Embed Size (px)
Citation preview

lable at ScienceDirect
European Journal of Medicinal Chemistry 46 (2011) 1827e1834
Contents lists avai
European Journal of Medicinal Chemistry
journal homepage: http: / /www.elsevier .com/locate/ejmech
Original article
Glycoglycerolipid analogues inhibit PKC translocation to the plasma membraneand downstream signaling pathways in PMA-treated fibroblasts and humanglioblastoma cells, U87MG
Diego Colombo 1, Cristina Tringali 1, Laura Franchini, Federica Cirillo, Bruno Venerando*
Department of Medical Chemistry, Biochemistry and Biotechnology, University of Milan, Via Saldini 50, 20133 Milan, Italy
a r t i c l e i n f o
Article history:Received 26 July 2010Received in revised form16 February 2011Accepted 17 February 2011Available online 23 February 2011
Keywords:ChemopreventionGlycoglycerolipidsPKCGlioblastoma
Abbreviations: PMA, phorbol 12-myristate-13 aceMGDG, monogalactosyldiacylglycerols; EBV-EA, EpsteDAG, diacylglycerol; FBS, fetal bovine serum; PBS, ph* Corresponding author. Department of Medical C
Biotechnology, University of Milan, Via F.lli Cervi 93,Tel.: þ39 (02) 50330361; fax: þ39 (02) 50330365.
E-mail address: [email protected] (B. Ven1 These authors contributed equally to this work.
0223-5234/$ e see front matter � 2011 Elsevier Masdoi:10.1016/j.ejmech.2011.02.043
a b s t r a c t
Glycoglycerolipid analogues, derived from 2-O-b-D-galactosylglycerol, have been synthesized on the baseof the structure of natural glycoglycerolipids showing anti-tumor and anti-inflammatory efficacy. Thesecompounds have been previously demonstrated to inhibit phorbol 12-myristate-13-acetate (PMA)induced tumor promotion in mouse skin, but their mechanism of action has never been elucidated. Inthis work, we studied the effects of glycoglycerolipid analogues on PKC activation induced by PMA and itsdownstream target molecules, in human fibroblasts. Our results proved that: a) the tested compoundswere able to block PKC translocation to the plasma membrane, promoted by PMA, in a dose-dependentmanner (IC50: 0.48 mM for the most active compound 2); b) the efficacy of these compounds was stronglyconnected to their acyl chain linked to galactose; in particular, the addition of hexanoyl and branchedchains enhanced PKC inhibition, the presence of a cyclohexane ring and an excessive length of the acylchain, or its lack, exerted a negative effect; c) the inhibition of PKC translocation blocked enzyme acti-vation and downstream signaling pathways, MAPK and FAK, involved in proliferation and adhesion/migration control. In addition, the branched glycoglycerolipid (compound 2) was able to inhibit PKCtranslocation and activation in naturally highly PKC activating glioblastoma cells, U87MG. As conse-quence, U87MG cell proliferation and, especially, migration potential resulted to be markedly reduced(�30% and �84%, respectively). Thus, these results reveal the role of a PKC-dependent mechanism inglycoglycerolipid analogues mediated protective effects and highlight their possible employment in thefield of prevention/treatment of cancer.
� 2011 Elsevier Masson SAS. All rights reserved.
1. Introduction
In the early 1980s, protein kinase C (PKC) was identified as themost important target of phorbol esters and the main signalingmolecule involved in their carcinogenic effects [1,2]. On these bases,a link between PKC and cancer was definitely traced. The PKCfamily comprises at least 12 Ser/Thr kinases that could be dividedinto three subgroups: classical PKCs (a, bI, bII, g), novel PKCs (d, e, h,q), and atypical PKCs (z, i/l) [3]. PKC isoforms behave as down-stream targets of several growth factor receptors, and, in turn,
tate; PKC, protein kinase C;ineBarr virus early antigen;osphate buffer saline.hemistry, Biochemistry and20090, Segrate, Milan, Italy.
erando).
son SAS. All rights reserved.
modify the activities of signaling proteins including receptors,kinases, cytoskeletal proteins, and transcription factors [4]. In thiscontext, many evidences supported a crucial role of these enzymesin ruling cell signaling pathways involved in cell proliferation,differentiation, survival, migration control, and in cancer cellinvasion mediated by integrin binding [5]. PKC downstream eventsfrequently involve MEK-ERK [6,7] and PI3K-Akt pathways [8].Hence, PKC subtly regulates many pathways involved in cancertransformation and plays a very complex function for the pheno-type of the disease. Accordingly, deregulation of PKC activity andexpression has been reported to occur in several malignanciesincluding breast, pancreatic, hematological cancers, glioblastomamultiforme, and, overall, increased PKC levels have been frequentlyrecorded [9]. Therefore, targeting PKC may represent an attractivestrategy for novel anticancer therapies and several inhibitors arecurrently being evaluated in clinical trials involving different cancertypes [3].
Among several classes of PKC inhibitors, an ever-increasinginterest has been devoted to naturally occurring compounds,

D. Colombo et al. / European Journal of Medicinal Chemistry 46 (2011) 1827e18341828
especially those obtained from fruits and vegetables. This strategycould be a safe and less toxic alternative to other chemo-therapeutical efforts and seems to provide interesting outcome.These agents include bryostatin [10e12] and curcumin [13]. Inparticular, curcumin, a dietary pigment responsible for the yellowcolor of curry, has been shown to be a potent inhibitor of PKC and iscurrently being evaluated in myeloma, pancreas, and colorectalcancer therapy and chemoprevention [14].
In this context, the search of new natural compounds to beemployed can acquire a great importance for the prevention and/ortreatment of several human cancers. Glycoglycerolipids are plantlipids consisting mainly of monogalactosyldiacylglycerols (MGDG)and digalactosyldiacylglycerols carrying saturated and/or unsatu-rated fatty acids chains [15]. The growing interest for the use ofMGDG as drugs or nutracetics is due to both their abundance inmany edible plants [16] and to their recently reported biologicalactivities, such as tumor growth and DNA polymerase inhibition[17,18], and anti-inflammatory action [19]. Moreover, glycoglycer-olipids demonstrated to be able to hamper tumor promotionshowing to be, likely, a useful tool to prevent cancer [20e24]. Thesereports prompted us to synthesize a large number of analogues ofsuch natural compounds, in order to deepen their structure-anti-tumor activity relationship [25]. The chemical synthesis of thesemolecules has been deeply described in a previous work byColombo et al. [25]. In brief, our compounds differ from the naturalones in some structural features, such as the linkage of glycerol togalactose and the position of the acyl chain, that was linked to thesugar instead of to glycerol (Fig. 1). They revealed low toxicity andhigh anti-tumor-promoting activity [25] in a short-term in vitroassay for the inhibition of EpsteineBarr virus early antigen (EBV-EA)activation induced by phorbol 12-myristate-13-acetate (PMA), inthe immortalized human lymphoblastoid Raji cell line [26]. More-over, they have been confirmed to inhibit PMA-induced tumorpromotion in a classical two-stage skin carcinogenesis assay inmice [25]. Despite their important chemopreventive efficacy andpromising employment in the prevention/treatment of cancer, theaction mechanism of these compounds is still largely unknown.Anyway PKC was supposed to be involved in the inhibition process[25], being its role in carcinogenesis and in neoplastic trans-formation induced by PMA. Actually, the activity of different PKCisoforms in different cells seems to be responsible for the overallresponse to PKC activation by different stimuli and the final balance(for example between PKCa/b and PKCd activity) could be related tothe genesis and progression of cancer [3].
Therefore, to check our previous hypotheses and identifythe molecular target of our glycoglycerolipid analogues, we testedthe effects of 2-O-b-D-galactosylglycerol (compound 1) and ofcompounds 2e4 carrying different acyl chains linked to galactose,shown in Fig. 1, on the main signaling pathways involved in PMA-induced tumor promotion in dermal fibroblasts, taken by humanskin, and employed as a suitable healthy cell model to study themolecular mechanisms underlying cancer transformation inducedby PMA. Moreover, we checked the effects of compound 2, whichrevealed the highest activity in our assays, on some malignantproperties of the highly PKC activating human glioblastoma U87MGcell line.
Fig. 1. Structural features of synthetic glycoglycerolipid analogues. The testedanalogues 1e5 (referred to as C.1-C.5 in the following figures) differ in the nature of theacyl chain (R).
Our results demonstrated that: a) glycoglycerolipid analoguesinhibit PKC translocation to the plasma membrane and, therefore,impair its activation and the triggering of key downstream signalingpathways involved in proliferation and adhesion/migration; b) thebiological activity of glycoglycerolipid analogues seems to bestrongly related to the structure of the acyl chain linked to galac-tose; c) glycoglycerolipid analogues are able to reduce significantlythe malignancy of U87MG cells, through the inhibition of PKC.
2. Results
2.1. Glycoglycerolipid analogues inhibit PKC activation in PMA-treated fibroblasts
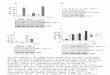
On the base of results previously obtained which demonstratedthat glycoglycerolipid analogues are able to reduce the tumorige-nicity promoted by PMA [25], in order to identify the intracellularsignaling pathways modulated by these compounds, we analyzedtheir effects using human dermal fibroblasts, as cell model.Therefore, we stimulated fibroblasts with 100 nM PMA, while, inparallel, control fibroblasts were treated with 0.005% ethanol. Asexpected, fibroblasts incubated with PMA underwent to a higherdegree of PKC redistribution from the cytosolic to the membranecompartment [2], indicating the occurrence of enzyme activation(þ49%) (Fig. 2). In order to determine the effects of glycoglycer-olipid analogues, before PMA stimulation, we pre-incubatedfibroblasts with glycoglycerolipid analogues at 0.2 or 2 mMconcentrations for 1 h. This treatment markedly inhibited thetranslocation of PKC to the plasmamembrane, in a dose-dependentmanner and differently according to the acyl chain linked togalactose. In particular, the 2-O-b-D-galactosylglycerol (compound1) (lacking the acyl chain) blocked the recruitment of PKC by 27% at0.2 mM, and by 50% at 2 mM (Fig. 2B); the “branched” compound 2inhibited PKC by 23% at 0.2 mM, and by 99% at 2 mM (Fig. 2C); the“cyclic” compound 3 inhibited PKC by 38% at 0.2 mM, and by 52% at2 mM (Fig. 2D); the “short chain” compound 4 inhibited PKC by 35%at 0.2 mM, and by 64% at 2 mM (Fig. 2E); the “long chain” compound5 inhibited PKC by 42% only at 2 mM but had no effects at 0.2 mM(Fig. 2F). These data suggest that the inhibitory effect of the testedcompounds on PKC translocation to the plasma membrane of PMAstimulated fibroblasts is strictly related to the presence of a 2-O-b-D-glycosylglycerol skeleton and modulated by an acyl chain. 2 wassignificantly the most active compound of the series (at the higherconcentration used) and the decanoyl derivative compound 5 theless one. The total amount of PKC was not affected by glycoglycer-olipid analogues (data not shown).
We assayed also the PKC inhibition activity of the most activecompound 2 at different concentrations. To this aim fibroblastswere incubated for 1 h, before PMA treatment, with 2 from 0.2 to2 mM (Fig. 3A,B).
PKC inhibition degree linearly increased according to theincrement of compound 2 concentration (Fig. 3B); IC50 was calcu-lated at 0.48 mM.
In parallel to the activation of PKC in fibroblasts, PMA-induceda slight increment of PKC expression as mRNA (isoform delta),possibly as a consequence of a higher employment of the enzyme(Fig. 3C). Significantly, the pre-incubation of fibroblasts with 2 mMcompound 2, also suppressed this effect (Fig. 3C), even if thisappears to be secondary to the inhibition of PKC enzyme activation.
2.2. Glycoglycerolipid analogues inhibit signaling pathwaysdownstream of PKC in PMA-treated fibroblasts
In order to better clarify the intracellular effects of glyco-glycerolipid analogues, we explored key signaling pathways for cell

Fig. 2. Inhibition of PKC recruitment to the plasma membrane promoted by glycoglycerolipid analogues. (A) Western blot and densitometric analysis of PKC linked to the plasmamembrane in fibroblasts. (BeF) Western blot and densitometric analysis of PKC linked to the plasma membrane in fibroblasts after stimulation with 100 nM PMA for 15 min, and infibroblasts pre-treated with 0.2 mM or 2 mM compound 1 (B), 2 (C), 3 (D), 4 (E) or 5 (F) for 1 h, and followed by 100 nM PMA treatment for 15 min; an equal amount of membraneproteins was loaded in each lane. Dotted line represents basal PKC activation in fibroblasts, with no PMA stimulation. Data are the means � S.D. of three experiments. Significanceaccording to Student’s t-test. *P < 0.05; **P < 0.01; ***P < 0.001.
D. Colombo et al. / European Journal of Medicinal Chemistry 46 (2011) 1827e1834 1829
homeostasis demonstrated to be stimulated by PKC activating PMA[27]. As previously stated, we pre-incubated fibroblasts with 2 mMcompound 2, for 1 h and, then, we added 100 nM PMA, for 15 min.The activation of the MAPK cascade and FAK was analyzed on thebase of their phosphorylation state, a reliable index of their activity.As shown in Fig. 4AeC, PMA-induced a marked increase of thephosphorylated forms of RAF, MEK, ERK1/2 in non pre-treatedfibroblasts, indicating the promotion by PKC of the MAPK pathway.Significantly, the pre-incubation of fibroblasts with compound 2inhibited the activation of MAPK. Moreover, compound 2 treatmentof fibroblasts partially inhibited the activation of FAK, promoted byPMA (Fig. 4D). Similar results were obtained, also, employing theother glycoglycerolipid analogues, according to the PKC inhibitiondegree promoted by them (data not shown).
2.3. Glycoglycerolipid analogue 2 inhibits PKC activation in humanglioblastoma cells, U87MG
On the base of previous results, we determined if glycoglycer-olipid analogues were able to inhibit PKC in human glioblastomacells, U87MG. It has been reported that over-expressed or hyper-active PKC is one of the malignant feature of glioblastoma multi-forme and strengthens tumor growth and migration [28e30]; alsoU87MG showed a high activation degree of PKC, detected as PKCassociated to the plasma membrane (Fig. 5A). As shown in Fig. 5A,
the treatment of U87MG cells with 2 mM compound 2 for 24 hinhibited the translocation of PKC to the plasma membrane by 58%.Significantly, compound 5 seemed not to be able to affect PKCactivation in these cells, and also the efficacy of the othercompounds was reduced (data not shown).
2.4. Glycoglycerolipid analogue 2 reduces proliferation rate andmigration in human glioblastoma cells, U87MG
We examined if the inhibition of PKC mediated by compound 2was associated with anti-tumor efficacy in U87MG cells. As shownin Fig. 5B, after treatment of U87MG cells with 2 mM compound 2for 24 h or 48 h, [3H] thymidine incorporation decreased by 25%after 24 h and by 30% after 48 h, providing evidence of a reductionof cell proliferation rate.
To evaluate the possibility for limiting glioblastoma cell migra-tion by a treatment with compound 2 and a subsequent inhibitionof PKC, we performed a scratchmigration assay in U87MG cells pre-treated with 2 mM compound 2 for 48 h, in comparison to controlU87MG cells pre-treated only with ethanol. Microphotographsshowing the width of the wound were taken 6 h, 24 h, and 30 hafter scratching (Fig. 6). Compound 2 treatment severely impairedmigration capability of U87MG cells: while control cells rapidlymoved into the wound, spreading more than half of the area after6 h and all the entirewound after 30 h, the size of thewound traced

Fig. 3. Inhibition of PKC translocation to the plasma membrane mediated by glyco-glycerolipid analogue 2. (A) Western blot image of membrane proteins probed withanti-PKC antibody of fibroblasts pre-treated with compound 2 at increasing concen-trations, for 1 h, followed by 100 nM PMA treatment for 15 min; (B) PKC inhibitioncurve of fibroblasts pre-treated with compound 2 at increasing concentrations for 1 h,followed by PMA treatment for 15 min; an equal amount of membrane proteins wasloaded in each lane; (C) Real-Time PCR analysis of PCK alpha and PKC delta in fibro-blasts pre-treated with compound 2 at increasing concentrations for 1 h, followed by100 nM PMA treatment for 15 min. The data represent the means � S.D of fourexperiments. Significance according to Student’s t-test. *P < 0.05.
D. Colombo et al. / European Journal of Medicinal Chemistry 46 (2011) 1827e18341830
in the plate containing U87MG cells pre-treated with compound 2was reduced only by 16% after 30 h, demonstrating a minormigration potential.
3. Discussion
Glycoglycerolipid analogues, derived from 2-O-b-D-gal-actosylglycerol, have been synthesized [25,31] based on the struc-tures of natural glycoglycerolipids and emphasizing some featurespossibly related to their anti-tumor activity [15e19,21e24]. Previ-ously, in an in vivo two-stage carcinogenesis assay, our glyco-glycerolipid analogues showed a high efficacy to inhibit the growthof mouse skin papillomas induced by PMA [25]. Moreover, theiranti-PMA action has been confirmed in a short-term in vitro assayfor EBV-EA activation, in Raji cells [25]. PMA constitutes theprototype of phorbol esters and is a well-known tumor promoter;in particular, it promotes alterations in various cell processesincluding cell growth, differentiation, survival, via the activationof PKC [1,2,32]. Therefore, given the crucial involvement of PKCin PMA-induced tumor promotion events [1,2] we investigated ifthe anti-tumor-promoting effects of glycoglycerolipid analogues
observed in mouse skin were mediated by PKC. As cell model, weemployed primary human dermal fibroblasts that show low gradeof PKC activation without stimulation. When cells were treatedwith PMA alone, PKC moved from the cytosolic fraction to theparticulate fraction. Significantly, pre-treating fibroblasts withglycoglycerolipid analogues before PMA stimulation, we inhibitedthe translocation of PKC to the plasma membrane in a dose-dependent manner. Overall, five compounds (1e5) differing in theacyl chain linked to galactose were tested and an interestingstructureeactivity relationship emerged. Firstly, the length of theacyl chain linked to galactose revealed an intrinsic importance forglycoglycerolipid analogue efficacy, as its absence (compound 1)reduced PKC inhibition aswell the presence of more than six carbonatoms in the chain (compound 5). Moreover, the branching of theacyl chain enhanced the PKC inhibition efficacy of the molecule(compound 2 vs compound 4), decreasing the membrane trans-location of PKC even below its basal level (without PMA stimula-tion), when these compounds were added at 2 mM. At thisconcentration, compound 2 almost completely inhibited PKCadhesion to plasmamembrane. On the contrary, the presence of thecyclohexane ring (compound 3) reduced the action. This evidencesuggests that an interaction between PKC and glycoglycerolipidanalogues could occur and that the process specifically couldinvolve the acyl chain. Although more biochemical details needa deeper elucidation, it might be hypothesized that thesecompounds could interact with PKC at the plasma membrane [33]or inside the cell, thus hampering its association to the plasmamembrane. This last event is mediated by two domains of PKC: theC1 domain which links diacylglycerol (DAG) or PMA [34e36] andthe C2 domainwhich interacts with membrane phosphatidylserine[37,38]. Basing on their chemical structure, it could be speculatedthat glycoglycerolipid analogues could compete with PMA, bindingto the C1 domain. In fact, the pivotal polar functional groups of PMAthat have been identified in the recognition of the C1 domaininclude hydroxyl groups and carbonyl groups [39,40]. Hydroxylgroups are carried by all the compounds and a carbonyl group iscarried by the acyl chain of compounds 2e5. Moreover, alsohydrophobic interactions are crucial for high affinity to C1 domain:the hydrophobic side chains of PMA or DAG are thought to interactwith hydrophobic amino acids surrounding the ligand binding cleft[41,42]. In this regard, both the absence and the not optimal lengthof glycoglycerolipid analogue acyl chains or the presence ofa cyclohexane ring could play a key role to explain why compounds1, 3, and 5 are less active. In this perspective, the engagement of C1domain upon glycoglycerolipid analogues binding could preventthe interaction with PMA and, therefore, the association of PKC tothe plasma membrane. Possibly secondary to enzyme inhibition,the pre-treatment of fibroblasts with glycoglycerolipid analoguesalso inhibited the slight increment of PKCd mRNA promoted byPMA. The inhibition of PKC-PMA interaction and the subsequentanchorage of the enzyme to the plasma membrane could block PKCactivation, as we demonstrated on downstream effector targets ofPKC, recording a reduced activation induced by PMA of the MAPKcascade and FAK. The blockage of these signaling pathways could belikely responsible for the PMA-induced anti-tumor-promotingactivity of such glycoglycerolipid analogues, previously demon-strated in the in vitro EBV-EA and in the in vivo two-stage mouseskin carcinogenesis assays [25].
In addition, glycoglycerolipid analogues were also active againstthe human glioblastoma cells U87MG which is characterized bya significant activation of PKC. PKC resulted to be severely impli-cated in glioblastoma multiforme malignancy and to contribute toenhance some features such as invasiveness and migration[28e30]. Only compound 2, which demonstrated the higheractivity in fibroblasts, was efficacious to inhibit PKC movement to

Fig. 4. Signaling pathways inhibited by glycoglycerolipid analogues: Western blot image and densitometric analysis of total cell lysate proteins probed with (A) anti-P-RAF, (B) anti-P-MEK, (C) anti-P-ERK1/2, (D) anti-P-FAK antibodies after treatment of fibroblasts with 2 mM compound 2 for 1 h, followed by 100 nM PMA for 15 min. An equal amount of cell lysateproteins was loaded in each lane. Dotted line represents basal RAF, MEK, ERK1/2, FAK phosphorylation in fibroblasts, with no PMA stimulation. The data represent the means � S.Dof four experiments. Significance according to Student’s t-test. **P < 0.01; ***P < 0.001.
D. Colombo et al. / European Journal of Medicinal Chemistry 46 (2011) 1827e1834 1831
the plasma membrane. This event had deep consequences onU87MG phenotype: the proliferation rate resulted to be reducedand, above all, the migration potential which is particularlyelevated in these cells, was completely inhibited, supporting thenotion that PKC could be a promising target in glioblastomatherapy.
In summary, glycoglycerolipid analogues decrease PKC proteintranslocation to membranes both induced by PMA (in fibroblasts)or DAG (in glioblastoma cells) and, hence, prevent signaling path-ways typically associated to a malignant phenotype. Thus, due totheir chemopreventive efficacy and the lack of toxicity [25], thisclass of modified natural compounds could have a promisingemployment in the field of prevention/treatment of cancer.
4. Material and methods
4.1. Materials
Pepstatin A, aprotinin, leupeptin, DMEM medium, fetal bovineserum (FBS), glutamine, penicillin, streptomycin, PMA wereprovided by Sigma Aldrich (St Louis, MO, USA); RNeasyMini kit wasprovided by Qiagen (Milan, Italy); iScript cDNA Synthesis kit and iQSYBR Green Supermix by Bio-Rad Laboratories (Richmond, VA,USA); PVDF membrane, SuperSignal West Dura Extended DurationSubstrate, and Comassie Protein Assay Reagent by Pierce Biotech-nology (Rockford, IL, USA); [3H]thymidine (20 Ci/mmol) by PerkinElmer (Waltham, MA, USA).

Fig. 5. Inhibition of PKC activation by glycoglycerolipid analogues in U87MG cells. (A) Western blot image and densitometric analysis membrane proteins probed with anti-PKCantibody, after 2 mM compound 2 or 5 treatment of U87MG cells, for 24 h. An equal amount of membrane proteins was loaded in each lane. The data represent the means � S.D ofthree experiments. Significance according to Student’s t-test. (B) Incorporation of [3H]thymidine in U87MG cells treated for 24 h or 48 h with 2 mM compound 2. The data representthe means � S.D of four experiments. Significance according to Student’s t-test. *P < 0.05; **P < 0.01; ***P < 0.001.
D. Colombo et al. / European Journal of Medicinal Chemistry 46 (2011) 1827e18341832
4.2. Cell culture
Primary human dermal fibroblasts were a kind gift by Dr. M.Sampaolesi and were cultured in DMEM high glucose supple-mented with 10% (v/v) FBS, 4 mM glutamine, 100 U/ml penicillin,and 100 mg/ml streptomycin. The human glioblastoma cell lineU87MG was purchased from ECACC (European Collection of CellCulture-Sigma Aldrich) and maintained in DMEM high glucosesupplemented with 10% (v/v) FBS, 4 mM glutamine, 100 U/mlpenicillin, and 100 mg/ml streptomycin.
4.3. Treatment of fibroblasts and U87MG cells withglycoglycerolipid analogues
Glycoglycerolipid analogues 1e5 were synthesized according toColombo et al. [25,31] andwere>95% pure (by 1H NMR). In order todetect the effect of these compounds on fibroblast signaling path-ways, 5 � 105 cells were plated in 100 mm plates in DMEM sup-plemented with 10% FBS. 24 h after seeding, fibroblasts weretreated with glycoglycerolipid analogues, dissolved in ethanol, at0.2e2 mM concentrations in 10% FBS DMEM for 1 h, at 37 �C. Then,100 nM PMA was added to all samples, for 15 min. In parallel, analiquot of fibroblasts were treated with 0.005% ethanol and 100 nMPMA. Control fibroblasts were incubated only with 0.005% ethanol.Finally, all fibroblasts were scraped and lysed by sonication.
For glioblastoma cell assays, 4 � 105 U87MG cells were plated in100 mm plates in DMEM supplemented with 10% FBS. After 24 h,U87MG cells were treated with glycoglycerolipid analogues at anoptimal concentration (2 mM), in 10% FBS DMEM, for 24 h, at 37 �C;then, they were scraped and lysed by sonication. Control U87MGcells were incubated with 0.005% ethanol.
4.4. Western blot
Fibroblasts and U87MG cells were scraped, harvested bycentrifugation and resuspended in PBS containing 1 mg/ml pep-statin A, 10 mg/ml aprotinin, and 10 mg/ml leupeptin. The cells werelysed by sonication in order to obtain the total extract. Cell lysatewas centrifuged at 200.000� g, for 20 min, on a TL100
Ultracentrifuge (Beckman, Fullerton, CA, USA) to obtain themembrane fraction. (25 mg) total or (10 mg) membrane proteinswere separated on 10% SDS-PAGE, and subsequently transferredonto PVDF membrane. The following antibodies were used for theassays: anti-PKC (Sigma Aldrich), anti-phospho-RAF, anti-phos-pho-MEK, anti-phospho-ERK, anti-phospho-FAK (Cell Signaling,Danvers, MA, USA).
The densitometric analysis of blots was performed using theQuantity One Software (Bio-Rad Laboratories).
4.5. RNA extraction and real-time RT-PCR
Total RNA was isolated using the RNeasy Mini kit (Bio-RadLaboratories), following the protocol suggested by the manufac-turer. 1 mg of RNA was reverse-transcribed employing the iScriptcDNA Synthesis kit (Bio-Rad Laboratories). Briefly,10 ng of total RNAwas used as template for real-time PCR performed using the iCyclerthermal cycler (Bio-Rad Laboratories). PCRmixture included 0.2 mMprimers, 50 mM KCl, 20 mM Tris/HCl, pH 8.4, 0.8 mM dNTPs, 0.7 UiTaq DNA Polymerase, 3 mMMgCl2, and SYBRGreen (iQ SYBRGreenSupermix from Bio-Rad Laboratories), in a final volume of 20 ml.Amplification and real-time data acquisition were performed usingthe following cycle conditions: initial denaturation at 95 �C for3 min, followed by 40 cycles of 10 s at 95 �C and 30 s at 58 �C. Thefold change in expression of the genes was normalized to theexpression of b-actin mRNA and was calculated by the equation(2)�DDCt. The primers used are: PKC delta (forward: 50-AAGGCCCT-CAAGAAGGATGT-30; reverse: 50CTGGAAGGTGCAGATGAGGT-30),PKC alpha (forward: 50-GTGGCAAAGGAGCAGAGAAC-30; reverse: 50-AACTGGGGGTTGACATACGA-30), b-actin (forward: 50-CGACAGGAT-GCAGAAGGAG-30; reverse: 50-ACATCTGCTGGAAGGTGGA-30). Theaccuracy was monitored by the analysis of the melting curves.
4.6. Proliferation assay
To assess [3H]thymidine incorporation after PKC inhibition,4 � 104 U87MG cells were plated and treated with 2 mM compound2 for 24 h or 48 h. 0.5 mCi/ml [3H]thymidine was added for 4 h at

Fig. 6. Phase contrast microphotographs of U87MG cells in scratch migration assay. Cells pre-treated with 2 mM of compound 2 (left panel) or ethanol (control) (right panel) for 48 hat (A) time 0 or (B) 6 h, (C) 24 h, and (D) 30 h post-scratching. Dotted lines represent the borders of wounds. Image magnification: 4�. a.u.: arbitrary units.
D. Colombo et al. / European Journal of Medicinal Chemistry 46 (2011) 1827e1834 1833
37 �C, and the radioactivity incorporated was determined in tri-chloroacetic insoluble material using a b-Counter (PerkinElmer).
4.7. Scratch migration assay
8 � 104 U87MG cells were plated in 6-well plate and treated for48 hwith 2 mMcompound 2 in 10% FBS DMEM. A scratch woundwascreated with a 1 ml tip (time 0). Then, the plates were washed twicewith PBS to remove detached cells and the cultured medium sup-plementedwith2mMcompound2was replaced. Imagesof thewoundarea were observed using an inverted fluorescence microscope (Ix50Olympus) and photographed 6 h, 24 h, and 30 h post-scratching. Thewidth of thewoundwasmeasured using the software Image ProPlus.
4.8. Statistical analysis
Values are presented as means � S.D. Statistical analyses wereperformed using Student’s t-test.
Acknowledgement
This work was supported by University of Milan and by PRIN (toBV) and the Italian Ministries of Health and Research.
References
[1] M. Castagna, Y. Takai, K. Kaibuchi, K. Sano, U. Kikkawa, Y. Nishizuka, Directactivation of calcium-activated, phospholipid-dependent protein kinase bytumor-promoting phorbol esters, J. Biol. Chem. 257 (1982) 7847e7851.
[2] U. Kikkawa, Y. Takai, R. Miyake, Y. Nishizuka, Protein kinase C as a possiblereceptor protein of tumor-promoting phorbol esters, J. Biol. Chem. 258 (1983)11442e11445.
[3] H.J. Mackay, C.J. Twelves, Targeting the protein kinase C family: are we thereyet? Nat. Rev. Cancer 7 (2007) 554e562.
[4] E.M. Griner, M.G. Kazanietz, Protein kinase C and other diacylglycerol effectorsin cancer, Nat. Rev. Cancer 7 (2007) 281e294.
[5] C. Rosse, M. Linch, S. Kermorgant, A. Cameron, K. Boeckeler, P. Parker, PKC andthe control of localized signal dynamics, Nat. Rev. Mol. Cell. Biol. 11 (2010)103e112.

D. Colombo et al. / European Journal of Medicinal Chemistry 46 (2011) 1827e18341834
[6] H. Cai, U. Smola, V. Wixler, I. Eisenmann-Tappe, M.T. Diaz-Meco, J. Moscat,U. Rapp, G.M. Cooper, Role of diacylglycerol-regulated protein kinase Cisotypes in growth factor activation of the Raf-1 protein kinase, Mol. Cell. Biol.17 (1997) 732e741.
[7] C.J. Marshall, Cell signaling. Raf gets it together, Nature 386 (1996) 127e128.[8] V.C. Grossoni, K.B. Falbo, M.G. Kazanietz, E.D. de Kier Joffè, A.J. Urtreger,
Protein kinase C delta enhances proliferation and survival of murinemammary cells, Mol. Carcinog. 46 (2007) 381e390.
[9] A. Ali, S. Ali, B. El-Rayes, P. Philip, F. Sarkar, Exploitation of protein kinase C:a useful target for cancer therapy, Cancer Treat. Rev. 35 (2009) 1e8.
[10] J.B. Smith, L. Smith, G.R. Pettit, Bryostatins: potent, new mitogens that mimicphorbol ester tumor promoters, Biochem. Biophys. Res. Commun. 132 (1985)939e945.
[11] P.A. Philip, J.A. Zonder, Pharmacology and clinical experience with bryostatin1: a novel anticancer drug, Expert Opin. Investig. Drug 8 (1989) 2189e2199.
[12] B.F. El-Rayes, S. Gadgeel, A.F. Shields, S. Manza, P. Lorusso, P.A. Philip, Phase Istudy of bryostatin 1 and gemcitabine, Clin. Cancer Res. 12 (2006) 7059e7062.
[13] J.K. Lin,Molecular targets of curcumin, Adv. Exp.Med. Biol. 595 (2007) 227e243.[14] S. Singh, A. Khar, Biological effects of curcumin and its role in cancer che-
moprevention and therapy, Anticancer Agent Med. Chem. 6 (2006) 259e270.[15] G. Holzl, P. Dormann, Structure and function of glycoglycerolipids in plants
and bacteria, Prog. Lipid Res. 46 (2007) 225e243.[16] E. Larsen, L.P. Christensen, Common vegetables and fruits as a source of 1,2-di-O-
a-linolenoyl-3-O-b-D-galactopyranosyl-sn-glycerol, a potential anti-inflamma-tory and antitumor agent, J. Food Lipids 14 (2007) 272e279.
[17] N. Maeda, Y. Kokai, S. Ohtani, T. Hada, H. Yoshida, Y. Mizushina, Inhibitoryeffects of preventive and curative orally administered spinach glycoglycer-olipid fraction on the tumor growth of sarcoma and colon in mouse graftmodels, Food Chem. 112 (2009) 205e210.
[18] Y. Matsui, T. Hada, N. Maeda, Y. Sato, Y. Yamaguchi, T. Takeuchi, M. Takemura,F. Sugawara, K. Sakaguchi, H. Yoshida, Structure and activity relationship ofmonogalactosyl diacylglycerols, which selectively inhibited in vitro mamma-lian replicative DNA polymerase activity and human cancer cell growth,Cancer Lett. 283 (2009) 101e107.
[19] A. Bruno, C. Rossi, G. Marcolongo, A. Di Lena, A. Venzo, C.P. Berrie, D. Corda,Selective in vivo anti-inflammatory action of the galactolipid monogalacto-syldiacylglycerol, Eur. J. Pharmacol. 524 (2005) 159e168.
[20] A. Murakami, H. Ohigashi, K. Koshimizu, Chemoprevention: insights into bio-logicalmechanismsandpromising foodfactors, FoodRev. Int.15(1999)335e395.
[21] H. Shirahashi, N. Murakami, M. Watanabe, A. Nagatsu, J. Sakakibara,H. Tokuda, H. Nishino, A. Iwashima, Glycolipids. VIII. Isolation and identifi-cation of antitumor-promoting principles from the fresh-water cyanobacte-rium Phormidium tenue, Chem. Pharm. Bull. 41 (1993) 1664e1666.
[22] N. Murakami, H. Imanura, J. Sakakibara, N. Yamada, Glycolipids. I. Seven newmonogalactosyl diacylglycerols isolated from the axenic cyanobacteriumPhormidium tenue, Chem. Pharm. Bull. 38 (1999) 3497e3499.
[23] A. Murakami, Y. Nakamura, K. Koshimizu, H. Ohigashi, Glyceroglycolipidsfrom Citrus hystrix, a traditional herb in Thailand, potently inhibit the tumor-promoting activity of 12-O-tetradecanoylphorbol 13-acetate in mouse skin,J. Agric. Food Chem. 43 (1995) 2779e2783.
[24] T. Morimoto, A. Nagatsu, N. Murakami, J. Sakakibara, H. Tokuda, H. Nishino,A. Iwashima, Anti-tumour-promoting glyceroglycolipids from the green alga,Chlorella vulgaris, Phytochemistry 40 (1995) 1433e1437.
[25] D. Colombo, L. Franchini, L. Toma, F. Ronchetti, R. Tanaka, J. Takayasu,H. Nishino, H. Tokuda, Cyclic and branched acyl chain galactoglycerolipids andtheir effect on anti-tumor-promoting activity, Eur. J. Med. Chem. 41 (2006)1456e1463 (and references herein reported).
[26] H. Tokuda, T. Konoshima, M. Kozuka, T. Kimura, Inhibitory effects of 12-O-tetradecanoylphorbol-13-acetate and teleocidin B-induced EpsteineBarrvirus by saponins and its related compounds, Cancer Lett. 40 (1988) 309e317.
[27] C.B. Weldon, A. McKee, W.B.M. Collins-Buro, L.I. Melnik, A.B. Scandurro,J.A. McLachlan, M.E. Burow, B.S. Beckman, PKC-mediated survival signaling inbreast carcinoma cells a role for MEK1-AP1 signaling, Int. J. Oncol. 26 (2005)763e768.
[28] H. Guo, F. Gu, W. Li, B. Zhang, R. Niu, L. Fu, N. Zhang, Y. Ma, Reduction ofprotein kinase C zeta inhibits migration and invasion of human glioblastomacells, J. Neurochem. 109 (2009) 203e213.
[29] M.H. Park, B.H. Ahn, Y.K. Hong, S. do Min, Overexpression of phospholipase Denhances matrix metalloproteinase-2 expression and glioma cell invasion viaprotein kinase C and protein kinase A/NF-kappaB/Sp1-mediated signalingpathways, Carcinogenesis 30 (2009) 356e365.
[30] R.M. Baldwin, D.A. Parolin, I.A. Lorimer, Regulation of glioblastoma cell inva-sion by PKC iota and RhoB, Oncogene 27 (2008) 3587e3595.
[31] D. Colombo, F. Ronchetti, A. Scala, L. Toma, Bioactive glycoglycerolipidanalogues: an expeditious enzymatic approach to mono- and diesters of 2-O-b-D-galactosylglycerol, Tetrahedron: Asymmetry 9 (1998) 2113e2119.
[32] P.M. Blumberg, In vitro studies on the mode of action of the phorbol esters,potent tumor promoters: part 1, Crit. Rev. Toxicol. 8 (1988) 153e197.
[33] K. Danker, W. Reutter, G. Semini, Glycosidated phospholipids: uncoupling ofsignaling pathways at the plasma membrane, Br. J. Pharmacol. 160 (2010)36e47.
[34] P.J. Parker, L. Coessens, N. Totty, L. Rhee, S. Young, E. Chen, S. Stabel,M.D. Waterfield, A. Ullrich, The complete primary structure of protein kinaseC, the major phorbol ester receptor, Science 233 (1986) 853e859.
[35] M.G. Kazanietz, Novel “nonkinase” phorbol ester receptors: the C1 domainconnection, Mol. Pharmacol. 61 (2002) 759e767.
[36] D. Ron, M.G. Kazanietz, New insight into the regulation of protein kinase C andnovel phorbol ester receptors, FASEB J. 13 (1999) 1658e1676.
[37] R.V. Stahelin, W. Cho, Roles of calcium ions in the membrane binding of C2domains, Biochem. J. 359 (2001) 679e685.
[38] J. Rizo, T.C. Sudhof, C2-domains, structure and fuction of a universal Ca2þ-binding domain, J. Biol. Chem. 273 (1998) 15879e15882.
[39] K. Sugita, C.F. Neville, M. Sodeoka, H. Sasai, M. Shibasaki, Stereocontrolledsyntheses of phorbol analogs and evaluation of their binding affinity to PKC,Tetrahedron Lett. 36 (1995) 1067e1070.
[40] G. Boijeaf Gennas, V. Talman, O. Aitio, E. Ekokoski, M. Finel, R. Tuominem,J. Yli-Kauhaluoma, Design, Synthesis, and biological activity of isophthalicacid derivatives targeted to the C1 domain of protein kinase C, J. Med. Chem.52 (2009) 3969e3981.
[41] G. Zhang, M.G. Kazanietz, P.M. Blumberg, J.H. Hurley, Crystal structure of thecys2 activator-binding domain of protein kinase c delta in complex withphorbol ester, Cell 81 (1995) 917e924.
[42] N.E. Lewin, D.K. Bhattacharyya, P.M. Blumberg, V.E. Marquez, Conformation-ally constrained analogues of diacylglycerol (DAG). How much structuralcomplexity is necessary for recognition and high binding affinity to proteinkinase C? J. Med. Chem. 43 (2000) 921e944.