Embed Size (px)
Citation preview

Aquaculture 358–359 (2012) 50–60
Contents lists available at SciVerse ScienceDirect
Aquaculture
j ourna l homepage: www.e lsev ie r .com/ locate /aqua-on l ine
Growth, metabolism and immune responses to evisceration andthe regeneration of viscera in sea cucumber, Apostichopus japonicus
Yuanqi Zang a, Xiangli Tian a,⁎, Shuanglin Dong a, Yunwei Dong b
a The Key Laboratory of Mariculture, Ministry of Education, Ocean University of China, Qingdao 266003, People's Republic of Chinab The State Key Laboratory of Marine Environmental Science, Xiamen University, Xiamen 361005, People's Republic of China
⁎ Corresponding author. Tel.: +86 532 8203 2117; faE-mail address: [email protected] (X. Tian).
0044-8486/$ – see front matter © 2012 Elsevier B.V. Alldoi:10.1016/j.aquaculture.2012.06.007
a b s t r a c t
a r t i c l e i n f oArticle history:Received 19 December 2011Received in revised form 11 April 2012Accepted 8 June 2012Available online 16 June 2012
Keywords:EviscerationRegenerationGrowth rateOxygen consumption rateImmune responsesApostichopus japonicus
Growth, metabolism and immune responses of sea cucumber, Apostichopus japonicus, following eviscerationinduced artificially, were investigated in this study. The body weight, specific growth rate (SGR), oxygen con-sumption rate (OCR), activities of immunoenzymes including acidic phosphatase (ACP), alkaline phosphatase(AKP) from hydrolytic system, and catalase (CAT), superoxide dismutase (SOD), total antioxidant capacity(T-AOC) and malonyl-dialdehyde (MDA) from antioxidant system in muscle, intestine and respiratory treewere measured to evaluate the physiological responses of A. japonicus to evisceration and regeneration of vis-cera. The results showed that body weight of sea cucumber significantly dropped following evisceration andthen increased gradually with digestive function resumed. Accelerated growth rates were observed in the re-generation group from the 10th day to 20th day, but there was no significant difference between the regen-eration group and control group at the end of the experiment. OCR reduced rapidly after evisceration andincreased gradually afterwards. Compared to the control, no significant difference in OCR was found on the45th day (P>0.05). The immune responses of A. japonicuswere highly tissue-specific during the regenerationof viscera. Both ACP and AKP activities in muscle peaked on the 10th day after evisceration, then decreasedgradually to normal on the 45th day (P>0.05), while those in the regenerated tissues exhibited a differentialrising trend from the 20th day after evisceration. SOD activity in muscle was not significantly influenced byevisceration; however, in the regenerated tissues it was enhanced and significantly higher than those in thecontrol on the 45th day (Pb0.01). CAT activity in muscle of the regeneration group was significantly higherthan in the control during the experiment (Pb0.05). Comparatively, CAT activity in the respiration tree in-creased invariably after evisceration, while in the intestine CAT activity was not significantly affected by evis-ceration (P>0.05). Significant increase was found in both T-AOC and MDA in all three tissues afterevisceration (Pb0.05); however, they all returned to normal at the end of the experiment. Results of thisstudy indicated that growth, metabolism and immune responses including hydrolytic and antioxidant en-zymes in sea cucumber were significantly influenced by evisceration. Although the regeneration group didnot catch up with the control in body weight, SGR, OCR, major non-specific immune parameters resumedto normal level within 45 days after evisceration, indicating that A. japonicus might have recovered physio-logically from evisceration. Results from this study first presented comprehensive and valuable data of phys-iological responses to evisceration and the regeneration of viscera in A. japonicus, which would haveimportant implications for those farmers engaged in sea cucumber culture.
© 2012 Elsevier B.V. All rights reserved.
1. Introduction
A phenomenon known in the representatives of holothurians such asApostichopus japonicus (Sun et al., 2011; Tan et al., 2008; Zheng et al.,2006), Holothuria glaberrima (Mashanov et al., 2012; Quiñones et al.,2002; Suárez-Castillo et al., 2004), etc., is spontaneous rejection of inter-nal organs, i.e., evisceration (Shukalyuk and Dolmatov, 2001). Eviscera-tion has been demonstrated in most cases as a response to artificial
x: +86 532 8203 2435.
rights reserved.
stimuli with fewer cases of its definite occurrence in natural conditionssuch as body injury, overcrowding or water pollution (Wilkie, 2001),and it has been recognized that regeneration occurs after these cases(Emson and Wilkie, 1980; Swan, 1961).
A wealth of information has been accumulated on the regenerationof tissues in several species of holothurians. Bertolini (1930) exploredthe cellular events that enable regeneration of the digestive tract andfirst accurately analyzed histological aspects of the regenerative pro-cess in Stichopus regalus. Dawbin (1949) studied the regeneration ofviscera in Stichopus mollis and Kille (1942) focused on the regenerationof reproductive system in Holothuria parvula. Over the last decade,more studies had been conducted to investigate the histological and

Table 1Chemical composition and energy content of the formulated diet.
Moisture (%) Protein (%) Dry matter (%) Lipid (%) Energy (kJ/g)
9.31±0.5 17.38±0.2 22.89±0.2 1.67±0.1 12.51±0.1
Values were given as mean±SE. Protein, lipid and dry matter were measured aspercentages on a dry matter basis.
51Y. Zang et al. / Aquaculture 358–359 (2012) 50–60
cellular aspects, including cell division, dedifferentiation, andmigrationwithin the intestine, body wall and other appendages (Dolmatov andGinanova, 2009; García-Arrarás et al., 2006; Murray and García-Arrarás, 2004; Sun and Zheng, 2005; Wang and Li, 2007; Zheng et al.,2006). More recently, the molecular events involved in regenerationhave also started to be uncovered (Mashanov et al., 2010, 2012; Sunet al., 2011). Particularly, an up-regulation expression of Wnt9, TCTP,and Bmp1/Tll in visceral regeneration of the sea cucumber H. glaberrimawas found, and it's the first attempt to map the expression domains ofWnt9 in a species outside the vertebrate lineage (Mashanov et al.,2012). However, to our knowledge, the responses of growth, metabo-lism and immune following the evisceration in holothurian specieshave not been fully studied until now.
The immune response in echinoderms includes the recognition offoreign materials (non-self), expulsion of non-self or rendering it in-nocuously, and wound healing (Nusetti et al., 2005; Ramírez-Gómezet al., 2008). These defense mechanisms are conducted by cellularand humoral immune responses, in which immune enzymes are com-monly involved as non-specific immunity. As hydrolytic enzymes, ACPand AKP participate in the degradation of foreign proteins, carbohy-drates and lipids (Ottaviani, 1984; Pipe et al., 1993; Xue and Renault,2000). Antioxidant system is important because reactive oxygen spe-cies (ROS) are continuously generated by a variety of cellular processes,such as superoxide anion (O2
−), hydrogen peroxide (H2O2), hydroxylradical (OH), and singlet oxygen (Roch, 1999). Antioxidant defensescan be measured by total antioxidant capacity (T-AOC). The key enzy-matic players include superoxide dismutase (SOD) and catalase (CAT),which detoxify O2
− and H2O2, respectively (Hermes-Lima et al., 1998).Terminal products includingMDA formed during the lipid peroxidationprocess which was induced by ROS, are very active and capable of,cross-linking of membrane proteins containing amino groups(Kehrer, 1993).
The sea cucumber, A. japonicus Selenka, is one of the most importantholothurian species cultured in China (Liao, 1980). In 2010 total pro-duction of the sea cucumbers was 13.03×104 t, within15.01×104 ha(Fisheries Bureau, Agriculture Department, China, 2011). However,average yield of A. japonicus was still very low (868.10 kg/ha or so in2010), due to limited knowledge of the species' eco-physiology Assame as other holothurian species, severe evisceration induced by var-ious natural and artificial factors commonly occurs during transport andculture for A. japonicus, causing highermortality and lower yield (Zhenget al., 2006). After expulsion of the internal organs, the growth of seacucumbers drops sharply, and due to loss of the respiratory tree, thebody wall will perform respiration for the whole animal (Choe, 1963).This could affect metabolism, as well as immune functions for differenttissues during the period of the regeneration of internal organ (Tan et
Fig. 1. Changes in body weight of A. japonicus during the regeneration of viscera. Each bar rvalues: *Pb0.05; **Pb0.01.
al., 2008). Regeneration of viscera in A. japonicus had significant effectson oxygen consumption rate and ammonia excretion, as well as bodyconsumption and energy content, as Tan et al. (2008) described.Ramírez-Gómez et al. (2008) reported that evisceration and the earlystages of regeneration in H. glaberrimawas characterized by tissue rup-ture and wound-healing, and the immune system of the animals wasprobably challenged to heal the wound and defend the organismagainst any microbial invasion. Therefore, it is important to investigatethe growth, metabolism and immune status following evisceration inA. japonicus to provide useful knowledge for farmers engaged in sea cu-cumber culture, including: (1) weight loss after evisceration, whichmight be principal aspects of commercial value and (2) immune status,which decides whether a special management should be conducted orimmunostimulants added and (3) the possible duration of regeneratingperiod.
The study was designed to investigate the growth, metabolismand immune status to evisceration and the regeneration of viscera inA. japonicus. Activities of acidic phosphatase (ACP), alkaline phospha-tase (AKP), catalase (CAT), superoxide dismutase (SOD), total antiox-idant capacity (T-AOC), and malonyl-dialdehyde (MDA) in muscles,intestines and respiratory trees were monitored as biomarkers ofthe regeneration.
2. Material and methods
2.1. Experimental animals and acclimation
Sea cucumbers were collected in May 2009 from the coastal cul-ture pond located in Jimo, Qingdao City, Shandong Province, P. R.China. The animals were cultured in a 1000-L fiberglass tank filledwith filtered seawater for 15 days to fully ensure thermal and envi-ronmental adaptation. During acclimation, the seawater replenisheddaily and maintained at 17 °C±0.5 °C and 28 to 30‰ salinity. The an-imals were fed to satiation once daily at 18:00 with a formulated feed(Sargassum spp., fish meal, sea mud, wheat, vitamin and mineral pre-mixes; 9.31±0.5% moisture, 17.38±0.2% protein, 22.89±0.2% drymatter, 1.67±0.1% lipid, 12.51±0.1 kJ g−1 energy; Table 1). The ex-periments were performed between 17 May 2009 and 30 June 2009.
epresents means±SE. Asterisks indicate significant differences with respect to control

Fig. 2. Changes in specific growth rate of A. japonicus during the regeneration of viscera.Each bar represents means±SE. Asterisks indicate significant differences with respectto control values: *Pb0.05; **Pb0.01, and NS indicates no significant difference.
52 Y. Zang et al. / Aquaculture 358–359 (2012) 50–60
2.2. Experimental design and management
After acclimation, 200 sea cucumbers of similar size (51.03±2.15 g) were arbitrarily collected and equally divided into the regener-ation and control groups. 100 sea cucumbers as one group were in-duced evisceration by injecting KCl 0.35 mol L−1 (2% body weight)into the coelom (Tan et al., 2008). One or two minutes later, sea cu-cumbers eviscerated and were randomly distributed into 20 rectangu-lar glass aquaria (55×30×35 cm, water volume of 45 L) with 5individuals per aquarium (regeneration group). The other 100 sea cu-cumbers, the control group, were not eviscerated. They were randomlydistributed into 20 aquaria with 5 individuals per aquarium as well.
Sea cucumbers in the regeneration group and the control werecultured with normal feed supply (feed over 5% of wet body weight)once a day (18:00 h) in aquaria during the period of the experiment.Uneaten food in the tank was collected 24 h after feeding bypipetting. Feces were collected twice daily by pipetting beforefeeding.
Seawater used in the experiment was filtered by a composite sandfilter, and 50% of the water in each aquarium was exchanged daily toensure good water quality. Aeration was provided continuously tomaintain dissolved oxygen above 6 mg L−1.
During the experiment, seawater temperature and salinity wascontrolled at 17±0.5 °C and 28–30‰ respectively. The ammoniawas less than 0.025 mg L−1 and pH ranged from 7.8 to 8.2. A simulat-ed natural photoperiod (14 h light:10 h dark) was used throughoutthe period of experiment.
Fig. 3. Changes in oxygen consumption rate of A. japonicus during the regeneration of viscerato control values: *Pb0.05; **Pb0.01, and NS indicates no significant difference.
2.3. Measurement of growth
Sea cucumbers in 5 aquaria for each treatment were assigned tomeasure the growth rate of animals. Sea cucumbers were weighedevery 5 days during the experiment. Before weighing, the animalswere fasted for 24 h to evacuate their guts, and then weighed as pre-viously described (Zhou et al., 2006).
2.4. Determination of oxygen consumption rate
Sea cucumbers in 5 aquaria for each treatment were used for theoxygen consumption rate determination (OCR). OCR was determinedevery 5 days for the regeneration and control groups during the ex-periment. Prior to the determination of oxygen consumption, sea cu-cumbers were starved for 24 h to reduce any associated metabolicresponses. In the regeneration and control groups, there were fivereplicates and one blank control to correct for the respiration of bac-teria in the water. The tested animals were put individually into a 1 Lconical flask. After a 4 h test period, the final level of dissolved oxygenin each chamber was determined using the Winkler method (Ji et al.,2008; Strickland and Parsons, 1968). Oxygen consumption rate (OCR)of sea cucumber was calculated with the following equation (Omoriand Ikeda, 1984):
OCR μg O2ð Þ � g−1 � h−1� ¼ DtVt−D0V0ð Þ=WTh
where, T, time duration (h); Dt, changes of the oxygen content (μg O2
L−1) before and after test in the test bottles; D0, changes of the oxy-gen content (μg O2 L−1) before and after test in the blank bottles;Vt, volumes of the test bottles (L); V0, volumes of the blank bottles(L); W, wet weight of the sea cucumber (g), determined as previouslydescribed (Zhou et al., 2006).
Regeneration procedure was considered finished when there wasno significant difference of OCR between the regeneration group andthe control group (Tan et al., 2008).
2.5. Tissue preparation and immune parameters assays
Sea cucumbers in 10 aquaria for each treatment were assigned tosample for immune parameters assays. Muscle, intestine and respira-tory tree samples were collected every 5 days, and five animals wererandomly sampled from 10 aquaria during the experiment. Musclewas removed from the posterior of the body. The whole intestinewas removed by an incision at the esophagus and cloaca. It was thencut longitudinally and washed thoroughly in ice-cold physiological
. Each bar represents means±SE. Asterisks indicate significant differences with respect

Fig. 4. Changes of ACP activity in muscle, intestine and respiratory tree of A. japonicus during the regeneration of viscera. Each bar represents means±SE. Asterisks indicate signif-icant differences with respect to control values: *Pb0.05; **Pb0.01, and NS indicates no significant difference.
53Y. Zang et al. / Aquaculture 358–359 (2012) 50–60
saline. After rinsing, the three tissues were blotted dry with filter paper,and each sample was frozen with liquid nitrogen in an Eppendorf tube(1.5 mL) and stored at−80 °C until analyzed.Muscle sampleswere col-lected throughout the experiment. Samples of intestine and respiratorytree in the regeneration group were collected from the 20th day afterevisceration due to unavailability of samples before then.
At each sampling interval, samples were thawed, weighed and ho-mogenized in 5 volumes of ice-cold physiological saline using a man-ual glass homogenizer. The homogenates were then centrifuged at10,000 g for 20 min at 4 °C. The supernatant was then pipetted intoclean centrifuge vials and stored at 4 °C until analyzed (less than12 h).
Protein content of the homogenates was measured following themethod of Spector (1978), using bovine serum albumin as the stan-dard. All assays for enzyme activities were carried out in duplicateand measured in the cuvette of a UV 2102PC spectrophotometer(Unico, Shanghai, China).
Acid and alkaline phosphatase (ACP and AKP) activity assays werecarried out according to Barrett's (1972) method using a commercialkit (Nanjing Jiancheng Biotech Company, China). Concentration ofphenol was measured spectrophotometrically at 520 nm after incu-bation at 37 °C for 30 min (for ACP), or 15 min (for AKP). ACP andAKP activity was defined as the amount of phenol (mol) producedper milligram protein.
Superoxide dismutase (SOD) activity was determined according toJi (1991) with an assay kit (Nanjing Jiancheng Biotech Company,China). Assay conditions were 65 μmol phosphate buffer, pH 7.8,
1 μmol hydrochloric hydroxylamine, 0.75 μmol xanthine and2.3×10−3 IU xanthine dismutase. 50 μL of the supernatant given noblank reaction were incubated in the system for 40 min at 37 °C,and terminated with 2 mL 3.3 g L−1 p-aminobenzene sulfonic acidand 10 g L−1 naphthylamine. An SOD unit is defined as the amountof enzyme that inhibits the superoxide-induced oxidation (monitoredat 550 nm) by 50%.
Catalase (CAT) activity was measured according to Góth (1991)with an assay kit (Nanjing Jiancheng Biotech Company, China). Thebase system including 4.60 μmol phosphate buffers, pH 7.4,6.5×10−3 μmol H2O2 was incubated 1 min at 37 °C. The reactionwas terminated immediately by 32.4 μmol ammonium molybdate. ACAT unit is defined as catalyzing the use of 1 μmol H2O2 per second.
Malondialdehyde (MDA) concentration was determined spectro-photometrically using the thiobarbituric acid method (Esterbauerand Cheeseman, 1990) with a commercial kit (Nanjing Jiancheng Bio-tech Company, China). The supernatant was added to an equal vol-ume of 1% thiobarbituric acid in a 90 °C water bath for 10 min. Aftercooling, thiobarbituric acid reactive substance (TBARS) levels wereestimated at 532 nm against a blank consisting of 5% cold trichloro-acetic acid mixed with 1% thiobarbituric acid. Malondialdehyde bisacetal was used as a standard. Concentrations of lipid peroxidationcompounds were expressed as nmol mg−1 protein.
The total antioxidant capacity (T-AOC) was measured by the col-orimetric technique as described by Miller et al. (1993) using a T-AOC commercial kit (Nanjing Jiancheng Biotech Company, China).These antioxidants reduce ferric ion (Fe3+) to ferrous ion (Fe2+).

Fig. 5. Changes of AKP activity in muscle, intestine and respiratory tree of A. japonicus during the regeneration of viscera. Each bar represents means±SE. Asterisks indicate signif-icant differences with respect to control values: *Pb0.05; **Pb0.01, and NS indicates no significant difference.
54 Y. Zang et al. / Aquaculture 358–359 (2012) 50–60
The latter combines with phenanthroline and produces a stable che-late, which can be measured spectrophotographically at 520 nm.The T-AOC was determined in units per mg of tissue protein.
2.6. Statistical analysis
Statistical analysis of data was performed with a statistical pack-age (SPSS 11.0 for windows, SPSS Inc., Richmond, CA, USA) Valueswere presented as means±standard error of the mean. Data for thebody weight, SGR, OCR and immune parameters were tested for ho-mogeneity of variances, then possible differences were comparedamong time intervals with one-way ANOVA followed by a Tukey'sHSD multiple comparison test Possible differences between the treat-ment and the control were tested by Student's t-test. Differenceswere considered significant or extremely significant among treat-ment groups at a probability level of Pb0.05 or Pb0.01, respectively.
3. Results
3.1. Growth
Body weight of A. japonicus was significantly affected by eviscer-ation (Fig. 1). Compared to the control group, body weight of ani-mals dropped sharply following evisceration and continued todecrease until the 10th day, then increased gradually as visceraregenerated. However, body weight of the regeneration group was
still significantly lower than the control group at the end of the ex-periment (Pb0.01).
The specific growth rate (SGR) of A. japonicus in regenerationgroup was much lower than the control by the 10th day after eviscer-ation (Pb0.01), then accelerated growth rates were observed fromthe 10th day to 20th day. This trend, however, dampened after the20th day. No significant difference was found between the regenera-tion group and the control from the 35th day to the end (P>0.05,Fig. 2).
3.2. Oxygen consumption rate
Oxygen consumption rates of A. japonicus were significantly af-fected by evisceration (Fig. 3). After evisceration, OCR of animalsdropped sharply, continue to decrease until the 5th day (2.50±0.43 μg O2·g−1·h−1) and then increased gradually until the 45thday (6.49±1.05 μg O2·g−1·h−1). No significant difference wasfound between the regeneration group and the control at the end ofthe experiment (P>0.05).
3.3. Immune parameters
Immune responses in muscle of A. japonicus were detected fromthe 1st day following evisceration to the end of the experiment,while those in intestine and respiratory tree were determined from

Fig. 6. Changes of SOD activity in muscle, intestine and respiratory tree of A. japonicus during the regeneration of viscera. Each bar represents means±SE. Asterisks indicate signif-icant differences with respect to control values: *Pb0.05; **Pb0.01, and NS indicates no significant difference.
55Y. Zang et al. / Aquaculture 358–359 (2012) 50–60
20th day after evisceration when those tissues regenerated sufficient-ly for detection .
3.3.1. Acidic phosphatase activitiesACP activity in different tissues of A. japonicus in the control was
highly tissue-specific and ranged from 10.20±0.82 to 14.63±0.64 U/g prot, 52.81±2.31 to 583.85±0.88 U/g prot and 71.96±2.31 to 90.33±2.01 U/g prot in muscle, intestine and respiratory tree,respectively (Fig. 4). ACP activity in the respiratory tree tissue was sig-nificantly higher than in intestine tissue, andmuscle tissue had the low-est ACP activity (Pb0.05). With evisceration, ACP activity in musclehighly significantly increased compared to the control (Pb0.01). Themaximum value occurred on the 10th day (22.41±0.431 U/g prot),then it decreased gradually until the 45th day. No significant differencewas found at the end of experiment (P>0.05); however, for theregenerated tissues, ACP activity in both intestine and respiratory treeexhibited a rising trend totally from the 20th day after evisceration.ACP activity in intestine was significantly higher than in the controlfrom the 30th day to the end of the experiment (Pb0.05), while thatin the respiratory treewas significantly lower than in the control excepton the 45th day (72.03±3.06 U/g prot, Pb0.05).
3.3.2. Alkaline phosphatase activitiesValues of AKP activity in muscle, intestine and respiratory tree
for the control ranged from 11.40±0.75 to 20.49±1.21 U/g prot,
88.26±3.20 to 104.08±2.81 U/g prot and 33.07±0.78 to 37.35±0.94 U/g prot, respectively (Fig. 5). AKP activity in intestine ofA. japonicus was the highest among the three different tissues, andthat in respiratory tree was significantly higher than in muscle(Pb0.05). Compared to the control, AKP activity inmuscle of the regener-ation group significantly increased after evisceration, and the maximumvalue (39.43±0.75 U/g prot) occurred on the 10th day. Furthermore,AKP activity decreased gradually, and no significant difference wasfound after the 30th day. AKP activity was significantly lower in theregenerated intestine than in the control on the 20th day after eviscera-tion (Pb0.01), then increased gradually. No significant difference wasfound between the regeneration group and the control at the end ofexperiment (P>0.05). In the regenerated respiratory tree, AKP activityfluctuated from the 20th day after evisceration. However, no significantdifference was found between the regeneration group and the controlgroup throughout the experiment (P>0.05).
3.3.3. Superoxide dismutase activitiesSOD activity in muscle of A. japonicus fluctuated after evisceration
(Fig. 6). During the regeneration of the viscera, there was no significantdifference between the regeneration group and the control group(P>0.05). For both regenerated intestine and respiratory tree, SOD ac-tivities were significantly higher than in the control (Pb0.01) on the20th day after evisceration, and decreased gradually thereafter.
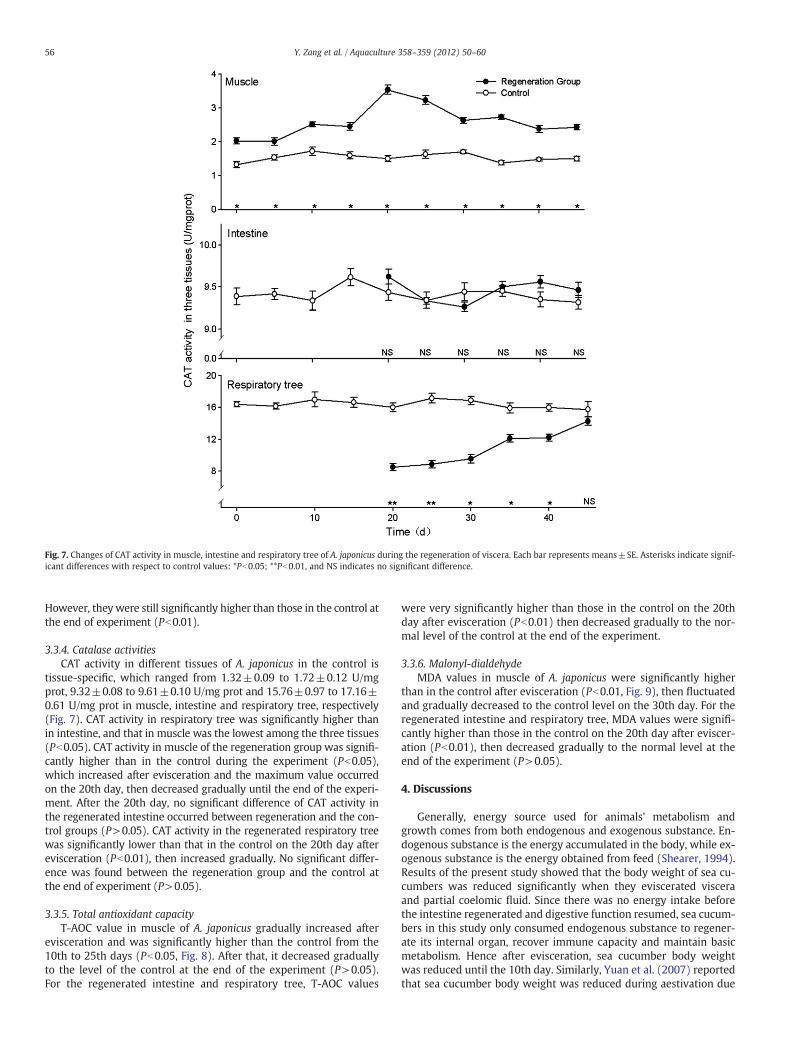
Fig. 7. Changes of CAT activity in muscle, intestine and respiratory tree of A. japonicus during the regeneration of viscera. Each bar represents means±SE. Asterisks indicate signif-icant differences with respect to control values: *Pb0.05; **Pb0.01, and NS indicates no significant difference.
56 Y. Zang et al. / Aquaculture 358–359 (2012) 50–60
However, theywere still significantly higher than those in the control atthe end of experiment (Pb0.01).
3.3.4. Catalase activitiesCAT activity in different tissues of A. japonicus in the control is
tissue-specific, which ranged from 1.32±0.09 to 1.72±0.12 U/mgprot, 9.32±0.08 to 9.61±0.10 U/mg prot and 15.76±0.97 to 17.16±0.61 U/mg prot in muscle, intestine and respiratory tree, respectively(Fig. 7). CAT activity in respiratory tree was significantly higher thanin intestine, and that in muscle was the lowest among the three tissues(Pb0.05). CAT activity in muscle of the regeneration group was signifi-cantly higher than in the control during the experiment (Pb0.05),which increased after evisceration and the maximum value occurredon the 20th day, then decreased gradually until the end of the experi-ment. After the 20th day, no significant difference of CAT activity inthe regenerated intestine occurred between regeneration and the con-trol groups (P>0.05). CAT activity in the regenerated respiratory treewas significantly lower than that in the control on the 20th day afterevisceration (Pb0.01), then increased gradually. No significant differ-ence was found between the regeneration group and the control atthe end of experiment (P>0.05).
3.3.5. Total antioxidant capacityT-AOC value in muscle of A. japonicus gradually increased after
evisceration and was significantly higher than the control from the10th to 25th days (Pb0.05, Fig. 8). After that, it decreased graduallyto the level of the control at the end of the experiment (P>0.05).For the regenerated intestine and respiratory tree, T-AOC values
were very significantly higher than those in the control on the 20thday after evisceration (Pb0.01) then decreased gradually to the nor-mal level of the control at the end of the experiment.
3.3.6. Malonyl-dialdehydeMDA values in muscle of A. japonicus were significantly higher
than in the control after evisceration (Pb0.01, Fig. 9), then fluctuatedand gradually decreased to the control level on the 30th day. For theregenerated intestine and respiratory tree, MDA values were signifi-cantly higher than those in the control on the 20th day after eviscer-ation (Pb0.01), then decreased gradually to the normal level at theend of the experiment (P>0.05).
4. Discussions
Generally, energy source used for animals' metabolism andgrowth comes from both endogenous and exogenous substance. En-dogenous substance is the energy accumulated in the body, while ex-ogenous substance is the energy obtained from feed (Shearer, 1994).Results of the present study showed that the body weight of sea cu-cumbers was reduced significantly when they eviscerated visceraand partial coelomic fluid. Since there was no energy intake beforethe intestine regenerated and digestive function resumed, sea cucum-bers in this study only consumed endogenous substance to regener-ate its internal organ, recover immune capacity and maintain basicmetabolism. Hence after evisceration, sea cucumber body weightwas reduced until the 10th day. Similarly, Yuan et al. (2007) reportedthat sea cucumber body weight was reduced during aestivation due

Fig. 8. Changes of T-AOC activity in muscle, intestine and respiratory tree of A. japonicus during the regeneration of viscera. Each bar represents means±SE. Asterisks indicate sig-nificant differences with respect to control values: *Pb0.05; **Pb0.01, and NS indicates no significant difference.
57Y. Zang et al. / Aquaculture 358–359 (2012) 50–60
to absence of digestive function and consumption of endogenous sub-stance. A previous study reported that it takes about two weeks forA. japonicus to regenerate a new intestinewith partial digestive function(Zheng et al., 2006). Likewise sea cucumber in this study regained partialability of ingestion and digestion on the 15th day and then altered fromconsuming solely endogenous substance to the combination of endoge-nous and exogenous substance. In this study, the body weight of a typi-cal sea cucumber increased gradually after the 15th day of evisceration.Although accelerated growth rates were observed in the regenerationgroup from the 10th day to 20th day, their final body weight did notequal or surpass the control's body weight on the 45th day afterevisceration.
Respiratory tree is the primary respiratory organ of the sea cu-cumber; cutaneous respiration is the other major form of respiration(Choe, 1963). Before regeneration, sea cucumbers obtain oxygen viacutaneous respiration to maintain their energy metabolism. However,as shown in a previous study (Tan et al., 2008), the accessory respira-tory organ cannot provide sufficient oxygen intake, and as a result,OCR reduced rapidly after evisceration. Afterwards, sea cucumberOCR increased gradually with new respiratory tree regenerated andpartial function resumed. On the 45th day after evisceration, whensea cucumber regenerated their respiratory tree, no significant differ-ence in OCR was found between the regeneration and control group,similar to result from a previous study (Zheng et al., 2006).
The health of animals might be evaluated by sensitiveimmunomarkers. The ideal immunomarker indicates not only thehealth of the animals but also the degree of environmental stress onimmune systems (Wang et al., 2008b). Susceptibility to disease may
increase if immunomarkers are below the normal standard. Eviscera-tion of sea cucumber is a protective behavior to cope with severe en-vironmental challenge, and sea cucumbers enter a regeneration stageafter evisceration (Fan et al., 2007; Zheng et al., 2006). Similar to mol-lusks, sea cucumbers cope with foreign materials via both cellular andhumoral mechanisms in which many non-specific, defense-relatedparameters are involved (Dyrynda et al., 1998; Renault et al., 2001;Wootton and Pipe, 2003). Thus, major immunoenzyme activities in-cluding ACP, AKP from hydrolytic system and SOD, CAT, MDA, T-AOC from antioxidant system, of sea cucumber could be monitoredto evaluate the immunity status during regeneration (Wang et al.,2008a, 2008b).
ACP is a phosphatase, a type of enzyme, used to free attached phos-phate groups from other molecules during digestion. It is also stored inlysosomes and functions when these fuse with endosomes (Cajaravilleet al., 2000; Rajalakshmi and Mohandas, 2005). AKP is a meta-lloenzyme, which catalyzes the nonspecific hydrolysis of phosphatemonoesters (Zhang et al., 2004). When exposed to a variety of environ-mental stress, as lysosomal enzymes, ACP and AKP also participate indegradation of foreign proteins, carbohydrates and lipids (Ottaviani,1984; Pipe et al., 1993; Xue and Renault, 2000). In the present study,evisceration significantly affected activities of hydrolases in sea cucum-ber. A significant increase of ACP activity in muscle was observed on the10th day after evisceration, and this is consistent with the increase ofAKP activity. The mutual increases in both ACP and AKP activities sug-gest not only enhancement of the capacity of degradation and defenseto foreign materials, but also improvement of metabolic intensity toprovide more energy for the regeneration of the viscera in sea

Fig. 9. Changes of MDA activity in muscle, intestine and respiratory tree of A. japonicus during the regeneration of viscera. Each bar represents means±SE. Asterisks indicate sig-nificant differences with respect to control values: *Pb0.05; **Pb0.01, and NS indicates no significant difference.
58 Y. Zang et al. / Aquaculture 358–359 (2012) 50–60
cucumber. As regeneration advanced, ACP and AKP activities in all threetissues returned to normal, and no significant difference was found be-tween the regeneration and control groups on the 45th day after evis-ceration except for ACP activity in intestine, which was significantlyhigher than the control probably due to the enhanced digestion in theregenerating intestine.
The adaptation of organisms to environmental stress depends on itsantioxidant defense, and those species with lower antioxidant efficien-cymay be less tolerant of conditions affecting oxidative damage (Regoliet al., 2004). Because oxygen radicals are continuously generated by avariety of cellular processes, all organisms have evolved antioxidant de-fenses which can be measured by total antioxidant capacity (T-AOC)with both enzymatic and non-enzymatic components. Enzymatic de-fenses include superoxide dismutase (SOD) and catalase (CAT), whichdetoxify O2
− and H2O2, respectively (Hermes-Lima et al., 1998). MDAis a measure of terminal products of lipid hydroperoxides, which re-flects oxyradicals stress on organisms. In the present study, T-AOCvalue in muscle increased and peaked on the 20th day after eviscera-tion, and those in the regenerated tissues elevated significantly aswell. SOD activities in the regenerated tissues significantly enhancedafter evisceration until the 45th day. During the whole period of the ex-periment, CAT activities in muscles of the regeneration group were sig-nificantly higher than in the control, which ascended after eviscerationand peaked on the 20th day. Correspondingly, a significant increase inMDA value for both muscle and regenerated tissues was observed onthe 20th day after evisceration. More interestingly, there is a clear op-posite trend in the response of CAT and SOD in the respiratory tree. Alow-level but enhancing CAT activity in the regenerating respiratorytree was detected, and returned to normal on the 45th day.
Comparatively, SOD activity in the respiratory tree significantly en-hanced on the 20th day after evisceration, then decreased gradually,but still significantly higher than that in the control at the end. As weknow, SOD catalyzes the conversion of O2
− into H2O2, while catalyzesthe decomposition of H2O2 to water and oxygen, which usually are animportant antioxidant defense in nearly all cells exposed to oxygen(Hermes-Lima et al., 1998). However, the invertebrate immune systemwas more than a collection of simple innate responses (H. glaberrima,Ramírez-Gómez et al., 2008). It has been revealed that H2O2 also canbe catalyzed by other peroxidases, such as glutathione peroxidase(GPX; Hermes-Lima et al., 1998). In the present study, the increasingCAT activity indicated that CAT might participated in parts of H2O2 de-toxification in the regenerating respiratory tree, however, if there wereany other specific peroxidases involved in the regeneration of the respi-ratory tree and their possible roles was still unknown, which deservedfurther research in the future.
The immune response to evisceration and the regeneration of vis-cera appeared to be an adaptation allowing the sea cucumber to man-age oxidative stress during regeneration. With regeneration of therespiration tree, OCR of sea cucumber increased to normality at the45th day. Thus, the sea cucumber displayed coordinate changes in anti-oxidant defenses including an increase before the 20th day and the de-crease afterwards mitigating oxidative stress that occurred as part ofnatural cycles of regeneration. With the MDA and T-AOC activities ret-urned to normal on the 45th day, the mission of the antioxidant de-fenses during regeneration were completed. Unfortunately, themolecular mechanisms that trigger and regulate changes in antioxidantenzyme activities in sea cucumbers are still unknown but could proveto have key relevance to regeneration of new tissues. In the other

59Y. Zang et al. / Aquaculture 358–359 (2012) 50–60
animals, the oxidative stress could play an important role in the onsetand progress of a reduced capacity for wound healing and tissue regen-eration, and an up-regulating of antioxidant enzymes were alsodetected during this stage in the polychaete Eurythoe complanata(Nusetti et al., 2005), which was similar with the present study.
The immune responses of A. japonicus to evisceration were tissue-specific and changed in different temporal patterns during the regener-ation of the viscera. Results from the present study demonstrated thatnon-specific immune parameters in muscle significantly enhancedafter evisceration generally. However, those in the regenerated intes-tine and respiratory tree showed different trends. SOD activity, T-AOCand MDA values were significantly higher than the control when mea-sured on the 20th day, while ACP activity in respiratory tree and AKPactivity in intestine were significantly lower, and no significant differ-ence was found in AKP activity for respiratory tree and CAT for intes-tine. On the 45th day after evisceration, if taking changes in SGR andOCR into account together, most immune parameters returned in dif-ferential temporal patterns to normal levels, which might indicatethat A. japonicus recovered to normal physiology from evisceration..
Previous studies showed that new intestine functioned for digestionon the 21th day in Holothuria glaberrlma (García-Arrarás et al., 1998)and on the 14th day in A. japonicus after evisceration (Zheng et al.,2006), respectively. However, at least 35 days are required for thenew intestine to return to normal (García-Arrarás et al., 1998; Zhenget al., 2006). Similar to results from Zheng et al. (2006), in the presentstudy, it was observed that new intestine resumed its normal digestivefunction on the 15th day and indicated that the first two or three weeksafter evisceration might be a critical period for the regeneration of theviscera in A. japonicus. This may have practical implications for sea cu-cumber farmers and suggests that management should eliminate possi-ble environmental stressors in the first two to three weeks afterevisceration. On the other hand, immunostimulants are suggestedadded during the regenerating course, such as lipopolysaccharides, asdescribed in H. glaberrima (Santiago-Cardona et al., 2003).
In conclusion, the results present valuable data regarding thegrowth, metabolism and non-specific immune parameters in responsesto evisceration and regeneration of viscera in A. japonicus. Although theeviscerated sea cucumbers did not catch up with the controls in bodyweight, their SGR, OCR and major non-specific immune parameters re-sumed to normal levels implying physiological recovery within 45 daysafter evisceration.
Acknowledgments
We thank Mr. William Bill Trimble and Dr. Li Li in the Departmentof Fisheries and Allied Aquaculture, Auburn University for their helpin preparing the paper. This research was supported by the NaturalScience Funds for Distinguished Young Scientists of Shandong Prov-ince (grant no. JQ201009), the Great Project of Scientific and Techni-cal Supporting Programs (grant no. 2011BAD13B03), the NationalNatural Science Foundation of China (grant no. 30771661), the PublicScience and Technology Research Funds Projects of Ocean, State Oce-anic Administration of the People's Republic of China (grant no.200905020), the Program for New Century Excellent Talents in Uni-versity (grant no. NCET-08-0503), and the 111 Project, China Ministryof Education (grant no. B08049).
References
Barrett, A.J., 1972. Lysosomal enzymes. In: Dingle, J.T. (Ed.), Lysosomes: A LaboratoryHandbook. North-Holland, Amsterdam, pp. 46–135.
Bertolini, F., 1930. Rigenerazione dell'apparato digerente nello Stichopus regalus.Pubblicazioni Stazione Zoologica di Napoli 10, 439–448.
Cajaraville, M.P., Bebianno, M.J., Blasco, J., Porte, C., Sarasquete, C., Viarengo, A., 2000.The use of biomarkers to assess the impact of pollution in coastal environmentsof the Iberian Peninsula: a practical approach. The Science of the Total Environ-ment 247, 295–311.
Choe, S., 1963. Biology of the Japanese Common Sea Cucumber Stichopus japonicusSelenka. National University, Pussan.
Dawbin, W.H., 1949. Auto-evisceration and the regeneration of viscera in the holothu-rian Stichopus mollis (Hutton). Transactions of the Royal Society of New Zealand 77,497–523.
Dolmatov, I.Y., Ginanova, T.T., 2009. Post-autotomy regeneration of respiratory trees inthe holothurian Apostichopus japonicus (Holothuroidea, Aspidochirotida). Cell andTissue Research 336, 41–58.
Dyrynda, E.A., Pipe, R.K., Burt, G.R., Ratcli, N.A., 1998. Modulations in the immune de-fences of mussels (Mytilus edulis) from contaminated sites in the UK. Aquatic Tox-icology 42, 169–185.
Emson, R.H., Wilkie, I.C., 1980. Fission and autotomy in echinoderms. Oceanographyand Marine Biology: An Annual Review 18, 155–250.
Esterbauer, H., Cheeseman, K.H., 1990. Determination of aldehydic lipid peroxidationproducts: malonaldehyde and 4-hydroxynonenal. Methods in Enzymology 186,407–421.
Fan, T.J., Du, Y.T., Cong, R.S., 2007. Research advances in echinoderm regeneration. Jour-nal of Ocean University of China 37 (4), 563–568 (in Chinese with Englishabstract).
Fisheries Bureau, Agriculture Department, China, 2011. China Fisheries Statistical Year-book. China Agricultural Press, Beijing.
García-Arrarás, J.E., Lourdes, S., Roberto, I.T., Irma, D.M., Lucy, T.A., Ilia, 1998. Cellularmechanisms of intestine regeneration in the sea cucumber, Holothuria glaberrlmaSalenka (Holothuroidea: Echinodermata). The Journal of Experimental Zoology281, 288–304.
García-Arrarás, J.E., Schenk, C., Rodrigues-Ramirez, R., Torres, I.I., Valentin, G.,Candelaria, A.G., 2006. Spherulocytes in the echinoderm Holothuria glaberrimaand their involvement in intestinal regeneration. Developmental Dynamics 235,3259–3267.
Góth, L., 1991. A simple method for determination of serum catalase activity and revi-sion of reference range. Clinica Chimica Acta 196, 143–151.
Hermes-Lima, M., Storey, J.M., Storey, K.B., 1998. Antioxidant defenses and metabolicdepression. The hypothesis of preparation for oxidative stress in land snails. Com-parative Biochemistry and Physiology. B 120, 437–448.
Ji, J.P., 1991. An ultramicroanalytic and rapid method for determination of superoxidedismutase activity. Journal of Nanjing Railway Medical College 10, 27–30 (in Chi-nese with English abstract).
Ji, T.T., Dong, Y.W., Dong, S.L., 2008. Growth and physiological responses in the sea cu-cumber, Apostichopus japonicus Selenka: aestivation and temperature. Aquaculture283, 180–187.
Kehrer, J.P., 1993. Free radicals as mediators of tissue injury and disease. Critical Re-views in Toxicology 23, 2148.
Kille, F.R., 1942. Regeneration of the reproductive system following binary fission in thesea-cucumber Holothuria parvula (Selenka). The Biological Bulletin 83, 55–66.
Liao, Y., 1980. The aspidochirote holothurians of China with erection of a new genus inechinoderms, present and past. In: Jangoux, M. (Ed.), Proceeding of European Col-loquium on Echinoderm. A.A Balkema Publishers, Rotterdam, Netherlands, pp.115–120.
Mashanov, V.S., Zueva, O.R., Rojas-Catagena, C., García-Arrarás, J.E., 2010. Visceral re-generation in a sea cucumber involves extensive expression of survivin and mor-talin homologs in the mesothelium. BMC Developmental Biology 10, 117.
Mashanov, V.S., Zueva, O.R., García-Arrarás, J.E., 2012. Expression of Wnt9, TCTP, andBmp1/Tll in sea cucumber visceral regeneration. Gene Expression Patterns 12,24–35.
Miller, N.J., Rice-Evans, C., Davies, M.J., Gopinathan, V., Milner, A., 1993. A novel methodfor measuring antioxidant capacity and its application to monitoring the antioxi-dant status in premature neonates. Clinical Science 84, 407–412.
Murray, G., García-Arrarás, J.E., 2004. Myogenesis during holothurian intestinal regen-eration. Cell and Tissue Research 318, 515–524.
Nusetti, O., Zapata-Vívenes, E., Esclapés, M.M., Rojas, A., 2005. Antioxidant enzymesand tissue regeneration in Eurythoe complanata (Polychaeta: Amphinomidae) ex-posed to used vehicle crankcase oil. Archives of Environmental Contaminationand Toxicology 48, 509–514.
Omori, M., Ikeda, T., 1984. Methods in Marine Zooplankton Ecology. John Wiley andSons Inc, New York, pp. 173–209.
Ottaviani, E., 1984. Composition of the serum haemolymph of Planorbis corneus (Gas-tropoda, Pulmonata). Comparative Biochemistry and Physiology. B 78B, 227–239.
Pipe, R.K., Porte, C., Livingstone, D.R., 1993. Antioxidant enzymes associated with theblood cells and hemolymph of the mussels Mytilus edulis. Fish & Shellfish Immu-nology 3, 221–233.
Quiñones, J.L., Rosa, R., Ruiz, D.L., García-Arrarás, J.E., 2002. Extracellular matrix remod-eling and metalloproteinase involvement during intestine regeneration in the seacucumber Holothuria glaberrima. Developmental Biology 250, 181–197.
Rajalakshmi, S., Mohandas, A., 2005. Copper-induced changes in tissue enzyme activityin a freshwater mussel. Ecotoxicology and Environmental Safety 62, 140–143.
Ramírez-Gómez, F., Ortiz-Pineda, P.A., Rojas-Cartagena, C., Suárez-Castillo, E.C., García-Arrarás, J.E., 2008. Immune-related genes associated with intestinal tissue in thesea cucumber Holothuria glaberrima. Immunogenetics 60, 57–71.
Regoli, F., Cerrano, C., Chierici, E., 2004. Seasonal variability of prooxidant pressure andantioxidant adaptation to symbiosis in the Mediterranean demosponge Petrosiaficiformis. Marine Ecology Progress Series 275, 129–137.
Renault, T., Xue, Q.G., Chilmonczyk, S., 2001. Flow cytometric analysis of European atoyster, Ostrea edulis, haemocytes using a monoclonal antibody specific forgranulocytes. Fish & Shellfish Immunology 11, 269–274.
Roch, P., 1999. Defense mechanisms and disease prevention in farmed marine inverte-brate. Aquaculture 172, 125–145.

60 Y. Zang et al. / Aquaculture 358–359 (2012) 50–60
Santiago-Cardona, P.G., Berríos, C.A., Ramírez, F., García-Arrarás, J.E., 2003. Lipopoly-saccharides induce intestinal serum amyloid A expression in the sea cucumberHolothuria glaberrima. Developmental and Comparative Immunology 27, 105–110.
Shearer, K.D., 1994. Factors affecting the proximate composition of cultured fishes withemphasis on salmonids. Aquaculture 119, 63–88.
Shukalyuk, A.I., Dolmatov, I.Yu., 2001. Regeneration of the digestive tube in the Holo-thurian Apostichopus japonicus after evisceration. Russian Journal of Marine Biolo-gy 27, 168–173.
Spector, T., 1978. Refinement of the coomassie blue method of protein quantitation.Analytical Biochemistry 86, 142–146.
Strickland, J.D.H., Parsons, T.R., 1968. A practical handbook of seawater analysis. Fisher-ies Research Board of Canada Bulletin 167, 1–11.
Suárez-Castillo, E.C., Medina-Ortíz, W.E., Roig-López, J.L., García-Arrarás, J.E., 2004.Ependymin, a gene involved in regeneration and neuroplasticity in vertebrates, isoverexpressed during regeneration in the echinoderm Holothuria glaberrima.Gene 334, 133–143.
Sun, X.Q., Zheng, F.X., 2005. Evisceration and regeneration in the holothurian. Journalof Ocean University of China 35 (5), 719–725 (in Chinese with English abstract).
Sun, L.N., Chen, M.Y., Yang, H.S., Wang, T.M., Liu, B.Z., 2011. Large scale gene expressionprofiling during intestine and body wall regeneration in the sea cucumber Apo-stichopus japonicus. Comparative Biochemistry and Physiology Part D 6, 195–205.
Swan, E.F., 1961. Seasonal evisceration in the sea cucumber Parastichopus californicus.Science 133, 1078.
Tan, Y.K., Li, X., Duan, J.J., 2008. Energy metabolism and biochemical composition ofApostichopus japonicus during the regeneration of viscera. Journal of Fishery Sci-ences of China 15 (4), 683–688 (in Chinese with English abstract).
Wang, X., Li, X., 2007. The morphological and histological observation of regenerationof alimentary tract in sea cucumber Apostichopus japonicus. Journal of Dalian Fish-eries University 22, 340–346.
Wang, F.Y., Yang, H.S., Gabr, H.R., Gao, F., 2008a. Immune condition of Apostichopusjaponicus during aestivation. Aquaculture 285, 238–243.
Wang, F.Y., Yang, H.S., Gao, F., Liu, G.B., 2008b. Effects of acute temperature or salinitystress on the immune response in sea cucumber, Apostichopus japonicus. Compar-ative Biochemistry and Physiology Part A 151, 491–498.
Wilkie, I.C., 2001. Autotomy as a prelude to regeneration in echinoderms. Journal of Mi-croscopy Research and Technique 55, 369–396.
Wootton, E.C., Pipe, R.K., 2003. Structural and functional characterization of the bloodcells of the bivalve mollusc, Scrobicularia plana. Fish & Shellfish Immunology 15,249–262.
Xue, Q., Renault, T., 2000. Enzymatic activities in European flat oyster, Ostrea edulis, andPacific oyster, Crassostrea gigas, hemolymph. Journal of Invertebrate Pathology 76,155–163.
Yuan, X.T., Yang, H.S., Wan, L.L., Zhou, Y., Zhang, T., Liu, Y., 2007. Effects of aestivationon energy budget of sea cucumber Apostichopus japonicus (Selenka)(Echinodermata: Holothuroidea). Acta Ecologica Sinica 27 (8), 3155–3161 (in Chi-nese with English abstract).
Zhang, M., Wang, L., Guo, Z.Y., Wang, B.J., 2004. Effect of lipopolysaccharide and Vibrioanguillarum on the activities of phosphatase, superoxide dismutase and the con-tent of hemocyanin in the serum of Fenneropenaeus chinensis. Marine Science 28,22–25 (in Chinese with English abstract).
Zheng, F.X., Sun, X.Q., Zhang, J.X., 2006. Histological studies on evisceration and regen-eration in Apostichopus japonicus. Journal of Fishery Sciences of China 13 (1),134–139 (in Chinese with English abstract).
Zhou, Y., Yang, H.S., Liu, S.L., Yuan, X.T., Mao, Y.Z., Liu, Y., Xu, X.L., Zhang, F.S., 2006.Feeding and growth on bivalve biodeposits by the deposit feeder Stichopusjaponicus Selenka (Echinodermata: Holothuroidea) co-cultured in lantern nets.Aquaculture 256, 510–520.





























