Embed Size (px)
Citation preview

JOURNAL OF VIROLOGY, Nov. 2004, p. 11726–11738 Vol. 78, No. 210022-538X/04/$08.00�0 DOI: 10.1128/JVI.78.21.11726–11738.2004Copyright © 2004, American Society for Microbiology. All Rights Reserved.
Functional Analysis of the Noncoding Regions of the Uukuniemi Virus(Bunyaviridae) RNA Segments
Kirsten Flick,1† Anna Katz,2†‡ Anna Overby,1,2† Heinz Feldmann,1,3 Ralf F. Pettersson,2and Ramon Flick1,2,3*
Special Pathogens Program, National Microbiology Laboratory, Health Canada, Canadian Science Centre for Human andAnimal Health (CSCHAH),1 and Department of Medical Microbiology, University of Manitoba,3 Winnipeg, Canada,
and Ludwig Institute for Cancer Research, Stockholm Branch, Karolinska Institute, Stockholm, Sweden2
Received 21 April 2004/Accepted 10 June 2004
The role of the variable portion of the noncoding regions (NCRs) of the three Bunyaviridae RNA segments(L, M, S) in transcription, replication, and packaging was studied using the recently developed plasmid-drivenRNA polymerase I minigenome system for Uukuniemi (UUK) virus, genus Phlebovirus (11), as a model.Comparison of the different segments showed that all NCRs were sufficient to mediate transcription/replicationof a minigenome but demonstrated decreased promoter strength in the order M > L > S. Chimeric minige-nomes with flanking NCRs from different genome segments revealed that the number of total base pairs withinthe inverted, partially complementary ends was important for transcription and replication. Point mutationsincreasing the base-pairing potential produced increased reporter expression, indicating that complementaritybetween the 5� and 3� ends is crucial for promoter activity. The role of the intergenic region (IGR) locatedbetween the two open reading frames of the ambisense UUK virus S segment was analyzed by inserting thissequence element downstream of the reporter genes. The presence of the IGR was found to enhance reporterexpression, demonstrating that efficient transcription termination, regulated by the IGR, is important foroptimal minigenome mRNA translation. Finally, genome packaging efficacy varied for different NCRs and wasstrongest for L followed by M and S. Strong reporter gene activity was still observed after seven consecutivecell culture passages, indicating a selective rather than random genome-packaging mechanism. In summary,our results demonstrate that the NCRs from all three segments contain the necessary signals to initiatetranscription and replication as well as packaging. Based on promoter strength, M-segment NCRs may be thepreferred choice for the development of reverse genetics and minigenome rescue systems for bunyaviruses.
Members of the Bunyaviridae family, which comprises morethan 300 viruses grouped into the five genera (Orthobunyavirus,Hantavirus, Nairovirus, Phlebovirus, and Tospovirus), are envel-oped viruses with a tripartite, single-stranded RNA genome ofnegative polarity. The L segment encodes the RNA-dependentRNA polymerase (L) (9), the M segment encodes the twoglycoproteins (GN and GC) and, in some viruses, a nonstruc-tural protein (NSM) (25), and the S segment encodes the nu-cleoprotein (N) and, in some viruses, a nonstructural protein(NSs) (8, 26, 29).
Viruses belonging to this family share several common fea-tures with other negative-strand RNA viruses. The templatesfor viral polymerase-catalyzed transcription and replication arethe ribonucleoproteins (RNP), which for bunyaviruses consistof the full-length RNA segments associated with the nucleo-protein N and the viral polymerase L. The genomic viralRNA (vRNA) segments contain genes in antisense orientationflanked by stretches of noncoding nucleotides (26). The regu-latory elements for viral transcription and replication, as wellas encapsidation and packaging signals, are thought to be lo-cated within these noncoding regions (NCRs) (26). The termi-
nal nucleotides are genus specific and highly conserved, andbecause of their partial inverted complementarities they canform double-stranded regions leading to circular RNAs (17),providing the functional promoter region for interaction withthe viral polymerase (12). The function of the remaining nu-cleotides of the NCRs is still not well understood, despite thefact that several reverse genetics and minigenome rescue sys-tems for different bunyaviruses have recently been developed(3, 4, 7, 10, 11, 13, 18, 24). However, an encapsidation site hasbeen characterized within the 5� NCR of the S vRNA segment,using the Orthobunyavirus Bunyamwera as a model (21). Stud-ies using influenza virus reverse genetics recently revealed apackaging signal within the highly conserved parts of the NCRs(30) and parts of the open reading frames (ORFs) (15, 32), aswell as nucleotides which influence segment transcription lev-els (14).
A special feature of viruses belonging to the genus Phlebo-virus is the ambisense S segment, which contains two geneslocated in different orientations: the N gene in antisense andthe S RNA-encoded nonstructural protein (NSs) gene in senseorientation. Therefore, only the N gene can be transcribed di-rectly from the genomic vRNA template, whereas the comple-mentary replication intermediate RNA (cRNA) serves as tem-plate for NSs gene transcription (26). In Uukuniemi (UUK)virus (a phlebovirus) the two S-segment-encoded genes areseparated by a 75-nucleotide (nt) intergenic region (IGR),which harbors transcription termination signals for the synthe-sis of the two subsegmental N and NSs mRNAs (29).
* Corresponding author. Present address: Department of Pathology,University of Texas Medical Branch, 301 University Blvd., Galveston,TX 77555-0609. Phone: (409) 747-2962. Fax: (409) 747-2437. E-mail:[email protected].
† K.F., A.K., and A.O. contributed equally to this work.‡ Present address: Department of Virology, Haartman Institute,
University of Helsinki, Helsinki, Finland.
11726
on April 11, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

We used a recently developed reverse genetics system forthe UUK virus (11) to analyze the function of the NCRs as wellas the influence of the IGR on viral transcription and replica-tion. Furthermore, we have compared the promoter activitiesas well as the packaging efficiencies of the L, M, and S vRNA-based minigenomes and investigated the influence of comple-mentarity between segment NCRs on transcription and repli-cation.
MATERIALS AND METHODS
Cells and virus. BHK-21 cells (American Type Culture Collection) weregrown on plastic dishes in Glasgow minimal essential medium (GMEM) supple-
mented with 5 to 10% fetal calf serum, 2 mM L-glutamine, 100 IU of penicillin/ml, and 100 �g of streptomycin/ml (Invitrogen/Life Technologies).
Construction of plasmids. Pol I-driven UUK virus minigenome constructswere generated by using pRF108 (12) as a murine (m) pol I promoter- andterminator-containing plasmid for inserting different PCR fragments. This plas-mid was used to deliver the pol I transcription cassette (Fig. 1).
UUK virus S-reporter gene constructs. For construction of S-segment-basedminigenomes, primers containing the complete 5� (25 nt, positions 1 to 25) and3� (34 nt, positions 1687 to 1720) vRNA NCRs of the UUK virus S segment wereused to amplify the chloramphenicol acetyltransferase (CAT) or green fluores-cent protein (GFP) reporter genes in antisense (CAT, primers RF289 andRF291; GFP, primers RF290 and RF292) or sense (CAT, primers RF294 andRF295; GFP, primers RF296 and RF297) orientation (Fig. 2). PCR fragments
FIG. 1. Schematic representation of the different UUK virus minigenomes used for reverse genetics studies. PCR-amplified UUK virusminigenome fragments were inserted into pRF108, a plasmid containing the murine pol I promoter and terminator for pol I-driven transcription(12). To facilitate readout of minigenome expression levels, the UUK virus RNA segment ORFs were replaced with the gene encoding CAT (leftpanel) or GFP (right panel). The reporter genes are flanked by the NCRs of the UUK virus S, M, and L segments. The lengths of the 5� and 3�NCRs are presented in Fig. 3A. Insertion of the intergenic region sequence is marked with IGR.
VOL. 78, 2004 FUNCTIONAL ANALYSIS OF UUK VIRUS NONCODING RNA REGIONS 11727
on April 11, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

FIG. 2. Oligonucleotide primers used to construct plasmids. Italics, UUK-specific sequences; shading, reporter or viral gene start and stopcodons; boldface, restriction endonuclease recognition sequences; underlining, introduced nucleotide changes.
11728 FLICK ET AL. J. VIROL.
on April 11, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

were inserted into the BsmBI/BbsI fragment of the pRF108 (12) plasmid (Fig. 1).This resulted in four constructs, two with an antisense (�) reporter gene(pRF287, pol I [m] UUK S-CAT [�]; pRF288, pol I [m] UUK S-GFP [�]) andtwo with a reporter gene in sense (�) orientation (pRF289, pol I [m] UUKS-CAT [�]; pRF290, pol I [m] UUK S-GFP [�]). It should be noted that allgenerated minigenomes, independent of reporter gene orientation, containedsimilar Kozak sequences (purine at positions �3 and �1) to avoid differences intranslation initiation efficiencies (pRF287 S-CAT [�], AGATCATGG; pRF289S-CAT [�], TAAGCATGG; pRF288 S-GFP [�], AGATCATGG; pRF290 S-GFP [�], TAAGCATGG [CAT gene start codons are underlined]).
UUK virus S-reporter gene constructs containing the IGR. For further anal-yses of the UUK virus S-segment NCRs, the IGR sequence (UUK virus S vRNA,positions 848 to 921) was inserted into the four different UUK virus S-reportergene minigenomes as described above (pRF287, pRF288, pRF289, and pRF290).PCR primers were designed containing the 5� UUK virus S NCR (RF327) andthe 3� UUK virus S NCR (RF326) adjacent to the IGR sequence, respectively(Fig. 2). By using the BbsI-restricted PCR fragment RF327/291 or RF326/RF294for insertion into the BsmBI/BbsI fragment of the pRF108 vector (Fig. 1), CAT re-porter gene-containing minigenome constructs were generated and named pRF312(pol I UUK virus S-CATIGR [�]) and pRF310 (pol I UUK virus S-CATIGR [�]),respectively. Similar constructs containing the GFP reporter gene flanked by theUUK virus S-segment NCRs and the IGR sequence were generated by combin-ing BbsI-restricted PCR product RF327/292 or RF326/RF296 with the BsmBI/BbsI fragment of pRF108, resulting in pRF313 (pol I UUK virus S-GFP [�]IGR) and pRF311 (pol I UUK virus S-GFP [�] IGR), respectively.
UUK virus L-reporter gene constructs. For analyzing UUK virus minigenomesencoding reporter genes flanked by the UUK virus L-segment NCRs, the 5� (95nt, vRNA positions 1 to 95) and 3� (18 nt, vRNA positions 6406 to 6423) NCRswere added to the CAT or GFP reporter genes in two consecutive PCRs. Primersused in the first PCR provided the complete 3� UUK virus L-NCR (CAT, RF299;GFP, RF301) and the last 35 nt of the 5� UUK virus L-NCR (CAT, RF316; GFP,RF317), whereas the second PCR, using the first amplification product PCRRF316/RF299 or PCR RF317/RF301 as a template (diluted 1:100), added theremaining first 60 nt of the 5� UUK virus L-NCR (CAT, RF298B/RF299; GFP,RF298B/RF301). Insertion of the BbsI-restricted PCR products into the BsmBI/BbsI fragment of pRF108 resulted in two different UUK virus L-segment-basedminigenome constructs, where the L ORF is exactly replaced by either the CATor GFP reporter genes pRF293 (pol I UUK virus L-CAT) and pRF294 (pol IUUK L-GFP).
Chimeric UUK virus minigenomes. For analyzing UUK virus minigenomescontaining NCRs derived from different genome segments, CAT gene-contain-ing UUK virus L, M, and S minigenome constructs pRF293, pRF200, andpRF312, respectively, were treated with EcoRI and NheI restriction endonucle-ases, resulting in two fragments of approximately 3,350 bp (containing the plas-mid backbone, including the pol I promoter and the 5� NCR) and 450 bp(containing the pol I terminator and the 3� NCR) in length. By combining thelarger fragment from one plasmid with the smaller fragment from anotherconstruct, UUK virus minigenomes with NCRs derived from different UUK virusRNA segments were generated: pRF367 (UUK S-CAT-M), pRF368 (UUKM-CAT-S), pRF369 (UUK L-CAT-M), pRF370 (UUK M-CAT-L), pRF371(UUK L-CAT-S), and pRF372 (UUK S-CAT-L).
Transfection and superinfection with UUK virus. BHK-21 and BSR cell lineswere seeded in 6-cm-diameter tissue culture dishes and were transfected with thedifferent minigenome plasmid DNAs using Lipofectamine 2000 reagent (Invitro-gen/Life Technologies). Transfections were performed as described previously(12). To determine the efficiency of transfection, the plasmid pHL2823 contain-ing an enhanced GFP under the control of the cytomegalovirus (CMV) promoter(R. Flick and G. Hobom, unpublished data) was similarly transfected. Trans-fected cells were superinfected 24 h posttransfection with UUK virus at a mul-tiplicity of infection (MOI) of 1 to 3. Briefly, 150 �l of virus stock, diluted in 350�l of cell culture medium (without fetal calf serum [FCS] and antibiotics), wasincubated for 1 h in 6-cm-diameter cell culture plates (37°C, 5% CO2). Unad-sorbed virus was removed and 5 ml of cell culture medium (with 5% FCS andantibiotics) was added.
Passaging of recombinant UUK virus. BHK-21 cells were transfected as de-scribed above and were superinfected 24 h later with UUK virus at an MOI of 1to 3. Cells were analyzed for reporter gene expression 72 hours postinfection(hpi), and the corresponding supernatants were used for virus passaging. Celldebris was removed by centrifugation at 3,000 � g for 10 min, and cells (approx-imately 3 � 106 BHK-21) were infected with 2 ml or 200 �l of undilutedsupernatant. After a 1-h incubation period (37°C, 5% CO2) the inoculum wasreplaced by fresh medium (GMEM, 5% FCS, antibiotics) and cells were incu-bated for 72 h. This procedure was repeated as indicated in successive passages.
CAT assays. Cell extracts were prepared as described by Gorman et al. (16),and CAT activity was determined using a commercially available Flash Cat kit(Molecular Probes, Eugene, Oreg.) as described previously (11, 12). The reactionproducts were visualized by UV illumination, documented by photography, andevaluated using WinCam software (Cybertech, Berlin, Germany) or QuantityOne (Bio-Rad). Ratios of activities were calculated based on at least threeindependent sets of serial dilutions of cell lysates down to a level of 30 to 50%product formation for better quantification within a linear range.
Cell fixation and UV microscopy. Cells transfected with GFP-containing mini-genome constructs and either cotransfected with pCMV UUK-L and pCMVUUK-N (expression plasmids under CMV control for UUK virus L and Nproteins [11]) or superinfected with UUK virus were fixed with 4% paraformal-dehyde. GFP expression was visualized using an Axioplan 2 microscope (Zeiss)and documented using a 3CDC color video camera (DXC-970MD; Sony) andthe imaging software Northern Eclipse 6 (Empix Imaging, Inc.). Alternatively,cells were trypsinized before fixation with 4% paraformaldehyde and were ana-lyzed using fluorescence-activated cell sorting (FACSCalibur; Becton Dickin-son).
RESULTS
Analysis of the terminal NCRs of the ambisense UUK virusS segment. To analyze the NCRs of the ambisense UUK virusS segment which are responsible not only for the S segmentreplication but also for transcription of the N and the NSsgenes, minigenomes containing the CAT or GFP reporter genein either sense or antisense orientation and replacing either theNSs or N ORF were generated (Fig. 1). This allowed us tostudy the cis-acting sequences for the two S-segment-carryinggenes separately. In a first series of experiments, constructswere generated where the CAT or GFP reporter gene wasinserted in sense or antisense orientation between the 25 nt ofthe 5� NCR and the 34 nt of the 3� NCR of the UUK virus SvRNA. Antisense-oriented reporter gene constructs (UUK S-CAT [�], pRF287; UUK S-GFP [�], pRF288) (Fig. 1) simu-late transcription of the N gene (vRNA3 [cRNA3 vRNA]3mRNA), whereas transcription of the NSs gene (vRNA 3cRNA 3 mRNA) was analyzed using constructs containingsense-oriented reporter genes (UUK S-CAT [�], pRF289;UUK S-GFP [�], pRF290) (Fig. 1). Pol I-driven (20, 33) mini-genome constructs were transfected into BHK-21 cells andwere cotransfected with pCMV UUK-L and pCMV UUK-N,which provide the viral L and N proteins. After pol I transcrip-tion in the nucleus, the artificial UUK virus minigenomes aretransported to the cytoplasm, where they are encapsidated bythe N protein and are replicated and transcribed by the Lpolymerase. The level of reporter gene expression reflects theefficiency of transcription and replication from the promoterregions located in the flanking NCRs. The analysis of all testedminigenome constructs resulted in reporter gene expression(Fig. 3A, panels c, d, g, and h; B, columns 3, 5, 7, and 9; insert,lanes 2 and 6), confirming that the terminal NCRs of the SRNA contain all regulatory elements for encapsidation, repli-cation, and transcription of viral genome S segments. No re-porter gene activity could be detected by omitting the UUK-Land UUK-N expression plasmids (Fig. 3A, panels a and b;insert, lanes 1 and 5). Similarly, CAT and GFP reporter geneexpression levels demonstrated equal promoter activity of the3� cRNA (5� vRNA) NCR (pRF289/pRF290), responsible forthe NSs gene transcription start, compared to the 3� vRNANCR (pRF287/pRF288) serving as the transcription start pointfor the N gene (Fig. 3A, panels c and d versus g and h; B, column3 versus 5 and column 7 versus 9; insert, lane 2 versus 6).
VOL. 78, 2004 FUNCTIONAL ANALYSIS OF UUK VIRUS NONCODING RNA REGIONS 11729
on April 11, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

Analysis of the IGR. To determine the role of the IGR in thetranscription and replication processes of the UUK virusS-derived minigenomes, the 75-nt-long sequence located be-tween the two S-segment-encoded N and NSs genes was
inserted into the UUK virus S-segment-based minigenomeconstructs described above (pRF287, pRF288, pRF289, andpRF290). For antisense constructs the IGR was inserted ex-actly between the stop codon of the reporter gene and the 5�
FIG. 3. Role of the intergenic region for minigenome expression levels. Different UUK virus S-segment-based minigenome constructs wereanalyzed for reporter gene expression to define the role of the IGR of the ambisense UUK virus S segment. Minigenomes were transfected intoBHK-21 cells and were analyzed 24 or 48 h posttransfection for CAT or GFP reporter gene expression, respectively. (A) Analyses of UUK virusGFP minigenome expression by FACS. Transfection efficiency was determined, and average GFP intensity was measured. In the dot plots eachdot represents 1 out of 10,000 analyzed cells; signals in the lower left corner represent the cells without GFP expression, and signals in the upperleft corner represent the GFP-expressing cells. In the FACS histogram the y axis represents the cell numbers (%) and the x axis represents the GFPintensities. (B) Quantification of GFP and CAT expression. Column 1, reporter gene background activity in BHK-21 cells upon pRF311 (UUKS-GFPIGR) transfection; column 2, reporter gene background activity after transfection of pRF310 (UUK S-CATIGR); columns 3 to 6, average GFPintensities after transfection with different GFP-containing UUK virus S-segment-based minigenomes and cotransfected UUK-L and -N expressionplasmids; columns 7 to 10, CAT activity in BHK-21 cells transfected with different CAT-containing UUK virus S-segment-based minigenomes andcotransfected with pCMV UUK-L and pCMV UUK-N. Insert lanes 1, 3, 5, and 7 show CAT reporter gene background activity after transfectionwith pol I-driven UUK virus S-CAT minigenomes, omitting the UUK-L and -N expression plasmids. Lanes 2, 4, 6, and 8 of the insert show CATactivity after transfection with the different UUK virus S-CAT constructs cotransfected with pCMV UUK-L and -N plasmids.
11730 FLICK ET AL. J. VIROL.
on April 11, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

vRNA NCR (Fig. 1). For UUK virus minigenomes with areporter gene in sense orientation, the IGR was inserted be-tween the 3� vRNA NCR and the stop codon of the reportergene (Fig. 1). This resulted in two CAT-containing (UUKS-CATIGR [�], pRF312; UUK S-CATIGR [�], pRF310) andtwo GFP-containing (UUK S-GFPIGR [�], pRF313; UUKS-GFPIGR [�], pRF311) minigenome constructs. Cotrans-fection of each of these plasmids with the expression plas-mids for the UUK virus L and N proteins (pCMV UUK-Land pCMV UUK-N) led to reporter gene expression, whichwas compared to the corresponding constructs without theinserted IGR sequence (Fig. 3). No reporter gene expres-sion could be detected in the absence of cotransfected UUKvirus L and N expression plasmids (Fig. 3B, lanes 1 and 2;insert, lanes 3 and 7), confirming that there is no back-ground for any of the different IGR-containing minige-nomes. For CAT analysis, cells were harvested 24 h post-transfection and CAT activity was determined from cell lysates.IGR-containing minigenomes both showed a strong increase inreporter gene expression levels compared to that of corre-sponding constructs without the IGR sequence for the anti-sense- and the sense-oriented reporter gene constructs (Fig.3B, column 7 versus 8 and column 9 versus 10; insert, lane 2versus 4 and lane 6 versus 8).
GFP expression levels, determined 48 h posttransfection byusing UV microscopy and FACS analysis, were significantlyhigher (average GFP intensity was 3 to 6 times stronger) aftertransfection of IGR-containing minigenomes compared to thatof constructs lacking the IGR sequence (Fig. 3A, panel c, paneld versus e, f, and g, panel h versus i, and panel j; B, column 3versus 4 and column 5 versus 6). These results suggest thatgene expression from UUK virus minigenomes based on theambisense S segment is improved by inserting the IGR se-quence downstream of the ORF.
Comparison of promoter activities between the three UUKvirus genome segment NCRs. In order to compare the activityand efficiency of cis-acting elements within the terminal NCRsof the three UUK virus RNA genome segments (Fig. 4A), wegenerated eight different pol I-driven UUK virus minigenomesby replacing the viral genes N, NSs, M, and L with CAT orGFP reporter genes (Fig. 1, pRF200 [M-CAT] [11], pRF312[SN-CAT], pRF310 [SNSs-CAT], pRF293 [L-CAT], pRF31 [M-GFP] [11], pRF313 [SN-GFP], pRF311 [SNSs-GFP], pRF294[L-GFP]). Each UUK virus minigenome construct was trans-fected into BHK-21 or BSR cells, and cells were cotransfectedwith the CMV-driven expression plasmids pCMV UUK-L andpCMV UUK-N. Cells transfected with the CAT-containingminigenomes were harvested 24 h posttransfection, and CATactivity was assayed from cell lysates (Fig. 4B, left panel). Inthe case of UUK virus GFP minigenomes, cells were fixed 48 hposttransfection with paraformaldehyde and GFP expressionwas determined using UV microscopy (data not shown) andFACS analysis (Fig. 4B, right panel). Transfection with alltested minigenome constructs resulted in reporter gene expres-sion, confirming that the NCRs of each segment contain allregulatory elements for encapsidation, replication, and tran-scription of the artificial viral genome segments. Similar CATactivities were measured for each construct in BHK-21 versusBSR cells (comparison not shown). No reporter gene activity
could be detected by omitting the UUK-L and UUK-N expres-sion plasmids (data not shown).
The UUK virus CAT-containing reporter gene flanked bythe UUK virus M segment NCRs (pRF200) showed the high-est reporter gene expression level compared to those of theCAT reporter flanked by the L (pRF293) and S segment NCRs(pRF310 and pRF312) (Fig. 4B, lane 1 versus 2, 3, and 4). TheL NCRs displayed a higher promoter activity compared to thatof the UUK virus S-CAT minigenomes (Fig. 4B, lane 2 versus3 and 4).
These results were confirmed by using the UUK virus mini-genomes containing GFP as the reporter gene. The M NCR-flanked GFP minigenome (pRF31) clearly showed the stron-gest GFP expression level, followed by the UUK virus L-GFP(pRF294) and the UUK virus S-GFP constructs (pRF311 andpRF313) (Fig. 4B, lane 11 versus 12 versus 13 and 14). It isnoteworthy that the different expression levels from the ana-lyzed UUK virus minigenomes were not based on different polI minigenome transcription levels (data not shown).
To determine the expression kinetics for each UUK virussegment-based minigenome, a time course experiment wasperformed. Pol I-driven minigenome plasmids were trans-fected together with UUK-L and -N expression plasmids. Cellswere harvested at 10 to 70 h posttransfection, and CAT activitywas determined from cell lysates (Fig. 4C). At 10 h posttrans-fection M- and L-segment-based minigenomes showed veryweak CAT activities, whereas no reporter gene activity couldbe detected at this time for the two S-segment-based con-structs. A strong increase in CAT expression could be observed22 h posttransfection. Minigenomes flanked by the L- or S-segment NCRs reached their maximum expression levels at28 h, whereas the M minigenome expression did not reach aplateau until 46 h, after which it remained high up to 70 hposttransfection (Fig. 4C).
For a more accurate comparison of the S-NCR-derived mini-genomes, dilution series of minigenome-transfected cell lysateswere used and analyzed for CAT expression. A minor but sig-nificant difference between the promoters responsible for theN or NSs gene expression could be observed (Fig. 4B, middlepanel, lane 5 versus 6, lane 7 versus 8, and lane 9 versus 10),demonstrating that the S-segment-based minigenome with theN gene replaced by a CAT gene is somewhat more efficiently ex-pressed than the constructs in which the NSs gene is replaced.
Taken together, comparison of UUK virus minigenomesbased on all three different UUK virus genome segments re-vealed that the strongest promoter activity is located within the185 nt of the 5� NCR and 17 nt of the 3� NCR of the UUK virusM segment.
Passaging of recombinant UUK viruses. We next analyzedwhether the minigenomes could be packaged into infectiousUUK virus particles and serially passaged to fresh cell cultures.As was recently described for the M-segment-based minige-nome (11), we expected that the reporter gene expression levelwould decrease with the number of transfers, due to the factthat there is no pressure for the virus to keep an additionalreporter gene-encoding segment.
To determine the influence of the different NCRs on pack-aging efficiency, BHK-21 cells were transfected with pol I-driven minigenomes containing the NCRs of any of the threegenomic RNA segments and were cotransfected with UUK-L
VOL. 78, 2004 FUNCTIONAL ANALYSIS OF UUK VIRUS NONCODING RNA REGIONS 11731
on April 11, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

FIG. 4. Comparison of promoter strengths of the three UUK virus RNA segments. RNA pol I-driven UUK virus minigenome constructscontaining the CAT or GFP reporter gene flanked by the NCRs of the three different RNA genome segments L, M, and S were transfected intoBHK-21 cells. Cotransfection of expression plasmids pCMV UUK-L and pCMV UUK-N provides the L and N proteins required for transcription
11732 FLICK ET AL. J. VIROL.
on April 11, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

and UUK-N expression plasmids. To provide the necessary packaging ma-chinery, cells were superinfected with UUK virus 24 h post-transfection and aliquots (2 ml or 200 �l) from supernatantswere transferred 72 hpi to fresh cell cultures. From the trans-fected and superinfected cells, as well as from each consecutivepassage, reporter gene activity was determined at 72 hpi. Thiswas repeated six times, resulting in a total of seven passages.
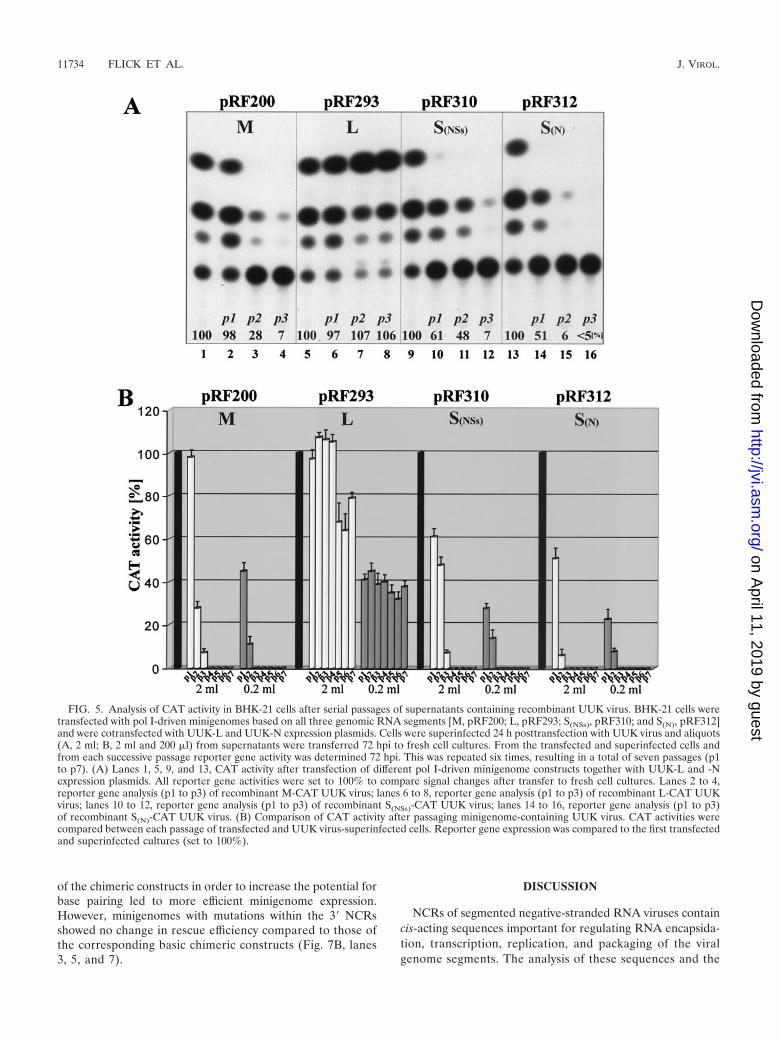
All minigenomes could be efficiently rescued by serially pas-saging tissue culture medium once (Fig. 5A, lanes 2, 6, 10, and14), demonstrating that cis-active signals responsible for RNAincorporation into UUK progeny viruses are located within theflanking NCRs of all three genomic RNA segments. Mini-genome constructs based on the M (pRF200) or S segment(pRF310 and pRF312) showed rapidly decreasing levels ofreporter gene expression during the successive transfers (Fig.5A, lanes 1 to 4, 9 to 12, and 13 to 16, respectively; see alsopanel B). After three passages, only weak CAT activities weredetectable for pol I UUK M-CAT (for pRF200, see Fig. 5A,lane 4, and B) and pol I UUK SNSs-CAT (for pRF310, see Fig.5A, lane 12, and B), whereas no reporter gene expression wasmeasurable for pol I UUK SN-CAT (for pRF312, see Fig. 5A,lane 16, and B). In contrast, the L-segment-based artificialvRNA (pol I UUK L-CAT [pRF293]) showed slightly in-creased CAT activities during the first four passages (Fig. 5A,lanes 5 to 8, and B), indicating a more efficient packaging eventfor this minigenome than for that of the M- and S-segment-based RNAs. Even after seven passages, about 75% of theCAT reporter expression level of the primary transfected andsuperinfected cells was observed (Fig. 5B).
In summary, the passaging experiments demonstrated thatcis-acting signals responsible for the incorporation of RNAsegments into UUK virus particles are located within the twoflanking NCRs of each genomic segment but that there areclear differences in packaging efficiencies (L � M � SNSs � SN).
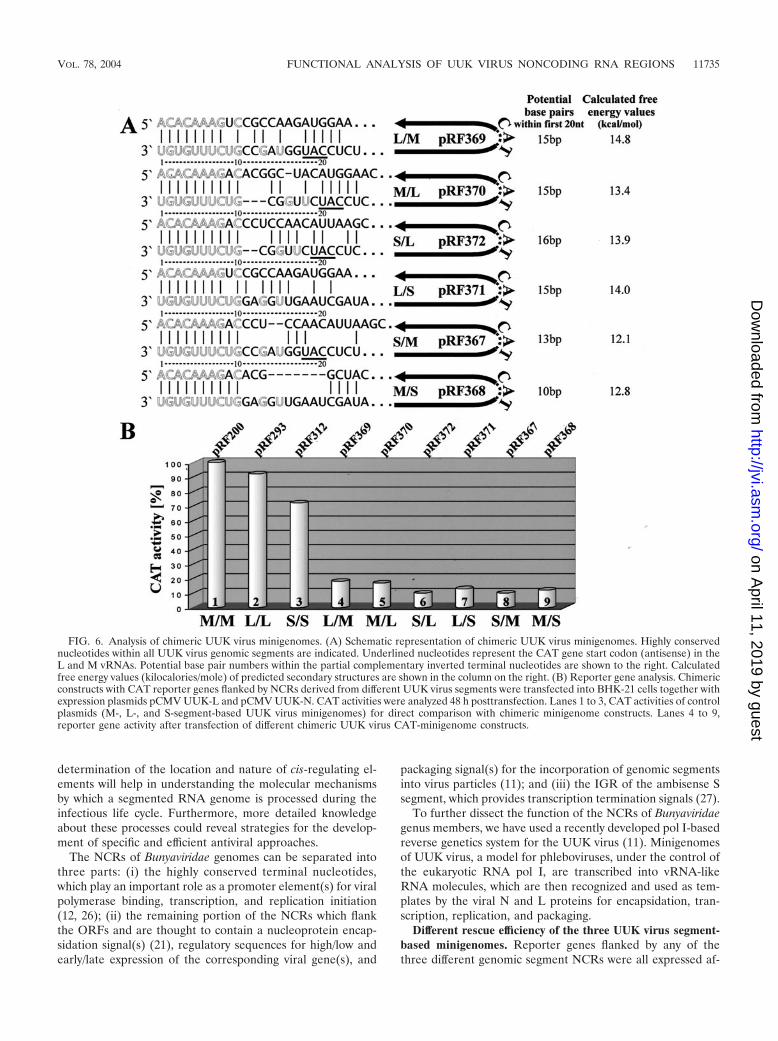
Chimeric UUK virus minigenomes. To further examine thebasis for the differing promoter activities within the three ge-nomic RNA segments, we generated chimeric constructs withthe reporter gene flanked by NCRs derived from differentUUK virus segments. The CAT gene-containing UUK virus L,M, and S minigenome constructs pRF293, pRF200, and pRF312,respectively, were treated with EcoRI and NheI restrictionendonucleases, resulting in two fragments approximately 3,350and 450 bp in length. By combining the larger fragment fromone plasmid with the smaller fragment from another construct,the following UUK virus minigenomes with NCRs derivedfrom different UUK virus RNA segments were generated:
pRF367 (UUK S-CAT-M), pRF368 (UUK M-CAT-S), pRF369(UUK L-CAT-M), pRF370 (UUK M-CAT-L), pRF371 (UUKL-CAT-S), and pRF372 (UUK S-CAT-L) (Fig. 6A). Theseplasmids were individually transfected into BHK-21 cells andcotransfected UUK-L and -N expression plasmids (pCMVUUK-L and pCMV UUK-N). Comparison of these chimericUUK virus minigenomes revealed that combining NCRs de-rived from different UUK virus segments resulted in an almostcomplete loss of reporter gene expression (Fig. 6B, lanes 4 to9) compared to that of the three wild-type UUK virus con-structs (Fig. 6B, lanes 1 to 3). No significant difference in CATexpression from the various chimeric constructs could be de-termined.
For structural comparison, the first and last 25 nt of thechimeric minigenome sequence, connected by a stretch of 20uracils, were used as a basis for secondary structure prediction(GeneBee) (5, 6). The calculated total free energy (kilocalo-ries/mole) revealed a substantially reduced predicted stabilityof the chimeric constructs compared to that of the wild-typeNCRs (Fig. 4A and 6A). The first 20 nt of both vRNA ends,which play a major role during the transcriptional start, canform 18 to 20 bp for the UUK virus L, M, and S segments (Fig.4A). However, the NCRs of the chimeric minigenomes havethe potential for only 10 to 16 bp (Fig. 6A). The missinginteraction of nucleotides from the segment ends can causedramatic changes in the secondary structure of the NCRs. Inparticular, the predicted length of stem structures and inter-rupting loops (data not shown) and, therefore, the correspond-ing free energy values (kilocalories/mole) of the RNA second-ary structures differ substantially between the chimeric andwild-type minigenomes (Fig. 4A and 6A). Based on these anal-yses we introduced point mutations into the first or last 20 ntof each chimeric minigenomes by using oligonucleotide-di-rected mutagenesis to increase the number of potential basepairs within the terminal segment regions. We either intro-duced multiple mutations (exchanges and deletions) into the 5�NCR, adjusting the sequence to that of the 3� NCR of thechimeric construct, or vice versa (Fig. 7A). The resulting con-structs contained 16 to 20 potential base pairs within the first20 terminal nucleotides (Fig. 7A). Cotransfection of these polI-driven chimeric minigenome plasmids with restored basepairing, together with expression plasmids encoding the viral Land N proteins, resulted in substantially increased reportergene expression for three of the constructs compared to that ofthe original chimeric constructs containing 5� NCR mutations(Fig. 7B, lanes 2, 4, and 6). Therefore, adjusting the two ends
and replication of the minigenomes. (A) Schematic representation of the terminal nucleotides of UUK virus genome segments. Highly conservednucleotides within all UUK virus genomic segments are indicated. Underlined nucleotides represent the gene start codon (antisense) in the L andM vRNAs. Nucleotide numbers within the flanking 5� or 3� NCRs are shown to the right. Calculated free energy values (kilocalories/mole) ofpredicted secondary structures are shown in the column on the right. (B) UUK virus CAT and GFP minigenomes. Twenty-four or 48 hposttransfection cells were harvested and analyzed for CAT activity or GFP expression, respectively. Lanes 1 to 4, CAT activity in BHK-21 cellsafter cotransfection of different UUK virus CAT minigenome constructs and UUK virus L and N expression plasmids; lanes 5 to 10, comparisonof CAT activity from the plasmids S(NSs) (pol I UUK S-CATIGR [�]) and S(N) (pol I UUK S-CATIGR [�]) to determine promoter activities of theambisense UUK virus S segment for NSs versus N transcription by using dilution series of extracts from BHK-21-transfected cells; lanes 11 to 14,FACS analysis of cells 48 h after transfection with different UUK virus GFP minigenomes. Average GFP intensities (from three independentexperiments) of BHK-21 cells after cotransfection of UUK virus GFP minigenome constructs and UUK virus L and N expression plasmids arepresented. (C) Time course experiment. BHK-21 cells were transfected with UUK virus genome segment-based reporter plasmids and werecotransfected with UUK virus L and N expression plasmids. Cells were harvested at the indicated time points posttransfection and were analyzedfor CAT activity. BHK-21 cells only transfected with minigenome served as a negative control (MOCK).
VOL. 78, 2004 FUNCTIONAL ANALYSIS OF UUK VIRUS NONCODING RNA REGIONS 11733
on April 11, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from
of the chimeric constructs in order to increase the potential forbase pairing led to more efficient minigenome expression.However, minigenomes with mutations within the 3� NCRsshowed no change in rescue efficiency compared to those ofthe corresponding basic chimeric constructs (Fig. 7B, lanes3, 5, and 7).
DISCUSSION
NCRs of segmented negative-stranded RNA viruses containcis-acting sequences important for regulating RNA encapsida-tion, transcription, replication, and packaging of the viralgenome segments. The analysis of these sequences and the
FIG. 5. Analysis of CAT activity in BHK-21 cells after serial passages of supernatants containing recombinant UUK virus. BHK-21 cells weretransfected with pol I-driven minigenomes based on all three genomic RNA segments [M, pRF200; L, pRF293; S(NSs), pRF310; and S(N), pRF312]and were cotransfected with UUK-L and UUK-N expression plasmids. Cells were superinfected 24 h posttransfection with UUK virus and aliquots(A, 2 ml; B, 2 ml and 200 �l) from supernatants were transferred 72 hpi to fresh cell cultures. From the transfected and superinfected cells andfrom each successive passage reporter gene activity was determined 72 hpi. This was repeated six times, resulting in a total of seven passages (p1to p7). (A) Lanes 1, 5, 9, and 13, CAT activity after transfection of different pol I-driven minigenome constructs together with UUK-L and -Nexpression plasmids. All reporter gene activities were set to 100% to compare signal changes after transfer to fresh cell cultures. Lanes 2 to 4,reporter gene analysis (p1 to p3) of recombinant M-CAT UUK virus; lanes 6 to 8, reporter gene analysis (p1 to p3) of recombinant L-CAT UUKvirus; lanes 10 to 12, reporter gene analysis (p1 to p3) of recombinant S(NSs)-CAT UUK virus; lanes 14 to 16, reporter gene analysis (p1 to p3)of recombinant S(N)-CAT UUK virus. (B) Comparison of CAT activity after passaging minigenome-containing UUK virus. CAT activities werecompared between each passage of transfected and UUK virus-superinfected cells. Reporter gene expression was compared to the first transfectedand superinfected cultures (set to 100%).
11734 FLICK ET AL. J. VIROL.
on April 11, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from
determination of the location and nature of cis-regulating el-ements will help in understanding the molecular mechanismsby which a segmented RNA genome is processed during theinfectious life cycle. Furthermore, more detailed knowledgeabout these processes could reveal strategies for the develop-ment of specific and efficient antiviral approaches.
The NCRs of Bunyaviridae genomes can be separated intothree parts: (i) the highly conserved terminal nucleotides,which play an important role as a promoter element(s) for viralpolymerase binding, transcription, and replication initiation(12, 26); (ii) the remaining portion of the NCRs which flankthe ORFs and are thought to contain a nucleoprotein encap-sidation signal(s) (21), regulatory sequences for high/low andearly/late expression of the corresponding viral gene(s), and
packaging signal(s) for the incorporation of genomic segmentsinto virus particles (11); and (iii) the IGR of the ambisense Ssegment, which provides transcription termination signals (27).
To further dissect the function of the NCRs of Bunyaviridaegenus members, we have used a recently developed pol I-basedreverse genetics system for the UUK virus (11). Minigenomesof UUK virus, a model for phleboviruses, under the control ofthe eukaryotic RNA pol I, are transcribed into vRNA-likeRNA molecules, which are then recognized and used as tem-plates by the viral N and L proteins for encapsidation, tran-scription, replication, and packaging.
Different rescue efficiency of the three UUK virus segment-based minigenomes. Reporter genes flanked by any of thethree different genomic segment NCRs were all expressed af-
FIG. 6. Analysis of chimeric UUK virus minigenomes. (A) Schematic representation of chimeric UUK virus minigenomes. Highly conservednucleotides within all UUK virus genomic segments are indicated. Underlined nucleotides represent the CAT gene start codon (antisense) in theL and M vRNAs. Potential base pair numbers within the partial complementary inverted terminal nucleotides are shown to the right. Calculatedfree energy values (kilocalories/mole) of predicted secondary structures are shown in the column on the right. (B) Reporter gene analysis. Chimericconstructs with CAT reporter genes flanked by NCRs derived from different UUK virus segments were transfected into BHK-21 cells together withexpression plasmids pCMV UUK-L and pCMV UUK-N. CAT activities were analyzed 48 h posttransfection. Lanes 1 to 3, CAT activities of controlplasmids (M-, L-, and S-segment-based UUK virus minigenomes) for direct comparison with chimeric minigenome constructs. Lanes 4 to 9,reporter gene activity after transfection of different chimeric UUK virus CAT-minigenome constructs.
VOL. 78, 2004 FUNCTIONAL ANALYSIS OF UUK VIRUS NONCODING RNA REGIONS 11735
on April 11, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

ter transfection of pol I-driven minigenomes into BHK-21 cellsexpressing the viral L and N proteins (Fig. 4). This demon-strated that all necessary signals for RNA encapsidation, tran-scription, and replication are located within these flankingNCRs. The use of minigenomes of similar lengths facilitatedthe comparison of expression efficiencies (promoter strengths)between the three segment-derived RNAs without the inter-ference of various RNA sizes. The M-segment-based con-structs showed the strongest reporter gene expression (CAT
and GFP) followed by the L-derived constructs, whereas therescue efficiencies of minigenomes containing S-segmentNCRs were the lowest (Fig. 4B). This result is surprising con-sidering that the N protein, a gene product of the S segment,is the most abundant protein in UUK virus-infected cells (31).The reason for this result could be partly due to the differentlengths of the wild-type genomic segments (L, 6,423 bp; M,3,229 bp; S, 1,720 bp) compared to the sizes of the analyzedminigenomes (L-CAT, 772 bp; M-CAT, 863 bp; S-CAT, 800
FIG. 7. Analysis of chimeric UUK virus minigenomes with restored complementary ends. (A) Schematic representation of mutated chimericUUK virus minigenomes. Point mutations were introduced into different chimeric UUK virus minigenomes using oligonucleotide-directedmutagenesis (RF508, RF512, and RF514 for 5� vRNA modifications; RF509, RF513, and RF515 for 3� vRNA modifications) (Fig. 2) to increasethe total number of base pairs within the terminal nucleotides. Highly conserved nucleotides within all UUK virus genomic segments arehighlighted. Underlined nucleotides represent the CAT gene start codon (antisense) in the L and M vRNAs. Introduced point mutations aremarked with a box. (B) Reporter gene analysis. BHK-21 cells were transfected with the chimeric reporter plasmids and cotransfected with UUK-Land -N expression plasmids. Reporter gene activity was analyzed 48 h posttransfection. Lane 1, CAT activity of cells transfected with S-segment-based UUK virus minigenomes (set at 100%); lanes 2, 4, and 6, chimeric UUK virus minigenomes with point mutations introduced in the 5� NCRsto restore base pairing with the 3�-terminal nucleotides; lanes 3, 5, and 7, chimeric UUK virus minigenomes with nucleotide changes in the 3� NCRto restore base pairing with 5�-terminal nucleotides.
11736 FLICK ET AL. J. VIROL.
on April 11, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

bp; L-GFP, 854 bp; M-GFP, 947 bp; S-GFP, 884 bp). In thewild-type RNA segments the overall length could influenceviral gene expression, as shown in similar studies for influenzavirus (M. Azzeh, R. Flick, and G. Hobom, unpublished data).Our results here suggest that the longer M and L segmentsneed stronger promoter activities than the S segment to com-pensate for their larger size. Interestingly, in a time courseexperiment comparison of reporter gene expression efficiencyrevealed that M minigenomes showed the strongest CAT ac-tivity at all time points examined, followed by the L-segment-based construct and the two S-segment-based constructs. How-ever, the different UUK virus minigenomes did not follow thesame kinetics. All UUK virus minigenomes showed a strongincrease of reporter gene activity between 10 and 22 h post-transfection. The M-segment-based minigenome showed a fur-ther increased reporter gene expression up to 46 h (Fig. 4C). Incontrast, L- and S-segment-based constructs reached a plateauat 22 to 28 h posttransfection. This is in agreement with thesituation in infected cells where M-segment-directed glycopro-tein expression increases during the late stage of infection forefficient assembly of progeny viruses. The ratio of vRNA seg-ments (L:M:S) in purified virions has been found to be about1:4:2 (22, 23), and similar ratios have been observed in infectedcells (31). This also supports the conclusion that the M RNA isthe most efficiently replicated segment. Using a different re-verse genetics system, Barr et al. (1) recently obtained verysimilar results for BUN virus (Orthobunyavirus genus). In theirstudy, a luciferase reporter gene was flanked by the NCRs fromthe three RNA segments. The relative replication efficiency(promoter strength) of the three minigenomes was also foundto be M � L � S.
To further analyze the reasons for the different rescue effi-ciencies of the three genome segments, chimeric minigenomeswere generated containing NCRs derived from different UUKvirus RNA segments. Surprisingly, all six possible combina-tions (L-S, S-L, M-S, S-M, L-M, and M-L) showed only verylow CAT expression levels (Fig. 6B), demonstrating that NCRsderived from different genome segments do not interact in afunctionally efficient way to regulate RNA encapsidation, tran-scription, and replication processes. Further analysis of thechimeric minigenomes using RNA secondary structure predic-tion software and stability calculation (GeneBee) (5, 6) of thebase-paired terminal nucleotides suggested that a decreasedbase-pairing potential between the terminal nucleotides ofboth NCRs could be the reason for the inefficient rescue ofthe chimeric minigenomes. We therefore introduced multiplepoint mutations (exchanges and deletions) to adjust the first 20nt of the 5� NCR to the last 20 nt of the 3� NCR to increase thebase-pairing potential between the ends (Fig. 7A). These base-paired chimeric constructs showed significantly increased CATsignals (Fig. 7B, lanes 2, 4, and 6) compared to the originalconstructs, indicating that interaction between the terminalnucleotides of each segment is crucial for efficient cis-regula-tory processes. Reporter gene activities from constructs with5�-mutated NCRs were restored to S-segment-based CAT lev-els (Fig. 7B). In contrast, UUK virus minigenomes with mul-tiple nucleotide mutations within the 3� NCR showed only lowrescue efficiencies, comparable to those of the correspondingchimeric constructs (Fig. 7B, lanes 3, 5, and 7). One explana-tion could be that the mutations influenced the Kozak se-
quence of the M and L 3� NCRs, thus interfering with trans-lation initiation. Furthermore, transcription and replicationinitiation takes place at the terminal 3� nucleotides of vRNAmolecules. It was shown previously that not only base pairingbut also the nucleotide sequence at the conserved highly base-paired region at the 3� end of the M RNA NCR were impor-tant for promoter activity (12). Our present results show thatmultiple mutations in the adjacent 3� NCR also affect pro-moter function, and they do so more severely than mutations inthe 5� end. It is noteworthy that restoration of the CAT activityto S-segment-based minigenomes was achieved by rather dras-tic changes of the sequences adjacent to the highly conservedpromoter region of the viral RNA segments by multiple (two tofour) nucleotide exchanges and/or deletions (two) at the 5�end. The results clearly demonstrate that base pairing betweenthe terminal nucleotides of the nonconserved NCRs is neces-sary for efficient transcription and replication processes.
Results similar to ours were recently reported for BUN virus(2). Using hybrid 3� and 5� ends flanking a luciferase reporter,no transcription or replication of reporter RNAs was observed.When base pairing was restored, RNA synthesis was againreadily detected. The authors concluded that the BUN virusNCRs do not act as independent RNA synthesis promoters;rather, the 3� and 5� sequences cooperate to form the func-tional promoter for both RNA replication and mRNA synthe-sis. Our results are in good agreement with these conclusions.
Analysis of S-segment-based minigenomes. To analyze thecis-acting signals responsible for viral N or NSs gene transcrip-tion and replication, we generated minigenomes by insertingreporter genes in sense or antisense orientation between thetwo terminal NCRs of the UUK virus S segment. Surprisingly,CAT and GFP levels were similar for all the constructs, dem-onstrating equal promoter activities for N and NSs gene tran-scription and replication (Fig. 4). However, dilution seriesrevealed that promoter elements in the vRNA orientation,responsible for N gene mRNA transcription, showed moder-ately stronger activities compared to those of cRNA NCRsresponsible for the NSs gene transcription (Fig. 4B). This is inagreement with protein levels during UUK virus infection,where the N protein is more abundant than NSs. In addition,NSs is much less stable than the N protein, contributing to thelower level of NSs observed in infected cells (28).
Additional S-segment-based minigenomes were generatedto analyze the effect of the IGR on minigenome rescue effi-ciency. The IGR harbors the transcription termination signalsfor N and NSs mRNA synthesis (27). The 75-nt-long IGR wasinserted downstream of the reporter gene stop codon. Com-parison of minigenomes with and without IGR revealed anenhancing effect of the IGR on reporter gene expression levels.This was independent of the orientation of the CAT/GFPgene, demonstrating that cis-acting signals located within theIGR play an important role for transcription of the N and NSsgenes.
Our present working hypothesis is that proper transcriptiontermination, regulated by signal elements within the IGR (27,28, 29), eliminates the encapsidation signal located in the 5�vRNA NCR (21) in the mRNA transcript, preventing the bind-ing of N proteins to the mRNA transcripts. In minigenomeswithout IGR no termination signal is available, resulting inmRNA transcripts containing the complete vRNA 5� NCR,
VOL. 78, 2004 FUNCTIONAL ANALYSIS OF UUK VIRUS NONCODING RNA REGIONS 11737
on April 11, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

including the N binding site. This leads to an encapsidatedmRNA transcript and subsequent inhibition of efficient trans-lation. In addition, mRNA transcripts with both completeflanking NCRs (as in the case of constructs without IGR) canform terminal base-paired regions (panhandle-like structures)interfering with translation initiation.
Based on these results, the IGR of ambisense RNA seg-ments should be included when developing minigenome rescuesystems based on the S-segment NCRs in order to improvereporter gene expression levels.
Analysis of packaging efficiencies by passaging experiments.To determine if cis-acting signals responsible for RNA segmentincorporation into virions are located within the two flankingNCRs, minigenomes derived from the three UUK virus geno-mic segments were used in passaging experiments. All minige-nomes were packaged into progeny virus particles as demon-strated by the successful transfer of CAT activity from transfectedand UUK virus-superinfected cells to fresh cell lines. This showedthat signals responsible for segment incorporation are locatedwithin the terminal NCRs; however, packaging efficiencies dif-fered between the minigenomes. M- and S-segment-based RNAscould be detected for two to three passages, whereas after sub-sequent passages no reporter gene activity could be measured(Fig. 5A and B). This was expected, because no system wasused to select for the recombinant viruses containing the CATminigenome against the wild-type virus used for the superin-fection experiment. Similar results were originally reported forinfluenza virus segments. Passaging of a reporter gene flankedby just the 3�- and 5�-terminal NCRs of vRNAs resulted in therapid loss of the transgene from the pool of progeny virus par-ticles (19). Surprisingly, minigenomes containing UUK virusL-segment-derived NCRs could be passaged three times withincreasing CAT signals, whereas subsequent passages (pas-sages 4 to 7) resulted in only a 20 to 30% decrease in reportergene expression levels. This outcome suggests that a stable poolof recombinant UUK viruses containing the L-segment-de-rived minigenomes was generated. Whether the minigenome isincorporated as an additional fourth segment or replaces thewild-type L segment remains to be determined. By analogywith influenza virus vRNA segments (15, 32), it is possible thatadditional packaging signals are present within the coding re-gions of each UUK virus segment. The pol I system shouldmake it possible to identify such putative signals.
ACKNOWLEDGMENTS
We thank Anita Bergstrom for excellent technical assistance. Fur-thermore, we are grateful to Erin Kinsella and Adrienne Meyers forassistance with cloning and Allison Groseth for reviewing the manu-script.
REFERENCES
1. Barr, J. N., R. M. Elliott, E. F. Dunn, and G. W. Wertz. 2003. Segment-specific terminal sequences of Bunyamwera bunyavirus regulate genomereplication. Virology 311:326–338.
2. Barr, J. N., and G. W. Wertz. 2004. Bunyamwera bunyavirus RNA synthesisrequires cooperation of 3�- and 5�-terminal sequences. J. Virol. 78:1129–1138.
3. Blakqori, G., G. Kochs, O. Haller, and F. Weber. 2003. Functional L poly-merase of La Crosse virus allows in vivo reconstitution of recombinantnucleocapsids. J. Gen. Virol. 84:1207–1214.
4. Bridgen, A., and R. M. Elliott. 1996. Rescue of a segmented negative-strandRNA virus entirely from cloned complementary DNAs. Proc. Natl. Acad.Sci. USA 93:15400–15404.
5. Brodsky, L. I., A. V. Vasiliev, L. Kalaidzidis, S. Osipov, R. L. Tatuzov, andS. I. Feranchuk. 1992. GeneBee: the program package for biopolymer struc-ture analysis. Dimacs 8:127–139.
6. Brodsky, L. I., V. V. Ivanov, L. Kalaidzidis, A. M. Leontovich, V. K. Nikolaev,S. I. Feranchuk, and V. A. Drachev. 1995. GeneBee-NET: internet-basedserver for analyzing biopolymers structure. Biochemistry 60:923–928.
7. Dunn, E. F., D. C. Pritlove, H. Jin, and R. M. Elliott. 1995. Transcription ofa recombinant Bunyavirus RNA template by transiently expressed Bunyavi-rus proteins. Virology 211:133–143.
8. Elliott, R. M., C. S. Schmaljohn, and M. S. Collett. 1991. Bunyaviridae genomestructure and gene expression. Curr. Top. Microbiol. Immunol. 169:91–141.
9. Elliott, R. M., E. Dunn, J. F. Simons, and R. F. Pettersson. 1992. Nucleotidesequence and coding strategy of the Uukuniemi virus L RNA. J. Gen. Virol.73:1745–1752.
10. Flick, K., J. Hooper, C. Schmaljohn, R. Pettersson, H. Feldmann, and R.Flick. 2003. Rescue of Hantaan virus minigenomes. Virology 306:219–224.
11. Flick, R., and R. F. Pettersson. 2001. Reverse genetics system for Uukuniemivirus (Bunyaviridae): RNA polymerase I-catalyzed expression of chimericviral RNAs. J. Virol. 75:1643–1655.
12. Flick, R., F. Elgh, G. Hobom, and R. F. Pettersson. 2002. Mutational analysisof the Uukuniemi virus (Bunyaviridae) promoter reveals two regions offunctional importance. J. Virol. 76:10849–10860.
13. Flick, R., H. Feldmann, and F. Elgh. 2003. Reverse genetics for Crimean-Congo hemorrhagic fever (CCHF) virus. J. Virol. 77:5997–6006.
14. Fodor, E., P. Palese, G. G. Brownlee, and A. Garcia-Sastre. 1998. Attenua-tion of influenza A virus mRNA levels by promoter mutations. J. Virol.72:6283–6290.
15. Fujii, Y., H. Goto, T. Watanabe, T. Yoshida, and Y. Kawaoka. 2003. Selectiveincorporation of influenza virus RNA. Proc. Natl. Acad. Sci. USA 100:2002–2007.
16. Gorman, C. M., L. F. Moffat, and B. H. Howard. 1982. Recombinant ge-nomes which express chloramphenicol acetyltransferase in mammalian cells.Mol. Cell. Biol. 2:1044–1051.
17. Hewlett, M. J., R. F. Pettersson, and D. Baltimore. 1977. Circular forms ofUukuniemi virion RNA: an electron microscopic study. J. Virol. 21:1085–1093.
18. Lopez, N., R. Muller, C. Prehaud, and M. Bouloy. 1995. The L protein of RiftValley fever virus can rescue viral ribonucleoproteins and transcribe syn-thetic genome-like RNA molecules. J. Virol. 69:3972–3979.
19. Luytjes, W., M. Krystal, M. Enami, J. D. Pavin, and P. Palese. 1989. Am-plification, expression, and packaging of a foreign gene by influenza virus.Cell 59:1107–1113.
20. Neumann, G., A. Zobel, and G. Hobom. 1994. RNA polymerase I-mediatedexpression of influenza viral RNA molecules. Virology 202:477–479.
21. Osborne, J. C., and R. M. Elliott. 2000. RNA binding properties of bun-yamwera virus nucleocapsid protein and selective binding to an element inthe 5� terminus of the negative-sense S segment. J. Virol. 74:9946–9952.
22. Pettersson, R., and L. Kaariainen. 1973. The ribonucleic acids of Uukuniemivirus, a noncubical tick-borne arbovirus. Virology 56:608–619.
23. Pettersson, R. F., M. J. Hewlett, D. Baltimore, and J. M. Coffin. 1977. Thegenome of Uukuniemi virus consists of three unique RNA segments. Cell11:51–63.
24. Prehaud, C., N. Lopez, M. J. Blok, V. Obry, and M. Bouloy. 1997. Analysisof the 3� terminal sequence recognized by the Rift Valley fever virus tran-scription complex in its ambisense S segment. Virology 227:189–197.
25. Ronnholm, R., and R. F. Pettersson. 1987. Complete nucleotide sequence ofthe M RNA segment of Uukuniemi virus encoding the membrane glycopro-teins G1 and G2. Virology 160:191–202.
26. Schmaljohn, C. S., and J. W. Hooper. 2001. Bunyaviridae: the viruses andtheir replication, p. 1581–1602. In D. M. Knipe and P. M. Howley (ed.),Fields virology, vol. 2. Lippincott Williams and Wilkins, Philadelphia, Pa.
27. Simons, J. F., and R. F. Pettersson. 1991. Host-derived 5� ends and over-lapping complementary 3� ends of the two mRNAs transcribed from theambisense S segment of Uukuniemi virus. J. Virol. 65:4741–4748.
28. Simons, J. F., R. Persson, and R. F. Pettersson. 1992. Association of thenonstructural protein NSs of Uukuniemi virus with the 40S ribosomal sub-unit. J. Virol. 66:4233–4241.
29. Simons, J. F., U. Hellman, and R. F. Pettersson. 1990. Uukuniemi virus SRNA: ambisense coding strategy, packaging of complementary strands intovirions, and homology to members of the genus Phlebovirus. J. Virol. 64:247–255.
30. Tchatalbachev, S., R. Flick, and G. Hobom. 2001. The packaging signal ofinfluenza viral RNA molecules. RNA 7:979–989.
31. Ulmanen, I., P. Seppala, and R. F. Pettersson. 1981. In vitro translation ofUukuniemi virus-specific RNAs: identification of a nonstructural protein anda precursor to the membrane glycoproteins. J. Virol. 37:72–79.
32. Watanabe, T., S. Watanabe, T. Noda, Y. Fujii, and Y. Kawaoka. 2003. Ex-ploitation of nucleic acid packaging signals to generate a novel influenzavirus-based vector stably expressing two foreign genes. J. Virol. 77:10575–10583.
33. Zobel, A., G. Neumann, and G. Hobom. 1993. RNA polymerase I catalysedtranscription of insert viral cDNA. Nucleic Acids Res. 21:3607–3614.
11738 FLICK ET AL. J. VIROL.
on April 11, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from





























