Embed Size (px)
Citation preview

Short Report
Genetic Clues About the Origin of Ache Hunter-Gatherers of ParaguaySIDIA M. CALLEGARI-JACQUES,1,2* KIM HILL,3 A. MAGDALENA HURTADO,3 LUCIANAT. RODRIGUES,1
CLAITON H.D. BAU,1 AND FRANCISCO M. SALZANO1
1Departamento de Genetica, Instituto de Biociencias, Universidade Federal do Rio Grande do Sul, 91501-970 Porto Alegre, RS, Brazil2Departamento de Estatıstica, Instituto de Matematica, Universidade Federal do Rio Grande do Sul, 91509-900, Porto Alegre, RS, Brazil3School of Human Evolution and Social Change, Arizona State University, Tempe, Arizona 85287
ABSTRACT Seventeen genetic studies performed among the until recently hunter-gatherer Ache were reviewedand the corresponding data integrated to general information about this group, and to statistical tests of hypothesisconcerning its origin(s). Two features of the Ache gene pool emerge: (a) the distinctiveness in relation to the general pat-tern of Amerindian genetic data; and (b) the reduced within population genetic variability. As for the origin of this peo-ple, the evidence suggests a general Tupian background with considerable introgression of Je genetic material, whichcould be explained by the Tupi warfare habit of absorption instead of extermination of defeated individuals. Am. J.Hum. Biol. 20:735–737, 2008. ' 2008 Wiley-Liss, Inc.
The Ache (or Guayaki) are one of the few remaininghunter-gatherers groups, living in socioecological condi-tions that prevailed for about 99% of our species’ history.These Tupi-speakers of Paraguay used to live in 60,000 km2
territory in the Parana River drainage of Paraguay,the process of more permanent contact with the outsidesociety occurring in 1959 and 1979 only.
At the time of contact they were described as havingwhite skin, light eye and hair color, beards, baldness, andstrong Asian characteristics. Cannibalism and other nonordinary behaviors were mentioned or imagined. Actually,their distinct morphological traits really occur, but theyintergrade with more common Amerindian features. Can-nibalism for ritual purposes stopped in the 1960s.
Small nomadic bands formed their demographic struc-ture, the two most culturally similar groups being theTupian hunter-gatherers Heta across the Parana riverand the Siriono/Yuki that live in Bolivian Chaco. By the9th century Guarani agriculturalists occupied the region,followed some centuries later by Europeans. After contact,severe epidemics occurred, with slow but steady recovery(Hill and Hurtado, 1996, 1999). Several demographic,physiological and other nongenetic investigations havebeen performed among them (Appendix 1, Supporting In-formation).
Ache’s origin was the subject of much fantasy. Two pos-sibilities have been envisaged: (a) they would be remnantsof Caaigua (‘‘forest people’’; usually identified as southernJes) who adopted the Guarani language and culture; or (b)they would be a fugitive Guarani group who took refuge inthe forest and completely lost agricultural skills (Hill andHurtado, 1996). The latter version is in accordance withAche’s mythology.
Genetic studies have presented contradictory results,some favoring a closer relationship with Tupi groups(Callegari-Jacques et al., 2007; Tsuneto et al., 2003; Wanget al., 2007), others showing a higher genetic similaritywith Je (Gaspar et al., 2002; Kohlrausch et al., 2005), andone posing the Ache as distinct from both Amerindiangroups (Battilana et al., 2002).
Our aims here are: (a) to summarize in a single sourcethe abundant published Ache genetic data and to comparethis group to the general Amerindian population; and (b)to verify which hypotheses about the Ache origin are sup-ported by genetic data.
SUBJECTS AND METHODS
Field and laboratory information
Currently the Ache population numbers about 1,200persons. Our sample collections were made in Arroyo Ban-dera (558500W, 238300S) and Chupa Pou (568300W,248100S), Paraguay. The data registered included age,family relationships, weight, stature, and color blindness.Physical examinations were performed and eventualhealth problems medicated. Blood samples were collectedfrom 99 individuals who were older than 10 years. Appro-priate informed consent was previously obtained. Thetribe has been frequently consulted on subsequent use ofgenetic samples, and receives a donation to their healthfund when genetic research is published.Eight categories grouping 48 genetic markers were used
in the present analyses of Ache origins (Table 1). The refer-ences and main original results are given in Appendices 2and 3, Supporting Information. Tribes pooled to composethe Tupi and Je populations, and sample sizes consideringthe different markers are presented in Appendices 4 and 5.
Statistical analysis
Tupi and Je gene frequencies are weighted averages ofall available data for these populations. As the group thatoriginated the Ache could have a marginal genetic consti-tution, we repeated all analyses using two tribes geo-graphically closer to the Ache (Guarani for the Tupi; Kain-gang for the Je).The relationship between Ache, Tupi, and Je was stud-
ied using the generalized hierarchical modeling (GHM)
Additional Supporting Information may be found in the online version ofthis article.
Contract grant sponsors: Conselho Nacional de Desenvolvimento Cientı-fico e Tecnologico; Institutos do Milenio e Apoio a Grupos de ExcelenciaPrograms, Fundacao de Amparo a Pesquisa do Estado do Rio Grandedo Sul.
*Correspondence to: Sidia M. Callegari-Jacques, Departamento deGenetica, Instituto de Biociencias, UFRGS, Caixa Postal 15053, 91501-970Porto Alegre, RS, Brazil. E-mail: [email protected]
Received 25 April 2008; Revision received 27 June 2008; Accepted 5 July2008
DOI 10.1002/ajhb.20816
Published online 12 September 2008 in Wiley InterScience (www.inter-science. wiley.com).
AMERICAN JOURNAL OF HUMAN BIOLOGY 20:735–737 (2008)
VVC 2008 Wiley-Liss, Inc.
approach described in Long and Kittles (2003), assuminga variable rate of evolution. Two tree-like models weretested: (1) Ache first links with Tupi, then Je connects tothis cluster; (2) Ache first clusters with Je, then Tupi joinsthe group. The degree of fit between the theoretical andobserved genetic variance-covariance matrices was meas-ured by a likelihood ratio statistic—lambda. Since lambdaideally follows a chi-square distribution, the sum of sev-eral such values will also be chi-square distributed, thusallowing for valid statistical tests. The observed variance-covariance matrix was generated using arcsine trans-formed gene frequencies and correction for sample size.The six sets of results obtained for biparental markers
were pooled by summing up the lambda statistics, aggre-gating the corresponding P-values using Fisher’s (1970)method, and analyzing all markers in a single run. Thearea beyond lambda in the distribution (P) was also usedto evaluate its displacement from zero (perfect fit).Admixture estimates were obtained using Chakra-
borty’s method (1975, 1986) after addition of Y-chromo-some and mtDNA data to the set of markers.
RESULTS
The results obtained in the physical examinations aretypical of rural South American populations, except forthe large number of adults with tuberculosis symptoms(Appendix 6, Supporting Information). Genetically, the
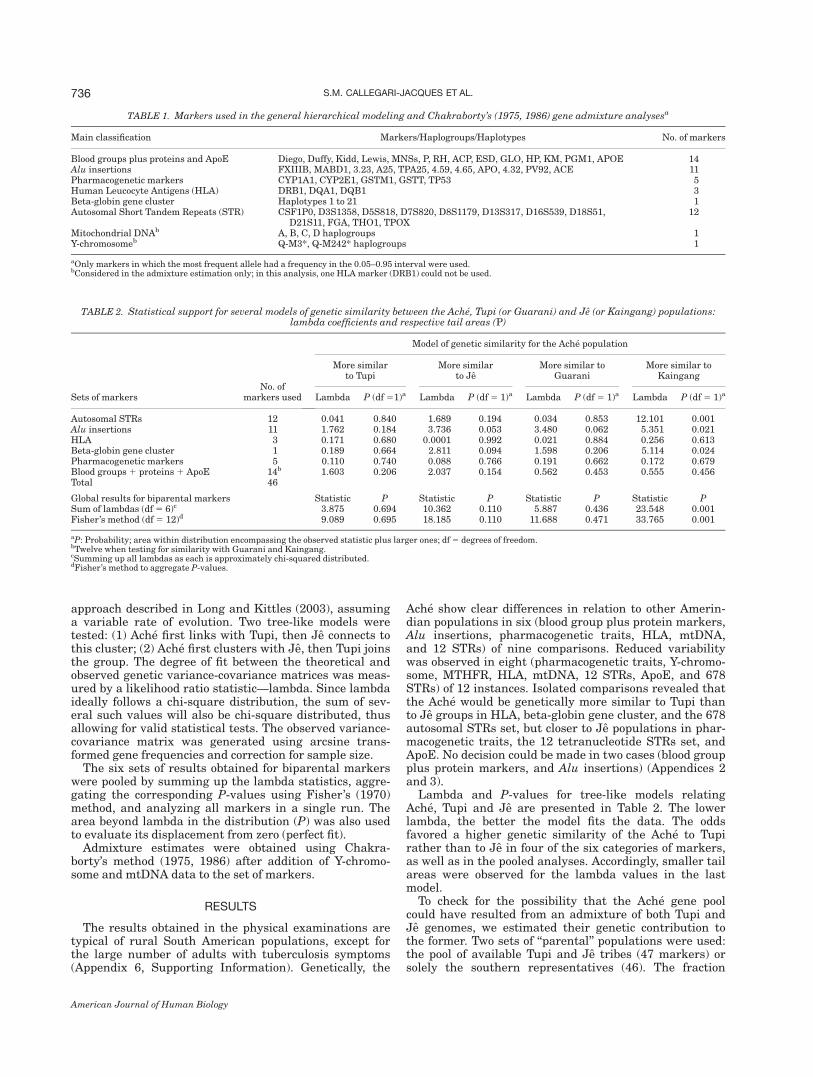
Ache show clear differences in relation to other Amerin-dian populations in six (blood group plus protein markers,Alu insertions, pharmacogenetic traits, HLA, mtDNA,and 12 STRs) of nine comparisons. Reduced variabilitywas observed in eight (pharmacogenetic traits, Y-chromo-some, MTHFR, HLA, mtDNA, 12 STRs, ApoE, and 678STRs) of 12 instances. Isolated comparisons revealed thatthe Ache would be genetically more similar to Tupi thanto Je groups in HLA, beta-globin gene cluster, and the 678autosomal STRs set, but closer to Je populations in phar-macogenetic traits, the 12 tetranucleotide STRs set, andApoE. No decision could be made in two cases (blood groupplus protein markers, and Alu insertions) (Appendices 2and 3).Lambda and P-values for tree-like models relating
Ache, Tupi and Je are presented in Table 2. The lowerlambda, the better the model fits the data. The oddsfavored a higher genetic similarity of the Ache to Tupirather than to Je in four of the six categories of markers,as well as in the pooled analyses. Accordingly, smaller tailareas were observed for the lambda values in the lastmodel.To check for the possibility that the Ache gene pool
could have resulted from an admixture of both Tupi andJe genomes, we estimated their genetic contribution tothe former. Two sets of ‘‘parental’’ populations were used:the pool of available Tupi and Je tribes (47 markers) orsolely the southern representatives (46). The fraction
TABLE 1. Markers used in the general hierarchical modeling and Chakraborty’s (1975, 1986) gene admixture analysesa
Main classification Markers/Haplogroups/Haplotypes No. of markers
Blood groups plus proteins and ApoE Diego, Duffy, Kidd, Lewis, MNSs, P, RH, ACP, ESD, GLO, HP, KM, PGM1, APOE 14Alu insertions FXIIIB, MABD1, 3.23, A25, TPA25, 4.59, 4.65, APO, 4.32, PV92, ACE 11Pharmacogenetic markers CYP1A1, CYP2E1, GSTM1, GSTT, TP53 5Human Leucocyte Antigens (HLA) DRB1, DQA1, DQB1 3Beta-globin gene cluster Haplotypes 1 to 21 1Autosomal Short Tandem Repeats (STR) CSF1P0, D3S1358, D5S818, D7S820, D8S1179, D13S317, D16S539, D18S51,
D21S11, FGA, THO1, TPOX12
Mitochondrial DNAb A, B, C, D haplogroups 1Y-chromosomeb Q-M3*, Q-M242* haplogroups 1
aOnly markers in which the most frequent allele had a frequency in the 0.05–0.95 interval were used.bConsidered in the admixture estimation only; in this analysis, one HLA marker (DRB1) could not be used.
TABLE 2. Statistical support for several models of genetic similarity between the Ache, Tupi (or Guarani) and Je (or Kaingang) populations:lambda coefficients and respective tail areas (P)
Sets of markersNo. of
markers used
Model of genetic similarity for the Ache population
More similarto Tupi
More similarto Je
More similar toGuarani
More similar toKaingang
Lambda P (df51)a Lambda P (df5 1)a Lambda P (df 5 1)a Lambda P (df 5 1)a
Autosomal STRs 12 0.041 0.840 1.689 0.194 0.034 0.853 12.101 0.001Alu insertions 11 1.762 0.184 3.736 0.053 3.480 0.062 5.351 0.021HLA 3 0.171 0.680 0.0001 0.992 0.021 0.884 0.256 0.613Beta-globin gene cluster 1 0.189 0.664 2.811 0.094 1.598 0.206 5.114 0.024Pharmacogenetic markers 5 0.110 0.740 0.088 0.766 0.191 0.662 0.172 0.679Blood groups 1 proteins 1 ApoE 14b 1.603 0.206 2.037 0.154 0.562 0.453 0.555 0.456Total 46
Global results for biparental markers Statistic P Statistic P Statistic P Statistic PSum of lambdas (df 5 6)c 3.875 0.694 10.362 0.110 5.887 0.436 23.548 0.001Fisher’s method (df5 12)d 9.089 0.695 18.185 0.110 11.688 0.471 33.765 0.001
aP: Probability; area within distribution encompassing the observed statistic plus larger ones; df5 degrees of freedom.bTwelve when testing for similarity with Guarani and Kaingang.cSumming up all lambdas as each is approximately chi-squared distributed.dFisher’s method to aggregate P-values.
736 S.M. CALLEGARI-JACQUES ET AL.
American Journal of Human Biology

(6SE) attributed to Tupi was 0.60 6 0.02 and to Guarani0.65 6 0.02, these two estimates not differing in a signifi-cant way.
DISCUSSION AND CONCLUSIONS
The evolutionary fate of a group basically depends ontwo parameters: birth and death. The probability of suchevents, on the other hand, is conditioned by the popula-tion’s external environment, its demographic structure,the resulting disease patterns, and cultural practices. Sal-zano (1985) enumerated several characteristics that dif-ferentiate hunter-gatherers from agriculturalists; ofthese, two features related to the Ache gene pool emerge:their distinctiveness in relation to the general pattern ofAmerindian genetic structure, and their reduced geneticvariability.
Special attention was given in the present communica-tion to the possible origin(s) of the Ache people. Our studysuggests a general Tupian background, with considerableintrogression of Je genetic material. The simple adoptionof a Tupian language by a Jean speaking group would notexplain the results of our analyses, neither would the hy-pothesis of the Ache being descendants of a Guaranigroup. Our results rather suggest admixture of Tupi andJe genomes, possibly due to incorporation of defeated Jeindividuals in warfare. Absorption into the group insteadof extermination of the conquered individuals was a gen-eral cultural trait of the Tupi-Guarani tribes. The oral tra-ditions of both Ache and Guarani account for possible con-quest/slavery relationships between these tribes (Hill andHurtado, 1996).
This hypothesis has some support in the admixturevalues obtained using selected genetic markers. The Jecontribution to the Ache gene pool was estimated as 0.276 0.02 when only biparental markers were used; theinclusion of mtDNA in the calculation raised this value to0.40 6 0.02, which signals for differential genetic contri-bution of Je women. This evidence did not change whenGuarani and Kaingang were used as parental popula-tions. Based on biparental markers, the Kaingang contri-bution was 0.29 6 0.01, changing to 0.35 6 0.02 with theaddition of mtDNA. In defeated bands or villages, womenwould be more likely to survive warfare and be slaved,
contributing eventually to the gene pool of the conquerors’population.Studies like those reported here may be important for
unraveling the factors that may influence the geneticstructure of hunter-gatherer populations.
LITERATURE CITED
Battilana J, Bonatto SL, Freitas LB, Hutz MH, Weimer TA, Callegari-Jac-ques SM, Batzer MA, Hill K, Hurtado AM, Tsuneto LT, Petzl-Erler ML,Salzano FM. 2002. Alu insertions versus blood group plus proteingenetic variability in four Amerindian populations. Ann Hum Biol29:334–347.
Callegari-Jacques SM, Crossetti SG, Kohlrausch FB, Salzano FM, TsunetoLT, Petzl-Erler ML, Hill K, Hurtado AM, Hutz MH. 2007. The b-globingene cluster distribution revisited—patterns in Amerindian popula-tions. Am J Phys Anthropol 134:190–197.
Chakraborty R. 1975. Estimation of race admixture—a new method. Am JPhys Anthropol 42:507–511.
Chakraborty R. 1986. Gene admixture in human populations: models andpredictions. Yearbk Phys Anthropol 29:1–43.
Fisher RA. 1970. Statistical methods for research workers, 14 ed. NewYork: Hafner.
Gaspar PA, Hutz MH, Salzano FM, Hill K, Hurtado MA, Petzl-Erler ML,Tsuneto LT, Weimer TA. 2002. Polymorphisms of CYP1A1, CYP2E1,GSTM1, GSTT1, and TP53 genes in Amerindians. Am J Phys Anthropol119:249–256
Hill K, Hurtado AM. 1996. Ache life history. The ecology and demographyof a foraging people. New York: Aldine de Gruyter.
Hill K, Hurtado AM. 1999. The Ache of Paraguay. In: Lee RB, Daly R, edi-tors. The Cambridge encyclopedia of hunters and gatherers. Cambridge:Cambridge University Press. p 92–96.
Kohlrausch FB, Callegari-Jacques SM, Tsuneto LT, Petzl-Erler ML, Hill K,Hurtado AM, Salzano FM, Hutz MH. 2005. Geography influences micro-satellite polymorphism diversity in Amerindians. Am J Phys Anthropol126:463–470.
Long JC, Kittles RA. 2003. Human genetic diversity and the nonexistenceof biological races. Hum Biol 75:449–471.
Salzano FM. 1985. Changing patterns of disease among South AmericanIndians. In: Chakraborty R, Szathmary EJE, editors. Diseases of com-plex etiology in small populations. Ethnic differences and researchapproaches. New York: Alan R.Liss. p 301–323.
Tsuneto LT, Probst CM, Hutz MH, Salzano FM, Rodriguez-Delfin LA, ZagoMA, Hill K, Hurtado AM, Ribeiro-dos-Santos AKC, Petzl-Erler ML.2003. HLA class II diversity in seven Amerindian populations. Cluesabout the origins of the Ache. Tiss Antigens 62:512–526.
Wang S, Lewis CM Jr, Jakobsson M, Ramachandran S, Ray N, Bedoya G,Rojas W, Parra MV, Molina JA, Gallo C, Mazzotti G, Poletti G, Hill K,Hurtado AM, Labuda D, Klitz W, Barrantes R, Bortolini MC, SalzanoFM, Petzl-Erler ML, Tsuneto LT, Llop E, Rothhammer F, Excoffier L,Feldman MW, Rosenberg NA, Ruiz-Linares A. 2007. Genetic variationand population structure inNative Americans. PLoSGenet 3:2049–2067.
737PARAGUAYAN ACHE ORIGIN
American Journal of Human Biology





























