Embed Size (px)
Citation preview

9th International Working Conference on Stored Product Protection
446
PS5-9 – 6305
Genetic differentiation at the inter- and intra-specific level of storedgrain insects using a simple molecular approach (RAPD)
F. Fleurat-Lessard1,*, V. Pronier1
1 INRA–Research Unit Mycology and Food Safety (MycSA), Pôle QUALIS, BP n° 81, F 33883 Villenave d’Ornon,France
* Corresponding author, E-mail: [email protected]
Abstract
Population diversity in stored grain insectslinked with geographical origin of the infestingstrains may be an issue in international trade. Itmay be that some imported strains exhibitphenotypes that may negatively impact controlstrategies available in the country at the destinationof imported grain lots. However, severalmolecular methods are routinely used to assessgenetic diversity among insect species or strainsin the same species. Therefore, the potential ofRAPD technique for the identification of differentstrains based on polymorphic DNA revealing wasinvestigated with grain insect species. The bandingpatterns observed from DNA extracts with sixdifferent primers were compared between severalgrain insect species: Sitophilus spp.,Oryzaephilus surinamensis, Rhyzoperthadominica, Tribolium castaneum, Ephestiakuehniella, Plodia interpunctella, and threebruchid beetles. Then a particular study with thesame molecular tools was performed for thedifferentiation between geographical strains in theSitophilus spp. complex, with a special attentionpaid to S. zeamais strains originating fromdifferent continents (Africa, South America andEurope). The repeatability of the banding patternswas tested on: i/ extracted DNA from poolsamples of different stages and generations in thesame strains; ii/ Extracted DNA from individualsof the same species and strain. The differencesobserved in band profiles of the different species
were highly significant and enabled the easydifferentiation between them. It was shownfrequent dissimilarities between the band profilesof geographical strains of S. zeamais. Thepotential for RAPD-PCR technique to provideuseful genetic data for the discrimination up tothe population level of insects found in storedproducts in international trade is discussed.
Key words: RAPD-PCR, genetic differentiation,grain insect pest, geographical strain, moleculartechnique.
Introduction
The investigation about genetic differencesamong populations of stored grain insect speciesmay be used as discriminating criteria for thedetermination of the geographical origin, thedistribution, and the migrations of thesepopulations at the regional, national orinternational level (Fields and Phillips, 2002). Thegenetic differences among the populations of asingle species are the result of the genetic variationof the founder population in response to theselection pressure resulting of novel conditions forlife and reproduction encountered after migrationto new environments. The difficulty to observepolymorphic DNA among species and populationsof stored grain insect is mainly related to the lackof information about DNA of the major noxiousspecies. Only the most studied species, i.e.

Biology, Behavior, and Pest Detection on Stored Grain
447
Curculionidae, Bostrichidae, and Bruchidae, havesome DNA sequences available in DNAdatabases. This issue may be overcome using thepolymerase chain reaction-based randomlyamplified polymorphic DNA (RAPD-PCR) whichdoes not require previous knowledge of a DNAsequence to design primers (Williams et al., 1990;1993). This technique has generated useful resultsin the differentiation between insect species andstrains (Hoy, 2003). This method has a greatpotential for the identification of a large numberof DNA polymorphisms in the genome quicklyand efficiently (Haymer and McInnis, 1994). ThePCR-RAPD fingerprints may be used fordetermining at what point in the marketing channela commodity became infested (Dowdy andMcGaughey, 1996). They may be also useful todifferentiate between species with very closemorphological characters e.g. at the larval stage(Hidayat et al., 1996) as well as between closelyrelated strains of some species (Welsh et al.,1990). Despite all these advantages importantquestions have been raised for a practicalapplication. One of the most important is thereproducibility of the results obtained with thesame primers across a variety of protocols andlaboratory condit ions and the level ofpolymorphism that is searched (inter-specific,intra-specific, between geographical strains, etc.).Because the most frequent cause of poor qualityRAPDs is poor quality DNA, it is important topurify DNA with methods that will yield pure andundegraded DNA. With the ready-to-use kitsavailable for genomic cells and tissue DNAextraction followed by ready-to-go RAPDanalysis systems the quality and reproducibilityof PCR-RAPD analysis may be greatly improved.However, the reproducibility in the banding patterndiscriminating among insect species or strains,when studied with several individuals in a pool orwith a single individual has not been thoroughlyinvestigated with these kits.
In the current study, we used commercial kitsto determine the polymorphism among species,and geographical strains of the same species, ona pool of several individuals or on the genomicDNA extracted from a single individual. Our firstgoal was the choice of the more adequate primers
for the differentiation between species andpopulations of weevil from different geographicalorigin. The second goal was the study of thevariability factors that can affect the bandingpatterns obtained with different species or thedifferent geographical populations of a singlespecies. Our study aimed to standardize theRAPD-PCR technique in view of the identificationof the origin of the infestation of stored productsthrough the similarities with reference bandingpatterns of geographical strains.
Materials and methods
Insect materials
The different insect species used in this studywere sampled in the collection of the INRA MycSAlaboratory. In a first experiment, the RAPD-PCRwas performed to differentiate genetic materialfrom species with the highest occurrence in cerealgrain and by products: the three grain weevils,Sitophilus granarius (L.), S. oryzae (L.) and S.zeamais Motschulsky; the three bruchid beetles,Acanthoscelides obtectus (Say), Bruchus lentis(L.) and Callosobruchus maculatus (F.); the redflour beetle, Tribolium castaneum (Herbst), thesowtoothed grain beetle, Oryzaephilussurinamensis (L.); the lesser grain borer beetle,Rhyzopertha dominica (F.); the Mediterraneanflour moth, Ephestia kuehniella (Zeller); the Indianmeal moth, Plodia interpunctella (Huebner).These species are reared in laboratory conditionsfor more than hundred generations after theirsampling from infested products. For the grainweevils, geographical strains were also introducedin the comparison: two S. granarius strainsreferenced Sgr1 and Sgr08; three S. oryzaestrains referenced Sor1, Sor10, Sor11; and threeS. zeamais strains referenced Sze11, Sze12,Sze16.
In a second experiment, the differentiationpotential of the RAPD-PCR kits was applied todifferent geographical strains of the maize weevilS. zeamais. Three geographical populationsoriginating from Argentina, Ivory coast, andFrance have been used. These geographical

9th International Working Conference on Stored Product Protection
448
strains have been reared for more than 15 yearsin the laboratory on the same maize grain variety,i.e. more than hundred generations.
Storing insects for DNA preservation
The technique of storage in absolute ethanol(EtOH) was applied in all circumstances (Quickeet al., 1999). The insects were dehydrated inabsolute EtOH in cryotubes. Then, EtOH wasremoved and replaced with fresh to reduce thedilution with endogenous water (Hoy, 2003). Priorto the use for PCR, the tubes were sealed toprevent the evaporation of EtOH and thedehydrated insects were stored in a refrigeratorat 4 °C. This technique is safe for the preservationof insect DNA for more than two years at least(Quicke et al., 1999).
Insect DNA isolation (extraction)
In the first experiment on different species ofbeetles, the DNA extraction was carried out on apool of five individuals. For the Lepidopteraspecies, a single larva was used. For the secondstudy of the variability factors a part of theRAPD-PCR analyses were carried out on DNAextracted from a single individual. In this particularcase, five replicates were compared to assess thestability level of the RAPD profiles on DNAextracted from different individuals. For DNAextracted on a group of five individuals, the RAPDanalysis was carried out on five DNA samplesfrom the same extract. Prior to DNA extraction,insects were removed from EtOH and leave todry on sterile absorbent paper for. Then, theprotocol of DNeasy kit (Qiagen S.A.,Courtaboeuf, France) was used. In brief, theinsects were introduced in a 1.5 mL Eppendorfätube in which 180 µL PBS 1x solution wereadded. Insect sample was homogenized using anelectric homogenizer with a disposable microtube-pestle. Then, 4 µL R-Nase A (100 mg mL-1) wereadded and the tubes were incubated at roomtemperature during 2 min. To the R-Nase Atreated lysate, 20 µL of protein precipitationsolution (proteinase K 20 mg mL-1) and 200 µLbuffer AL were added followed by a thorough
vortexing. The tubes were then incubated in awater bath at 70 °C for 10 min.
Then, 200 µL EtOH (96 to 100 %) wereadded and tubes were thoroughly mixed byvortexing. The mixture was transferred into a minispin column placed in a 2 mL collection tube. Thetubes were centrifuged 1 min at 6,000 g. The minispin column was placed in a new 2 mL collectiontube and 500 µL washing buffer 1 were addedbefore a centrifugation for 1 min at 6,000 g. Then,the mini spin column was placed in a new 2 mLcollection tube. 500 µL washing buffer 2 wereadded and the tube was centrifuged for 3 min at20,000 g to dry the membrane.
For the DNA elution step, after the mini spincolumn was transferred with caution in a 1.5 mLnew tube, 200 µL eluting buffer were addeddirectly onto the membrane before an incubationat room temperature for 1 min followed by acentrifugation for 1 min at 6,000 g. This elutionstep was repeated once in the same conditions.The tubes with re-hydrated DNA were stored at4 °C in a refrigerator before PCR.
RAPD-PCR primers
The choice of discriminating primers was doneamong the six primers included in the “Ready-To-Go RAPD analysis kit” from AmershamBiosciences (GE Healthcare Europe GmbH,Orsay, France). Each kit contains 2.5 mmol ofeach of the following primers:
P1GGTGCGGGAAP2GTTTCGCTCCP3GTAGACCCGTP4AAGAGCCCGTP5AACGCGCAACP6CCCGTCAGCA
After the first experiment with the six primers,only P3 and P5 were used in the otherexperiments. This choice was based onfavourable characteristics of the banding patternsobserved: a limited number of major bands forRAPD analysis with these two primers and a highpotential of differentiation observed up to thestrain level with the species for which severalpopulations were compared.

Biology, Behavior, and Pest Detection on Stored Grain
449
PCR protocol
In each tube of RAPD analysis with a micro-bead the following products were included: 5 µLof a single RAPD primer (25 pmol); 2 µL of insectDNA extract (yield of the extraction assessed to5 to 50 ng per mg of insect body weight); bi-distilled water added to total of 25 µL. Twodifferent Escherichia coli DNAs were used asstandard DNA “extract” control for each RAPDanalysis run (in each electrophoresis gel). The tubecontent was gently mixed by vortexing or byrepeatedly pipeting up and down. A briefcentrifugation is needed to seat the mix in thebottom of the tube. The samples were placed inthe thermal cycler using the following PCRconditions: one cycle 95 °C, 5 min; 45 cycles95 °C, 1 min; one cycle 36 °C, 1 min; one cycle72 °C, 2 min.
Gel analysis
The electrophoresis revealing the bandingpattern of the RAPD-PCR amplified DNA wascarried out on a 2 % agarose gel using 1x TBEbuffer containing 0.5 µg mL-1 ethidium dibromide.In each well of the gel, 5 µL of PCR amplifiedproduct and 1 µL of 6x tracking buffer wereloaded. The electrophorese is performed untilgood separation of RAPD bands and the settingdepended of the capacity of the electrophoresissystem (e.g. 150 V for 3 h).
Variability factors investigated from bandingpattern interpretation
The similarities from one generation to anotherin the same strain were studied on the differentgeographical populations of S. zeamais.Additionally, the differences related to the sexwere revealed through the comparison of bandsobtained with the two parents issued from the F1progeny (considered as F2 generation) and thatwere used to obtain inbreed lines for F3generation. The variability in banding pattern withthese F3 inbreed lines was observed at theindividual level. The RAPD-PCR analysis was
carried out on DNA extracted from differentstages of S. zeamais of the same four“continental” strains (South-American, Africanand the two European populations from Portugaland France) used in all species comparison. Thus,the similarity of RAPD-PCR profiles from DNAextracted from individual larva, pupa and adultsampled in the same generation of S. zeamaisstrain ref.11 was checked. Finally, the stability ofthe banding patterns observed on successivegenerations of different S. zeamais strains wasstudied as well as the polymorphism related tothe sex from individual DNA extract.
Results
Differentiation between species
On the RAPD products observed in the 11different species which were studied none wasidentical to another (Figure 1). Each species hasits own profile that is specific and allows a cleardiscrimination with another species (Table 1). Forthe three species of the Bruchidae Family,Bruchus lentis, Acanthoscelides obtectus andCallosobruchus maculatus, only one RAPD-PCR amplified products with P3 primer iscommon to the three species (approximately680 bp). All other fragments amplified either withP3 or P5 primers are differently distributed(Table 1). For the geographical strains of the threeweevil species (S. oryzae, S. granarius, and S.zeamais), the specific combination of theamplified fragments for each strain enabled thedifferentiation between populations of S. zeamaisoriginating from different continents. For S.oryzae and S. granarius strains originating fromdifferent geographical locations in France, theprofiles were very similar either with P3 or P5primer (Figure 1 and Table 1). In this last situation,the differentiation between the origin of “domestic”populations originating from different areas of thesame country seems more difficult, even if somedissimilarities were detected.
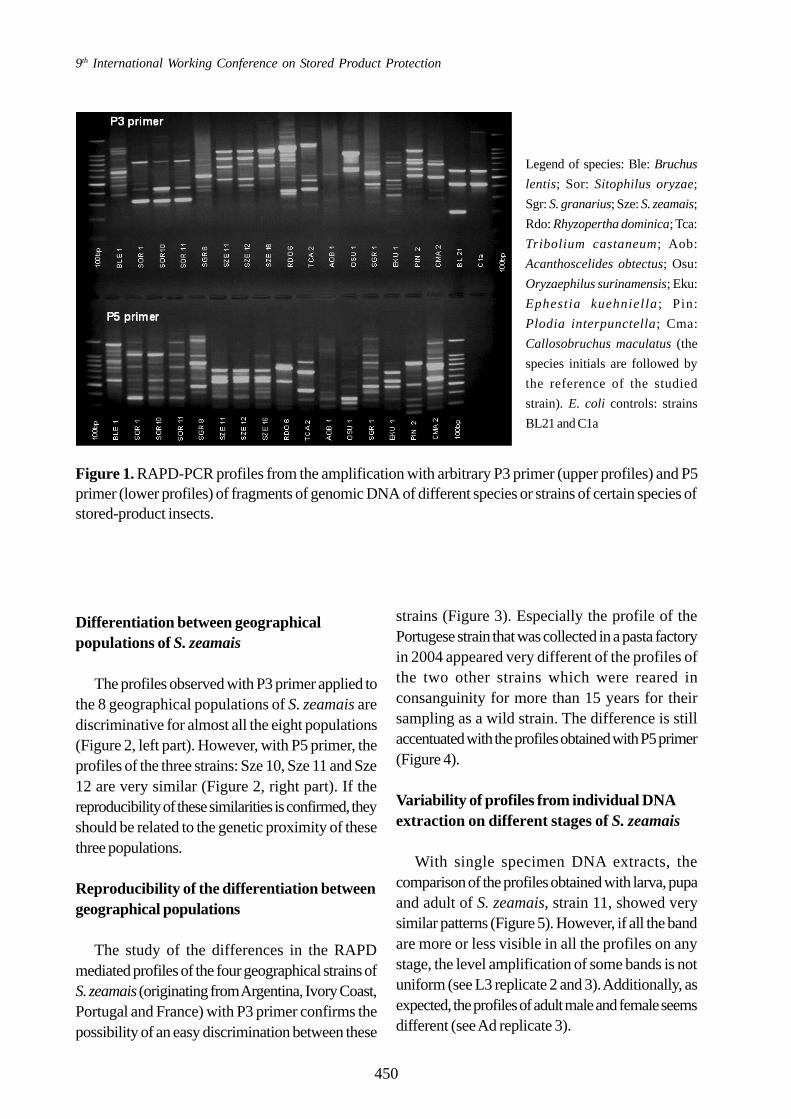
9th International Working Conference on Stored Product Protection
450
Figure 1. RAPD-PCR profiles from the amplification with arbitrary P3 primer (upper profiles) and P5primer (lower profiles) of fragments of genomic DNA of different species or strains of certain species ofstored-product insects.
Legend of species: Ble: Bruchus
lentis; Sor: Sitophilus oryzae;
Sgr: S. granarius; Sze: S. zeamais;
Rdo: Rhyzopertha dominica; Tca:
Tribolium castaneum; Aob:
Acanthoscelides obtectus; Osu:
Oryzaephilus surinamensis; Eku:
Ephestia kuehniel la; Pin:
Plodia interpunctella; Cma:
Callosobruchus maculatus (the
species initials are followed by
the reference of the studied
strain). E. coli controls: strains
BL21 and C1a
Differentiation between geographicalpopulations of S. zeamais
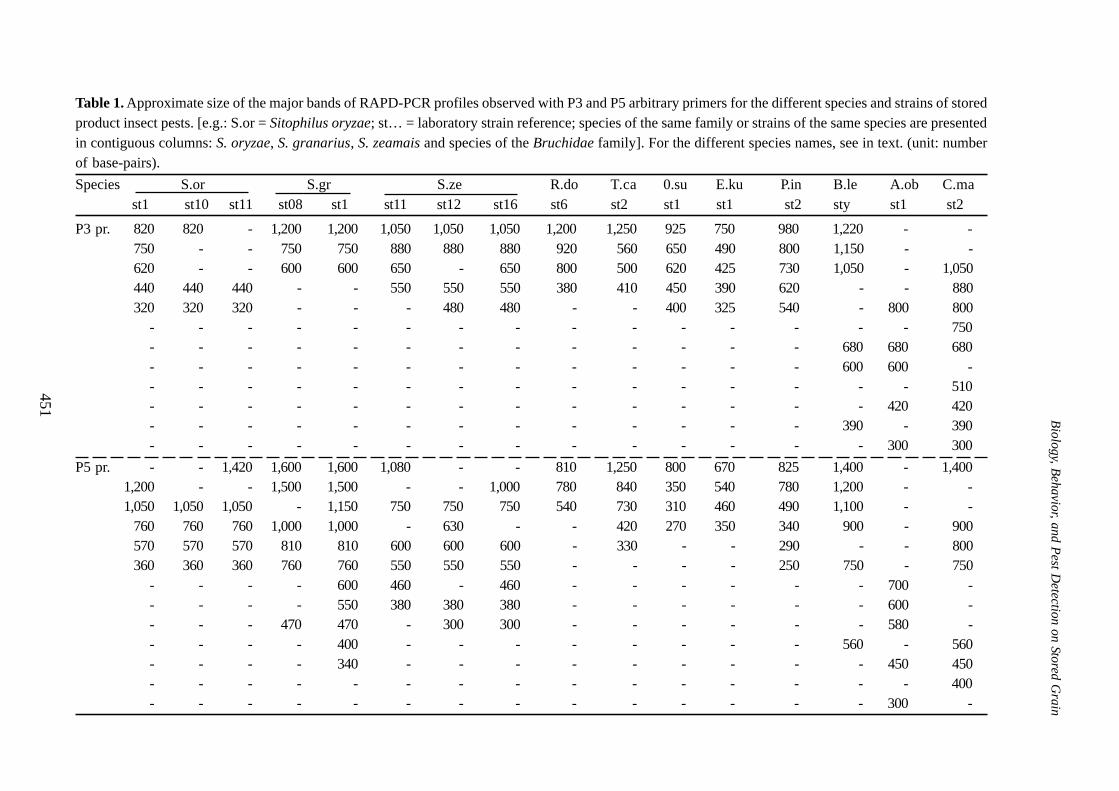
The profiles observed with P3 primer applied tothe 8 geographical populations of S. zeamais arediscriminative for almost all the eight populations(Figure 2, left part). However, with P5 primer, theprofiles of the three strains: Sze 10, Sze 11 and Sze12 are very similar (Figure 2, right part). If thereproducibility of these similarities is confirmed, theyshould be related to the genetic proximity of thesethree populations.
Reproducibility of the differentiation betweengeographical populations
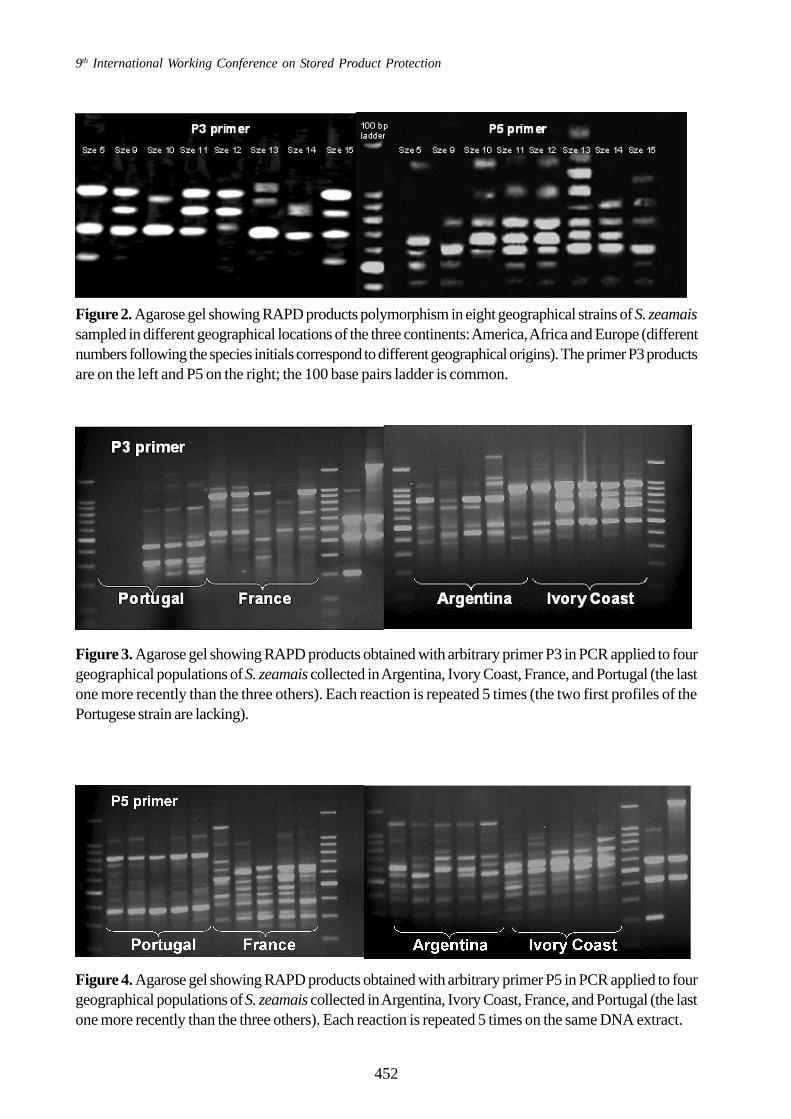
The study of the differences in the RAPDmediated profiles of the four geographical strains ofS. zeamais (originating from Argentina, Ivory Coast,Portugal and France) with P3 primer confirms thepossibility of an easy discrimination between these
strains (Figure 3). Especially the profile of thePortugese strain that was collected in a pasta factoryin 2004 appeared very different of the profiles ofthe two other strains which were reared inconsanguinity for more than 15 years for theirsampling as a wild strain. The difference is stillaccentuated with the profiles obtained with P5 primer(Figure 4).
Variability of profiles from individual DNAextraction on different stages of S. zeamais
With single specimen DNA extracts, thecomparison of the profiles obtained with larva, pupaand adult of S. zeamais, strain 11, showed verysimilar patterns (Figure 5). However, if all the bandare more or less visible in all the profiles on anystage, the level amplification of some bands is notuniform (see L3 replicate 2 and 3). Additionally, asexpected, the profiles of adult male and female seemsdifferent (see Ad replicate 3).
Bio
log
y, Be
ha
vior, a
nd
Pe
st De
tectio
n o
n S
tore
d G
rain
451
Table 1. Approximate size of the major bands of RAPD-PCR profiles observed with P3 and P5 arbitrary primers for the different species and strains of storedproduct insect pests. [e.g.: S.or = Sitophilus oryzae; st… = laboratory strain reference; species of the same family or strains of the same species are presentedin contiguous columns: S. oryzae, S. granarius, S. zeamais and species of the Bruchidae family]. For the different species names, see in text. (unit: numberof base-pairs).
Species S.or S.gr S.ze R.do T.ca 0.su E.ku P.in B.le A.obC.mast1 st10 st11 st08 st1 st11 st12 st16 st6 st2 st1 st1 st2 sty st1 st2
P3 pr. 820 820 - 1,200 1,200 1,050 1,050 1,050 1,200 1,250 925 750 980 1,220 - -750 - - 750 750 880 880 880 920 560 650 490 800 1,150 - -620 - - 600 600 650 - 650 800 500 620 425 730 1,050 - 1,050440 440 440 - - 550 550 550 380 410 450 390 620 - - 880320 320 320 - - - 480 480 - - 400 325 540 - 800 800
- - - - - - - - - - - - - - - 750- - - - - - - - - - - - - 680 680 680- - - - - - - - - - - - - 600 600 -- - - - - - - - - - - - - - - 510- - - - - - - - - - - - - - 420 420- - - - - - - - - - - - - 390 - 390- - - - - - - - - - - - - - 300 300
P5 pr. - - 1,420 1,600 1,600 1,080 - - 810 1,250 800 670 825 1,400 - 1,4001,200 - - 1,500 1,500 - - 1,000 780 840 350 540 780 1,200 - -1,050 1,050 1,050 - 1,150 750 750 750 540 730 310 460 490 1,100 - -
760 760 760 1,000 1,000 - 630 - - 420 270 350 340 900 - 900570 570 570 810 810 600 600 600 - 330 - - 290 - - 800360 360 360 760 760 550 550 550 - - - - 250 750 - 750
- - - - 600 460 - 460 - - - - - - 700 -- - - - 550 380 380 380 - - - - - - 600 -- - - 470 470 - 300 300 - - - - - - 580 -- - - - 400 - - - - - - - - 560 - 560- - - - 340 - - - - - - - - - 450 450- - - - - - - - - - - - - - - 400- - - - - - - - - - - - - - 300 -
9th International Working Conference on Stored Product Protection
452
Figure 2. Agarose gel showing RAPD products polymorphism in eight geographical strains of S. zeamaissampled in different geographical locations of the three continents: America, Africa and Europe (differentnumbers following the species initials correspond to different geographical origins). The primer P3 productsare on the left and P5 on the right; the 100 base pairs ladder is common.
Figure 3. Agarose gel showing RAPD products obtained with arbitrary primer P3 in PCR applied to fourgeographical populations of S. zeamais collected in Argentina, Ivory Coast, France, and Portugal (the lastone more recently than the three others). Each reaction is repeated 5 times (the two first profiles of thePortugese strain are lacking).
Figure 4. Agarose gel showing RAPD products obtained with arbitrary primer P5 in PCR applied to fourgeographical populations of S. zeamais collected in Argentina, Ivory Coast, France, and Portugal (the lastone more recently than the three others). Each reaction is repeated 5 times on the same DNA extract.

Biology, Behavior, and Pest Detection on Stored Grain
453
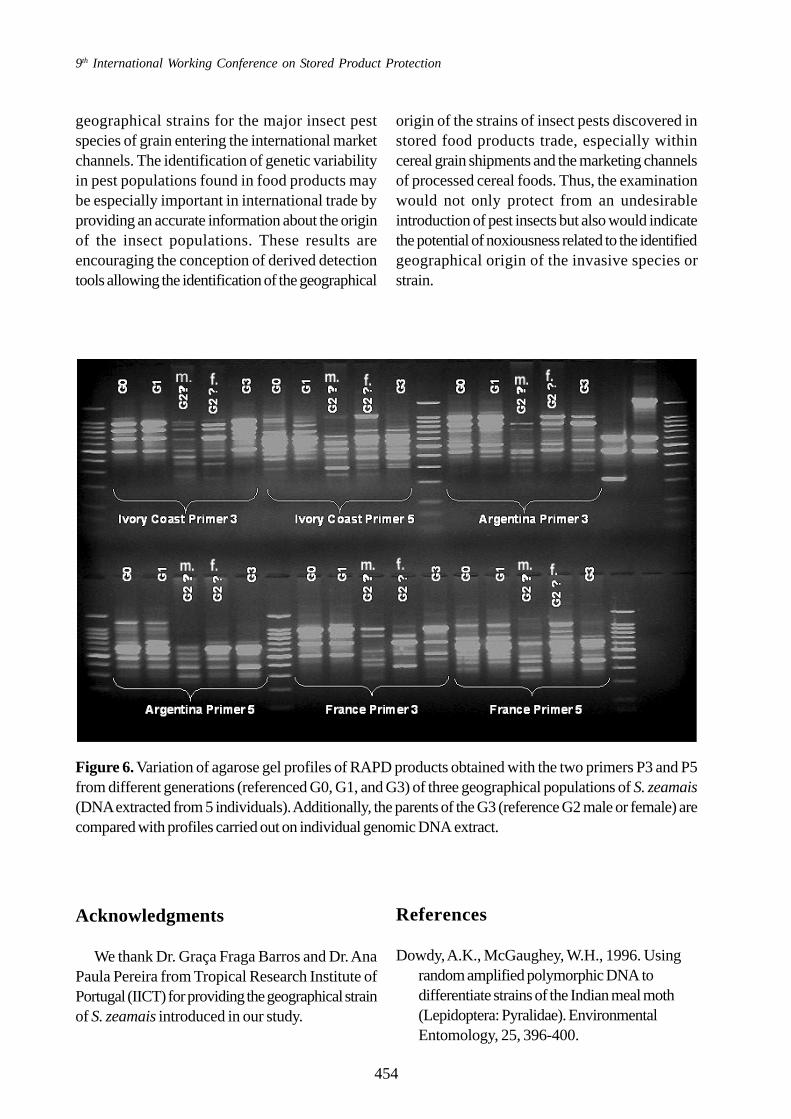
Variability from one generation to another
From the banding patterns in RAPD productsobserved with different generations of threedifferent geographical strains (“continentalstrains”), it was observed that the specific profilesallowing the differentiation between thesegeographical strains were stable along thegenerations, when DNA was extracted from 5individual (generations G0, G1 and G3) (Figure6). However, when DNA was extracted on asingle individual, as it is the case with the parents(male and female) of the G3 generation(referenced G2 male or female on Figure 6), thebanding pattern showed some different bandsbetween the two sexes.
Discussion
It was showed that RAPD-PCR may detectsmall differences in the genomes of storedproduct insects (Coleoptera or Lepidoptera),either at the species or the intra species level, suchas between geographical populations of a
cosmopolitan species. It was shown that this“fingerprinting” can be easily carried out onindividual insects and that the banding profileswere little dependant of the development stage.Moreover, this technique has already beenrecommended for the differentiation between thetwo species of the grain weevils (S. oryzae andS. zeamais) at the larval stage whenmorphological criteria are lacking (Hidayat et al.,1996). However, the male and female have notexactly the same pattern with this technique andthis difference become visible during the pupalstage. This technique seems especially useful forthe discrimination between biotypes among aspecies since RAPD-PCR pattern are correlatedwith the evolutionary adaptation of sub-populations when these are introduced in newenvironments or in stressing physical conditions(Espinasa and Borowsky, 1998).
Further investigation will be started to test thecorrespondence between these differences andthe geographical origin, at least at the continentallevel. The next step will be the constitution of acollection of a strain database of DNA fingerprintsallowing the comparison of genetic proximity in
Figure 5. Agarose gel showing the reproducibility of the RAPD products obtained on the same strain of S.zeamais on extracts from three different development stages: larva (L3), pupa (Pu), and adult (Ad). Eachreaction is repeated (Ri) on 5 different DNA extracts from a single individual.
9th International Working Conference on Stored Product Protection
454
geographical strains for the major insect pestspecies of grain entering the international marketchannels. The identification of genetic variabilityin pest populations found in food products maybe especially important in international trade byproviding an accurate information about the originof the insect populations. These results areencouraging the conception of derived detectiontools allowing the identification of the geographical
Figure 6. Variation of agarose gel profiles of RAPD products obtained with the two primers P3 and P5from different generations (referenced G0, G1, and G3) of three geographical populations of S. zeamais(DNA extracted from 5 individuals). Additionally, the parents of the G3 (reference G2 male or female) arecompared with profiles carried out on individual genomic DNA extract.
origin of the strains of insect pests discovered instored food products trade, especially withincereal grain shipments and the marketing channelsof processed cereal foods. Thus, the examinationwould not only protect from an undesirableintroduction of pest insects but also would indicatethe potential of noxiousness related to the identifiedgeographical origin of the invasive species orstrain.
Acknowledgments
We thank Dr. Graça Fraga Barros and Dr. AnaPaula Pereira from Tropical Research Institute ofPortugal (IICT) for providing the geographical strainof S. zeamais introduced in our study.
References
Dowdy, A.K., McGaughey, W.H., 1996. Usingrandom amplified polymorphic DNA todifferentiate strains of the Indian meal moth(Lepidoptera: Pyralidae). EnvironmentalEntomology, 25, 396-400.

Biology, Behavior, and Pest Detection on Stored Grain
455
Espinasa, L., Borowsky, R., 1998. Evolutionarydivergence of AP-PCR (RAPD) patterns.Molecular Biology and Evolution, 15, 408-414.
Fields, P.G., Phillips, T.W., 2002. The distributionand PCR-based fingerprints of Rhyzoperthadominica (F.) in Canada. In: Credland, P.F.,Armitage, D.M., Bell, C.H., Cogan, P.M.,Highley, E. (Eds.). Proceedings of the 8th
international Working Conference on StoredProduct Entomology, York, UK, July 22-26,2002, CAB International, Wallingford, UK,pp. 517-522.
Haymer, D.S., McInnis, D.O., 1994. Resolutionof populations of the Mediterranean fruit fly atthe DNA level using random primers for thepolymerase chain reaction. Genome, 37, 244-248.
Hidayat, P., Ffrench-Constant, R.H., Phillips,T.W., 1996. Molecular and morphologicalmarkers for diagnosis of Sitophilus oryzaeand S. zeamais (Coleoptera: Curculionidae).In: Highley, E., Wright, E.J., Banks, H.J.,Champ, B.R. (Eds.) Proceedings of the 6th
International Working Conference on StoredProduct Protection, Canberra (Australia),
April 6-11, 1994, CAB International,Wallingford, UK, I, pp. 528-532.
Hoy, M.A., 2003. Insect molecular genetics.Academic Press, San Diego CA, USA, 2nd
Edition, 544 p.
Quicke, D.L.J., Belshaw, R., Lopez-Vaamonde,C., 1999. Preservation of hymenopteranspecimens for subsequent molecular andmorphological study. Norvegian Academy ofSciences Letters in Zoologia Scripta, 28, 261-267.
Welsh, J., Petersen, C., McClelland, M., 1990.Polymorphisms generated by arbitrary primerPCR in the mouse; application to strainidentification and genetic mapping. NuvcleicAcids Research, 19, 303-306.
Williams, J.G.K., Kubelik, A.R., Livak, K.J.,Rafalski, J.A., Tingey, S.V., 1990. DNApolymorphisms amplified by arbitrary primersare useful as genetic markers. Nucleic AcidsResearch, 18, 6531-6535.
Williams, J.G.K., Hanafey, M.K., Rafalski, J.A.,Tingey, S.V., 1993. Genetic analysis usingrandom amplified polymorphic DNA markers;Methods in Enzymology, 218, 704-7





























