Embed Size (px)
Citation preview

REVIEW PAPER
Genetic syndromes caused by mutations in epigenetic genes
Marıa Berdasco • Manel Esteller
Received: 10 December 2012 / Accepted: 18 January 2013 / Published online: 31 January 2013
� Springer-Verlag Berlin Heidelberg 2013
Abstract The orchestrated organization of epigenetic
factors that control chromatin dynamism, including DNA
methylation, histone marks, non-coding RNAs (ncRNAs)
and chromatin-remodeling proteins, is essential for the
proper function of tissue homeostasis, cell identity and
development. Indeed, deregulation of epigenetic profiles
has been described in several human pathologies, including
complex diseases (such as cancer, cardiovascular and
neurological diseases), metabolic pathologies (type 2 dia-
betes and obesity) and imprinting disorders. Over the last
decade it has become increasingly clear that mutations of
genes involved in epigenetic mechanism, such as DNA
methyltransferases, methyl-binding domain proteins, his-
tone deacetylases, histone methylases and members of the
SWI/SNF family of chromatin remodelers are linked to
human disorders, including Immunodeficiency Centro-
meric instability Facial syndrome 1, Rett syndrome,
Rubinstein–Taybi syndrome, Sotos syndrome or alpha-
thalassemia/mental retardation X-linked syndrome, among
others. As new members of the epigenetic machinery are
described, the number of human syndromes associated with
epigenetic alterations increases. As recent examples, muta-
tions of histone demethylases and members of the non-
coding RNA machinery have recently been associated with
Kabuki syndrome, Claes-Jensen X-linked mental retardation
syndrome and Goiter syndrome. In this review, we describe
the variety of germline mutations of epigenetic modifiers that
are known to be associated with human disorders, and dis-
cuss the therapeutic potential of epigenetic drugs as pallia-
tive care strategies in the treatment of such disorders.
Introduction
Chromatin dynamism is critical to basic cellular processes
such as gene transcription, DNA replication, DNA
recombination and DNA repair. DNA accessibility is
modulated by epigenetic mechanisms that ultimately alter
the structure of the chromatin and provide binding sites
for a wide variety of regulatory proteins. The orchestrated
organization of epigenetic factors, including DNA meth-
ylation, histone marks, non-coding RNAs (ncRNAs), and
their associated chromatin proteins, is essential for
development and cellular differentiation. For instance,
extensive chromatin remodeling occurs on a global level
during early development. DNA methylation patterns
undergo genome-wide alterations that occur immediately
after fertilization and during early-preimplantation devel-
opment, together with histone modification changes, such
as increased H3K9me with differentiation (Reik 2007).
Epigenetic factors also guarantee the activation and
maintenance of specific differentiation programs in adult
somatic cells (Berdasco and Esteller 2010). The active
role of epigenetic factors in controlling cellular differen-
tiation is supported by spontaneous cell differentiation
after treatment with demethylating agents or histone
M. Berdasco � M. Esteller (&)
Cancer Epigenetics Group, Cancer Epigenetics and Biology
Program (PEBC), Bellvitge Biomedical Research Institute
(IDIBELL), 3rd Floor, Hospital Duran i Reynals, Av. Gran
Via 199-203, 08908 L’Hospitalet de LLobregat Barcelona,
Catalonia, Spain
e-mail: [email protected]
M. Esteller
Department of Physiological Sciences II, School of Medicine,
University of Barcelona, Barcelona, Catalonia, Spain
M. Esteller
Institucio Catalana de Recerca I Estudis Avancats (ICREA),
08010 Barcelona, Catalonia, Spain
123
Hum Genet (2013) 132:359–383
DOI 10.1007/s00439-013-1271-x

deacetylase inhibitors (reviewed in Berdasco and Esteller
2011). Treatment with the demethylation agent 5-aza-20-deoxycytidine promotes differentiation of different types
of adult stem cells into cardiac myogenic or osteogenic
cells by enhancing the expression of lineage genes. In a
similar manner, histone deacetylase inhibitor trichostatin
enhances chondrogenic or neural differentiation of stem
cells, reinforcing the epigenetic control of differentiation.
Furthermore, the essential role of these factors is reflected
in the fact that altered profiles of epigenetic marks often
lead to defaults in cellular homeostasis and development
of human diseases. Genetic alterations could explain the
causes of several monogenic diseases. However, the
genetic basis underlying the origin of complex and mul-
tifactorial diseases remains largely unknown and the
importance of the role of non-genetic mechanisms,
including epigenetic mechanisms or posttranslational
protein modifications, is increasingly being realized.
Cancer has been the best characterized complex human
disease associated with epigenetic defects (Berdasco and
Esteller 2010), but the list of complex diseases carrying
epigenetic defaults has been increasing rapidly in recent
years. Epigenetic studies have now been made of complex
diseases such as obesity, type 2 diabetes mellitus, car-
diovascular diseases and neurological disorders. These
pathogenic mechanisms are particularly interesting
because the epigenetic effects may also be affected by
aspects of the environment such as diet and lifestyle,
raising the possibility of ‘‘resetting’’ the altered epigenetic
marks. Deleterious epigenetic profiles could be a conse-
quence of mutations in the ‘‘writers’’, that is to say,
dysfunctional enzymes that are responsible for putting in
and out the epigenetic marks. Defective epigenetic
machinery has been observed in cancer initiation and
progression. Furthermore, germline mutations of epige-
netic modifiers contribute to the development of human
diseases including intellectual disability (review in: Fro-
yen et al. 2006; Kramer and van Bokhoven 2009;
Franklin and Mansuy 2011). The aim of the present
review is to provide an overview of these disorders
grouped by the type of epigenetic change involved:
(i) alterations in DNA methylation players; (ii) mutations
in histone modifiers; (iii) disruption of chromatin-remod-
eling complexes, and (iv) mutations in non-coding RNA
processing machinery.
Genetic disorders linked to DNA methylation defects
DNA methylation, or the addition of a methyl group to a
cytosine, is a key epigenetic player that has long been
considered the genome’s fifth base (Portela and
Esteller 2010). In mammals this reaction occurs at CpG
(deoxycytidine-phosphate-deoxyguanosine) sites located
throughout the genome, but there are certain areas, known
as CpG islands, that are enriched in CpG dinucleotides
(especially in promoter regions). Non-CpG islands of the
human genome are usually methylated and prevent geno-
mic instability phenomena, such as the movement of
transposable elements (Berdasco and Esteller 2010). Nor-
mal methylation at these sequences is also necessary for
X-chromosome inactivation in females and genomic
imprinting. Conversely, CpG islands are usually unme-
thylated, being closely related to the expression of house-
keeping genes. It is estimated that only 6 % of the human
CpG islands are methylated and, consequently, silenced,
being essential for maintaining tissue-specific patterns
during development and differentiation. By our current
understanding, this ‘‘DNA methylation code’’ seems to be
an oversimplification, since, in recent years, new genomic
contexts outside of CpG islands, known as CpG shores,
have emerged as candidates for regulating gene expression
of tissue-specific genes. The technological advance in
studying DNA methylation will provide insight into the
role of 5-methylcytosine patterns with respect to their
density, location and function, amongst other features.
Additionally the recently discovered cytosine modification
5-hydroxymethyl-20-deoxycytidine (5hmC) needs to be
further studied to determine its implications for normal and
diseased epigenetic regulation.
The enzymes responsible for introducing the methyl
group into a cytosine are DNA methyltransferases
(DNMTs). Three major proteins with DNMT activity have
been identified in mammals: DNMT1, DNMT3A and
DNMT3B. DNMT1 is a widely expressed maintenance
DNMT that recognizes hemimethylated DNA and is
responsible for maintaining the existing methylation pat-
terns after DNA replication. By contrast, DNMT3 enzymes
are de novo DNMTs that introduce methyl groups into
previously unmethylated cytosines. These enzymes intro-
duce a methyl group into the genome, but this ‘‘writing’’
must be interpreted (read) by the rest of the cellular
machinery (i.e., transcription factors, DNA polymerases,
chromatin-remodeling proteins, epigenetic enzymes, etc.).
Additional members of the DNMT family without meth-
yltransferase activity have been reported, such as DNMT2
or DNMT3L. DNMT3L lacks the amino acid sequence
necessary for methyltransferase but it seems to be required
for the establishment of maternal genomic imprints
(Aapola et al. 2002).
Methyl-CpG-binding domain (MBD) proteins are one of
the DNA methylation-associated proteins that could be
recruited to methylated DNA and in turn facilitate the
recruitment of histone modifiers and chromatin-remodeling
complexes (Portela and Esteller 2010). Evidence is
mounting of the role of DNA methylation in modulating
360 Hum Genet (2013) 132:359–383
123

cognitive functions of the central nervous system, such as
learning and memory, and of how dysregulation of DNMTs
activities can give rise to neurological disorders (Liu et al.
2009; Urdinguio et al. 2009; Feng et al. 2010). Some of the
genetic syndromes featuring mutations in the DNA-related
machinery that often cause neurological disorders are dis-
cussed in this section (Table 1).
DNMT1 mutations and disorders of the central
and peripheral nervous system
Hereditary sensory and autonomic neuropathy type 1 with
dementia and hearing loss (HSAN1; MIM #614116) is a
degenerative disorder of the central and peripheral nervous
system. Its clinical manifestations consist of sensory
impairment, sudomotor dysfunction (loss of sweating),
dementia and sensorineural hearing loss. HSAN1 is
inherited in an autosomal dominant manner, although the
proportion of patients with de novo mutations is unknown,
DNMT1 being the only gene in which mutations of exons
20 and 21 are known to cause HSAN1 (Klein et al. 2011;
Winkelmann et al. 2012). Molecular genetic testing to
screen for three mutations in exon 21 of DNMT1
(p.Ala570Val, p.Gly605Ala and p.Val606phe) is available
for research purposes. Mutations are present within the
targeting-sequence domain of DNMT1 that regulates the
binding of the enzyme to chromatin during the S-phase and
is responsible for maintaining this association during the
G2-M phases (Fatemi et al. 2001; Song et al. 2012).
DNMT1 is strongly expressed in postmitotic neurons and
plays important roles in neuronal differentiation, migration
and central neural connection (Feng et al. 2010). The
functional involvement of DNMT1 mutations has been
assessed in in vitro studies in HeLa cells (Klein et al.
2011), so that cells carrying mutations in the DNMT1
targeting sequence showed abnormal heterochromatin
binding of DNMT1 during the G2 phase and were pre-
maturely degraded. Abolishing DNMT1 function affects
DNA methylation cellular levels: first, a lower level of
global methylation (8 %) has been measured in mutant
cells (i.e., satellite 2 methylation was reduced); second,
site-specific hypermethylation at specific loci has also been
found (Klein et al. 2011). Additionally, DNMT1 is required
for CD4? differentiation into T regulatory cells (Jose-
fowicz et al. 2009), and a link between the absence of
CD4? T regulatory cells and the autoimmune response in
neurological syndromes has been proposed (Winkelmann
et al. 2012). The findings from several studies together
suggest that DNMT1 participates in a precise mechanism
of dynamic regulation of neuronal survival, but additional
efforts will be needed to elucidate its pathogenic mecha-
nisms and to explain the phenotypic variation observed
between individuals bearing different mutations.
DNMT3 mutations and immunodeficiency centromeric
instability facial syndrome 1
Immunodeficiency centromeric instability facial syndrome
1 (ICF1, MIM #242860) is a rare autosomal recessive
disorder characterized by immune defects in association
with centromere instability and facial anomalies. Several
chromosomal abnormalities have been described, including
the juxtacentromeric heterochromatin formation of chro-
mosomes 1, 9 and 16, an increased frequency of somatic
recombination between the arms of these chromosomes,
and a marked tendency to form multibranched configura-
tions (Ehrlich 2003). 60 % of ICF1 patients carry muta-
tions in the de novo DNA methyltransferase DNMT3B
(Xu et al. 1999; Lana et al. 2012). Mutations in the ZBTB24
gene, which encodes a transcription factor, are responsible
for the ICF type 2 phenotype (de Greef et al. 2011).
Hypomethylation in ICF patients commonly affect specific
non-coding repetitive sequences (satellites 2 and 3, subt-
elomeric sequences and Alu sequences), imprinted genes
and genes located in constitutive and facultative hetero-
chromatin (Xu et al. 1999; Yehezkel et al. 2008; Brun et al.
2011), causing chromatin decondensation and chromosome
instability. Recently, whole-genome bisulfite sequencing of
an ICF patient harboring mutated DNMT3B and one heal-
thy control have been performed to assess DNA methyla-
tion at base pair resolution (Heyn et al. 2012). The authors
concluded that ICF patients have 42 % less global DNA
methylation, especially in inactive heterochromatic regions
(in accordance with previous studies). Interestingly, the
methylation status of transcriptional active loci and rRNA
repeats did not change, suggesting that there is a selective
pressure to maintain the stability of these genomic struc-
tures (Heyn et al. 2012). In addition to methylation studies,
the altered expression of more than 700 genes in ICF1
patients has been described, especially genes related to
immune function, development and neurogenesis (Jin et al.
2008). Interestingly, half the upregulated genes were hy-
pomethylated (compared with normal cells) in parallel with
the loss of the histone repressive H3K27 trimethylation
mark and the gain of the histone active marks H3K9
acetylation and H3K4 trimethylation (Jin et al. 2008). Not
only the protein-coding genes are altered; a dramatic loss
of methylation (from 80 to 30 %) was found in hetero-
chromatic genes, which are usually aberrantly hypome-
thylated in cancer cells, although the hypomethylation was
not always associated with their activation (Brun et al.
2011). In contrast to the dysregulation of protein-coding
genes, no changes in histone marks associated with het-
erochromatic genes could be found (Brun et al. 2011).
Finally, the genomic instability generated in ICF patients
also resulted in replication defects, including shortening of
the S-phase, a higher global replication fork speed and
Hum Genet (2013) 132:359–383 361
123
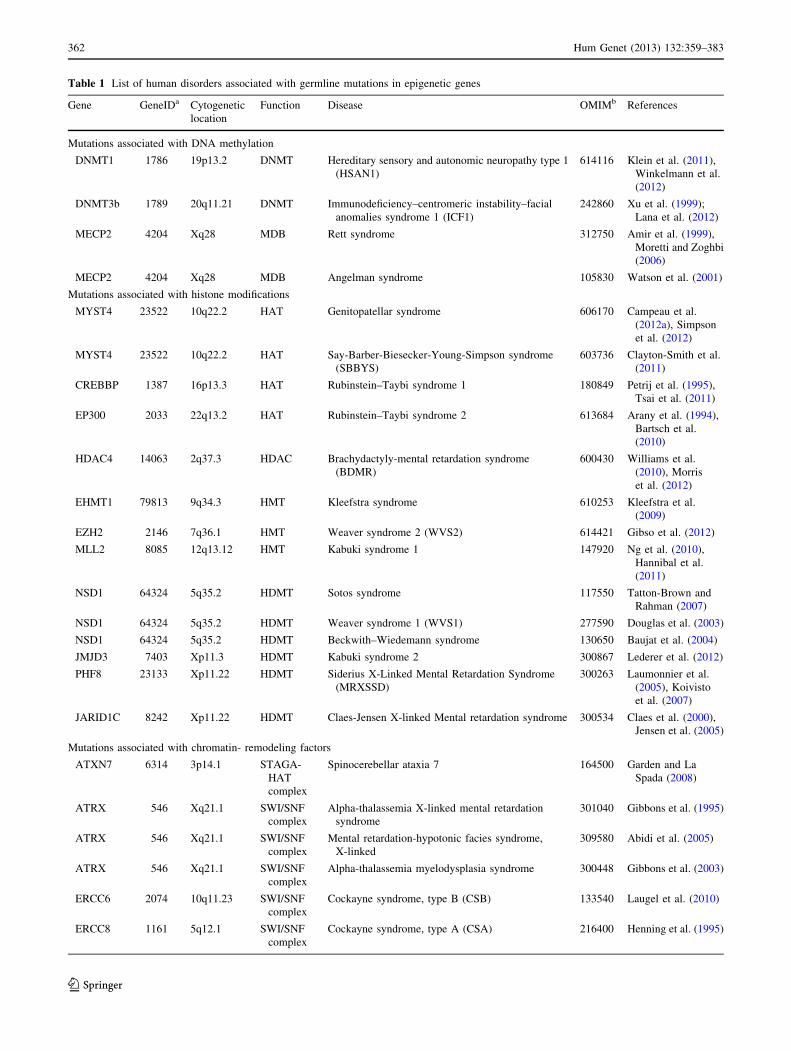
Table 1 List of human disorders associated with germline mutations in epigenetic genes
Gene GeneIDa Cytogenetic
location
Function Disease OMIMb References
Mutations associated with DNA methylation
DNMT1 1786 19p13.2 DNMT Hereditary sensory and autonomic neuropathy type 1
(HSAN1)
614116 Klein et al. (2011),
Winkelmann et al.
(2012)
DNMT3b 1789 20q11.21 DNMT Immunodeficiency–centromeric instability–facial
anomalies syndrome 1 (ICF1)
242860 Xu et al. (1999);
Lana et al. (2012)
MECP2 4204 Xq28 MDB Rett syndrome 312750 Amir et al. (1999),
Moretti and Zoghbi
(2006)
MECP2 4204 Xq28 MDB Angelman syndrome 105830 Watson et al. (2001)
Mutations associated with histone modifications
MYST4 23522 10q22.2 HAT Genitopatellar syndrome 606170 Campeau et al.
(2012a), Simpson
et al. (2012)
MYST4 23522 10q22.2 HAT Say-Barber-Biesecker-Young-Simpson syndrome
(SBBYS)
603736 Clayton-Smith et al.
(2011)
CREBBP 1387 16p13.3 HAT Rubinstein–Taybi syndrome 1 180849 Petrij et al. (1995),
Tsai et al. (2011)
EP300 2033 22q13.2 HAT Rubinstein–Taybi syndrome 2 613684 Arany et al. (1994),
Bartsch et al.
(2010)
HDAC4 14063 2q37.3 HDAC Brachydactyly-mental retardation syndrome
(BDMR)
600430 Williams et al.
(2010), Morris
et al. (2012)
EHMT1 79813 9q34.3 HMT Kleefstra syndrome 610253 Kleefstra et al.
(2009)
EZH2 2146 7q36.1 HMT Weaver syndrome 2 (WVS2) 614421 Gibso et al. (2012)
MLL2 8085 12q13.12 HMT Kabuki syndrome 1 147920 Ng et al. (2010),
Hannibal et al.
(2011)
NSD1 64324 5q35.2 HDMT Sotos syndrome 117550 Tatton-Brown and
Rahman (2007)
NSD1 64324 5q35.2 HDMT Weaver syndrome 1 (WVS1) 277590 Douglas et al. (2003)
NSD1 64324 5q35.2 HDMT Beckwith–Wiedemann syndrome 130650 Baujat et al. (2004)
JMJD3 7403 Xp11.3 HDMT Kabuki syndrome 2 300867 Lederer et al. (2012)
PHF8 23133 Xp11.22 HDMT Siderius X-Linked Mental Retardation Syndrome
(MRXSSD)
300263 Laumonnier et al.
(2005), Koivisto
et al. (2007)
JARID1C 8242 Xp11.22 HDMT Claes-Jensen X-linked Mental retardation syndrome 300534 Claes et al. (2000),
Jensen et al. (2005)
Mutations associated with chromatin- remodeling factors
ATXN7 6314 3p14.1 STAGA-
HAT
complex
Spinocerebellar ataxia 7 164500 Garden and La
Spada (2008)
ATRX 546 Xq21.1 SWI/SNF
complex
Alpha-thalassemia X-linked mental retardation
syndrome
301040 Gibbons et al. (1995)
ATRX 546 Xq21.1 SWI/SNF
complex
Mental retardation-hypotonic facies syndrome,
X-linked
309580 Abidi et al. (2005)
ATRX 546 Xq21.1 SWI/SNF
complex
Alpha-thalassemia myelodysplasia syndrome 300448 Gibbons et al. (2003)
ERCC6 2074 10q11.23 SWI/SNF
complex
Cockayne syndrome, type B (CSB) 133540 Laugel et al. (2010)
ERCC8 1161 5q12.1 SWI/SNF
complex
Cockayne syndrome, type A (CSA) 216400 Henning et al. (1995)
362 Hum Genet (2013) 132:359–383
123
earlier replication of heterochromatic genes in S-phase
(Lana et al. 2012). To conclude, the loss of DNMT3B
function and the interaction of DNMT3B with histone
modifications, together with the variable clinical features
of the patients, make ICF samples an ideal model for
investigating the epigenetic network and their molecular
consequences in several biological pathways (gene tran-
scription, DNA replication and recombination, among
others).
MeCP2 genetic alterations and Rett syndrome
Rett syndrome (MIM #312750) is a progressive neurode-
velopmental disorder characterized by arrested develop-
ment between 6 and 18 months of age, regression of
acquired skills, loss of speech, unusual stereotyped move-
ments and intellectual disability (Zachariah and Rastegar
2012). It affects predominantly females, occurring at a
frequency of 1:10,000 live births, but male patients with
Rett syndrome and variable phenotype (i.e., severe to
moderate congenital encephalopathy, infantile death or
psychiatric manifestations) have also been described (Ravn
et al. 2003; Moretti and Zoghbi 2006). Mutations in the
X-linked gene encoding methyl-CpG-binding protein 2
(MeCP2), including missense, frameshift and nonsense
mutations and intragenic deletions, account for the condi-
tion in up to 96 % of Rett syndrome patients (Amir et al.
1999; Moretti and Zoghbi 2006). Interestingly, the increase
in MeCP2 dosage due to duplications of the locus and
surrounding areas also causes the neurological disorder
Lubs X-linked mental retardation syndrome (MRXSL,
MIM #300260) (Van Esch et al. 2005). MeCP2 mutations
in Rett syndrome patients arise in the germline although the
phenotypic alterations in the neurological system appear at
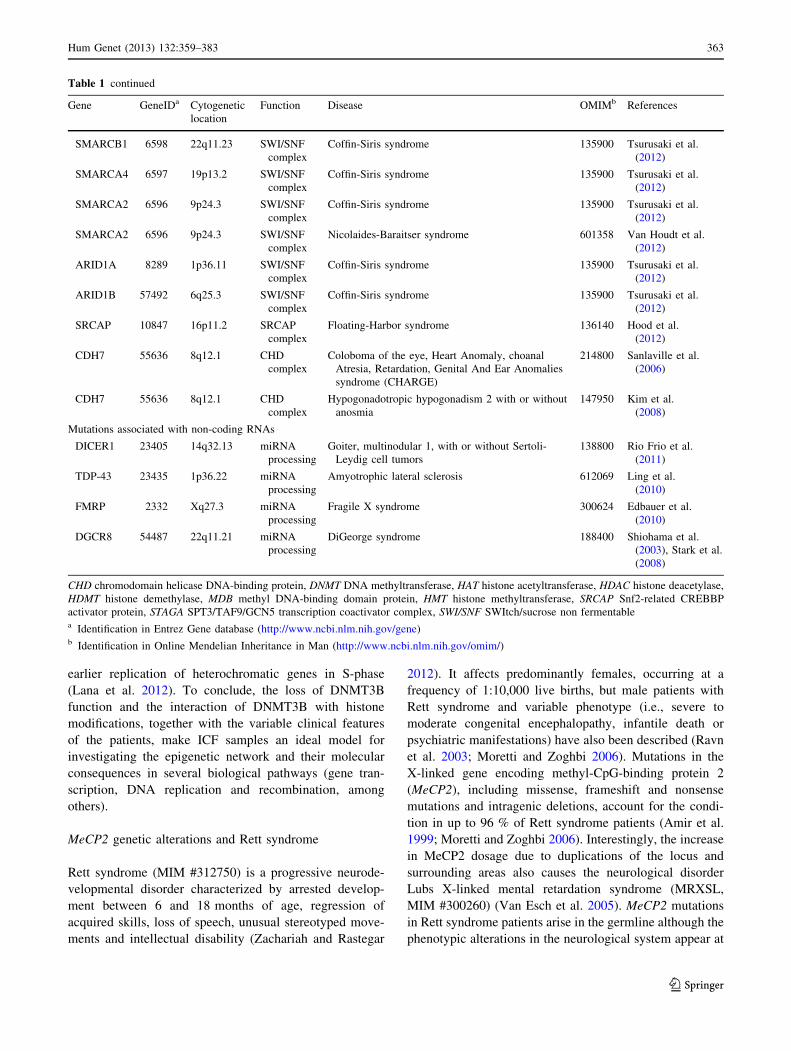
Table 1 continued
Gene GeneIDa Cytogenetic
location
Function Disease OMIMb References
SMARCB1 6598 22q11.23 SWI/SNF
complex
Coffin-Siris syndrome 135900 Tsurusaki et al.
(2012)
SMARCA4 6597 19p13.2 SWI/SNF
complex
Coffin-Siris syndrome 135900 Tsurusaki et al.
(2012)
SMARCA2 6596 9p24.3 SWI/SNF
complex
Coffin-Siris syndrome 135900 Tsurusaki et al.
(2012)
SMARCA2 6596 9p24.3 SWI/SNF
complex
Nicolaides-Baraitser syndrome 601358 Van Houdt et al.
(2012)
ARID1A 8289 1p36.11 SWI/SNF
complex
Coffin-Siris syndrome 135900 Tsurusaki et al.
(2012)
ARID1B 57492 6q25.3 SWI/SNF
complex
Coffin-Siris syndrome 135900 Tsurusaki et al.
(2012)
SRCAP 10847 16p11.2 SRCAP
complex
Floating-Harbor syndrome 136140 Hood et al.
(2012)
CDH7 55636 8q12.1 CHD
complex
Coloboma of the eye, Heart Anomaly, choanal
Atresia, Retardation, Genital And Ear Anomalies
syndrome (CHARGE)
214800 Sanlaville et al.
(2006)
CDH7 55636 8q12.1 CHD
complex
Hypogonadotropic hypogonadism 2 with or without
anosmia
147950 Kim et al.
(2008)
Mutations associated with non-coding RNAs
DICER1 23405 14q32.13 miRNA
processing
Goiter, multinodular 1, with or without Sertoli-
Leydig cell tumors
138800 Rio Frio et al.
(2011)
TDP-43 23435 1p36.22 miRNA
processing
Amyotrophic lateral sclerosis 612069 Ling et al.
(2010)
FMRP 2332 Xq27.3 miRNA
processing
Fragile X syndrome 300624 Edbauer et al.
(2010)
DGCR8 54487 22q11.21 miRNA
processing
DiGeorge syndrome 188400 Shiohama et al.
(2003), Stark et al.
(2008)
CHD chromodomain helicase DNA-binding protein, DNMT DNA methyltransferase, HAT histone acetyltransferase, HDAC histone deacetylase,
HDMT histone demethylase, MDB methyl DNA-binding domain protein, HMT histone methyltransferase, SRCAP Snf2-related CREBBP
activator protein, STAGA SPT3/TAF9/GCN5 transcription coactivator complex, SWI/SNF SWItch/sucrose non fermentablea Identification in Entrez Gene database (http://www.ncbi.nlm.nih.gov/gene)b Identification in Online Mendelian Inheritance in Man (http://www.ncbi.nlm.nih.gov/omim/)
Hum Genet (2013) 132:359–383 363
123

early postnatal stages (Zachariah and Rastegar 2012).
Recently, abrogation of MeCP2 function in adult mice has
been found to result in severe neurological symptoms
commonly observed in Rett syndrome, such as global
shrinkage of the brain, increased neuronal cell density,
retraction of dendritic arbors, reduction of synaptic proteins
and altered astrocytic development, among others (Nguyen
et al. 2012). This is further evidence of the involvement of
MeCP2 in regression from a normal mature brain to a Rett-
like brain. This dynamism is of vital importance and sug-
gests opportunities for reverting the Rett phenotype. In this
regard, Rett syndrome is not a neurodegenerative disorder,
neurons do not die, opening the opportunities of phenotypic
reversion by means of MeCP2 restoration (Giacometti et al.
2007; Guy et al. 2007; Tropea et al. 2009).
MeCP2 protein is widely expressed in several tissues but
the highest level of expression has been observed in the
brain (Zachariah and Rastegar 2012). Although a higher
level of MeCP2 expression has been described in neurons,
especially in postmitotic neurons, deregulation of MeCP2
expression in glia cells also contributes to the progression
of Rett syndrome (Ballas et al. 2009). MeCP2 was initially
identified as a transcriptional repressor that binds to
methylated CpG dinucleotides and recruits corepressors
such as mSin3 and HDACs (Jones et al. 1998). However, in
recent years, studies have concluded that MeCP2 may act
either as a repressor or an activator, depending on its
interaction proteins (Chahrour et al. 2008; Yasui et al.
2007). Chahrour et al. (2008) examined the gene expres-
sion profiles in the hypothalamus of mice lacking or
overexpressing MeCP2 and, contrary to expectation, con-
firmed that MeCP2 occupancy preferentially occurs in
active genes (85 %) and is associated with binding to the
transcriptional activator CREB1. In accordance with this
finding, similar genome-wide analyses of MeCP2 binding,
CpG methylation and gene expression showed that MeCP2
binds methylated and unmethylated DNA preferentially to
actively expressed genes (Yasui et al. 2007). Several
MeCP2 target genes (UBE3A, DLX5, BDNF and PRODH)
have been identified (Samaco et al. 2005; Horike et al.
2005; Chang et al. 2006; Urdinguio et al. 2008), although
no direct link between MeCP2-dependent expression of
these genes and the phenotypic abnormality of Rett syn-
drome has so far been found. A role for oxidative stress as
a mechanism underlying the Rett phenotype has been
suggested (De Felice et al. 2012) mainly on the basis of
two observations: (i) oxidation of either a single guanine to
8-oxoG or of a single 5mC to 5hmC, significantly inhibits
(by at least an order of magnitude) binding of MeCP2 to
the oligonucleotide duplex (Valinluck et al. 2004); (ii)
several MeCP2 target genes affect the oxidative stress
response, such as BDNF, CREB or Prodh (Chang et al.
2006; Urdinguio et al. 2008). Finally, analysis of the
posttranslational modifications of MeCP2 proteins could
also provide insight into the effect on synaptic plasticity
mediated by the regulation of specific genes. Recent work
has shown that phosphorylation of MeCP2 at serine 421 is
induced by membrane depolarization and leads to the
regulation of BDNF transcription (Zhou et al. 2006).
Interestingly, a mouse model showed that phosphorylation
at serine 421 has widespread effects on synaptic plasticity
(Li et al. 2011). Neuronal activity also influences dephos-
phorylation of MeCP2 at serine 80, which alters the tran-
scription of several genes (Tao et al. 2009). Considering
these results together suggests that further research is
warranted into the role of these MeCP2 posttranslational
modifications (and other phosphorylation sites) in neuronal
plasticity. It should be stressed that recent evidence sug-
gests that MeCP2 is not only involved in transcriptional
regulation, but also possibly in RNA splicing (Young et al.
2005), chromatin condensation (similar to H1 function)
(Ishibashi et al. 2008) and the silencing of repetitive ele-
ments (Muotri et al. 2010). These findings demonstrate that
MeCP2 function and its involvement in Rett syndrome
might be more complex than previously appreciated.
Finally, MeCP2 mutations are also linked to a broad
spectrum of neurological disorders (Van Esch et al. 2005;
Villard 2007), and to autism (Carney et al. 2003), Angel-
man syndrome (Watson et al. 2001) and Prader–Willi
syndrome (Samaco et al. 2004). As mentioned before,
duplications or triplications on chromosome Xq28 con-
taining the MeCp2 region are also associated with the Lubs
X-linked mental retardation syndrome (Van Esch et al.
2005). Taking together, it must be highlighted that both
down- and overexpression of MeCP2 result in altered
neuron function, an aspect that must be especially con-
sidered for therapeutic purposes based on MeCP2 restora-
tion. Although it is clear that MeCP2 deficiency affects the
brain function, a definitive molecular pathology of the
MeCP2-associated disorders remains elusive. In this con-
cern, progresses are currently being carried out mainly due
to the development of appropriate experimental systems,
such as stem cell-based system allowing the synchronous
differentiation of neuronal progenitors in wild-type or
mutant MeCP2 (Yazdani et al. 2010) or by the develop-
ment of several mouse models that reproduce many traits
of Rett syndrome (Na et al. 2012). Hopefully their findings
will help us better understand the many facets of the
pathobiology of the disease.
Genetic alteration of histone modifiers
The histone modification network is very complex.
Post-transcriptional histone modifications can occur in
various histone proteins (e.g., H2B, H3, H4) and variants
364 Hum Genet (2013) 132:359–383
123

(e.g., H3.3) and affect different histone residues (lysine,
arginine, serine) located in their N-terminal tails (Esteller
2008; Bannister and Kouzarides 2011). Several chemical
groups [methyl, acetyl, phosphate, small ubiquitin-related
modifier (SUMO) and ADP-ribose] may be added in dif-
ferent degrees depending on the chemical modification
(mono-, di- or trimethylation) (Bannister and Kouzarides
2011). Cross-talk between histone marks can occur within
the same residue, in the same tail or among different his-
tone tails (Portela and Esteller 2010) and, as a conse-
quence, the functional significance of histone modifications
depends on the combination of all marks (the ‘‘histone
code’’). Furthermore, we must not forget that an additional
level of complexity exists due to the communication
between the epigenetic marks involving DNA, histone and
chromatin-related proteins. Histone modifications are
involved in gene transcription, although the consequence of
each mark depends on the residue affected and the type of
modification. In general, acetylation of lysines is associated
with transcriptional activation. However, methylation of
lysine 4 of the H3 histone is associated with active tran-
scription whereas methylation of lysines 9 and 27 is
associated with gene silencing (Bannister and Kouzarides
2011). Gene transcription is not the only characteristic that
is controlled in this way; histone modifications are a
mechanism for controlling chromatin structure and they
also affect more global biological processes such as DNA
repair, DNA replication, alternative splicing and chromo-
some condensation (Portela and Esteller 2010).
Addition of chemical groups to histone residues is a very
dynamic and reversible process catalyzed by two sets of
enzymes (and their protein complexes) that have antago-
nistic activities, enzymes that covalently attach the chem-
ical groups and others for removing them (Fig. 1).
Acetyltransferases (HATs) and histone deacetylases
(HDACs) are among the least specific histone modifiers
because they are able to modify several residues. Con-
versely, histone methyltransferases (HMTs), histone
demethylases (HDMTs) and kinases have higher specificity
(Portela and Esteller 2010). Genetic alterations of histone
modifier enzymes are frequently linked to human diseases.
In this regard, aberrations in the histone modification
profiles associated with cancer could be a consequence of
the genetic disruption of the epigenetic machinery
Normal development
…K… DRLVKRHRKAGGKGLGKGGKGRGS91 135812161820
- N- terminal
P PMeAcAc
Ac
MeAc
Ac
Me
CBP
P300
pCAFSWI/SNF
DNMT3B
PRMT1
CARM1
HDAC1HDAC2
SINA3MeCP2
CH3
H4
Pri-mRNA
Pre-mRNA
DroshaXPO5/mRNA
DICER
miRNA
RISC
Genetic alterations of epigenetic genes
Writers
Ac
Me
P
5mC DNMTs
HATs
HMTs
Kinases
Readers
Ac
Me
Bromodomains
ChromodomainsPHD domainsPWWP domainsTudor domains
5mC MBDs
Erasers
Ac
Me
P
HDACs
HDMTs
Phosphatases
Epigenetic therapies
HDAC inhibitorsHDMT inhibitors
DNA demethylating agentsOthers
miRNA
Fig. 1 Epigenetic mechanisms disrupted in human disorders. Epige-
netic mechanisms regulate chromatin function and cell identity.
Appropriate activity of enzymes controlling DNA methylation,
histone modifications and non-coding RNAs controls the temporal
and spatial patterns of gene expression, DNA repair and DNA
replication. Their deregulation may contribute to human diseases.
Epigenetic-based therapies, such as histone deacetylase inhibitors, can
partially alter the phenotype of the disease by recovering the aberrant
epigenetic patterns, and are a promising area in pharmacological
research. Chemical modifications at histone H4 are shown as a
representative example of histone marks. 5-mC; 5-methylcytosine; Achistone acetylation, DNMTs DNA methyltransferases, HATs histone
acetyltransferases, HDACs histone deacetylases, HDMTs histone
demethylases, HMTs histone methyltransferases, MBDs methyl-
binding domain proteins, Me histone methylation, P histone phos-
phorylation, PHD plant homeodomain, PWWP proline–tryptophan–
tryptophan–proline domain, XPO5 exportin-5
Hum Genet (2013) 132:359–383 365
123

(Berdasco and Esteller 2010). Several hematological
malignancies can be associated with chromosomal trans-
locations in the coding region of HATs (i.e., CBP-MOZ) or
HMTs (i.e., mixed-lineage leukemia 1 (MLL1), or nuclear
receptor binding SET domain protein 1 (NSD1). In solid
tumors, both HMT genes [such as EZH2, mixed-lineage
leukemia 2 (MLL2)] and DNMTs [i.e., Jumonji domain-
containing protein 2C (JMJD2C/GASC1)] are known to be
amplified (Berdasco and Esteller 2010). In this section we
will focus on non-tumoral human diseases in which the
epigenetic profile changes as a consequence of genetic
alterations in histone modifiers (Table 1).
MYST4 acetyltransferase (KAT6B) mutations
in Genitopatellar syndrome and Say-Barber-Biesecker-
Young-Simpson syndrome
Genitopatellar syndrome (GPS, MIM #606170) is a rare
skeletal dysplasia consisting of microcephaly, severe psy-
chomotor retardation and craniofacial defects, associated
with congenital flexion contractures of the lower extremi-
ties, abnormal or missing patellae, and urogenital anoma-
lies (Reardon 2002). To date it has been described in only
18 subjects (Campeau et al. 2012a). Whole-exome
sequencing identified mutations in the KAT6B acetyl-
transferase that lead to protein truncation (Campeau et al.
2012a; Simpson et al. 2012). All mutations are heterozy-
gous, exhibit autosomal dominant inheritance and occur in
the proximal portion of the last exon (Campeau et al.
2012a). KAT6B is a member of the MYST family of
proteins containing a conserved acetyltransferase domain
(Champagne et al. 1999). The absence of decreased
expression of the KAT6B transcript has been described in
GPS patients (Campeau et al. 2012a); however, GPS sub-
jects were characterized by decreased global acetylation of
histone H3 and H4 (Simpson et al. 2012).
Heterozygous mutations of KAT6B have also been
described in patients with Say-Barber-Biesecker-Young-
Simpson syndrome (SBBYS, MIM #603736) (Clayton-
Smith et al. 2011). The main clinical features associated
with the syndrome are distinctive facial appearance, severe
hypotonia and feeding problems, associated skeletal prob-
lems, cardiac defects, severe intellectual disability, delayed
motor milestones, and significantly impaired speech
(Clayton-Smith et al. 2011). Unlike with GPS syndrome,
mutations in SBBYS patients are located throughout the
gene or more distally in the last exon. Although GPS and
SBBYS share several clinical features, such as severe
intellectual disability, cardiac defects and genital abnor-
malities, the range of phenotypic alteration varies between
the two syndromes (Campeau et al. 2012b). Campeau and
collaborators created a database for establishing correla-
tions between phenotype and genotype in patients carrying
KAT6B mutations. Their preliminary results suggested that
the common features are a consequence of haploinsuffi-
ciency of the C-terminal region, whereas the unique phe-
notypes of GPS could arise from the expression of a
truncated protein that acquires new cellular functions
(Campeau et al. 2012b).
The molecular mechanisms of abrogation of KAT6B
function that lead to defective neural development are
not currently known. The Querkopf mouse model that
only expressed a 5 % of the MYST4 levels quantified for
wild-type mice leads to a phenotype commonly observed
in human syndromes: brain development defects, facial
dysmorphisms and alteration of bone growth (Thomas
et al. 2000), supporting the role of KAT6B in cerebral
cortex development. According to this idea, KAT6B is
expressed in developing cerebral cortex, adult neural
stem cells, osteoblasts and germ cells (Merson et al.
2006). A study performed in one Noonan-like phenotype
patient, with a chromosomal breakpoint in KAT6B
resulting in 50 % gene expression, and in Querkopf mice,
described a reduction of H3 acetylation that specifically
dysregulates the expression of genes in the MAPK
pathway (Kraft et al. 2011).
Genetic disruption of ep300/CBP acetyltransferases
in Rubinstein–Taybi syndrome
The Rubinstein–Taybi syndrome (RSTS, MIM #180849) is
a well-defined disease characterized by postnatal growth
deficiency, microcephaly, specific facial characteristics,
broad thumbs, big toes and intellectual disability. An
increased risk of tumors (mainly leukemia in childhood and
meningioma in adulthood) has been observed (Hennekam
2006). Although the exact molecular etiology of RSTS is
not clearly understood, it is widely accepted that RSTS is
associated with breakpoints, translocations, mutations and
microdeletions of chromosome 16p13.3 (Lacombe et al.
1992). Petrij et al. (1995) were the first to report that
mutations in the gene encoding the CREB binding protein
(CBP), located in the aforementioned region, could cause
RSTS. Recent investigations employing larger series of
RSTS samples detected CBP mutations in 45–55 % of
patients (Roelfsema et al. 2005; Tsai et al. 2011).
Furthermore, CBP has a homolog located at 22q13.2,
known as the E1A-binding protein p300 (p300) (Arany
et al. 1994). The exact frequency of genetic alterations of
p300 in RTS is not yet known, but some sources estimate it
to be 3 % (Bartholdi et al. 2007; Bartsch et al. 2010). Since
a cytogenetic or molecular abnormality in p300/CBP could
be detected in about 55 % of patients, further work to
explain the cause of the syndrome in the remaining 45 % of
patients without a ‘‘classic’’ genetic abnormality is still
needed.
366 Hum Genet (2013) 132:359–383
123

CREB binding protein was first identified as a nuclear
transcription coactivator that binds specifically to CREB
when it is phosphorylated (Chrivia et al. 1993), while p300
was originally described by protein-interaction assays with
the adenoviral E1A oncoprotein (Eckner et al. 1994).
Although both proteins are highly homologous (63 %
homology at the amino acid level) and have common
interaction partners, they have distinct cellular functions
and cannot always replace one another (Viosca et al. 2010).
There are two biological mechanisms whereby defects in
p300/CBP function could cause the RTS symptoms:
(i) CBP and p300 proteins act as cofactors for over 300
transcriptional factors, including several regulators
involved in neuronal activity, such as c-Fos, c-Jun, CREB
or NF-jß, known oncoproteins (myb), transforming viral
proteins (E1A, E6, large T antigen) and tumor-suppressor
proteins (p53, E2F, RB, Smads, RUNX, BRCA1) (Chan
and La Thangue 2001; Kasper et al. 2011); (ii) both pro-
teins have HAT activity that targets the N-terminal tails of
histones and contributes to transcriptional activation by
relaxing the structure of the nucleosomes (Ogryzko et al.
1996). CBP and ep300 have some common activities, such
as the acetylation of H4K5, H3K14, H3K18, H3K27 and
H3K56 (Das et al. 2009; Jin et al. 2011). However, they
also have unique properties such as substrate specificity
profiles that could explain functional differences of both
enzymes (McManus and Hendzel 2003). RSTS disorder
has been modeled in mice, and several heterozygous
cbp ± mice (homozygous cbp -/- mutants are embryonic
lethal) have been generated (Bourtchouladze et al. 2003;
Alarcon et al. 2004). These model animals exhibit deficits
in long-term memory and cognitive impairments reminis-
cent of human RSTS neural symptoms, confirming the role
of CBP in the etiology of the disease (Alarcon et al. 2004;
Korzus et al. 2004). At the molecular level, these mice
have reduced HAT activity, decreased acetylation of spe-
cific histone proteins and impaired CBP-dependent gene
expression (Alarcon et al. 2004; Korzus et al. 2004). The
role of CBP in neural differentiation and development has
been recently demonstrated in CBP genetically modified
models (Wang et al. 2010). Phosphorylation of CBP by
atypical protein kinase C f is necessary for CBP binding to
neural promoters, followed by histone acetylation and
transcriptional activation, leading to neural differentiation
of stem cell precursors (Wang et al. 2010). This mechanism
could explain how CBP alterations can result in cognitive
dysfunction of RSTS patients.
HDAC4 histone deacetylase mutations
in Brachydactyly-mental retardation syndrome
Brachydactyly-mental retardation syndrome (BDMR, MIM
#600430) is a complex disease that presents a wide
spectrum of clinical features, such as intellectual disabilities,
developmental delays, sleep disturbance, craniofacial and
skeletal abnormalities (including brachydactyly type E),
cardiac defects and autism (Aldred et al. 2004). Hetero-
zygous mutations in the HDAC4 deacetylase gene located
on chromosome 2q37.2 have been reported in BMRS
subjects (Williams et al. 2010; Morris et al. 2012). HDAC4
acts as a corepressor for transcription factors regulating the
expression of genes from the osteogenic, chondrogenic,
myogenic and neurogenic differentiation pathways (Miska
et al. 2001; Arnold et al. 2007; Chen and Cepko 2009).
HDAC4 is essential for the repression of RUNX2 and
MEF2 transcription factors in normal bone development
(Arnold et al. 2007). Indeed, mice with a deleted MEFC2
gene have impaired chondrogenic and osteogenic devel-
opment that antagonizes the phenotype of the Hdac4
mutant mice, which is similar to the human BDMR phe-
notype (Arnold et al. 2007; Rajan et al. 2009). These
results suggest that haploinsufficiency of HDAC4 causes
BDMR through its ability to regulate important master
genes of cellular differentiation.
Mutations in histone methyltransferase EHMT1
in Kleefstra syndrome
Kleefstra syndrome (MIM #610253), previously known as
9q subtelomeric deletion syndrome, is characterized
by severe intellectual disability, hypotonia, brachy(micro)
cephaly, epileptic seizures, flat face with hypertelorism,
synophrys, anteverted nares, everted lower lip, carp mouth
with macroglossia, and heart defects (Willemsen et al.
2012). The mutational landscape of Kleefstra patients
includes a microdeletion in the distal long arm of chro-
mosome 9q or intragenic loss of function mutations in the
histone methyltransferase EHMT1 (Kleefstra et al. 2006,
2009), both leading to haploinsufficiency of EHMT1.
EHMT1 is a specific HMT for lysine 9 at histone H3 and is
involved in gene repression (i.e., the NF-kB gene) (Ogawa
et al. 2002; Ea et al. 2012). It has not been established how
EHMT1 disruption results in the phenotypic skills of
Kleefstra syndrome, but recent research on EHMT mutants
in Drosophila melanogaster demonstrates that learning and
memory defects could be restored after re-expression of
EHMT (Kramer et al. 2011). Interestingly, the same
authors have recently identified new genetic mutations
affecting epigenetic modifiers in Kleefstra syndrome sub-
jects without mutations in the EHMT1 gene, including the
methyl-binding domain MBD5, the histone methyltrans-
ferase MLL3 or the chromatin-remodeling factor
SMARCB1 (Kleefstra et al. 2012). These findings highlight
the crucial role of epigenetic modifiers in brain develop-
ment and strongly emphasize the need to explore this area
of research further.
Hum Genet (2013) 132:359–383 367
123

NSD1 histone methyltransferase genetic alterations
in Sotos syndrome
Sotos syndrome (MIM #117550) is an autosomal dominant
condition characterized by overgrowth that results in tall
stature and macrocephaly, a distinctive facial appearance,
learning disability (Lapunzina 2005; Tatton-Brown and
Rahman 2007), and an increased incidence of malignant
neoplasms (Rahman 2005). The distinctive head shape and
size has led to Sotos syndrome sometimes being called
cerebral gigantism (Tatton-Brown and Rahman 2007).
Most cases of NSD1 mutational mechanisms, including
truncating, missense and splice-site mutations and
deletions, result in loss of function of the NSD1 protein
(Tatton-Brown and Rahman 2007). Mutations in the
nuclear receptor SET domain containing protein-1 gene
(NSD1), which encodes a histone methyltransferase of
lysine residues H3K36 and H4-K20, are found in patients
exhibiting the clinical symptoms of Sotos syndrome
(Kurotaki et al. 2002; Rayasam et al. 2003). Recently,
mutations in the NFIX gene were associated with a Sotos
syndrome-like phenotype without NSD1 mutations (Yon-
eda et al. 2012).
NSD1 is essential for early postimplantation of embryos
and NSD1 homozygous mutants are embryonic lethal,
although heterozygous mutant NSD1 are viable and fertile
(Rayasam et al. 2003). The NSD1 protein contains a
su(var)3-9, enhancer-of-zeste, trithorax (SET) domain
responsible for HMT activity and other functional domains,
including plant homeodomain (PHD) and proline–trypto-
phan–tryptophan–proline (PWWP) domains, both of which
are involved in a protein–protein interaction (Kurotaki
et al. 2001). Additionally, the potential of NSD1 to mono-
and dimethylate lysines K218 and K221 of the p65 subunit
of the immune response gene NF-kB has been noted (Lu
et al. 2010). NSD1 has previously been shown to interact
with nuclear receptors, such as Nizp1, a DNA-binding
transcriptional repressor (Huang et al. 1998; Nielsen et al.
2004), but to date there has been no evidence clarifying
whether NSD1 mutations contribute to deregulation of
brain function. It has become clear that NSD1 is a versatile
protein that can act as a corepressor or coactivator,
depending on the cellular context (Huang et al. 1998;
Pasillas et al. 2011). According to this idea, binding of
NSD1 PHD domains to target genes is guided by the
presence of specific histone marks of promoters (Pasillas
et al. 2011), specifically methylation at lysine H3K4 and
H3K9. By binding to trimethylated H3K9, NSD1 can
recognize genes that are transcriptionally repressed and
interact with other repression complexes (i.e., DNMT1 or
HP1), whereas its interaction with trimethylated H3K4
allows binding to active genes (Pasillas et al. 2011). In
addition, patients with genetic disruption of the NSD1 gene
have an increased risk of developing malignancy before
adulthood, including neuroblastoma, Wilms tumors and
hematological malignancies (Rahman 2005). NSD1 also
has a tumor-suppressor function (Berdasco et al. 2009).
NSD1 function is abrogated in neuroblastoma and glioma
cells by transcriptional silencing associated with CpG
island-promoter hypermethylation, and restoration of its
expression demonstrates that its tumor-suppressor features
are mediated by a mechanism dependent on MEIS1
expression (Berdasco et al. 2009).
Finally, occasional individuals have NSD1 defects that
overlap clinically with Sotos syndrome and other condi-
tions, such as Weaver syndrome 1 (WVS1, MIM #277590)
(Douglas et al. 2003). Beckwith–Wiedemann syndrome
(BWS, MIM #130650) is, like Sotos syndrome, an over-
growth syndrome. It is cause by deregulation of imprinted
growth-regulatory genes within the 11p15 region. Inter-
estingly, correlations between the two syndromes have
been found: first, unexplained Beckwith–Wiedemann
patients could be related to NSD1 deletions or mutations
(2/52 cases) and secondly, 11p15 anomalies (including the
KCNQ10T1 imprinting center) were identified in Sotos
syndrome cases (2/52) (Baujat et al. 2004; Mayo et al.
2012). A potential role for NSD1 in imprinting of the
11p15 region is suggested, although the molecular basis for
this association is not known.
EZH2 histone methyltransferase mutations
and Weaver syndrome
Weaver syndrome 2 (WVS2, MIM #614421) is an over-
growth syndrome characterized by tall stature, advanced
bone age, macrocephaly, hypertelorism, learning disabili-
ties and dysmorphic facial features (Weaver et al. 1974). A
predisposition to hematological malignancies has also been
reported (Basel-Vanagaite 2010). Heterozygous mutations
in the histone methyltransferase EZH2 gene on chromo-
some 7q36.1 have been identified in Weaver syndrome
patients (Gibson et al. 2012). EZH2 protein is a member of
the polycomb repressive complex 2 (PRC2), together with
SUZ12 and EED, which catalyses the trimethylation of
lysine H3K27 (Kirmizis et al. 2004). Mammalian EZH2
has critical roles in X-chromosome inactivations, genomic
imprinting during germline development, stem cell main-
tenance and cell lineage determination, including osteo-
genesis, myogenesis and hematogenesis (Chou et al. 2011;
Wyngaarden et al. 2011). A role for EZH2 in regulating the
circadian-clock functions has been suggested (Etchegaray
et al. 2006). In addition, mice with targeted mutant EZH2
beta cells have reduced beta cell proliferation and beta cell
mass (Chen et al. 2009), whereas mice with EZH2 mutant
satellite cells exhibit defects in muscle regeneration (Juan
et al. 2011). Some characteristics of these phenotypes are
368 Hum Genet (2013) 132:359–383
123

shared with that of human Weaver syndrome, such as
defects in limb development (Wyngaarden et al. 2011).
However, to date, few studies have provided any insight into
the specific contribution of EZH2 mutations in the Weaver
phenotype. As described above, some patients with Weaver
syndrome have a mutated NSD1 gene that is responsible for
the overgrowth Sotos syndrome (Douglas et al. 2003). There
are some clinical features common to both syndromes (i.e.,
developmental delay, overgrowth and macrocephaly), but
some features are specific to Weaver syndrome 2 (i.e., ret-
rognathia with a prominent chin crease or carpal bone age)
(Gibson et al. 2012). Indeed, EZH2 protein acts in the PI3K/
mTOR pathway, which has been associated with growth
defects. This suggests that the pathways through NSD1 and
EZH2 HTMs that contribute to overgrowth disorders can be
different (Tatton-Brown et al. 2011).
Histone methyltransferase (MLL2) and demethylase
(JMJD3) mutations in Kabuki syndrome
Kabuki syndrome 1 (MIM #147920) is an autosomal
dominant intellectual disability syndrome with additional
features, including a highly distinctive recognizable facial
phenotype characterized by long palpebral fissures with
eversion of the lateral third of the lower eyelids, a broad
and depressed nasal tip, large prominent earlobes, scoliosis,
short fifth finger, persistence of fingerpads, radiographic
abnormalities of the vertebrae, hands, and hip joints, and
others (Niikawa et al. 1981). Mutations in the histone
methyltransferase MLL2 gene are a major cause of Kabuki
syndrome (type 1) (Ng et al. 2010; Hannibal et al. 2011),
but mutations in the histone demethylase KDM6A have
also been found in Kabuki syndrome (type 2, MIM
#300867) (Lederer et al. 2012). MLL2 is a lysine H3K4-
specific histone methyltransferase that belongs to the SET1
family of proteins (Dillon et al. 2005), whereas KDM6A
(JMJD3) is a histone demethylase that specifically acts in
mono- di- and trimethylated lysine H3K27 (Hong et al.
2007; Lan et al. 2007). Both enzymes help regulate genes
from the myogenic lineage during embryogenesis (Aziz
et al. 2010; Seenundun et al. 2010). The identification of
mutations in MLL2 and KDM6A suggests a crucial effect
of altered histone methylation profiles on the phenotype of
Kabuki syndrome.
Histone demethylase PHF8 mutations in Siderius
X-linked mental retardation syndrome
Siderius X-linked mental retardation syndrome (MRXSSD,
MIM #300263) is an inherited condition that was first
described in 1999 as a heterogeneous intellectual disability
syndrome associated with cleft lip/cleft palate (Siderius
et al. 1999). Mutations in the PHD finger protein 8 (PHF8)
gene located on the Xp11 chromosome have been identi-
fied in subjects with MRXSSD (Laumonnier et al. 2005;
Abidi et al. 2007; Koivisto et al. 2007). The PHF8 protein
contains a PHD-type domain zinc finger domain and a
Jumanji domain, the latter conferring histone demethylat-
ing catalytic activity with specificity for: mono- and
dimethylated histone H3 at lysine K9 (Yu et al. 2010) and
mono-methyl histone H4 at lysine K20 (Qi et al. 2010).
PHF8 transcript is strongly expressed in the embryonic and
early postnatal steps of brain development (Laumonnier
et al. 2005), implying a connection between PHF8 muta-
tions and the MRXSSD phenotype. Some evidences of the
functional role of PHF8 in different experimental models
exist (Liu et al. 2010; Qi et al. 2010). In zebrafish, it has
been described that PHF8 regulates brain apoptosis and
craniofacial development through the transcriptional gene
regulation (Qi et al. 2010); in Caenorhabditis elegans, a
RNA interference-based functional genomic project iden-
tified PHF8 as a gene involved in controlling cellular
growth and differentiation during embyogenesis (Fernan-
dez et al. 2005); in human HeLa cells, PHF8 deficiency by
siRNA mechanisms leads to a delay in G1/S transition and
its dissociation from chromatin in early mitosis which
demonstrates an active role of PHF8 in the control of cell
cycle (Liu et al. 2010). Interestingly, PHF8 interacts with
another MRXSSD protein, the transcription factor ZNF711
(Kleine-Kohlbrecher et al. 2010). PHF8 and ZNF11 pro-
teins share a set of target genes, being of special relevance
the interaction of both proteins with JARID1C (KDM5C)
(also involved in alterations of the intelectual ability)
(Kleine-Kohlbrecher et al. 2010; Claes et al. 2000). In
addition to MRXSSD syndrome, defects affecting the
chromosomal region of PHF8 (a larger Xp11.22 deletion
that includes the FAM120C and WNK3 genes) have also
been associated with autism (Qiao et al. 2008).
Histone demethylase JARID1C (KDM5C) mutations
in Claes-Jensen X-linked mental retardation syndrome
Mutations in the histone demethylase JARID1C (KDM5C)
were first identified in individuals with X-linked mental
retardation syndrome with additional features of progres-
sive spastic paraplegia, facial hypotonia, aggressive
behavior and strabismus (MIM #300534) (Claes et al.
2000; Jensen et al. 2005). Heterogeneous clinical features
associated with XLMR and mutated KDM5C have been
consistently identified since then (Abidi et al. 2008; Santos-
Reboucas et al. 2011; Ounap et al. 2012). KDM5C gene
encodes a specific histone H3 demethylase at lysine K4
(Tahiliani et al. 2007). It is ubiquitously expressed
although fetal brain tissues have higher KDM5C expres-
sion levels than other tissues (Xu et al. 2008). The tran-
scriptional repressor activity of KDM5C is mediated by the
Hum Genet (2013) 132:359–383 369
123

Re-1 silencing transcription factor (REST) complex
(Tahiliani et al. 2007). Interestingly, knock-out of the
KDM5C complex results in increased trimethylation at
H3K4 and gain of expression of SCN2A and SYN1 neuro-
logical genes, and provides evidence of the contribution of
KDM5C complex to X-linked mental retardation defects
(Tahiliani et al. 2007).
Mutations in chromatin remodelers
Nucleosome positioning and, consequently, DNA accessi-
bility may be controlled by mechanisms that are indepen-
dent of histone-modifying enzymes. Several groups of
protein complexes (‘‘chromatin-remodeling complexes’’)
are known to restructure nucleosomes in an ATP hydro-
lysis-dependent manner. To date, four families of chro-
matin remodelers have been described in eukaryotes: SWI/
SNF, ISWI, NURD/Mi-2/CHD, and INO80/SWR1 (Har-
greaves and Crabtree 2011). The ATPase domain is a
common feature, but the composition of the different
subunits comprising the complex is highly variable. In a
similar manner, each ATPase domain may be targeted to
specific domains (i.e., bromodomain, DNA helicase, etc.).
Together, the binding affinity and the complex composition
confer unique features on chromatin remodelers in a wide
range of biological processes and genomic contexts.
SWI/SNF is one of the best studied chromatin-remod-
eling complexes in human cells and is composed of at least
15–20 subunits, including ATPases, catalytic subunits (i.e.,
SMARCA2 and SMARCA4) and structural components
involved in target recognition or stabilization functions
(i.e., ARID1A, ARID1B and SMARCC1) (Hargreaves and
Crabtree 2011). All members of this complex contain either
SMARCA2 (also known as Brahma protein) or SMARCA4
(also known as BRG1) as a catalytic unit. The two proteins
share 75 % amino acid homology (Santen et al. 2012). The
SWI/SNF complex plays a crucial role in cell differentia-
tion (Ho et al. 2009), cell cycle (Nagl et al. 2007) and DNA
repair (Park et al. 2006). In the human ISWI (imitation
switch) family of chromatin remodelers, the catalytic sub-
unit is represented by SNF2H and SNF2L proteins (Flaus
and Owen-Hughes 2011). ISWI complexes participate in
biological functions such as chromatin assembly, nucleo-
some spacing, DNA replication and activation or repres-
sion of transcriptional regulation (Erdel and Rippe 2011).
The CHD family also contains a SNF2L ATPase domain
together with tandem chromodomains in the N-terminal
region (Murawska and Brehm 2011). CHD complexes are
highly versatile and although they are involved in tran-
scriptional regulation, various CHD regulatory complexes
are involved in the initiation, elongation or termination of
transcription. Furthermore, specific CHD complexes, such
as Mi-2/nuRD, contain HDAC and MBD proteins in the
same complex (Murawska and Brehm 2011). The INO80
subfamily is the most recently identified SWI/SNF family
of chromatin remodelers. Mammalian INO80 complex
comprises the INO80 catalytic unit and Snf2-related CBP
activator protein (SRCAP) and p400 subunits (Morrison
and Shen 2009). The INO80 subfamily is the most evolu-
tionarily conserved of all the chromatin-remodeling com-
plexes due to the high degree of homology of its ATPase
subunit (Morrison and Shen 2009). Apart from regulation
of transcription, the INO80 complex is involved in genome
stability pathways, such as DNA repair, replication, telo-
mere regulation and centromere stability (Ho and Crabtree
2010). In summary, ATP-dependent enzymes that remodel
chromatin are important regulators of chromatin dyna-
mism. Evidence is emerging that alterations in such chro-
matin-remodeling complexes have consequences for
normal development. Some examples of genetic mutations
of chromatin-remodeling complexes in human diseases are
summarized in this section (Table 1).
ATXN7 mutation in Spinocerebellar Ataxia 7
Spinocerebellar Ataxia Type 7 (SCA7; MIM 164500) is an
autosomal dominant inherited neurodegenerative disorder
characterized by progressive cerebellar ataxia, including
dysarthria and dysphagia, and cone-rod and retinal dys-
trophy with progressive central visual loss resulting in
blindness in affected adults. The disease is caused by an
expanded CAG trinucleotide repeat encoding a polygluta-
mine tract in ataxin-7 (ATXN7) gene, from 4 to 35 repeats
in normal gene to a variable expansion of 36–306 repeats in
pathogenic ATXN7 variants (Garden and La Spada 2008).
ATXN7 protein is a transcription factor with important
roles in chromatin regulation through its effect on histone
modification and histone deubiquitination. It is a member
of the transcription coactivator complex STAGA (SPT3/
TAF9/GCN5) with acetyltransferase activity, but may also
be found in the USP22 deubiquitination complex (Sopher
et al. 2011). Although the nuclear expression of ATXN7 is
necessary for transcriptional regulation, a functional role
for cytoplasmic ATXN7 in the regulation of cytoskeletal
dynamics (mediated by its interaction with microtubules)
has recently been proposed (Nakamura et al. 2012).
Defects of the nuclear ATXN7 gene have been correlated
with the SCA7 phenotype (Chen et al. 2012a, b), although
the molecular mechanisms underlying the disease are not
clearly understood and the effect of epigenetic dysregula-
tion of target genes is still a matter of debate. Some results
from yeast and mice indicate that loss of the Gcn5 ace-
tyltransferase function triggered by polyQ-Atxn7, resulting
in chromatin structure changes, could be involved in the
SCA7 phenotype (Yoo et al. 2003; McMahon et al. 2005;
370 Hum Genet (2013) 132:359–383
123

Helmlinger et al. 2006). In contrast, loss of Gcn5 functions
in mice bearing polyQ-Atxn7 accelerates neuronal dys-
function in a mechanism that is independent of gene
expression changes (Chen et al. 2012a, b). Deciphering the
exact causal consequences of ATXN7 dysregulation in
SCA7 disease through its role as nuclear transcription
regulator or cytoplasmic function will need further
research.
ATRX mutations in alpha-thalassemia X-linked mental
retardation syndrome
ATR-X syndrome (MIM #301040) is an X-linked disorder
comprising severe psychomotor retardation, characteristic
facial features, genital abnormalities, and the blood disease
alpha-thalassemia (Gibbons et al. 1995). Mutations in the
ATRX gene located at Xq21.1 and coding for a member of
the SWI/SNF chromatin-remodeling family of proteins
underpin the molecular genetics of the disease (Gibbons
et al. 1995). The X-linked mental retardation-hypotonic
facies syndrome (MIM #309580) and the alpha-thalassemia
myelodysplasia syndrome (MIM #300448) are also asso-
ciated with mutations in the ATRX gene (Abidi et al. 2005;
Gibbons et al. 2003). The N-terminus contains a globular
domain, called ADD (ATRX-DNMT3-DNMT3L) that can
bind to the N-terminal of histone H3 (Argentaro et al.
2007). Indeed, ATRX is known to be required for the
incorporation of the histone H3.3 specifically at telomeric
sequences (Lewis et al. 2010). On the other hand, the
C-terminus contains seven helicase/ATPase domains that
share sequence homology with the SNF2 family of proteins
(Picketts et al. 1996). Through this domain, ATRX shows
in vitro ATP-dependent nucleosome remodeling and DNA
translocase activities (Gibbons et al. 2003). The ATRX
protein is expressed genome-wide, but is enriched at telo-
meric and subtelomeric regions (Law et al. 2010). In this
context, decreased ATRX expression is associated with
altered expression of telomere-associated RNA (Goldberg
et al. 2010) and the DNA-damage response during S-phase
at telomeric regions of pluripotent stem cells (Wong et al.
2010). The mechanisms by which ATRX is recruited to
telomeric regions are not fully understood, although ATRX
binding depends on trimethylation at K9H3 (Kourmouli
et al. 2005). It binds to tandem repeat sequences with
G-rich motifs and has been predicted to form non-B DNA
structures (Law et al. 2010). More importantly, the size of
the tandem repeats located in specific genes influences their
expression (Law et al. 2010), providing a molecular
explanation of how the same mutation at the ATRX gene
can result in different phenotypes. Furthermore, ATRX
mutations have been correlated with alterations in the DNA
methylation patterns of highly repeated sequences,
including rDNA, Y-specific satellite and subtelomeric
repeats (Gibbons et al. 2000). The interplay between
ATRX and the DNA methylation machinery is reinforced
by the discovery that the methyl-binding domain protein
MECP2 targeted the C-terminal helicase domain of ATRX
to heterochromatic foci (Nan et al. 2007). MeCP2 is also
mutated in Rett syndrome, so the finding suggests that
alteration of the MECP2–ATRX interaction leads to path-
ological changes that contribute to the intellectual dis-
ability phenotype observed in both syndromes.
Mutations in ERCC6 and Cockayne syndrome
Cockayne syndrome types A (CSA; MIM #216400) and B
(CSB; MIM #133540) are autosomal recessive disorders
caused by mutation in the ERCC8 and ERCC6 genes,
respectively (Henning et al. 1995; Laugel et al. 2010).
Approximately 62 % patients diagnosed with Cockayne
syndrome carry mutations of the ERCC6 gene (Laugel
et al. 2010). The syndromes are characterized by severe
postnatal growth failure, progressive neurological dys-
function and traits reminiscent of normal aging, such as
visual impairment and sensorineural hearing loss and loss
of adipose tissue (Licht et al. 2003). ERCC (excision
repair cross-complementing) genes are part of the nucle-
otide excision repair (NER) pathway, which are respon-
sible for removing DNA lesions such as UV-induced
DNA damage. ERCC6 is a nuclear protein containing a
SWI/SNF-like ATPase domain, a nucleotide-binding
domain and an ubiquitin-binding domain (Anindya et al.
2010). Apart from NER functions, ERCC6 is also
involved in transcription regulation, chromatin mainte-
nance and remodeling (Newman et al. 2006). At the
transcriptional level, CSB cooperates with the NurD/
CHD4 complex for controlling transcription of rRNA
genes (Xie et al. 2012). In this regard, CHD4/NuRD is
involved in maintaining silenced rRNA genes but in
permissive contexts (‘‘poised’’ for transcription), whereas
CSB mediates the transition from the permissive to the
active state (Xie et al. 2012). CSB-mediated activation
could be due, at least in part, in conjunction with the
CSB–G9a interaction, to an increase in trimethylated
K9H3 and recruitment of Pol-I to chromatin (Yuan et al.
2007). A new chromatin connection for CSB has recently
been proposed (Batenburg et al. 2012). Primary fibro-
blasts derived from a CSB patient had a dysfunctional
telomere structure (Batenburg et al. 2012). CSB knock-
down was accomplished with alterations in TERRA, a
large non-coding telomere repeat-containing RNA,
resulting in alterations of telomere length and integrity
(Batenburg et al. 2012). Finally, a role for CSB in con-
trolling key mitochondrial functions in addition to the
nucleolus function has been proposed (Berquist et al.
2012).
Hum Genet (2013) 132:359–383 371
123

SRCAP mutations and Floating-Harbor syndrome
Floating-Harbor syndrome (FHS; MIM #136140) is a rare
condition characterized by proportionate short stature,
delayed osseous maturation, language deficits and a typical
facial appearance. Mutations in the SNF2-related CBP
activator protein (SRCAP) cause the FHS syndrome (Hood
et al. 2012). SRCAP expression has also been linked to
cancer, whereby it positively modulates PSA antigen
expression and promotes proliferation in prostate cancer
cells (Slupianek et al. 2010) and potentiates Notch-
dependent gene activation (Eissenberg et al. 2005). With
regard to its molecular activity, SCARP catalyzes in vitro
incorporation of the histone variant H2A.Z into chromatin
(Ruhl et al. 2006), a histone with a well-known function in
transcription regulation and cell-cycle progression. As an
example, SRCAP expression in yeast is important for the
deposition of such histone variants in specific promoters
like SP-1, G3BP and FAD synthetase (Wong et al. 2007).
Interestingly, the demethylation effect after 5-Aza-20-deoxycytidine treatment, a drug approved by the US Food
and Drug Administration (FDA) for the treatment of
hematological malignancies, requires the activity of
SCARP to introduce H2A.Z, which facilitates the acqui-
sition of nucleosome-free regions (Yang et al. 2012). On
the other hand, SRCAP is also an interaction partner of the
histone acetyltransferase CBP, meaning that SRCAP-CBP
colocalization may occur at transcriptionally active sites
(Monroy et al. 2001).
CHD7 mutations and CHARGE syndrome
The acronym CHARGE (MIM # 214800) stands for colo-
boma of the eye, heart anomaly, choanal atresia, retarda-
tion of mental and somatic development, genital and/or
urinary abnormalities and ear abnormalities and/or deaf-
ness (Sanlaville et al. 2006). It is an autosomal dominant
condition with genotypic heterogeneity, although most
cases are due to the mutation or deletion of the chromod-
omain helicase DNA-binding domain protein-7 (CHD7), a
member of SNF2-like ATP-dependent chromatin-remod-
eling enzymes (Sanlaville et al. 2006). CHD7 mutations
have been also identified in Kallmann syndrome, a devel-
opmental disorder that shares with CHARGE some phe-
notypic features such as impaired olfaction and
hypogonadism (Kim et al. 2008). Mice with heterozygous
mutations in CHD7 are a good model for studying
CHARGE syndrome, and analyses of mouse mutant phe-
notypes have demonstrated a role in the development and
function of the neuronal system. CHD7 is necessary for
mammalian olfactory tissue development and function
(Layman et al. 2009), proliferation of inner ear neuroblasts
and inner ear morphogenesis in mice (Hurd et al. 2010),
promotion of the formation of multipotent migratory neural
crest that gives rise to craniofacial bones and cartilages,
and the peripheral nervous system, amongst others (Bajpai
et al. 2010). Recent in vitro studies have suggested that
CHD7 may directly regulate BMP4 expression, a protein
involved in cartilage and bone formation, by binding with
an enhancer element downstream of the BMP4 locus (Jiang
et al. 2012). More CHD7 targets have been identified, such
as the CHD7-dependent regulation (in association with
BRG1) of SOX9 and TEIST1 genes in human neural crest
cells (Bajpai et al. 2010). Mechanisms in which CHD7
regulates downstream genes vary in a tissue- and cell-
specific manner and depend on specific binding to meth-
ylated histone H3 lysine 4 in enhancer regions (Schnetz
et al. 2009). Although further research is needed, all these
findings suggest that mutations in CHD7 could transcrip-
tionally deregulate tissue-specific genes and developmental
genes resulting in the CHARGE phenotype.
Mutations in SWI/SNF complex family genes
in Coffin-Siris syndrome and mental retardation
Coffin-Siris syndrome (CSS, MIM #135900) or ‘‘fifth
digit’’ syndrome is a multiple congenital anomaly-mental
retardation syndrome characterized by severe develop-
mental delay, coarse facial features, hirsutism and absent
fifth fingernails, toenails and distal phalanges (Santen et al.
2012). In a recent study, 87 % of patients with CSS carried
a mutation in one or more members of the SWI/SNF family
of genes, which includes SMARCB1, SMARCA4, SMAR-
CA2, SMARCE1, ARID1A and ARID1B (Santen et al. 2012;
Tsurusaki et al. 2012). Interestingly, CSS patients carrying
different genetic mutations of the SWI/SNF chromatin-
remodeling factors gave rise to similar CSS phenotypes
(Tsurusaki et al. 2012), suggesting a general role for these
complexes in coordinating chromatin conformation and
gene expression. Deregulation of the SWI/SNF complexes
is also a common feature of tumorigenesis through its
function in mammalian differentiation, proliferation and
DNA repair (Reisman et al. 2009). However, the link
between SWI/SNF mutations and intellectual disorders is
still unclear. SMARCA2 and SMARCA4 are catalytic
subunits with ATPase activity, while ARID1A and
ARID1B are structural subunits involved in target recog-
nition and protein–protein interactions (Hargreaves and
Crabtree 2011). Both types of subunit are necessary to
regulate the transcription of several genes, such as c-FOS,
vimentin, CD44, cyclins, E-cadherin and important tran-
scription factors that have been functionally linked to SWI/
SNF (Reisman et al. 2009; Santen et al. 2012).
Mutations in SMARCA2 have also been described in
patients with the Nicolaides–Baraitser syndrome (NBS,
MIM # 601358), which is characterized by severe
372 Hum Genet (2013) 132:359–383
123

intellectual disability, early-onset seizures, short stature,
dysmorphic facial features and sparse hair (Van Houdt
et al. 2012; Wolff et al. 2012). Interestingly, Harikrishnan
et al. (2005) found that SMARCA2 associates with MECP2
and regulates FMR1 gene repression in mouse fibroblasts
and human T-lymphoblastic leukemia cells. Methylation at
promoter sites specified the recruitment of MECP2/
SMARCA2; while inhibition of methylation was associated
with complex release (Harikrishnan et al. 2005). These
results highlight an interesting link between epigenetic
marks and ATPase-dependent chromatin remodeling. How
SWI/SNF deregulation produces altered expression pat-
terns of genes associated with the CSS or NBS phenotype
is not clear, although some clues about the role of the
complex in neural differentiations could help interpret
some features (Seo et al. 2005; Lessard et al. 2007).
Mutations in non-coding RNA machinery
Non-coding RNAs, defined as functional RNA molecules
that are not translated into a protein, may also contribute to
the genesis of many human disorders (Table 1) (Esteller
2011). The best characterized ncRNAs in human condi-
tions are microRNAs (miRNAs) (Croce 2009), although
other ncRNAs members are emerging, such as small
nucleolar RNAs (snoRNAs), PIWI-interacting RNAs
(piRNAs), large intergenic non-coding RNAs (lincRNAs),
long non-coding RNAs (lncRNAs) and transcribed ultra-
conserved regions (T-UCRs), among others. If we focus on
miRNAs biogenesis, miRNAs are transcribed as individual
units (named primary miRNA (pri-miRNA)). After pro-
cessing by the Drosha complex, precursor miRNAs (pre-
miRNAs) are exported from the nucleus by the protein
exportin 5 (XPO5). Further processing by Dicer and TAR
RNA-binding protein 2 (TARBP2) generates mature
miRNAs, which are included into the RNA-induced
silencing complex (RISC). Once in this complex, miRNAs
could exert their function through degradation of protein-
coding transcripts or by translational repression. ncRNAs
profiles are frequently disrupted in different types of cancer
and in non-tumoral disorders, such as imprinting disorders,
rheumatoid arthritis, Rett syndrome and Alzheimer’s dis-
ease (Esteller 2011). Widespread alterations of ncRNAs
profiles could be also a consequence of genetic mutations
of the ncRNA-associated machinery. Some recent exam-
ples in this area are discussed in this section.
TARDBP mutations and amyotrophic lateral sclerosis
10 with or without frontotemporal lobar degeneration
Amyotrophic lateral sclerosis 10 (ALS10, MIM #612069)
with or without frontotemporal lobar degeneration (FTLD)
is an autosomal dominant neurodegenerative disorder
characterized by death of the motor neurons in the brain,
brainstem and spinal cord, resulting in fatal paralysis and
respiratory failure with a typical disease course of
1–5 years. Heterozygous mutations in the TAR DNA-
binding protein 43 (TARDBP) on chromosome 1p36,
encoding for the TDP43 protein, are present in a 50 % of
individuals affected by ALS10 (Ling et al. 2010). TARD-
BP is a member of the miRNA machinery. It has a double
effect on miRNA pathway. First, it enhances precursor
miRNA (pre-miRNA) production by both interacting with
the nuclear complex of Drosha or by direct binding to the
primary miRNAs (pri-miRNAs) in the nucleus; and sec-
ondly, it binds to the terminal loops of pre-miRNAs in the
cytoplasm by interaction with the Dicer complex (Kawa-
hara and Mieda-Sato 2012). Under non-pathological con-
ditions, TARDBP is mainly localized inside the nucleus,
but altered cellular distributions, including neuronal cyto-
plasmic, intranuclear inclusions and dystrophic neurites or
glial cytoplasmic locations are found in ALS10 and FTLD
(Arai et al. 2006; Neumann et al. 2006; Lagier-Tourenne
et al. 2010). Mutant TARDBP mice models developed a
similar phenotype than human TARDBP mutation
(Wegorzewska et al. 2009). Interestingly, no cytoplasmic
aggregates were found in mice mutants; suggesting that
other mechanisms rather than toxic cytoplasmic aggrega-
tion are underlying the molecular basis of ALS degenera-
tion (Wegorzewska et al. 2009). Results are in accordance
with a recent paper in which TDP-43 depletion in differ-
entiated Neruo2a results in decreased expression of miR-
132-3p and miR-132-5p. Further research of the specific
involvement of the aforementioned miRNAS (and others to
be explored) will strongly contribute to the understanding
of the pathogenesis of ALS10.
DGCR8 mutations and DiGeorge syndrome
DiGeorge syndrome (MIM #188400) is a complex disorder
characterized by learning disabilities, characteristic facial
appearance, submucous cleft palate, conotruncal heart
defects, thymic hypoplasia or aplasia, neonatal hypocal-
cemia, psychiatric illness and susceptibility to infection
due to a deficit of T cells (Goodship et al. 1998; Shiohama
et al. 2003). DiGeorge syndrome is caused by a 1.5 to 3.0-
Mb hemizygous deletion of chromosome 22q11.2 com-
prising the DiGeorge syndrome critical region gene 8
(DGCR8) (Shiohama et al. 2003) that encodes a double
stranded RNA-binding protein that is essential for miRNA
biogenesis. Specifically, DGCR8 is required in miRNA
maturation for processing pri-miRNAs to release pre-
miRNAs in the nucleus (Han et al. 2006). Genetic modified
mouse models carrying a hemizygous chromosomal defi-
ciency on chromosome 16 that spans a segment syntenic to
Hum Genet (2013) 132:359–383 373
123

the 1.5-Mb 22q11.2 microdeletion showed alterations in
the biogenesis of a set of miRNAs in the brain (Stark et al.
2008). Furthermore, DGCR8 deficiency resulted in altera-
tions of dendritic morphology, impaired sensorimotor gatin
and memory alterations similar to human DiGeorge phe-
notype (Stark et al. 2008). Additionally, it has been also
described that inactivation of a Dgcr8 conditional allele in
neural crest cells results in cardiovascular defects (Chapnik
et al. 2012). Similar results have been found in DGCR8
conditional knock-out mice embryos and knockout vascu-
lar smooth muscle cells (Chen et al. 2012a, b). DGCR8
deficiency was associated with down-regulation of the
miR-17/92 and miR-143/145 clusters in vascular smooth
muscle cells, reduced cell proliferation and increased
apoptosis (Chen et al. 2012a, b). These data provide spe-
cific explanations for cardiovascular and neuronal defects
that could explain, at least in part, the DiGeorge syndrome
phenotype.
DICER mutations in multinodular 1 Goiter disease
Autosomal dominant multinodular Goiter (MNG, MIM
#138800) is a disorder characterized by nodular over-
growth of the thyroid gland. In MNG type 1, some females
may also develop Sertoli–Leydig ovary tumors (Rio Frio
et al. 2011). Heterozygous mutations in DICER, a gene
encoding an RNase III endonuclease essential for micr-
oRNA processing, have recently been linked to MNG
pathogenesis (Rio Frio et al. 2011). Mutations in DICER
are also associated with pleuropulmonary blastoma (Hill
et al. 2009) and play a critical role in normal cardiac
function (Chen et al. 2008). DICER contains two RNase III
domains and a PAZ domain, a module that binds the end of
double-strand RNA (Macrae et al. 2006). Familial MNG
shows clear selective disruption of the PAZ domain, sug-
gesting a potential role of this domain in thyroid devel-
opment (Rio Frio et al. 2011). At the functional level,
lymphoblasts taken from MNG patients showed altered
miRNA compared with control profiles (i.e., LET7A and
miR345), suggesting a dysregulation of gene expression
patterns (Rio Frio et al. 2011). Further determination of the
consequences of specific microRNAs synthesis could be an
important topic for future research into MNG and other
DICER-associated pathologies.
Therapeutic applications of epigenetics
One of the main characteristics of epigenetic mechanisms
is their reversibility, making them potentially powerful
tools for curative pharmacological therapy (Fig. 1). Reac-
tivation of epigenetically silenced genes has been possible
for years by the treatment with DNA demethylation drugs,
such as zebularine or 5-aza-20-deoxycytidine (5-ADC), or
with histone deacetylase (HDAC) inhibitors, including
SAHA (suberoylanilide hydroxamic acid), valproic acid
(VPA) and trichostatin A (TSA). Indeed, some of these
drugs have significant antitumoral activity and the FDA has
approved the use of several of them for treating patients
(Kaminskas et al. 2005; Fiskus et al. 2008; Scuto et al.
2008). This approval has sparked a dramatic increase in the
development and trials of ‘‘epigenetic drugs’’ for treating
cancer and neural diseases (Kazantsev and Thompson
2008; Heyn and Esteller 2012). Although the most
advanced clinical trials are those corresponding to cancer
treatment, interest has grown in the fields of neurological
and neurodegenerative diseases in recent years (Day and
Sweatt 2012). The possibilities have only just begun to be
explored in human patients, but the basis of this therapy
has been confirmed in animal models.
Rubinstein–Taybi syndrome (RSTS) is probably the best
model for studying the therapeutic uses of HDAC inhibi-
tors that could compensate for the deficiency of HAT
activity (CBP mutations). RSTS has been modeled in mice,
and several heterozygous cbp ± mice (homozygous
cbp -/- mutants are embryonic lethal) have been gener-
ated (Alarcon et al. 2004). These models feature deficits in
long-term memory and cognitive impairments reminiscent
of human RTS neural symptoms, confirming the role of
CBP in the etiology of the disease. At the molecular level,
these mice have reduced HAT activity, decreased acetyla-
tion of specific histone proteins and impaired CBP-depen-
dent gene expression (Alarcon et al. 2004). Treatment with
HDAC inhibitors, such as SAHA or TSA, ameliorate defi-
cits in synaptic plasticity and cognition in cbp ± mice
(Hallam and Bourtchouladze 2006; Vecsey et al. 2007) by
enhancing transcriptional expression of specific neuronal
genes. In a similar manner, immortalized human lympho-
cytes derived from patients with RSTS showed reduced
acetylation levels, which primarily affect histones H2A and
H2B, compared to the histone acetylation levels of
immortalized human lymphocytes derived from patients
with Cornelia de Lange syndrome (a neurological disorder)
or healthy controls (Lopez-Atalaya et al. 2012). Interest-
ingly, the acetylation deficits in RSTS cells were rescued by
treatment with TSA (Lopez-Atalaya et al. 2012).
Some therapeutic approaches have been investigated in
Rett syndrome. Mecp2 knock-out mice also exhibit the
neurodevelopmental phenotype characteristic of human
Rett syndrome (Shahbazian et al. 2002). The cognitive
defect of MeCp2-deficient mice can be reverted by MecP2-
induced overexpression in mice (Collins et al. 2004) and in
MeCp2-deficient astrocytes (Lioy et al. 2011), suggesting
that this might well be an effective treatment for Rett
syndrome. Apart from this gene therapy strategy, which is
not really applicable in humans, pharmacological treat-
ments based on epigenetic targets are beginning to be
374 Hum Genet (2013) 132:359–383
123

explored as more feasible therapeutic interventions. Since
MeCP2 binds directly to methylated promoters in associ-
ation with the corepressor complex Sin3 and HDAC,
another therapeutic strategy could be based on targeting
HDAC activity. It has been widely demonstrated that
HDAC inhibitors enhance memory formation and neuronal
postnatal formation in various experimental models
(Kazantsev and Thompson 2008). There are some exam-
ples of the benefits of such treatments in the literature: the
alleviation of motor deficits in mouse models of Hunting-
ton disease by injections of SAHA (Hockly et al. 2003); the
delay of neurodegeneration and tauopathy in a mouse
model of Alzheimer’s disease through the activation of
SIRT1 by means of resveratrol treatments (Kilgore et al.
2010); and the reactivation of the frataxin genes by treat-
ments with benzamide-based HDAC inhibitors in immor-
talized cultured cells from Friedrich’s Ataxia syndrome
patients (Herman et al. 2006).
However, many questions remain unsolved concerning
drug potency, selectivity and permeability. Treatment with
HDAC inhibitors results in the re-expression of a limited
number of memory-associated genes (Vecsey et al. 2007).
This raises a question: are HDAC inhibitors’ treatments
potentially able to restore all types of genes? Or by contrast
do more ‘‘easier reverted’’ genes exist that can be phar-
macologically manipulated? If reversion is conditioned to
any factor (i.e., their genomic context or the acetylation
level) HDAC treatments would be more effective in certain
diseases than others. Multiple epigenetic enzymes could
contribute to a specific mark (especially histone acetyla-
tion) and furthermore, manipulating histone modifications
could also affect DNA methylation. Which is the best
target for inducing specific gene re-expression? Should
systemic or enzyme-targeted drugs be employed? This lack
of specificity means that almost none of the available
epigenetic drugs are isoform-specific and, for instance,
HDAC inhibitors have binding affinities for different
HDACs isoforms (Kilgore et al. 2010). It is expected that
development of subclass-specific inhibitors might contrib-
ute to minimize the side effects of the epigenetic therapy.
On the other hand, the tissue also influences its effective-
ness. DNA demethylating drugs are deoxycytidine analogs
that must be incorporated into the DNA after replication
cycles. As a consequence, the ‘‘speed’’ of base substitution
is strongly influenced by the rate of cell division. It is to be
expected that such DNA demethylating treatments will not
be very effective in neuronal cells with few or no cell
divisions. Further knowledge about the pathogenesis and
identification of the molecular/regulatory pathways that are
altered in the disease will enable the optimal target to be
identified and the therapeutic potential of epigenetic-based
drugs to be exploited. Despite these difficulties, epigenetic-
based therapy may become a successful intervention in the
treatment of human disorders associated with epigenetic
alterations.
Acknowledgments This work was supported by grants from the
European Research Council (Advanced EPINORC), the Fondo de
Investigaciones Sanitarias (PI08-1345 and PI10/02267), the Ministe-
rio de Ciencia e Innovacion (SAF2011-22803), the Fundacio La
Marato de TV3 (111430/31), European COST Action TD09/05, and
the Health Department of the Catalan Government (Generalitat de
Catalunya). M.E. is an Institucio Catalana de Recerca i Estudis
Avancats Research Professor.
References
Aapola U, Liiv I, Peterson P (2002) Imprinting regulator DNMT3L is
a transcriptional repressor associated with histone deacetylase
activity. Nucleic Acids Res 30(16):3602–3608
Abidi FE, Cardoso C, Lossi AM, Lowry RB, Depetris D, Mattei MG,
Lubs HA, Stevenson RE, Fontes M, Chudley AE, Schwartz CE
(2005) Mutation in the 5-prime alternatively spliced region of
the XNP/ATR-X gene causes Chudley-Lowry syndrome. Europ
J Hum Genet 13:176–183
Abidi FE, Miano MG, Murray JC, Schwartz CE (2007) A novel
mutation in the PHF8 gene is associated with X-linked mental
retardation with cleft lip/cleft palate. Clin Genet 72:19–22
Abidi FE, Holloway L, Moore CA, Weaver DD, Simensen RJ,
Stevenson RE, Rogers RC, Schwartz CE (2008) Mutations in
JARID1C are associated with X-linked mental retardation, short
stature and hyperreflexia. J Med Genet 45:787–793
Alarcon JM, Malleret G, Touzani K, Vronskaya S, Ishii S, Kandel ER,
Barco A (2004) Chromatin acetylation, memory, and LTP are
impaired in CBP ± mice: a model for the cognitive defect in
Rubinstein-Taybi syndrome and its amelioration. Neuron
42:947–959
Aldred MA, Sanford ROC, Thomas NS, Barrow MA, Wilson LC,
Brueton LA, Bonaglia MC, Hennekam RCM, Eng C, Dennis
NR, Trembath RC (2004) Molecular analysis of 20 patients with
2q37.3 monosomy: definition of minimum deletion intervals for
key phenotypes. J Med Genet 41:433–439
Amir RE, Van den Veyver IB, Wan M, Tran CQ, Francke U, Zoghbi
HY (1999) Rett syndrome is caused by mutations in X-linked
MECP2, encoding methyl-CpG-binding protein 2. Nat Genet
23(2):185–188
Anindya R, Mari PO, Kristensen U, Kool H, Giglia-Mari G,
Mullenders LH, Fousteri M, Vermeulen W, Egly JM, Svejstrup
JQ (2010) A ubiquitin-binding domain in Cockayne syndrome B
required for transcription-coupled nucleotide excision repair.
Mol Cell 38(5):637–648
Arai T, Hasegawa M, Akiyama H, Ikeda K, Nonaka T, Mori H, Mann
D, Tsuchiya K, Yoshida M, Hashizume Y, Oda T (2006) TDP-43
is a component of ubiquitin-positive tau-negative inclusions in
frontotemporal lobar degeneration and amyotrophic lateral
sclerosis. Biochem Biophys Res Commun 351(3):602–611
Arany Z, Sellers WR, Livingston DM, Eckner R (1994) E1A-
associated p300 and CREB-associated CBP belong to a
conserved family of coactivators. Cell 77(6):799–800
Argentaro A, Yang JC, Chapman L, Kowalczyk MS, Gibbons RJ,
Higgs DR, Neuhaus D, Rhodes D (2007) Structural conse-
quences of disease-causing mutations in the ATRX-DNMT3-
DNMT3L (ADD) domain of the chromatin-associated protein
ATRX. Proc Natl Acad Sci USA 104(29):11939–11944
Arnold MA, Kim Y, Czubryt MP, Phan D, McAnally J, Qi X, Shelton
JM, Richardson JA, Bassel-Duby R, Olson EN (2007) MEF2C
Hum Genet (2013) 132:359–383 375
123

transcription factor controls chondrocyte hypertrophy and bone
development. Dev Cell 12(3):377–389
Aziz A, Liu QC, Dilworth FJ (2010) Regulating a master regulator:
establishing tissue-specific gene expression in skeletal muscle.
Epigenetics 5(8):691–695
Bajpai R, Chen DA, Rada-Iglesias A, Zhang J, Xiong Y, Helms J,
Chang CP, Zhao Y, Swigut T, Wysocka J (2010) CHD7
cooperates with PBAF to control multipotent neural crest
formation. Nature 463(7283):958–962
Ballas N, Lioy DT, Grunseich C, Mandel G (2009) Non-cell
autonomous influence of MeCP2-deficient glia on neuronal
dendritic morphology. Nat Neurosci 12(3):311–317
Bannister AJ, Kouzarides T (2011) Regulation of chromatin by
histone modifications. Cell Res 21(3):381–395
Bartholdi D, Roelfsema JH, Papadia F, Breuning MH, Niedrist D,
Hennekam RC, Schinzel A, Peters DJ (2007) Genetic heteroge-
neity in Rubinstein-Taybi syndrome: delineation of the pheno-
type of the first patients carrying mutations in EP300. J Med
Genet 44(5):327–333
Bartsch O, Labonte J, Albrecht B, Wieczorek D, Lechno S, Zechner
U, Haaf T (2010) Two patients with EP300 mutations and facial
dysmorphism different from the classic Rubinstein-Taybi
syndrome. Am J Med Genet 152A:181–184
Basel-Vanagaite L (2010) Acute lymphoblastic leukemia in Weaver
syndrome. Am J Med Genet 152A:383–386
Batenburg NL, Mitchell TR, Leach DM, Rainbow AJ, Zhu XD (2012)
Cockayne Syndrome group B protein interacts with TRF2 and
regulates telomere length and stability. Nucleic Acids Res
40(19):9661–9674
Baujat G, Rio M, Rossignol S, Sanlaville D, Lyonnet S, Le Merrer M,
Munnich A, Gicquel C, Cormier-Daire V, Colleaux L (2004)
Paradoxical NSD1 mutations in Beckwith-Wiedemann syn-
drome and 11p15 anomalies in Sotos syndrome. Am J Hum
Genet 74:715–720
Berdasco M, Esteller M (2010) Aberrant epigenetic landscape in
cancer: how cellular identity goes awry. Dev Cell 19(5):698–711
Berdasco M, Esteller M (2011) DNA methylation in stem cell renewal
and multipotency. Stem Cell Res Ther 2(5):42
Berdasco M, Ropero S, Setien F, Fraga MF, Lapunzina P, Losson R,
Alaminos M, Cheung NK, Rahman N, Esteller M (2009)
Epigenetic inactivation of the Sotos overgrowth syndrome gene
histone methyltransferase NSD1 in human neuroblastoma and
glioma. Proc Natl Acad Sci USA 106(51):21830–21835
Berquist BR, Canugovi C, Sykora P, Wilson DM 3rd, Bohr VA
(2012) Human Cockayne syndrome B protein reciprocally
communicates with mitochondrial proteins and promotes tran-
scriptional elongation. Nucleic Acids Res 40(17):8392–8405
Bourtchouladze R, Lidge R, Catapano R, Stanley J, Gossweiler S,
Romashko D, Scott R, Tully T (2003) A mouse model of
Rubinstein-Taybi syndrome: defective long-term memory is
ameliorated by inhibitors of phosphodiesterase 4. Proc Natl Acad
Sci USA 100:10518–10522
Brun ME, Lana E, Rivals I, Lefranc G, Sarda P, Claustres M,
Megarbane A, De Sario A (2011) Heterochromatic genes
undergo epigenetic changes and escape silencing in immunode-
ficiency, centromeric instability, facial anomalies (ICF) syn-
drome. PLoS ONE 6(4):e19464
Campeau PM, Kim JC, Lu JT, Schwartzentruber JA, Abdul-Rahman
OA, Schlaubitz S, Murdock DM, Jiang MM, Lammer EJ, Enns
GM, Rhead WJ, Rowland J, Robertson SP, Cormier-Daire V,
Bainbridge MN, Yang XJ, Gingras MC, Gibbs RA, Rosenblatt
DS, Majewski J, Lee BH (2012a) Mutations in KAT6B,
encoding a histone acetyltransferase, cause Genitopatellar syn-
drome. Am J Hum Genet 90(2):282–289
Campeau PM, Lu JT, Dawson BC, Fokkema IF, Robertson SP,
Gibbs RA, Lee BH (2012b) The KAT6B-related disorders
genitopatellar syndrome and Ohdo/SBBYS syndrome have
distinct clinical features reflecting distinct molecular mecha-
nisms. Hum Mutat 33(11):1520–1525
Carney RM, Wolpert CM, Ravan SA, Shahbazian M, Ashley-Koch A,
Cuccaro ML, Vance JM, Pericak-Vance MA (2003) Identifica-
tion of MeCP2 mutations in a series of females with autistic
disorder. Pediat Neurol 28:205–211
Chahrour M, Jung SY, Shaw C, Zhou X, Wong ST, Qin J, Zoghbi HY
(2008) MeCP2, a key contributor to neurological disease,
activates and represses transcription. Science 320(5880):
1224–1229
Champagne N, Bertos NR, Pelletier N, Wang AH, Vezmar M, Yang
Y, Heng HH, Yang XJ (1999) Identification of a human histone
acetyltransferase related to monocytic leukemia zinc finger
protein. J Biol Chem 274:28528–28536
Chan HM, La Thangue NB (2001) p300/CBP proteins: HATs for
transcriptional bridges and scaffolds. J Cell Sci 114:2363–2373
Chang Q, Khare G, Dani V, Nelson S, Jaenisch R (2006) The disease
progression of Mecp2 mutant mice is affected by the level of
BDNF expression. Neuron 49(3):341–348
Chapnik E, Sasson V, Blelloch R, Hornstein E (2012) Dgcr8 controls
neural crest cells survival in cardiovascular development. Dev
Biol 362(1):50–56
Chen B, Cepko CL (2009) HDAC4 regulates neuronal survival in
normal and diseased retinas. Science 323(5911):256–259
Chen JF, Murchison EP, Tang R, Callis TE, Tatsuguchi M, Deng Z,
Rojas M, Hammond SM, Schneider MD, Selzman CH, Meissner
G, Patterson C, Hannon GJ, Wang DZ (2008) Targeted deletion
of Dicer in the heart leads to dilated cardiomyopathy and heart
failure. Proc Natl Acad Sci USA 105(6):2111–2116
Chen H, Gu X, Su IH, Bottino R, Contreras JL, Tarakhovsky A, Kim
SK (2009) Polycomb protein Ezh2 regulates pancreatic beta-cell
Ink4a/Arf expression and regeneration in diabetes mellitus.
Genes Dev 23(8):975–985
Chen YC, Gatchel JR, Lewis RW, Mao CA, Grant PA, Zoghbi HY,
Dent SY (2012a) Gcn5 loss-of-function accelerates cerebellar
and retinal degeneration in a SCA7 mouse model. Hum Mol
Genet 21(2):394–405
Chen Z, Wu J, Yang C, Fan P, Balazs L, Jiao Y, Lu M, Gu W, Li C,
Pfeffer LM, Tigyi G, Yue J (2012b) DiGeorge syndrome critical
region 8 (DGCR8) protein-mediated microRNA biogenesis is
essential for vascular smooth muscle cell development in mice.
J Biol Chem 287(23):19018–19028
Chou RH, Yu YL, Hung MC (2011) The roles of EZH2 in cell lineage
commitment. Am J Transl Res 3(3):243–250
Chrivia JC, Kwok RP, Lamb N, Hagiwara M, Montminy MR,
Goodman RH (1993) Phosphorylated CREB binds specifically to
the nuclear protein CBP. Nature 365:855–859
Claes S, Devriendt K, Van Goethem G, Roelen L, Meireleire J,
Raeymaekers P, Cassiman JJ, Fryns JP (2000) Novel syndromic
form of X-linked complicated spastic paraplegia. Am J Med
Genet 94:1–4
Clayton-Smith J, O’Sullivan J, Daly S, Bhaskar S, Day R, Anderson
B, Voss AK, Thomas T, Biesecker LG, Smith P, Fryer A,
Chandler KE, Kerr B, Tassabehji M, Lynch SA, Krajewska-
Walasek M, McKee S, Smith J, Sweeney E, Mansour S,
Mohammed S, Donnai D, Black G (2011) Whole-exome-
sequencing identifies mutations in histone acetyltransferase gene
KAT6B in individuals with the Say-Barber-Biesecker variant of
Ohdo syndrome. Am J Hum Genet 89:675–681
Collins AL, Levenson JM, Vilaythong AP, Richman R, Armstrong
DL, Noebels JL, David Sweatt J, Zoghbi HY (2004) Mild
overexpression of MeCP2 causes a progressive neurological
disorder in mice. Hum Mol Genet 13(21):2679–2689
Croce CM (2009) Causes and consequences of microRNA dysreg-
ulation in cancer. Nat Rev Genet 10(10):704–714
376 Hum Genet (2013) 132:359–383
123

Das C, Lucia MS, Hansen KC, Tyler JK (2009) CBP/p300-mediated
acetylation of histone H3 on lysine 56. Nature 459(7243):
113–117
Day JJ, Sweatt JD (2012) Epigenetic treatments for cognitive
impairments. Neuropsychopharmacology 37(1):247–260
De Felice C, Signorini C, Leoncini S, Pecorelli A, Durand T, Valacchi
G, Ciccoli L, Hayek J (2012) The role of oxidative stress in Rett
syndrome: an overview. Ann N Y Acad Sci 1259(1):121–135
De Greef JC, Wang J, Balog J, den Dunnen JT, Frants RR,
Straasheijm KR, Aytekin C, van der Burg M, Duprez L, Ferster
A, Gennery AR, Gimelli G, Reisli I, Schuetz C, Schulz A,
Smeets DF, Sznajer Y, Wijmenga C, van Eggermond MC, van
Ostaijen-Ten Dam MM, Lankester AC, van Tol MJ, van den
Elsen PJ, Weemaes CM, van der Maarel SM (2011) Mutations in
ZBTB24 are associated with immunodeficiency, centromeric
instability, and facial anomalies syndrome type 2. Am J Hum
Genet 88(6):796–804
Dillon SC, Zhang X, Trievel RC, Cheng X (2005) The SET-domain
protein superfamily: protein lysine methyltransferases. Genome
Biol 6(8):227
Douglas J, Hanks S, Temple IK, Davies S, Murray A, Upadhyaya M,
Tomkins S, Hughes HE, Cole TR, Rahman N (2003) NSD1
mutations are the major cause of Sotos syndrome and occur in
some cases of Weaver syndrome but are rare in other overgrowth
phenotypes. Am J Hum Genet 72(1):132–143
Ea CK, Hao S, Yeo KS, Baltimore D (2012) EHMT1 binds to NF-jB
p50 and represses gene expression. J Biol Chem 287(37):
31207–31217
Eckner R, Ewen ME, Newsome D, Gerdes M, DeCaprio JA,
Lawrence JB, Livingston DM (1994) Molecular cloning and
functional analysis of the adenovirus E1A-associated 300-kD
protein (p300) reveals a protein with properties of a transcrip-
tional adaptor. Genes Dev 8:869–884
Edbauer D, Neilson JR, Foster KA, Wang CF, Seeburg DP, Batterton
MN, Tada T, Dolan BM, Sharp PA, Sheng M (2010) Regulation
of synaptic structure and function by FMRP-associated microR-
NAs miR-125b and miR-132. Neuron 65(3):373–384
Ehrlich M (2003) The ICF syndrome, a DNA methyltransferase 3B
deficiency and immunodeficiency disease. Clin Immunol
109:17–28
Eissenberg JC, Wong M, Chrivia JC (2005) Human SRCAP and
Drosophila melanogaster DOM are homologs that function in the
notch signaling pathway. Mol Cell Biol 25(15):6559–6569
Erdel F, Rippe K (2011) Chromatin remodelling in mammalian cells
by ISWI-type complexes–where, when and why? FEBS J
278(19):3608–3618
Esteller M (2008) Epigenetics in cancer. N Engl J Med
358(11):1148–1159
Esteller M (2011) Non-coding RNAs in human disease. Nat Rev
Genet 12(12):861–874
Etchegaray JP, Yang X, DeBruyne JP, Peters AH, Weaver DR,
Jenuwein T, Reppert SM (2006) The polycomb group protein
EZH2 is required for mammalian circadian clock function. J Biol
Chem 281(30):21209–21215
Fatemi M, Hermann A, Pradhan S, Jeltsch A (2001) The activity of
the murine DNA methyltransferase Dnmt1 is controlled by
interaction of the catalytic domain with the N-terminal part of
the enzyme leading to an allosteric activation of the enzyme after
binding to methylated DNA. J Mol Biol 309(5):1189–1199
Feng J, Zhou Y, Campbell SL, Le T, Li E, Sweatt JD, Silva AJ, Fan G
(2010) Dnmt1 and Dnmt3a maintain DNA methylation and
regulate synaptic function in adult forebrain neurons. Nat
Neurosci 13(4):423–430
Fernandez AG, Gunsalus KC, Huang J, Chuang LS, Ying N, Liang
HL, Tang C, Schetter AJ, Zegar C, Rual JF, Hill DE, Reinke V,
Vidal M, Piano F (2005) New genes with roles in the C. elegans
embryo revealed using RNAi of ovary-enriched ORFeome
clones. Genome Res 15(2):250–259
Fiskus W, Wang Y, Joshi R, Rao R, Yang Y, Chen J, Kolhe R, Balusu
R, Eaton K, Lee P, Ustun C, Jillella A, Buser CA, Peiper S,
Bhalla K (2008) Cotreatment with vorinostat enhances activity
of MK-0457 (VX-680) against acute and chronic myelogenous
leukemia cells. Clin Cancer Res 14(19):6106–6115
Flaus A, Owen-Hughes T (2011) Mechanisms for ATP-dependent
chromatin remodelling: the means to the end. FEBS J
278(19):3579–3595
Franklin TB, Mansuy IM (2011) The involvement of epigenetic
defects in mental retardation. Neurobiol Learn Mem 96(1):61–67
Froyen G, Bauters M, Voet T, Marynen P (2006) X-linked mental
retardation and epigenetics. J Cell Mol Med 10(4):808–825
Garden GA, La Spada AR (2008) Molecular pathogenesis and cellular
pathology of spinocerebellar ataxia type 7 neurodegeneration.
Cerebellum 7(2):138–149
Giacometti E, Luikenhuis S, Beard C, Jaenisch R (2007) Partial
rescue of MeCP2 deficiency by postnatal activation of MeCP2.
Proc Natl Acad Sci USA 104(6):1931–1936
Gibbons RJ, Picketts DJ, Villard L, Higgs DR (1995) Mutations in a
putative global transcriptional regulator cause X-linked mental
retardation with alpha-thalassemia (ATR-X syndrome). Cell
80:837–845
Gibbons RJ, McDowell TL, Raman S, O’Rourke DM, Garrick D,
Ayyub H, Higgs DR (2000) Mutations in ATRX, encoding a
SWI/SNF-like protein, cause diverse changes in the pattern of
DNA methylation. Nat Genet 24:368–371
Gibbons RJ, Pellagatti A, Garrick D, Wood WG, Malik N, Ayyub H,
Langford C, Boultwood J, Wainscoat JS, Higgs DR (2003)
Identification of acquired somatic mutations in the gene encod-
ing chromatin-remodeling factor ATRX in the alpha-thalassemia
myelodysplasia syndrome (ATMDS). Nat Genet 34:446–449
Gibson WT, Hood RL, Zhan SH, Bulman DE, Fejes AP, Moore R,
Mungall AJ, Eydoux P, Babul-Hirji R, An J, Marra MA, FORGE
Canada Consortium, Chitayat D, Boycott KM, Weaver DD,
Jones SJM (2012) Mutations in EZH2 cause Weaver syndrome.
Am J Hum Genet 90:110–118
Goldberg AD, Banaszynski LA, Noh KM, Lewis PW, Elsaesser SJ,
Stadler S, Dewell S, Law M, Guo X, Li X, Wen D, Chapgier A,
DeKelver RC, Miller JC, Lee YL, Boydston EA, Holmes MC,
Gregory PD, Greally JM, Rafii S, Yang C, Scambler PJ, Garrick
D, Gibbons RJ, Higgs DR, Cristea IM, Urnov FD, Zheng D
(2010) Allis CD (2010) Distinct factors control histone variant
H3.3 localization at specific genomic regions. Cell
140(5):678–691
Goodship J, Cross I, LiLing J, Wren C (1998) A population study of
chromosome 22q11 deletions in infancy. Arch Dis Child
79:348–351
Guy J, Gan J, Selfridge J, Cobb S, Bird A (2007) Reversal of
neurological defects in a mouse model of Rett syndrome.
Science 315(5815):1143–1147
Hallam TM, Bourtchouladze R (2006) Rubinstein-Taybi syndrome:
molecular findings and therapeutic approaches to improve
cognitive dysfunction. Cell Mol Life Sci 63:1725–1735
Han J, Lee Y, Yeom KH, Nam JW, Heo I, Rhee JK, Sohn SY, Cho Y,
Zhang BT, Kim VN (2006) Molecular basis for the recognition
of primary microRNAs by the Drosha-DGCR8 complex. Cell
125(5):887–901
Hannibal MC, Buckingham KJ, Ng SB, Ming JE, Beck AE, McMillin
MJ, Gildersleeve HI, Bigham AW, Tabor HK, Mefford HC,
Cook J, Yoshiura K, Matsumoto T, Matsumoto N, Miyake N,
Tonoki H, Naritomi K, Kaname T, Nagai T, Ohashi H,
Kurosawa K, Hou JW, Ohta T, Liang D, Sudo A, Morris CA,
Banka S, Black GC, Clayton-Smith J, Nickerson DA, Zackai
EH, Shaikh TH, Donnai D, Niikawa N, Shendure J, Bamshad MJ
Hum Genet (2013) 132:359–383 377
123

(2011) Spectrum of MLL2 (ALR) mutations in 110 cases of
Kabuki syndrome. Am J Med Genet 155A:1511–1516
Hargreaves DC, Crabtree GR (2011) ATP-dependent chromatin
remodeling: genetics, genomics and mechanisms. Cell Res
21(3):396–420
Harikrishnan KN, Chow MZ, Baker EK, Pal S, Bassal S, Brasacchio
D, Wang L, Craig JM, Jones PL, Sif S, El-Osta A (2005) Brahma
links the SWI/SNF chromatin-remodeling complex with
MeCP2-dependent transcriptional silencing. Nat Genet 37:
254–264
Helmlinger D, Hardy S, Abou-Sleymane G, Eberlin A, Bowman AB,
Gansmuller A, Picaud S, Zoghbi HY, Trottier Y, Tora L, Devys
D (2006) Glutamine-expanded ataxin-7 alters TFTC/STAGA
recruitment and chromatin structure leading to photoreceptor
dysfunction. PLoS Biol 4(3):e67
Hennekam RCM (2006) Rubinstein-Taybi syndrome. Europ. J Hum
Genet 14:981–985
Henning KA, Li L, Iyer N, McDaniel LD, Reagan MS, Legerski R,
Schultz RA, Stefanini M, Lehmann AR, Mayne LV, Friedberg
EC (1995) The Cockayne syndrome group A gene encodes a WD
repeat protein that interacts with CSB protein and a subunit of
RNA polymerase II TFIIH. Cell 82:555–564
Herman D, Jenssen K, Burnett R, Soragni E, Perlman SL, Gottesfeld
JM (2006) Histone deacetylase inhibitors reverse gene silencing
in Friedreich’s ataxia. Nat Chem Biol 2(10):551–558
Heyn H, Esteller M (2012) DNA methylation profiling in the clinic:
applications and challenges. Nat Rev Genet 13(10):679–692
Heyn H, Vidal E, Sayols S, Sanchez-Mut JV, Moran S, Medina I,
Sandoval J, Simo-Riudalbas L, Szczesna K, Huertas D, Gatto S,
Matarazzo MR, Dopazo J, Esteller M (2012) Whole-genome
bisulfite DNA sequencing of a DNMT3B mutant patient.
Epigenetics 7(6):542–550
Hill DA, Ivanovich J, Priest JR, Gurnett CA, Dehner LP, Desruisseau
D, Jarzembowski JA, Wikenheiser-Brokamp KA, Suarez BK,
Whelan AJ, Williams G, Bracamontes D, Messinger Y, Good-
fellow PJ (2009) DICER1 mutations in familial pleuropulmonary
blastoma. Science 325:965
Ho L, Crabtree GR (2010) Chromatin remodelling during develop-
ment. Nature 463(7280):474–484
Ho L, Jothi R, Ronan JL, Cui K, Zhao K, Crabtree GR (2009) An
embryonic stem cell chromatin remodeling complex, esBAF, is
an essential component of the core pluripotency transcriptional
network. Proc Natl Acad Sci USA 106(13):5187–5191
Hockly E, Richon VM, Woodman B, Smith DL, Zhou X, Rosa E,
Sathasivam K, Ghazi-Noori S, Mahal A, Lowden PA, Steffan JS,
Marsh JL, Thompson LM, Lewis CM, Marks PA, Bates GP
(2003) Suberoylanilide hydroxamic acid, a histone deacetylase
inhibitor, ameliorates motor deficits in a mouse model of
Huntington’s disease. Proc Natl Acad Sci USA 100(4):
2041–2046
Hong S, Cho YW, Yu LR, Yu H, Veenstra TD, Ge K (2007)
Identification of JmjC domain-containing UTX and JMJD3 as
histone H3 lysine 27 demethylases. Proc Natl Acad Sci USA
104(47):18439–18444
Hood RL, Lines MA, Nikkel SM, Schwartzentruber J, Beaulieu C,
Nowaczyk MJ, Allanson J, Kim CA, Wieczorek D, Moilanen JS,
Lacombe D, Gillessen-Kaesbach G, Whiteford ML, Quaio CR,
Gomy I, Bertola DR, Albrecht B, Platzer K, McGillivray G, Zou
R, McLeod DR, Chudley AE, Chodirker BN, Marcadier J,
FORGE Canada Consortium, Majewski J, Bulman DE, White
SM, Boycott KM (2012) Mutations in SRCAP, encoding SNF2-
related CREBBP activator protein, cause Floating-Harbor syn-
drome. Am J Hum Genet 90(2):308–313
Horike S, Cai S, Miyano M, Cheng JF, Kohwi-Shigematsu T (2005)
Loss of silent-chromatin looping and impaired imprinting of
DLX5 in Rett syndrome. Nat Genet 37(1):31–40
Huang N, vom Baur E, Garnier JM, Lerouge T, Vonesch JL, Lutz Y,
Chambon P, Losson R (1998) Two distinct nuclear receptor
interaction domains in NSD1, a novel SET protein that exhibits
characteristics of both corepressors and coactivators. EMBO J
17(12):3398–3412
Hurd EA, Poucher HK, Cheng K, Raphael Y, Martin DM (2010) The
ATP-dependent chromatin remodeling enzyme CHD7 regulates
pro-neural gene expression and neurogenesis in the inner ear.
Development 137(18):3139–3150
Ishibashi T, Thambirajah AA, Ausio J (2008) MeCP2 preferentially
binds to methylated linker DNA in the absence of the terminal
tail of histone H3 and independently of histone acetylation.
FEBS Lett 582(7):1157–1162
Jensen LR, Amende M, Gurok U, Moser B, Gimmel V, Tzschach A,
Janecke AR, Tariverdian G, Chelly J, Fryns JP, Van Esch H,
Kleefstra T, Hamel B, Moraine C, Gecz J, Turner G, Reinhardt
R, Kalscheuer VM, Ropers HH, Lenzner S (2005) Mutations in
the JARID1C gene, which is involved in transcriptional regu-
lation and chromatin remodeling, cause X-linked mental retar-
dation. Am J Hum Genet 76:227–236
Jiang X, Zhou Y, Xian L, Chen W, Wu H, Gao X (2012) The
mutation in Chd7 causes misexpression of Bmp4 and develop-
mental defects in telencephalic midline. Am J Pathol 181(2):
626–641
Jin B, Tao Q, Peng J, Soo HM, Wu W, Ying J, Fields CR, Delmas
AL, Liu X, Qiu J, Robertson KD (2008) DNA methyltransferase
3B (DNMT3B) mutations in ICF syndrome lead to altered
epigenetic modifications and aberrant expression of genes
regulating development, neurogenesis and immune function.
Hum Mol Genet 17(5):690–709
Jin Q, Yu LR, Wang L, Zhang Z, Kasper LH, Lee JE, Wang C,
Brindle PK, Dent SY, Ge K (2011) Distinct roles of GCN5/
PCAF-mediated H3K9ac and CBP/p300-mediated H3K18/27ac
in nuclear receptor transactivation. EMBO J 30(2):249–262
Jones PL, Veenstra GJ, Wade PA, Vermaak D, Kass SU, Landsberger
N, Strouboulis J, Wolffe AP (1998) Methylated DNA and
MeCP2 recruit histone deacetylase to repress transcription. Nat
Genet 19(2):187–191
Josefowicz SZ, Wilson CB, Rudensky AY (2009) Cutting edge: TCR
stimulation is sufficient for induction of Foxp3 expression in the
absence of DNA methyltransferase 1. J Immunol 182(11):
6648–6652
Juan AH, Derfoul A, Feng X, Ryall JG, Dell’Orso S, Pasut A, Zare H,
Simone JM, Rudnicki MA, Sartorelli V (2011) Polycomb EZH2
controls self-renewal and safeguards the transcriptional identity
of skeletal muscle stem cells. Genes Dev 25(8):789–794
Kaminskas E, Farrell A, Abraham S, Baird A, Hsieh LS, Lee SL,
Leighton JK, Patel H, Rahman A, Sridhara R, Wang YC, Pazdur
R (2005) FDA, Approval summary: azacitidine for treatment of
myelodysplastic syndrome subtypes. Clin Cancer Res 11(10):
3604–3608
Kasper LH, Thomas MC, Zambetti GP, Brindle PK (2011) Double
null cells reveal that CBP and p300 are dispensable for p53
targets p21 and Mdm2 but variably required for target genes of
other signaling pathways. Cell Cycle 10(2):212–221
Kawahara Y, Mieda-Sato A (2012) TDP-43 promotes microRNAbiogenesis as a component of the Drosha and Dicer complexes.
Proc Natl Acad Sci USA 109(9):3347–3352
Kazantsev AG, Thompson LM (2008) Therapeutic application of
histone deacetylase inhibitors for central nervous system disor-
ders. Nat Rev Drug Discov 7(10):854–868
Kilgore M, Miller CA, Fass DM, Hennig KM, Haggarty SJ, Sweatt
JD, Rumbaugh G (2010) Inhibitors of class 1 histone
deacetylases reverse contextual memory deficits in a mouse
model of Alzheimer’s disease. Neuropsychopharmacology
35(4):870–880
378 Hum Genet (2013) 132:359–383
123

Kim HG, Kurth I, Lan F, Meliciani I, Wenzel W, Eom SH, Kang GB,
Rosenberger G, Tekin M, Ozata M, Bick DP, Sherins RJ, Walker
SL, Shi Y, Gusella JF, Layman LC (2008) Mutations in CHD7,
encoding a chromatin-remodeling protein, cause idiopathic
hypogonadotropic hypogonadism and Kallmann syndrome. Am
J Hum Genet 83:511–519
Kirmizis A, Bartley SM, Kuzmichev A, Margueron R, Reinberg D,
Green R, Farnham PJ (2004) Silencing of human polycomb
target genes is associated with methylation of histone H3 Lys 27.
Genes Dev 18(13):1592–1605
Kleefstra T, van Zelst-Stams WA, Nillesen WM, Cormier-Daire V,
Houge G, Foulds N, van Dooren M, Willemsen MH, Pfundt R,
Turner A, Wilson M, McGaughran J, Rauch A, Zenker M, Adam
MP, Innes M, Davies C, Lopez AG, Casalone R, Weber A,
Brueton LA, Navarro AD, Bralo MP, Venselaar H, Stegmann
SP, Yntema HG, van Bokhoven H, Brunner HG (2009) Further
clinical and molecular delineation of the 9q subtelomeric
deletion syndrome supports a major contribution of EHMT1
haploinsufficiency to the core phenotype. J Med Genet 46:
598–606
Kleefstra T, Brunner HG, Amiel J, Oudakker AR, Nillesen WM,
Magee A, Genevieve D, Cormier-Daire V, van Esch H, Fryns JP,
Hamel BC, Sistermans EA, de Vries BB, van Bokhoven H
(2006) Loss-of-function mutations in euchromatin histone
methyl transferase 1 (EHMT1) cause the 9q34 subtelomeric
deletion syndrome. Am J Hum Genet 79:370–377
Kleefstra T, Kramer JM, Neveling K, Willemsen MH, Koemans TS,
Vissers LE, Wissink-Lindhout W, Fenckova M, van den Akker
WM, Kasri NN, Nillesen WM, Prescott T, Clark RD, Devriendt
K, van Reeuwijk J, de Brouwer AP, Gilissen C, Zhou H, Brunner
HG, Veltman JA, Schenck A, van Bokhoven H (2012) Disrup-
tion of an EHMT1-Associated Chromatin-Modification Module
Causes Intellectual Disability. Am J Hum Genet 91(1):73–82
Klein CJ, Botuyan MV, Wu Y, Ward CJ, Nicholson GA, Hammans S,
Hojo K, Yamanishi H, Karpf AR, Wallace DC, Simon M,
Lander C, Boardman LA, Cunningham JM, Smith GE, Litchy
WJ, Boes B, Atkinson EJ, Middha S, Dyck B, Parisi JE, Mer G,
Smith DI, Dyck PJ (2011) Mutations in DNMT1 cause
hereditary sensory neuropathy with dementia and hearing loss.
Nat Genet 43:595–600
Kleine-Kohlbrecher D, Christensen J, Vandamme J, Abarrategui I,
Bak M, Tommerup N, Shi X, Gozani O, Rappsilber J, Salcini
AE, Helin K (2010) A Functional Link between the Histone
Demethylase PHF8 and the Transcription Factor ZNF711 in
X-Linked Mental Retardation. Mol Cell 38(2–2):165–178
Koivisto AM, Ala-Mello S, Lemmela S, Komu HA, Rautio J, Jarvela
I (2007) Screening of mutations in the PHF8 gene and
identification of a novel mutation in a Finnish family with
XLMR and cleft lip/cleft palate. Clin Genet 72:145–149
Korzus E, Rosenfeld MG, Mayford M (2004) CBP histone acetyl-
transferase activity is a critical component of memory consol-
idation. Neuron 42:961–972
Kourmouli N, Sun YM, van der Sar S, Singh PB, Brown JP (2005)
Epigenetic regulation of mammalian pericentric heterochromatin
in vivo by HP1. Biochem Biophys Res Commun 337(3):901–907
Kraft M, Cirstea IC, Voss AK, Thomas T, Goehring I, Sheikh BN,
Gordon L, Scott H, Smyth GK, Ahmadian MR, Trautmann U,
Zenker M, Tartaglia M, Ekici A, Reis A, Dorr HG, Rauch A,
Thiel CT (2011) Disruption of the histone acetyltransferase
MYST4 leads to a Noonan syndrome-like phenotype and
hyperactivated MAPK signaling in humans and mice. J Clin
Invest 121(9):3479–3491
Kramer JM, van Bokhoven H (2009) Genetic and epigenetic defects
in mental retardation. Int J Biochem Cell Biol 41(1):96–107
Kramer JM, Kochinke K, Oortveld MA, Marks H, Kramer D, de Jong
EK, Asztalos Z, Westwood JT, Stunnenberg HG, Sokolowski
MB, Keleman K, Zhou H, van Bokhoven H, Schenck A (2011)
Epigenetic regulation of learning and memory by Drosophila
EHMT/G9a. PLoS Biol 9(1):e1000569
Kurotaki N, Harada N, Yoshiura K, Sugano S, Niikawa N, Matsumoto
N (2001) Molecular characterization of NSD1, a human
homologue of the mouse Nsd1 gene. Gene 279(2):197–204
Kurotaki N, Imaizumi K, Harada N, Masuno M, Kondoh T, Nagai T,
Ohashi H, Naritomi K, Tsukahara M, Makita Y, Sugimoto T,
Sonoda T, Hasegawa T, Chinen Y, Tomita Ha HA, Kinoshita A,
Mizuguchi T, Yoshiura Ki K, Ohta T, Kishino T, Fukushima Y,
Niikawa N, Matsumoto N (2002) Haploinsufficiency of NSD1
causes Sotos syndrome. Nat Genet 30(4):365–366
Lacombe D, Saura R, Taine L, Battin J (1992) Confirmation of
assignment of a locus for Rubinstein-Taybi syndrome gene to
16p13.3. Am J Med Gent 44:126–128
Lagier-Tourenne C, Polymenidou M, Cleveland DW (2010) TDP-43
and FUS/TLS: emerging roles in RNA processing and neurode-
generation. Hum Mol Genet 19(R1):R46–R64
Lan F, Bayliss PE, Rinn JL, Whetstine JR, Wang JK, Chen S, Iwase
S, Alpatov R, Issaeva I, Canaani E, Roberts TM, Chang HY, Shi
Y (2007) A histone H3 lysine 27 demethylase regulates animal
posterior development. Nature 449(7163):689–694
Lana E, Megarbane A, Tourriere H, Sarda P, Lefranc G, Claustres M,
De Sario A (2012) DNA replication is altered in Immunodefi-
ciency Centromeric instability Facial anomalies (ICF) cells
carrying DNMT3B mutations. Eur J Hum Genet 20(10):
1044–1050
Lapunzina P (2005) Risk of tumorigenesis in overgrowth syndromes:
a comprehensive review. Am J Med Genet C Semin Med Genet
137C(1):53–71
Laugel V, Dalloz C, Durand M, Sauvanaud F, Kristensen U, Vincent
MC, Pasquier L, Odent S, Cormier-Daire V, Gener B, Tobias ES,
Tolmie JL, Martin-Coignard D, Drouin-Garraud V, Heron D,
Journel H, Raffo E, Vigneron J, Lyonnet S, Murday V, Gubser-
Mercati D, Funalot B, Brueton L, Sanchez Del Pozo J, Munoz E,
Gennery AR, Salih M, Noruzinia M, Prescott K, Ramos L, Stark
Z, Fieggen K, Chabrol B, Sarda P, Edery P, Bloch-Zupan A,
Fawcett H, Pham D, Egly JM, Lehmann AR, Sarasin A, Dollfus
H (2010) Mutation update for the CSB/ERCC6 and CSA/ERCC8
genes involved in Cockayne syndrome. Hum Mutat 31(2):
113–126
Laumonnier F, Holbert S, Ronce N, Faravelli F, Lenzner S, Schwartz
CE, Lespinasse J, Van Esch H, Lacombe D, Goizet C, Phan-
Dinh Tuy F, van Bokhoven H, Fryns JP, Chelly J, Ropers HH,
Moraine C, Hamel BC, Briault S (2005) Mutations in PHF8 are
associated with X linked mental retardation and cleft lip/cleft
palate. J Med Genet 42:780–786
Law MJ, Lower KM, Voon HP, Hughes JR, Garrick D, Viprakasit V,
Mitson M, De Gobbi M, Marra M, Morris A, Abbott A, Wilder
SP, Taylor S, Santos GM, Cross J, Ayyub H, Jones S, Ragoussis
J, Rhodes D, Dunham I, Higgs DR, Gibbons RJ (2010) ATR-X
syndrome protein targets tandem repeats and influences allele-
specific expression in a size-dependent manner. Cell 143:
367–378
Layman WS, McEwen DP, Beyer LA, Lalani SR, Fernbach SD, Oh E,
Swaroop A, Hegg CC, Raphael Y, Martens JR, Martin DM
(2009) Defects in neural stem cell proliferation and olfaction in
Chd7 deficient mice indicate a mechanism for hyposmia in
human CHARGE syndrome. Hum Mol Genet 18(11):1909–1923
Lederer D, Grisart B, Digilio MC, Benoit V, Crespin M, Ghariani SC,
Maystadt I, Dallapiccola B, Verellen-Dumoulin C (2012)
Deletion of KDM6A, a histone demethylase interacting with
MLL2, in three patients with Kabuki syndrome. Am J Hum
Genet 90:119–124
Lessard J, Wu JI, Ranish JA, Wan M, Winslow MM, Staahl BT, Wu
H, Aebersold R, Graef IA, Crabtree GR (2007) An essential
Hum Genet (2013) 132:359–383 379
123

switch in subunit composition of a chromatin remodeling
complex during neural development. Neuron 55(2):201–215
Lewis PW, Elsaesser SJ, Noh KM, Stadler SC, Allis CD (2010) Daxx
is an H3.3-specific histone chaperone and cooperates with
ATRX in replication-independent chromatin assembly at telo-
meres. Proc Natl Acad Sci USA 107(32):14075–14080
Li H, Zhong X, Chau KF, Williams EC, Chang Q (2011) Loss of
activity-induced phosphorylation of MeCP2 enhances synapto-
genesis. LTP and spatial memory. Nat Neurosci 14(8):
1001–1008
Licht CL, Stevnsner T, Bohr VA (2003) Cockayne syndrome group B
cellular and biochemical functions. Am J Hum Genet 73:
1217–1239
Ling SC, Albuquerque CP, Han JS, Lagier-Tourenne C, Tokunaga S,
Zhou H, Cleveland DW (2010) ALS-associated mutations in
TDP-43 increase its stability and promote TDP-43 complexes
with FUS/TLS. Proc Natl Acad Sci USA 107(30):13318–13323
Lioy DT, Garg SK, Monaghan CE, Raber J, Foust KD, Kaspar BK,
Hirrlinger PG, Kirchhoff F, Bissonnette JM, Ballas N, Mandel G
(2011) A role for glia in the progression of Rett’s syndrome.
Nature 475(7357):497–500
Liu L, van Groen T, Kadish I, Tollefsbol TO (2009) DNA
methylation impacts on learning and memory in aging. Neuro-
biol Aging 30(4):549–560
Liu W, Tanasa B, Tyurina OV, Zhou TY, Gassmann R, Liu WT, Ohgi
KA, Benner C, Garcia-Bassets I, Aggarwal AK, Desai A,
Dorrestein PC, Glass CK, Rosenfeld MG (2010) PHF8 mediates
histone H4 lysine 20 demethylation events involved in cell cycle
progression. Nature 466(7305):508–512
Lopez-Atalaya JP, Gervasini C, Mottadelli F, Spena S, Piccione M,
Scarano G, Selicorni A, Barco A, Larizza L (2012) Histone
acetylation deficits in lymphoblastoid cell lines from patients
with Rubinstein-Taybi syndrome. J Med Genet 49(1):66–74
Lu T, Jackson MW, Wang B, Yang M, Chance MR, Miyagi M,
Gudkov AV, Stark GR (2010) Regulation of NF-kappaB by
NSD1/FBXL11-dependent reversible lysine methylation of p65.
Proc Natl Acad Sci USA 107(1):46–51
Macrae IJ, Zhou K, Li F, Repic A, Brooks AN, Cande WZ, Adams
PD, Doudna JA (2006) Structural basis for double-stranded RNA
processing by Dicer. Science 311(5758):195–198
Mayo S, Garin I, Monfort S, Rosello M, Orellana C, Oltra S, Zazo C,
de Naclares GP, Martınez F (2012) Hypomethylation of the
KCNQ1OT1 imprinting center of chromosome 11 associated to
Sotos-like features. J Hum Genet 57(2):153–156
McMahon SJ, Pray-Grant MG, Schieltz D, Yates JR 3rd, Grant PA
(2005) Polyglutamine-expanded spinocerebellar ataxia-7 protein
disrupts normal SAGA and SLIK histone acetyltransferase
activity. Proc Natl Acad Sci USA 102(24):8478–8482
McManus KJ, Hendzel MJ (2003) Quantitative analysis of CBP- and
P300-induced histone acetylations in vivo using native chroma-
tin. Mol Cell Biol 23(21):7611–7627
Merson TD, Dixon MP, Collin C, Rietze RL, Bartlett PF, Thomas T,
Voss AK (2006) The transcriptional coactivator Querkopf
controls adult neurogenesis. J Neurosci 26(44):11359–11370
Miska EA, Langley E, Wolf D, Karlsson C, Pines J, Kouzarides T
(2001) Differential localization of HDAC4 orchestrates muscle
differentiation. Nucleic Acids Res 29(16):3439–3447
Monroy MA, Ruhl DD, Xu X, Granner DK, Yaciuk P, Chrivia JC
(2001) Regulation of cAMP-responsive element-binding protein-
mediated transcription by the SNF2/SWI-related protein.
SRCAP. J Biol Chem 276(44):40721–40726
Moretti P, Zoghbi HY (2006) MeCP2 dysfunction in Rett syndrome
and related disorders. Curr Opin Genet Dev 16(3):276–281
Morris B, Etoubleau C, Bourthoumieu S, Reynaud-Perrine S, Laroche
C, Lebbar A, Yardin C, Elsea SH (2012) Dose dependent
expression of HDAC4 causes variable expressivity in a novel
inherited case of brachydactyly mental retardation syndrome.
Am J Med Genet A 158A(8):2015–2020
Morrison AJ, Shen X (2009) Chromatin remodelling beyond
transcription: the INO80 and SWR1 complexes. Nat Rev Mol
Cell Biol 10(6):373–384
Muotri AR, Marchetto MC, Coufal NG, Oefner R, Yeo G, Nakashima
K, Gage FH (2010) L1 retrotransposition in neurons is modu-
lated by MeCP2. Nature 468(7322):443–446
Murawska M, Brehm A (2011) CHD chromatin remodelers and the
transcription cycle. Transcription 2(6):244–253
Na ES, Nelson ED, Kavalali ET, Monteggia LM (2012) The Impact
of MeCP2 Loss- or Gain-of-Function on Synaptic Plasticity.
Neuropsychopharmacology. doi:10.1038/npp.2012.116
Nagl NG Jr, Wang X, Patsialou A, Van Scoy M, Moran E (2007)
Distinct mammalian SWI/SNF chromatin remodeling complexes
with opposing roles in cell-cycle control. EMBO J 26(3):
752–763
Nakamura Y, Tagawa K, Oka T, Sasabe T, Ito H, Shiwaku H, La
Spada AR, Okazawa H (2012) Ataxin-7 associates with micro-
tubules and stabilizes the cytoskeletal network. Hum Mol Genet
21(5):1099–1110
Nan X, Hou J, Maclean A, Nasir J, Lafuente MJ, Shu X, Kriaucionis
S, Bird A (2007) Interaction between chromatin proteins MECP2
and ATRX is disrupted by mutations that cause inherited mental
retardation. Proc Nat Acad Sci USA 104:2709–2714
Neumann M, Sampathu DM, Kwong LK, Truax AC, Micsenyi MC,
Chou TT, Bruce J, Schuck T, Grossman M, Clark CM,
McCluskey LF, Miller BL, Masliah E, Mackenzie IR, Feldman
H, Feiden W, Kretzschmar HA, Trojanowski JQ, Lee VM (2006)
Ubiquitinated TDP-43 in frontotemporal lobar degeneration and
amyotrophic lateral sclerosis. Science 314(5796):130–133
Newman JC, Bailey AD, Weiner AM (2006) Cockayne syndrome
group B protein (CSB) plays a general role in chromatin
maintenance and remodeling. Proc Natl Acad Sci USA
103(25):9613–9618
Ng SB, Bigham AW, Buckingham KJ, Hannibal MC, McMillin MJ,
Gildersleeve HI, Beck AE, Tabor HK, Cooper GM, Mefford HC,
Lee C, Turner EH, Smith JD, Rieder MJ, Yoshiura K,
Matsumoto N, Ohta T, Niikawa N, Nickerson DA, Bamshad
MJ, Shendure J (2010) Exome sequencing identifies MLL2
mutations as a cause of Kabuki syndrome. Nat Genet 42:
790–793
Nguyen MV, Du F, Felice CA, Shan X, Nigam A, Mandel G,
Robinson JK, Ballas N (2012) MeCP2 Is Critical for Maintaining
Mature Neuronal Networks and Global Brain Anatomy during
Late Stages of Postnatal Brain Development and in the Mature
Adult Brain. J Neurosci 32(29):10021–10034
Nielsen AL, Jørgensen P, Lerouge T, Cervino M, Chambon P, Losson
R (2004) Nizp1, a novel multitype zinc finger protein that
interacts with the NSD1 histone lysine methyltransferase through
a unique C2HR motif. Mol Cell Biol 24(12):5184–5196
Niikawa N, Matsuura N, Fukushima Y, Ohsawa T, Kajii T (1981)
Kabuki make-up syndrome: a syndrome of mental retardation,
unusual facies, large and protruding ears, and postnatal growth
deficiency. J Pediat 99:565–569
Ogawa H, Ishiguro K, Gaubatz S, Livingston DM, Nakatani Y (2002)
A complex with chromatin modifiers that occupies E2F- and
Myc-responsive genes in G0 cells. Science 296(5570):
1132–1136
Ogryzko VV, Schiltz RL, Russanova V, Howard BH, Nakatani Y
(1996) The transcriptional coactivators p300 and CBP are
histone acetyltransferases. Cell 87:953–959
Ounap K, Puusepp-Benazzouz H, Peters M, Vaher U, Rein R, Proos
A, Field M, Reimand T (2012) A novel c.2T [ C mutation of the
KDM5C/JARID1C gene in one large family with X-linked
intellectual disability. Eur J Med Genet 55(3):178–184
380 Hum Genet (2013) 132:359–383
123

Park JH, Park EJ, Lee HS, Kim SJ, Hur SK, Imbalzano AN, Kwon J
(2006) Mammalian SWI/SNF complexes facilitate DNA double-
strand break repair by promoting gamma-H2AX induction.
EMBO J 25(17):3986–3997
Pasillas MP, Shah M, Kamps MP (2011) NSD1 PHD domains bind
methylated H3K4 and H3K9 using interactions disrupted by
point mutations in human Sotos syndrome. Hum Mutat
32(3):292–298
Petrij F, Giles RH, Dauwerse HG, Saris JJ, Hennekam RC, Masuno
M, Tommerup N, van Ommen GJ, Goodman RH, Peters DJ
(1995) Rubinstein-Taybi syndrome caused by mutations in the
transcriptional co-activator CBP. Nature 376(6538):348–351
Picketts DJ, Higgs DR, Bachoo S, Blake DJ, Quarrell OW, Gibbons
RJ (1996) ATRX encodes a novel member of the SNF2 family of
proteins: mutations point to a common mechanism underlying
the ATR-X syndrome. Hum Molec Genet 5:1899–1907
Portela A, Esteller M (2010) Epigenetic modifications and human
disease. Nat Biotechnol 28(10):1057–1068
Qi HH, Sarkissian M, Hu GQ, Wang Z, Bhattacharjee A, Gordon DB,
Gonzales M, Lan F, Ongusaha PP, Huarte M, Yaghi NK, Lim H,
Garcia BA, Brizuela L, Zhao K, Roberts TM, Shi Y (2010)
Histone H4K20/H3K9 demethylase PHF8 regulates zebrafish
brain and craniofacial development. Nature 466(7305):503–507
Qiao Y, Liu X, Harvard C, Hildebrand MJ, Rajcan-Separovic E,
Holden JJ, Lewis ME (2008) Autism-associated familial mic-
rodeletion of Xp11.22. Clin Genet 74(2):134–144
Rahman N (2005) Mechanisms predisposing to childhood overgrowth
and cancer. Curr Opin Genet Dev 15(3):227–233
Rajan I, Savelieva KV, Ye GL, Wang CY, Malbari MM, Friddle C,
Lanthorn TH, Zhang W (2009) Loss of the putative catalytic
domain of HDAC4 leads to reduced thermal nociception and
seizures while allowing normal bone development. PLoS ONE
4(8):e6612
Ravn K, Nielsen JB, Uldall P, Hansen FJ, Schwartz M (2003) No
correlation between phenotype and genotype in boys with a
truncating MECP2 mutation. J Med Genet 40(1):e5
Rayasam GV, Wendling O, Angrand PO, Mark M, Niederreither K,
Song L, Lerouge T, Hager GL, Chambon P, Losson R (2003)
NSD1 is essential for early post-implantation development and
has a catalytically active SET domain. EMBO J 22(12):
3153–3163
Reardon W (2002) Genitopatellar syndrome: a recognizable pheno-
type. Am J Med Genet 111:313–315
Reik W (2007) Stability and flexibility of epigenetic gene regulation
in mammalian development. Nature 447(7143):425–432
Reisman D, Glaros S, Thompson EA (2009) The SWI/SNF complex
and cancer. Oncogene 28(14):1653–1668
Rio Frio T, Bahubeshi A, Kanellopoulou C, Hamel N, Niedziela M,
Sabbaghian N, Pouchet C, Gilbert L, O’Brien PK, Serfas K,
Broderick P, Houlston RS, Lesueur F, Bonora E, Muljo S,
Schimke RN, Bouron-Dal Soglio D, Arseneau J, Schultz KA,
Priest JR, Nguyen VH, Harach HR, Livingston DM, Foulkes
WD, Tischkowitz M (2011) DICER1 mutations in familial
multinodular goiter with and without ovarian Sertoli-Leydig cell
tumors. JAMA 305:68–77
Roelfsema JH, White SJ, Ariyurek Y, Bartholdi D, Niedrist D,
Papadia F, Bacino CA, den Dunnen JT, van Ommen GJ,
Breuning MH, Hennekam RC, Peters DJ (2005) Genetic
heterogeneity in Rubinstein-Taybi syndrome: mutations in both
the CBP and EP300 genes cause disease. Am J Hum Genet
76(4):572–580
Ruhl DD, Jin J, Cai Y, Swanson S, Florens L, Washburn MP,
Conaway RC, Conaway JW, Chrivia JC (2006) Purification of a
human SRCAP complex that remodels chromatin by incorpo-
rating the histone variant H2A.Z into nucleosomes. Biochemistry
45:5671–5677
Samaco RC, Nagarajan RP, Braunschweig D (2004) LaSalle JM
(2004) Multiple pathways regulate MeCP2 expression in normal
brain development and exhibit defects in autism-spectrum
disorders. Hum Mol Genet 13(6):629–639
Samaco RC, Hogart A, LaSalle JM (2005) Epigenetic overlap in
autism-spectrum neurodevelopmental disorders: MECP2 defi-
ciency causes reduced expression of UBE3A and GABRB3.
Hum Mol Genet 14(4):483–492
Sanlaville D, Etchevers HC, Gonzales M, Martinovic J, Clement-Ziza
M, Delezoide AL, Aubry MC, Pelet A, Chemouny S, Cruaud C,
Audollent S, Esculpavit C, Goudefroye G, Ozilou C, Fredouille
C, Joye N, Morichon-Delvallez N, Dumez Y, Weissenbach J,
Munnich A, Amiel J, Encha-Razavi F, Lyonnet S, Vekemans M,
Attie-Bitach T (2006) Phenotypic spectrum of CHARGE
syndrome in fetuses with CHD7 truncating mutations correlates
with expression during human development. J Med Genet
43:211–217
Santen GW, Aten E, Sun Y, Almomani R, Gilissen C, Nielsen M,
Kant SG, Snoeck IN, Peeters EA, Hilhorst-Hofstee Y, Wessels
MW, den Hollander NS, Ruivenkamp CA, van Ommen GJ,
Breuning MH, den Dunnen JT, van Haeringen A, Kriek M
(2012) Mutations in SWI/SNF chromatin remodeling complex
gene ARID1B cause Coffin-Siris syndrome. Nat Genet
44:379–380
Santos-Reboucas CB, Fintelman-Rodrigues N, Jensen LR, Kuss AW,
Ribeiro MG, Campos M Jr, Santos JM, Pimentel MM (2011) A
novel nonsense mutation in KDM5C/JARID1C gene causing
intellectual disability, short stature and speech delay. Neurosci
Lett 498:67–71
Schnetz MP, Bartels CF, Shastri K, Balasubramanian D, Zentner GE,
Balaji R, Zhang X, Song L, Wang Z, Laframboise T, Crawford
GE, Scacheri PC (2009) Genomic distribution of CHD7 on
chromatin tracks H3K4 methylation patterns. Genome Res
19(4):590–601
Scuto A, Kirschbaum M, Kowolik C, Kretzner L, Juhasz A, Atadja P,
Pullarkat V, Bhatia R, Forman S, Yen Y, Jove R (2008) The
novel histone deacetylase inhibitor, LBH589, induces expression
of DNA damage response genes and apoptosis in Ph- acute
lymphoblastic leukemia cells. Blood 111(10):5093–5100
Seenundun S, Rampalli S, Liu QC, Aziz A, Palii C, Hong S, Blais A,
Brand M, Ge K, Dilworth FJ (2010) UTX mediates demethyl-
ation of H3K27me3 at muscle-specific genes during myogenesis.
EMBO J 29(8):1401–1411
Seo S, Richardson GA, Kroll KL (2005) The SWI/SNF chromatin
remodeling protein Brg1 is required for vertebrate neurogenesis
and mediates transactivation of Ngn and NeuroD. Development
132(1):105–115
Shahbazian M, Young J, Yuva-Paylor L, Spencer C, Antalffy B,
Noebels J, Armstrong D, Paylor R, Zoghbi H (2002) Mice with
truncated MeCP2 recapitulate many Rett syndrome features and
display hyperacetylation of histone H3. Neuron 35:243–254
Shiohama A, Sasaki T, Noda S, Minoshima S, Shimizu N (2003)
Molecular cloning and expression analysis of a novel gene
DGCR8 located in the DiGeorge syndrome chromosomal region.
Biochem Biophys Res Commun 304:184–190
Siderius LE, Hamel BC, van Bokhoven H, de Jager F, van den
Helm B, Kremer H, Heineman-de Boer JA, Ropers HH,
Mariman EC (1999) X-linked mental retardation associated
with cleft lip/palate maps to Xp11.3-q21.3. Am J Med Genet
85:216–220
Simpson MA, Deshpande C, Dafou D, Vissers LE, Woollard WJ,
Holder SE, Gillessen-Kaesbach G, Derks R, White SM, Cohen-
Snuijf R, Kant SG, Hoefsloot LH, Reardon W, Brunner HG,
Bongers EM, Trembath RC (2012) De novo mutations of the
gene encoding the histone acetyltransferase KAT6B cause
Genitopatellar syndrome. Am J Hum Genet 90(2):290–294
Hum Genet (2013) 132:359–383 381
123

Slupianek A, Yerrum S, Safadi FF, Monroy MA (2010) The
chromatin remodeling factor SRCAP modulates expression of
prostate specific antigen and cellular proliferation in prostate
cancer cells. J Cell Physiol 224(2):369–375
Song J, Teplova M, Ishibe-Murakami S, Patel DJ (2012) Structure-
based mechanistic insights into DNMT1-mediated maintenance
DNA methylation. Science 335(6069):709–712
Sopher BL, Ladd PD, Pineda VV, Libby RT, Sunkin SM, Hurley JB,
Thienes CP, Gaasterland T, Filippova GN, La Spada AR (2011)
CTCF regulates ataxin-7 expression through promotion of a
convergently transcribed, antisense noncoding RNA. Neuron
70:1071–1084
Stark KL, Xu B, Bagchi A, Lai WS, Liu H, Hsu R, Wan X, Pavlidis P,
Mills AA, Karayiorgou M, Gogos JA (2008) Altered brain
microRNA biogenesis contributes to phenotypic deficits in a
22q11-deletion mouse model. Nat Genet 40:751–760
Tahiliani M, Mei P, Fang R, Leonor T, Rutenberg M, Shimizu F, Li J,
Rao A, Shi Y (2007) The histone H3K4 demethylase SMCX
links REST target genes to X-linked mental retardation. Nature
447:601–605
Tao J, Hu K, Chang Q, Wu H, Sherman NE, Martinowich K, Klose
RJ, Schanen C, Jaenisch R, Wang W, Sun YE (2009)
Phosphorylation of MeCP2 at Serine 80 regulates its chromatin
association and neurological function. Proc Natl Acad Sci USA
106(12):4882–4887
Tatton-Brown K, Rahman N (2007) Sotos syndrome. Eur J Hum
Genet 15(3):264–271
Tatton-Brown K, Hanks S, Ruark E, Zachariou A, Duarte Sdel V,
Ramsay E, Snape K, Murray A, Perdeaux ER, Seal S, Loveday
C, Banka S, Clericuzio C, Flinter F, Magee A, McConnell V,
Patton M, Raith W, Rankin J, Splitt M, Strenger V, Taylor C,
Wheeler P, Temple KI, Cole T; Childhood Overgrowth Collab-
oration, Douglas J, Rahman N (2011) Germline mutations in the
oncogene EZH2 cause Weaver syndrome and increased human
height. Oncotarget 2(12):1127-1133
Thomas T, Voss AK, Chowdhury K, Gruss P (2000) Querkopf, a
MYST family histone acetyltransferase, is required for
normal cerebral cortex development. Development 127(12):
2537–2548
Tropea D, Giacometti E, Wilson NR, Beard C, McCurry C, Fu DD,
Flannery R, Jaenisch R, Sur M (2009) Partial reversal of Rett
Syndrome-like symptoms in MeCP2 mutant mice. Proc Natl
Acad Sci USA106(6):2029-2034
Tsai AC, Dossett CJ, Walton CS, Cramer AE, Eng PA, Nowakowska
BA, Pursley AN, Stankiewicz P, Wiszniewska J, Cheung SW
(2011) Exon deletions of the EP300 and CREBBP genes in two
children with Rubinstein-Taybi syndrome detected by aCGH.
Europ J Hum Genet 19:43–49
Tsurusaki Y, Okamoto N, Ohashi H, Kosho T, Imai Y, Hibi-Ko Y,
Kaname T, Naritomi K, Kawame H, Wakui K, Fukushima Y,
Homma T, Kato M, Hiraki Y, Yamagata T, Yano S, Mizuno S,
Sakazume S, Ishii T, Nagai T, Shiina M, Ogata K, Ohta T,
Niikawa N, Miyatake S, Okada I, Mizuguchi T, Doi H, Saitsu H,
Miyake N, Matsumoto N (2012) Mutations affecting components
of the SWI/SNF complex cause Coffin-Siris syndrome. Nat
Genet 44:376-378
Urdinguio RG, Lopez-Serra L, Lopez-Nieva P, Alaminos M, Diaz-
Uriarte R, Fernandez AF, Esteller M (2008) Mecp2-null mice
provide new neuronal targets for Rett syndrome. PLoS ONE
3(11):e3669
Urdinguio RG, Sanchez-Mut JV, Esteller M (2009) Epigenetic
mechanisms in neurological diseases: genes, syndromes, and
therapies. Lancet Neurol 8(11):1056–1072
Valinluck V, Tsai HH, Rogstad DK, Burdzy A, Bird A, Sowers LC
(2004) Oxidative damage to methyl-CpG sequences inhibits the
binding of the methyl-CpG binding domain (MBD) of methyl-
CpG binding protein 2 (MeCP2). Nucleic Acids Res
32(14):4100–4108
Van Esch H, Bauters M, Ignatius J, Jansen M, Raynaud M, Hollanders
K, Lugtenberg D, Bienvenu T, Jensen LR, Gecz J, Moraine C,
Marynen P, Fryns JP, Froyen G (2005) Duplication of the
MECP2 region is a frequent cause of severe mental retardation
and progressive neurological symptoms in males. Am J Hum
Genet 77(3):442–453
Van Houdt JK, Nowakowska BA, Sousa SB, van Schaik BD,
Seuntjens E, Avonce N, Sifrim A, Abdul-Rahman OA, van den
Boogaard MJ, Bottani A, Castori M, Cormier-Daire V, Deardorff
MA, Filges I, Fryer A, Fryns JP, Gana S, Garavelli L, Gillessen-
Kaesbach G, Hall BD, Horn D, Huylebroeck D, Klapecki J,
Krajewska-Walasek M, Kuechler A, Lines MA, Maas S,
Macdermot KD, McKee S, Magee A, de Man SA, Moreau Y,
Morice-Picard F, Obersztyn E, Pilch J, Rosser E, Shannon N,
Stolte-Dijkstra I, Van Dijck P, Vilain C, Vogels A, Wakeling E,
Wieczorek D, Wilson L, Zuffardi O, van Kampen AH, Devriendt
K, Hennekam R, Vermeesch JR (2012) Heterozygous missense
mutations in SMARCA2 cause Nicolaides-Baraitser syndrome.
Nat Genet 44:445–449
Vecsey CG, Hawk JD, Lattal KM, Stein JM, Fabian SA, Attner MA,
Cabrera SM, McDonough CB, Brindle PK, Abel T, Wood MA
(2007) Histone deacetylase inhibitors enhance memory and
synaptic plasticity via CREB: CBP-dependent transcriptional
activation. J Neurosci 27:6128–6140
Villard L (2007) MECP2 mutations in males. J Med Genet 44:
417–423
Viosca J, Lopez-Ayala JP, Olivares R, Eckner R, Barco A (2010)
Syndromic features and mild cognitive impairment in mice with
genetic reduction on p300 activity: differential contribution of
p300 and CBP to Rubinstein-Taybi syndrome etiology. Neuro-
biol Dis 37:186–194
Wang J, Weaver IC, Gauthier-Fisher A, Wang H, He L, Yeomans J,
Wondisford F, Kaplan DR, Miller FD (2010) CBP histone
acetyltransferase activity regulates embryonic neural differenti-
ation in the normal and Rubinstein-Taybi syndrome brain. Dev
Cell 18(1):114–125
Watson P, Black G, Ramsden S, Barrow M, Super M, Kerr B,
Clayton-Smith J (2001) Angelman syndrome phenotype associ-
ated with mutations in MECP2, a gene encoding a methyl CpG
binding protein. J Med Genet 38:224–228
Weaver DD, Graham CB, Thomas IT, Smith DW (1974) A new
overgrowth syndrome with accelerated skeletal maturation,
unusual facies, and camptodactyly. J Pediat 84:547–552
Wegorzewska I, Bell S, Cairns NJ, Miller TM, Baloh RH (2009)
TDP-43 mutant transgenic mice develop features of ALS and
frontotemporal lobar degeneration. Proc Natl Acad Sci USA
106(44):18809–18814
Willemsen MH, Vulto-van Silfhout AT, Nillesen WM, Wissink-
Lindhout WM, van Bokhoven H, Philip N, Berry-Kravis EM,
Kini U, van Ravenswaaij-Arts CM, Delle Chiaie B, Innes AM,
Houge G, Kosonen T, Cremer K, Fannemel M, Stray-Pedersen
A, Reardon W, Ignatius J, Lachlan K, Mircher C, Helderman van
den Enden PT, Mastebroek M, Cohn-Hokke PE, Yntema HG,
Drunat S, Kleefstra T (2012) Update on Kleefstra Syndrome.
Mol Syndromol 2(3–5):202–212
Williams SR, Aldred MA, Der Kaloustian VM, Halal F, Gowans G,
McLeod DR, Zondag S, Toriello HV, Magenis RE, Elsea SH
(2010) Haploinsufficiency of HDAC4 causes brachydactyly
mental retardation syndrome, with brachydactyly type E, devel-
opmental delays, and behavioral problems. Am J Hum Genet
87:219–228
Winkelmann J, Lin L, Schormair B, Kornum BR, Faraco J, Plazzi G,
Melberg A, Cornelio F, Urban AE, Pizza F, Poli F, Grubert F,
Wieland T, Graf E, Hallmayer J, Strom TM, Mignot E (2012)
382 Hum Genet (2013) 132:359–383
123

Mutations in DNMT1 cause autosomal dominant cerebellar
ataxia, deafness and narcolepsy. Hum Mol Genet 21(10):
2205–2210
Wolff D, Endele S, Azzarello-Burri S, Hoyer J, Zweier M, Schanze I,
Schmitt B, Rauch A, Reis A, Zweier C (2012) In-frame deletion
and missense mutations of the C-terminal helicase domain of
SMARCA2 in three patients with Nicolaides-Baraitser syn-
drome. Molec Syndromol 2:237–244
Wong MM, Cox L, Chrivia J (2007) The chromatin remodeling
protein SRCAP is critical for deposition of the histone variant
H2A.Z at promoters. J Biol Chem 282:26132–26139
Wong LH, McGhie JD, Sim M, Anderson MA, Ahn S, Hannan RD,
George AJ, Morgan KA, Mann JR, Choo KH (2010) ATRX
interacts with H3.3 in maintaining telomere structural integrity
in pluripotent embryonic stem cells. Genome Res 20(3):351–360
Wyngaarden LA, Delgado-Olguin P, Su IH, Bruneau BG, Hopyan S
(2011) Ezh2 regulates anteroposterior axis specification and
proximodistal axis elongation in the developing limb. Develop-
ment 138(17):3759–3767
Xie W, Ling T, Zhou Y, Feng W, Zhu Q, Stunnenberg HG, Grummt I,
Tao W (2012) The chromatin remodeling complex NuRD
establishes the poised state of rRNA genes characterized by
bivalent histone modifications and altered nucleosome positions.
Proc Natl Acad Sci USA 109(21):8161–8166
Xu GL, Bestor TH, Bourc’his D, Hsieh CL, Tommerup N, Bugge M,
Hulten M, Qu X, Russo JJ, Viegas-Pequignot E (1999)
Chromosome instability and immunodeficiency syndrome
caused by mutations in a DNA methyltransferase gene. Nature
402:187–191
Xu J, Deng X, Disteche CM (2008) Sex-specific expression of the
X-linked histone demethylase gene Jarid1c in brain. PLoS ONE
3(7):e2553
Yang X, Noushmehr H, Han H, Andreu-Vieyra C, Liang G, Jones PA
(2012) Gene reactivation by 5-aza-20-deoxycytidine-induced
demethylation requires SRCAP-mediated H2A.Z insertion to
establish nucleosome depleted regions. PLoS Genet 8(3):e1002604
Yasui DH, Peddada S, Bieda MC, Vallero RO, Hogart A, Nagarajan
RP, Thatcher KN, Farnham PJ, Lasalle JM (2007) Integrated
epigenomic analyses of neuronal MeCP2 reveal a role for long-
range interaction with active genes. Proc Natl Acad Sci USA
104(49):19416–19421
Yazdani M, Deogracias R, Guy J, Poot RA, Bird A, Barde YA (2010)
Disease modeling using embryonic stem cells: MeCP2 regulates
nuclear size and RNA synthesis in neurons. Stem Cells
30(10):2128–2139
Yehezkel S, Segev Y, Viegas-Pequignot E, Skorecki K, Selig S
(2008) Hypomethylation of subtelomeric regions in ICF syn-
drome is associated with abnormally short telomeres and
enhanced transcription from telomeric regions. Hum Mol Genet
17(18):2776–2789
Yoneda Y, Saitsu H, Touyama M, Makita Y, Miyamoto A, Hamada
K, Kurotaki N, Tomita H, Nishiyama K, Tsurusaki Y, Doi H,
Miyake N, Ogata K, Naritomi K, Matsumoto N (2012) Missense
mutations in the DNA-binding/dimerization domain of NFIX
cause Sotos-like features. J Hum Genet 57(3):207-211
Yoo SY, Pennesi ME, Weeber EJ, Xu B, Atkinson R, Chen S,
Armstrong DL, Wu SM, Sweatt JD, Zoghbi HY (2003) SCA7
knockin mice model human SCA7 and reveal gradual accumu-
lation of mutant ataxin-7 in neurons and abnormalities in short-
term plasticity. Neuron 37(3):383–401
Young JI, Hong EP, Castle JC, Crespo-Barreto J, Bowman AB, Rose
MF, Kang D, Richman R, Johnson JM, Berget S, Zoghbi HY
(2005) Regulation of RNA splicing by the methylation-depen-
dent transcriptional repressor methyl-CpG binding protein 2.
Proc Natl Acad Sci USA102(49):17551-17558
Yu L, Wang Y, Huang S, Wang J, Deng Z, Zhang Q, Wu W, Zhang
X, Liu Z, Gong W, Chen Z (2010) Structural insights into a
novel histone demethylase PHF8. Cell Res 20(2):166–173
Yuan X, Feng W, Imhof A, Grummt I, Zhou Y (2007) Activation of
RNA polymerase I transcription by cockayne syndrome group B
protein and histone methyltransferase G9a. Mol Cell
27(4):585–595
Zachariah RM, Rastegar M (2012) Linking epigenetics to human
disease and Rett syndrome: the emerging novel and challenging
concepts in MeCP2 research. Neural Plast 2012:415825
Zhou Z, Hong EJ, Cohen S, Zhao WN, Ho HY, Schmidt L, Chen WG,
Lin Y, Savner E, Griffith EC, Hu L, Steen JA, Weitz CJ,
Greenberg ME (2006) Brain-specific phosphorylation of MeCP2
regulates activity-dependent Bdnf transcription, dendritic
growth, and spine maturation. Neuron 52(2):255–269
Hum Genet (2013) 132:359–383 383
123






![Behavioural Implications of Genetic Syndromes [Read-Only] · 2015-11-24 · 1 Demystifying Syndromes: Behavioural Implications of Genetic Syndromes Dr. Karen Baker Psychologist Regional](https://img.dokumen.tips/doc/110x75/5f3aa6ca0bfcaa6c88001557/behavioural-implications-of-genetic-syndromes-read-only-2015-11-24-1-demystifying.jpg)






















