of feed consumed but the efficiency of feed
utilization and feed conversion into body tissue which underpins the growth rate.
Broilers producers tend to plump for white feathered strains because they result in a ‘cleaner-
looking ’carcass after processing. But there are instances where production management
considerations outweigh this and coloured-feathered strains are preferred. Examples include
broiler production in countries with high rainfall and the indigenous soil is red. In these
situations, red/brown Rohde Island Reds may be the most sensible choice. Feather cover
must be good to maintain insulation and restrict heat loss from the body, as well as
minimizing incidence of skin blistering which ruins marketability of processed birds. Many
modern strains of broilers will produce yellow fat because they have been custom-bred for
the American market. In markets where yellow fat is undesirable, producers should remove
carotene and carotenoid pigments (coloured chemical) from the ration. Similarly, factors that
determine carcass quality in one country may not suit another. For instance, consumers in
some countries may consider the body conformation, texture and taste of carcasses high
quality by ‘Western’ standards to offer an unattractive and insufficiently chewy eating
experience. For supermarket sales in general, breast meat should be broad and deep. Many

5
such problems are overcome by incorporating local strains into cross breeding programmes
to produce appropriate broiler parents stock.
2.3. Nutrients Requirement for Broilers
Poultry diets must be formulated to provided all the bird’s nutrient requirements if optimum
growth and production is to be achieved. On the basis of physiology structures of the poultry,
a good knowledge of the digestive system of birds could put the producer in a serious
problem. Nutrient can be defined as chemical substances found in feed ingredients which
when made available to the animal are used for maintenance, production and health of the
animals (Atteh, 2002). It can also be described as a specific chemical element or compound
supplied by or derived from the diet and absorbed into the body tissues to support
physiological processes. These nutrients are essential to be in appropriate amounts and
proportions in the diets of intensively housed birds, because they have no access to other
sources of nutrients. The essential nutrients required by the poultry birds are carbohydrates,
fat and oil (mostly served as energy givers); protein (amino acid), minerals (micro and
macro), vitamins and water (Pond et al., 1995; Olomu, 1995 and Smith, 2001).
The dietary requirement of nutrients varies with age, bodyweight and rate of growth of the
birds. Thus the requirement for any nutrient may be defined as the amount of that nutrient
which must be supplied in the diet to meet the needs of the normal healthy animal given on
otherwise completely adequate diet in an environment compatible with good health (Olomu,
1995). Such level of nutrient must meet the requirement for maintenance, growth, productive
and reproductive potential of the animal (NRC, 1994).
2.3.1. Energy
Energy is not a nutrient but a property of energy- yielding nutrients when they are oxidized
during metabolism. The dietary energy levels have been shown to affect broiler chickens’
feed intake. Nahashan et al. (2001) reported that as dietary energy level increases, birds
satisfy their energy needs by decreasing feed intake. Decreased in feed intake with high
energy levels in the diets of broiler chickens have also been reported by Leeson and
Summers (2001) and Veldkamp et al.,(2005). Thus, in formulating poultry diets, the nutrient
requirements of broiler chickens have frequently been expressed per unit of dietary
metabolisable energy (Gonzalez and Pesti, 1993). This practice is based on the theory that

6
birds will adjust their feed intake according to their metabolisable energy requirements and
was summarized by the NRC (1994) as an absolute requirement for energy in terms of
kilocalories per kilogram of diet cannot be stated because poultry adjust their feed intake to
obtain their necessary daily requirement.
Also, based on a re-evaluation of numerous research data, the NRC (1994) has revised their
previous conclusions by stating that the practice of relating nutrient concentrations as a
function of dietary metabolisable energy concentration while, as a result of the over-
consumption of energy on diets with a high metabolisable energy concentration, the
application of specific nutrient-to-metabolisable energy rations in broiler chickens and
turkeys should be re-evaluated. Leeson et al. (1996) showed that broiler chickens fed up to
25 and 49 days of age were able to adjust their feed intake to a constant energy intake over a
range of dietary metabolisable energy levels from 11.29 to 13.80 MJ ME/kg DM, which
indicated that broiler chickens retain an innate ability to eat to a fixed energy requirement
rather than to physical capacity as was suggested by Newcombe and Summers (1984).
However, on closer observation of the data of Leeson et al (1996), it can be seen that early
feed intake to 25 days of age was not greatly affected by dietary metabolisable energy
concentrations over the range of 12.13 to 13.80 MJ ME/kg DM that a significant increase in
feed intake occurred. Also, the effects of metabolisable energy concentration on feed intake
were very different between the early (0-25 days) and later (26-49 days) growth periods, with
the metabolisable energy concentration having a far greater effect on increasing feed intake
during the grower-finisher phase. This led to the overall conclusion by these authors that
broiler chickens do indeed eat to a constant metabolisable energy intake when viewed over
the entire 49-day growing period.
Contrary to the observation above, Richard (2003) observed that modern broiler chickens
selected for rapid growth do not regulate voluntary feed intake to achieve energy balance.
This altered ability of broiler chickens to adjust feed intake due to differences in
metabolisable energy density of the diet was postulated to result from continued selection for
rapid juvenile growth rates, which may have altered hypothalamic mechanisms that regulate
feed intake in broiler chickens (Bokkers and Koene, 2003). Other reports have also shown no
effect of dietary metaboisable energy concentration on feed intake between two groups of

7
broiler chickens fed ad-libitum diets containing two energy levels of 13.38 and 15 MJ ME/
kg DM.
2.3.2. Protein and Amino Acid
Dietary requirements for protein are actually requirements for the amino acids contained in
the dietary protein. Amino acids obtained from dietary protein are used by poultry to fulfill a
diversity of functions. For example, amino acids, as protein, are primary constituents of
structural and protective tissues, such as feather, bone matrix and ligaments, as well as of the
soft tissues, including organs and muscles. Also, amino acids and small peptides resulting
from digestion-absorption may serve a variety of metabolic functions and as precursors of
many important nonprotein body constituents. Because body proteins are in a dynamic state,
with synthesis and degradation occurring continuously, an adequate intake of dietary amino
acids is required. If dietary protein (amino acids) is inadequate, there is a reduction or
cessation of growth or productivity and a withdrawal of protein from less vital body tissues
to maintain the functions of more vital tissues.
There are 22 amino acids in body proteins and all are physiologically essential (NRC, 1994).
Nutritionally, ten of these are indispensable because chickens are unable to synthesize them
or cannot synthesize them at a rate sufficient to meet their needs. These are methionine,
lysine, threonine, leucine, valine, isoleucine, arginine, phenylalanine, histidine and
tryptophan (Austic, 1995; NRC, 1994).
The amino acid requirements of poultry represent the requirements for the indispensable
amino acids plus sufficient nitrogen in an appropriate chemical form for synthesis of the
dispensable amino acids. Chickens are sensitive to the dietary balance of these amino acids
(Austic, 1995). For the diet to be used with maximum efficiency the chicken must receive the
indispensable amino acids in the correct quantities and sufficient amino acids for metabolic
demands must be available. The presence of adequate amounts of nonessential amino acids in
the diets reduces the necessity of synthesizing them from essential amino acids. Amino acid
requirements may be classified as those for maintenance, carcass growth, egg production and
feather growth on the basis of their respective amino acid profiles (Hurwitz et al., 1978). In

8
order for the bird to realize its genetic potential and achieve the best levels of performance
through maximum rates of protein synthesis, amino acids must be provided in the necessary
quantities, avoiding both excesses and deficiencies (Sainbury, 1984). Thus, stating dietary
requirements for both protein and essential amino acids is an appropriate ways to ensure that
all amino acids needed physiologically are provided. Protein and amino acid requirements
vary considerably according to the physiological state of the bird, that is, the rate of growth
or egg production. Other factors contributing to variations in amino acid requirements of the
chickens include age, body size, sex and breed. Amino acid requirements decrease with age
and at the same time, the ideal balance of amino acids changes gradually to reflect those of
maintenance (Zubair and Leeson, 1996). For instance, the percentage of amino acid required
in the diet is the highest for young growing animals and gradually to maturity, when only
enough amino acid to maintain body tissue is required (Pond et al., 1995). The balance of
amino acids needed for maintenance is not proportional to the balance of amino acids in a
bird’s tissue, but rather reflects the relative rate of obligatory loss of each individual amino
acid (Gous et al., 1987).
Because of this reason, the balance needed for maintenance is considerably different from
that needed for growth or egg production (Nemavhola, 2001). Dietary amino acid levels
slightly below maintenance can sustain life, but muscle mass and functions are impaired
(Leeson et al., 1996). Matching the amino acid profile of the diet with animal requirements is
crucial for maximizing animal performance. For instance, turkey poults and broiler chickens
have high amino acid requirements to meet the needs for rapid growth while indigenous
chickens such as the Venda breed will require less amino acid to meet their needs because of
their slow growth rate and small body size. Because the contributions of maintenance and
growth to total amino acid requirement change with body size and the ideal amino acid
profiles for maintenance and growth are different, the composition of the ideal amino acid
pattern will change continuously during the growth period (Mack et al., 1999).
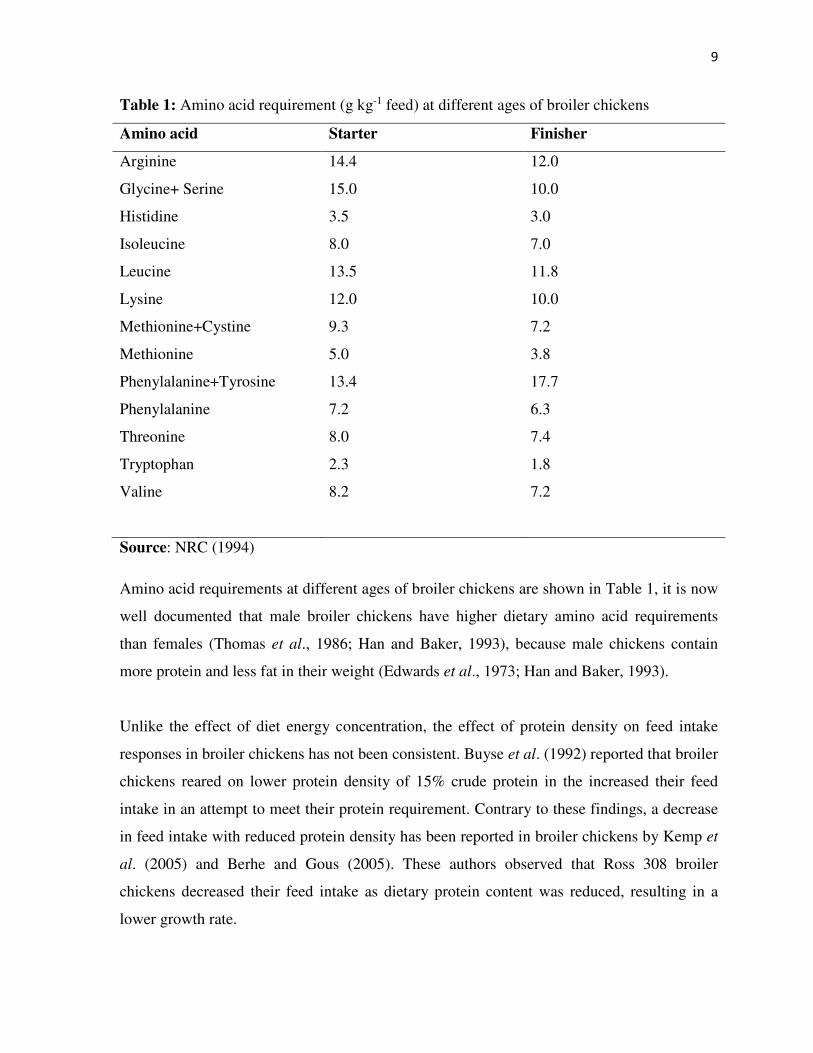
9
Table 1: Amino acid requirement (g kg-1 feed) at different ages of broiler chickens
Amino acid Starter Finisher
Arginine 14.4 12.0
Glycine+ Serine 15.0 10.0
Histidine 3.5 3.0
Isoleucine 8.0 7.0
Leucine 13.5 11.8
Lysine 12.0 10.0
Methionine+Cystine 9.3 7.2
Methionine 5.0 3.8
Phenylalanine+Tyrosine 13.4 17.7
Phenylalanine 7.2 6.3
Threonine 8.0 7.4
Tryptophan 2.3 1.8
Valine 8.2 7.2
Source: NRC (1994)
Amino acid requirements at different ages of broiler chickens are shown in Table 1, it is now
well documented that male broiler chickens have higher dietary amino acid requirements
than females (Thomas et al., 1986; Han and Baker, 1993), because male chickens contain
more protein and less fat in their weight (Edwards et al., 1973; Han and Baker, 1993).
Unlike the effect of diet energy concentration, the effect of protein density on feed intake
responses in broiler chickens has not been consistent. Buyse et al. (1992) reported that broiler
chickens reared on lower protein density of 15% crude protein in the increased their feed
intake in an attempt to meet their protein requirement. Contrary to these findings, a decrease
in feed intake with reduced protein density has been reported in broiler chickens by Kemp et
al. (2005) and Berhe and Gous (2005). These authors observed that Ross 308 broiler
chickens decreased their feed intake as dietary protein content was reduced, resulting in a
lower growth rate.

10
2.3.3 Water
Water is one of the most important nutrients needed by broilers. Water constitutes more than
half of the weight of poultry meat and about two-third of the weight of egg (Ranjhan, 1981).
The newborn animal contains 750 to 800g of water per kg weight, though this falls to about
500g/kg in the matured fat animal (McDonald et al., 1995). Oluyemi and Roberts (2007)
reported that drinking water for poultry should be free from salt and toxic substances. He also
reported that the ratio of dry matter intake by poultry in a temperate environment is normally
1:2, but does depend to some extent on the diet. Thus, increase in protein level also increases
the water consumption of the birds.
Water is available in the feed (metabolisable water) but the bulk of water must be provided
separately in drinking cans or troughs. The water must be provided ad-libitum, clean and free
from excessive salts, which might have a laxative effect and must be cool. Olomu (1995)
suggested that 2 to 2.5 liters of water in the first two weeks is adequate for 100 chicks, this
doubled after another 2 weeks. In practice, the daily water consumption per 1000 broilers
ranges from 20 liters at one week of age to 380 liters at 8weeks of age. On very hot days, the
water consumption may be increased as much as three times of the water intake on cooler
days.
2.3.4 Minerals
Minerals are mainly inorganic components of feed. The body of animals contains large
number of mineral elements, which occur in combination with the organic constituents.
Chickens body contains about 4% mineral matter (Ranjhan, 1981). Phosphorus and calcium
are the two most abundant mineral elements in the bone and other skeletal components of the
birds.
Olomu (1995) classified mineral elements as macro and micro depending on the quantitative
requirement for each. Macro elements include; calcium, phosphorus, sodium, chlorine,
sulphur, magnesium and potassium while some of the micro-elements are iodine, zinc,
cobalt, iron, copper and molybdenum. Excess or deficiency of these mineral elements is
detrimental to the health and development of the birds. Minerals function in many ways such

11
as in body structural components and acid-base balance. Calcium and phosphorus have been
found to have closely related metabolic functions. The requirement of these minerals appears
to be higher for warm climates than for cold climates (Olomu, 1995). It is therefore
recommended that the level of calcium be increased from 1-1.2% to 4-4.5% and the level of
total phosphorus from 0.7-0.8% to 1-1.1% during periods of hot weather. Ranjhan (1981)
stated that mineral deficiency and over supply have serious adverse effects and could lead to
death of chicks. Thus, in practice 0.30-0.35% common salt or sodium chloride would take
care of the requirements for sodium and chlorine which must be supplied from sources other
than the normal ingredients used in formulating poultry diets.
2.3.5 Vitamins
Vitamins are organic compounds that cannot be synthesized in the poultry body and they are
required in extremely small quantities but absolutely essential for normal growth and health.
Oluyemi and Robert (2007) listed the vitamins required in poultry diets as follows: the fat
soluble- vitamins A, D, E and K and the water soluble vitamins- thiamine (B1), niacin,
riboflavin (B2), pyridoxine (B6), biotin, pantothenic acid, folic acid, choline, and vitamin B12.
Vitamin C can be synthesized in the body of birds but they are required as antistress in hot
environment.
The deficiency of these vitamins in the feed consumed by birds or non-availability of these
vitamins to the birds usually result in disease symptoms, such as rickets which is caused by
vitamin D deficiency associated with low or imbalance of calcium and or phosphrous level in
the diet; curled toe paralysis (Vitamin B12) and perosis associated with deficiency of choline
and biotin (Olomu, 1995). Deficiency of folic acid cause poor feathering, poor feather
pigmentation and anemia (Obioha, 1992).
2.4 Lipids in poultry Nutrition
Lipids are substances soluble in non-polar organic solvents (chloroform and ether) and
insoluble in water, which can be divided into four categories: simple, compound, derived and
terpenoids.
Simple lipids are esters of fatty acids and certain alcohols, particularly glycerol and
cholesterol. These can be divided into three classes: triglycerides, steroids, and waxes.

12
Triglycerides are constituted by glycerol esterified with three fatty acids and represent more
than 90% of the foods used in animal feeding. Steroids are lipids that cannot be saponified,
which means that heating in the presence of alkalis does not hydrolyze them; therefore soap
is not produced from their fatty acids. Sterols are the most abundant steroids, and cholesterol
is the main sterol of animal tissues (Ferreira, 1999). Waxes are esters of fatty acids with long-
chain alcohols and constitute the natural protecting coverage of leaves, stems, insects, skin,
feathers, hairs and also the structural material of beehives. They have no nutritional value,
since they are hydrophobic and cannot be degraded by the digestive enzymes of superior
animals (Ferreira, 1999). Compound lipids are esters of glycerol that contain two fatty acids
plus another chemical group such as choline or serine. The most important groups are the
phospholipids, which contain phosphate in their structures, whereas glycolipids have one
carbohydrate replacing a fatty acid, and lipoproteins, which constitute the principal means by
which lipids are transported in the blood (Leeson and Summers, 2001). Derived lipids
include substances with fatty acid, glycerol and with other alcohols that derive from simple
and compound lipids after hydrolysis (Ferreira, 1999). Terpenoids are characterized by a
repeating unity, which is the isopropene. This group includes the carotenoids, xantophylls,
tocopherols and vitamins A and K (Ferreira, 1999).
According to Fats in animal feed (1985), the so-called free fatty acids are not linked to
another organic component as the glycerol. The free fatty acids comprise a small fraction of
total lipids in natural foods. The physical and chemical properties of lipids are determined by
the composition of their fatty acids, and the length and saturation degree of the carbonic
chain.
The term saturated means the absence of double bonds, whereas unsaturated indicates the
presence of one or more double bonds. The identification and nomenclature of the main fatty
acids are represented on.
Birds are not able to synthesize all fatty acids and thus, some are considered essential fatty
acids. Linoleic (18:2, n-6) and linolenic (18:3, n-3) fatty acids are recognized as
metabolically essential. However, the linoleic acid is the only essential fatty acid whose diet
requirement has been demonstrated (National Research Council, 1994).

13
The increase in the length of the carbonic chain of saturated fatty acids increases the melting
point of the fat and the presence of the double bond decreases the melting point. The longer
is the chain, the smaller is the number of double bonds, and less soluble it will be in water.
The geometry of the double bond also influences the melting point. Trans fatty acids have
higher melting point than their cis isomers. The importance of knowing the fatty acid
composition of each lipidic source is as fundamental as knowing the carbonic chain length,
its saturation degree, and the position of the double bond. All of these factors influence the
digestion process (Fats in animal feeds, 1985).
2.4.1. Lipid Digestion
The digestion and absorption of fat in the chicken occurs mainly in the small intestine (Scott,
et al., 1982). The presence of digesta, with intact dietary fat in the duodenum, stimulates
cholecystokinin secretion, which in turn induces the secretion of pancreatic enzymes and bile
(Friedman and Nylund, 1980). While bile salts emulsify fat along with co-lipase providing
more surface area for the enzymes to act upon, lipase hydrolyses the emulsified triglycerides
on sn-1 and sn-3 positions to release monoglycerides and free fatty acids (Mu and Høy,
2004). Cholesterol esterase hydrolyses cholesterol-fatty acid esters into cholesterol and free
fatty acids (Mu and Høy, 2004). Also large amounts of endogenous phospholipids enter
through bile, predominantly phosphatidylcholine in addition to dietary phospholipids.
Phospholipase A2 cleaves phospholipids at the sn-2 position to release lysolecithins and free
fatty acids (Scott, et al., 1982).
2.4.2. Lipid Absorption
The main factor that affects the metabolizable energy value of oils and fats is their
digestibility, which is dependent on the following factors: the length of the carbonic chain,
the number of double bonds, the presence or absence of ester bonds (triglyceride or free fatty
acid), the specific arrangement of the saturated and unsaturated fatty acids on the glycerol
backbone, the composition of the free fatty acid, the composition of the diet, the type and
quantity of triglycerides supplemented in the diet, the intestinal flora, the sex and age of the
birds (Renner & Hill, 1961a; Leeson and Summers, 2001, Nascif et al., 2004).

14
The lipids arrive intact in the duodenum. The presence of food in the duodenum stimulates
the secretion of cholecystokinin, which induces the contraction of the gall bladder and the
secretion of the pancreatic juice, making the chyme. Colipase binds to the oil-water interface
and produces the emulsion. Afterwards, triglycerides undergo hydrolysis under the action of
the pancreatic lipase, and micelles are formed by monoglycerides, diglycerides and free fatty
acids. Micelles are able to solubilize high amounts of fatty acids of low polarity and fat-
soluble vitamins.
Monoglycerides and long-chain unsaturated fatty acids when linked to the conjugated bile
salts promptly form the micelles, whereas saturated fatty acids have less ability to form
micelles due to their characteristic low polarity. Since long-chain unsaturated fatty acids have
greater ability to form micelles, they may act synergistically in the absorption of saturated
fatty acids when mixed with them (Ferreira, 1999). Once inside the enterocytes,
monoglycerides and free fatty acids are re-esterified and combined with free and esterified
cholesterol, lipoprotein and phospholipids to form the so-called lipoproteins or chylomicrons
or still portomicrons. Thus, re-esterified triglycerides are taken from the digestive system to
the circulatory system of the bird in this form (Leeson & Summers, 2001).
As the lymphatic system of the birds is not well developed, the absorption route of the lipids
is through the portal venous system. Very Low density lipoproteins (VLDL) are
quantitatively more important in birds, particularly during the laying period, because they are
the proteins that carry fat from the liver to the extra hepatic tissues, as for example, to the
ovary, where they will be used for egg yolk synthesis. Triglycerides that are not utilized in
the liver or not incorporated into the egg yolk are utilized in other tissues (heart, muscle) or
are stored in the adipose tissue (Escribano, 1991). During absorption and transportation
processes, no alterations occur in the composition of fatty acids. Therefore, there is a great
similarity between the dietary fat fed to birds and the body fat that is deposited. Triglycerides
can be synthesized from the glucose resulting from the digestion of the carbohydrates. The
composition of the fat synthesized through this route is characterized by an elevated content
of palmitic acid, estearic acid, and oleic acid, which constitute 90% of the total fatty acids.
The presence of fat in the diet inhibits the synthesis of fat from carbohydrates. The final
composition of body fat is a pondered average among the fat produced endogenously from

15
glucose, the quantity and composition of the fatty acids in the diet and the fat resulting from
catabolism through b-oxidation (Ferreira, 1999, Sanz et al., 2000). Carew et al. (1972)
evaluated the absorption of corn oil and tallow in light male birds and observed that
absorption of both fats was lower in younger birds. The capability to absorb corn oil
increased from 84 to 95% from the first to the second week of age and absorption of tallow
has increased from 40 to 79%. These results indicated that newly hatched chicks do not have
complete physiological ability to absorb fat. Nevertheless, this ability develops quickly after
the first days of life. Broiler breeder males showed lower apparent digestibility of fat in the
first week of life than in the second and third weeks of age (Mahagna et al., 1988).
Despite the existing contradictions in the literature and the common sense between the
nutritionists that the chicks on the first days of age are not able to digest oils, Zelenka et al.
(1997) has used 2.9% and 3.3% of ether extract in the diet in two experiments and
demonstrated that the apparent digestibility of the fat is high during the first week of age, low
on the second and high after the third week of life. Noy and Sklan (1995) studied the
digestion and absorption of fats in young birds (from 1 to 21 days) and have reported that the
true digestibility of the unsaturated fat in four-day-old birds was higher than 85%, increasing
a little on the subsequent days. This demonstrates that the activity of lipases and bile salts on
the fourth day of age were enough for the complete fat digestion. It was concluded that fat
digestibility is not a limiting factor for the growth of young birds. According to Freitas
(1999), broilers show high digestibility of fat on the first week of life and the inclusion of oil
in the initial diet promotes a better performance of the chickens until 21 days of age. During
the first three weeks of age, chicks fed diets with oil have shown higher apparent digestibility
values of ether extract than the ones that received rations without oil (Cançado, 1999).
2.5 Dietary Fats in Nutrition
Fats are usually added to the feed for meat-type poultry to increase overall energy
concentration and, in turn, improve productivity and feed efficiency. Oxidation of fat is an
efficient means to obtain energy for the cell in large quantity, whereas anabolic use involves
direct incorporation into the body as a part of growth. Lipid accrual is most obvious in
adipose tissue; however, cell multiplication also requires an array of lipids to form associated

16
membrane. These two uses can occur simultaneously; however. the extent of each may vary
considerably.
2.5.1 Importance of Dietary Fat to Poultry
Scott et al.,(1982) have concluded that the net amount of energy obtained by chicks is 60% of
the metabolizable energy of proteins, 75% of the metabolizable energy of carbohydrates,, and
90% of the metabolizable energy of fats (Scott, et al., 1982), underscoring the high efficiency
of metabolizable energy contributed by fat. Baião and Lara observed that the inclusion of oil
in the starter diet increased the fat digestibility in broiler chicks during the first week of life
and resulted in better performance through 21 days of age, compared to the chicks received
rations without oil (Baião and Lara, 2005). Furthermore, chicks utilized metabolizable energy
more efficiently for growth if part of the carbohydrate portion of the diet was replaced
isocalorically with corn oil (Carew and Hill, 1964) or acidulated soybean soap stock
(Lipstein et al., 1965). For growing chicks, the net availability of metabolizable energy from
corn oil was about 10% higher than that of carbohydrate-rich feed ingredients; yellow corn
and milo (De Groóte, 1968). Also, chicks fed diets devoid of supplemental fat had higher
levels of lipogenesis and increased adipose fat deposition (Dvorin et al., 1998).
2.5.2 Advantages of Dietary Fat Supplementation
1. Essential Fatty Acids
Birds are not able to synthesize all fatty acids and thus, some are considered metabolically
essential viz. linoleic (18:2, n-6) and linolenic (18:3, n-3) fatty acids. However, to date the
NRC has only defined the dietary requirements for linoleic acid (NRC, 1994). These
essential fatty acids are absorbed from the feed, in which dietary fat is the major source
(Dvorin, et al., 1998). Fatty acids obtained from crude soybean oil and corn oil are very high
in unsaturated fatty acids, with high levels of linoleic acid (Scragg et al., 1987). Any
deficiency of linoleic acid adversely affects egg production, and it has been proven that
supplementation of linoleic acid, present in vegetable oils, increases egg weight (Shutze and
Jensen, 1963; Balnave, 1971). Depressed growth (especially in male chickens) may be the
first sign of an inadequate supply of essential fatty acids (Wiseman and Salvador 1991). Zero
hatchability represents the most acute deficiency of linoleic acid that has been reported to

17
date. It was manifested by increased susceptibility to respiratory infections, poor egg
production, small egg size, and low fertility (Miller, et al., 1963; Menge, et al., 1965).
Furthermore, the conjugated isomers of linoleic acid have been reported to be effective in
minimizing reduced growth rates by preventing the catabolic effects of immune stimulation
(Cook et al., 1993).
2. Micronutrients
Dietary fats play a significant role in the absorption of fat-soluble vitamins (A, D, E and K)
by acting as their "carriers" - apart from serving as major sources of these vitamins (Iqbal and
Hussain, 2009). The fat-soluble vitamins are incorporated into portomicrons for
transportation after absorption, and are also deposited in body lipid stores (Drevon, 1991).
Hence impairment of the absorption of fat-soluble vitamins is the most serious consequence
of a dietary deficiency of fat (Jacob et al., 2011).
Vitamin A: Acid oil and corn oil are rich vegetable sources of carotenoids which are
precursors for the synthesis of vitamin A. Vitamin A plays a significant role in regulating cell
growth, specifically epithelial cells. It is also necessary for bone metabolism through
involvement with osteoblasts (bone forming cells) and osteoclasts (cells involved in bone
resorption) (Surai et al., 2001). Vitamin A influences both antibody production and T-
lymphocyte proliferation responses, thus a deficiency can result in decreased immune
responses (Friedman and Sklan, 1989a, b). Moreover it has been observed that for maximal
immune response in growing chickens, it may be beneficial to include dietary vitamin A at
higher levels than NRC (NRC, 1994) recommendations (Sklan et al., 1994) and dietary fats
would be an ideal option in doing so.
Vitamin D: Vitamin D is required for normal calcium absorption and utilization, hence
inadequate levels of vitamin D induces calcium deficiency resulting in decreased egg
production (Fritts and Waldroup, 2003; Jacob et al., 2011). Vitamin D also plays a vital role
in the immune responses of chickens, particularly cellular immune responses (Aslam et al.,
1998). Moreover vitamin D prevents tibial dyschondroplasia by increasing bone health
substantially (Edwards Jr, 1990). Recently, it has also been reported that vitamins A and D

18
regulate bile acid synthesis, thus controlling fat absorption as well as their own absorption
(Schmid, et al., 2010).
Vitamin E: Lipid and myoglobin oxidation are major causes of meat quality deterioration
during storage (Jensen et al., 1998) and poultry meat has higher sensitivity to oxidation
damages among all meats, which may be ranked according to oxidation susceptibility in the
following order: turkey > chicken > pork > beef > lamb (Tichivangana and Morrissey, 1985).
It has also been successfully demonstrated that dietary vitamin E supplementation
significantly increased the α-tocopherol content of muscle membranes in many animals
(Lauridsen et al., 1997). α-tocopherol functions as a lipid antioxidant and free radical
scavenger (Hsu and Guo, 2002). Moreover vitamin A and E also have a profound effect on
alleviating heat stress in chickens (Sahin et al., 2001). The cost factor of dietary vitamin E
supplementation demands efficient utilization of dietary vitamin E as well as lipid
metabolism (Zouari et al., 2010).
3. Lecithin
Lecithin (phosphatidylcholine) is a phospholipid that is extracted commercially from
soybeans. It promotes the incorporation of fatty acids into micelles by acting as an emulsifier,
facilitating fat absorption (Cho et al., 2008). Dietary lecithin plays an important role in the
transport of triglycerides out of intestinal mucosa by providing surfactant lecithin for the PM
envelope and supporting mucosal protein biosynthesis (O'Doherty et al., 1973). Lecithin also
alters the serum hormone levels, affects hepatic gene expression, and thereby regulates fat
metabolism of broilers (Huang et al., 2008). Dietary supplementation of lecithin increased
the metabolizable energy of added fat, but parallel to lipid availability (Sibbald et al., 2000).
Lecithin has also been observed to improve the tenderness of meat (Collins et al., 2011).
When supplemented at 6%, lecithin significantly increased yolk percentage, improved haugh
unit score (albumen density) and yolk color, while increasing fat digestibility and plasma
total lipids (Attia et al., 2009).

19
4. Milling Benefits
Added fat in the diet acts as a binder, improving pellet quality and providing optimal
durability (Wilson and Bayer, 2000). It has also been observed for many years that dietary fat
reduces dusting and wastage of feed during the milling process (Wilson and Bayer, 2000).
5. Diet-Induced Thermogenesis
The reduced heat increment of fat-containing diets, which appears to be one of the primary
causes of the extra-caloric effect, has been explained (Edwards, 1969) as follows: the
conversion of carbohydrates and protein to fatty acids is a heat-producing process, while the
presence of an adequate level of dietary fat prevents this process. A high proportion of the
dietary fatty acids can be used directly in the synthesis of tissue triglycerides, while fatty
acids formed from carbohydrates must pass through a series of heat-producing metabolic
transformations. The heat increment lost to the environment constitutes a loss of expensive
energy utilized to produce fatty acids. Moreover, this additional heat produced during the
fatty acid synthesis from dietary carbohydrate may also accentuate the effects of heat stress,
under hot weather conditions. In high-temperature environments adequate amounts of dietary
fat should be present in order to inhibit the conversion of dietary carbohydrates to fatty acids,
a heat-producing process (Edwards, 1969). The beneficial effects of vegetable oil on the
efficiency of energy utilization was due to a reduced heat increment rather than to a reduction
in the basal component of the heat production (Shannon and Brown, 1969).
2.5.3 Fat Digestibility in Young Chicks
It has been proven that fat digestibility is not a limiting factor in the growth of young broilers
(Noy and Sklan, 1996). These authors reported that the true digestibility of the unsaturated
fatty acids in four-day old birds was over 85%, which demonstrates that the activity of
lipases and bile salts by the fourth day of age were sufficient for near complete fat digestion.
It has also been shown that fats of vegetable origin are more digestible in young birds. For
example, the capability of broiler chicks to absorb vegetable oil increased from 84% during
the first week of age to 95% during the second, while the absorption of tallow increased from
40 to 79% over the same time period (Carew Jr et al., 1972). The metabolizable energy of a
corn-soy diet supplemented with soybean oil was maximized as early as two weeks of age in

20
broilers (Batal and Parsons, 2002). Day-old broiler chicks had increased apparent fat
digestibility when fed with soy oil at a dietary inclusion level of 3.5% (Zollitsch et al., 1997).
2.5.4 Efficient Utilization of Dietary Fat
The main factor affecting the metabolizable energy value of fats is their digestibility, which
is dependent on the length of carbon chain and the degree of saturation of the constituent
fatty acids (Renner and Hill, 1961a; Leeson and Summers, 1976; Waldroup et al., 1995;
Leeson and Summers, 1997). Interactions between the degrees of saturation, relative to fat
absorption have been suggested by several authors (Young and Garrett, 1963; Leeson and
Summers, 1976). The poultry grease and palm oil samples consistently had the lowest
metabolizable energy values, while blends apparently based on soybean oil consistently had
the highest metabolizable energy values (Pesti et al., 2002). Renner and Hill (1961b) asserted
that fats which had previously been used for deep frying such as restaurant grease have
substantially lower and more variable metabolizable energy values. This decrease depends to
a great extent on the duration of heating, temperature used, and fatty acid profile. Heating
increases the loss of poly-unsaturated fatty acids, as they are much more sensitive to
oxidative degradation than saturated fatty acids, and it changes the fatty acid profile (Renner
and Hill, 1961a). Metabolizable energy values may also be reduced by the presence of
damaged fats including, for example, dimeric or polymerized fatty acids. This same pattern is
applicable to animal fats such as tallow, choice white grease and poultry fat, which typically
have very low poly-unsaturated fatty acid levels with high saturated fatty acid content. These
"animal" renderers' fats have wide variation not only in metabolizable energy value, but also
in chemical characteristics. Consequently, the metabolizable energy of these saturated fats
may be increased if the basal diet contains fats high in poly-unsaturated fatty acids (Lewis
and Payne, 1966; Ketels et al., 1987; Wiseman and Salvador, 1991). It has been proven that
the best way to use fats with more saturated fatty acids, viz. stearic and palmitic acids, such
as animal and palm fats, is by blending them with unsaturated fatty acids (Renner and Hill,
1961a, b). Oleic acid (C18:1) plays a more direct role in the absorption process, either in the
lumen or in the mucosa cells, to facilitate the increased absorption of saturated fatty acids
(Young and Garrett, 1963). These researchers have usually found a synergism between fatty

21
acids that improve fat digestibility, and the resulting impact on its metabolizable energy
content, particularly at high levels of dietary inclusion.
2.6 Sources of Dietary Fat in poultry Nutrition
Cotton oil
The utilization of cotton oil is limited by the presence of the natural yellow pigment
denominated gossypol, which is a toxic or anti-nutritional element. It causes iron deficiency
and lysine unavailability due to Maillard reactions, thus reducing the nutritional value of
proteins (when utilized in the meal form). Ferrous sulphate must be added to broiler diets in
which cotton oil is included, because it chelates gossypol, preventing its absorption in the
digestive tract and thus neutralizes its deleterious effects. Broilers tolerate levels higher than
100 ppm of free gossypol without performance impairment. It is not recommended to use
gossypol in laying hen diets. The presence of cyclopropenoic fatty acids (malvalic and
sterculic) intensifies the effects of gossypol, resulting in green yolk and pink coloration of the
albumen. When high amounts of these fatty acids are ingested, the yolk develops redness and
viscous appearance after short time of refrigeration. This alteration is caused by the increase
in the concentration of saturated fatty acids in the yolk as a function of the inhibition of the
enzymes 6 e 5 desaturase, which hinders the denaturation of stearic and palmitic acids
and their corresponding monounsaturated fatty acids. Besides the ability of changing the
composition of yolk fatty acids, the cyclopropenoic fatty acids alter the permeability of the
vitelline membrane and cause iron diffusion from the yolk to the albumen, where
ovotransferrin acts as iron chelator. This reaction between ovotransferrin and iron is
responsible for the pink coloration of the albumen. On the other hand, some ovotransferrin
may diffuse to the yolk, combine with iron and also cause red yolk coloration. The pink
albumen coloration is seen more often in eggs stored for long periods of time. In breeder
flocks, besides yolk discoloration, high levels of gossypol and the diffusion of ovotransferrin
can reduce egg hatchability (Leeson & Summers, 2001).

22
Canola oil or Rapeseed oil
Canola is the name given to rapeseed that contains less than 2% of erucic acid (docosenoic
acid, C22:1, -9) in relation to the total fatty acid and less than 30 umoles of glucosinolates
per gram of free oil on seed dry matter basis (Leeson & Summers, 2001).
In birds, the adverse effects of adding erucic acid to the diets are reflected on intake, growth
and the apparent digestibilities of total lipid and individual fatty acids ( Leeson & Summers,
2001). Furthermore, chicks fed with diets containing erucic acid deposit less fat and utilize
energy from this lipid less frequently ( Leeson & Summers, 2001).
Female broilers fed diets containing two different forms of canola oil showed better growth
rate when compared to females fed diets containing tallow and acidulated soybean oil
soapstock. This observation confirms the advantage of using vegetal oils instead of tallow
and acidulated soybean oil soapstock as energy sources for birds. The better growth rates are
a result from the higher percentage of long chain fatty acids and higher contents of
triglycerides (Thacker et al., 1994). Andreotti et al. (2001) observed similar performance,
carcass yield and cut yields of broilers with 49 days old fed poultry fat, canola, sunflower,
corn, soybean or lard. To evaluate the effects of different lipidic sources on the physical and
chemical characteristics of thigh meat, broilers were fed diets containing soybean oil,
sunflower oil, canola oil, corn oil, poultry fat or lard. Lard and corn oil resulted in more red-
colored meat when compared to the meat of birds fed with oil of canola, sunflower or
soybean, but did not differ from the poultry fat (Souza et al., 2001).
Sunflower oil
Alao and Balnave (1984) reported better development and feed conversion of birds fed with
diets containing sunflower oil when compared to birds fed olive oil. It was suggested that the
difference was a function of the fatty acid composition of the different vegetable oils. Sanz et
al. (1999) fed broilers with sunflower oil or a mixture of beef tallow/lard and the utilization
of saturated fats resulted in greater abdominal fat deposits than unsaturated fats. According to
Sanz et al. (2000a), the utilization of a source of unsaturated lipids reduces fat and increases
protein on the broiler carcasses. The difference in protein accretion was attributed to the level

23
of saturation of the fat, since the energy derived from unsaturated fat may be used for other
metabolic purposes, whereas the energy derived from saturated sources is less promptly
utilized and accumulates as body fat. In general, corporal fat accumulation may be
considered the result of the balance between the fat absorbed from the diet, the endogenous
synthesis of fat (lipogenesis) and the catabolism of fat by -oxidation (lipolysis).Therefore,
in cases in which the quantity of absorbed fat is the same, lower fat depositions may be
attributed to an increase in catabolism or endogenous decrease in the synthesis of fatty acids,
or both.
Sanz et al. (2000b) evaluated inclusion levels of 8% of two lipid sources, one saturated (beef
tallow) and one unsaturated (sunflower oil). There was a significant reduction in the
abdominal fat of birds fed diets with sunflower oil (Sanz et al.,2000b). Provided that the
quantity of absorbed fat was higher in birds fed diets containing sunflower oil, the reduction
in the fat deposit of these birds seemed to be a function of higher fat oxidation rates
(catabolism) and lower synthesis of fatty acids. Using 8% of inclusion of three lipidic sources
(sunflower oil, fish oil and beef tallow) in broiler feeding, Newman et al. (2002) observed a
higher proportion of linoleic acid and higher polyunsaturated/saturated ratio in the
composition of the fat of birds fed with sunflower oil.
Linseed oil
The effects of soybean, canola, sunflower and linseed oils on the nutritive and organoleptic
traits of the meat and the profile of fatty acids were evaluated in 5-week-old broilers (López
Ferrer et al., 1999). The parameters of meat quality were not different between treatments,
even though meat juiciness of the samples from birds fed with canola oil has been inferior to
the other treatments. The values of TBARS were higher in fresh abdominal fat of birds fed
linseed oil and soybean oil and in the breast muscle stored for 12 months when compared to
the samples of fresh breast. Higher levels of omega-3 in both tissues were observed in birds
fed linseed oil.

24
Rosa (1999) used three types of oils (linseed, soybean and a mixture of linseed and fish) at
inclusion levels of 1, 2 and 3% in broiler diets and observed that the composition of fatty
acids of the rations influenced the fatty acid profile of the breast and thigh.
It has been suggested that the size of fat deposits may be changed according to the fatty acid
profile of the diet. The use of beef tallow, olive oil, sunflower oil and linseed oil in broiler
diets evidenced that fat sources rich in polyunsaturated fatty acids produce less fat deposits
than the ones rich in saturated or monounsaturated fatty acids (Crespo and Esteve-Garcia
(2002).
Palm oil
According to Rodriguez et al. (2002), palm oil or mixtures of palm oil, fatty acids distilled
from the palm and calcic soap are sources of vegetal oils with a fatty acid profile that might
replace animal fats without any kind of negative impact on carcass quality.
Soybean oil
The raw soybean oil has on its composition several substances considered as impurities that
must be removed using several processes (filtration, hydration and degum). These substances
are solid residues from the extraction process, and they are phospholipids, gums, metallic
complexes, free fatty acids, peroxides, polymers, secondary products from oxidation and
pigments (Beauregard et al., 1996).
Degummed Soybean oil
The degummed soybean oil is a product obtained from the raw oil after degumming, which
consists of centrifugation the soybean oil to separate it into the two following parts:
degummed oil and unsaponifiable matters (Baiao and Lara 2005).
Scaife et al. (1994) fed female broilers with rations containing different sources of lipids
(beef tallow, soybean oil, canola oil, marine fish oil or a mixture of these oils) and observed
that live weight was significantly higher when soybean oil was used. Birds fed with canola
oil also showed higher intake and higher weight. Birds fed beef tallow had the poorest
conversion rate.

25
Vieira et al. (2002) evaluated broiler rations containing 0, 4 and 8% of soybean oil and
acidulated soybean oil soapstock, and observed similar weight gain between the different
lipidic sources and improved feed conversion in birds fed with soybean oil in comparison to
the birds fed with acidulated soybean oil soapstock. They also observed a reduction in feed
intake of birds fed with acidulated soybean oil soapstock when the inclusion level was
increased from 4 to 8%, whereas no reduction in feed intake was observed in birds fed with
soybean oil.
Lara et al. (2004) fed male broilers with different lipidic sources (degummed soybean oil,
poultry fat, acidulated soybean oil soapstock, a mixture of poultry fat/soybean oil and other
mixture of soybean oil/ acidulated soybean oil soapstock). It was observed better weight gain
and intake in birds fed with soybean oil in relation to the birds fed with acidulated soybean
oil soapstock. The different lipidic sources had no influence on the levels of moisture, ether
extract, and protein of the breast, thigh and whole carcasses. According to Moura (2003), the
inclusion of soybean oil in broiler diets does not affect the moisture and ether extract in the
breast and thigh muscles. Furthermore, the deposition of fat on the breast muscle and viscera
is not affected by the inclusion of the oil in the diet.
Acidulated soybean oil soapstock
The acidulated soybean oil soapstock, also denominated as soybean fatty acid, is a sub-
product of the industry of soybean oil. This sub-product is obtained through the alkaline
neutralization of the raw oil, which produces a raw soap (a mixture of soaps, neutral oil,
water, sterols, pigments, and other constituents). This unstable product is converted in
acidulated soybean oil soapstock after a treatment of sulfuric acid in hot aqueous solution.
Compared to soybean oil, acidulated soybean oil soapstock contains high levels of free fatty
acids (50%), unsaponifiable matter, and oxidized fatty acids, besides being also rich in
carotenoids (Bornstein and Lipstein, 1963; Lipstein et al., 1965; Pardio et al.,2001). Free
fatty acids affect absorption negatively and, consequently, also the nutritive value of the fats.
The ratio between free fatty acids and intact triglycerides is important, since supplements
with free fatty acids are absorbed less efficiently than the free fatty acids in form of
triglycerides. The reason for this is that monoglycerides are essential to incorporate insoluble

26
fatty acids in the micellar complex. There are not enough monoglycerides to combine with
all free fatty acids when they are offered as the only fat source and absorption is then
impaired (Fats in animal feeds, 1985; Blanch et al., 1995).
Wiseman and Salvador (1991) worked with diets for broilers supplemented with 3 different
sources of lipids (tallow, soybean oil and palm oil) and their respective acid oils, with
different levels of free fatty acids. The reduction in metabolizable energy values of the fats
with higher contents of free fatty acids tend to be more pronounced with greater inclusion
levels of lipids.
Besides, reduction is also greater with greater saturation levels. The increase in free fatty acid
levels reduces progressively the ME values of the energy source, and such effect is more
pronounced in young birds.
The use of refined soybean oil, raw soybean oil or acidulated soybean oil soapstock showed
an increase on the levels of linoleic fatty acid on the carcass of birds fed with refined soybean
oil and raw soybean oil in relation to the acidulated soybean oil (Cascabulho, 2000).
The value of nitrogen-corrected metabolizable energy of acidulated soybean oil soapstock
(8.144 kcal/kg DM) determined for 28-day-old broilers was 5% inferior than the value of raw
soybean oil. The inclusion of acidulated soybean oil soapstock in rations for animals depends
on the maintenance of a minimum of quality control, so that the product has surely not been
adulterated by the inclusion of other low-quality oils (Vieira et al., 2002).
The values of nitrogen-corrected metabolizable energy of the acidulated soybean oil
soapstock suggested for young and adult birds were 7.788 and 8.610 cal/kg DM, respectively
(Freitas et al., 2002).
Palm kernel oil
Palm kernel oil, similarly to coconut oil, is high in saturated fats and is more saturated than
palm oil. Palm kernel oil is high in lauric acid which has been shown to raise blood
cholesterol levels, both as LDL-C (cholesterol contained in low-density lipoprotein) and
HDL-C (cholesterol contained in high-density lipoprotein) Cascabulho (2000). Palm kernel
oil does not contain cholesterol or trans fatty acids (Rodriguez et al., 2002).

27
Animal fats
Deaton et al. (1981) used diets with similar nutritive values added with 4, 7 and 10% of
animal fat, and observed that the increasing fat level of the diet increased the quantity of
abdominal fat, corroborating results reported by Yalçin et al. (1998).
Poultry fat
Poultry fat is also known as viscera oil and is obtained after the extraction of fat by
autoclaving or in a percolator tank and expeller. After extraction, the fat is placed into a
decantation tank to extract the acidulated soapstock and moisture excess. At this point, it is
ready to be used in ration or to be refined. Product yield varies from 1.3 to 1.6% of the live
weight of the bird (Mano et al., 1999). This range depends on the level and source of energy
used in the ration, besides bird sex, age and weight at slaughter. Higher percentages of fat are
obtained when higher levels of energy are used, older birds are slaughtered and consequently
with higher live weight; moreover, females produce more fat than males, independent of
dietary energy level and age at slaughter. Assessing the effects of the mixtures of vegetal and
animal fats, corn oil and poultry fat on the proportions of 0, 3, 6 and 9% in the alimentation
of broilers,
Griffiths et al. (1977) observed that the birds fed with corn oil and poultry fat were
significantly heavier than birds non-supplemented with fat. Weight gain, feed intake and feed
conversion of broilers fed with diets containing 4% of poultry fat, 4% of soybean oil or a
mixture of 2% of soybean oil and 2% of poultry fat were not affected by oil sources (Dutra Jr
et al., 1991). However, there was a decrease in feed intake and weight when the mixture was
used.
The effects of increasing inclusion levels (0, 2, 4, 6, 8 and 10%) of refined palm oil (8,000
kcal/kg ME), palm oil (8,500 kcal/kg ME), corn oil (9,000 kcal/kg ME) and poultry fat
(9,000 kcal/kg ME) in diets for broiler were studied by Valencia et al. (1993). There were no
effects of the sources of oil on weight gain and feed conversion. On the other hand,
performance was proportionally better with increasing inclusion levels of the oil sources.

28
Performance parameters were evaluated in broilers fed different lipidic sources (raw soybean
oil, poultry fat and acidulated soybean soapctock and its mixtures). There were no significant
differences in weight gain, feed intake, feed conversion and viability when soybean oil and
poultry fat were used (Lara et al., 2003).
The effects of the oxidized poultry fat on the values of metabolizable energy for broilers
from 31 to 34 days old was evaluated by Racanicci et al. (2000). Oxidized poultry fat is
produced by heating from 80 to 90oC, with constant aeration for 40 days. The birds fed with
rations containing 10% of fresh poultry fat or oxidized poultry fat showed apparent
metabolizable energy and apparent adjusted energy for nitrogen values of 9,240, 9,150, 7,700
and 7,595kcal/kg, respectively. According to Edwards et al. (1973), the inclusion of poultry
fat in broiler diets resulted in an increase in the deposition of oleic acid and a reduction in the
levels of the linoleic acid in the adipose tissue of the birds. The use of cotton oil or acid
cotton oil increased the deposition of linoleic acid. Similar results were obtained by Lara et
al. (2003), who observed a higher deposition of monounsaturated fatty acids on the carcass of
broilers fed with poultry fat when compared to birds fed with raw soybean oil and acidulated
soybean oil soapstock.
Beef tallow
Newman et al. (2002) fed broilers with 8% of sunflower oil, fish oil or beef tallow in the diet
and observed poorer feed conversion in the birds fed with beef tallow. There was a positive
relationship between the composition of the fatty acid present in diet and the composition of
the fatty acid of the breast, thigh and skin of broilers fed corn oil, beef tallow or a mixture of
both (Marion and Woodroof, 1963). The type of fat added to the diet has a significant
influence on the profile of fatty acids of the abdominal fat, and birds fed tallow have shown
higher concentrations of saturated fatty acids and lower concentrations of unsaturated fatty
acids comparing to the birds fed with acidulated soybean oil soapstock (Thacker et al., 1994).
Sanz et al. (1999) formulated broiler diets containing sunflower oil and bovine/swine fat and
reported that the inclusion of saturated fats produced higher accumulation of intramuscular
fat and abdominal fat. In a study using one saturated (beef tallow) and one unsaturated
(sunflower oil) lipid source at 8% of inclusion, Sanz et al. (2000b) observed a significant
reduction in the deposit of abdominal fat in the birds that received diets with sunflower oil.

29
According to Crespo & Esteve-Garcia (2002b), the location of fat deposition depends on the
kind of fatty acid added to the diet (saturated and polyunsaturated). Birds fed with diets rich
on animal saturated fatty acids tend to have proportionally larger abdominal and mesenteric
fat than other fat deposits.
Lard
Working with broilers fed diets containing sunflower oil and mixture of bovine tallow/swine
fat, Sanz et al. (2000a) did not observed effects of the lipidic sources on feed intake, weight
gain and feed conversion of the birds. No effects on performance was observed when broilers
were fed from 21 to 49 days with diets containing one of the following lipid sources: poultry
fat, refined soybean oil, refined canola oil, refined sunflower oil, lard and refined corn oil
(Andreotti et al., 2001). Souza et al. (2001) evaluated six different lipid sources in rations for
broilers and observed that the meat of birds fed with lard presented a more pronounced red
coloration in relation to the meat from birds fed with soybean oil, sunflower oil and canola
oil, but it was not different from birds fed poultry fat.
Fish oil
Fish oil is produced by the compression of whole fishes and sub-products for industries of
fishery. This kind of oil contains high percentages of long-chain polyunsaturated fatty acids,
which accounts for the oxidative instability and the transference of characteristic fish flavor
to the meat of animals fed fish oil. In general, fish oils are rich sources of omega- 3 fatty
acids and poor sources of omega-6, and the contents of linoleic acid are also low (<2%).The
fatty acid profile of the different oils varies with the period of the year, processing method
and predominant fish species that was captured (Fedna, 1999). Some authors reported the
unpleasant flavor of fish in the meat of broilers fed with 1.5% to 2.5% of fish oil (Miller &
Robisch, 1969). Phetteplace and Watkins (1990) evaluated different relations of poultry fat
and fish oil (Menhaden) in broiler feeding. Birds fed with diets containing more fish oil have
deposited more quantities of unsaturated fatty acids in the abdominal fat, as well as more n-3
fatty acids. On the other hand, both total n-6 fatty acids and the relation of n-6:n-3 were
higher in the abdominal fat of birds fed with rations containing higher quantities of poultry
fat.

30
Chanmugam et al. (1992) also demonstrated that the content of -3 fatty acids in the thigh of
broilers might be increased by the addition of linseed oil or fish oil in the diet (Menhaden).
Therefore, it has been suggested the inclusion of low dietary levels of fish oil together with
another source of linoleic fatty acid to obtain an acceptable product with increased ratio
between -3:-6 fatty acids. Scaife et al. (1994) have utilized beef tallow, soybean oil, colza
oil, oil of marine fish or mixtures of these oils in female broiler diets. The relation -6:-3
was significantly increased with the inclusion of soybean oil and decreased with the inclusion
of marine fish oil. Rosa (1999) observed that the composition of fatty acids of the rations has
influenced the profile of fatty acids of the breast and thigh using linseed oil, soybean oil and
a mixture of linseed plus fish oil at inclusions levels of 1, 2 an 3% in diets for broilers.
Diet inclusion of 8% of sunflower oil, fish oil or beef tallow resulted in lower deposition of
corporal fat on birds fed with fish oil and sunflower oil. It was concluded that feeding
broilers with sources of n-3 and n-6 produced less carcass fat, and also improved feed
conversion. The results are interesting both from an economical point of view and from the
point of view of the health of consumers (Newman et al., 2002).
2.7 Effects of Dietary Fats on Poultry Performance
A number of different fat sources are available for poultry from both vegetable and animal
sources and from the rendering industry. (Sanz et al., 2000). It was observed that the
replacement of tallow by vegetable fats rich in polyunsaturated fatty acids like sunflower oil,
soybean oil, or linseed oil resulted in a decrease of abdominal fat deposition in broilers
(Newman et al., 2002; Ferrini et al., 2008). In a study, Sanz et al., (1999) fed broilers with
tallow in diet and found that the utilization of saturated fats resulted in greater abdominal fat
deposits compared to birds fed unsupplemented diet. Newman et al. (2002) reported a
significant depression of feed efficiency in broilers fed 8% beef tallow compared to birds fed
sunflower or fish oil. Furthermore, addition of 3% canola oil in broiler diet resulted in a
significant improvement in body weight and feed conversion ratio when compared to birds
fed animal fat (Newman et al., 2002). On the other hand, Shahryar et al. (2011) found that
the addition of 6% fat in broiler lad to an increase of abdominal fat and gizzard weight in
comparison with those of birds fed unsupplemented diets.

31
The improvement in body weight for age of the modern broiler chicken, due to increased
growth rate and associated higher nutrient supply, have lead to frequent occurrence of
metabolic and skeletal disorders (Robinson et al.,1992) or increased carcass fat deposition
(Yu and Robinson, 1992). The two main metabolic disorders affecting broiler chickens are
ascitis disease and sudden death syndrome, which is closely related to the rapid rate and
increased feed intake, particularly overconsumption of dietary energy. These pathological
conditions commonly occur in fast growing broilers chickens, fed ad-libitum.
The findings of Hrdinka et al. (2002) showed that feeding with feed-grade PG resulted in
significantly smaller abdominal fat pads compared to the other fat sources as SO, palm oil
and restaurant grease. Previous reports indicate that broiler chickens fed with diets enriched
with PUFA have less abdominal fat deposition than broiler chickens fed with diets containing
SFA. This is of particular interest in the rearing of female broilers, because the amount of
abdominal fat at slaughter age may represent >3% of the live weight.
Azman et al (2005) noted that final body weight was significantly affected by dietary fat
sources. Daily weight gain and daily feed intake was significantly higher in the beef tallow
group compared to the other groups. The feed conversion ratio of the poultry grease group
was better compared to the other groups. In the soybean oil supplemented group, the amount
of long-chain polyunsaturated fatty acids was significantly higher in thigh skins, breast
muscle and abdominal fat pad. In the thigh and breast muscle of the poultry grease fed group
oleic acid content was significantly higher, and beef tallow caused increased accumulation
of saturated fatty acid in thigh skins and abdominal fat pad.
Crespo and Esteve Garcia (2002) found that chickens fed dietary fat showed higher live
weight when compared to birds fed diet with no supplemental fat.
Oils of plant origin such as soybean oil contain high level of unsaturated fatty acids and are
more completely digested by fowls than animal fats such as tallow. (Leeson and Atteh 2003).
Sanz et al (2000) have reported that broiler chickens fed with diets enriched of
polyunsaturated fatty acids have less abdominal fat or total body fat deposition than do
broilers chickens fed with diets containing saturated fatty acids.

32
2.8 Lipid Profile
A lipid profile is a measurement of various lipids that are found in the blood. This kind of
blood test is often used to assess risk of heart disease (www.wikipedia.com). There are two
common concerns people have about lipids in diet: One is their high caloric value, which
may lead to undesired weight gain. The other is their association with high total cholesterol
levels, which are a risk factor for cardiovascular disease. Limiting the intake of fat and oil in
the diet, especially saturated fats, may help keep cholesterol levels low and thus lower one’s
risk of heart disease (Alfred, 2002).
One reason the USDA recommends that 10% or fewer of one’s calories come from saturated
fats is because the amount of saturated fat in one’s diet correlates strongly with cholesterol
levels. Saturated fats are generally solid at room temperature. Fat from animals (e.g. butter
and lard) is almost always saturated, but some oils from plants are saturated, too (e.g. palm
oil or coconut oil.)
A lipid profile contains information about several different kinds of lipid that normally
circulate in the blood. Values are numerical, but in order to simplify explanation, ranges of
numerical values are often placed into categories such as ‘low risk,’ or ‘high risk.’ For
example, a total cholesterol level over 240 mg/dl is said to be ‘high risk’, but that doesn’t
mean a reading of 238 is fine. With total cholesterol and LDL cholesterol the higher the
number, the higher the risk. Conversely, the lower the LDL cholesterol, the lower the risk.
However, a low number is not a guarantee against heart disease. The population with low
cholesterol is at lower risk of heart disease, but heart disease is not absent in this population.
Triglycerides:
According to (Nomenclature of Lipids), triglyceride (TG, triacylglycerol, TAG, or
triacylglyceride) is an ester derived from glycerol and three fatty acids (tri- + glyceride).
Triglycerides are the main constituent of body fat in humans and animals, as well as
vegetable fat (Nelson and Cox, 2000). They are also present in the blood to enable the
bidirectional transference of adipose fat and blood glucose from the liver, and are a major
component of human skin oils (Alfred, 2002).

33
There are many different types of triglyceride, with the main division being between
saturated and unsaturated types. Saturated fats are "saturated" with hydrogen – all available
places where hydrogen atoms could be bonded to carbon atoms are occupied. These have a
higher melting point and are more likely to be solid at room temperature (Davidson, 2008).
Unsaturated fats have double bonds between some of the carbon atoms, reducing the number
of places where hydrogen atoms can bond to carbon atoms. These have a lower melting point
and are more likely to be liquid at room temperature
This is the most common type of lipid formed in animals. Fat tissue is primarily for the
storage of this form of lipid. Triglyceride levels vary quite a bit over short time periods. A
meal high in sugar, fat, or alcohol can raise the triglyceride level drastically, so the most
repeatable measures of this lipid are taken after 12 hours of fasting. Even though sugar and
alcohol are not lipids, your body will convert any form of excess calories into triglycerides
for long-term storage. A value below 150 mg/dl indicates no increased risk, 150 -200
indicates a slight risk, and over 200 mg/dl is a high risk.
Cholesterol: It is a sterol (or modified steroid), a lipid molecule and is biosynthesized by all
animal cells because it is an essential structural component of all animal (not plant or
bacterial) cell membranes that is required to maintain both membrane structural integrity and
fluidity. Cholesterol enables animal cells to not need a cell wall (like plants and bacteria) to
protect membrane integrity and cell viability, thus are able to change shape and move about
(unlike bacteria and plant cells which are restricted by their cell walls). In addition to its
importance within cells, cholesterol also serves as a precursor for the biosynthesis of steroid
hormones, bile acids, and vitamin D (Westover et al., 2003). Cholesterol is the principal
sterol synthesized by animals. All kinds of cells in animals can produce it. In vertebrates the
hepatic cells typically produce greater amounts than other cells. It is almost completely
absent among prokaryotes (bacteria and archaea), although there are some exceptions such as
Mycoplasma, which require cholesterol for growth (Rizos, 2011). Cholesterol is present in
the blood in three forms. The three defined below are all combinations of protein,
cholesterol, and triglyceride. Cholesterol is a lipid and is insoluble in water. It is transported
through the blood encased in a soluble protein.

34
Cholesterol is a necessary molecule in human metabolism (Gérard et al., 2007). It is a
component of cell membranes, and is a building block of bile, estrogen and testosterone. The
cholesterol necessary for normal metabolism is manufactured by the liver. Generally, a level
less than 200 mg/dl is considered desirable. Between 200 mg/dl and 240 mg/dl is considered
borderline high, and over 240 mg/dl is considered high.
Low-density lipoprotein (LDL)
LDL is one of the five major groups of lipoproteins. These groups, from least dense to most
dense, are chylomicrons, very low-density lipoprotein (VLDL), intermediate-density
lipoprotein (IDL), low-density lipoprotein and high-density lipoprotein (HDL), all of which
are far smaller than human cells. In nutrition, LDL is sometimes referred to as the "bad
cholesterol"; however, this is a misleading misnomer. LDL and cholesterol are completely
different molecules – LDL is about 10 times larger in diameter than cholesterol (~200
angstroms compared with ~20 angstroms).
Lipoproteins transfer fats around the body in the extracellular fluid, can be sampled from
blood and allow fats to be taken up by the cells of the body by receptor-mediated endocytosis
(Dashty et al., 2014). Lipoproteins are complex particles composed of multiple proteins
which transport all fat molecules (lipids) around the body within the water outside cells.
They are typically composed of 80-100 proteins/particle (organized by a single
apolipoprotein B for LDL and the larger particles). A single LDL particle is about 220-275
angstroms in diameter (Segrest et al., 2001), typically transporting 3,000 to 6,000 fat
molecules/particle, varying in size according to the number and mix of fat molecules
contained within. The fats carried include cholesterol, phospholipids, and triglycerides;
amounts of each vary considerably.LDL particles pose a risk for cardiovascular disease when
they invade the endothelium and become oxidized, since the oxidized forms are more easily
retained by the proteoglycans. A complex set of biochemical reactions regulates the
oxidation of LDL particles, chiefly stimulated by presence of necrotic cell debris and free
radicals in the endothelium (http://www.rpi.edu/dept/bcbp/molbiochem.htm). Increasing
concentrations of LDL particles are strongly associated with increasing rates of accumulation
of atherosclerosis within the walls of arteries over time, eventually resulting in sudden plaque

35
ruptures and triggering clots within the artery opening, or a narrowing or closing of the
opening, i.e. cardiovascular disease, stroke, and other vascular disease complications
(http://www.nejm.org).
LDL particles (though far different from cholesterol per se) are sometimes referred to as bad
cholesterol because they can transport their content of fat molecules into artery walls, attract
macrophages, and thus drive atherosclerosis. In contrast, HDL particles (though far different
from cholesterol per se) are often called good cholesterol or healthy cholesterol because they
can remove fat molecules from macrophages in the wall of arteries (Kumar et al., 2011).
This is sometimes referred to as the “bad cholesterol.” This form contains the highest amount
of cholesterol. A value between 130 – 159 mg/dl is borderline high, and over 160 mg/dl is
considered ‘high” (Zhang et al., 2008).
High-density lipoprotein (HDL)
HDL is one of the five major groups of lipoproteins. Lipoproteins are complex particles
composed of multiple proteins which transport all fat molecules (lipids) around the body
within the water outside cells. They are typically composed of 80-100 proteins/particle
(organized by one, two or three ApoA; more as the particles enlarge picking up and carrying
more fat molecules) and transporting up to hundreds of fat molecules/particle. Unlike the
larger lipoprotein particles which deliver fat molecules to cells, HDL particles remove fat
molecules from cells which want to export fat molecules (Sorrentino, 2011). The fats carried
include cholesterol, phospholipids, and triglycerides; amounts of each quite variable.
Increasing concentrations of HDL particles are strongly associated with decreasing
accumulation of atherosclerosis within the walls of arteries (Stephens et al., 2012). This is
important because atherosclerosis eventually results in sudden plaque ruptures,
cardiovascular disease, stroke and other vascular diseases. HDL particles are sometimes
referred to as "good cholesterol" because they can transport fat molecules out of artery walls,
reduce macrophage accumulation, and thus help prevent or even regress atherosclerosis.
However, studies have shown that HDL-lacking mice still have the ability to transport
cholesterol to bile, suggesting that there are alternative mechanisms for cholesterol removal
(Toth, 2005)

36
This is sometimes called “good cholesterol” the higher the number, the better. A value below
40 mg/dl is considered a risk factor. A value above 60 mg/dl is considered protective against
heart disease. HDL cholesterol is cholesterol that is packaged for delivery to the liver, where
the cholesterol is removed from the body.
Very-low-density lipoprotein (VLDL)
This is a type of lipoprotein made by the liver (Gibbons et al., 2004). VLDL is one of the five
major groups of lipoproteins (chylomicrons, VLDL, low-density lipoprotein, intermediate-
density lipoprotein, high-density lipoprotein) that enable fats and cholesterol to move within
the water-based solution of the bloodstream. VLDL is assembled in the liver from
triglycerides, cholesterol, and apolipoproteins. VLDL is converted in the bloodstream to low-
density lipoprotein (LDL). VLDL particles have a diameter of 30-80 nm. VLDL transports
endogenous products, whereas chylomicrons transport exogenous (dietary) products.
Recently both the lipid composition (Dashti et al., 2011) and protein composition (Dashty et
al., 2014) of this lipoprotein were characterised in great detail.
This form contains the highest amount of triglyceride. Like LDL, this is considered “bad
cholesterol.” A value less than 32 mg/dl is desirable. VLDL is usually not measured directly,
but is estimated from the triglyceride count by dividing the triglyceride count by 5. This
mathematical way to estimate VLDL is not valid when the triglyceride is above 400 mg/dl.

37
CHAPTER THREE
MATERIALS AND METHODS
3.1 Location and duration of the Study
The study was carried out at the Poultry Unit of the Department of Animal Science Teaching
and Research Farm, University of Nigeria, Nsukka. Nsukka lies in the Derived Savanna
region, and is located on longitude 60 25□ N and latitude 70 24□ E (Offomata, 1975) and at
an altitude of 430m above the sea level (Breinholt etal., 1981). The climate of the study area
is a typical humid tropical type, with a relative humidity range of 56.01-103.83%. Average
diurnal minimum temperature ranges from 220C-24.70C while the average maximum
temperature ranges from 330C- 370C (0konkwo and Akubuo, 2007, Energy Center, UNN,
2008). Annual rainfall ranges from 1567.05mm to 1846.98mm (Meteorological Center, Crop
Science Department, University of Nigeria Nsukka, 2009 unpublished). The study lasted
(8wks).
3.2. Experimental materials
Palm oil, soybean oil, palm kernel oil and groundnut oil and other feed ingredients were used
for the study.
3.3. Experimental Diets
In the starter and finisher phases of the experiment five diets were formulated. The
percentage compositions of the diets are shown in Tables 1 and 2, respectively.
3.4 Experimental Animals and Management
In the starter phase, one hundred and twenty day-old broiler chicks were used for the study.
The birds were randomly allotted to five treatment groups of 24 birds each, and assigned to
five diets (Table 1). Each treatment group was replicated three times with 8 birds per
replicate. The birds in each replicate were brooded in a deep litter pen measuring 1.50m x
1.50m in the experimental poultry house. The poultry house is an open-sided one; the sides
and demarcations between individual pens are covered with wire gauze. The litter material
was fresh wood shavings. Heat was provided with charcoal pots placed under metal hovers.
Feed and water were provided to the birds ad libitum while additional light was provided at

38
night using kerosene powered lamps to enable the birds eat at night. The chicks were
vaccinated against Newcastle and Gumboro diseases as at and when due.
In the finisher phase, birds from the starter phase were randomly assigned to 5 treatment
groups with 24 birds per group. Each group were replicated three times with eight birds per
replicate placed on deep litter of fresh wood shavings in open sided house partitioned into
pens measuring 1.50m x 1.50m. The birds were randomly assigned to 5 diets (table 2).
Table 2: Percentage composition of broiler starter diets
Ingredients (%)Diets 1 2 3 4 5
Maize 22.5 22.5 22.00 22.5 22.00 Cassava root meal 21.5 20.00 20.00 20.5 19.5 Wheat offal 5.0 5.5 6.3 5.0 6.8 Soybean meal 14.00 14.00 14.00 14.00 14.00 Groundnut cake 24.00 24.00 23.07 24.00 23.07 Palm kernel cake 5.00 5.00 5.00 5.00 5.00 Fish meal 3.00 3.00 3.00 3.00 3.00 Bone meal 4.00 4.00 4.00 4.00 4.00 Salt 0.25 0.25 0.25 0.25 0.25 Vit-min premix* 0.25 0.25 0.25 0.25 0.25 Methionine 0.25 0.25 0.25 0.25 0.25 Lysine 0.25 0.25 0.25 0.25 0.25 Palm oil - 1.00 - - - Soybean oil - - 1.00 - - Palm kernel oil - - - 1.00 - Groundnut oil - - - - 1.00 Total 100 100 100 100 100 Calculated Composition: Crude protein (%) 24.01 24.08 24.02 24.03 24.07 Energy(Mcal/kg ME) 2.89 2.89 2.89 2.89 2.89 Crude fibre(%) 4.84 4.85 4.90 4.81 4.94
* Vit A – 10,000.00 iu., D3-2,000 iu., B1-0.75g., B2-5g., Nicotinic acid – 25g., K3-2.5g., E-25g., Biotin – 0.05g., Folic acid – 1g., Choline Cholride 250g., Cobalt –0.400g., Copper 8g., Managnese 64g., Iron –32g., Zn – 40g., Iodine – 0.8g., Flavomycin – 100g., Spiramycin 5g 3-Nitro-50g., DL-Methionie –50g., Selenium 0.6g., Lysine 120g., BAT-5g.

39
Table 3: Percentage composition of broiler finisher diets
Ingredients(%)/Diets 1 2 3 4 5
Maize 34.00 35.00 35.00 36.00 35.00
CRM 20.04 17.00 16.00 17.4 16.00
Wheat offal 5.00 6.00 7.0 5.0 7.0
SBM 12.42 11.80 11.50 12.20 11.50
GNC 16.00 16.00 16.00 16.00 16.00
PKC 5.4 6.2 6.50 5.4 6.5
Fish meal 2.00 2.00 2.00 2.00 2.00
Bone meal 4.00 4.00 4.00 4.00 4.00
Salt 0.25 0.25 0.25 0.25 0.25
Vit-min premix* 0.25 0.25 0.25 0.25 0.25
Methionine 0.25 0.25 0.25 0.25 0.25
Lysine 0.25 0.25 0.25 0.25 0.25
Palm oil - 1.00 - - -
Soybean oil - - 1.00 - -
Palm kernel oil - - - 1.00 -
Groundnut oil - - - - 1.00
Total 100 100 100 100 100
Calculated Composition:
Crude protein (%) 20.13 20.12 20.14 20.11 20.14
Energy(Mcal/kg ME) 2.93 2.93 2.93 2.92 2.93
Crude fibre(%) 4.51 4.54 4.63 4.39 4.63
* Vit A – 10,000.00 iu., D3-2,000 iu., B1-0.75g., B2-5g., Nicotinic acid – 25g., Calcium pantothenate 12.5g., B12-0.015g., K3-2.5g., E-25g., Biotin – 0.050g., Folic acid –1g., Manganese 64g., Choline chloride 250g. Cobalt-0.8g., Copper 8g., Manganese 64g., Iron –32G., Zn-40g., Iodine-0.8g., Flavomycin-100g., Spiramycin 5g., Dl-methionie-50g, Selenium 0.6g., Lysine 120g., BAT-5g.

40
3.5 Data Collection and measurements
Body weight, feed intake were measured. Feed intake was done daily by a weigh back
technique, in which feed remaining at the end of the day was subtracted from the initial feed
offered to the birds. Body weight of all the birds was recorded on a weekly basis. Data
obtained were used to calculate feed conversion ratio (FCR), average daily weight gain and
protein efficiency ratio.
3.6 Carcass and organ evaluation
At the end of the experimental period, two birds per treatment were randomly selected from
each treatment, starved overnight and weighed for carcass and organ evaluation. The birds
were slaughtered by severing the jugular vein, scalded in warm water for a minute and de-
feathered by manual plucking. The birds were eviscerated and weighed to obtain their
dressed carcass weight. The kidney, abdominal fat, liver, gizzard, heart, spleen and
proventiculus were removed and weighed using a sensitive scale and their weigts were also
recorded. The dressed carcass and organ weight were expressed as percentages of live
weight.
3.7 Blood collection and Analysis
Blood sample for serum triglyceride and cholesterol were collected into anticoagulant free
test tubes and later centrifuged at 3000 revolutions per minute(rpm) for 10 minutes at room
temperature. The serum was separated into different test tubes leaving the thick blood
residue. Total cholesterol, triglycerides, HDL-C, LDL-C and VLDL were determined
thereafter.
Parameters determined
The parameters determined include as follows
i. Final body weight (g)
ii. Daily feed intake (g)
iii. Average daily weight gain (g)
iv. Feed conversion ratio (g)
v. Protein efficiency ratio (g)
vi. Cost of feed / kg weight gain

41
Carcass Traits
i. Live weight
ii. Dressed carcass
iii. Organ weight
iv. Carcass dressing %
v. Relatives organ weight %
Lipid profile
i. Triglyceride , mg/dl
ii. Total cholesterol
iii. HDL- cholesterol mg/dl
iv. LDL- cholesterol mg/dl
v. VLDL- cholesterol mg/dl
3.8 Experimental Design
The study was conducted in a Completely Randomized Design experiment with five
treatment groups (T1, T2, T3, T4, and T5). Each diet (treatment) was repeated three times,
with each replicate comprising 8 birds. The dietary treatments consist of 1% added fat from
four different sources and were arranged according to the following treatments: T1- birds in
this treatment served as the control group and received diet with no added fat. T2- birds in
this treatment 2 received diet containing 1% palm oil, T3 – birds in this treatment 3 received
diet containing 1% soybean oil, T4- birds in this treatment received diet containing 1% palm
kernel oil and T5- birds in this treatment received diet containing 1% groundnut oil.
The experimental model is as shown.
Xi j= μ + τi + εij
Where: Xi j= individual observation
μ = population mean
τi= effect of dietary fat sources
εij= experiment or random error

42
3.9 Proximate analysis
Feed samples were assayed for their proximate composition at the laboratory of the
Department of Animal Science, University of Nigeria Nsukka. Feed and excreta samples
were assayed for proximate composition by the method of AOAC (1990).
3.10 Statistical analysis
Data generated were subjected to analysis of variance (ANOVA) in CRD using the SPSS
computer package (Snedecor and Cochram, 1980). Significantly different means were
separated using Duncan’s New Multiple Range test. (Duncan, 1955).

43
CHAPTER FOUR
RESULTS AND DISCUSSION
4.1 Results
4.1.1 Proximate composition of experimental Diets
Result of the proximate composition of the experimental diets is presented in Table 4
Table 4: Proximate composition of experimental diet
Treatments*
T1 T2 T3 T4 T5
Dry matter(g) 88.5 91.0 89.0 90.5 89.5
Ash (%) 7.91 6.59 9.55 8.84 9.50
Crude Fibre (%) 5.08 6.04 6.33 4.42 5.03
Crude protein (%) 22.47 21.85 21.78 22.39 23.12
Ether Extract (%) 4.52 3.85 2.25 2.76 5.59
Nitrogen Free Extract
(%)
60.02 61.67 60.09 61.59 56.76
T1 = No oil; T2 = Palm oil; T3 = Soybean oil; T4 = Palm kernel oil; T5 = Groundnut oil.
The crude protein contents of the diets are 23.12%, 22.47%, 22.39 % 21.85% and 21.78 for
diets 5,1,4,2 and 3, respectively. Dietary treatment 5 had the highest crude protein value.
Diets 1,2,3,4 and 5 contained 5.08%, 6.04%, 6.33%, 4.42 % and 5.03% crude fibre,
respectively. The crude fibre content of treatment 3 was the highest while diet 4 had the least
fibre content. The nitrogen- free extract contents of diets 1,2,3,4 and 5 were 60.02%, 61.67%.
60.09%, 61.59%, and 56.76 %, respectively. Treatment had the highest NFE value while
treatment 5 had the least value of NFE. Treatment 5 had the highest content of ether extract
while treatment 3 had the least ether extract content. Treatment 2 had the least ash content of
6.59%, while treatment 3 had the highest ash content of 9.55%.
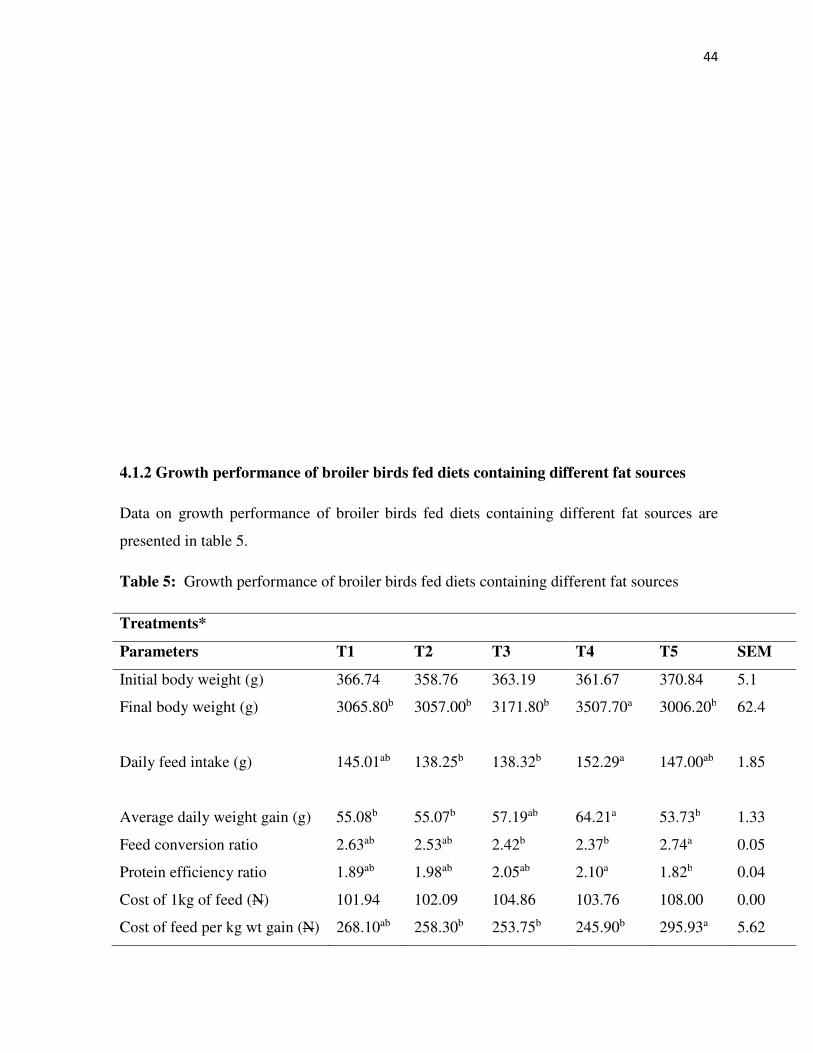
44
4.1.2 Growth performance of broiler birds fed diets containing different fat sources
Data on growth performance of broiler birds fed diets containing different fat sources are
presented in table 5.
Table 5: Growth performance of broiler birds fed diets containing different fat sources
Treatments*
Parameters T1 T2 T3 T4 T5 SEM
Initial body weight (g) 366.74 358.76 363.19 361.67 370.84 5.1
Final body weight (g) 3065.80b 3057.00b 3171.80b 3507.70a 3006.20b 62.4
Daily feed intake (g) 145.01ab 138.25b 138.32b 152.29a 147.00ab 1.85
Average daily weight gain (g) 55.08b 55.07b 57.19ab 64.21a 53.73b 1.33
Feed conversion ratio 2.63ab 2.53ab 2.42b 2.37b 2.74a 0.05
Protein efficiency ratio 1.89ab 1.98ab 2.05ab 2.10a 1.82b 0.04
Cost of 1kg of feed (N) 101.94 102.09 104.86 103.76 108.00 0.00
Cost of feed per kg wt gain (N) 268.10ab 258.30b 253.75b 245.90b 295.93a 5.62

45
Mortality (%) 4.17c 8.33b 16.67a 8.33b 0.00d 2.38
a,b,c,d Means on the same row with different superscript are significantly (P < 0.05) different
SEM= Standard error of mean.
T1 = No oil; T2 = Palm oil; T3 = Soybean oil; T4 = Palm kernel oil; T5 = Groundnut oil.
Final body weight (FBW) varied significantly (p < 0.05) among the treatments. Birds fed diet
containing palm oil (T4) had high FBW than the birds on the other treatments whose values
were statistically the same. Daily feed intake (DFI), average daily weight gain (ADWG),
feed conversion ratio (FCR), protein efficiency ratio (PER), cost feed per kg weight gain
(CFG) and mortality were all significantly (p<0.05)affected by dietary treatments. Birds on
treatment 4 had similar DFI as birds on treatment 5 and 1 but higher than those of on
treatments 2 and 3.
Birds fed diet containing with palm kernel oil (treatment 4) had similar (p>0.05) average
daily weight gain value as birds on treatment 4 (soybean oil) and this was significantly
(p<0.05) higher than the average weight gain value of birds on treatments 1, 2 and 5. The
average daily weight gain value of birds on treatments 1, 2, 3 and 5 were similar (p>0.05).
The feed conversion ratio values of birds on treatment 5 were similar (p>0.05) to those of
birds on treatments 1 and 2 and this was significantly (p<0.05) higher than the feed
conversion ratio value of birds on treatments 3 and 4. Birds on treatments 1, 2, 3 and 4 had
similar values in fed conversion ratio.
Birds on treatment 4 had similar (p>0.05) protein efficiency ratio value as those of birds on
treatments 1, 2 and 3, and this was significantly (p<0.05) higher than the protein efficiency
ratio value of birds on treatment 5. The PER values of birds on treatments 1, 2, 3, and 5 were
similar (p>0.05).
Birds fed diet containing groundnut oil had similar (p>0.05) cost of feed per kg weight gain
with those on treatment 1, and this was significantly higher than the cost of feed per kg

46
weight gain of birds on treatments 2, 3 and 4. The cost of feed per kg gain values of birds on
treatment 1, 2, 3 and 4 were similar (p>0.05).
Birds on treatment 3 had the highest (p<0.05) % mortality, followed by birds on treatment 2
and 4 whose values were however similar (p>0.05), followed by birds on treatment 1, while
birds on treatment 5 had the lowest (p<0.05) % mortality.
4.1.3 Carcass and organ characteristics of broiler birds fed diets containing different fat
sources
Data on carcass and organ characteristics of broiler birds fed diets containing different fat
sources are presented in table 6.
Table 6: Carcass and Organ Characteristics of broiler birds fed diets containing different fat
sources.
Treatments*
Parameters T1 T2 T3 T4 T5 SEM
Live weight (kg) 3.43a 3.05ab 3.13ab 3.58a 2.55b 0.14
Carcass weight (kg) 2.35ab 2.05b 2.35ab 2.90a 1.75b 0.14
Dressing % 70.68bc 67.26c 74.94ab 81.10a 68.52bc 1.77
Kidney (g) 1.70b 1.20b 3.70ab 5.60a 2.25b 0.58
Abdominal fat (g) 37.35b 37.35b 16.05c 46.10a 11.05c 5.58
Spleen (g) 4.80a 3.45b 3.60b 4.21a 2.30c 0.38
Proventiculus (g) 13.70a 11.55ab 12.60ab 13.85a 9.45b 0.65
Heart (g) 13.75a 11.15ab 13.05a 14.15a 8.55b 0.78

47
Liver (g) 69.45a 59.35ab 58.65ab 59.75ab 48.00b 2.86
Gizzard (g) 56.35a 51.20ab 50.15ab 46.45b 40.85b 2.65 a,b,c,d Means on the same row with different superscript are significantly (P < 0.05) different
SEM= Standard error of mean.
T1 = Control; T2 = Palm oil; T3 = Soybean oil; T4 = Palm kernel oil; T5 = Groundnut oil
The result showed that there were significant differences (p<0.05) in all the carcass and
organ parameters measured.
Birds on treatments 1 and 4 had similar (p>0.05) live weight values with those on treatments
2 and 3 and these were significantly (p<0.05) higher than the mean live weight value of birds
on treatment 5. Treatments 2, 3 and 5 birds had similar (p>0.05) mean live weight values.
Birds on treatments 1, 3 and 4 had similar (p>0.05) mean values of carcass weight, while
birds on treatments 1, 2, 3 and 5 had similar (p>0.05) mean values. Treatment 4 birds had
significantly (p<0.05) mean higher from dressed carcass weight value than birds on
treatments 2 and 5.
Birds on treatments 3and 4 had significantly (p<0.05) higher mean dressing % value than
those on treatments 1, 2 and 5 which had similar mean dressing values.
Birds on treatment 4 had similar (p>0.05) mean kidney value with those on treatment 3 , and
this was significantly (p<0.05) higher than the mean kidney values of birds on treatment 1,2
and 5. The mean kidney values of birds on treatments 1, 2, 3 and 5 were similar (p>0.05).
Birds on treatment 4 had the highest (p<0.05) mean abdominal fat value. Birds on treatment
1 had similar (p>0.05) abdominal fat value with those on treatment 2, and these were
significantly (p<0.05) higher than the abdominal fat values of birds on treatment 3 and 5.
The mean spleen values of birds on treatments 1 and 2 are similar (p>0.05), and are
significantly (p<0.05) higher than those of the birds on treatments 2 and 4 which were also
similar (p>0.05), but were significantly (p<0.05) higher than the abdominal fat value of birds
on treatment 5 which had the least mean value of (2.30g).

48
Birds on (p<0.05) higher than the mean proventiclous value of birds on treatment 5. The
mean proventiculous values of birds on treatments 2 , 3, and 5were similar.
Birds on treatment 1 had similar (p>0.05) mean proventiculous weight value with those on
treatments 4,3 and 2and this was significantly weight value of birds on treatment 1 was
similar (p<0.05) to the mean heart weight values of birds on treatments 2,3 and 4,and this
was significantly (p<0.05) higher than the mean heart weight value of those on treatment
5.Birds on treatments 2 and 3 had similar (p<0.05) values.
Birds on treatment 1 had similar (p>0.05) mean liver weight value with those on treatments
2, 3 and 4, and this was significantly (p<0.05) higher than the liver weight value of birds on
treatment 5. Birds on treatments 2, 3,4 and 5 had similar values. The liver value of birds on
treatment 1 were significant different those on treatment 5. Treatment 1 birds had the highest
mean value of (69.45g) and treatment 5 had the least mean value of (48.00g).
The gizzard weight value of birds on treatment 1 was similar (p<0.05) to the mean gizzard
weight values of those on treatments 2 and 3, and this was significantly (p<0.05) higher than
the gizzard value of birds on treatments 4 and 5. Treatments 2, 3, 4 and 5 birds had similar
(p<0.05) gizzard weight values.
4.1.4 Relative Organ Weight of broiler birds fed diets containing different fat sources
Data on relative organ weights of broiler birds fed diets containing different fat sources are
presented in Table 7.
Table 7: Relative Organ Weight of broiler birds fed diet containing different fat
sources.
Treatments*
Parameters T1 T2 T3 T4 T5 SEM
Gizzard (%) 1.70 1.68 1.64 1.30 1.59 1.85
Heart (%) 0.42a 0.37ab 0.42a 0.40ab 0.33b 0.01
Kidney (%) 0.05c 0.04c 0.12ab 0.16a 0.09bc 0.02
Abdominal fat (%) 1.12 1.23 0.56 1.29 0.44 0.16
Spleen (%) 0.15 0.12 0.12 0.12 0.09 0.01

49
Proventiculus(%) 0.42 0.38 0.41 0.39 0.37 0.01
Liver (%) 2.09 1.95 1.89 1.68 1.88 0.06
a,b,c,d Means on the same row with different superscript are significantly (P < 0.05) different
SEM = Standard Error of Mean.
T1 = Control; T2 = Palm oil; T3 = Soybean oil; T4 = Palm kernel oil; T5 = Groundnut oil
While no significant (p>0.05) differences existed among treatments in relative gizzard
weight, relative abdominal fat weight, relative spleen weight, relative proventicles weight
and relative liver weight, there were significant (p<0.05) differences among treatments in
relative heart weight and relative kidney weight.
The relative heart weight values of birds on treatments 1 and 3 were significantly (p<0.05)
higher than the RHW values of birds on treatment 5. However birds on the treatments 1 and
3 had similar (p>0.05) RHW values with those on treatments 2 and 4. The RHW value of
birds on treatments 2, 4 and 5 were also similar (p>0.05).
Birds on treatment 4 had similar (p>0.05) relative kidney weight value as those on treatment
3 but significantly (p<0.05) higher than the RKW of birds on treatments 1,2 and 5 which had
similar RKW values. The RKW of birds on treatments 3 and 5 were also similar (p>0.05).
4.1.5 Lipid profile of broiler birds fed diets containing different fat sources
Data on Lipid Profile of broiler birds fed diets containing different fat sources are presented
in Table 8.
Table 8: Lipid profile of broiler birds fed diet containing different fat sources.
Treatments*
Parameters T1 T2 T3 T4 T5 SEM
Total
cholesterol(mmol/l)
3.34b 3.09b 3.84ab 4.58a 3.22b 0.21
HDL-C (mmol/l) 0.74ab 0.93a 0.55b 0.68ab 0.61b 0.05
LDL-C(mmol/l) 1.37c 1.88bc 2.19b 3.88a 1.83bc 0.29
Trigys(mmol/l) 1.48a 1.39a 0.53b 1.36a 1.39a 0.14
VLDL(mmol/l) 0.75a 0.70a 0.27b 0.69a 0.70a 0.07

50
a,b,c,d Means on the same row with different superscript are significantly (P < 0.05) different
SEM= standard error of mean
T1 = Control; T2 = Palm oil; T3 = Soybean oil; T4 = Palm kernel oil; T5 = Groundnut oil.
HDL-C= High density lipoprotein cholesterol, LDL-C= Low density lipoprotein cholesterol,
Trigs= Triglycerides, VLDL= Very low density lipoprotein.
The result showed that significant differences (p<0.05) existed among treatments in all the
lipid profile parameters measured.
Birds on treatment 4 had similar (p>0.05) total cholesterol value as those on treatment 3, but
was significantly (p<0.05) higher than the mean total cholesterol values of birds on
treatments 1, 2, and 5.The total cholesterol value of birds on treatments 1, 2,3 and 5 were also
similar(p>0.05).
The mean HDL values of birds on treatment 2 was similar (p>0.05) to those on treatments 1
and 4, but was significantly (p<0.05) higher than the mean HDL values of birds on
treatments 3 and 5. The mean HDL values of birds on treatments 1, 3, 4 and 5 were similar
(p<0.05).
The mean LDL value of birds on treatment 4 was significantly (p<0.05) higher than the mean
LDL values of birds on treatments 1, 2, 3 and 5 followed, birds on treatment 3 whose LDL
values was similar (p>0.05) to those of birds on treatment 2 and 5,but significantly (p>0.05)
higher than the mean LDL value of birds treatment 1.
Birds on treatments 1, 2, 4 and 5 had similar (p> 0.05) triglycerides values which were
however, significantly (p<0.05) higher than the triglycerides value of birds on treatment 3.
Also, the mean VLDL value of birds on treatments 1, 2, 4 and 5 were similar (p>0.05) but
significantly (p<0.05) higher than the VLDL value of birds on treatment 3.

51
4.2 Discussion
4.2.1 Growth performance of broiler birds fed diets containing different fat sources.
As shown in Table 2, there were significant differences (p<0.05) among dietary treatments
on growth performance of broilers. In the present study, it was observed that birds fed diets
with supplemented fat sources improved in body weight and feed efficiency than the control
group.
The significant improvement in growth performance of birds fed diets containing dietary fat
from palm kernel oil and soybean oil could be attributed to the ability of these fat sources to
reduce the passage rate of the digesta through the gastrointestinal tract, thereby allowing for
better nutrient absorption and utilization (Latshaw, 2008). It has been previously reported
that dietary supplementation with vegetable or animal fats did not affect feed passage rate
through the digestive tract in Leghorn chicken (Golian and Polin, 1984). However, the
dissimilarity between the report of Golian and Polin (1984) and the present results could be
attributed to the differences in dietary fat sources and also to the type of birds used in the two
studies. Although the inclusion of fat in broiler feed is beneficial to the growth of birds, one
of the major concerns with regards to dietary fat supplementation is the actual metabolizable
energy (ME) value that can be assigned to each fat source. However, the improvement in
utilization of dietary fat components in bird is enhanced by different sources regardless of
ME content (Jeffri et al., 2010). Early research on the inclusion of fats in poultry diets
indicated that plant-derived oils had higher ME value when compared to animal sources with
high free fatty acid content (Waldroup et al., 1995). Contrally to the results obtained in the
present study, Pesti et al. (2002), Ghazalah et al., (2008) and Mohammadu et al. (2011)
indicated that there were no significant differences among treatments in all the growth
parameters evaluated when broilers were fed diets containing fat from different sources.
The result showed that there were marked differences in daily. Birds on diets supplemented
with palm oil and soybean oil (treatments 2 and 3) had lower values than the rest of the
treatments, which might be as a result of the energy content of these dietary fat sources
(Jeffri et al., 2002). The observed result on feed intake thus could indicate that the
improvement in body weight may have partially been due to improved nutrient digestibility

52
and better nutrient absorption as reflected by improved feed efficiency. It was also found that
broilers fed diets containing palm kernel oil had better growth rate than those in the other
treatments, especially the control group. This observation confirms the advantage of adding
vegetable oil in birds’ diet (Gallardo et al.2012). The improved growth rate may be due to
the higher percentage of long chain fatty acids and higher contents of triglycerides in
vegetable oil (Thacker et al., 1994). The observed similarities in protein efficiency ratio
(PER) of birds in the various treatments may be attributed to the fact that vitamins present in
most dietary fat sources are involved in amino acid metabolism. Vitamin B12 for instance is
required for the production and development of blood cells (sources: Rita T. dela Cruz of
www.bar.gov.ph).
As shown in Table 2, the percentage mortality of birds in treatment 5 was significantly lower
than the values observed in the rest of the treatments. It does seem that groundnut oil had
high level of monounsaturated fatty acid and reduced level of saturated fatty acid which
might have helped to decrease the level of cholesterol. Groundnut oil also has antioxidants
such as vitamin E which might also play an important role in reducing stress in the birds fed
dietary groundnut oil (Anyasor et al. 2009).
4.2.2 Carcass and Organ characteristics of broiler birds fed diets containing different
fat sources.
As shown in table 3, birds fed diet containing palm kernel oil (treatment 4) had significantly
(p<0.05) higher dressing % than those on treatments 1(control), 2 (palm oil) and 5
(groundnut oil). The results are similar to the findings of Felton and Kerley (2004), but
differed from the results reported by Alaedein (2013) which showed that there were no
significant difference in dressing percentage of birds fed soybean oil. The results are also in
consonance with that reported by Andretti et al .(2001) which showed that broilers fed diets
containing poultry fat, canola, sunflower oil, corn, soybean and lad, respectively had similar
carcass yield values. However, the authors observed significant differences (p>0.05) among
treatments in the organ weights measured.
Table 3 also shows that broilers fed diet containing palm kernel oil had the highest
abdominal fat value, while the least values were observed in treatments 3 and 5(diet

53
containing soybean oil and groundnut oil, respectively). Among available fat sources in
poultry nutrition, it has been recognized that soybean oil has adequate essential fatty acids,
unsaturated fatty acid (such as linolenic acid) that can improve broiler performance, and also
linolenic acid can be converted to longer chain omega -3 fatty acids that is an important
factor in animal feeding as health promoter (Bezard et al., 1994). The increase in abdominal
fat observed in birds that consumed the diet containing palm kernel oil is in line with the
results reported by Shahyar et al .(2011) which showed that the addition of 6% fat caused a
significant increase in abdominal fat weight of broilers as compared to those fed control diet.
The results reported by Sanz et al. (2000b) in a study in which the effect of two lipid sources
(beef tallow, sunflower and soybean oil) were evaluated, showed that there was a significant
reduction in the abdominal fat of birds fed diet containing sunflower oil. This shows that the
treated broilers were able to utilize the unsaturated lipids in the vegetable oils very
efficiently. This may have resulted in the reduction of the abdominal fat with the resultant
increase in the lean carcass weight of broilers. This is in agreement with the result of the
present study were birds fed diets with soybean oil and groundnut oil has the least abdominal
fat deposition. Crespo and Esteve- Grarcia (2002) suggested that reduction in the abdominal
fat of broilers fed diets containing soybean oil and groundnut oil seems to be a consequence
of higher lipid oxidation despite the higher synthesis of endogenous fatty acids. Soybean oil
contains high levels of unsaturated acids and are more completely digested of by fowls.
The dressed carcass weight and most of the organ weights (kidney, spleen, heart,
proventriculus, liver and gizzard weights) were significantly (p<0.05) affected by the
inclusion of fat from different sources when compared to the control group. The spleen,
heart, proventriculus, liver and gizzard weights were significantly (p<0.05) higher in the
control as compared to the group fed diet containing groundnut oil. Similar observation had
been reported by Tabiedian et al. (2005) in a study that investigated the effect dietary fats on
organ characteristics of birds. The difference observed in the gizzard weight for instance
might be due to decrease in the passage rate of food which might have gradually increased
gizzard function with the resultant increase in gizzard muscular mass, a condition known as
muscular hypertrophy. This suggestion is in agreement with the observations of Lopez-Ferrer
et al. (2001) and Habib et al. (2011). The liver and to some extent the heart play a major role

54
in detoxification, their similarity in weight tends to suggest that the different dietary
treatments did not have any toxicity effect on these organs. This is in line with the findings of
Diara et al. (2014).
4.2.3 Relative organ weights of broiler birds fed diets containing different sources
Table 4 shows that there were significant differences (p<0.05) among treatments in the
relative weights of heart and kidney of broiler birds fed dietary fat sources. This result
contradicts the result reported by Aydin (2007) which showed that there were no significant
differences among treatments in relative organ weights of birds fed diets containing oil and
linoleic acid.
4.2.4 Lipid profile of broiler birds fed diets containing different fat sources
As shown in Table 5, dietary treatments had significant (p<0.05) effects on all the
parameters (total cholesterol, HDL-C, LDL-C, triglycerides and VLDL) evaluated. The
present results are in agreement with the findings reported by Ozdogan and Aksit (2003).
Birds on treatment 4(diet containing palm kernel oil) had significantly (p<0.05) higher level
of total cholesterol, while birds on treatment 3(diet containing soybean oil) had significantly
(p<0.05) lower level of triglycerides when compared to the control group. Palm kernel oil is
high in lauric acid which has been shown to raise blood cholesterol (Cascaboulho, 2000).
Cholesterol serves as a precursor for biosynthesis of steroid hormones, bile acid and vitamin
D (Weslover et al., 2003). According to (Blanch et al., 1995), blood cholesterol values are
influenced by dietary ingredients. This could be the reason for the various changes seen in
the blood cholesterol parameters of birds fed different dietary fat sources in this study. The
results (Table 5) reveal that a negative relationship existed between cholesterol and
triglyceride values as seen in some of the treatment groups. For instance the total cholesterol
levels of birds fed diet containing 0% oil, palm oil and groundnut oil were low while the
triglyceride levels were higher. In previous studies, animals fed diets rich in cholesterol or
saturated fat had elevated carcass and blood cholesterol levels. However, dietary ingredients
can be manipulated to change the blood values (Blanch et al., 1995; Verma et al; 1995). Low
density lipoprotein (LDL) cholesterol and high density lipoprotein (HDL) cholesterol were
significantly affected by dietary fat sources. This result is in agreement with that of Nobar et
al. (2010) in a study which investigated the effect of canola oil as dietary fat source on lipids

55
profiles of turkey. The lowest value of LDL was observed in the control group, although
birds fed diets containing palm oil and groundnut oil had similar values. It does seem that the
inclusion of dietary fat from soybean and palm kernel oil increased significantly the serum
LDL. The LDL particles pose a risk for cardiovascular diseases, and increase in the
concentration of LDL particles is strongly associated with increasing rates of accumulation of
atherosclerosis within the walls and may eventually lead to sudden plaque rupture and trigger
clot within arterial opening (www.nejm.org). The HDL was higher in the treatment group
that was fed dietary palm oil, although the HDL value was similar to the values observed in
the control group and the group on palm kernel oil. It has been demonstrated that increases in
dietary fat correlate closely with the secretion of lipases and levels of high density lipoprotein
(Coring et al., 1989). However, there is relatively little information regarding the effects of
the quality of the dietary fat on the secretory activity of the pancreas, liver and other vital
organs in monogastrics. Ozdogan and Aksit (2003) also reported significant differences in
LDL and HDL of broilers fed different fat sources which had different energy levels.
According to Sorrentino (2011), unlike other larger lipoprotein particles which deliver fat
molecules to cells, HDL particles remove fat molecule from cells which are involved in the
export of fat molecules. Stephen et al. (2012) reported that increasing concentrations of HDL
particles are strongly associated with decreasing accumulation of atherosclerosis which
eventually results in sudden plaque ruptures, cardiovascular disease and other vascular
diseases. The inclusion of fat from palm oil and palm kernel oil in broiler diets seem to be
beneficial since this leads to increase in HDL. It is therefore safe to consume meat from
broilers fed diets containing vegetable oil, especially palm oil. The VLDL values of birds on
all the treatments were significantly (p<0.05) different, with birds on treatment 3 (soybean
oil) having the least value of 0.27mg/dl. A value of less than 32mg/dl is desirable (Gibbons
et al., 2004). The value obtained in the present study is therefore below normal range and this
does not pose any danger to the animals or those who consume the meat. It is known that
VLDL transport endogenous products and is converted in the blood stream to LDL (Dashti et
al., 2011). The result observed is in agreement with that reported by Ozdogan and Aksit
(2003).

56
CHAPTER FIVE
SUMMARY AND CONCLUSION
5.1 Summary
In an eight-week study conducted at the Poultry Unit of the Department of Animal
Science Teaching and Research Farm, University of Nigeria, Nsukka with one hundred and
twenty day-old broiler chicks, the effect of dietary fat sources on growth performance,
carcass trait, organ characteristics and lipid profile of broilers was evaluated. Results showed
that there were significant differences (p<0.05) among dietary treatments on growth
performance of broilers. Birds supplemented with fat sources improved in body weight and
feed efficiency than the control group, especially treatment 4 (palm kernel oil), and treatment
3 (soybean oil). The observed significant improvement in growth performance of birds was
attributed to the ability of these fat sources to reduce the passage rate of the digesta through
the gastrointestinal tract, thereby allowing for better nutrient absorption and utilization. The
results also showed that there were marked differences in daily and total feed intake. Birds on
diets supplemented with palm oil and soybean oil (treatments 2 and 3) had lower values than
the rest of the treatments.
Birds fed diet containing palm kernel oil (treatment 2) had significantly (p<0.05) higher
dressing % than those on treatments 1(control), 2 (palm oil) and 5 (groundnut oil). The
results also showed that broilers fed diet containing palm kernel oil had the highest
abdominal fat value, while the least values were observed in treatments 3 and 5(diet
containing soybean oil and groundnut oil, respectively. The dressed carcass weight and most
of the organ weights (kidney, spleen, heart, proventriculus, liver and gizzard weights) were
significantly (p<0.05) affected by the inclusion of fat from different sources when compared
to the control group. The spleen, heart, proventriculus, liver and gizzard weights were
significantly (p<0.05) higher in the control as compared to the group fed diet containing
groundnut oil. There were significant differences (p<0.05) among treatments in the relative
weights of heart and kidney of broiler birds fed dietary fat sources.

57
Dietary treatments had significant (p < 0.05) effects on all the parameters (total cholesterol,
HDL-C, LDL-C, triglycerides and VLDL) evaluated. Birds on treatment 4(diet containing
palm kernel oil) had significantly (p<0.05) higher level of total cholesterol, while birds on
treatment 3(diet containing soybean oil) had significantly (p<0.05) lower level of
triglycerides when compared to the control group. Low density lipoprotein (LDL) cholesterol
and high density lipoprotein (HDL) cholesterol were significantly affected by dietary fat
sources. The HDL was higher in the treatment group that was fed dietary palm oil, although
the HDL value was similar to the values observed in the control group and the group on palm
kernel oil. The VLDL values of birds on all the treatments were significantly (p<0.05)
different, with birds on treatment 3 (soybean oil) having the least value of 0.27mg/dl.
5.2 Conclusion
It is evident from the results obtained in the present study that addition of vegetable fat from
palm kernel oil and groundnut oil in broiler diets enhanced the growth performance, carcass
and organ weights of birds. Birds that consumed the diets containing soybean oil had the
least very low density lipoprotein, while those that consumed diet containing palm kernel oil
had similar High Density Lipoprotein Cholesterol value with birds on control.
5.3 Recommendation
Based on the findings of the present study it is recommended that farmers should add any of
vegetable oils investigated, especially soybean oil in the diets of broiler starters and finishers
because such addition will enhance growth performance and carcass yield of birds, result
in less abdominal fat, induce the production of high levels of unsaturated fatty acids and
high density lipoprotein cholesterol and reduce the level of very low lipoprotein
cholesterol.

58
REFERENCES
Alaedein M. A (2013). Feeding alternative source of fat to broilers. Reseacher, 5 (10): 84-87. Alao, S.J and Balnave D. (1984) Growth and carcass composition of broiler fed sunflower
and olive oil. British Poultry Science; 25:209-219. Alfred Thomas (2002). "Fats and Fatty Oils". Ullmann's Encyclopedia of Industrial
Chemistry. Ullmann's Encyclopedia of Industrial Chemistry. Weinheim: Wiley-VCH. doi:10.1002/14356007.a10_173. ISBN 3527306730.
Andreotti, M.O, Junqueira O.M, Cancherini, L.C, Rodrigues, E.A, Sakomura, N.K.(2001)
Valor nutricional de algumas fontes de gordura para frangos de corte. Anais da 38th Reunião Anual da Sociedade Brasileira de Zootecnia; Piracicaba, SP. Piracicaba, SBZ.
Anyasor G. N., Ogunwenwo K. O., Oyelana O. A., Ajayi D. and Dangana J. (2009). Chemical analysis of Groundnut (Arachis hypogeal oil). Pakistan journal of nutrition. 8 (3): 269-272.
AOAC (1990). Association of official Analytical Chemists. Official Methods of Analysis (21th edition) Washington D.C.USA.
Aslam, S. M., J. D. Garlich, and M. A. Qureshi. (1998). Vitamin D deficiency alters the immune responses of broiler chicks. Poult. Sci. 77:842-849.
Atteh, J.O.( 2002). Principle and practice of livestock feed manufacturing. Adlek printers,
Ilorin, Nigeria, pp:13-17. Attia, Y. A., A. S. Hussein, A. E. Tag El-Din, E. M. Qota, A. I. Abed El-Ghany, and A. M.
El-Sudany.( 2009). Improving productive and reproductive performance of dual-purpose crossbred hens in the tropics by lecithin supplementation. Trop. Anim. Health and Prod. 41:461-475.
Austic, R.E., (1995). Poultry In: Basic Animal nutrition and feeding pond. W.G., D.G. Church and K. R. Pond (Eds). John Wiley and sons Canada, ltd.New York, pp:495-515.
Aydin, R. (2007). Effect of dietary oils and conjugated linoleic acid on the growth performance of broilers vaccinated with the La Sota Newcastle vaccine. South
African Journal of Animal Science. 37 (2):74- 80.
Azman,M.A., Ubrahim, H., ER, U., Nurg. B.(2005).Effects of various dietary fat sources on performance and body fatty acid composition of broiler chickens. Turk. .J. Vet Anim Sci 29: 811-819.

59
Baião, N.C.and Lara, L.J.C. (2005). Oil and fat in broiler nutrition. Braz J Poult Sci.;1:129–141.
Balnave, D. (1971). Response of laying hens to dietary supplementation with energetically equivalent amounts of maize starch or maize oil. J Sci. Food Agri. 22:125-128.
Batal, A. B., and C. M. Parsons. (2002). Effects of age on nutrient digestibility in chicks fed different diets. Poult. Sci. 81:400-407.
Beauregard, L., Moustafa, A., Sampaio, J M. (1996) Puntos críticos a considerer en la refinación de aceites para la producción de grasas y margarinas. Soya Notícias; 16:10-15.
Berhe, E.T. and R.M. Gous, (2005). Effect of dietary protein content on allometric
relationships between carcass portions and body protein in cob and Ross broilers. Proceedings of the 24th Conference of south African Branch of WPSA, Pretina, Jaboticabal, UNESP.
Bezard J, Blond JP, Bernard A and Clouet P, (1994). The metabolism and availability of essential fatty acids in animal and human tissues. Reprod Nutr Dev.;34:539–568. [PubMed]
Blanch A, Barroeta AC, Baucells MD and Puchal F (1995). The nutritive value of dietary fats in relation to their chemical composition. Apparent fat availability and metabolizable energy in two-week-old chicks. Poultry Science; 74:1335-1340.
Blanch, A. and Grashorn, M.A., (1995). Effect of different dietary fat sources on general
performance and carcass yield in broiler chickens. Proc. 12th Eur. Symp. Qual. Poult. Meat, Zaragoza, Spain. pp. 71-75.
Bornstein S, Lipstein B. (1963) Some unusual waste vegetable oils or fat supplements in practical broiler rations. Poultry Science 1963; 42: 172-184.
Breinholt, K.A., Gowen, F.A.Y. and Nwosu, C.C. (1981). Influence of environmental and
animal factors on day and night grazing activity of imported Holstein-Friesian cows in the humid lowland tropics of Nigeria. Tropical Animal Production 6 (4):328-336.
Buyse, J., E. Decuypere, L. Berghman, E.R. Kuln and F. Vandesande, 1992. The effect of dietary protein content on episodic growth hormorn secretion and on heat production of male broilers chickens. Br. Poult. Sci., 33:1101-1109.
Cançado, S.V.(1999) Efeito do jejum pré-alojamento de pintos de corte sobre o desempenho, órgãos digestivos e digestibilidade da ração [tese]. Belo Horizonte: Escola de Veterinária, UFMG.
Carew Jr, L. B., R. H. Machemer Jr, R. W. Sharp, and D. C. Foss. (1972). Fat absorption by
the very young chick. Poult. Sci. 51:738-742.

60
Carew, L. B., and F. W. Hill. (1964). Effect of corn oil on metabolic efficiency of energy utilization by chicks. J Nutr. 83:293.
Cascabulho, A.R.(2000) Efeitos de diferentes óleos de soja na composição de gordura da carcaça de frango de corte [dissertação]. Belo Horizonte: Escola de Veterinária, UFMG.
Celebi S, Utlu N. (2004). Laying performance, serum lipoproteins, cholesterol and triglyceride of hens as influenced by dietary fat sources. J Appl Anim Res.;25:121–124.
Chanmugam P, Boudreau M, Boutte T, Park RS, Hebert J, Berrio L, Hwang DH.(1992) Incorporation of different types of n-3 fatty acids into tissue lipids of poultry. Poultry Science ; 71:516 521.
Cho, J. H., Y. J. Chen, J. S. Yoo, W. T. Kim, I. B. Chung, and I. H. Kim. (2008). Evaluation
of fat sources (lecithin, mono-glyceride and mono-diglyceride) in weaned pigs: Apparent total tract and ileal nutrient digestibilities. Nutr. Res. Practice 2:130.
Collins, C. L., A. K. Lealiifano, H. Akit, F. T. Fahri, P. C. Baskett, F. R. Dunshea, R. Australia, and N. S. W. Corowa. (2011). Influence of soyabean lecithin on carcass weight and dressing percentage
Cook, M. E., C. C. Miller, Y. Park, and M. Pariza. (1993). Immune modulation by altered nutrient metabolism: nutritional control of immune-induced growth depression. Poult. Sci. 72:1301-1305.
Coring, T., Juste, and Lhoste, E.F. (1989).Nutritional regulation of pancreatic and biliary secretions.Nutr. Res. Rev. 2 : 161-180.
Crespo N, and Esteve-Garcia E. (2002). Nutrient and fatty acid deposition in broilers fed
different fatty acid profiles. Poult Sci.81:1533–1542b. [PubMed]
Crespo, N., and Esteve-Garcia E.(2002) Dietary polyunsaturated fatty acids decrease fat deposition in separable fat depots but not in the remainder carcass. Poultry Science; 81:512-518a.
Dashty M, Kulik W, Hoek F, Veerman EC, Peppelenbosch MP, Rezaee F. (2011). "A phospholipidomic analysis of all defined human plasma lipoproteins.". Sci Rep. 1 (139). doi:10.1038/srep00139. PMC 3216620. PMID 22355656.
Dashty M, Motazacker MM, Levels J, de Vries M, Mahmoudi M, Peppelenbosch MP, Rezaee F. (2014). "Proteome of human plasma very low-density lipoprotein and low-density lipoprotein exhibits a link with coagulation and lipid metabolism.". Thromb Haemost. 23 (111): 518–530. doi:10.1160/TH13-02-0178. PMID 24500811.

61
De Groóte, G. (1968). Energetic evaluation of unstabilized and stabilized fish meals in terms of metabolizable energy and net energy for maintenance and growth. Feedstuffs 40:26-27.
Deaton, J.W, Mcnaughton, J.L, Reece, F.N, Lott, B.D.(1981).Abdominal fat of broilers as influenced by dietary level of animal fat. Poultry Science; 60:1250-1253.
Diarra M.S., Delaquis P., Pempel H., Bach S., Harltin C., Aslam M. (2014). Antibiotic
resistance and diversity of salmonella entercia associated with broiler chickens. J.
food prdt. 77, 40-99.
Drevon, C. A. (1991). Absorption, transport and metabolism of vitamin E. Free Radical Res. 14:229-246.
Duncan, D.B. (1955). New Multiple Range Test Biometrics, 11:1-42.
Dutra, Jr, W.M., Ariki, J, Kronka, S.N, Junqueira, O.M.(1991) Óleo de abatedouro avícola em comparação ao óleo de soja na alimentação de frangos de corte. Revista da Sociedade Brasileira de Zootecnia; 20(5):471-475.
Dvorin, A., Z. Zoref, S. Mokady, and Z. Nitsan. (1998). Nutritional aspects of hydrogenated
and regular soybean oil added to diets of broiler chickens. Poult. Sci. 77:820-825.
Edwards, H. M. (1969). Factors influencing the efficiency of energy utilization of growing chickens, with special reference to fat utilisation. Feedstuffs, Minneapp 41:14-15.
Edwards, H.M, Denmamn F, Abou-Ashour A, Nugara D.(1973). Influences of age, sex and type of dietary fat supplementation on total carcass and fatty acid composition. Poultry Science 1973;52:934-948.
Edwards Jr, H. M. (1990). Efficacy of several vitamin D compounds in the prevention of
tibial dyschondroplasia in broiler chickens. J Nutr. 120:1054.
Energy Center, UNN, 2008.
Escribano, F.(1991) Fisiologia digestiva y metabolismo de las grasas e hidratos de carbono en gallinas ponedoras. In: Nutrición y alimnetación de gallinas ponedoras. Madrid: Ediciones Mundiprensa.
Fats in animal feeds. Illinois: Milk Specialties; 1985. 22p. Fedna, N.(1999). FEDNA para la formulación de piensos compuestos. Madrid: Ediciones
Peninsular. Felton, E.E., and M.S. Kerley. (2004). Performance and carcass quality of steers fed different
sources of dietary fat. J. Anim. Sci. 82:1794-1805.

62
Ferreira, W.M. (1999).Digestão e metabolismo dos lipídios. Belo Horizonte: Escola de Veterinária da UFMZ. p.1-34.
Ferrini G, Baucells MD, Esteve-Garcia E, and Barroeta AC. (2008). Dietary polyunsaturated
fat reduces skin fat as well as abdominal fat in broiler chickens. Poult Sci.;87:528–535.
Freitas, E.R, Sakomura, N.K Neme R, dos Santos A.L.(2002). Valores de energia metabolizável do óleo ácido de soja para aves. In: Anais da 39º Reunião Anual da Sociedade Brasileira de Zootecnia; Recife, PE.
Freitas, B.C.F.(1999) Digestibilidade da gordura nas primeiras semanas de vida e seu efeito
sobre o desempenho do frango de corte [dissertação]. Belo Horizonte: Escola de Veterinária, UFMG.
Friedman, A., and D. Sklan.(1989a). Antigen-specific immune response impairment in the
chick as influenced by dietary vitamin A. The Journal of Nutrition 119:790.
Friedman, A., and D. Sklan.(1989b). Impaired T lymphocyte immune response in vitamin A
depleted rats and chicks. Br. J. Nutr 62:439-449.
Friedman, H. I., and B. Nylund. (1980). Intestinal fat digestion, absorption, and transport. A review. The American journal of clinical nutrition 33:1108.
Fritts, C. A., and P. W. Waldroup. (2003). Effect of source and level of vitamin D on live performance and bone development in growing broilers. J Appl. Poult. Res. 12:45-52.
Gallardo, M.A, Perez, D.D and Leighton F.M.(2012). Modification of fatty acid composition in broiler chickens fed canola oil. Biol Res. 45:149–161.
Gérard P, Lepercq P, Leclerc M, Gavini F, Raibaud P, Juste C (2007). "Bacteroides sp. strain D8, the first cholesterol-reducing bacterium isolated from human feces". Appl. Environ. Microbiol. 73 (18): 5742–9. doi:10.1128/AEM.02806-06. PMC 2074900. PMID 17616613.
Ghazalah, A.A, Abd-Elsamee M.O, Ali A.M.(2008). Influence of dietary energy and poultry fat on the response of broiler chicks to heat therm. Int J. Poult Sci. 2;7:355–359.
Gibbons GF, Wiggins D, Brown AM, Hebbachi AM. (2004). "Synthesis and function of hepatic very-low-density lipoprotein.". Biochem Soc Trans. 32 (Pt 1): 59–64. doi:10.1042/bst0320059. PMID 14748713.
Golian A, and Polin D.(1984). Passage rate of feed in very young chicks. Poult. Sci.63:1013–1019.
Gonzales, M.J. and G.M. Pesti, (1993). Evaluation of the protein to energy ratio concept in broiler and turkey nutrition. Poult. Sci. J., 72:2115-2123.

63
Gous, R.M., M.Griessel and T.R. Morries, (1987). Effect of dietary energy concentration on the response of laying hens to amino acids, Br. Poult Sci., 28:427-436.
Griffiths, L, Lesson S, and Summers J.D.(1977). Influence of energy system and level of various fat sources on performance and carcass composition of broilers. Poultry
Science ; 56:1018-1026. Habib D.F, Fahmi A.A, Kholousy N.M, Amin A.I, Shalaby M, Ahmed M.M, Shanab A.M.
(2011). The role of liver in leptin metabolism in experimental nephrotic syndrome. EXCLI J;10:322-31.
Habib, A., Ramin S., and Alireza A. (2011). Effect of dietary supplemented canola and poultry fat on the performance and carcass characteristics of broiler chickens. Current Res. J. Biol. Sci., 3:388-392.
Han, Y. and D.H. Baher, (1993). Effects of sex, heat stress, body weight and genetic strain on
thr lysine requirement of broiler chicks. Poult. SCI., 72:701-708.
Hrdinka, C., Zollitsch W., Knaus W and Lettner F. (2002). Effects of dietary fatty acid pattern on melting point and composition of adipose tissues and intramuscular fat of broiler carcass. Poult. Sci., 75:208-215.
Hsu, P. C., and Y. L. Guo. (2002). Antioxidant nutrients and lead toxicity. Toxicology 180:33-44.
Huang, J., D. Yang, S. Gao, and T. Wang. (2008). Effects of soy-lecithin on lipid metabolism and hepatic expression of lipogenic genes in broiler chickens. Livestock Sci. 118:53-60.
Hurwitz, S., D. Sklan and I. Bartou, (1978). New formal approaches to the determination of energy and amino acid requirements of chicks. Poult. Sci., 57:197-205.
Iqbal, J., and M. M. Hussain. (2009). Intestinal lipid absorption. Am. J Physiol. Endocrinol.
Metab. 296:E1183-E1194.
Jacob, J. P., H. R. Wilson, R. D. Miles, G. D. Butcher, and F. B. Mather. (2011). Factors affecting egg production in backyard chicken flocks.
Jeffri D, Firman H, Kamyab A, (2010). Comparison of soybean oil with an animal/vegetable blend at four energy levels in broiler rations from hatch to market. Int Poult Sci.; 9:1027–1030.
Jensen, C., C. Lauridsen, and G. Bertelsen. (1998). Dietary vitamin E: quality and storage stability of pork and poultry. Trends Food Sci. Technol. 9:62-72.
Kemp, C., C. Fisher and M. Kenny,(2005). Genotype-nutrition interactions on broilers response to balanced protein in two commercial strains. Proceedings of the 15th European Symposium of Poultry Nutrition, Sept.25-29,Balatonfured, Hungary, pp: 54-56.

64
Ketels, E., G. Huyghebaert, and G. Groote. (1987). The nutritional value of commercial fat blends in broiler diets, 1: Effect of the incorporation level on the metabolizable energy content. Arch Geflugelkd 51.
Kumar, M., Bussmann, RW., Joshi, M., Kumar, P. (2011). Ethnomedicinal uses of plants close to rural habitation in Garhwal Himalaya, India. Journal of Medicinal Plants Research. 5(11), 2252-2260.
Lara, L.J.C. (2004). Efeito da fonte lipídica em dietas para frangos de corte sobre o desempenho, rendimento e composição da carcaça [dissertação]. Belo Horizonte: Escola de Veterinária, UFMG.
Latshaw, J.D.(2008). Daily energy intake of broiler chickens is altered by proximate nutrient
content and form of the diet. Poult Sci;87:89–95.
Lauridsen, C., D. J. Buckley, and P. A. Morrissey. (1997). Influence of dietary fat and vitamin E supplementation on [alpha]-tocopherol levels and fatty acid profiles in chicken muscle membranal fractions and on susceptibility to lipid peroxidation. Meat Sci. 46:9-22.
Leeson S, Summers J.D. (2001). Nutrition of the chicken. 4th ed. Ontario: University Books; 413p.
Leeson, S. and Atteh J.O. (2003). Ultilization of fats and fatty acids by turkey poults. Sci., 74.
Leeson, S.(2002). Is feed efficiency still a useful measure of broiler performance: Department of Animal and Poultry Science, University of Gvelp, Ministry of Agriculture, Food and Rural Affairs, Canada.
Leeson, S., and J. D. Summers. (1976). Fat ME values: the effect of fatty acid saturation. Feedstuffs 48:26-28.
Leeson, S., and J. D. Summers. (1997). Commercial Poultry Nutrition. 2nd ed. University Books, Guelph, ON, Canada.
Leeson, S., L. Caston and J.D. Summers. (1996). Broiler response to diet energy. Poult. Sci., 75:529-535.
Lewis, D., and C. G. Payne. (1966). Fats and amino acids in broiler rations. Br. Poult. Sci. 7:209-218.
Lipstein B, Budowski P, and Bornstein S.(1965). Effect of autoxidation on the nutritive value of acidulated soybean soapstock in chicks. Poultry Science; 44:1480-1488.

65
López-Ferrer S, Baucells MD, Barroeta AC and Grashorn MA. (1999). Influence of vegetable oil sources on quality parameters of broiler meat. Archiv für Geflügelkunde; 63(1):29-35.
López-ferrer S., Baucells M.D., Barroeta A.C., Galobart J., Grashorn M.A. (2001). N-3
Enrichment of Chicken meat.2. use or precursors of lay-chain polyunsaturated fatty acid: linseed oil. Poultry sci. 80, 753-761.
Mack, S., D. Bercovici, G. de Groote, B. Leclercq and M. Lppens et al., (1999). Ideal amino acid profile and dietary lysine specification for broiler chickens of 20-40 days of age Br. Poult. Sci., 40:257-265.
Mahagna M, Said N, Nir I, Nitsan Z.(1988) Development of digestibility of some nutrients and of energy utilization in young broiler chickens. In: Proceedings of the XVIII World´s Poultry Congress; Nagaya, JP. p.250-251.
Mano S, Quiroz M, Pardi H and Padilha A. (1999) Apostila de tecnologia de aves e
derivados. Rio de Janeiro:Faculdade de Veterinária, Universidade Federal Fluminense.
Marion JE, Woodroof JG. (1963). The fatty acid composition of breast, thigh and skin tissues
of chicken broilers as influenced by dietary fats. Poultry Science; 42:1202-1207. McDonald, P., Edwards, R.A., Green Halgh, J.F.D and Morgan, C.A. (1995). Animal
Nutrition. 5th Edition. Longman Scientific and Technical, N.Y. USA. 284-312. Menge, H., C. C. Calvert, and C. A. Denton. (1965). Further studies of the effect of linoleic
acid on reproduction in the hen. J Nutr. 86:115.
Meterological Center, Crop Science Department University of Nigeria Nsukka, 2009 unpublished. Miller D and Robisch P. (1969).Effect of herring, menhaden and safflower oils on the w-3
and w-6 fatty acid content of broiler tissue. Journal of Food Science; 34:136-141. Miller, E. C., H. Menge, and C. A. Denton. (1963). Effects of long-term feeding of a fat-free
diet to laying hens. J Nutr. 80:431-440.
Moav, R., (1995). Fat supplementation to poultry diet. Wrld Poult. Misset. 11, 57-58.
Mohammadi M, Lavvaf A, Hajian E, Jaberzadeh V. (2011). The effect of different levels of poultry by product meal on carcass quality in broiler. Res Opin Anim Vet Sci.1:697–699.
Moura B.H.S. (2003) Desempenho e composição da carcaça de frangos de corte alimentados com diferentes níveis energéticos com e sem óleo [dissertação]. Belo Horizonte: Escola de Veterinária, UFMG.

66
Mu, H., and C. E. Høy.( 2004). The digestion of dietary triacylglycerols. Prog. Lipid Res. 43:105-133.
Nahashan , S.N., N. Adefobe, A. Amenyeriu and D. Wright, (2001). Effect of varying metabolisable energy and crude protein concentrations in diets of pearl gray guinea fowl pullets 1. Growth performance. Poult. Sci. J., 85:1847-1854.
Nascif CCC, Gomes PC, Albino LFT, Rostagno HS.(2004) Determinação dos valores energéticos de alguns óleos e gorduras para pintos de corte machos e fêmeas aos 21 dias de idade. Revista Brasileira de Zootecnia; 33(2):375-385.
National Research Council. (1994) . Nutrient requirements of poultry. National Academic
Press; Washington, DC, USA.
Nelson, D.L and Micheal M. Cox (2000). Lehninger principles of biochemistry, 3rd edition. ISBN: 1-57259-9316. 1255 pp.
Nemavhola, Z., (2001). The effect of different diets on production performance and meat quality of the indigenous chickens. Masters of Agricultural Management (Animal production) Dissertation, University of Limpopo, South Africa.
Newcombe, M. and J.D Summers; (1984), effect of previous diet on feed intake and body weight gain of broiler and leghorn chicks. Poult. Sci J., 63: 1237.1242.
Newman,R.E, Bryden, W.L, Fleck, E, Ashes,J.R, Buttemer, W.A, Storlien, L.H, Downing, J.A. (2002). Dietary n-3 and n-6 fatty acids alter avian metabolism: molecular-species composition of breast-muscle phospholipids. Br J Nutr.88: 19–28.
Nobar, R.S., Gorbani A., Nazaradi K, Ayazi A., and Hamidiyan A (2010). Beneficial effects of canola oil on breast fatty acids profile and some biochemical parameters of Iranian native turkey. Journal of cell and animal biology vol 4(8) Pp 125-130.
Noy Y, Sklan D. (1995) Digestion and absortion in the young chick. Poultry Science 74(2):366-373.
Noy, Y., and D. Sklan. (1996). Uptake capacity in vitro for glucose and methionine and in
situ for oleic acid in the proximal small intestine of posthatch chicks. Poult. Sci. 75:998-1002.
NRC. 1994. Nutrient requirements of poultry. 9th Edn., National Academy press, Washington, DC., USA., ISBN-13: 9780309048927, pages:155.
Obioha, F.C. (1992). A Guide to poultry production in the tropics Acena Publishers, Enugu, Nigeria. Pp.8-15.
O'Doherty, P. J. A., G. Kakis, and A. Kuksis. 1973. Role of luminal lecithin in intestinal fat absorption. Lipids 8:249-255.

67
Ofomata, G.E.K. (1975). Soil Erosion. In: Nigeria in maps. Eastern States. Benin, Ethopia Pub. House.43-45.
Okonkwo, W.I. and Akubuo, C.O. (2007) Trombe wall system for poultry Brooding Int .J. Poultry sci. 6:125-130.
Olomu, J.M. (1995). Monogastric Animal Nutrition, Principles and practice. Pp.70-72.
Jachem publication, Benin. Oluyemi, J.A. and Roberts, F.A. (2007). Poultry production in warm-wet climate. Golden
wallet press, Ibadan. Pp. 12-19.
Ozdogan M. and M. Aksit (2003). Effect of feeds containing different fats on carcass and blood parameters of broilers. Poultry Science Association, Inc.
Palmquist, D.L.,(2002). An appraisal of fats and fatty acid. In: Poultry Feedstuff: Supply, Composition and Nutritive Value. Chapter 5, pp. 87-97.
Pardio, V.T, Landin, L.A, Waliszewski, K.N, Badillo C, Perez-gil F.(2001). The effect of
acidified soapstock on feed conversion and broiler skin pigmentation. Poultry Science; 80:1236-1239.
Peebles, E.D, Zumwalt, C.D, Doyle, S.M, Gerard, P.D, Latour, M.A, Boyle, C.R, Smith,
T.W. (2000) Effects of breeder age and dietary fat source and level on broiler hatching egg characteristics. Poult Sci;79:698–704.
Pesti G.N, Bakalli R.I, Qiao M, Sterling K.G. (2002). A comparison of eight grades of fats as broiler feed ingredients. Poult Sci;81:382–390.
Phetteplace, H.W and Watkins A.(1990) Lipid measurements in chickens fed different combinations of chicken fat and Menhaden oil. Journal Agricultural of Food
Chemistry ; 38:1848-1853. Pond, W.G., D.C. Church and K.R. Pond. (1995). Basic Animal Nutrition and feeding. 4th
Edn., John Wiley and Sons, New York, USA., pp:615
Racanicci, A.M.C, Menten, J.F.M, Regitano, D. Arce, M, Gaiotto, J.B, Longo F.A, Pedroso A.A, Sorbara J.O.B. (2000). Oxidação lipídica reduz o conteúdo de energia metabolizável do óleo de vísceras de aves para frangos de corte na fase de crescimento. In: Anais da 37º Reunião Anual da Sociedade Brasileira de Zootecnia; Viçosa:SBZ; CDROM.
Ranjhan, S.K.(1981). Animal nutrition in thetropics (Second revised edition). Vikas
publishing house. London. Pp. 65-73. Renner, R. and F. W. Hill. (1961a). Factors affecting the absorbability of saturated fatty acids
in the chick. J Nutr. 74:254.

68
Renner, R and Hill F.W. (1961b) Utilization of fatty acids by the chickenJournal of Nutrition; 74:259-264.
Richard,O,K and Church, D.C (1998). Livestock feeds and feeding. Fourth edition. Pretice
Hall, Upper Saddle River, New Jersey 07458. Pp 16-29.
Richards, M. P., 2003. Genetic regulation of feed intake and energy balance in poultry. Sci., 82:907-916.
Rita, T. dela Cruz of www.bar.gov.ph
Rizos, C.V. (2011). "Effects of Thyroid Dysfunction on Lipid Profile". The Open
Cardiovascular Medicine Journal 5 (1): 76–84. doi:10.2174/1874192401105010076.
Robinson L.J., Pang S., Harris D.S., Heuser J., James D.E. (1992) Translocation of the glucose transporter (GLUT4) to the cell surface in permeablized 3T3–L1 adipocytes: effects of ATP, insulin and GTPγS and localization of GLUT 4 to clathrin lattices. J.
Cell Biol. 117:1181–1196.
Rodriguéz M.A, Crespo, N.P, Cortés M, CREUS E.(2002), Medel P. Efecto del tipo de grasa de la dieta en la alimentacion del broiler, con enfasis en los productos derivados del aceite de palma. Selecciones avícolas ; 44(10):693-702.
Rosa F.C.(1999). Teor de ácidos graxos poliinsaturados ômega-3 no peito e coxa de frangos
de corte alimentados com rações contendo três fontes de óleo [dissertação]. Lavras: Universidade Federal de Lavras.
Rose, S. P. (2001). Principles of Poultry Science. CABI Publishing.
Sadeghi, 1 and S.A. Tabiedian (2005). Effect of different energy to protein ratio and tallow supplementation on broiler performance GH. International Journal of Poultry Science 4 (12): 976-981.
Sahin, K., N. Sahin, M. Onderci, S. Yaralioglu, and O. Kucuk. (2001). Protective role of supplemental vitamin E on lipid peroxidation, vitamins E, A and some mineral concentrations of broilers reared under heat stress. Veterinari Medicina Praha 46:140-144.
Sainbury, D., (1984). Poultry Health and Management. Granada publishing. Great Britian.
Sanz M, Flores A. and Lopez-Bote CJ. (2000a). The metabolic use of energy from dietary fat in broilers is affected by fatty acid saturation. British Poult Sci. 41:61–68.
Sanz M, Flores A, Perez DE, Ayala P, Lopez-Bote CJ.(1999). Higher lipid accumulation in broilers fed on saturated fats than in those fed unsaturated fats. British Poultry Science ; 40:95-101.

69
Sanz M, Lopez-Bote CJ, Menoyo D D and Bautista JM.(2000b) Abdominal fat deposition
and fatty acid synthesis are lower and β-oxidation is higher in broiler chickens fed diets containing unsaturated rather than saturated fat. J Nutr.130:3034–3037.
Scaife, J.R, Moyo, J, Galbraith H, Michie W, Campbell, V.(1994) Effect of different dietary supplemental fats and oils on the tissue fatty acid composition and growth of female broilers. British Poultry Science; 35:107-118.
Schmidt, D. R., S. R. Holmstrom, K. Fon Tacer, A. L. Bookout, S. A. Kliewer, and D. J.
Mangelsdorf.( 2010). Regulation of Bile Acid Synthesis by Fat-soluble Vitamins A and D. J Biol. Chem. 285:14486-14494.
Scott, M. L., M. C. Nesheim, and R. J. Young. (1982). Nutrition of the Chicken. 3rd ed. M. L. Scott and Associates. Ithaca, NY.
Scragg, R. H., N. B. Logan, and N. Geddes. (1987). Response of egg weight to the inclusion of various fats in layer diets. Br. Poult. Sci. 28:15-21.
Segrest J.P, Jones M.K, De Loof H, Dashti N (2001). "Structure of apolipoprotein B-100 in low density lipoproteins".Journal of Lipid Research 42 (9): 1346–67. PMID 11518754.
Shahryar, H.A, Salamatdoustnobar R, Lak A, Lotfi A.R.(2011). Effect of dietary supplemented canola oil and poultry fat on the performance and carcass characterizes of broiler chickens. Curr Res J Biol Sci.3:388–392.
Shannon, D. W. F., and W. O. Brown. (1969). Calorimetric studies on the effect of dietary energy source and environmental temperature on the metabolic efficiency of energy utilization by mature Light Sussex cockerels. J Agri. Sci. 72:479-489.
Shutze, J. V., and L. S. Jensen. (1963). Influence of linoleic acid on egg weight. Poult. Sci. 42:921-924.
Sibbald, I. R., J. D. Summers, and S. J. Slinger. (2000). Factors affecting the metabolizable energy content of poultry feeds. Poult. Sci. 39:544-556.
Sklan, D., D. Melamed, and A. Friedman. (1994). The effect of varying levels of dietary vitamin A on immune response in the chick. Poult. Sci. 73:843-847.
Smith, A.J. (2001). Poultry. The tropical Agriculture. 2nd (revised) ed. Macmillan education ltd. U.k. pp 1-73.
Snedecor, G.W and Cochran, W.G. (1980), Statistical methods, seventh edition, Ames: Lowa state University press: Pp 23-25.
Sorrentino M. J. (2011), "Non-HDL-Cholesterol", Hyperlipidemia in Primary Care, pp. 86–87, ISBN 9781603275026

70
Souza, P.A, Souza H.B.A, Oba A, Leonel F.R, Pelicano E.R.L, Norkus E.A, Junqueira O.M and Andreotti M.O.(2001) Características físicas e químicas da carne da coxa de frangos de corte produzidos com diferentes fonts de óleo. In: Anais da 38º Reunião Anual da Sociedade Brasileira de Zootecnia; Piracicaba, SP.
Stephens NA, Kieft R, Macleod A, Hajduk SL (2012). "Trypanosome resistance to human innate immunity: targeting Achilles' heel". Trends in Parasitology 28 (12): 539–45. doi:10.1016/j.pt.2012.09.002. PMID 23059119.
Surai, P. F., B. K. Speake, and N. H. C. Sparks. (2001). Carotenoids in avian nutrition and embryonic development. 1. Absorption, availability and levels in plasma and egg yolk. J Poult. Sci. 38:1-27.
Tabeidian, A., Sadeghi, G.H. and Pourreza, J., (2005). Effect of dietary protein levels and soybean oil supplementation on broiler performance. Int. J. Poult. Sci. 4, 799-803.
Thacker, P.A, Campbell, G.L and Xu, Y.(1994) Composition and nutritive value of acidulated fatty acids, degummed canola oil and tallow as energy sources for starting broiler chicks. Anim Feed Sci Technol. 46:251–260.
Thomas, O.P., A.I Zuckerman, M. Farran and C.B. Tampling, (1986). Updated amino acid requirements of broilers. Proceedings of the Maryland Nutrition Conference, (Muc’ 86), College park, MD, pp:79-85.
Tichivangana, J. Z., and P. A. Morrissey. (1985). Metmyoglobin and inorganic metals as pro-oxidants in raw and cooked muscle systems. Meat Sci. 15:107-116.
Toth, P.P (2005). "Cardiology patient page. The "good cholesterol": high-density lipoprotein". Circulation 111 (5): e89–e91. doi:10.1161/01.CIR.0000154555.07002.CA. PMID 15699268.
Valencia, .M.E, Watkins, S.E, Waldroup, A.L, Waldroup, P.W.(1993). Utilization of crude and refined palm and palm kernel oils in broiler diets. Poultry Science 1993; 72:2200-2215.
Veldkamp, T., R.P. Kwakkel, P.R. Ferket and M.W.A. Verstgen, (2005). Growth response to
dietary energy and lysine at high and low ambient temperature in male Turkets. Poult Sci. J., 84:273-282.
Verma, N.D., Panda, J.N., Singh, K.B. and Shrivastav, A.K., (1995). Effect of feeding cholesterol and fat on serum cholesterol of Japanese quail. Indian J. Poult. Sci. 30, 218-223.
Vieira, S.L, Ribeiro , A.M.L, Kessler, A.M, Fernandes LM, Ebert AR, Eichner G.(2002) Utilização da energia de dietas para frangos de corte formulados com óleo ácido de soja. Revista Brasileira Ciência Avícola ; 4(2):1-13.

71
Waldroup, P.W, Watkins S.E, Saleh, E.A.(1995). Comparison of 2 blended animal-vegetable fats having low or high free fatty acid content. J Appl Poult Res.4:41–48.
Westover, E.J, Covey, D.F, Brockman, H.L, Brown, R.E, Pike LJ (2003). "Cholesterol
depletion results in site-specific increases in epidermal growth factor receptor phosphorylation due to membrane level effects. Studies with cholesterol enantiomers". J. Biol. Chem. 278 (51): 51125–33. doi:10.1074/jbc.M304332200. PMC 2593805. PMID 14530278.
Wilson, K. J. & Bayer, R. S. (2000). Poultry Nutrition Information for Small Flock.
http://www.oznet.ksu.edu/library/lvstk2/ep80.pdf. Wiseman, J. (1984). Fats in animal nutrition. Butterworths, London, UK. Wiseman, J., and F. Salvador. (1991).The influence of free fatty acid content and degree of
saturation on the apparent metabolizable energy value of fats fed broilers. Poultry
Science; 70:573-582. Www.nejm.org. Yalçin, S, Ozkan S and Açikgoz Z.(1998). Influence of dietary energy on bird performance,
cacass parts yield and nutrient composition of breast meat of heterozygous naked neck broilers reared at natural optimum and summer temperatures. British Poultry
Science; 39(5):633-638. Young, R. J., and R. L. Garrett. (1963). Effect of oleic and linoleic acids on the absorption of
saturated fatty acids in the chick. J Nutr. 81:321. Yu,M.W., and Robinson, F.E. (1992). The application of short-term feed restriction to
broiler chicken production: A review: J.Appl. Poult. Res.1:147-153. Zelenka W, Knaus W, Aichinger F, Lettner F.(1997) Effects of different dietary fat sources
on performance and carcass characteristics of broilers. Animal Feed Science Tecnology; 66:63-73.
Zhang, Da-Wei; Garuti, Rita; Tang, Wan-Jin; Cohen, Jonathan C. and Hobbs, Helen H. (2008). "Structural requirements for PCSK9-mediated degradation of the low-density lipoprotein receptor". Proceedings of the National Academy of Sciences of the United States of America 105 (35): 13045–13050. doi:10.1073/pnas.0806312105. ISSN 0027-8424. PMC 2526098. PMID 18753623.
Zollitsch, W., W. Knaus, F. Aichinger, and F. Lettner. (1997). Effects of different dietary fat sources on performance and carcass characteristics of broilers. Anim. Feed Sci.
Technol. 66:63-73.

72
Zouari, N., F. Elgharbi, N. Fakhfakh, A. B. Bacha, Y. Gargouri, and N. Miled. (2010). Effect of dietary vitamin E supplementation on lipid and colour stability of chicken thigh meat. Afr. J. Biotechnol. 9:2276-2283.
Zubair, A.K. and S. leeson (1996). Compensatory growth in the broiler chicken: a review.
World’s Poult. Sci.J., 52:192-201.

73
APPENDIX 1
ANOVA Table on Growth performance of broilers fed diets containing different fat sources
Sum of Squares Df Mean Square F Sig.
TFI Between Groups 1037759.064 4 259439.766 3.758 .041
Within Groups 690351.921 10 69035.192
Total 1728110.986 14
DFI Between Groups 431.976 4 107.994 3.756 .041
Within Groups 287.556 10 28.756
Total 719.532 14
FBW Between Groups 492562.480 4 123140.620 3.786 .040
Within Groups 325216.294 10 32521.629
Total 817778.774 14
IBW Between Groups 262.571 4 65.643 .126 .970
Within Groups 5195.444 10 519.544
Total 5458.015 14
TWG Between Groups 502451.019 4 125612.755 3.280 .058
Within Groups 383018.797 10 38301.880
Total 885469.816 14
ADWG Between Groups 209.129 4 52.282 3.228 .060
Within Groups 161.968 10 16.197
Total 371.097 14
FCR Between Groups .283 4 .071 2.559 .104
Within Groups .276 10 .028
Total .559 14
PER Between Groups .153 4 .038 2.457 .114
Within Groups .156 10 .016
Total .309 14
FCKG Between Groups 3436.162 4 859.041 3.335 .062
Within Groups 2317.910 9 257.546
Total 5754.073 13

74
APPENDIX 2:
ANOVA Table on Carcass and Organ characteristics of broilers fed diets containing different fat sources
Sum of Squares Df Mean Square F Sig.
KIDN Between Groups 25.364 4 6.341 6.162 .036
Within Groups 5.145 5 1.029 Total 30.509 9
ABFAT Between Groups 1838.606 4 459.652 2.394 .182
Within Groups 960.175 5 192.035 Total 2798.781 9
SPLN Between Groups 6.997 4 1.749 1.424 .348
Within Groups 6.141 5 1.228 Total 13.139 9
PRVT Between Groups 26.226 4 6.557 2.902 .137
Within Groups 11.295 5 2.259 Total 37.521 9
LIVER Between Groups 462.004 4 115.501 2.108 .217
Within Groups 273.980 5 54.796 Total 735.984 9
DCWT Between Groups 1.456 4 .364 5.200 .050
Within Groups .350 5 .070 Total 1.806 9
DPCT Between Groups 252.907 4 63.227 10.457 .012
Within Groups 30.230 5 6.046 Total 283.137 9
HTW Between Groups 42.656 4 10.664 4.434 .067
Within Groups 12.025 5 2.405 Total 54.681 9
LW Between Groups 1.254 4 .313 3.453 .103
Within Groups .454 5 .091 Total 1.707 9
GIZ Between Groups 266.220 4 66.555 .915 .521
Within Groups 363.580 5 72.716 Total 629.800 9

75
APPENDIX 3:
ANOVA Table on Relative Organ weight of broilers fed diets containing different fat sources
Sum of Squares df Mean Square F Sig.
GIZ Between Groups 133.557 4 33.389 .950 .506
Within Groups 175.647 5 35.129 Total 309.203 9
HRT Between Groups .011 4 .003 2.891 .137
Within Groups .005 5 .001 Total .015 9
KID Between Groups .018 4 .004 7.716 .023
Within Groups .003 5 .001 Total .021 9
ABFAT Between Groups 1.283 4 .321 1.781 .270
Within Groups .900 5 .180 Total 2.183 9
SPLN Between Groups .003 4 .001 .535 .718
Within Groups .007 5 .001 Total .010 9
PVRT Between Groups .003 4 .001 .660 .646
Within Groups .006 5 .001 Total .009 9
LIVR Between Groups .183 4 .046 1.460 .339
Within Groups .157 5 .031
Total .340

76
APPENDIX 4:
ANOVA Table on Lipid profile of broilers fed diets containing different fat sources
Sum of Squares Df Mean Square F Sig.
TCHOL Between Groups 2.972 4 .743 4.469 .066
Within Groups .831 5 .166
Total 3.804 9
HDL Between Groups .166 4 .042 3.752 .090
Within Groups .055 5 .011
Total .221 9
LDL Between Groups 7.467 4 1.867 40.994 .001
Within Groups .228 5 .046
Total 7.695 9
TRGL Between Groups 1.239 4 .310 3.244 .114
Within Groups .477 5 .095
Total 1.716 9
VLDL Between Groups .317 4 .079 3.165 .119
Within Groups .125 5 .025
Total .443 9